
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Ecol. Evol. , 15 May 2018
Sec. Chemical Ecology
Volume 6 - 2018 | https://doi.org/10.3389/fevo.2018.00061
This article is part of the Research Topic IC-E3 - Insect Chemical Ecology, Ethology and Evolution View all 15 articles
 Tina Boddum1*†
Tina Boddum1*† Béla P. Molnár1,2†
Béla P. Molnár1,2† Sharon R. Hill1
Sharon R. Hill1 Göran Å. O. Birgersson1
Göran Å. O. Birgersson1 Bill S. Hansson3
Bill S. Hansson3 Kibrom B. Abreha1Erik Andreasson1‡Ylva Hillbur1‡
Kibrom B. Abreha1Erik Andreasson1‡Ylva Hillbur1‡Gall midges (Diptera: Cecidomyiidae) are a speciose family that, as adults, are short lived (lasting only a few days), they use olfactory cues for host and mate localization, and their host plant specificity is a key characteristic of the family. These traits make them good models with which to study the role of olfaction in speciation. The overall objective of this study was to analyze the host selection behavior of the gall midge, Contarinia nasturtii, a crucifer specialist that also is a crucifer pest. Here, we demonstrate that the host specificity of gravid C. nasturtii females was initiated by the olfactory-driven host plant choice during oviposition. Olfactory preference of the female, while narrow, encompassed more plants than were accepted for egg-laying, indicating that other factors following the initial olfactory attraction are involved in ultimate host choice. Similarly, C. nasturtii showed flexibility in host plant choice depending on which plants were available for oviposition. Larvae developed on host plants selected by females for oviposition. This slightly broader range of olfactory preference over acceptance, and the flexibility in host choice, might be the basis for the rapid speciation reported in the gall midge family. Furthermore, we assessed whether ubiquitous and/or family-specific plant odors are involved in the attraction of gravid C. nasturtii to their hosts. For that, we used the crucifer Arabidopsis thaliana, which has a broad range of defined ecotypes and large number of mutants. The attraction of C. nasturtii to two A. thaliana-types were tested; one that does not produce the ubiquitous green leaf volatiles (Columbia, Col-0), and a knock-out mutant which does not produce the crucifer-specific glucosinolates. Surprisingly, C. nasturtii was attracted to both types, indicating that neither of these compounds, nor their breakdown products (e.g., isothiocyanates), are essential for C. nasturtii host attraction.
With more than 6,000 described species (Gagné, 2010), the gall midges (Diptera: Cecidomyiidae) form a species-rich insect family with a rate of speciation that is considered to be more rapid than other dipterans (Harris and Foster, 1999). The vast majority of the gall midge species are monophagous herbivores (Gagné, 2004, 2010) specialized on a single plant family, plant species, plant organ or plant part (Stireman et al., 2008; Mishima et al., 2014).
Due to low larvae mobility and short lifespan (1–2 days on average) (Harris and Foster, 1999), females are under strong selection pressure to quickly select suitable (and ideally high quality) host plants (Delobel, 1982; Larsson and Strong, 1992; Renwick and Chew, 1994). Given the successful development of offspring, the misidentification of the preferred host plant, or acceptance of an alternative host plant, may interrupt gene flow between populations associated with the alternate host plant and the original host plant and can therefore be the first step in the formation of host races (Nylin and Janz, 2009). Host shift related formation of host races and sibling species has been described and suggested for several gall midges (e.g., Tabuchi and Amano, 2003; Cook et al., 2011; Molnár et al., 2018).
Gall midges make good models for studying speciation via host selection behavior. For this study, we chose the swede midge, Contarinia nasturtii (Kieffer, Diptera: Cecidomyiidae), as our model. Among the cecidomyiids, the genus Contarinia has a comparatively broad host range (Gagné, 2004) and is one of the largest genera with at least 300 species (Yukawa et al., 2005). Compared to other gall-inducing insects, Contarinia species are less closely associated with their hosts. Most of them do not induce host-specific galls, but live freely in flower heads or gregariously in leaf rolls or leaf fold galls (Yukawa, 2000; Gagné, 2004). This type of host association might be linked to an ability of female Contarinia to accept a broader range of hosts for oviposition and, consequently, to the rapid speciation observed in gall midges compared with other gall-forming insects.
The overall objective of this study is to analyze the host selection behavior of C. nasturtii females to assess the specificity of their host attraction and selection, evaluate the potential cues used in host attraction, and evaluate the potential for shifts among related plant taxa. Contarinia nasturtii has a preference for crucifers, in particular cabbage, on which it is an economically significant pest (Readshaw, 1965; Thygesen, 1966; Hallett, 2007; Olfert et al., 2009). To investigate the level of host selectivity among gravid C. nasturtii, we exposed these females to a range of natural plant stimuli, including crucifers and unrelated plants, in both choice and no-choice assays. We determined host plant preference of females based on the presence of eggs, and acceptance of the host based on the presence of larvae. We also investigated whether there is variation in the ability of females to assess host plant quality as reflected by different oviposition strategies.
Host plant attraction in herbivorous insects is predominantly guided by olfaction (Galanihe and Harris, 1997; Crook and Mordue, 1999; Chapman, 2003; Birkett et al., 2004; Balkenius et al., 2006; Couty et al., 2006; Hall et al., 2012; Saveer et al., 2012) and several studies have demonstrated host selection through the detection of host plant-specific ratios of ubiquitous volatiles (e.g., green leaf volatiles; GLVs; Bruce et al., 2005). There are, however, examples of insect specialists that are attracted to species- or family-specific volatile emissions, most notably insects that use the host-plant family-specific volatile cues of the crucifers (Brassicae spp.), the glucosinolates and isothiocyanates, for host plant recognition (Hopkins et al., 2009). The second part of this study is focused on the impact of olfactory cues on female swede midge host selection. Using well-characterized mutants of the crucifer Arabidopsis thaliana (L.) Heynhold (Brassicales: Brassicaceae), we provide insights into the mechanism underlying host attraction by C. nasturtii by investigating the role of the ubiquitous GLVs, and the family-specific glucosinolates in attracting gravid females (D'Auria and Gershenzon, 2005; Leonelli, 2007).
Contarinia nasturtii used in this study originated from a laboratory culture established in August 2000 at the Swedish University of Agricultural Sciences, Alnarp, Sweden. The laboratory cultures were supplemented with field-collected C. nasturtii annually. The laboratory culture consisted of cages housing up to 50 adult midges per cage. When the culture was supplemented, ~200 midges were added, distributed among the cages. The insects were kept in ventilated glass cages (50 × 50 × 50 cm) in a climate chamber at 22°C and 75% relative humidity with a 16:8 h light-dark photoperiod and were reared on cauliflower.
The host plants used in the oviposition choice, larval presence, egg deposition, and initial host odor attraction studies were cauliflower (Brassica oleracea Linnaeus, Brassicales: Brassicaceae), rapeseed (Brassica napus Linnaeus, Brassicales: Brassicaceae), wheat (Triticum aestivum Linnaeus, Poales: Poaceae), lettuce (Lactuca sativa Linnaeus, Asterales: Asteraceae), and thale cress (Arabidopsis thaliana ecotype Colombia-0; Col-0). All of the plants were grown in a climate chamber at 20°C and 75% relative humidity with a 10:14 h light-dark photoperiod. To control sciarid flies (Sciaridae), all of the seeds were treated with the nematodes Steinernema feltiae Filipjev (Rhabditida: Steinernematidae; Koppert, Netherlands) immediately after planting. The age of the plants when used was: cauliflower, 10 weeks; lettuce, 6 weeks; rapeseed (rosette stage), 7 weeks; rapeseed (ripening stage), 4 months; wheat (heading completed), 4 months; and A. thaliana, 8 weeks old. Three-week-old wheat and rapeseed plants were transferred to a climate chamber held at 4°C for 5 weeks, in order to reach the stage suitable for the experiments. The plants were subsequently transferred back to the original chamber to complete their development. Two stages of rapeseed were tested, as C. nasturtii are found on both stages in nature. The rosette stage most resembles the cauliflower (a preferred host plant of C. nasturtii) in size and shape.
For the odor profiling and behavioral experiments, two differing genotypes of A. thaliana plants were grown: the ecotype without green leaf volatiles (GLV), Col-0 (Duan et al., 2005), and the quadruple glucosinolate knockout mutant, cyp79B2cyp79B3myb28myb29 (QKO, Sun et al., 2009). The conditions differed slightly from those above, with the plants grown at 20 ± 1°C, 65 ± 5% relative humidity with 10:14 h light-dark photoperiod with fluorescent lamp light intensity of 200 μmol m−2 s−1. To ensure the same biomass and physiological status, QKO plants (which grow slower under these growth conditions than Col-0) were 1week older (4 weeks) than the Col-0 (3 weeks) at the time of the experiments. Cauliflower was grown for 8 weeks in a greenhouse under natural light conditions (12 ± 2:12 ± 2 h light-dark photoperiod).
The oviposition choice of female C. nasturtii was tested in mesh cages (200 × 70 × 70 cm) placed in a climate chamber at 22°C and 75% relative humidity. All insects emerged in the climate chamber where the experiments were conducted. The female C. nasturtii were provided with ample opportunities to mate prior to the experiments and were 1-day post-emergence at the onset of experiments. Host choice and larval presence was determined by enclosing 15 females with two plants of either cauliflower, rapeseed (ripening), rapeseed (rosette), A. thaliana, or lettuce for 24 h. The presence/absence of eggs was recorded after 3 days and presence/absence of larvae was recorded late in the larval stage, i.e., after 16 days. Six replicates were performed.
In the no-choice assay, two test plants of the same kind, i.e., rapeseed (rosette) or cauliflower, were placed at one end of the enclosure and the mated females were released at the other end. In the two-choice experiments, two plants of one type were placed at one end of the cage while two plants of the other type were placed at the other end. The insects were released in the center of the cage. In both experiments, 15 mated females (treated as above) and 5 males were used. The males were released to provide all possible opportunities for the all of the females in the assay to be mated. The females were given 24 h to oviposit. The plants were then moved to another climate chamber and kept at 22°C and 75% relative humidity. After 3 days, one of each plant type was examined for eggs and after 16 days the second plant was examined for larvae. Six replicates were performed per assay and plant-type combination.
One mated female was placed in a cage (50 × 50 × 50 cm) together with either six cauliflower or six rapeseed (rosette) plants. We recorded whether a female did or did not oviposit on the plants, and whether the female oviposited on a single plant or on several plants after 3 days. After infestation, the plants were kept for 14 days in a climate chamber at 22°C and 75% relative humidity, and then examined for damage caused by gall midge infestation. If any damage caused by larval feeding was observed, it was concluded that the plant was accepted as host by the female. Twenty replicates were conducted for cauliflower and 26 replicates for rapeseed.
Female C. nasturtii attraction to the plants was tested in a glass Y-tube olfactometer described by Andersson et al. (2009). The air was filtered and humidified by pumping it through two 250 ml gas-wash bottles, one with granulated activated charcoal and one with distilled water, prior to dividing into two streams. The airflow (0.6 l min−1) through the two Teflon tubes passed into polyethylene cooking bags (45 × 55 cm, Toppits, Germany) which contained the test stimulus. Cooking bags containing plants were compared to either an empty cooking bag (blank) or to another plant. To control for position effects, empty bags were compared to empty bags. The air now containing the stimulus passed through more Teflon tubing into the arms of the Y-tube olfactometer. Visual cues were excluded by placing the Y-tube in a box (40 × 40 × 40 cm) covered with a white cotton cloth. The flight of the C. nasturtii was activated by illumining the set-up with a lamp (Massive, 906609, 400 W, Belgium) placed 30 cm from the set-up in the upwind direction. Experiments were conducted in a climate chamber at 22°C and 75% relative humidity and 16:8 h light-dark photoperiod. Insects emerged in this climate chamber prior to the experiments.
The gall midges were introduced into the Y-tube at the downwind entry arm. The experiments started at 4 pm and lasted until 9 am the following day. The light was on between 4 and 8 p.m. to provide the same conditions as in the climate chamber. To avoid escapes from the Y-tube, insect netting covered the entry and a capture cage was connected to each arm. After each experiment, all glassware was placed at 350°C for 8 h, and both the Teflon tubing and the connections were washed in 70% ethanol. To avoid left-right bias the “blank arm” was exchanged between experiments. In this set-up, attraction to the following plants were compared to the blank: cauliflower, rapeseed (ripening), A. thaliana (CO-1), rapeseed (rosette), wheat, and lettuce. Furthermore, the attraction to the following plants were compared: cauliflower to A. thaliana (CO-1), cauliflower to rapeseed (rosette), and A. thaliana (CO-1) to A. thaliana (QKO). In each replication, 10–20 mated females were released simultaneously in the entry arm. The number of emerging midges varied from day-to-day, which accounts for the variable number of females in each replicate. Each treatment was replicated 5–6 times, until a total of 90–110 females were tested under each condition.
Plants for odor collection were grown in pots completely wrapped in aluminum foil, except for small holes above the seeds through which the plants grew. To damage A. thaliana and cauliflower plants for odor collection and behavioral assays, we cut 10 of the major leaves to the mid-rib at three random locations with a pair of micro-scissors just prior to experiments. Odor collections were performed over a 4 h period. Damaged and undamaged plants were wrapped in polyethylene cooking bags (45 × 55 cm, Toppits, Germany) with a solid phase microextraction (SPME) fiber (Supelco, Sigma-Aldrich Inc., St. Louis, MO, USA 75 μm Carboxen®/Polydimethyl-siloxane; CAR/PDMS) inserted close to the leaves. Immediately after odor collection, the fiber was injected into a combined gas chromatograph and mass spectrometer (GC-MS; HP 6890 GC and 5975 MS, Agilent Technologies Inc., Santa Clara, CA, USA), operated in the electron impact ionization mode at 70 eV. The GC was equipped with fused silica capillary columns (30 m, 0.25 mm, d.f. 1/4 0.25 mm), DB-wax (J&W Scientific, Folsom, CA, USA) or HP-5MS (Agilent Technologies). The GC-MS and the post-run data analyses were performed according to Saveer et al. (2012). A helium mobile phase was used at an average linear flow rate of 35 cm s−1. Each sample (2 μl) was injected (splitless mode, 30 s, injector temperature 225°C). For both columns, the GC oven temperature was programmed from 30°C (3 min hold) at 8°C min−1 to 225°C (5 min hold). Compounds were tentatively identified by matching their mass spectra with those in the MS Libraries (NIST 11 and Wiley) and were verified by comparing with injected synthetic reference compounds with the published Kovat's index (Ki) values.
Data from the choice test “female oviposition choice and larval presence” were analyzed using the Wilcoxon signed-rank test (Minitab, Version 14, Minitab Inc., State College, PA). Mann-Whitney tests were used to analyze the results from the olfactory-based host choice assays, and the assays comparing female attraction to A. thaliana host plants lacking either GL Vs or glucosinolates, as well as to cauliflower (Minitab, Version 14, Minitab Inc., State College, PA). The results from the distribution of eggs by individual females were analyzed using a nominal logistic regression (JMP® Pro, Version 12, SAS Institute Inc., Cary, NC, 1989–2007).
The females oviposited on cauliflower, rapeseed (ripening), and rapeseed (rosette) (Table 1). All three plants supported some larval development, as assessed by larval presence 16 days post-oviposition. No eggs were deposited on either A. thaliana or lettuce, and therefore, no larvae were observed (Table 1).

Table 1. Presence (+) or absence (–) of eggs and larvae on cauliflower, rapeseed (ripening), rapeseed (rosette), Arabidopsis thaliana and lettuce after 15 female Contarinia nasturtii had been enclosed with the plants for 24 h in a no-choice assay.
Both cauliflower and rosette stage rapeseed were found to be acceptable as host plants to female C. nasturtii in the no-choice assay (Figure 1A). There was no difference found in the number of larvae on either cauliflower or rosette stage rapeseed (U = 12.0; P = 0.368, n = 6; Figure 1A). However, cauliflower was found to be the preferred host in a two-choice assay, with more larvae found on the cauliflower than on rosette stage rapeseed (U = 33.0; P = 0.047, n = 6; Figure 1B).
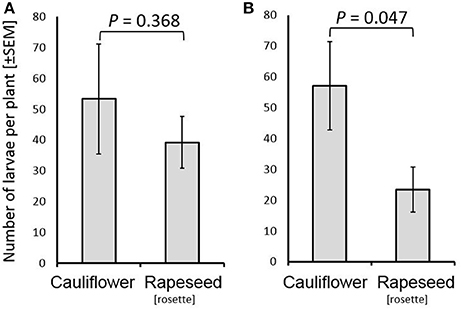
Figure 1. Number of Contarinia nasturtii larvae on cauliflower and rapeseed (rosette) in a no-choice assay (A), and on cauliflower and rapeseed (rosette) in a two-choice assay (B). Larval presence on the two host types was compared by Mann-Whitney tests for arcsine-transformed data. P-values are displayed over each graph. n = 6 for each choice type. Fifteen females were released in each replicate.
To test whether the female swede midges are able to evaluate oviposition sites, one female was presented with six cauliflower or rosette stage rapeseed plants. We recorded whether the female did not oviposit, only oviposited on one plant, or chose multiple oviposition sites. For both rapeseed and cauliflower, more females oviposited on several plants compared to females that did not oviposit at all (χ2 = 7.37, P = 0.007 and χ2 = 7.37, P = 0.007, respectively; Figure 2). Regardless of host species, approximately half of the swede midge females laid eggs on more than one plant (Figure 2). This suggests that host species does not impact the bet-hedging strategy of laying eggs across multiple plants (Figure 2). The choice between ovipositing on a single plant or not at all, however, appears to be affected by the host plant. Similar proportions were observed in the rapeseed rosette stage assays between those containing plants without an infestation, and those with only a single plant infested (Figure 2). In the cauliflower assays, the proportion of single plant infestations was similar to the proportion of multiple plants infestations, with <10% of the trials exhibiting no infestation (Figure 2).
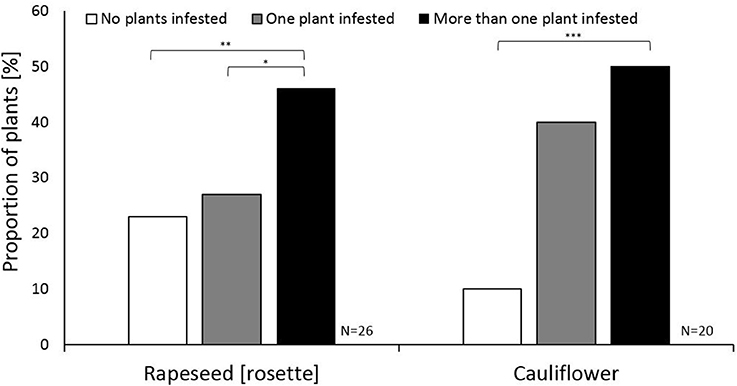
Figure 2. The host acceptance strategy of individual Contarinia nasturtii females on cauliflower and rapeseed (rosette) plants in the host acceptance assay. White, gray, and black bars indicate the proportion of C. nasturtii laying no eggs, ovipositing on only one plant, or ovipositing on more than one plant. The Y-axis shows the proportion of females choosing a certain oviposition strategy. Nominal logistic regression was used for comparing cauliflower and rapeseed with significant differences marked with asterisks as follows: *P < 0.05; **P < 0.01; ***P < 0.001. n = 20–26 for each plant type.
Female swede midges were more attracted to the volatile emissions from cauliflower (W = 39.0, P = 0.02), ripening rapeseed (W = 15.5, P = 0.016), and A. thaliana Col-0 ecotype (W = 56.0, P = 0.008) than to the blank in the Y-tube olfactometer (Figures 3A–C). In addition, the volatiles of A. thaliana Col-0 ecotype and cauliflower were equally attractive to female midges (W = 56.5, P = 0.66; Figure 3D). The response to the younger, rosette stage rapeseed plants (W = 35.0, P = 0.03; Figure 3E) was less than the response to the blank, and the females were more attracted to cauliflower than the rosette rapeseed (Figure 3F; W = 36, P = 0.04). The response to wheat (Figure 3G) or lettuce (Figure 3H) (i.e., non-crucifers) did not differ from blanks (W = 33.0, P = 0.38 and W = 24.0, P = 0.531, respectively), nor was there a difference when two empty arms were compared (W = 27.5, P = 1.0, Figure 3I).
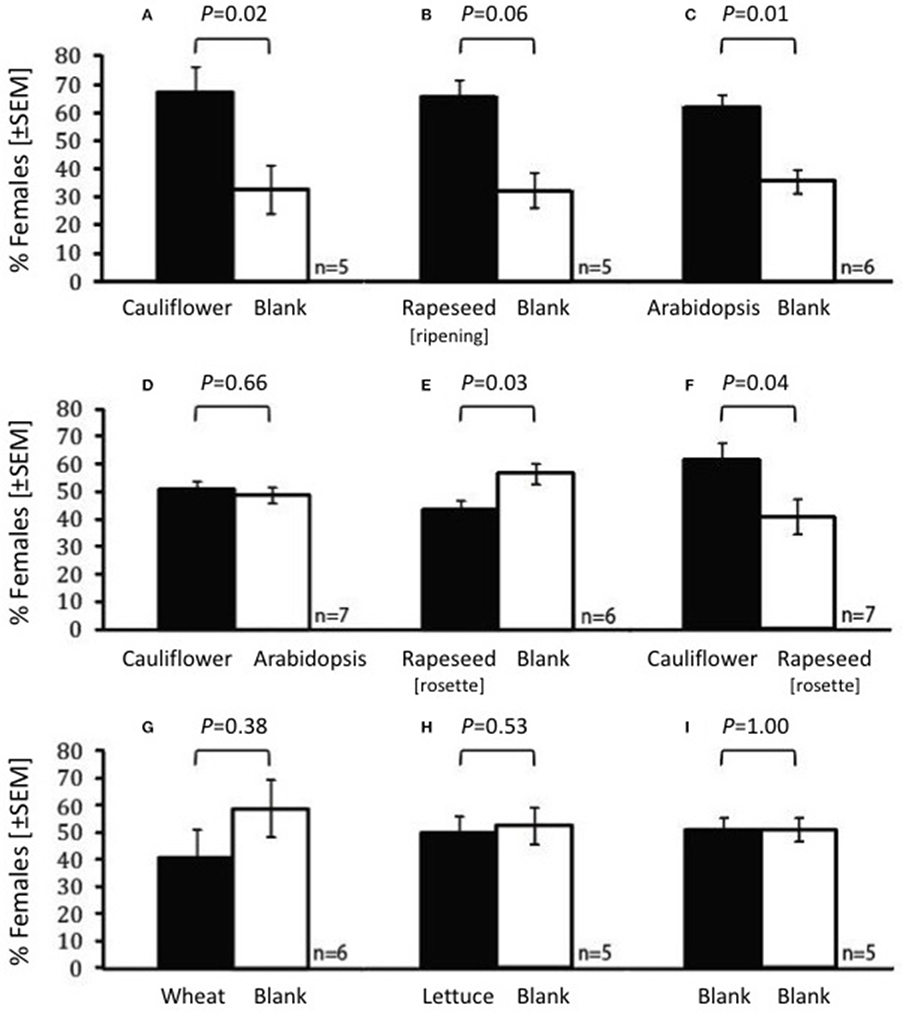
Figure 3. Olfactory preference of female Contarinia nasturtii in the olfactometer when the following are compared: (A) cauliflower vs. blank, (B) rapeseed (ripening) vs. blank, (C) Arabidopsis thaliana vs. blank, (D) cauliflower vs. A. thaliana, (E) rapeseed (rosette) vs. blank, (F) cauliflower vs. rapeseed (rosette), (G) wheat vs. blank, (H) lettuce vs. blank and (I) blank vs. blank. Female attraction to the two odors was compared by Mann-Whitney test. P-values are displayed over each comparison. n = 5–7 per combination, each using a total of 94–107 females.
As no eggs were found on the GLV-lacking A. thaliana (CO-1; Table 1), the expectation was that the odors from this plant would not be attractive to gravid females. This was not, in fact, what was observed. Female gall midges were not significantly less attracted to the odors emanating from the GLV-lacking A. thaliana Col-0 ecotype than to the cauliflower odors (Figure 3D). In addition, female midges were attracted to A. thaliana Col-0 ecotype volatiles when compared with a blank (W = 72.0; P = 0.02; Figure 4i), just as was observed for the females to cauliflower volatiles (W = 70.5; P = 0.03, respectively; Figure 4ii). A direct comparison between the GLV-lacking volatiles of Col-0 and glucosinolate-lacking volatiles of QKO plants revealed that they were equally attractive to swede midge females (W = 53.0; P = 1; Figure 4iii).
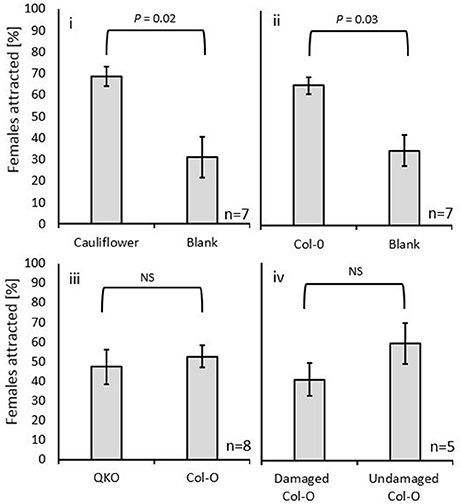
Figure 4. The attraction of the C. nasturtii in Y-tube bioassays to (i) cauliflower vs. blank; (ii) Arabidopsis thaliana Co1-0 (no green leaf volatiles) vs. blank; (iii) A. thaliana Co1-0 vs. A. thaliana glucosinolate knockout plants (QKO); and (iv) undamaged vs. mechanically damaged A. thaliana Co1-0. Female attraction to the two odors was compared by Wilcoxon signed-rank. P-values are displayed over each comparison.
We found no differences in the attraction of female swede midges to volatiles of damaged vs. undamaged Col-0 plants (W = 6.25; P = 94.8; Figure 4iv). An analysis of the volatile profiles of Col-0 and cauliflower plants revealed five compounds that were shared among damaged and undamaged cauliflower and A. thaliana Col-0 plants, namely decanal, 3-carene, sulcatone, limonene, and α-pinene (Figure 5).
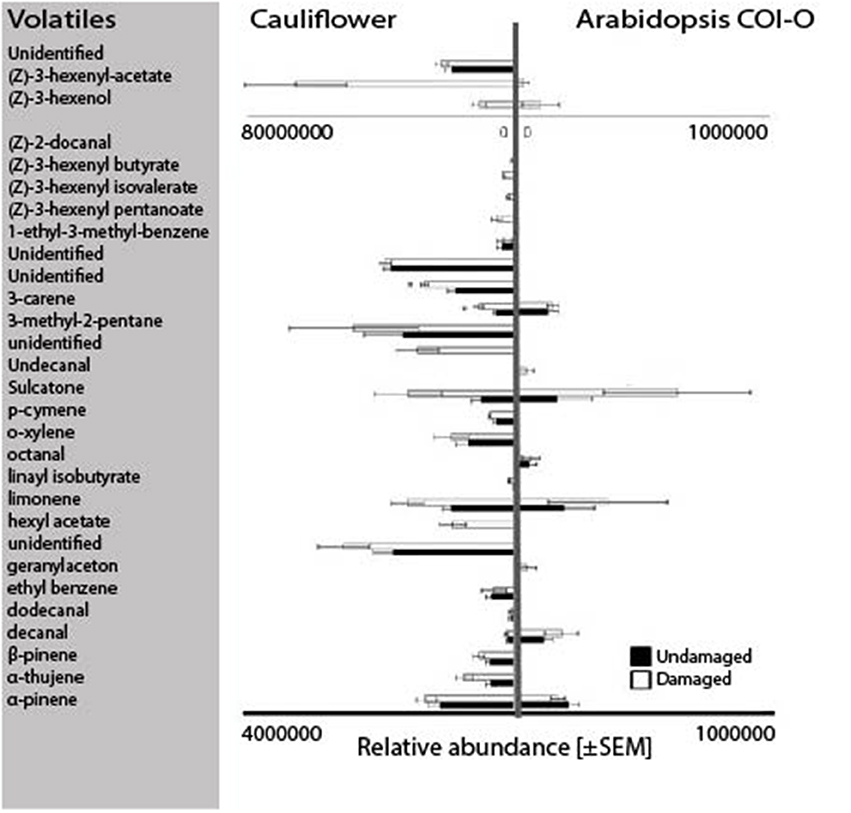
Figure 5. The volatile profiles of cauliflower (n = 5) and Arabidopsis thaliana Col-0 (n = 3) for undamaged (black) and damaged (white) plants. The relative abundance of the volatiles is based on solid phase microextraction (SPME) odor collection analyzed using GC-MS. Statistics: Mann-Whitney test with significant differences marked with an asterisk (P < 0.05). Note: the differing scale bars for A. thaliana Col-0 (0-1 × 106) and for cauliflower (0-4 × 106 or 0–8 × 107), due to large variation in emitted amounts.
Olfactory cues play a strong role in the host plant recognition of the swede midge, C. nasturtii. Females were not only able to differentiate between plant species solely based on olfactory cues, but also responded differently to the volatiles of different phenological stages of the same plant species.
Of the two predominating hypotheses concerning how herbivorous insects locate their host plants, both the attraction to ubiquitous volatiles (e.g., GLVs; Bruce et al., 2005) and attraction to family-specific volatiles (e.g., glucosinolates; Hopkins et al., 2009) appeared to be plausible for swede midge attraction to the crucifers. However, female attraction to an A. thaliana ecotype lacking GLVs does not differ from their attraction to their host plant cauliflower, or to an A. thaliana mutant that does not produce glucosinolates. From this it is reasonable to propose that other volatile compounds emitted by these plants mediate attraction. By comparing the volatiles emitted from two of the attractive plants, cauliflower and A. thaliana Col-0, we identified five shared compounds that are potentially involved in swede midge host recognition: decanal, sulcatone, 3-carene, limonene, and α-pinene. Sulcatone, 3-carene and α-pinene are also host cues for another gall midge species, the orange wheat blossom midge (Sitodiplosis mosellana; Birkett et al., 2004; Bruce and Pickett, 2011). In the current study, C. nasturtii were not attracted to wheat. This indicates that these gall midges utilize similar compounds, however in specific combinations with other compounds for host recognition.
No females oviposited on lettuce, possibly because its odor profile differed to such an extent from that of the plants in the swede midge host range that it was not even recognized as a host; lettuce emitted only limonene and not decanal, 3-carene, sulcatone, or α-pinene (Lonchamp et al., 2009) of the candidate volatiles in putative C. nasturtii attractive blend. The use of ubiquitous volatiles (potentially shared between host plants and non-host plants) may increase the adaptive potential for C. nasturtii. In a host shift situation, the olfactory system does not have to adapt to the host specific volatiles. With the frequent host shift of Contarinia species, utilizing compounds that are common for many plant genera would provide an evolutionary advantage. Heritable changes in insect plant recognition mechanisms have been proposed as the primary event in the evolution of new insect-plant associations (Dres and Mallet, 2002). A change in how the odor is perceived, e.g., from attractive to neutral or repellent, can be altered by relatively small genetic changes, such as different receptor expression, altered sensitivity and coding of key odors (Olsson S. et al., 2006), or new arborizations of olfactory receptor neurons (Karpati et al., 2008). Thus, a genetic change in gall midge host preference is, in principle, all that may be required to initiate a successful host shift. The odor-driven host selection behavior of C. nasturtii demonstrated here can be part of the explanation for the rapid speciation in the gall midge family.
When females had been allowed to oviposit on a single species of host plant—either on rapeseed in the rosette stage or on cauliflower—comparable numbers of larvae could be found on both hosts. However, when the female midge could choose between the two hosts, cauliflower was preferred. For an insect with short life span and limited mobility, such as C. nasturtii, plasticity in host preference is a good evolutionary strategy (Wiklund, 1975, 1981; Larsson and Ekbom, 1995). A female that can distinguish a suitable from an unsuitable host (but still accept an alternative host when nothing else is available) will have increased fitness compared to a solely discriminative female (if the alternative plant supports larval development). Although attractive, swede midges did not oviposit on A. thaliana. Such a lack of suitability as a host could be attributed to physical plant defenses (e.g., trichomes on the leaves and stems) (Mauricio and Rausher, 1997; Schoonhoven et al., 2005).
An association between female host plant preference and larval performance has been reported for other herbivorous gall midges (Åhman, 1985), while other studies demonstrate weak preference-performance associations (Larsson and Ekbom, 1995; Nyman et al., 2011). The relevant question, from the perspective of natural selection, is to what extent swede midges “hedge their bets.” We assessed the female oviposition behavior by testing individual females in a no-choice situation. For both plants, 50% of the tested females oviposited on multiple plants. The other females displayed highly selective behavior, depositing either all of their eggs on one plant or not ovipositing at all. This selective behavior appeared to be host-specific, as the proportion of females ovipositing on one plant did not differ from the proportion ovipositing on multiple plants in cauliflower, however fewer females laid all their eggs on a single plant when presented with rapeseed. In addition, more females did not oviposit when presented with rapeseed compared to cauliflower. The combination of variable and selective oviposition choice strategies within a population favors a host plant shift as a response to a situation in which the preferred host is not available (Wiklund, 1981). The findings in this paper indicate that in the absence of the preferred crucifer host (Thygesen, 1966; Olfert et al., 2009), other available plants might be used as host, maintaining a low level of infestation in a region, awaiting the return of the preferred host in subsequent years (e.g., during a crop rotation scheme). Furthermore, the identified variability in host preference in a C. nasturtii population could lead to C. nasturtii becoming a pest on other crops with an overlapping volatile profile.
The current pest management of C. nasturtii is based on pheromone traps that predominantly attract males (Hillbur et al., 2005). This study has demonstrated some candidate volatile mixtures that are likely attractive to female swede midges, which could be used to develop volatile lures to control the female population. This could be the first step toward the development of a trap for gravid females to supplement the pheromone traps.
TB, YH, and BM: Designed the study; TB and BM: Conducted the experiments and wrote the manuscript; KA: Helped design the A. thaliana experiments and write the manuscript; SH, GB, BH, EA, and YH: Supervised the experiments and supported the writing of the manuscript.
This study was supported by the Linnaeus initiative Insect Chemical Ecology, Ethology and Evolution ICE3 (The Swedish Research Council Formas, SLU).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The QKO seeds were kindly provided by Barbara Ann Halkier, University of Copenhagen.
Åhman, I. (1985). Larval feeding period and growth of Dasineura brassicae (Diptera) on Brassica host plants. Oikos 44, 191–194. doi: 10.2307/3544061
Andersson, M., Haftmann, J., Stuart, J., Cambron, S., Harris, M., Foster, S., et al. (2009). Identification of sex pheromone components of the Hessian fly, Mayetiola destructor. J. Chem. Ecol. 35, 81–95. doi: 10.1007/s10886-008-9569-1
Balkenius, A., Rosen, W., and Kelber, A. (2006). The relative importance of olfaction and vision in a diurnal and a nocturnal hawkmoth. J. Comparat. Physiol. A Sensory Neural Behav. Physiol. 192, 431–437. doi: 10.1007/s00359-005-0081-6
Birkett, M. A., Bruce, T. J. A., Martin, J. L., Smart, L. E., Oakley, J., and Wadhams, L. J. (2004). Responses of female orange wheat blossom midge, Sitodiplosis mosellana, to wheat panicle volatiles. J. Chem. Ecol. 30, 1319–1328. doi: 10.1023/B:JOEC.0000037742.05022.9f
Bruce, T. J. A., and Pickett, J. A. (2011). Perception of plant volatile blends by herbivorous insects-finding the right mix. Phytochemistry 72, 1605–1611. doi: 10.1016/j.phytochem.2011.04.01
Bruce, T., Wadhams, L., and Woodcock, C. (2005). Insect host location: a volatile situation. Trends Plant Sci. 10, 269–274. doi: 10.1016/j.tplants.2005.04.003
Chapman, R. F. (2003). Contact chemoreception in feeding by phytophagous insects. Annu. Rev. Entomol. 48, 455–484. doi: 10.1146/annurev.ento.48.091801.112629
Cook, M. A., Ozeroff, S. N., Fitzpatrick, S. M., and Roitberg, B. D. (2011). Host–associated differentiation in the reproductive behaviour of cecidomyiid midges on cranberry and blueberry. Entomol. Exp. Appl. 141, 8–14. doi: 10.1111/j.1570-7458.2011.01166.x
Couty, A., Van Emden, H., Perry, J. N., Hardie, J., Pickett, J. A., and Wadhams, L. J. (2006). The roles of olfaction and vision in host-plant finding by the diamondback moth, Plutella xylostella. Physiol. Entomol. 31, 134–145. doi: 10.1111/j.1365-3032.2006.00499.x
Crook, D. J., and Mordue, A. J. (1999). Olfactory responses and sensilla morphology of the blackcurrant leaf midge Dasineura tetensi. Entomol. Exp. Appl. 91, 37–50. doi: 10.1046/j.1570-7458.1999.00464.x
D'Auria, J., and Gershenzon, J. (2005). The secondary metabolism of Arabidopsis thaliana: growing like a weed. Curr. Opin. Plant Biol. 8, 308–316. doi: 10.1016/j.pbi.2005.03.012
Delobel, A. G. L. (1982). Oviposition and larval survival of the sorghum shoot fly, Atherigona soccata rond, as influenced by the size of its host plant (Diptera, Muscidae). Entomol. J. Appl. Entomol. 93, 31–38. doi: 10.1111/j.1439-0418.1982.tb03567.x
Dres, M., and Mallet, J. (2002). Host races in plant-feeding insects and their importance in sympatric speciation. Philos. Trans. R. Soc. B Biol. Sci. 357, 471–492. doi: 10.1098/rstb.2002.1059
Duan, H., Huang, M., Palacio, K., and Schuler, M. (2005). Variations in CYP74B2 (hydroperoxide lyase) gene expression differentially affect hexenal signaling in the Columbia and Landsberg Erecta ecotypes of Arabidopsis. Plant Physiol. 139, 1529–1544. doi: 10.1104/pp.105.067249
Gagné, R. J. (2004). A Catalog of the Cecidomyiidae (Diptera) of the World. Washington, DC: The Entomological Society of Washington.
Gagné, R. J. (2010). A catalog of the Cecidomyiidae (Diptera) of the world. digital version 1. Mem. Entomol. Soc. Washingt. 25, 1–544.
Galanihe, L. D., and Harris, M. O. (1997). Plant volatiles mediate host-finding behavior of the apple leafcurling midge. J. Chem. Ecol. 23, 2639–2655. doi: 10.1023/A:1022546506965
Hall, D., Amarawardana, L., Cross, J., Francke, W., Boddum, T., and Hillbur, Y. (2012). The chemical ecology of Cecidomyiid midges (Diptera: Cecidomyiidae). J. Chem. Ecol. 38, 2–22. doi: 10.1007/s10886-011-0053-y
Hallett, R. H. (2007). Host plant susceptibility to the swede midge (Diptera: Cecidomyiidae). J. Econ. Entomol. 100, 1335–1343. doi: 10.1093/jee/100.4.1335
Harris, M. O., and Foster, S. P. (1999). “Gall midges,” in Pheromones of Non-Lepidopteran Insects Associated With Agricultural Plants, ed J. Hardie (Oxford: CAB International), 27–49.
Hillbur, Y., Celander, M., Baur, R., Rauscher, S., Haftman, J., Franke, S., et al. (2005). Identification of the sex pheromone of the swede midge, Contarinia nasturtii. J. Chem. Ecol. 31, 1807–1828. doi: 10.1007/s10886-005-5928-3
Hopkins, R., van Dam, N., and van Loon, J. (2009). Role of glucosinolates in insect-plant relationship and multitrophic interactions. Annu. Rev. Entomol. 54. 57–83. doi: 10.1146/annurev.ento.54.110807.090623
Karpati, Z., Dekker, T., and Hansson, B. S. (2008). Reversed functional topology in the antennal lobe of the male european corn borer. J. Exp. Biol. 211, 2841–2848. doi: 10.1242/jeb.017319
Larsson, S., and Ekbom, B. (1995). Oviposition mistakes in herbivorous insects -confusion or a step toward a new host plant. Oikos 72, 155–160. doi: 10.2307/3546051
Larsson, S., and Strong, D. R. (1992). Oviposition choice and larval survival of Dasineura marginemtorquens (Diptera, Cecidomyiidae) on resistant and susceptible Salix viminalis. Ecol. Entomol. 17, 227–232. doi: 10.1111/j.1365-2311.1992.tb01051.x
Leonelli, S. (2007). Arabidopsis, the botanical Drosophila: from mouse cress to model organism. Endeavour 31, 34–38. doi: 10.1016/j.endeavour.2007.01.003
Lonchamp, J., Barry-Ryan, C., and Devereux, M. (2009). Identification of volatile quality markers of ready-to-use lettuce and cabbage. Food Res. Int. 42, 1077–1086. doi: 10.1016/j.foodres.2009.05.002
Mauricio, R., and Rausher, M. (1997). Experimental manipulation of putative selective agents provides evidence for the role of natural enemies in the evolution of plant defense. Evolution 51, 1435–1444. doi: 10.1111/j.1558-5646.1997.tb01467
Mishima, M., Sato, S., Tsuda, K., and Yukawa, J. (2014). Sexual isolation between two known intraspecific populations of Hartigiola (Diptera: Cecidomyiidae) that induce leaf galls on upper and lower surfaces of Fagus crenata (Fagales: Fagaceae), indicating possible diversification into sibling species. Arthropod Biol. 107, 789–798. doi: 10.1603/AN13125
Molnár, B. P., Boddum, T., Hill, S. R., Hansson, B. S., Ylva, H., and Göran, B. (2018). Ecological and phylogenetic relationships shape the peripheral olfactory systems of highly specialized gall midges (Cecidomiiydae). Front. Physiol. 9:323. doi: 10.3389/fphys.2018.00323
Nylin, S., and Janz, N. (2009). Butterfly host plant range: an example of plasticity as a promoter of speciation? Evol. Ecol. 23, 137–146. doi: 10.1007/s10682-007-9205-5
Nyman, T., Paajanen, R., Heiska, S., and Julkunen-Tiitto, R. (2011). Preference-performance relationship in the gall midge Rabdophaga rosaria: insights from a common-garden experiment with nine willow clones. Ecol. Entomol. 36, 200–211. doi: 10.1111/j.1365-2311.2011.01260.x
Olfert, O., Hallett, R., Weiss, R. M., Soroka, J., and Goodfellow, S. (2009). Potential distribution and relative abundance of swede midge, Contarinia nasturtii, an invasive pest in Canada. Entomol. Exp. Appl. 129, 221–228. doi: 10.1111/j.1570-7458.2006.00445.x
Olsson, S., Linn, C. E., and Roelofs, W. L. (2006). The chemosensory basis for behavioral divergence involved in sympatric host shifts. I. Characterizing olfactory receptor neuron classes responding to key host volatiles. J. Compar. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 192, 279–288. doi: 10.1007/s00359-005-0069-2
Readshaw, J. (1965). The ecology of the swede midge, Contarinia nasturtii (kieff.) (Diptera, Cecidomyiidae). Life-history and influence of temperature and moisture on development. Bull. Entomol. Res. 58, 25–29. doi: 10.1017/S0007485300056686
Renwick, J. A. A., and Chew, F. S. (1994). Oviposition behavior in Lepidoptera. Annu. Rev. Entomol. 39, 377–400. doi: 10.1146/annurev.en.39.010194.002113
Saveer, A. M., Kromann, S. H., Birgersson, G., Bengtsson, M., Lindblom, T., Balkenius, A., et al. (2012). Floral to green: mating switches moth olfactory coding and preference. Proc. R. Soc. B Biol. Sci. 279, 2314–2322. doi: 10.1098/rspb.2011.2710
Schoonhoven, J. M., van Loon, J. J. A., and Dicke, M. (2005). Insect-Plant Biology, 2nd Edn. Oxford; New York, NY: Oxford University Press.
Stireman, J. O. III., Janson, E. M., Carr, T. G., Devlin, H., and Abbot, P. (2008). Evolutionary radiation of asteromyia carbonifera (Diptera: Cecidomyiidae) gall morphotypes on the goldenrod Solidago Altissima (Asteraceae). Biol. J. Linnean Soc. 95, 840–858. doi: 10.1111/j.1095-8312.2008.01101.x
Sun, J., Sonderby, I., Halkier, B., Jander, G., and de Vos, M. (2009). Non-volatile intact indole glucosinolates are host recognition cues for ovipositing Plutella xylostella. J. Chem. Ecol. 35, 1427–1436. doi: 10.1007/s10886-009-9723-4
Tabuchi, K., and Amano, H. (2003). Host-associated differences in emergence pattern, reproductive behavior and life history of Asteralobia sasakii (Monzen) (Diptera: Cecidomyiidae) between populations on Ilex crenata and I. integra (Aquifoliaceae). Appl. Entomol. Zool 38, 501–508. doi: 10.1303/aez.2003.501
Thygesen, T. (1966). The swede midge (Contarinia nasturtii kieff). Observation on biology, survey of economic importance and control experiments. Tidsskr Planteavl 70, 179–197.
Wiklund, C. (1975). Evolutionary relationship between adult oviposition preferences and larval host plant range in Papilio machaon L. Oecologia 18, 185–197. doi: 10.1007/BF00345421
Wiklund, C. (1981). Generalist vs specialist oviposition behavior in Papilio machaon (Lepidoptera) and functional-aspects on the hierarchy of oviposition preferences. Oikos 36, 163–170. doi: 10.2307/3544441
Yukawa, J. (2000). Synchronization of gallers with host plant phenology. Popul. Ecol. 42, 105–463. doi: 10.1007/PL00011989
Keywords: Arabidopsis, host choice, gall midge, olfaction, specificity
Citation: Boddum T, Molnár BP, Hill SR, Birgersson GÅO, Hansson BS, Abreha KB, Andreasson E and Hillbur Y (2018) Host Attraction and Selection in the Swede Midge (Contarinia nasturtii). Front. Ecol. Evol. 6:61. doi: 10.3389/fevo.2018.00061
Received: 12 October 2017; Accepted: 23 April 2018;
Published: 15 May 2018.
Edited by:
Xavier Martini, University of Florida, United StatesReviewed by:
Lauren E. Des Marteaux, Biology Centre (ASCR), CzechiaCopyright © 2018 Boddum, Molnár, Hill, Birgersson, Hansson, Abreha, Andreasson and Hillbur. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tina Boddum, dGluYWJvZGR1bUBnbWFpbC5jb20=
†Tina Boddum and Béla P. Molnár share first authorship.
‡Erik Andreasson and Ylva Hillbur share senior authorship.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.