 Jessica Henneken
Jessica Henneken Jason Q. D. Goodger
Jason Q. D. Goodger Therésa M. Jones
Therésa M. Jones Mark A. Elgar
Mark A. Elgar- The School of Biosciences, The University of Melbourne, Melbourne, VIC, Australia
Diet is arguably the most significant environmental factor shaping chemical signals in animals. In rare cases, dietary components are converted directly into pheromones or signature mixtures but more generally variation in an individual's diet influences their overall condition and thus capacity to synthesize the signal. Typically, diet is variable between individuals of the same species and this can lead to variation in signals. This variation presents specific challenges to receivers, who must be able to recognize and respond to a greater range of signals. However, such variation might also provide the receiver with key information about the signaller, allowing them to respond to the signal advantageously. Here we investigate how dietary-mediated pheromones and signature mixtures can provide the receiver with reliable information about the signaller, ultimately benefiting the receiver in a way not achievable by a static signal.
Introduction
Pheromones (from the Greek pherein, to carry or transfer and hormon to excite) are chemical signals that transmit information between individuals of the same species. Typically, pheromones consist of particular blends of chemicals, some of which are species-specific, while others may be produced by multiple and often very different species (Wyatt, 2014). Pheromones alter the behavior of another organism and evolve precisely because of the nature of this effect (Maynard Smith and Harper, 2003). The receiver's capacity to detect and respond to the signal has similarly evolved to maximize detection (see Peso et al., 2015), thereby optimizing the effectiveness of pheromones. Wyatt (2010) distinguishes between pheromones, which typically elicit a predisposed response in the receiver, and signature mixtures that first require the receiver to learn the chemical signal. In contrast to pheromones, signature mixtures allow receivers to identify the signaller as a specific individual (Wyatt, 2010). Both pheromones and signature mixtures are employed in communication between individuals of the same species, but signature mixtures are used to distinguish individuals, for example as siblings or non-siblings or to distinguish between nest-mates and non-nest-mates.
Successful communication between animals requires accurate signals that are correctly perceived. Correct perception requires, in the first instance, the receiver to recognize the signal. If the signal varies between individuals within a group or species, we may expect receivers to experience impaired recognition of the signal. However, most signals, regardless of their modality, demonstrate some level of variation, resulting in chemical signals that may vary between populations (Palacio Cortés et al., 2010) or even individuals (Svensson et al., 1997) of the same species. Variation in chemical signals can be genetically determined and, in some animal systems, the signal's chemical profile is highly heritable (Svensson et al., 2002; Tabata et al., 2006; Thomas and Simmons, 2008). However, increasing evidence suggests that the environment can also influence an individual's ability to synthesize and emit chemical signals. Environmental factors that may influence chemical signals include temperature (Ono, 1993), humidity (Hock et al., 2014), and population-specific influences such as density (Anderbrant et al., 1985). Variation in chemical signals, whether genetically or environmentally driven, has significant implications for the receiver (see also Symonds and Elgar, 2008). Should the receiver respond to all variants of the signal and, if so, should that response be consistent?
Diet is likely one of the most significant environmental factors shaping chemical signals. Numerous studies demonstrate dietary influences on the chemical blend of a pheromone or signature mixture and, in most cases, subsequent variation in the receiver's response (Table 1). As demonstrated in Table 1, diet-mediated pheromones and signature mixtures span a wide range of taxa and the functional role of these signals are also diverse. Less well appreciated is the fact that dietary driven variation in pheromones and signature mixtures presents specific challengers to receivers, requiring them to recognize and respond correctly to a wider range of signals. In some cases this appears to decrease the reliability of the signal (Vonshak et al., 2009), causing the receiver to respond in a way that may be detrimental to both the signaller and the receiver. Nevertheless, in most cases, dietary-mediated signals appear to provide the receiver with information about the signaller, eliciting a beneficial behavioral response from the receiver (Martín and López, 2006; South et al., 2011; Liedo et al., 2013).
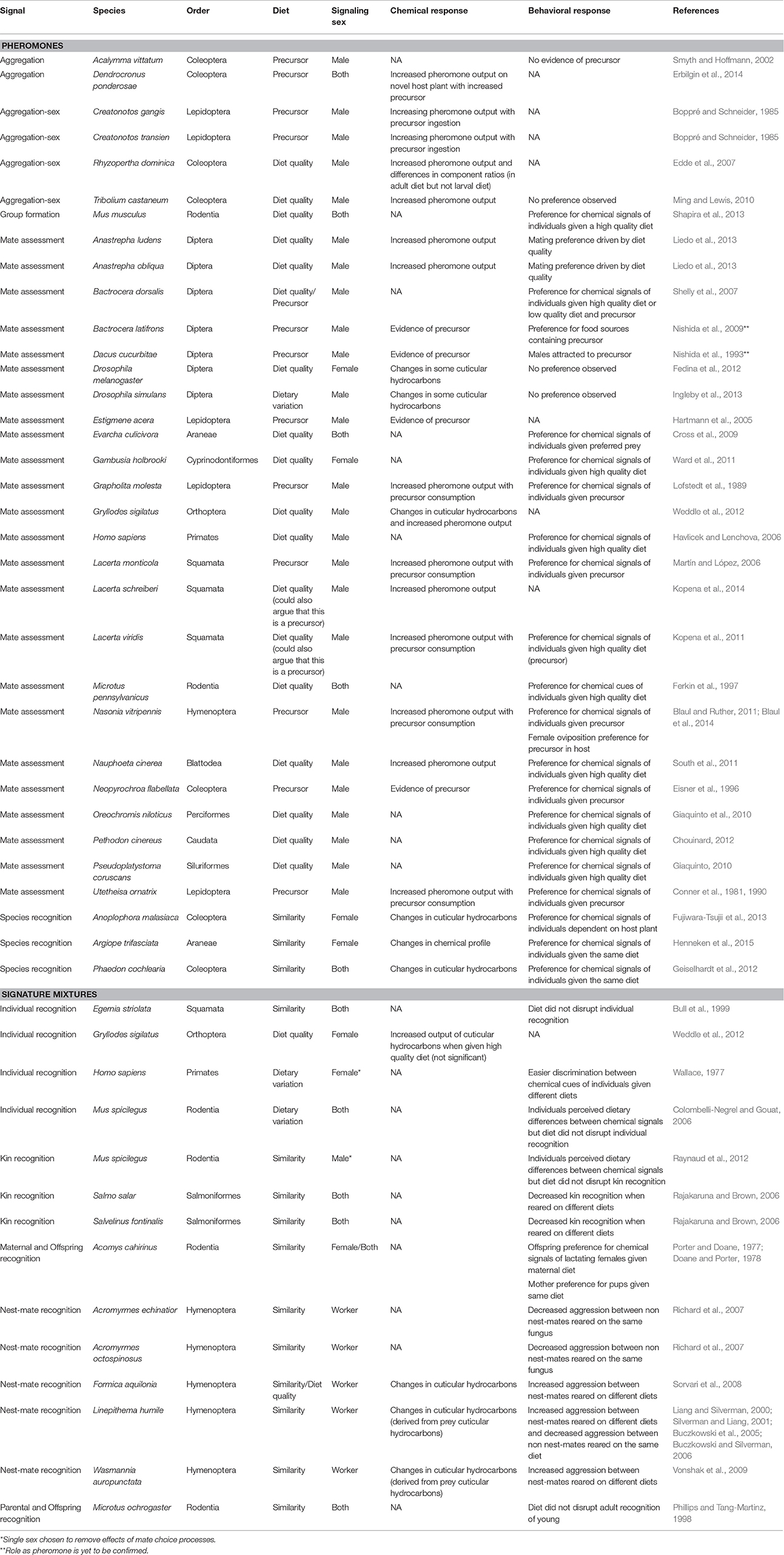
Table 1. Chemical and behavioral responses to dietary influences on pheromones and signature mixtures.
In this review, we examine the degree to which diet-mediated variation in pheromones and signature mixtures provides the receiver with both reliable and beneficial information. This information can be broadly classified into three types: information about the quality of the signaller; information about the quality of a resource provided by the signaller, and information on the identity of the signaller. In each case, dietary driven variation enforces signal reliability, allowing the receiver to respond advantageously. For example, a pheromone that relies on a high quality diet to be attractive to the receiver should inform the receivers about the nutritional state of the signaller (i.e., whether the signaller has been able to obtain adequate nutrition to produce an attractive pheromone). Ultimately, we argue that it is the receiver's ability to exploit this variation that allows for an evolutionarily stable non-static signal.
Dietary Influences on Pheromone Profiles and Signature Mixtures
There are three ways in which diet might contribute to quantitative and/or qualitative differences in a chemical signal. First, pheromone output might be influenced by diet quality and the accumulation of specific macro (South et al., 2011) or micro (Kopena et al., 2014) nutrients. In these cases, individuals consuming a high quality diet typically increase pheromone output, leading to a more conspicuous, and/or attractive signal (Liedo et al., 2013). Second, components of the pheromone that cannot be synthesized entirely de novo might be sequestered from dietary sources and then transformed, or directly incorporated into the pheromone (Blaul et al., 2014). These components are referred to as pheromone precursors, and the production of an attractive pheromone is mostly, if not entirely, dietary dependent (i.e., without dietary sequestration of the pheromone precursor, pheromone production is limited, or unavailable; Eisner and Meinwald, 2003). Finally, diet may influence the profile of the pheromone or signature mixture more directly, determining the individual chemical components present and their relative ratios, which may have consequences for recognition systems that rely on phenotype matching, or self-referral mechanisms (Buczkowski and Silverman, 2006).
Diet and Signal Reliability
If diet-mediated variation in pheromones and signature mixtures provides reliable (or honest) information about the signaller, we should be able to link this information back to the signaller's diet. In some cases, this is straightforward because specific signal components may be sequestered from dietary sources. In other cases, diet-mediated variation in the signal may provide indirect information about the signaller's genotype, such as the genes responsible for an individual's foraging ability (Figure 1). Characterizing the link between an individual's diet and their pheromone or signature mixture is key to understanding how diet-mediated variation in the chemical signal benefits the receiver.
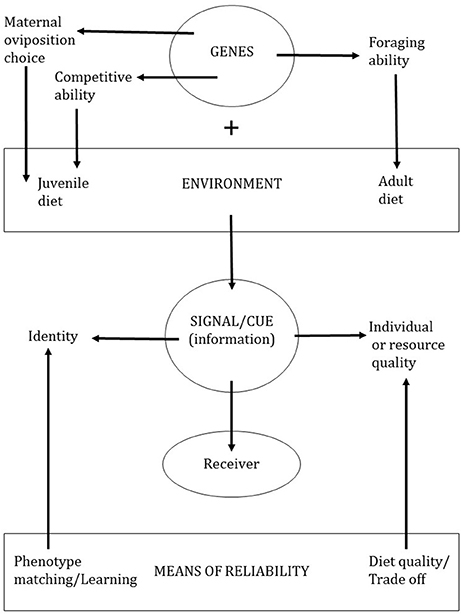
Figure 1. An animal's genotype and diet interact to produce a reliable chemical signal for a receiver.
Linking Diet and Individual Quality
Pheromones involved in the process of mate choice or assessment (Johansson and Jones, 2007) provide the receiver with information about the quality of the individual as a reproductive partner. Numerous studies demonstrate a role of diet quality for mate assessment pheromones across a wide range of taxa, including insects (Shelly et al., 2007; South et al., 2011; Fedina et al., 2012; Weddle et al., 2012; Liedo et al., 2013); mammals (Ferkin et al., 1997; Havlicek and Lenchova, 2006); reptiles (Kopena et al., 2011, 2014; Chouinard, 2012); arachnids (Cross et al., 2009; Baruffaldi and Andrade, 2015); and fish (Giaquinto, 2010; Giaquinto et al., 2010; Hayward and Gillooly, 2011; Table 1). Nonetheless, comprehensive studies that include both behavioral and chemical assays are largely limited to insect systems. For example, studies on mammals are typically behavioral, and although this is consistent with other pheromone research, future studies should consider combining behavioral assays with chemical analyses in these groups (Peso et al., 2015).
Laboratory studies investigating the link between diet and the qualities of the signaller typically provide the signaller with a diet that is either abundant or lacking in one or more macronutrients (i.e., proteins, carbohydrates, or lipids), and they then examine how this affects the attractiveness of the signaller (through behavioral assays) and/or pheromone output (through chemical analyses). Typically, such studies reveal an increase in total pheromone output or attractiveness in individuals given a high quality diet, but find no change to the relative proportions of chemical components present in the pheromone. For example, male cockroaches, Nauphoeta cinerea, provided with a diet rich in carbohydrates were more attractive to females and had greater pheromone output for three active pheromonal components, but there were no significant difference in the relative proportion in each of these components (South et al., 2011). Such diet-mediated chemical cues may provide the receiver with indirect information about the signaller's genotype or direct information about the signaller's reproductive potential through biological pathways and trade-offs.
Diet-Mediated Pheromones and Signaller Quality via Indirect Information
Pheromones are ultimately influenced by a combination of environmental and genetic factors. Diet is one the most significant environmental factors shaping a pheromone profile. The interaction between diet and the pheromone profile can be bi-directional, as an individual's genotype may influence its environment (and thus diet). For example, the quality of an individual's diet likely depends on their foraging ability, which can be a highly heritable trait (Karino et al., 2005; Missoweit et al., 2007). Pheromones that are determined by diet quality ensure that only individuals capable of securing the requisite quantity of appropriate nutrients can produce the attractive signal necessary for reproductive success. Further, if foraging ability is a heritable trait, then receivers that select partners based on diet-mediated signals may gain indirect information on the signaller's potential genetic contribution (through foraging) to future offspring.
The links between diet, pheromone and fitness are not always simple to elucidate. In some systems, the pheromone profile of adults is predominantly determined in the juvenile stages. For example, females of the parasitoid wasp Nasonia vitripennis prefer males that produce large quantities of a pheromone that requires the precursor linoleic acid for its production. Males consume linoleic acid during larval development within their animal host, or it can be synthesized from oleic acid, which is also ingested from hosts during this life stage (Blaul et al., 2014). It is unlikely that these pheromones provide information on the foraging ability of the males, because the dietary environment of the larvae depends entirely on the oviposition choices of their mother. Nevertheless, females prefer to lay their eggs in hosts with high linoleic acid content, which provides offspring with a source of this precursor and thereby increases their attractiveness to mates and ultimately their reproductive success (Blaul and Ruther, 2011). In this species, the male-derived pheromone provides receiving females with information about the oviposition choices of the male's mother, a trait that is likely heritable and determines the reproductive success of the receiving female's sons and daughters.
Diet-Mediated Pheromones and Signaller Quality via Direct Information
Signaling theory (Searcy and Nowicki, 2010) predicts that if the signal has significant costs, then it may provide the receiver with a reliable indication of individual quality, because the investment trade-off between the signal and other biological functions will keep the signal reliable. The specific costs of pheromone production have rarely been explicitly identified (Johansson et al., 2005; Foster and Anderson, 2015; Umbers et al., 2015), perhaps reflecting a widely held assumption that pheromone production is cheap (Alberts, 1992) and the difficulties of measuring the amount of pheromone released in model species such as moths (Umbers et al., 2015). Nonetheless, pheromone production could have significant physiological costs if its profile depends on nutrients that have other important biological functions. For example, vitamin E is an important antioxidant in vertebrates, but in some lizard species it is also linked with enhanced pheromone production and subsequent attractiveness (Kopena et al., 2011, 2014). Kopena et al. (2011) not only demonstrated that dietary vitamin E is incorporated in the femoral secretions of male European green lizards, Lacerta viridis, but that females prefer the secretions of males given a vitamin E supplement (Kopena et al., 2011). The vitamin E content of the pheromone is likely an honest indication of male quality. Only individuals of high quality can obtain sufficient vitamin E to allocate the micronutrient for pheromone production without experiencing the potential physiological costs.
More generally, a nutrient and energy-rich diet might simply provide more resources that can be diverted to a range of physiological processes, including pheromone production, without compromising vital biological pathways. Under such circumstances, well-fed individuals may be more capable of meeting the costs of pheromone production. Although some aspects of an individual's diet may reveal its genotype (such as their foraging ability), the capacity to produce particular pheromones will depend largely on the availability of resources. Several studies have demonstrated mating preference for chemical signals produced by well-fed individuals compared with their starved counterparts (Fisher and Rosenthal, 2006; Giaquinto, 2010; Baruffaldi and Andrade, 2015). Additionally, under-nourished males may have low sperm counts, poor sperm quality (Dunn and Moss, 1992; Izquierdo et al., 2001; Guan et al., 2014), or lower resistance to disease (Moret and Schmid-Hempel, 2000). Pheromones revealing information about male nutritional health may also be significant for females: recent studies link paternal nutritional health at conception to offspring health (Veenendaal et al., 2013) and fertility (McPherson et al., 2014), effects that may be trans-generational (see also Lane et al., 2015).
Linking Diet and Resource Quality
Some pheromones provide information about a potential resource provided by the signaller and, in turn, the resource may indicate individual quality as a reproductive partner. Nonetheless, it is not always clear whether the receiver is assessing individual or resource quality. Examples of these pheromones have not been widely demonstrated, and appear to occur only in a small number of insect genera.
Diet-Mediated Pheromones and Defensive Resources
Males of several species of Lepidoptera, including the butterflies Danaus chrysippus (Schneider et al., 1975), D. gilippus (Dussourd et al., 1989), Idea leuconoe (Nishida et al., 1996), and the moth Utetheisa ornatrix (Conner et al., 1981, 1990), provide females with a defensive chemical during courtship. Males sequester a pyrrolizidine alkaloid from their host plant as an adult or during the larval stage, some of which is stored in somatic tissue, likely as a form of chemical defense, and some is chemically modified to the male sex pheromone (Schneider et al., 1975; Conner et al., 1981). Females typically prefer males that produce large quantities of this pheromone, and males reared without access to the alkaloid are rendered less attractive or unattractive to females. The amount of pheromone produced by the male is a reliable signal of the amount of alkaloid he has accumulated and stored (see also Nishida, 2002). Females benefit from mating with males that produce high amounts of the alkaloid, because the male then transfers some of the stored alkaloid to the female during mating, and this is incorporated into her eggs protecting them from predation (see also Eisner and Meinwald, 2003). Additionally, male Cosmosoma myrodora moths cover female mates with a sequestered alkaloid during coupling, providing both partners with protection from predators, such as spiders, during lengthy copulations (Conner et al., 2000). The role of this alkaloid as a pheromone is unclear, but like other species where a pheromonal role of the alkaloid has been established (see above examples), C. myrodora females receive male-sequestered alkaloids via semen and transfer some of the alkaloid to their eggs for protection.
Interestingly, males of other Lepidoptera, including Phragmotobia fuliginosa and Pyrrhoraitia isabella, can sequester alkaloids as precursors for pheromone production, but this is not obligatory as they can mate successfully without access to this chemical (Krasnoff and Roelofs, 1989). One explanation for this unexpected result is that females of these species employ cryptic mate choice and can select against sperm from “alkaloid-limited” males after mating occurs. Cryptic mate choice is thought to occur in another alkaloid sequestering species, U. ornatrix (LaMunyon and Eisner, 1993) but, this strategy has not been tested in P. fuliginosa or P. isabella.
The importance of the sequestered alkaloid for these species is illustrated in laboratory studies of U. ornatrix, demonstrating that “alkaloid-limited” caterpillars resort to cannibalism in order to increase their alkaloid supply (Bogner and Eisner, 1991, 1992). Surprisingly, these caterpillars do not discriminate between kin and non-kin when engaging in cannibalistic behaviors (Hare and Eisner, 1995).
Diet-Mediated Pheromones and Host Plant Resources
Aggregation pheromones are often produced by one or both sexes and have evolved to attract conspecifics to a resource. For bark beetles (Curculionidae), an economically crucial pest, the resource is a host plant, and the aggregation of conspecifics enables successful colonization of the host and subsequent brood formation. Additionally, many species produce an anti-aggregation pheromone to arrest recruitment once the host tree has reached capacity. In the genera Dendroctonus and Ips (Renwick et al., 1976), beetles consume the monoterpene α-pinene from the bark of their host plant which is then converted into several related pheromonal components (specifically verbenol, verbenone or verbenene), that act as aggregation or anti-aggregation pheromones (Blomquist et al., 2010). As monoterpene constituents vary with host plant species, pheromone production can vary between beetle populations (Erbilgin et al., 2014). Further, as pheromone production relies on monoterpenes derived from the host tree, a highly attractive pheromone signal is limited to those individuals feeding on a suitable host. Receivers thus obtain direct information about the identity of the host from the signal produced. For example, in D. ponderosae, a pioneer female begins tree colonization by attacking a suitable host. She then transforms host derived α-pinene into the aggregation pheromone (-)-trans-verbenol, which attracts conspecifics of both sexes for host colonization and mating (Taft et al., 2015). Recruited males produce an additional aggregation pheromone (exo-brevicomin) via de novo synthesis. Lastly, once the host tree has reached maximum beetle capacity, two anti-aggregation pheromones are produced by the brood. Both sexes produce (-)-verbenone from host derived α-pinene via microbial transformation and males alone produce the second anti-aggregation pheromone, frontalin (Taft et al., 2015).
A key challenge is to determine whether the chemical compounds contributing to the pheromone are synthesized or sequestered from dietary sources (Blomquist et al., 2010). In some species, components are obtained directly from the host, but this is not the rule (Smyth and Hoffmann, 2002). Indeed, many insects produce pheromones containing components that are similar to those present within their host plants but they are synthesized by the insect entirely de novo (Miller et al., 1976; Seybold and Tittiger, 2003). The chemical concordance of pheromone components with host plants may have evolved to take advantage of pre-existing receptor cells of the receiver, which are already sensitive to these compounds as food or oviposition sources (see also Landolt and Phillips, 1997).
Linking Diet and Identity
Chemical communication that provides information about the membership of individuals to particular classes utilizes chemical profiles that are learnt or rely on self-referral mechanisms such as phenotype matching (Wyatt, 2014). These chemical signals are predicted to be more robust to environmental influences, since deviations would create a mismatch between the phenotype of individuals in the same class, or require individuals to relearn the new signal. Nevertheless, some degree of diet-mediated chemical signals are evident in these communication systems (Liang and Silverman, 2000). Where this type of variation occurs, individuals within a group recognize one another because they share the same diet and thus chemical profile, or they have learnt the chemical profile of others. Diet manipulation results in shifts in the signature mixture or pheromone, often resulting in receivers failing to recognize and/or respond appropriately to the new chemical profile (Porter and Doane, 1977).
Diet-Mediated Pheromones and Species Recognition
Being able to distinguish a mate of the right species is a crucial component of reproductive success in dioecious species. Species recognition pheromones are used by many animals to distinguish between conspecific and heterospecific individuals. Here we use the term pheromone, rather than signature mixture, as it appears unlikely that these chemical signals are learnt. However, it is important to note that whether these responses are predisposed or require learning is not known (Verzijden et al., 2012). Pheromones that reveal species identity should be relatively consistent between individuals of the same species, although populations sometimes differ in their pheromone profiles (Fornasiero et al., 2011). Despite being rarely explored beyond the theoretical argument that these pheromones are expected to be consistent between all individuals within a species, these data suggest that an environmental factor, such as diet, may have a strong influence on the profile of these pheromones. This may lead to reproductive isolation between populations, if receivers do not recognize (or respond to) all pheromone profiles. Empirical studies in this area are currently lacking and further research is clearly required.
The chemical composition of insect pheromones is often closely associated with their host plant, which may facilitate the diversification of insect species (Landolt and Phillips, 1997; Reddy and Guerrero, 2004; Nishida, 2014), because changes in pheromone signals that are a consequence of feeding on novel host plants may introduce reproductive isolation and speciation via assortative mating (Smadja and Butlin, 2009). For example, insect cuticular hydrocarbons are often employed to signal species (or nest-mate in the case of social insects) identity, and the cuticular hydrocarbon profiles of both male and female mustard leaf beetles Phaedon cochleariae change if they feed on a new host plant (Geiselhardt et al., 2012). Males have a strong mating preference for females given the same diet treatment, and the cuticular hydrocarbon profile can change within 2 weeks of the diet manipulation (Geiselhardt et al., 2012). Similar patterns have been documented in the white-spotted longicorn beetle, Anoplophora malasiaca (Fujiwara-Tsujii et al., 2013) and the orb-weaving spider Argiope trifasciata (Henneken et al., 2015). A diet-mediated pheromone can provide receivers with reliable information on species identity if diet is relatively consistent within the species (or between conspecifics within the same population). A diet-mediated signaling system may be more likely in species in which the majority of individuals within the communicating pool have a similar diet—with all individuals either monophagous or exploiting a similarly catholic diet. Further, it may also be favored in species that do not migrate or disperse widely, thereby constraining an individual's diet.
Diet-Mediated Signature Mixtures and Familial Recognition
In some species signature mixtures are used to recognize family members, such as offspring, parents and kin. Kin odor-based recognition, in particular, are thought to facilitate inbreeding avoidance or altruistic behavior (see Gadagkar, 1985), while maternal and offspring odor-based recognition are more likely to evolve in altricial and/or colonial species.
The role of signature mixtures in familial recognition is somewhat controversial. While chemical cues are undoubtedly employed in familial recognition, it is unclear if these are cues or signals (sensu Maynard Smith and Harper, 2003). Distinguishing between cues and signals is important because it determines the nature of the selection pressures acting on the signaller: signals evolve specifically to transfer information, and thus typically benefit both signaller and receiver (dishonest chemical signals which are detrimental to the receiver are one exception, for example chemical mimicry used in predator-prey interactions; Gemeno et al., 2000; Byers, 2015), while cues provide information to the receiver but have not evolved to benefit the signaller (Lehmann et al., 2014). For example, diet-related variation in chemical cues emanating from predators can be used by prey as cues revealing the presence of particular predators (Sharp et al., 2015). In this case, there has been selection on the receiver to distinguish between these cues, but clearly not on the signaller to provide such information. In contrast, diet-mediated chemical cues may contribute to the formation of social groups (Kleinhappel et al., 2014), in which case the nature of the chemical cue, as signal or cue, is ambiguous if signallers and receivers benefit from this behavior. Whether the following examples of diet-mediated signature mixtures would be better classified as diet-mediated signals or cues requires further investigation. Nonetheless, these studies demonstrate the potential consequences of employing a diet-mediated signal (or cue) in a recognition system.
In two landmark studies, Porter and Doane (1977), Doane and Porter (1978) manipulated the diet of captive female mice, Acomys cahinnus, and demonstrated that maternal and offspring recognition odors were strongly driven by diet (Porter and Doane, 1977; Doane and Porter, 1978), a mechanism that was also reported in the laboratory rat, Rattus norvegious (Leon, 1975). In these rodents, maternal recognition odors emanate from feces and variation in the maternal odor may reflect diet-mediated differences in bacterial flora (Brown and Schellinck, 1992). These, and other studies (e.g., Skeen and Thiessen, 1977; Galef, 1981), provide strong evidence of the role of diet in shaping maternal and offspring signature mixtures, a pattern likely to be widespread among rodents. The prairie vole, Microtus ochrogaster, may be an exception to this pattern. Phillips and Tang-Martinz (1998) demonstrated a role for odor in parent-offspring recognition in M. ochrogaster, but there was no significant effect of diet on their recognition behaviors (Phillips and Tang-Martinz, 1998). The authors report a non-significant trend toward a dietary-mediated odor (less aggression between unfamiliar individuals on the same diet). However, in contrast with previous studies, they did not consider the recognition behavior of related individuals on different diets, which may have been more informative. Logic predicts that diet-mediated maternal and offspring recognition odors would only provide receivers with reliable information about identity if an individual's diet is consistent. However, this is yet to be explicitly tested and it is also probable that under natural conditions, mothers and offspring recognize dietary driven changes in each other's odors by their near continuous exposure to one another.
More broadly, diet-mediated kin recognition has been investigated across a range of taxa (Table 1). Rajakaruna and Brown (2006) observed a decline in kin discrimination in two species of fish, Salmo salar and Salvelinus fontinalis, when individuals were presented with non-kin reared on the same diet (as the focal fish) and kin reared on a different diet (Rajakaruna and Brown, 2006). Further, fish of both species preferred the chemical cues of non-kin reared on the same diet over non-kin reared on a different diet. Similar patterns in kin recognition have been observed in toad (Scaphiopus spp.) larvae (Pfennig, 1990; Hall et al., 1995). Whilst it is likely that juvenile nest mates share the same diet, adult siblings that often disperse are expected to have less similar diets. Accordingly, these early recognition behaviors in amphibians may be driven by preference for familiar food or habitat cues rather than kin recognition odors (Pfennig, 1990). The patterns for the fish may even be driven by the oddity effect, where shoaling with like individuals reduces the risk of predation (Kleinhappel et al., 2014).
Diet-Mediated Signature Mixtures and Nest-Mate Recognition
Social insects rely on chemical signals to communicate important information such as colony identity, tasks, and status. The integrity of social insect colonies depends crucially on the capacity of individuals to distinguish between members of their own colony and other individuals (either conspecifics or otherwise). Social insects, including bees, wasps, ants, and termites, use cuticular hydrocarbons as signals to identify colony or nest identity (e.g., van Wilgenburg et al., 2011; Kather and Martin, 2015). The mechanism that allows individuals to recognize the identity of others remains unclear, but likely involves a self-referral mechanism. In the little fire ant, Wasmannia auropunctata, a species that forms vast super-colonies, worker ants are rarely aggressive to conspecific workers from different colonies. Workers recently collected from the field, however, react aggressively to workers that had been maintained in the laboratory for a 4 month period (Vonshak et al., 2009). Analysis of the cuticular hydrocarbons revealed differences between these two groups of ants that could be directly traced back to the laboratory diet (Vonshak et al., 2009). Similar observations have been recorded in other ants, including Formica aquilonia (Sorvari et al., 2008) and Linepithema humile (Liang and Silverman, 2000). In some of these systems, diet is so crucial to the nest-mate recognition system that the converse pattern also occurs: previously hostile colonies experience decreased aggression when reared on the same diet (Buczkowski et al., 2005; Richard et al., 2007). Intriguingly, the cuticular hydrocarbon profile of the myrmecophile spider Cosmophasis bitaeniata, which feeds on the larval stages of the ant Oecophylla smaragdina, changes with diet (Elgar and Allan, 2004), presumably allowing the spider to mimic the colony profile and thus remain undetected by its prey (Elgar and Allan, 2006).
Diet-mediated nest-mate recognition odors provide receivers with reliable information of identity and are most effective when diet varies between, but is consistent within, colonies. It is interesting to note that the response behavior between ants is not binary—in some cases, different colonies reared on the same diet were able to maintain their aggression toward each other, suggesting that dietary influences do not entirely “conceal” the signal (Buczkowski et al., 2005).
Conclusion and Future Directions
Diet-mediated variation in pheromones or signature mixtures provides an ideal opportunity to study signal reliability, as in many cases we can directly link diet with the information provided by the signal. Not only is diet relatively easy to manipulate in laboratory environments, it can also be manipulated to a finer scale, not readily available for other environmental influences. We can limit or completely withhold specific macro- or micro-nutrients, provide desirable nutrients ad libitum and even manipulate the energy content of the diet. Further, chemical analyses of the signal and an understanding of the biological pathways involved may allow us to consider the cost/benefit trade-offs underlying the evolutionary maintenance of the signal.
Consistent with much of the research on chemical communication, studies on diet-mediated variation in pheromones and signature mixtures outside of insect taxa are lacking. Although, there is behavioral evidence of diet-mediated variation in odors in mammals (e.g., Doane and Porter, 1978; Ferkin et al., 1997), including humans (Wallace, 1977; Havlicek and Lenchova, 2006), chemical analysis is surprisingly rarely attempted. Studies on chemical communication in mammals typically employ sweat, urine or feces in their behavioral assays, and characterization of the signal components of these secretions are required to confirm these dietary influences. Nevertheless, several studies on lizards have successfully demonstrated dietary driven variation in the chemical signal, consistent with results from behavioral assays (Martín and López, 2006; Kopena et al., 2011, 2014).
Despite the numerous studies demonstrating a role for dietary influences on mate assessment pheromones, caution is still required when interpreting these studies. This is particularly true for chemicals that are suspected pheromone precursors, which are then directly incorporated into the pheromone. Similar components from host plants may evolve as pheromonal components that take advantage of pre-existing receptors (Landolt and Phillips, 1997). To demonstrate unequivocally that a chemical is a pheromone precursor requires that individuals are reared on diets completely devoid of the suspected precursor and, ideally, this should be coupled with chemical analysis of the pheromone to confirm the complete absence of the pheromonal component.
Empirical demonstrations of an explicit link between a diet-mediated pheromone and the signaller's genotype are also rarely published. Thus, in addition to demonstrating the influence of diet on pheromone production, future studies should also consider how these diets are influenced by individual foraging abilities or maternal oviposition choices. There may also be suitable species for heritability studies, given the breadth of taxa for which diet-mediated mate assessment pheromones have been demonstrated (Table 1).
In some communication systems involving recognition of types of individuals, the consequence of diet-mediated variation in chemical signals may cause individuals to fail to recognize and/or respond correctly to the signal (Porter and Doane, 1977; Sorvari et al., 2008). It is important to note that this result emerged, in many of these studies, in response to radical changes in diet and that such extreme dietary variation rarely occurs in nature. Nevertheless, individuals are likely to experience different diets, and so we expect receivers to be able to accommodate some concomitant variation in the signal. For example, the receptor cells of receivers may not be sensitive enough to detect very small deviations to a chemical signal or selection may not act on the receiver's ability to perceive these small deviations. Additionally, some chemical components within a pheromone or signature mixture may be more robust to dietary variation (Ingleby et al., 2013). Future studies could investigate this by feeding individuals diets with varying degrees of dissimilarity and then assessing the reliability of the chemical signal in behavioral assays.
Finally, care should be taken with studies involving captive animals, especially where there is a dramatic change in diet, or even where all individuals are provided with a similar diet. Both instances may have consequences for communication systems involving chemical signaling including pheromone-based mate choice or odor-based recognition. Given the compelling evidence of a role for diet in shaping these chemical signals, both behavioral and chemical studies should take into account potential dietary effects on the signal's profile.
Author Contributions
JH was the primary author, wrote the first draft of the document, contributed to the concept and design of the manuscript and was the primary editor of the final draft. JG, TJ, and ME contributed to the concept/design of the manuscript, were involved in editing drafts and the final document and contributed knowledge.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The Australian Research Council (DP0987360 and DP120100162 to ME), Holsworth Wildlife Research Endowment managed by ANZ trustees, Jasper Loftus- Hill Memorial Fund, and Australian Government Research Training Program Scholarship (to JH) support our research.
References
Alberts, A. C. (1992). Constraints on the design of chemical communication systems in terrestrial veterbrates. Am. Nat. 139, S62–S89.
Anderbrant, O., Schlyter, F., and Birgersson, G. (1985). Intraspecific competition affecting parents and offspring in the bark beetle Ips typographus. Oikos 45, 89–98. doi: 10.2307/3565226
Baruffaldi, L., and Andrade, M. C. B. (2015). Contact pheromones mediate male preference in black widow spiders: avoidance of hungry sexual cannibals? Anim. Behav. 102, 25–32. doi: 10.1016/j.anbehav.2015.01.007
Blaul, B., and Ruther, J. (2011). How parasitoid females produce sexy sons: a causal link between oviposition preference, dietary lipids and mate choice in Nasonia. Proc. R. Soc. B Biol. Sci. 278, 3286–3293. doi: 10.1098/rspb.2011.0001
Blaul, B., Steinbauer, R., Merkl, P., Merkl, R., Tschochner, H., and Ruther, J. (2014). Oleic acid is a precursor of linoleic acid and the male sex pheromone in Nasonia vitripennis. Insect Biochem. Mol. Biol. 51, 33–40. doi: 10.1016/j.ibmb.2014.05.007
Blomquist, G. J., Figueroa-Teran, R., Aw, M., Song, M. M., Gorzalski, A., Abbott, N. L., et al. (2010). Pheromone production in bark beetles. Insect Biochem. Mol. Biol. 40, 699–712. doi: 10.1016/j.ibmb.2010.07.013
Bogner, F., and Eisner, T. (1991). Chemical basis of egg cannibalism in a caterpillar (Utetheisa ornatrix). J. Chem. Ecol. 17, 2063–2075. doi: 10.1007/BF00987992
Bogner, F., and Eisner, T. (1992). Chemical basis of pupal cannibalism in a caterpillar (Utetheisa ornatrix). Experientia 48, 97–102. doi: 10.1007/BF01923618
Boppré, M., and Schneider, D. (1985). Pyrrolizidine alkaloids quantitatively regulate both scent organ morphogenesis and pheromone biosynthesis in male Creatonotos moths (Lepidoptera: Arctiidae). J. Comp. Physiol. A 157:569.
Brown, R. E., and Schellinck, H. M. (1992). “Interactions among the MHC, Diet and bacteria in the production of social odors in rodents,” in Chemical Signals in Vertebrates 6, eds R. Dotyand and D. Müller-Schwarze (New York, NY: Springer), 175–181.
Buczkowski, G., Kumar, R., Suib, S. L., and Silverman, J. (2005). Diet-related modification of cuticular hydrocarbon profiles of the Argentine ant, Linepithema humile, diminishes intercolony aggression. J. Chem. Ecol. 31, 829–843. doi: 10.1007/s10886-005-3547-7
Buczkowski, G., and Silverman, J. (2006). Geographical variation in Argentine ant aggression behaviour mediated by environmentally derived nestmate recognition cues. Anim. Behav. 71, 327–335. doi: 10.1016/j.anbehav.2005.04.012
Bull, M. C., Griffin, L. C., and Perkins, V. M. (1999). Some properties of a pheromone allowing individual recognition, from the scats of an Australian lizard, Egernia striolata. Acta Ethol. 2, 35–42. doi: 10.1007/PL00012230
Byers, J. A. (2015). Earwigs (Labidura riparia) mimic rotting-flesh odor to deceive vertebrate predators. Sci. Nat. 102, 1–10. doi: 10.1007/s00114-015-1288-1
Chouinard, A. (2012). Rapid onset of mate quality assessment via chemical signals in a woodland salamander (Plethodon cinereus). Behav. Ecol. Sociobiol. 66, 765–775. doi: 10.1007/s00265-012-1324-5
Colombelli-Negrel, D., and Gouat, P. (2006). Male and female mound-building mice, Mus spicilegus, discriminate dietary and individual odours of conspecifics. Anim. Behav. 72, 577–583. doi: 10.1016/j.anbehav.2005.11.015
Conner, W. E., Boada, R., Schroeder, F. C., González, A., Meinwald, J., and Eisner, T. (2000). Chemical defense: bestowal of a nuptial alkaloidal garment by a male moth on its mate. Proc. Natl. Acad. Sci. U.S.A. 97, 14406–14411. doi: 10.1073/pnas.260503797
Conner, W. E., Eisner, T., Robert, K. V. M., Guerrero, A., and Meinwald, J. (1981). Precopulatory sexual interaction in an arctiid moth (Utetheisa ornatrix): role of a pheromone derived from dietary alkaloids. Behav. Ecol. Sociobiol. 9, 227–235. doi: 10.1007/BF00302942
Conner, W. E., Roach, B., Benedict, E., Meinwald, J., and Eisner, T. (1990). Courtship pheromone production and body size as correlates of larval diet in males of the arctiid moth, Utetheisa ornatrix. J. Chem. Ecol. 16, 543–552. doi: 10.1007/BF01021785
Cross, F. R., Jackson, R. R., and PoIiard, S. D. (2009). How blood-derived odor influences mate-choice decisions by a mosquito-eating predator. Proc. Natl. Acad. Sci. U.S.A. 106, 19416–19419. doi: 10.1073/pnas.0904125106
Doane, H. M., and Porter, R. H. (1978). The role of diet in mother-infant reciprocity in the spiny mouse. Dev. Psychobiol. 11, 271–277. doi: 10.1002/dev.420110311
Dunn, T. G., and Moss, G. E. (1992). Effects of nutrient deficiencies and excesses on reproductive efficiency of livestock. J. Anim. Sci. 70, 1580–1593. doi: 10.2527/1992.7051580x
Dussourd, D. E., Harvis, C. A., Meinwald, J., and Eisner, T. (1989). Paternal allocation of sequestered plant pyrrolizidine alkaloid to eggs in the danaine butterfly, Danaus gilippus. Experientia 45, 896–898. doi: 10.1007/BF01954068
Edde, P. A., Phillips, T. W., Robertson, J. B., and Dillwith, J. W. (2007). Pheromone output by Rhyzopertha dominica (Coleoptera: Bostrichidae), as affected by host plant and beetle size. Ann. Entomol. Soc. Am. 100, 83–90. doi: 10.1603/0013-8746(2007)100[83:POBRDC]2.0.CO;2
Eisner, T., and Meinwald, J. (2003). “Alkaloid-derived pheromones and sexual selection in Lepidoptera,” in Insect Pheromone Biochemistry and Molecular Biology, eds G. Blomquist and R. Vogt (Amsterdam; Boston, MA: Elsevier/Academic Press), 341–368. doi: 10.1016/B978-012107151-6/50014-1
Eisner, T., Smedley, S. R., Young, D. K., Eisner, M., Roach, B., and Meinwald, J. (1996). Chemical basis of courtship in a beetle (Neopyrochroa flabellata): cantharidin as precopulatory “enticing” agent. Proc. Natl. Acad. Sci. U.S.A. 93, 6494–6498. doi: 10.1073/pnas.93.13.6494
Elgar, M. A., and Allan, R. A. (2004). Spider chemical mimics acquire colony-specific cuticular hydrocarbons from their ant model prey. Naturwissenschaften 91, 143–147. doi: 10.1007/s00114-004-0507-y
Elgar, M. A., and Allan, R. A. (2006). Colony specific mimicry of the weaver ant Oecophylla smaragdina by the myrmecophilous salticid spider Cosmophasis bitaeniata. J. Ethol. 24, 239–246. doi: 10.1007/s10164-005-0188-9
Erbilgin, N., Ma, C., Whitehouse, C., Shan, B., Najar, A., and Evenden, M. (2014). Chemical similarity between historical and novel host plants promotes range and host expansion of the mountain pine beetle in a naïve host ecosystem. New Phytol. 201, 940–950. doi: 10.1111/nph.12573
Fedina, T. Y., Kuo, T.-H., Dreisewerd, K., Dierick, H. A., Yew, J. Y., and Pletcher, S. D. (2012). Dietary effects on cuticular hydrocarbons and sexual attractiveness in Drosophila. PLoS ONE 7:e49799. doi: 10.1371/journal.pone.0049799
Ferkin, M. H., Sorokin, E. S., Johnston, R. E., and Lee, C. J. (1997). Attractiveness of scents varies with protein content of the diet in meadow voles. Anim. Behav. 53, 133–141. doi: 10.1006/anbe.1996.0284
Fisher, H. S., and Rosenthal, G. G. (2006). Female swordtail fish use chemical cues to select well-fed mates. Anim. Behav. 72, 721–725. doi: 10.1016/j.anbehav.2006.02.009
Fornasiero, S., Dendi, F., Bresciani, E., Cecchinelli, E., and Zuffi, M. A. L. (2011). The scent of the others: chemical recognition in two distinct populations of the European whip snake, Hierophis viridiflavus. Amphib. Reptil. 32, 39–47. doi: 10.1163/017353710X541850
Foster, S. P., and Anderson, K. G. (2015). Sex pheromones in mate assessment: analysis of nutrient cost of sex pheromone production by females of the moth Heliothis virescens. J. Exp. Biol. 218, 1252–1258. doi: 10.1242/jeb.119883
Fujiwara-Tsujii, N., Yasui, H., and Wakamura, S. (2013). Population differences in male responses to chemical mating cues in the white-spotted longicorn beetle, Anoplophora malasiaca. Chemoecology 23, 113–120. doi: 10.1007/s00049-013-0126-1
Gadagkar, R. (1985). Kin recognition in social insects and other animals—a review of recent findings and a consideration of their relevance for the theory of kin selection. Proc. Anim. Sci. 94, 587–621. doi: 10.1007/bf03191863
Galef, J. B. G. (1981). Preference for natural odors in rat pups: implications of a failure to replicate. Physiol. Behav. 26, 783–786. doi: 10.1016/0031-9384(81)90099-8
Geiselhardt, S., Otte, T., and Hilker, M. (2012). Looking for a similar partner: host plants shape mating preferences of herbivorous insects by altering their contact pheromones. Ecol. Lett. 15, 971–977. doi: 10.1111/j.1461-0248.2012.01816.x
Gemeno, C., Yeargan, K. V., and Haynes, K. F. (2000). Aggressive chemical mimicry by the bolas spider Mastophora hutchinsoni: identification and quantification of a major prey's sex pheromone components in the spider's volatile emissions. J. Chem. Ecol. 26, 1235–1243. doi: 10.1023/A:1005488128468
Giaquinto, P. C. (2010). Female pintado catfish choose well-fed males. Behaviour 147, 391–332. doi: 10.1163/000579509x12535339073761
Giaquinto, P. C., da Silva Berbert, C. M., and Delicio, H. C. (2010). Female preferences based on male nutritional chemical traits. Behav. Ecol. Sociobiol. 64, 1029–1035. doi: 10.1007/s00265-010-0918-z
Guan, Y., Malecki, I. A., Hawken, P. A. R., Linden, M. D., and Martin, G. B. (2014). Under-nutrition reduces spermatogenic efficiency and sperm velocity, and increases sperm DNA damage in sexually mature male sheep. Anim. Reprod. Sci. 149, 163–172. doi: 10.1016/j.anireprosci.2014.07.014
Hall, J. A., Larsen, J. H., Miller, D. E., and Fitzner, R. E. (1995). Discrimination of kin-based and diet-based cues by larval spadefoot toads, Scaphiopus intermontanus (Anura Pelobatidae), under laboratory conditions. J. Herpetol. 29, 233–243. doi: 10.2307/1564560
Hare, J. F., and Eisner, T. (1995). Cannibalistic caterpillars (Utetheisa ornatrix; Lepidoptera: Arctiidae) fail to differentiate between eggs on the basis of kinship. Psyche 102, 27–33. doi: 10.1155/1995/84147
Hartmann, T., Theuring, C., Beuerle, T., Klewer, N., Schulz, S., Singer, M. S., et al. (2005). Specific recognition, detoxification and metabolism of pyrrolizidine alkaloids by the polyphagous arctiid Estigmene acrea. Insect Biochem. Mol. Biol. 35, 391–411. doi: 10.1016/j.ibmb.2004.12.010
Havlicek, J., and Lenchova, P. (2006). The effect of meat consumption on body odor attractiveness. Chem. Senses 31, 747–752. doi: 10.1093/chemse/bjl017
Hayward, A., and Gillooly, J. F. (2011). The cost of sex: quantifying energetic investment in gamete production by males and females. PLoS ONE 6:e16557. doi: 10.1371/journal.pone.0016557
Henneken, J., Jones, T. M., Goodger, J. Q. D., Dias, D. A., Walter, A., and Elgar, M. A. (2015). Diet influences female signal reliability for male mate choice. Anim. Behav. 108, 215–221. doi: 10.1016/j.anbehav.2015.07.023
Hock, V., Chouinard, G., Lucas, É., Cormier, D., Leskey, T., Wright, S., et al. (2014). Establishing abiotic and biotic factors necessary for reliable male pheromone production and attraction to pheromones by female plum curculios Conotrachelus nenuphar (Coleoptera: Curculionidae). Can. Entomol. 146, 528–547. doi: 10.4039/tce.2014.1
Ingleby, F. C., Hunt, J., and Hosken, D. J. (2013). Heritability of male attractiveness persists despite evidence for unreliable sexual signals in Drosophila simulans. J. Evol. Biol. 26, 311–324. doi: 10.1111/jeb.12045
Izquierdo, M. S., Fernández-Palacios, H., and Tacon, A. G. J. (2001). Effect of broodstock nutrition on reproductive performance of fish. Aquaculture 197, 25–42. doi: 10.1016/S0044-8486(01)00581-6
Johansson, B. G., and Jones, T. M. (2007). The role of chemical communication in mate choice. Biol. Rev. 82, 265–289. doi: 10.1111/j.1469-185X.2007.00009.x
Johansson, B. G., Jones, T. M., and Widemo, F. (2005). Cost of pheromone production in a lekking Drosophila. Anim. Behav. 69, 851–858. doi: 10.1016/j.anbehav.2004.08.007
Karino, K., Utagawa, T., and Shinjo, S. (2005). Heritability of the algal-foraging ability: an indirect benefit of female mate preference for males' carotenoid-based coloration in the guppy, Poecilia reticulata. Behav. Ecol. Sociobiol. 59:1. doi: 10.1007/s00265-005-0001-3
Kather, R., and Martin, S. (2015). Evolution of cuticular hydrocarbons in the hymenoptera: a meta-analysis. J. Chem. Ecol. 41, 871–883. doi: 10.1007/s10886-015-0631-5
Kleinhappel, T. K., Burman, O. H. P., John, E. A., Wilkinson, A., and Pike, T. W. (2014). Diet-mediated social networks in shoaling fish. Behav. Ecol. 25, 374–377. doi: 10.1093/beheco/aru006
Kopena, R., Lopez, P., and Martin, J. (2014). Relative contribution of dietary carotenoids and vitamin E to visual and chemical sexual signals of male Iberian green lizards: an experimental test. Behav. Ecol. Sociobiol. 68, 571–581. doi: 10.1007/s00265-013-1672-9
Kopena, R., Martín, J., López, P., and Herczeg, G. (2011). Vitamin E supplementation increases the attractiveness of males' scent for female European green lizards. PLoS ONE 6:e19410. doi: 10.1371/journal.pone.0019410
Krasnoff, S. B., and Roelofs, W. L. (1989). Quantitative and qualitative effects of larval diet on male scent secretions of Estigmene acrea, Phragmatobia foliginosa, and Pyrrharctia isabella (Lepidoptera: Arctiidae). J. Chem. Ecol. 15, 1077–1093. doi: 10.1007/BF01015201
LaMunyon, C. W., and Eisner, T. (1993). Postcopulatory sexual selection in an arctiid moth (Utetheisa ornatrix). Proc. Natl. Acad. Sci. U.S.A. 90, 4689–4692. doi: 10.1073/pnas.90.10.4689
Landolt, P. J., and Phillips, T. W. (1997). Host plant influences on sex pheromone behavior of phytophagous insects. Annu. Rev. Entomol. 42, 371–391. doi: 10.1146/annurev.ento.42.1.371
Lane, M., Zander-Fox, D. L., Robker, R. L., and McPherson, N. O. (2015). Review: peri-conception parental obesity, reproductive health, and transgenerational impacts. Trends Endocrinol. Metab. 26, 84–90. doi: 10.1016/j.tem.2014.11.005
Lehmann, K. D. S., Goldman, B. W., Dworkin, I., Bryson, D. M., and Wagner, A. P. (2014). From cues to signals: evolution of interspecific communication via aposematism and mimicry in a predator-prey system. PLoS ONE 9:e91783. doi: 10.1371/journal.pone.0091783
Leon, M. (1975). Dietary control of maternal pheromone in the lactating rat. Physiol. Behav. 14, 311–319. doi: 10.1016/0031-9384(75)90039-6
Liang, D., and Silverman, J. (2000). “You are what you eat”: diet modifies cuticular hydrocarbons and nestmate recognition in the Argentine ant, Linepithema humile. Naturwissenschaften 87, 412–416. doi: 10.1007/s001140050752
Liedo, P., Orozco, D., Cruz-López, L., Quintero, J. L., Becerra-Pérez, C., del Refugio Hernández, M., et al. (2013). Effect of post-teneral diets on the performance of sterile Anastrepha ludens and Anastrepha obliqua fruit flies. J. Appl. Entomol. 137, 49–60. doi: 10.1111/j.1439-0418.2010.01568.x
Lofstedt, C., Vickers, N. J., Roelofs, W. L., and Baker, T. C. (1989). Diet related courtship success in the oriental fruit moth, Grapholita molesta (Tortricidae). Oikos 55, 402–408. doi: 10.2307/3565601
Martín, J., and López, P. (2006). Vitamin D supplementation increases the attractiveness of males' scent for female Iberian rock lizards. Proc. R. Soc. Lond. B Biol. Sci. 273, 2619–2624. doi: 10.1098/rspb.2006.3619
McPherson, N. O., Fullston, T., Bakos, H. W., Setchell, B. P., and Lane, M. (2014). Obese father's metabolic state, adiposity, and reproductive capacity indicate son's reproductive health. Fertil. Steril. 101, 865–873. doi: 10.1016/j.fertnstert.2013.12.007
Miller, J. R., Baker, T. C., Carde, R. T., and Roelofs, W. L. (1976). Reinvestigation of oak leaf roller sex pheromone components and the hypothesis that they vary with diet. Science 192, 140–143. doi: 10.1126/science.1257758
Ming, Q.-L., and Lewis, S. M. (2010). Pheromone production by male tribolium castaneum (Coleoptera: Tenebrionidae) is influenced by diet quality. J. Econ. Entomol. 103, 1915–1919. doi: 10.1603/EC10110
Missoweit, M., Engels, S., and Sauer, K. P. (2007). Foraging ability in the scorpionfly panorpa vulgaris: individual differences and heritability. Behav. Ecol. Sociobiol. 61, 487–492. doi: 10.1007/s00265-006-0277-y
Moret, Y., and Schmid-Hempel, P. (2000). Survival for immunity: the price of immune system activation for bumblebee workers. Am. Soc. Adv. Sci. 290, 1166–1168. doi: 10.1126/science.290.5494.1166
Nishida, R. (2002). Sequestration of defensive substances from plants by Lepidoptera. Annu. Rev. Entomol. 47, 57–92. doi: 10.1146/annurev.ento.47.091201.145121
Nishida, R. (2014). Chemical ecology of insect-plant interactions: ecological significance of plant secondary metabolites. Biosci. Biotechnol. Biochem. 78, 1–13. doi: 10.1080/09168451.2014.877836
Nishida, R., Enomoto, H., Shelly, T. E., and Ishida, T. (2009). Sequestration of 3-oxygenated α-ionone derivatives in the male rectal gland of the solanaceous fruit fly, Bactrocera latifrons. Entomol. Exp. Appl. 131, 85–92. doi: 10.1111/j.1570-7458.2009.00835.x
Nishida, R., Iwahashi, O., and Tan, K. H. (1993). Accumulation of Dendrobium superbum (orchidaceae) fragrance in the rectal glands by males of the melon fly, Dacus cucurbitae. J. Chem. Ecol. 19, 713–722. doi: 10.1007/BF00985003
Nishida, R., Schulz, S., Kim, C. S., Fukami, H., Kuwahara, Y., Honda, K., et al. (1996). Male sex pheromone of a giant danaine butterfly, Idea leuconoe. J. Chem. Ecol. 22, 949–972. doi: 10.1007/BF02029947
Ono, T. (1993). Effect of rearing temperature on pheromone component ratio in potato tuberworm moth, Phthorimaea operculella, (Lepidoptera: Gelechiidae). J. Chem. Ecol. 19, 71–81. doi: 10.1007/BF00987472
Palacio Cortés, A. M., Zarbin, P. H. G., Takiya, D. M., Bento, J. M. S., Guidolin, A. S., and Consoli Fernando, L. (2010). Geographic variation of sex pheromone and mitochondrial DNA in Diatraea saccharalis (Fab., 1794) (Lepidoptera: Crambidae). J. Insect Physiol. 56, 1624–1630. doi: 10.1016/j.jinsphys.2010.06.005
Peso, M., Elgar, M. A., and Barron, A. B. (2015). Pheromonal control: reconciling physiological mechanism with signalling theory. Biol. Rev. 90, 542–559. doi: 10.1111/brv.12123
Pfennig, D. W. (1990). ‘Kin recognition’ among spadefoot toad tadpoles: a side-effect of habitat selection? Evolution 44, 785–789.
Phillips, M. L., and Tang-Martinz, Z. (1998). Parent-offspring discrimination in the prairie vole and the effects of odors and diet. Can. J. Zool. 76, 711–716. doi: 10.1139/z97-242
Porter, R. H., and Doane, H. M. (1977). Dietary-dependent cross-species similarities in maternal chemical cues. Physiol. Behav. 19, 129–131. doi: 10.1016/0031-9384(77)90169-X
Rajakaruna, R. S., and Brown, J. A. (2006). Effect of dietary cues on kin discrimination of juvenile Atlantic salmon (Salmo salar) and brook trout (Salvelinus fontinalis). Can. J. Zool. 84, 839–845. doi: 10.1139/z06-069
Raynaud, J., Messaoudi, F., and Gouat, P. (2012). Reliability of odour–genes covariance despite diet changes: a test in mound-building mice. Biol. J. Linn. Soc. 106, 682–688. doi: 10.1111/j.1095-8312.2012.01888.x
Reddy, G. V. P., and Guerrero, A. (2004). Interactions of insect pheromones and plant semiochemicals. Trends Plant Sci. 9, 253–261. doi: 10.1016/j.tplants.2004.03.009
Renwick, J. A. A., Hughes, P. R., and Krull, I. S. (1976). Selective production of cis- and trans-verbenol from (-)-and (+)-α-pinene by a bark beetle. Science 191, 199–201. doi: 10.1126/science.1246609
Richard, F.-J., Poulsen, M., Hefetz, A., Errard, C., Nash, D. R., and Boomsma, J. J. (2007). The origin of the chemical profiles of fungal symbionts and their significance for nestmate recognition in Acromyrmex leaf-cutting ants. Behav. Ecol. Sociobiol. 61, 1637–1649. doi: 10.1007/s00265-007-0395-1
Schneider, D., Boppré, M., Schneider, H., Thompson, W., Boriack, C., Petty, R., et al. (1975). A pheromone precursor and its uptake in male Danaus butterflies. J. Comp. Physiol. 97, 245–256. doi: 10.1007/BF00617545
Searcy, W. A., and Nowicki, S. (2010). The Evolution of Animal Communication: Reliability and Deception in Signaling Systems. Princeton, NJ; Oxford: Princeton University Press.
Seybold, S. J., and Tittiger, C. (2003). Biochemistry and molecular biology of de novo isoprenoid pheromone production in the Scolytidae. Annu. Rev. Entomol. 48, 425–453. doi: 10.1146/annurev.ento.48.091801.112645
Shapira, I., Brunton, D., Shanas, U., and Raubenheimer, D. (2013). Conspecific attraction in invasive wild house mice: effects of strain, sex and diet. Appl. Anim. Behav. Sci. 147, 186–193. doi: 10.1016/j.applanim.2013.05.009
Sharp, J. G., Garnick, S., Elgar, M. A., and Coulson, G. (2015). Parasite and predator risk assessment: nuanced use of olfactory cues. Proc. R. Soc. B Biol. Sci. 282, 1–5. doi: 10.1098/rspb.2015.1941
Shelly, T. E., Edu, J., and Pahio, E. (2007). Condition-dependent mating success in male fruit flies: ingestion of a pheromone precursor compensates for a low-quality diet. J. Insect Behav. 20, 347–364. doi: 10.1007/s10905-007-9082-3
Silverman, J., and Liang, D. (2001). Colony disassociation following diet partitioning in a unicolonial ant. Naturwissenschaften 88, 73–77. doi: 10.1007/s001140000198
Skeen, J. T., and Thiessen, D. D. (1977). Scent of gerbil cuisine. Physiol. Behav. 19, 11–14. doi: 10.1016/0031-9384(77)90151-2
Smadja, C., and Butlin, R. K. (2009). On the scent of speciation: the chemosensory system and its role in premating isolation. Heredity 102, 77–97. doi: 10.1038/hdy.2008.55
Smyth, R. R., and Hoffmann, M. P. (2002). Correspondence between rates of host plant consumption and responses to the Acalymma vittatum male-produced aggregation pheromone. Physiol. Entomol. 27, 235–242. doi: 10.1046/j.1365-3032.2002.00293.x
Sorvari, J., Theodora, P., Turillazzi, S., Hakkarainen, H., and Sundstrom, L. (2008). Food resources, chemical signaling, and nest mate recognition in the ant Formica aquilonia. Behav. Ecol. 19, 441–447. doi: 10.1093/beheco/arm160
South, S. H., House, C. M., Moore, A. J., Simpson, S. J., and Hunt, J. (2011). Male cockroaches prefer a high carbohydrate diet that makes them more attractive to females: implications for the study of condition dependence. Evolution 65, 1594–1606. doi: 10.1111/j.1558-5646.2011.01233.x
Svensson, G. P., Ryne, C., and Lofstedt, C. (2002). Heritable variation of sex pheromone composition and the potential for evolution of resistance to pheromone-based control of the Indian meal moth, Plodia interpunctella. J. Chem. Ecol. 28, 1447–1461. doi: 10.1023/A:1016204820674
Svensson, M. G. E., Bengtsson, M., and Löfqvist, J. (1997). Individual variation and repeatability of sex pheromone emission of female turnip moths Agrotis segetum. J. Chem. Ecol. 23, 1833–1850. doi: 10.1023/B:JOEC.0000006454.84415.5f
Symonds, M. R. E., and Elgar, M. A. (2008). The evolution of pheromone diversity. Trends Ecol. Evol. 23, 220–228. doi: 10.1016/j.tree.2007.11.009
Tabata, J., Hoshizaki, S., Tatsuki, S., and Ishikawa, Y. (2006). Heritable pheromone blend variation in a local population of the butterbur borer moth Ostrinia zaguliaevi (Lepidoptera: Crambidae). Chemoecology 16, 123–128. doi: 10.1007/s00049-006-0337-9
Taft, S., Najar, A., and Erbilgin, N. (2015). Pheromone production by an invasive bark beetle varies with monoterpene composition of its naïve host. J. Chem. Ecol. 41, 540–549. doi: 10.1007/s10886-015-0590-x
Thomas, M. L., and Simmons, L. W. (2008). Cuticular hydrocarbons are heritable in the cricket Teleogryllus oceanicus. J. Evol. Biol. 21, 801–806. doi: 10.1111/j.1420-9101.2008.01514.x
Umbers, K. D. L., Symonds, M. R. E., and Kokko, H. (2015). The mothematics of female pheromone signaling: strategies for aging virgins. Am. Nat. 185, 417–432. doi: 10.1086/679614
van Wilgenburg, E., Symonds, M. R. E., and Elgar, M. A. (2011). Evolution of cuticular hydrocarbon diversity in ants. J. Evol. Biol. 24, 1188–1198. doi: 10.1111/j.1420-9101.2011.02248.x
Veenendaal, M. V. E., Painter, R. C., de Rooij, S. R., Bossuyt, P. M. M., van der Post, J. A. M., Gluckman, P. D., et al. (2013). Transgenerational effects of prenatal exposure to the 1944–45 Dutch famine. BJOG 120, 548–554. doi: 10.1111/1471-0528.12136
Verzijden, M. N., ten Cate, C., Servedio, M. R., Kozak, G. M., Boughman, J. W., and Svensson, E. I. (2012). The impact of learning on sexual selection and speciation. Trends Ecol. Evol. 27, 511–519. doi: 10.1016/j.tree.2012.05.007
Vonshak, M., Dayan, T., Foucaud, J., Estoup, A., and Hefetz, A. (2009). The interplay between genetic and environmental effects on colony insularity in the clonal invasive little fire ant Wasmannia auropunctata. Behav. Ecol. Sociobiol. 63, 1667–1677. doi: 10.1007/s00265-009-0775-9
Wallace, P. (1977). Individual discrimination of humans by odor. Physiol. Behav. 19, 577–579. doi: 10.1016/0031-9384(77)90238-4
Ward, A. J. W., Herbert-Read, J. E., and Simpson, S. J. (2011). Diets and decisions: the potential use of food protein cues in dietary, sexual and social decisions by mosquitofish. Anim. Behav. 82, 783–790. doi: 10.1016/j.anbehav.2011.07.010
Weddle, C. B., Mitchell, C., Bay, S. K., Sakaluk, S. K., and Hunt, J. (2012). Sex-specific genotype-by-environment interactions for cuticular hydrocarbon expression in decorated crickets, Gryllodes sigillatus: implications for the evolution of signal reliability. J. Evol. Biol. 25, 2112–2125. doi: 10.1111/j.1420-9101.2012.02593.x
Keywords: diet, pheromones, signal reliability, animal communication, signal variation, signature mixtures
Citation: Henneken J, Goodger JQD, Jones TM and Elgar MA (2017) Diet-Mediated Pheromones and Signature Mixtures Can Enforce Signal Reliability. Front. Ecol. Evol. 4:145. doi: 10.3389/fevo.2016.00145
Received: 02 November 2016; Accepted: 23 December 2016;
Published: 10 January 2017.
Edited by:
Sasha Raoul Xola Dall, University of Exeter, UKReviewed by:
Ximena J. Nelson, University of Canterbury, New ZealandTristram D. Wyatt, University of Oxford, UK
Copyright © 2017 Henneken, Goodger, Jones and Elgar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mark A. Elgar, bS5lbGdhckB1bmltZWxiLmVkdS5hdQ==