 David M. Logue
David M. Logue Daniel Brian Krupp
Daniel Brian Krupp- 1Department of Psychology, University of Lethbridge, Lethbridge, AB, Canada
- 2SALT Lab, Department of Psychology and Department of Mathematics and Statistics, Queen's University, Kingston, ON, Canada
- 3Program in Evolution and Governance, One Earth Future Foundation, Broomfield, CO, USA
Mated birds of many species vocalize together, producing duets. Duetting behavior occurs at two levels of organization: the individual level and the pair level. Individuals initiate vocalizations, answer their mates' vocalizations, and control the structure and timing of their own vocalizations. Pairs produce duets that vary with respect to duration, temporal coordination, and phrase-type combinations, among other properties. To make sense of this hierarchical structure, organize duetting research, and identify new avenues of investigation, we advocate a “collective behavior” approach to the study of duets. We critically review key terminology in the duetting literature in light of this approach, and elucidate six insights that emerge from the collective behavior approach: (1) Individual-level behaviors describe pair-level behaviors, but the opposite is not true; (2) The level of organization informs how we test for the rules that govern behavior; (3) Functional hypotheses about duetting must distinguish individual from group characters; (4) Stimulus-response, cybernetics, and entrainment offer alternative hypotheses for the cognitive control of duetting behavior; (5) Avian duetting has the potential to be a model system for the ontogeny of vocal interaction; and (6) The collective behavior approach suggests new avenues of research. Ultimately, we argue that nearly every aspect of duetting research stands to benefit from adopting a collective behavior approach. This approach also has applications to other forms of interactive vocal communication in birds and primates, including humans.
Introduction
In hundreds of species of birds, mated pairs sing duets (Hall, 2004, 2009). The black-bellied wren (Pheugopedius fasciatoventris) is one such species. Researchers might take one of two approaches to describe its duetting behavior. They might say that both sexes initiate song and both answer their mates' songs to form duets (Logue and Gammon, 2004). They might also note that the mates' prior phrase-type influences both the answerer's phrase-type and, in the case of females, the latency to answer. In short, they could describe what the individuals do during the act of duetting. Alternatively, the researchers might quantify the proportion of duets initiated by each pair member, the repertoire of duets that each pair can produce, or the amount of overlap between male and female phrases. That is, the researchers could describe what the pair does during the act of duetting.
As this example illustrates, duetting occurs at two levels of organization: The individual level and the pair level. The properties of duets (e.g., duet-type, duet duration) are pair-level characters because they arise from the integration of individual phenotypes. Song initiation and song answering are the individual-level behaviors that generate duets, so variation in duet structure is caused by variation in initiation and answering. Important dimensions of variation in song initiation include the decision to initiate and the choice of phrase-type. Dimensions of variation in answering include the decision to answer, the choice of answer phrase-type, and the fine-scale timing of the answer.
We can codify the relationship between the levels of behavior by saying that duetting is a collective behavior that arises from the interaction of two individual-level behaviors, song initiation and song answering. The term “collective behavior” describes behavior at the group level, like murmurations of starlings (Sturnus vulgaris; King and Sumpter, 2012), fish shoals (Couzin and Krause, 2003), or huddling rat pups (Schank and Alberts, 1997). Collective behavior can arise from local interactions (Couzin, 2009), where local can refer to both space and time. As such, functional behaviors can emerge at the group level without any global blueprint. Research on collective behavior often involves large collectives of individuals that follow simple rules, resulting in group-level behaviors that would be difficult to predict based on the individual-level behaviors (Goldstein, 1999; Couzin, 2009). In contrast, duets are produced by small collectives of individuals, resulting in group-level behaviors that are not necessarily difficult to predict based on the individual-level behaviors. Neither of these differences, however, prohibit the application of the collective behavior approach to duetting. To the contrary, as the simplest possible groups, two-individual collectives are tractable test cases that can inform collective behavior research.
Our thesis is that it is critical to acknowledge that duetting occurs at both the individual and pair levels. This idea has ample precedent in the literature. Although early work implicitly assumed that pair mates' interests were perfectly aligned (Hall, 2004), the 1980s and 1990s saw a turn toward an individual-level approach to functional research on duetting (Wickler, 1980; Sonnenschein and Reyer, 1983; Levin, 1996a,b). In the 2000s, Logue (2006) argued that an improved understanding of the individual-level behaviors that structure duets would “elucidate previously unconsidered functional links and operational constraints” (pp. 326–327). Shortly thereafter, Brumm and Slater (2007) described duetting as “collective signaling,” and hypothesized that functionality might emerge at the pair level. In her influential review of vocal duetting, Hall (2009) summarized the state of the collective behavior approach and called for its broad application: “Distinguishing between pair-level aspects of duetting and the underlying individual behaviors … is essential to understanding how and why duetting occurs.” (pp. 67, 69). Here, we clarify and expand on these ideas.
The utility of the collective behavior approach should be evaluated in comparison to the alternative approach, which is to ignore the distinction between individual-level and group-level behavior. Acknowledging that duetting occurs at two levels offers two kinds of benefits:
1. It provides a logical framework to guide duetting research. Research on duetting is complicated by the fact there are two possible levels of organization. The collective behavior approach forces researchers to specify the level of interest, encouraging precise definitions of problems, questions, and research objectives and reducing ambiguity. For example, many papers address the “function of duetting.” That term could refer to the function of initiation, answering, or both behaviors for the male, the female, or both pair members. The collective behavior paradigm requires a specification of the level of organization, eliminating this kind of ambiguity.
2. It suggests new avenues for research. Distinguishing between the levels of organization reveals areas in need of additional research. Moreover, the collective behavior approach encourages work on the relationship between individual-level behavior and the properties of group-level phenomena, like duets (Brumm and Slater, 2007). Finally, this approach facilitates adoption of the theoretical and empirical toolkits that have been developed to study collective behavior.
A comprehensive review of the duetting literature according to this framework would be largely redundant with reviews by Hall (2004, 2009) and Dahlin and Benedict (2014). Instead, we offer a critical review of duet terminology (Box 1), and six insights that can be gained by treating duetting as a collective behavior. Collectively, these insights demonstrate that nearly every aspect of duetting research stands to benefit from the collective behavior approach. This report is part of a special issue on female song in birds, so it is worthwhile to articulate its relevance to the study of female song per se. Female song is often given in the context of vocal duets (Langmore, 1998), so the biology of female song is intimately related to that of duetting. We invoke the collective behavior approach in an attempt to clarify this relationship.
BOX 1. A critical review of terminology describing duets and duetting behavior.
Answer: A vocalization in response to, and in temporal coordination with, a vocalization produced by the duetting partner (Wickler and Sonnenschein, 1989; Riegert et al., 2004; Hall, 2009; Dahlin and Benedict, 2014). Initiation and answering are the individual-level behaviors that are necessary and sufficient to generate duets (Hall, 2009). Answer is synonymous with “reply” (Diamond and Terbourgh, 1968; Mann et al., 2003), but the former term is more common in the contemporary literature. The individual that answers is the “answerer” or “responder.”
Answering rule: Individual-level psychological property linking the duetting partners' previous vocalization(s) to the structure of the focal individual's answer (Hall, 2009). Some previous definitions also included rules governing phrase timing (Logue, 2006, 2007b; Logue et al., 2008), which we now treat separately (see “Timing rule”).
Call: A vocalization that is not a song. Many species duet using calls.
Contribution: All parts of a duet produced by one individual (e.g., “the male's contribution comprised three phrases”).
Coordination: The regularity of the temporal relationship between the male and female duet contributions. Coordination can be measured at the individual level and at the pair level.
Duet: A pair-level vocalization comprising one initiation and one or more answers. In nearly all duetting bird species, duet partners are pair-mates. Notable exceptions are the male-male duets in cooperatively displaying manakins of the genus Chiroxiphia (Trainer et al., 2002).
Duet code: A set of answering rules that links the phrase-type a focal individual hears to the phrase-type that the focal individual uses to answer (Logue and Gammon, 2004; Logue, 2006, 2007b). A duet code is an individual-level property, but some or all of the links comprising a code may be shared between partners or among other individuals. To answer according to a duet code is to “adhere” to a duet code (Logue and Gammon, 2004). The term “song fitting” is synonymous with adhering to a code (Rogers et al., 2006).
Duet train: A duet that includes two or more answers (e.g., male-female-male; Brown and Lemon, 1979).
Duet repertoire: In species with duet-types, the set of types that a pair or population produces is its “duet repertoire” or “duet-type repertoire” (Levin, 1996a; Hall, 2009).
Duet-type: A structurally distinct duet. Duet-type is a pair-level property that is generated by individual-level answering rules. Duet-type is a useful term for species with highly stereotyped duets (e.g., Sonnenschein and Reyer, 1983; Rogers, 2005).
Gap: A silent interval between the phrases of a duet. Gaps can be measured at both the individual level (e.g., the female's average latency relative to the end of the prior male phrase) and the pair level (e.g., the total gap duration over a whole duet).
Initiation: A vocalization that is not an answer. Initiations constitute the beginning of a duet if the partner answers, otherwise they are solo vocalizations (songs or calls). Initiation is an individual-level behavior (e.g., “the male initiated 13.2 vocalizations per hour”). The sex that initiates a duet, however, is a pair-level property (e.g., “the pair produced 3.2 male-initiated duets per hour”; Grafe and Bitz, 2004; Hall, 2004; Logue and Gammon, 2004).
Non-random phrase-type combination: A statistical association between the phrase-types that partners use in duets. Non-random phrase-type combinations at the level of the whole duet are pair-level phenomena. Adherence to a duet code by one or both sexes is one of several ways that pairs can generate non-random phrase-type combinations (Logue, 2006, 2007b).
Overlapping: The simultaneous production of sound by both duetting partners. Overlap can be measured at both the individual level and the pair level. Duets in which there is no overlap between subsequent phrases are sometimes described as “alternating” (Hall, 2009).
Phrase: The minimum vocal unit that an individual contributes to a duet (Mann et al., 2003). Phrases are structurally identical to solo songs in many species. The term “phrase” is less likely to cause confusion than alternatives, such as “song” (sensu Levin et al., 1996), which can also describe whole duets or solo songs.
Phrase repertoire: The set of phrase-types that can be produced by an individual, a pair, or a population. Phrase repertoire should not to be confused with “duet repertoires” (as in Voigt et al., 2006).
Phrase-type: A structurally distinctive class of phrases (Mann et al., 2003; Hall, 2009).
Song: A vocalization involved in mate attraction and territory ownership. Songs are usually louder and longer than calls, and usually exhibit a strong annual and diel rhythm (Catchpole and Slater, 2003). Songs are used as duet phrases in many species. In the duetting literature, the term “song” has been used to mean a solo song, a phrase, and a whole duet. To avoid confusion, we suggest that authors use modifiers to specify their meaning (e.g., “male solo song”) when using the term with respect to duet-singing birds.
Termination: The final phrase of a duet, or the act of singing that phrase. Duet termination is a pair-level property. For example, consider the duet “male-female-male.” The male terminates because the female does not answer his second phrase.
Timing rule: Individual-level psychological property linking the timing of one or more previous vocalizations to the timing of the focal individual's answer.
Insight 1: Individual-Level Behaviors Describe Pair-Level Behaviors, but the Opposite is not True
Description is the first phase of most research programs on duetting systems. Here, we consider description in light of the collective behavior framework.
Initiation and Answering
Answering (as defined in Box 1) is necessary and, along with initiation, sufficient to generate a duet. After the initiation, all subsequent phrases by either partner in the duet are answers. In some cases, it is useful to distinguish between the initial answer and “secondary answers” that prolong the duet.
The individual that initiates a vocalization does not directly control whether it will be answered, so rates of duet initiation (as in Illes, 2015) are pair-level properties (Hall, 2009). The term “initiation” describes an individual-level behavior but, perhaps counter-intuitively, “solo singing” is a pair-level property because the mate must refrain from answering for an initiation to be a solo song. Thus, the relative rates of duets and of solo songs (Grafe and Bitz, 2004; Mennill and Vehrencamp, 2005; Mennill, 2006) are pair-level properties. So too is the duration of the duet train (Logue, 2007b), which emerges from the answering behavior of both partners. The preceding statements assume that solo vocalizations are potential duet initiations. Researchers should test this assumption, to avoid lumping classes of solo vocalizations that are never answered with those that are. Care should also be taken to distinguish phrases that are never used in duets from those that change structure in response to being answered (Tobias and Seddon, 2009).
The proportion of duets initiated by males and females are widely reported pair-level statistics in the duetting literature (Gill et al., 2005; Mennill and Vehrencamp, 2005; Hall, 2006; Mennill, 2006; Topp and Mennill, 2008; Mann et al., 2009; Elie et al., 2010; van den Heuvel et al., 2014). Hall (2009, p. 77) warned that “the percentage of duets initiated by one sex or the other is not a good measure of the propensity of the opposite sex to duet, as it does not consider the unanswered (solo) songs of either sex and probably just reflects sex differences in song initiation rates.” We agree with the first part of this statement, but further note that differences in duet initiation rates could arise from differences in the individual-level behaviors initiation rate, answer rate, or a combination of the two (Table 1).
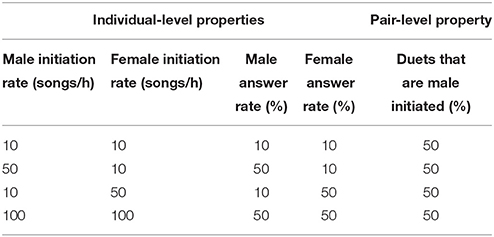
Table 1. A sample of individual-level property sets that produce an equivalent percentage of male initiated duets.
Many properties of duets depend on both partners' song initiation and song answering behaviors. These include the occurrence of duets, duet rate, duet initiation rates (including the proportion of duets initiated by males and females), duet termination rates, duet train duration, solo singing rates (if solo songs are potential duet initiations), the relative rates of duetting and solo singing, and the proportion of songs that occur in duets.
The individual-level properties, male and female initiation rates and answer rates, can be used to unambiguously reconstruct pair-level properties. Pair-level attributes, however, cannot be used to unambiguously reconstruct individual-level behavior. For example, based on the statement “50% of duets were initiated by the male” it is impossible to estimate the magnitude of the individual-level attributes “initiation rate” and “answer rate” or their relative values in the two sexes (Table 1). Because of this ambiguity, individual-level behaviors should constitute the main descriptors of duetting behavior (Hall, 2009). Benefits of this approach include clarity (these metrics leave no ambiguity at either level of organization), simplicity (only four numbers are required to describe patterns of initiation and answering, more if secondary answer rates are included) and comparability (metrics can be compared across studies).
Our recommendation to focus on individual-level descriptors of initiation and answering runs counter to some other recommendations for duet description, which include both individual- and pair-level descriptions (Farabaugh, 1982; Dahlin and Benedict, 2014). Indeed, pair-level descriptors should be reported if they are hypothesized to be a salient aspect of the signal (e.g., if extra-pair receivers are known to be indifferent to the individual-level processes that generate duets). Of course, pair-level metrics are the only option in systems where it is impossible to distinguish the partners' duet contributions (Benedict and McEntee, 2009; Benedict, 2010). In such systems, authors should report pair-level metrics and explain their limitations.
Phrase-Type Choice
Analogous to “song-type,” the term “duet-type” describes a discrete structural class of duets. Individual patterns of answering and phrase-type choice determine this pair-level property. Duet-types may be shared among all pairs, or distinctive to each pair. When duet-types are shared, pairs may “match” the duets of neighboring pairs, much like individual birds match song-types (Grafe and Bitz, 2004; Marshall-Ball and Slater, 2004).
Duet-type is a useful construct for species with discrete duet structures (Sonnenschein and Reyer, 1983; Rogers, 2005). In many duetting species, however, the exact sequence of phrase-types comprising a duet varies a great deal, prohibiting the identification of discrete types (Vencl and Soucek, 1976; Brown and Farabaugh, 1991; Logue, 2007b; Dahlin and Benedict, 2014; Table 2). For example, duets in wrens (Troglodytidae) are characterized by variation in the individual that initiates the duet, the number of answers in the duet trains, and the pattern of phrase-type switching within duets (Marshall-Ball and Slater, 2004). Such patterns are to be expected if duet organization is driven by local, individual-level processes (like adherence to duet codes) rather than global, pair-level processes (like shared, whole duet templates). If duets do not belong to structurally discrete types, pair-level descriptors like “duet repertoire” and “duet sharing” fall short of the desired level of precision. In such cases, we suggest that researchers interested in characterizing variation in duets phrase combinations focus on individual-level properties that influence duet structure, such as answering rules, and pair-level properties that characterize and constrain duetting, such as sharing of answering rules and phrase-types between mates, and the pair's phrase-type repertoire.
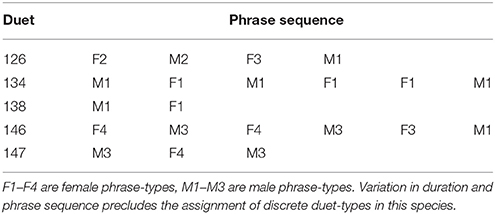
Table 2. Phrase-type sequences representing five duets from one mated pair of black-bellied wrens (Pheugopedius fasciatoventris).
Fine-Scale Timing
Most pair-level properties that depend on the interaction of the duetting individuals' timing rules fall under the umbrella of “temporal coordination” (Farabaugh, 1982; Levin, 1996a; Hall, 2009; Dahlin and Benedict, 2014). Coordination can be characterized with respect to phrase overlapping, silent gaps between phrases and variation in these properties (Elie et al., 2010).
In many duetting systems, a pair-level description of duet coordination does not unambiguously specify the individual-level behaviors responsible for phrase timing. For example, an antiphonal duet with high temporal coordination (minimal gaps and overlapping sections) can be achieved by both sexes basing the timing of their phrases on the timing of the mate's previous phrase or by one sex adhering to an endogenous repetition rate and the other sex adjusting its timing in response. As above, however, it is possible to unambiguously infer duet properties from individual-level patterns of behavior. We therefore recommend prioritizing descriptions of duet timing at the individual level. Pair-level properties should also be reported when they bear on research hypotheses.
To conclude this section, the way in which duets are described depends on the level of organization. Many descriptions of duetting in the literature (e.g., Mann et al., 2009) and recommendations to describe duets (Farabaugh, 1982; Dahlin and Benedict, 2014) emphasize the (pair-level) structure of the whole duet. Pair-level descriptions alone are rarely adequate for reconstructing the individual-level behaviors that produce duets. Individual-level behaviors, however, can be used to unambiguously reconstruct pair-level phenomena. We therefore recommend that authors report duetting behavior at the individual level when possible. Some pair-level descriptions may also be worth including (e.g., for ease of comparison between studies, for specific research hypotheses), when space permits.
Insight 2: The Level of Organization Informs how we Test for the Rules that Govern Behavior
The collective behavior approach to duetting research emphasizes the relationship between individual-level behavioral rules and the structure of pair-level duets. Researchers have attempted to identify individual-level answering rules with both observational methods (von Helversen and Wickler, 1971; Vencl and Soucek, 1976; Levin, 1996a; Mann et al., 2003; Mennill and Vehrencamp, 2005; Hall, 2006; Rogers et al., 2006; Valderrama et al., 2013), and interactive playback experiments (Logue, 2006, 2007b; Templeton et al., 2013). Analyses of observational data can be classified as “directional” analyses that measure phrase transitions within duets, or “holistic” analyses that measure associations at the level of the whole duet. Directional associations are measured between a focal phrase and some preceding referent (usually the mate's prior phrase). The order of the phrases is taken as evidence of causation (i.e., earlier events are assumed to influence later ones). Holistic associations are measured non-directionally, at the level of the whole duet (Mann et al., 2003; Mennill and Vehrencamp, 2005).
Phrase-type associations at the level of the whole duet are clearly a pair-level property, as evidenced by the single phrase association matrix that can be produced for each pair. Even in directional studies, however, observational methods cannot unambiguously demonstrate the individual-level answering rules that structure duets. This is because they cannot rule out the possibilities that male and female phrase-types are both influenced by some extrinsic cue, that phrase-type choice in one or both sexes is influenced by a non-vocal signal from the pair mate, or that the mates' intrinsic phrase-type switching programs are correlated (Logue, 2006). Thus, failure to find non-random phrase-type associations in an observational study is evidence against answering rules, but positive findings leave open several possible explanations. Playback experiments in which birds answer randomly ordered duet phrase stimuli allow researchers to rule out alternative explanations, so they provide the strongest evidence of the causal effects of the partner's phrases on the actor's phrase-type choice.
Like answering rules, timing rules can also be investigated observationally (Rivera-Cáceres, 2015) or experimentally (Templeton et al., 2013). Observational studies infer timing rules by modeling the delay to answer as a function of prior events in the duet (Logue et al., 2008; Rivera-Cáceres, 2015). The order of events is assumed to be evidence of the direction of influence. The song answering experimental protocol used to test for answering rules can also be used to characterize timing rules (Logue et al., 2008; Templeton et al., 2013).
Like observational studies of phrase choice, observational studies of timing can be directional or holistic. Directional studies, which are much more common, measure the timing of a focal phrase relative to one or more preceding referents. Such studies may be effective at determining individual-level behaviors, as evidenced by the high concordance between the results of directional observational and experimental approaches in a study that used both (Logue et al., 2008). As with studies of phrase choice, however, experiments provide more compelling causal explanations at the individual level than do observational studies. Holistic studies characterize the coordination of the whole duet, but provide little information about individual-level processes.
In conclusion, observational studies are well-suited to characterize pair-level properties of duets, with respect to both structure and timing. We encourage researchers studying the individual-level rules that govern duet participation to adopt the experimental approach.
Insight 3: Functional Hypotheses About Duetting must Distinguish Individual from Group Characters
As we have discussed, aspects of a duet can be described as individual phenotypic characters and as group characters. However, it is not always clear which description is relevant to a particular research hypothesis. We suggest that causal thinking can help resolve this issue.
Scientists are chiefly interested in determining the cause of an effect; an association between two variables is rarely considered meaningful unless it sheds light on their causal relationship. In evolutionary biology, a relevant concern is whether a given character caused differences in fitness in the past, giving rise to selection and adaptation. This causal link between character and fitness is the foundation for all questions of evolved function.
Figure 1 formalizes the causal relationships between characters and fitness with path diagrams (Pearl, 2009), in which solid arrows reflect direct causal effects and dotted lines reflect correlations between variables. Consider a focal individual i and her partner j who express, respectively, traits pi and pj and, together, a group trait P. There are three possible ways that i's fitness, wi, may be affected by the expression of these traits: only individual characters directly affect fitness (solid arrows between pi and wi and between pj and wi, as in Figure 1A), only the group character directly affects fitness (solid arrow between P and wi, as in Figure 1B), or both individual and group characters directly affect fitness (solid arrows between pi and wi, between pj and wi, and between P and wi, as in Figure 1C). According to the logic above, P has functional relevance only in the diagrams in Figures 1B,C, because it is only in these diagrams that the group character directly influences fitness. Although Figure 1A also contains a relationship between group character and fitness, it is correlational rather than causal.
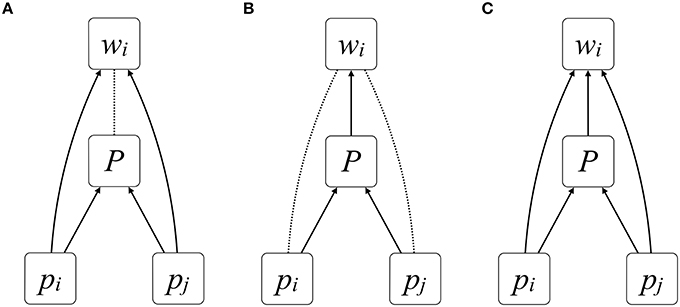
Figure 1. Causation, correlation, and individual vs. group characters. Solid arrows denote direct causal effects of one variable on another, whereas dotted lines denote correlations between them. Fitness, wi, can be directly affected by (A) individual traits pi and pj alone, (B) the group trait P alone, or (C) both individual and group traits pi, pj, and P. The group trait P is functionally relevant in (B,C) because it has a direct causal effect on fitness and, therefore, selection; it is not relevant in (A) because it exerts no influence on fitness.
For example, the decision to initiate a vocalization may influence fitness directly, or it may influence fitness indirectly, via its effects on the group character. At the individual level, initiation may have some of the same costs and benefits as solo singing. Indeed, initiation is solo singing if the behavior fails to elicit an answer from the partner. However, initiation also affects a group character: pairs with two initiators will duet more frequently than pairs with one initiator; likewise, pairs with one initiator will duet more frequently than those without any initiator. If the frequency of duetting also has a direct causal effect on individual fitness—or, if fitness cannot be measured, on a reasonable proxy thereof—it is a necessary part of the duet “story.” Hence, we have two characters under selection: an individual character (the propensity to initiate) and a group one (duet frequency). As in Figure 1C, the individual character influences the group character, and both influence fitness.
We offer three clarifications about this argument. First, it is independent of the concept of group selection. Group selection is contingent on direct causal effects of group character on group fitness, but group characters can cause changes in either individual or group fitness (Okasha, 2006, 2012; Krupp, in press). This is why Figure 1 and the example of duet frequency as a causal group character given above are able to make reference only to individual fitness, wi.
Second, this argument is independent of the concepts of synergy, non-additivity, and non-linearity (Queller, 1984, 1985; Taylor and Maciejewski, 2012; Taylor, 2013). Consider again the example above, relating duet frequency to individual fitness, and let the additional fitness caused by the duet frequency of pairs that have zero, one, or two initiators be, respectively, w0, w1, and w2. If w0 = 0, w1 = 1, and w2 = 2, we would have an additive effect of duet frequency and, if w0 = 0, w1 = 1, and w2 = 3, we would have a synergistic effect. Neither effect, however, has any bearing on the functional relevance of a group character. What matters is contained in the phrase “the fitness effect caused by the duet frequency of pairs,” above: that the group character, linear, or otherwise, directly causes differences in fitness (Krupp, in press).
Third, there is no purely statistical method to identify the effect of a group character. This is because causality depends on more than the joint distribution of character and fitness (Pearl, 2009). Group characters depend on individual characters, and so both individual and group characters will predict fitness even though only one may actually cause it (Okasha, 2006, in press; Krupp, in press). Statistical analysis alone cannot determine which of these models describes the true character-fitness causal structure, because each character is connected, either by causation, or by correlation, to fitness.
Signal receivers are major selective agents of signaling behaviors (Owren et al., 2010; Bradbury and Vehrencamp, 2011). A key step toward understanding the function of duetting behavior is figuring out whether receivers attend to individual-level properties, pair-level properties, or both. For example, we might ask “do receivers attend to the overall coordination of a duet or to the fine-scale timing of answers by the duetting individuals?” It is possible to conduct playback experiments to address this question, but we are not aware of any existing studies that attempt to do this with respect to duet timing or any of the other properties discussed above.
The relative salience of individual- and pair-level properties also has implications for the evolution of duetting behavior. If, for example, extra-pair receivers respond to individual-level temporal coordination, selection would drive both sexes toward higher coordination. Alternatively, if the overall temporal coordination is all that matters, one sex might evolve to compensate for the other.
The concept of causality disciplines our thinking about the biological significance of duet-level properties, such as duet duration, duet coordination, and duet-type. At one extreme, if fitness effects are entirely due to individual-level characters, then the duet-level properties of the pair exert no fitness consequences. At the other extreme, duet-level properties may be the only direct causal forces. If this is true, any combination of individual-level behaviors that results in a given duet structure will be functionally equivalent. It seems logical that individual-level functionality could apply to either within-pair or extra-pair receivers, whereas pair-level functionality would tend to apply to communication between the duetting pair and extra-pair receivers. Brumm and Slater (2007, p. R521) touched on the idea of group-level functionality when they wrote, “the interplay between the single components produced by different individuals can form a new quality, which, in turn, may act as a signal itself. This sort of meta-parameter could be formed by the particular phrases that individuals combine … or the temporal patterning between the individual contributions of the collective signal.” In any given duetting system both individual-level behaviors and duet-level properties may be salient to signal receivers (including the duetting birds themselves). This is, of course, an empirical question. Incorrect assignment of the level of behavioral relevance can distort research (Bell, 2014; Pellis and Pellis, 2015), so care should be taken to test the effects of behavioral variation at both levels.
Insight 4: Stimulus-Response, Cybernetics, and Entrainment Offer Alternative Hypotheses for the Cognitive Control of Duetting Behavior
Research in collective behavior aims to understand the individual-level decision processes that produce organized group-level phenomena. Duetting birds are ideal systems for research on the cognitive control of group-level behavior because they respond to stimuli immediately, and with high temporal and structural specificity (Thorpe, 1963; Laje and Mindlin, 2003; Amador et al., 2005; Logue, 2007b; Fortune et al., 2011). The relative paucity of studies on the cognitive control of interactive vocal communication in duetting birds contrasts with the large and growing number of such studies in humans and other primates. Drawing from that literature and other approaches to the control of behavior, we lay out three testable hypotheses about the cognitive control of duetting behavior, and describe an approach to link these mechanisms to pair-level duet properties.
Research in Behavioral Ecology generally assumes that the control of behavior can be described by the stimulus-response (S-R) model (Bell, 2014). It is unclear, however, whether S-R accurately models the cognitive control of song answering. A key prediction of the S-R model is that the structure and timing of answers (responses) should depend entirely on the phrase or phrases that are necessary to stimulate the answer (stimuli). The finding that the fine-scale timing of answers depends, in part, on an endogenous rhythm that can be measured in the answerer's previous phrases is evidence against a pure S-R model of the control of song answering (Hall and Magrath, 2007; Logue et al., 2008; Templeton et al., 2013; Rivera-Cáceres, 2015).
Cybernetic theory (Bell, 2014) offers an alternative model for the control of song answering in which individuals attempt to satisfy the “goal state” of hearing a complete phrase pair. For example, suppose that when a female hears Male Phrase X, she pursues the goal state of hearing the phrase pair “Male Phrase X → Female Phrase Y.” A distinctive prediction of the cybernetic control hypothesis could be tested by playing the female a stimulus comprising Male Phrase X followed by the first few notes of Female Phrase Y. According to the S-R hypothesis, the female should sing all of Female Phrase Y, but according to the cybernetic hypothesis, she should (at least sometimes) sing only the part of Phrase Y that is not included in the stimulus. One appealing characteristic of the cybernetic model is that it specifies both phrase structure and timing rules, eliminating the need for separate explanations for the evolution of answering rules and timing rules.
A third hypothesis for the cognitive control of duetting behavior sees duet partners as entrained oscillators (Hasson et al., 2012; Takahashi et al., 2013). Hasson et al. (2012) argue that during dyadic vocal interaction, oscillations in acoustic signals entrain with patterns of neural activity within and between interactants. Entrainment might functionally amplify signals and facilitate turn taking. Some existing data are consistent with the hypothesis that the temporal coordination of avian duets results from the coupled oscillator mechanism, as opposed to temporally local call-and-response mechanisms. Answer timing in wrens is influenced by both the focal individual's previous phrases and the mate's phrases, as would be expected given coupling (Logue et al., 2008; Rivera-Cáceres, 2015). Rufous hornero duets offer the best evidence of a coupled oscillator mechanism in birds. As the male hornero's song accelerates, the female locks into a series of rhythmic states relative to the male (Laje and Mindlin, 2003; Amador et al., 2005).
Chief among the methods used to study collective behavior are agent-based models (ABMs; Goldstone and Janssen, 2005). In ABMs, autonomous agents follow behavioral rules triggered by their local environment. These models are well-suited to test hypotheses about the control of answering behaviors. For example, one could program real or virtual duetting robots to follow duetting rules based on S-R, template matching, or entrainment. The ABMs could generate populations of duets, the properties of which could be compared to those of real duets. It would be particularly interesting to see how models behaved over variable inter-individual distances (Logue, 2007a; Templeton et al., 2013) or when more than two individuals answer, as occurs in chorusing species (Mann et al., 2006).
Insight 5. Avian Duetting has the Potential to be a Model System for the Ontogeny of Vocal Interaction
Song learning research has advanced our understanding of word acquisition at the behavioral (Lipkind et al., 2013), neurological (Doupe and Kuhl, 1999), and genetic (Fisher and Scharff, 2009; Pfenning et al., 2014) levels. Although an excellent model of word acquisition, song learning per se is not a model for the development of interactive vocal communication because it does not specify the way that individuals use their vocal signals during interactions. An animal model of vocal interaction could promote advances in academic disciplines, like Linguistics and Ethology, as well as applied fields like human-computer interactions and speech pathology. We suggest duetting in songbirds (Order: Passeriformes, Suborder: Passeri) as a model of interactive vocal communication in humans (Todt and Naguib, 2000; Logue and Stivers, 2012). Both processes involve orderly turn taking, both involve listening and rapidly responding with a stimulus-appropriate answer, and both appear to require learning, as we argue below. Unfortunately, we know very little about the ontogeny of duetting behavior, much less its neurological and genetic underpinnings. The collective behavior approach helps to advance this line of research by emphasizing that pair-level development is a consequence of individual-level development on the part of one or both pair mates.
We focus on the ontogeny of duetting behavior during two life phases. Although duetting may develop at other life phases (e.g., during dispersal, while territorial but unmated) existing evidence suggests that early life and the extended pair bonding period (potentially lasting the duration of the pair bond) are particularly important. In the first phase, juvenile birds develop the ability to initiate and answer song, and the rules that structure duets. The second phase of duet development occurs in the context of the developing pair bond. During this phase, individuals may adjust their own duetting behavior in response to the mate's (e.g., to better coordinate, temporally, or structurally, with the mate).
Four lines of observational evidence indicate that juvenile songbirds learn the details of their duetting behavior. First, songbirds learn their songs, including the phrases used in duets (Wickler and Sonnenschein, 1989; Levin et al., 1996; Catchpole and Slater, 2003). Although it is not possible to rule out a scenario in which birds apply non-learned answering rules to learned phrases, parsimony favors the hypothesis that answering rules are also learned. Second, duet codes in many species are individually distinctive (Hall, 2009), but in at least one species same-sex individuals share elements of their codes (Logue, 2006). This pattern of partial code sharing is a key prediction of the hypothesis that codes are learned. Third, juvenile birds have been observed duetting and chorusing with their parents, often mimicking the same-sex parent's role in the duet (Farabaugh, 1983; Brown and Farabaugh, 1991; Rivera-Cáceres and Guerrero Esmeralda, 2014; Figure 2). Finally, a recent study of plain wrens offers a comprehensive analysis of the ontogeny of answering rules and timing rules. It shows that juveniles duet with poorer temporal coordination than do adults, answer according to their parents' duet codes, and improve their adherence to that code over time (Rivera-Cáceres and Guerrero Esmeralda, 2014).
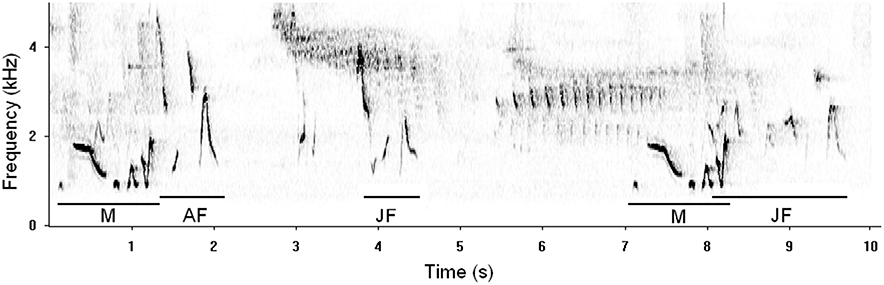
Figure 2. A spectrogram of a field observation that may depict duet code learning. A duet between an adult male (M) and adult female (AF) begins at 0 s. At 4 s the pair's juvenile daughter (JF) approximates the adult female's song. After the male repeats his song at 7 s, the daughter duets with him, again approximating the mother's answer. Spectrograph drawn from a field recording by D.M.L (Gamboa, Republic of Panamá, 2001).
Three studies focus on the ontogeny of pair-level duet properties during the extended pair-bonding period. In a cross-sectional study of plain wrens (Cantorchilus modestus), pairs that had been together for at least 2 years exhibited stronger associations between male and female phrase-types than did new pairs (Marshall-Ball et al., 2006). Neither Arrowood (1988) nor Benedict (2010) found evidence of changes in temporal coordination over the duration of the pair bond in white-winged parakeets (Brotogeris versicolurus) and California towhees (Pipilo crissalis), respectively. The pair-level approach to duet ontogeny used by these studies can tell us about developmental changes (or lack thereof) in whole duets that may be salient to receivers. They do not, however, reveal the individual-level ontogenetic processes (e.g., learning) that shape duetting behavior. For example, in the study of plain wrens, it is not possible to determine whether males, females, or both sexes changed their answering rules. Because individuals (rather than pairs) learn, studies aimed at understanding rule learning should focus on the individual level.
Three other studies bear on individual-level duet ontogeny in mated birds. Levin (1996a) conducted a mate exchange experiment on bay wrens (Cantorchilus nigricapillus). All four birds in the study answered with more variable timing immediately after re-mating than they did with their previous (established) mate, but only one (a female) met the criterion of statistical significance. The author concluded that “individual repertoires and duet precision do not change following a change in mates” (p. 1093). Our confidence in this conclusion is limited by the study's small sample size (two pairs), the short duration of post-pairing observations (3 days), and the focus on answering rules in males (to the exclusion of females). Logue (2007b) found that female black-bellied wrens adhere strictly to duet codes, whereas males adhere weakly to the reciprocal of their mates' codes. These findings suggest the hypothesis that females enter the pair-bond with a crystallized code, and males learn their mates' codes over time. Hall and Magrath's (2007) cross-sectional analysis of magpie lark (Grallina cyanoleuca) duets provides evidence that the fine-scale timing of answers changes over the duration of the pair bond. Both males and females in well-established pairs answered with lower temporal variability than did same-sex birds with new partners (see Figure 2 in Hall and Magrath, 2007).
Thus, the most compelling evidence that answering rules and timing rules develop in mated adult birds comes from cross-sectional studies (Marshall-Ball et al., 2006; Hall and Magrath, 2007). Cross-sectional analyses do not reveal causality, so it is possible that duetting behavior does not develop in mated adults, but rather (a) duet coordination influences pair duration, or (b) some unmeasured factor influences both duet coordination and pair duration.
In conclusion, the ontogeny of duetting behavior has the potential to serve as a model of the development of the rules that govern vocal interaction. Although pair-level research on duet ontogeny can provide biologically relevant information about changes in duet structure, it cannot reveal the development of the individual-level processes that govern duetting. Evidence is accumulating that songbirds learn answering rules and timing rules in early life. We currently lack strong evidence of duet ontogeny in mated individuals. The field stands to benefit from individual-level longitudinal studies of duetting behavior over the course of the pair bond.
Insight 6: The Collective Behavior Approach Suggests New Avenues of Research
Explicitly distinguishing the pair and individual levels suggests novel questions about duetting behavior. Here we offer three examples.
Evolution of Individual-Level Behavior
Several studies adopt the comparative approach to address evolutionary hypotheses about duetting (Farabaugh, 1982; Malacarne et al., 1991; Logue, 2005; Benedict, 2007; Hall, 2009; Mann et al., 2009; Logue and Hall, 2014). All of these studies implicitly focus on the pair-level by categorizing species as either “duetting” or “non-duetting.” Although comparative analyses rely on population- or species-level character scores, it is nevertheless possible to score the individual-level traits that characterize each population.
Analysis of the evolution of collective properties at the individual level can reveal evolutionary patterns that are not apparent from group-level analyses (Mank et al., 2005; Price et al., 2009). In the case of duetting, pair-level analyses obscure whether males, females, or both sexes initiate and answer. If answering functions differently in the two sexes (Tobias and Seddon, 2009) or if the cost-benefit ratio of answering differs between the sexes (Logue and Gammon, 2004), lumping the sexes in evolutionary analyses is likely to obscure sex differences in evolutionary patterns of duetting. Further, evolutionary reconstructions at the pair level provide no information about the interaction between answering in the two sexes (e.g., whether the evolution of answering in one sex tends to coincide with the evolution of answering in the other). Comparative studies that treat answering in the sexes separately would remedy this gap in the literature. Analyzing the evolution of initiation rates and answering rates as continuous variables would provide even more information, by addressing the high degree of natural variation in answer rates. Answering is not the only individual-level behavior that has been neglected by comparative studies. No existing studies have looked at the evolution of phrase-type choice or fine-scale timing at the individual level.
Complex and Dynamic Duetting Rules
Duetting rules specify the individual-level decision processes that structure group-level displays. Some species produce complex duets in which each partner produces multiple phrase-types (Vencl and Soucek, 1976; Wickler, 1976; Morton and Gonzalez Alonso, 1982; Brown and Farabaugh, 1991). Others are less complex, but still highly variable with respect to duration and phrase composition (Mann et al., 2003; Logue, 2007b). Yet the simple, first-order duet code is the only experimentally demonstrated answering rule (but see Wright and Dahlin, 2007). In principle, the phrase-type used for answering could be influenced by factors other the mate's previous phrase-type, such as the position of the answer in the duet, or the sequence of prior phrases. It is not clear whether species that produce complex duets abide by complex answering rules, or alternatively, if intrinsic patterns of phrase-type switching combined with simple duet codes explain the structure of their duets. We also know little about how answering rates and answering rules are affected by the answer's position in the duet train.
Although adherence to duet codes is seldom complete (Mann et al., 2003; Mennill and Vehrencamp, 2005; Logue, 2007b), there has not been any published work on the reasons, functional, or otherwise, that adult birds deviate from their duet codes. One possibility is that they simply make mistakes. Another is that codes are more complex than we have realized, so apparent “deviations” are actually defects in the researcher's hypothesized coding rules. A tantalizing speculation is that answering rules include contingencies based on the communicative goal of answering. If answerers choose phrase-types to control the communicative effect of answering, a certain vocal gesture might, for example, request incubation relief or suggest a location for joint foraging. The presence, structure, or timing of answers could communicate the answerers' decision or preference (Flack, 2013).
Answering as a Model of Perception
Individuals' abilities to perceive and distinguish variation in timing and phrase structure influence interactions between duetting individuals, and thus the properties of whole duets. The predictability of rule-based answering suggests the value of duetting birds as models of perception and categorization. Playbacks of manipulated phrase stimuli could identify the structures birds require to identify song types (Sinnott et al., 1980; Weary, 1990) and the robustness of song classification in noise (Wiley and Richards, 1982; Guilford and Dawkins, 1991). For example, if a subject consistently responds to Stimulus X with Answer Y, one could systematically manipulate the properties of the stimulus to determine the structural space that the subject recognizes as X. Logue's (2007b) chimeric song playback is the only study that we know of to take this approach.
Discussion
Duetting behavior occurs at two levels of organization: the individual and the pair. Our thesis is that this distinction is foundational to developing a correct and complete understanding of duetting. It is also a reminder to think and write precisely. Vague terms, like the often-used “function of duetting,” should be replaced by precise ones, like “the function of song answering in males.” Students of collective behavior, the control of behavior, vocal interaction, and the evolution of social behavior have developed theory and research methods to address behavior across levels of organization. Duetting researchers can adopt and, where necessary, modify these ideas to apply them to vocal duetting systems. Not only will this approach advance duetting research, it will allow advances within duetting research to feed back into related fields. Such cross-fertilization has the potential to transform duetting research from a “niche market” into a more broadly relevant field of research.
The research framework and insights in this paper have applications outside of avian duetting research. Vocal chorusing involves all of the same behaviors as duetting (initiation, answering, phrase choice, and fine-scale timing), and so virtually all of the ideas that we discuss above may be applied to chorusing research. Duetting also has strong parallels with countersinging between neighboring territory holders. As Hall (2009) recognized, duetting and countersinging are both “interactive processes that involve time- and pattern-specific relationships among the exchanged signals” (p. 170; see also Todt and Naguib, 2000). Research on countersinging (e.g., song type matching, overlapping, communication networks) might benefit from many the ideas we have discussed. For example, agent-based models might be used to model communication networks, and the ontogeny of the rules that govern countersinging might be studied in both early life and as adult birds develop relationships with their neighbors.
The ideas discussed in this paper are also applicable to duetting and chorusing primates, such as gibbons (Geissmann, 2002), monkeys (Müller and Anzenberger, 2002), and lemurs (Baker-Médard et al., 2013). Of course, there is another group of primates that has evolved an extraordinarily complex form of interactive vocal communication. In spite of the obvious parallels, however, the study of conversation in humans has had minimal crossover with duetting research (Salwiczek and Wickler, 2004; Logue and Stivers, 2012; Ghazanfar and Takahashi, 2014). It is our hope that with increased attention to the levels of organization, research on vocal duetting will help us understand this most human form of collective behavior.
Author Contributions
DL conceived of the paper, wrote the first draft of everything but Insight #3, and edited the entire MS. DBK wrote the first draft of Insight #3, and edited the entire MS.
Funding
This work was funded in part by a Discovery Grant from the Natural Sciences and Engineering Research Council of Canada (RGPIN-2015-06553) to DL.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors thank Sergio Pellis for discussions about cybernetics. Chinthaka Kaluthota and Karan Odom offered valuable feedback on earlier versions of the manuscript.
References
Amador, A., Trevisan, M. A., and Mindlin, G. B. (2005). Simple neural substrate predicts complex rhythmic structure in duetting birds. Phys. Rev. E Stat. Nonlin Soft. Matter Phys. 72:031905. doi: 10.1103/PhysRevE.72.031905
Arrowood, P. C. (1988). Duetting, pair bonding and agonistic display in parakeet pairs. Behaviour 106, 129–157. doi: 10.1163/156853988X00133
Baker-Médard, M. S., Baker, M. C., and Logue, D. M. (2013). Chorus song of the indri (Primates, Lemuridae): group differences and analysis of within-group vocal interactions. Int. J. Comp. Psychol. 26, 241–255.
Bell, H. C. (2014). Behavioral variability in the service of constancy. Int. J. Comp. Psychol. 27, 338–360.
Benedict, L. (2007). Occurrence and life history correlates of vocal duetting in North American passerines. J. Avian Biol. 39, 57–65. doi: 10.1111/j.0908-8857.2008.04103.x
Benedict, L. (2010). California towhee vocal duets are multi-functional signals for multiple receivers. Behaviour 147, 953–978. doi: 10.1163/000579510X498633
Benedict, L., and McEntee, J. P. (2009). Context, structural variability and distinctiveness of California towhee (Pipilo crissalis) vocal duets. Ethology 115, 77–86. doi: 10.1111/j.1439-0310.2008.01583.x
Bradbury, J., and Vehrencamp, S. L. (2011). Principles of Animal Communication, 2nd Edn. Sunderland, MA: Sinauer.
Brown, E. D., and Farabaugh, S. M. (1991). Song sharing in a group-living songbird, the Australian magpie, Gymnorhina tibicen. Part III: sex specificity and individidual specificity of vocal parts in communal chorus and duet songs. Behaviour 118, 244–274. doi: 10.1163/156853991X00319
Brown, R. N., and Lemon, R. E. (1979). Structure and evolution of song form in the wrens Thryothorus sinaloa and T. felix. Behav. Ecol. Sociobiol. 5, 111–131. doi: 10.1007/BF00293301
Brumm, H., and Slater, P. (2007). Animal communication: timing counts. Curr. Biol. 17, R521–R523. doi: 10.1016/j.cub.2007.04.053
Catchpole, C. K., and Slater, P. J. B. (2003). Bird Song: Biological Themes and Variations. Cambridge, UK: Cambridge University Press.
Couzin, I. D. (2009). Collective cognition in animal groups. Trends Cogn. Sci. (Regul. Ed). 13, 36–43. doi: 10.1016/j.tics.2008.10.002
Couzin, I. D., and Krause, J. (2003). Self-organization and collective behavior in vertebrates. Adv. Study Behav. 32, 1–75. doi: 10.1016/S0065-3454(03)01001-5
Dahlin, C. R., and Benedict, L. (2014). Angry birds need not apply: a perspective on the flexible form and multifunctionality of avian vocal duets. Ethology 120, 1–10. doi: 10.1111/eth.12182
Diamond, J. M., and Terbourgh, J. W. (1968). Dual singing by New Guinea birds. Auk 85, 62–82. doi: 10.2307/4083625
Doupe, A. J., and Kuhl, P. K. (1999). Birdsong and human speech: common themes and mechanisms. Ann. Rev. Neurosci. 22, 567–631. doi: 10.1146/annurev.neuro.22.1.567
Elie, J. E., Mariette, M. M., Soula, H. A., Griffith, S. C., Mathevon, N., and Vignal, C. (2010). Vocal communication at the nest between mates in wild zebra finches: a private vocal duet? Anim. Behav. 80, 597–605. doi: 10.1016/j.anbehav.2010.06.003
Farabaugh, S. M. (1982). “The ecological and social significance of duetting,” in Acoustic Communication in Birds, Vol.2, eds D. E. Kroodsma and E. H. Miller (New York, NY: Academic Press), 85–124.
Farabaugh, S. M. (1983). A Comparative Study of Duet Song in Tropical Thryothorus Wrens. Ph.D. dissertation, University of Maryland, College Park, MD.
Fisher, S. E., and Scharff, C. (2009). FOXP2 as a molecular window into speech and language. Trends Genet. 25, 166–177. doi: 10.1016/j.tig.2009.03.002
Flack, J. C. (2013). Animal communication: hidden complexity. Curr. Biol. 23, R967–R969. doi: 10.1016/j.cub.2013.10.001
Fortune, E. S., Rodríguez, C., Li, D., Ball, G. F., and Coleman, M. J. (2011). Neural mechanisms for the coordination of duet singing in wrens. Science 334, 666–670. doi: 10.1126/science.1209867
Geissmann, T. (2002). Duet-splitting and the evolution of gibbon songs. Biol. Rev. 77, 57–76. doi: 10.1017/S1464793101005826
Ghazanfar, A. A., and Takahashi, D. Y. (2014). The evolution of speech: vision, rhythm, cooperation. Trends Cogn. Sci. (Regul. Ed). 18, 543–553. doi: 10.1016/j.tics.2014.06.004
Gill, S. A., Vonhof, M. J., Stutchbury, B. J. M., Morton, E. S., and Quinn, J. S. (2005). No evidence for acoustic mate-guarding in duetting buff-breasted wrens (Thryothorus leucotis). Behav. Ecol. Sociobiol. 57, 557–565. doi: 10.1007/s00265-004-0893-3
Goldstein, J. (1999). Emergence as a construct: history and issues. Emergence 1, 49–72. doi: 10.1207/s15327000em0101_4
Goldstone, R. L., and Janssen, M. A. (2005). Computational models of collective behavior. Trends Cogn. Sci. (Regul. Ed). 9, 424–430. doi: 10.1016/j.tics.2005.07.009
Grafe, T. U., and Bitz, J. H. (2004). Functions of duetting in the tropical boubou, Laniarius aethiopicus: territorial defence and mutual mate guarding. Anim. Behav. 68, 193–201. doi: 10.1016/j.anbehav.2003.11.003
Guilford, T., and Dawkins, M. S. (1991). Receiver psychology and the evolution of animal signals. Anim. Behav. 42, 1–14. doi: 10.1016/S0003-3472(05)80600-1
Hall, M. L. (2004). A review of hypotheses for the functions of avian duetting. Behav. Ecol. Sociobiol. 55, 415–430. doi: 10.1007/s00265-003-0741-x
Hall, M. L. (2006). Convergent vocal strategies of males and females are consistent with a cooperative function of duetting in Australian magpie-larks. Behaviour 143, 425–449. doi: 10.1163/156853906776240623
Hall, M. L. (2009). A review of vocal duetting in birds. Adv. Study Behav. 40, 67–121. doi: 10.1016/s0065-3454(09)40003-2
Hall, M. L., and Magrath, R. D. (2007). Temporal coordination signals coalition quality. Curr. Biol. 17, R406–R407. doi: 10.1016/j.cub.2007.04.022
Hasson, U., Ghazanfar, A. A., Galantucci, B., Garrod, S., and Keysers, C. (2012). Brain-to-brain coupling: a mechanism for creating and sharing a social world. Trends Cogn. Sci. 16, 114–121. doi: 10.1016/j.tics.2011.12.007
Illes, A. E. (2015). Context of female bias in song repertoire size, singing effort, and singing independence in a cooperatively breeding songbird. Behav. Ecol. Sociobiol. 69, 139–150. doi: 10.1007/s00265-014-1827-3
King, A. J., and Sumpter, D. J. (2012). Murmurations. Curr. Biol. 22, R112–R114. doi: 10.1016/j.cub.2011.11.033
Krupp, D. B. (in press). Causality the levels of selection. Trends Ecol. Evol. doi: 10.1016/j.tree.2016.01.008.
Laje, R., and Mindlin, G. (2003). Highly structured duets in the song of the South American hornero. Phys. Rev. Lett. 91:258104. doi: 10.1103/PhysRevLett.91.258104
Langmore, N. E. (1998). Functions of duet and solo songs of female birds. Trends Ecol. Evol. 13, 136–140. doi: 10.1016/S0169-5347(97)01241-X
Levin, R. N. (1996a). Song behaviour and reproductive strategies in a duetting wren, Thryothorus nigricapillus: I. Removal experiments. Anim. Behav. 52, 1093–1106. doi: 10.1006/anbe.1996.0257
Levin, R. N. (1996b). Song behaviour and reproductive strategies in a duetting wren, Thryothorus nigricapillus: II. Playback experiments. Anim. Behav. 52, 1107–1117. doi: 10.1006/anbe.1996.0258
Levin, R. N., Paris, T. I., and Bester, J. K. (1996). Social versus innate influences on the development of sex-specific song in a tropical duetting wren. Am. Zool. 36, 92A.
Lipkind, D., Marcus, G. F., Bemis, D. K., Sasahara, K., Jacoby, N., Takahasi, M., et al. (2013). Stepwise acquisition of vocal combinatorial capacity in songbirds and human infants. Nature 498, 104–108. doi: 10.1038/nature12173
Logue, D. M. (2006). The duet code of the female black-bellied wren. Condor 108, 326–335. doi: 10.1650/0010-5422(2006)108[326:TDCOTF]2.0.CO;2
Logue, D. M. (2007a). Duetting in space: a radio-telemetry study of the black-bellied wren. Proc. R. Soc. B Biol. Sci. 274, 3005–3010. doi: 10.1098/rspb.2007.1005
Logue, D. M. (2007b). How do they duet? Sexually dimorphic behavioural mechanisms structure duet songs in the black-bellied wren. Anim. Behav. 73, 105–113. doi: 10.1016/j.anbehav.2006.05.011
Logue, D. M., Chalmers, C., and Gowland, A. H. (2008). The behavioural mechanisms underlying temporal coordination in black-bellied wren duets. Anim. Behav. 75, 1803–1808. doi: 10.1016/j.anbehav.2007.10.036
Logue, D. M., and Gammon, D. E. (2004). Duet song and sex roles during territory defence in a tropical bird, the black-bellied wren, Thryothorus fasciatoventris. Anim. Behav. 68, 721–731. doi: 10.1016/j.anbehav.2003.10.026
Logue, D. M., and Hall, M. L. (2014). Migration and the evolution of duetting in songbirds. Proc. R. Soc. B Biol. Sci. 281:20140103. doi: 10.1098/rspb.2014.0103
Logue, D. M., and Stivers, T. (2012). Squawk in interaction: a primer of conversation analysis for students of animal communication. Behaviour 149, 1283–1298. doi: 10.1163/1568539X-00003031
Malacarne, G., Cucco, M., and Camanni, S. (1991). Coordinated visual displays and vocal duetting in different ecological situations among Western Palearctic non-passerine birds. Ethol. Ecol. Evol. 3, 207–219. doi: 10.1080/08927014.1991.9525369
Mank, J. E., Promislow, D. E., Avise, J. C., and Yoder, A. (2005). Phylogenetic perspectives in the evolution of parental care in ray-finned fishes. Evolution 59, 1570–1578. doi: 10.1111/j.0014-3820.2005.tb01806.x
Mann, N. I., Dingess, K. A., and Slater, P. J. (2006). Antiphonal four-part synchronized chorusing in a Neotropical wren. Biol. Lett. 2, 1–4. doi: 10.1098/rsbl.2005.0373
Mann, N. I., Dingess, K., Slater, P., Graves, J., and Barker, K. (2009). A comparative study of song form and duetting in neotropical Thryothorus wrens. Behaviour 146, 1–43. doi: 10.1163/156853908X390913
Mann, N. I., Marshall-Ball, L., and Slater, P. J. B. (2003). The complex song duet of the plain wren. Condor 105, 672–682. doi: 10.1650/7208
Marshall-Ball, L., Mann, N. I., and Slater, P. J. B. (2006). Multiple functions to duet singing: hidden conflicts and apparent cooperation. Anim. Behav. 71, 823–831. doi: 10.1016/j.anbehav.2005.05.021
Marshall-Ball, L., and Slater, P. J. (2004). Duet singing and repertoire use in threat signalling of individuals and pairs. Biol. Lett. 271(Suppl. 6), S440–S443. doi: 10.1098/rsbl.2004.0186
Mennill, D. J. (2006). Aggressive responses of male and female rufous-and-white wrens to stereo duet playback. Anim. Behav. 71, 219–226. doi: 10.1016/j.anbehav.2005.05.006
Mennill, D. J., and Vehrencamp, S. L. (2005). Sex differences in singing and duetting behavior of neotropical rufous-and-white wrens (Thryothorus rufalbus). Auk 122, 175–186. doi: 10.1642/0004-8038(2005)122[0175:SDISAD]2.0.CO;2
Morton, E. S., and Gonzalez Alonso, H. J. (1982). The biology of the Torreornis inexpectata I. A comparison of vocalizations in T.i. inexpectata and T.i. sigmani. Wilson Bull. 94, 433–446.
Müller, A. E., and Anzenberger, G. (2002). Duetting in the titi monkey Callicebus cupreus: structure, pair specificity and development of duets. Folia Primatol. 73, 104–115. doi: 10.1159/000064788
Okasha, S. (2012). Emergence, hierarchy and top-down causation in evolutionary biology. Interface Focus. 2, 49–54. doi: 10.1098/rsfs.2011.0046
Okasha, S. (in press). The relation between kin multilevel selection: an approach using causal graphs. Br. J. Philos. Sci. axu047. doi: 10.1093/bjps/axu047.
Owren, M. J., Rendall, D., and Ryan, M. J. (2010). Redefining animal signaling: influence versus information in communication. Biol. Philos. 25, 755–780. doi: 10.1007/s10539-010-9224-4
Pellis, S. M., and Pellis, V. C. (2015). Are agonistic behavior patterns signals or combat tactics—Or does it matter? Targets as organizing principles of fighting. Physiol. Behav. 146, 73–78. doi: 10.1016/j.physbeh.2015.03.038
Pfenning, A. R., Hara, E., Whitney, O., Rivas, M. V., Wang, R., Roulhac, P. L., et al. (2014). Convergent transcriptional specializations in the brains of humans and song-learning birds. Science 346, 1333. doi: 10.1126/science.1256846
Price, J. J., Lanyon, S. M., and Omland, K. E. (2009). Losses of female song with changes from tropical to temperate breeding in the New World blackbirds. Proc. R. Soc. B Biol. Sci. 276, 1971–1980. doi: 10.1098/rspb.2008.1626
Queller, D. C. (1984). Kin selection and frequency dependence: a game theoretic approach. Biol. J. Linnean Soc. 23, 133–143. doi: 10.1111/j.1095-8312.1984.tb00133.x
Queller, D. C. (1985). Kinship, reciprocity and synergism in the evolution of social behaviour. Nature 318, 366–367. doi: 10.1038/318366a0
Riegert, J., Pribylova, A., and Sedlacek, O. (2004). Notes on calls of the Yellow-breasted Boubou (Laniarius atroflavus) during the breeding season in NW Cameroon. Biol. Lett. 41, 181–184.
Rivera-Cáceres, K. D. (2015). Plain wrens Cantorchilus modestus zeledoni adjust their singing tempo based on self and partner's cues to perform precisely coordinated duets. J. Avian Biol. 46, 361–368. doi: 10.1111/jav.00575
Rivera-Cáceres, K. D., and Guerrero Esmeralda, Q. (2014). “Duet coordination and duet code in juvenile plain wrens,” in Poster Presented at the Meeting of the Animal Behavior Society (Princeton, NJ).
Rogers, A. C. (2005). Male and female song structure and singing behaviour in the duetting eastern whipbird, Psophodes olivaceus. Aust. J. Zool. 53, 157–166. doi: 10.1071/ZO04083
Rogers, A. C., Mulder, R. A., and Langmore, N. E. (2006). Duet duels: sex differences in song matching in duetting eastern whipbirds. Anim. Behav. 72, 53–61. doi: 10.1016/j.anbehav.2005.08.019
Salwiczek, L. H., and Wickler, W. (2004). Birdsong: an evolutionary parallel to human language. Semiotica 151, 163–182. doi: 10.1515/semi.2004.066
Schank, J. C., and Alberts, J. R. (1997). Self-organized huddles in rat pups modeled by simple rules of individual behavior. J. Theor. Biol. 189, 11–25. doi: 10.1006/jtbi.1997.0488
Sinnott, J. M., Sachs, M. B., and Hienz, R. D. (1980). Aspects of frequency discrimination in passerine birds and pigeons. J. Comp. Physiol. Psychol. 94, 401. doi: 10.1037/h0077681
Sonnenschein, E., and Reyer, H.-U. (1983). Mate-guarding and other functions of antiphonal duets in the slate-coloured boubou (Laniarius funebris) 1. Z. Tierpsychol. 63, 112–140. doi: 10.1111/j.1439-0310.1983.tb00083.x
Takahashi, D. Y., Narayanan, D. Z., and Ghazanfar, A. A. (2013). Coupled oscillator dynamics of vocal turn-taking in monkeys. Curr. Biol. 23, 2162–2168. doi: 10.1016/j.cub.2013.09.005
Taylor, P. (2013). Inclusive and personal fitness in synergistic evolutionary games on graphs. J. Theor. Biol. 325, 76–82. doi: 10.1016/j.jtbi.2013.02.002
Taylor, P., and Maciejewski, W. (2012). An inclusive fitness analysis of synergistic interactions in structured populations. Proc. R. Soc. B Biol. Sci. 279:20121408. doi: 10.1098/rspb.2012.1408
Templeton, C. N., Mann, N. I., Ríos-Chelén, A. A., Quiros-Guerrero, E., Macías Garcia, C., and Slater, P. J. B. (2013). An experimental study of duet integration in the happy wren, Pheugopedius felix. Anim. Behav. 86, 821–827. doi: 10.1016/j.anbehav.2013.07.022
Thorpe, W. H. (1963). Antiphonal singing in birds as evidence for avian auditory reaction time. Nature 197, 774–776. doi: 10.1038/197774a0
Tobias, J. A., and Seddon, N. (2009). Signal jamming mediates sexual conflict in a duetting bird. Curr. Biol. 19, 577–582. doi: 10.1016/j.cub.2009.02.036
Todt, D., and Naguib, M. (2000). Vocal interactions in birds: the use of song as a model in communication. Adv. Study Behav. 29, 247–296. doi: 10.1016/S0065-3454(08)60107-2
Topp, S. M., and Mennill, D. J. (2008). Seasonal variation in the duetting behaviour of rufous-and-white wrens (Thryothorus rufalbus). Behav. Ecol. Sociobiol. 62, 1107–1117. doi: 10.1007/s00265-007-0538-4
Trainer, J. M., McDonald, D. B., and Learn, W. A. (2002). The development of coordinated singing in cooperatively displaying long-tailed manakins. Behav. Ecol. 13, 65–69. doi: 10.1093/beheco/13.1.65
Valderrama, S. V., Molles, L. E., and Waas, J. R. (2013). Effects of population size on singing behavior of a rare duetting songbird. Conserv. Biol. 27, 210–218. doi: 10.1111/j.1523-1739.2012.01917.x
van den Heuvel, I. M., Cherry, M. I., and Klump, G. M. (2014). Crimson-breasted Shrike females with extra pair offspring contributed more to duets. Behav. Ecol. Sociobiol. 68, 1245–1252. doi: 10.1007/s00265-014-1735-6
Vencl, F. V., and Soucek, B. (1976). Structure and control of duet singing in the white-crested laughing thrush (Garrulax leucolophus). Behaviour 57, 206–225. doi: 10.1163/156853976X00523
Voigt, C., Leitner, S., and Gahr, M. (2006). Repertoire and structure of duet and solo songs in cooperatively breeding white-browed sparrow weavers. Behaviour 143, 159–182. doi: 10.1163/156853906775900739
von Helversen, V. D., and Wickler, W. (1971). Uber den Duettgesang des afrikanischen Drongo Dicrurus adsimilis Bechstein. Z. Tierpsychol. 29, 301–321. doi: 10.1111/j.1439-0310.1971.tb01739.x
Weary, D. M. (1990). Categorization of song notes in great tits: which acoustic features are used and why? Anim. Behav. 39, 450–457. doi: 10.1016/S0003-3472(05)80408-7
Wickler, W. (1976). The ethological analysis of attachment. Z. Tierpsychol. 42, 12–28. doi: 10.1111/j.1439-0310.1976.tb00953.x
Wickler, W. (1980). Vocal duetting and the pair bond. I. Coyness and partner commitment. A hypothesis. Z. Tierpsychol. 52, 201–209. doi: 10.1111/j.1439-0310.1980.tb00711.x
Wickler, W., and Sonnenschein, E. (1989). Ontogeny of song in captive duet-singing slate-coloured boubous (Laniarius funebris). A study of birdsong epigenesis. Behaviour 111, 220–233. doi: 10.1163/156853989X00673
Wiley, R. H., and Richards, D. G. (1982). “Adaptations for acoustic communication in birds: sound transmission and signal detection,” in Acoustic Communication in Birds, Vol. 1, eds D. E. Kroodsma and E. H. Miller (Ithica, NY: Cornell University Press), 131–181. doi: 10.1016/B978-0-08-092416-8.50014-0
Keywords: animal communication, chorus, conversation, countersinging, cybernetics, causality, female song, levels of organization
Citation: Logue DM and Krupp DB (2016) Duetting as a Collective Behavior. Front. Ecol. Evol. 4:7. doi: 10.3389/fevo.2016.00007
Received: 26 October 2015; Accepted: 18 January 2016;
Published: 05 February 2016.
Edited by:
Naomi Langmore, Australian National University, AustraliaReviewed by:
James Dale, Massey University, New ZealandLauryn Benedict, University of Northern Colorado, USA
Copyright © 2016 Logue and Krupp. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David M. Logue, ZGF2aWQubG9ndWVAdWxldGguY2E=