 Belén Fadrique
Belén Fadrique Kenneth J. Feeley
Kenneth J. Feeley
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
GENERAL COMMENTARY article
Front. Ecol. Evol. , 12 April 2016
Sec. Biogeography and Macroecology
Volume 4 - 2016 | https://doi.org/10.3389/fevo.2016.00033
A commentary on
Novel competitors shape species' responses to climate change
by Alexander, J. M., Diez, J. M., and Levine, J. M. (2015). Nature 525, 515–518. doi: 10.1038/nature14952
There is a growing appreciation of the need to understand the effects of climate change on species interactions and how changes in interactions can influence the ability of species to persist in the face of climate change (Araújo and Luoto, 2007; Thuiller et al., 2008; Svenning et al., 2014). However, empirical or experimental studies investigating species interactions under climate change remain extremely scarce. Alexander et al. (2015) use experimental transplants of European alpine plant species and communities to provide valuable insight into some of the novel competitive interactions that may emerge as species migrate upslope to keep pace with rising temperatures. More specifically, they look at performance of plant species under simulated upslope migrations into preexisting higher-elevation plant communities as well as the performance of plant species that fail to migrate and find themselves competing with new suites of species migrating into their community from below. This is a useful approximation of some of the scenarios that are already being created by the unequal responses of species to climate and the creation of novel communities.
One limitation of the study by Alexander et al. (2015) is the omission of the earliest phases of establishment when processes such as dispersal and germination are crucial in the encroachment of initial populations of migrant species into the new locations (Hampe, 2011). In particular, the experimental set up fails to account for one of the potentially most important drivers of community assembly—priority effects. Priority effects refer to the observation that early colonists will often inhibit, or alternatively facilitate, the establishment of subsequent colonizers (Connell and Slatyer, 1977). The concept of priority effect has been assimilated into community assembly theory to explain the fact that some communities may exist in alternative stable states depending on the timing and order of the arriving species and their traits, such as competitive ability or fecundity (Young et al., 2001). Priority effects can facilitate establishment of new species through changes in the environment or mutualisms. For example, in alpine communities, cushion plants can ameliorate environmental conditions and thereby increase the survival of other species leading to higher species richness (Cavieres et al., 2014). On the other hand, inhibitory priority effects can occur through legacies in the plant-soil feedbacks (Grman and Suding, 2010) or size-mediated priority effects and asymmetric competition (Weiner, 1990; Rasmussen et al., 2014). Size-mediated or size-asymmetric priority effects is the unequal competitive ability between different stages of the established and the novel species, such that incumbent species with adult individuals have an inherent competitive advantage over seeds or seedlings from the incoming species which must germinate and establish. As an example of inhibitory priority effects, Körner et al. (2008) found a persistent difference in the final composition and biomass of experimental communities with grasses, herbs, and legume species, depending on the order that species were planted.
The experimental transplants conducted by Alexander et al. (2015) do not allow for the possible consequences of priority effects to be considered. In their experimental setup, the novel competitors were transplanted into a community with plants of the same growth form and size. This methodological approach eliminates any size- or stage-based asymmetric competition that migrating species will often have to face during the earliest stages of establishment and growth in newly-suitable habitats (scenarios 2–4).
Indeed, the experiments of Alexander et al. (2015) exclude any consideration of germination which is clearly an important filter for any plant migrating or expanding its range to include newly-suitable areas (Rehm and Feeley, 2013). Germination success is determined by both abiotic and biotic conditions. Assuming that species are remaining at equilibrium with climate and are migrating into areas where the abiotic conditions are suitable, then biotic interactions and priority effects will be of heightened importance. As species arrive into the newly-suitable areas they will have to germinate into communities of established individuals and thus may be at an immediate competitive disadvantage. For example, seeds of migrating species can be out-shaded by larger established individuals even if those individuals are of species that are experiencing decreased performance due to climate change. Conversely, the germination of arriving seeds could be facilitated by environmental amelioration (Cavieres et al., 2014). In the experiments of Alexander et al. (2015), all individuals of both the migrating and incumbent species are planted simultaneously and at a similar growth stage, thus excluding any possibility of inhibitory or facilitative priority effects.
Focusing on inhibitory priority effects, some of the ways that they could affect the four scenarios proposed by Alexander et al. (2015) are listed in Table 1. Priority effects will affect the establishment of focal species and competitors when migrating into new elevations. On the other hand, focal species will benefit from priority effects when other species attempt to migrate into their range. It is therefore possible that a species that would normally be outcompeted by a stronger competitor can persist longer than expected as a consequence of simply already being established. In other words, a weaker, inferior competitor can prevent the establishment of the later arriving stronger competitors thereby preventing species migrations.
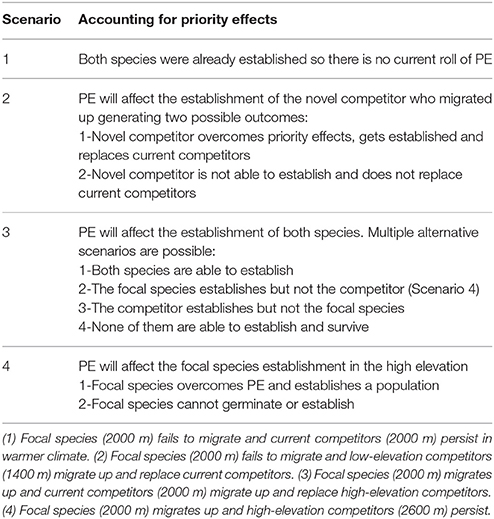
Table 1. The 4 scenarios proposed by Alexander et al. (2015) and some possible outcomes if priority effects are incorporated.
While few studies have looked at priority effects and competitive interactions within the framework of climate change, novel competitive interactions have in fact been extensively studied within the context of species introductions and invasive species (Stevens and Fehmi, 2011). Specifically, research on the establishment of introduced species has provided insight into establishment of species in novel ecosystems. From these studies we know that the differential timing of germination between established and novel plant species can be a primary factor determining the success of the invading species and the final composition of the community (Dickson et al., 2012). Research that was developed under the invasive species framework should be applied to future studies investigating the novel communities that are being created as a consequence of climate change.
Research on the synergistic effects of climate change and species interactions is conceptually and experimentally complex. Alexander et al. (2015) experimentally assess the performance of four alpine species under four proposed scenarios of species migrations. However, priority effects, excluded from the experiment, have a potentially-crucial but as of yet unknown role in the establishment of species and therefore the formation of new community assemblages. Inhibitory priority effects mediated by size or stage-asymmetric competition could potentially alter the outcome of the interactions by preventing the establishment of migrating novel competitors or by extending the persistence of the incumbent species even when they are weaker competitors. In order to better understand how species and communities will be affected by climate change it is clear that we need a more complete understanding of the complex ways that species interact and compete.
BF and KF contributed equally to the manuscript.
The authors are supported by the U.S. National Science Foundation (DEB-1350125).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Alexander, J. M., Diez, J. M., and Levine, J. M. (2015). Novel competitors shape species' responses to climate change. Nature 525, 515–518. doi: 10.1038/nature14952
Araújo, M. B., and Luoto, M. (2007). The importance of biotic interactions for modelling species distributions under climate change. Glob. Ecol. Biogeogr. 16, 743–753. doi: 10.1111/j.1466-8238.2007.00359.x
Cavieres, L. A., Brooker, R. W., Butterfield, B. J., Cook, B. J., Kikvidze, Z., Lortie, C. J., et al. (2014). Facilitative plant interactions and climate simultaneously drive alpine plant diversity. Ecol. Lett. 17, 193–202. doi: 10.1111/ele.12217
Connell, J. H., and Slatyer, R. O. (1977). Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 111, 1119–1144. doi: 10.1086/283241
Dickson, T. L., Hopwood, J. L., and Wilsey, B. J. (2012). Do priority effects benefit invasive plants more than native plants? An experiment with six grassland species. Biol. Invasions 14, 2617–2624. doi: 10.1007/s10530-012-0257-2
Grman, E., and Suding, K. N. (2010). Within-year soil legacies contribute to strong priority effects of exotics on native california grassland communities. Restor. Ecol. 18, 664–670. doi: 10.1111/j.1526-100X.2008.00497.x
Hampe, A. (2011). Plants on the move: the role of seed dispersal and initial population establishment for climate-driven range expansions. Acta Oecol. 37, 666–673. doi: 10.1016/j.actao.2011.05.001
Körner, C., Stöcklin, J., Reuther-Thiébaud, L., and Pelaez-Riedl, S. (2008). Small differences in arrival time influence composition and productivity of plant communities. New Phytol. 177, 698–705. doi: 10.1111/j.1469-8137.2007.02287.x
Rasmussen, N. L., Van Allen, B. G., and Rudolf, V. H. W. (2014). Linking phenological shifts to species interactions through size-mediated priority effects. J. Anim. Ecol. 83, 1206–1215. doi: 10.1111/1365-2656.12203
Rehm, E. M., and Feeley, K. J. (2013). Forest patches and the upward migration of timberline in the southern Peruvian Andes. For. Ecol. Manage. 305, 204–211. doi: 10.1016/j.foreco.2013.05.041
Stevens, J. M., and Fehmi, J. S. (2011). Early establishment of a native grass reduces the competitive effect of a non-native grass. Restor. Ecol. 19, 399–406. doi: 10.1111/j.1526-100X.2009.00565.x
Svenning, J. C., Gravel, D., Holt, R. D., Schurr, F. M., Thuiller, W., Münkemüller, T., et al. (2014). The influence of interspecific interactions on species range expansion rates. Ecography 37, 1198–1209. doi: 10.1111/j.1600-0587.2013.00574.x
Thuiller, W., Albert, C., Araújo, M. B., Berry, P. M., Cabeza, M., Guisan, A., et al. (2008). Predicting global change impacts on plant species' distributions: future challenges. Perspect. Plant Ecol. Evol. Syst. 9, 137–152. doi: 10.1016/j.ppees.2007.09.004
Keywords: priority effects, plant migration, climate change, mountain forest, species interactions
Citation: Fadrique B and Feeley KJ (2016) Commentary: Novel competitors shape species' responses to climate change. Front. Ecol. Evol. 4:33. doi: 10.3389/fevo.2016.00033
Received: 20 January 2016; Accepted: 21 March 2016;
Published: 12 April 2016.
Edited by:
Marco A. Molina-Montenegro, University of Talca, ChileReviewed by:
Marta A. Jarzyna, Yale University, USACopyright © 2016 Fadrique and Feeley. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Belén Fadrique, YmpmYWRyaXF1ZUBob3RtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.