 Charline Ragon1
Charline Ragon1 Christian Vérard2
Christian Vérard2 Jérôme Kasparian1
Jérôme Kasparian1 Hendrik Nowak3,4
Hendrik Nowak3,4 Evelyn Kustatscher4
Evelyn Kustatscher4 Maura Brunetti1*
Maura Brunetti1*- 1Group of Applied Physics and Institute for Environmental Sciences, University of Geneva, Geneva, Switzerland
- 2Section of Earth and Environmental Sciences, University of Geneva, Geneva, Switzerland
- 3School of Biosciences, University of Nottingham, Nottingham, United Kingdom
- 4Museum of Nature South Tyrol, Bolzano, Italy
Terrestrial ecosystems underwent extreme shifts in composition, following extensive degassing associated with the Siberian Traps near the Permian–Triassic boundary (PTB). These climatic perturbations are recorded in land plant macrofossil assemblages, which reflect complex changes in major biomes at the stage level. In this study, we quantitatively compare the major biomes reconstructed from the plant macrofossil assemblage data with those derived from coupled climate–vegetation simulations across the PTB. We focus on five stages across the PTB, from the Wuchiapingian to the Anisian. Our findings indicate that a shift from a cold climatic state to one with a mean surface temperature approximately
1 Introduction
The Permian–Triassic boundary (PTB) mass extinction occurred ca. 252 million years ago and was marked by the most severe biotic crisis in the Phanerozoic (Raup and Sepkoski, 1982; Stanley, 2016). Extreme reductions in both marine and terrestrial animals were recorded, most likely caused by extensive degassing associated with the Siberian Traps (Renne and Basu, 1991; Renne et al., 1995; Sibik et al., 2015; Burgess et al., 2017; Svensen et al., 2018; Davydov, 2021; Callegaro et al., 2021). In the PTB aftermath, life on Earth had to drastically adjust to repeated changes in climate and the carbon cycle for several million years. This crisis in the faunal realm was coeval to complex shifts in composition in terrestrial ecosystems that were not limited to a single event (Looy et al., 2001; Hochuli et al., 2016; Schneebeli-Hermann, 2020). Recent studies show that land plant macro- and microfossil (spores and pollen) records of the Early Triassic do not provide strong evidence for a sudden and catastrophic biodiversity loss coeval with the faunal diversity loss at the PTB (Nowak et al., 2019). Nonetheless, major changes in regional and global environmental conditions and flora composition occurred throughout the Early Triassic (Hochuli et al., 2016; Fielding et al., 2019; Schneebeli-Hermann, 2020; Mays et al., 2021; Mays and McLoughlin, 2022). This includes the abrupt extirpation of the primary coal-forming carbon sinks, such as the Glossopteris biome of Gondwana (Mays et al., 2019; Vajda et al., 2020; Mays et al., 2021) and the tropical gigantopterid forests of East Asia at the end of the Permian (Chu et al., 2020). Shifts between a lycophyte-dominated (spore-producing vascular plants) and a gymnosperm-dominated (seed-producing plants, including conifers) vegetation coincided, respectively, with a succession of warm and cold climatic conditions (Galfetti et al., 2007b; Schneebeli-Hermann, 2020). Changes from gymnosperm- to lycophyte-dominated vegetation, as recorded in palynomorph assemblages from subtropical locations, occurred at the PTB and during the earliest Triassic Induan stage (at the Griesbachian–Dienerian substage boundary), whereas the shift from lycophyte- to gymnosperm-dominated vegetation occurred in the subsequent Olenekian stage (at the middle-late Smithian boundary), with transient regimes observed before and after this transition. Spores and pollen from higher latitudes record similar changes in relative abundance during the Early Triassic (Hochuli et al., 2010; Hochuli et al., 2016).
The macrofossil assemblages described by Nowak et al. (2019) were later used to reconstruct major biomes (Nowak et al., 2020), defined as areas with comparable, climatically controlled plant and animal assemblages (Ziegler, 1990; Walter, 2012). A reduction in biome diversity from the Permian to the Early Triassic was observed and was associated with a climate shift characterized by an increase in seasonality. This period of high variability was followed by a stable phase that lasted through the Middle Triassic, with comparable biomes from the Olenekian to the Ladinian. Microfossils were excluded from the reconstruction of the biome distribution as their source areas can be vast (regional) and not representative of the depositional site (Nowak et al., 2020).
The analysis provided by Nowak et al. (2020) was performed with a temporal resolution at the stage level from the Wuchiapingian to the Ladinian. In the present paper, we will expand upon this analysis using climate simulations obtained by the offline coupling between a general circulation model (MITgcm) and a vegetation model (BIOME4). The climate simulations were performed using the paleogeographic configuration provided by PANALESIS (Vérard, 2019; Vérard, 2021) for the PTB, as described by Ragon et al. (2024). Interestingly, three alternative climatic steady states (denoted as cold, warm, and hot attractors) have been found for the same boundary conditions, with mean surface air temperatures (SATs) ranging from
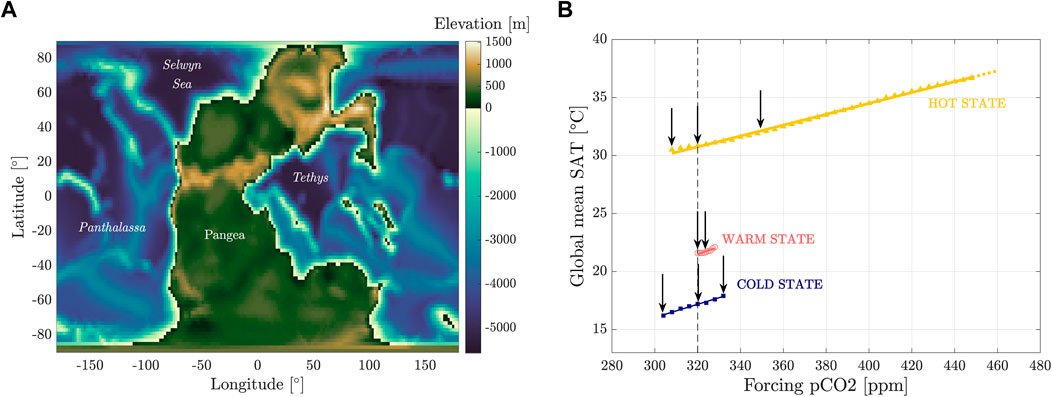
Figure 1. Permian–Triassic paleogeography (A) and corresponding bifurcation diagram (B) in terms of the equilibrium values of the global mean surface air temperature vs. the atmospheric
The presence of alternative attractors and the structure of the bifurcation diagram in Figure 1 suggest a potential explanation for shifts in composition observed in terrestrial ecosystems across the PTB as shifts between attractors. These transitions may have induced strong climatic variations in both atmospheric and oceanic circulations, impacting the whole climate system (Ragon et al., 2024; Rogger et al., 2024). Perturbations of the carbon cycle, as a consequence of the outgassing associated with the Siberian Traps, may have triggered not only bifurcation-induced tipping between the cold and hot states, with repeated activation of the hysteresis loop between these two states, but also noise- or rate-induced tipping between the three attractors (Ashwin et al., 2012; Brunetti and Ragon, 2023; Feudel, 2023).
The aim of this paper is to assess whether the changes in global vegetation patterns, as recorded in land–plant macrofossil assemblages at the stage level and presented by Nowak et al. (2020), can be explained by tipping between the simulated climatic attractors. We will quantitatively compare the modeled major vegetational biomes for the hot, warm, and cold states with the land–plant macrofossil assemblages compiled by Nowak et al. (2019) and Nowak et al. (2020), spanning from the Lopingian (starting at 259.51 Ma and including the Wuchiapingian and the Changhsingian) to the early Middle Triassic (
2 Methods
2.1 Paleogeographic reconstruction
The Permian–Triassic paleogeography is derived from PANALESIS (Vérard, 2019; Vérard, 2021), a global plate tectonic model providing maps every 10 Myr from 888 Ma (Tonian) to the present. The PANALESIS paleogeography for the PTB, which we use as a fixed boundary condition in our climate simulations, has proven to be in good agreement with geochemical and paleontological records (Chablais et al., 2011; Peyrotty et al., 2020; Bucur et al., 2020; Le Houedec et al., 2024), particularly in locating elements within the intertropical zone. The location of island arcs, continental ribbons, and even parts of Pangea, such as South China, remains, however, subject to uncertainties, with latitudes potentially varying up to ca.
2.2 Plant fossil records
The study by Nowak et al. (2020) was based on a dataset of plant macrofossil assemblages from Nowak et al. (2019), which is a compilation of previously published and unpublished plant fossil collections. This dataset is used in the present paper to determine major biomes at the stage level, spanning from the Wuchiapingian to the Anisian. Each plant genus is associated with the major biome(s) it could potentially occur in, possibly exclusively (i.e., when a plant genus is known to be limited to habitats aligned with a certain biome; see Supplementary Table S2 in Nowak et al. (2020) for details).
The records are aggregated within a moving 100-km radius, where each genus is assigned a weight corresponding to the inverse of the number of major biome(s) it is associated with. For each aggregated assemblage, a vote for all possible biomes is then tallied based on the weights of all present genera. The final major biome is determined based on either i) the exclusive major biome if a characteristic representative is present, ii) the major biome whose combined weight is
The classification of major biomes used in this study comprises six categories, which are mostly adapted from those in Nowak et al. (2020), which, in turn, followed the set of biomes introduced by Ziegler (1990) as far as they could be applied to the fossil dataset at hand. The resulting simplified major biomes are briefly described as follows: the tropical everwet major biome includes various vegetation types developing under constantly hot and humid conditions, typically near the equator, but it can extend up to mid-latitudes. The tropical summerwet major biome represents intermediate vegetation between tropical everwet and desert, found in middle to low latitudes with marked seasonality and wet summers. The desert major biome includes both subtropical and mid-latitude deserts but is generalized to all latitudes in this study and is marked by water deficiency. The warm-to-cool temperate major biome encompasses vegetation ranging from evergreen (i.e., plants that keep their needles or leaves all year) to deciduous (i.e., plants that shed their leaves in autumn), affected by seasonal changes in climatic conditions. The cold temperate major biome is associated with areas with low evaporation, where the short growing season is mainly controlled by temperature and sunshine. The tundra major biome has a very short growing season and was not recorded by Nowak et al. (2020).
2.3 Vegetation distribution along the stable branches
The simulated vegetation distribution is different between the three attractors but also varies along each stable branch, together with the atmospheric
The biomes resulting from the simulations (28 biomes; see Kaplan, 2001) are grouped in the same major biomes described in Section 2.2 to facilitate the comparison with the geological records provided by Nowak et al. (2020) and eliminate perturbations coming from small differences between similar biomes. The corresponding classification is described in Table 1.
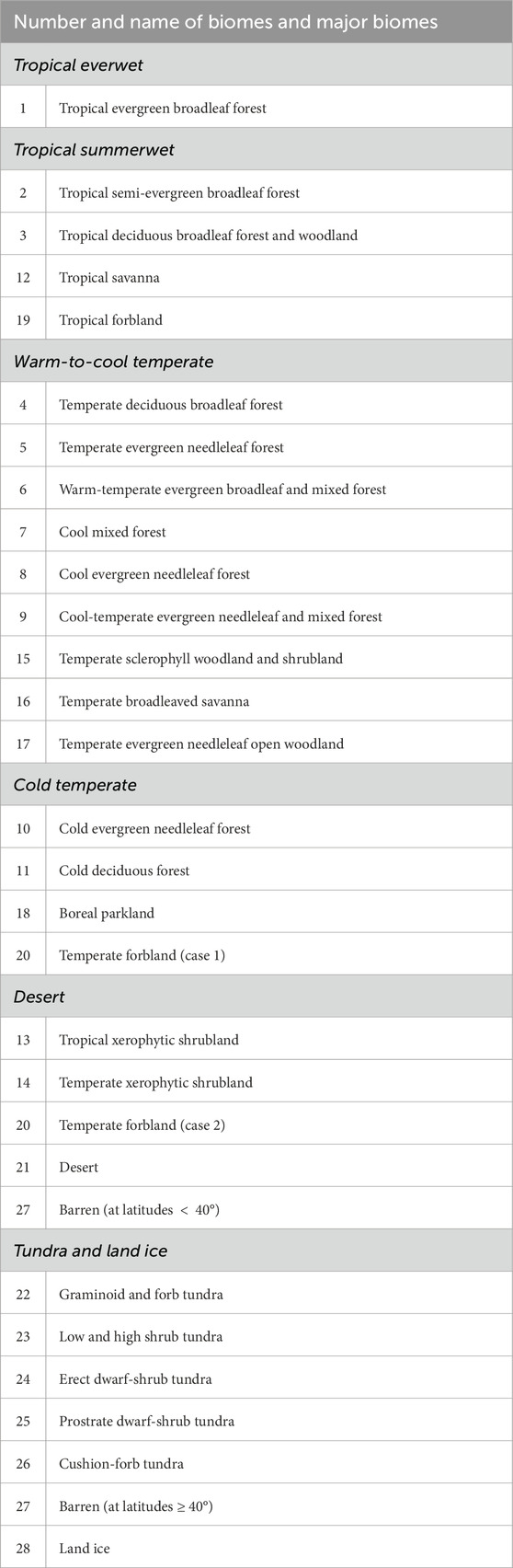
Table 1. Correspondence between the 28 biomes represented in the BIOME 4 model and the major biomes adapted from Nowak et al. (2020) (in italics). Cases 1 and 2 refer to alternative classifications of the temperate forbland biome.
The present-day “grassland” is replaced by the herbaceous non-graminoid “forbland” because graminoids only appeared during the Cretaceous–Cenozoic (Gradstein and Kerp, 2012). There is consequently no direct equivalence between simulated grasslands and any fossil assemblage or fossil-based biome from the Permian and Triassic, but we can infer likely correspondences based on the climatic conditions they represent. Both tropical savanna and tropical forbland biomes have been included in the tropical summerwet major biome. Even if they slightly differ from the broadleaf forest category, their seasonality has been considered the main argument for their classification. The classification of barren, a desertic biome mainly associated with polar regions in present-day vegetation, is distinguished by the latitude of formation: the desert major biome is found below 40
The criteria for temperate forbland biome in BIOME 4 correspond to desert-like conditions, more than everwet or seasonally wet. However, our simulations show that this biome is mostly formed in high latitudes,
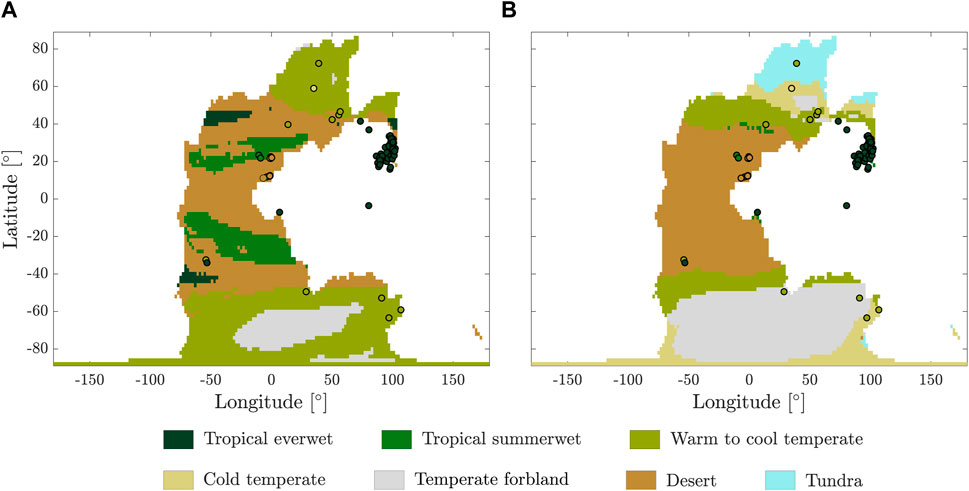
Figure 2. Comparison of plant fossil assemblages of Wuchiapingian age (259.51–254.14 Ma) superposed with major biomes modeled at 320 ppm for the (A) hot state and (B) cold state.
2.4 Similarity between geological records and modeled biomes
The paleontological record provides local information on vegetation, while the model simulates broader areas, posing a challenge for the direct comparison between the two. Visual comparisons (see example in Figure 2) can offer valuable insights into how well the simulation aligns with the records; however, this approach is not quantitative and does not always provide a clear answer.
Each plant fossil assemblage record is assigned to the model-grid cell corresponding to its location. Although the single outcrops are considered independent, their spatial resolution is sometimes higher than that of the model [i.e., records fall within the same grid cell of
The similarity between the plant macrofossil record at a given stage and the simulated vegetation distribution of an attractor is estimated as follows. For each reported assemblage of fossil plants, we compute the smallest geodesic distance
where
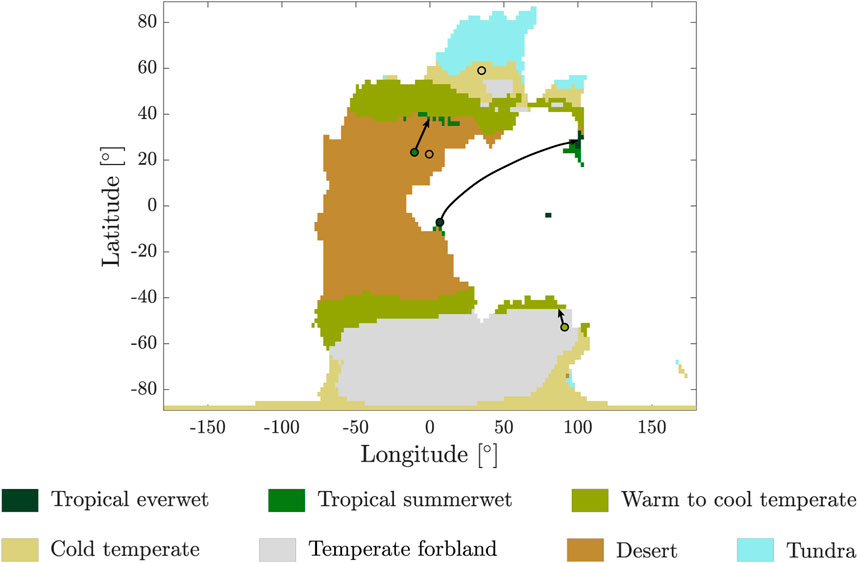
Figure 3. Example of the calculation of the smallest geodesic distance between geological records (circles with black contours) and the nearest region where the same major biome has been predicted by the simulation.
We define the weighted mean distance
where
2.5 Statistical tests
The distances
The utilization of several methods (on means and medians), together with the classification of the temperate forbland biome into two different major biomes (cases 1 and 2 in Table 1), allows us to test the robustness of the results (see also Supplementary Appendix SB).
2.5.1 Testing difference between means:
We assume that the distributions of
where
where Z0 is given in Equation 4 and
2.5.2 Testing difference between medians: Mood-test
The
The Mood-test is applied to determine whether the difference in the median value of the distances
A 5% threshold is used, as for the
3 Results
3.1 Simulation maps
The simulated distribution of the major biomes corresponding to the attractors at various atmospheric
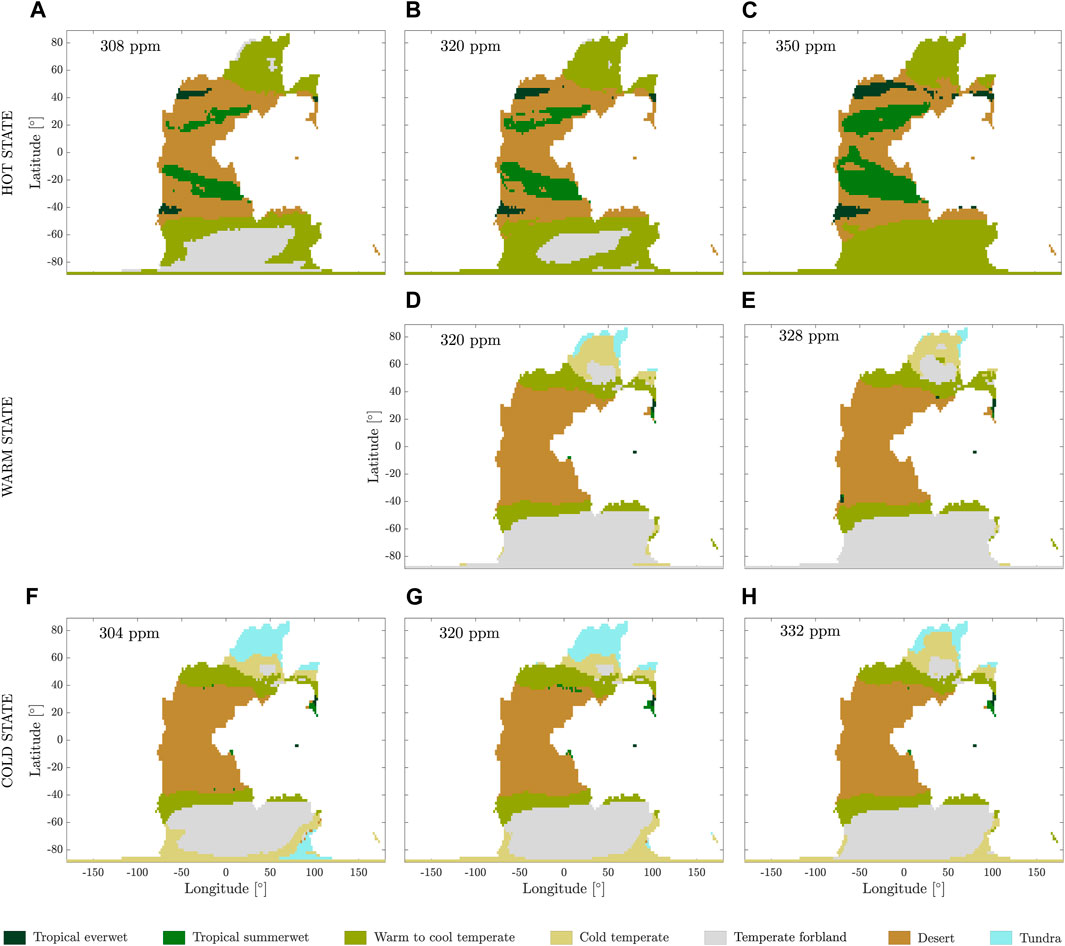
Figure 4. Distribution of the major biomes (see Table 1) for (A–C) hot, (D, E) warm, and (F–H) cold states, with the atmospheric
The simulated major biomes in the cold and warm states (Figures 4D–H) are both dominated by desert at tropical and subtropical latitudes. Along 60
In the hot state, the warm-to-cool temperate major biome is also present but shifted poleward compared to the other attractors, so it dominates in the northern polar region. The southern polar region is occupied by both the warm-to-cool temperate major biome and temperate forbland, with the extent of the latter reducing as the atmospheric
3.2 Test on the mean values
Figure 5 shows, at the five considered stages, the mean distance
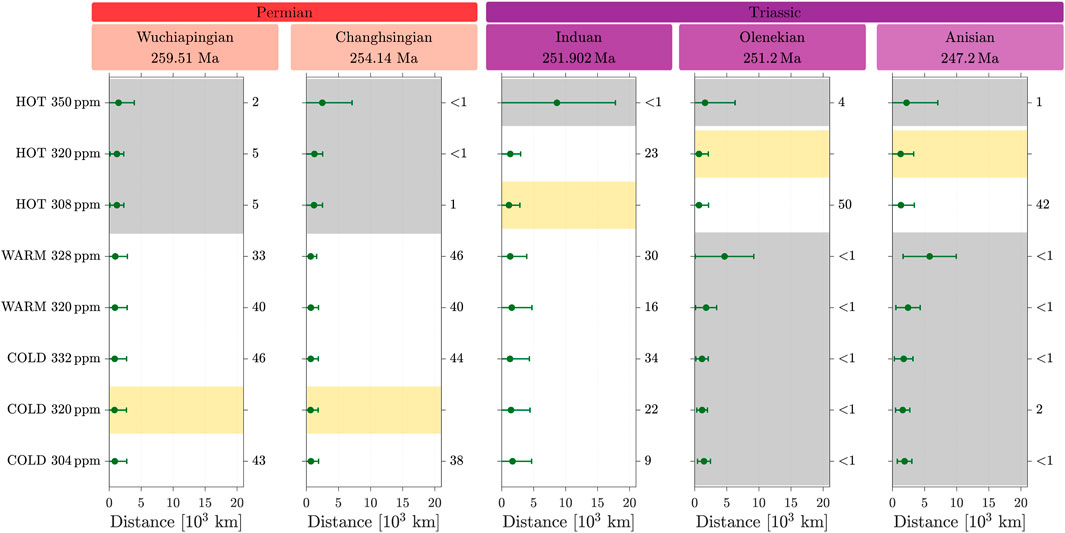
Figure 5. Weighted-mean and standard deviation of the distances computed between each of the eight simulations and geological records for the five stages. For each stage, the yellow area indicates the simulation minimizing the mean distance. The numbers on the right correspond to the
For the two oldest stages, the Wuchiapingian and the Changhsingian (Lopingian), the vegetation distribution modeled for the cold state at 320 ppm provides the best match with the fossil records. However, it is not significantly different from the vegetation distribution modeled at other positions on the cold branch or from the warm state. In contrast, the vegetation simulated in the hot state is significantly different, and thus, it is not a good candidate to reproduce the records of these stages.
In the case of the Induan, the hot state at 308 ppm minimizes
These results suggest a transition from the cold or warm state in the Lopingian to the hot state in the Olenekian, in concordance with the global warming recorded from the Lopingian to the Early Triassic (Retallack, 1999; Joachimski et al., 2012). The hot state persists during the Anisian, thus marking the stabilization of the climate system, as recorded, for example, in
3.3 Test on the median values
The test on medians shows a trend comparable to what is observed for that on means, as shown in Figure 6; Supplementary Table S2. For both the Wuchiapingian and the Changhsingian, the cold state, not statistically different along the branch or from the warm state, has a smaller median and thus matches better with the data, while the hot state has a significantly larger median and thus can be excluded.
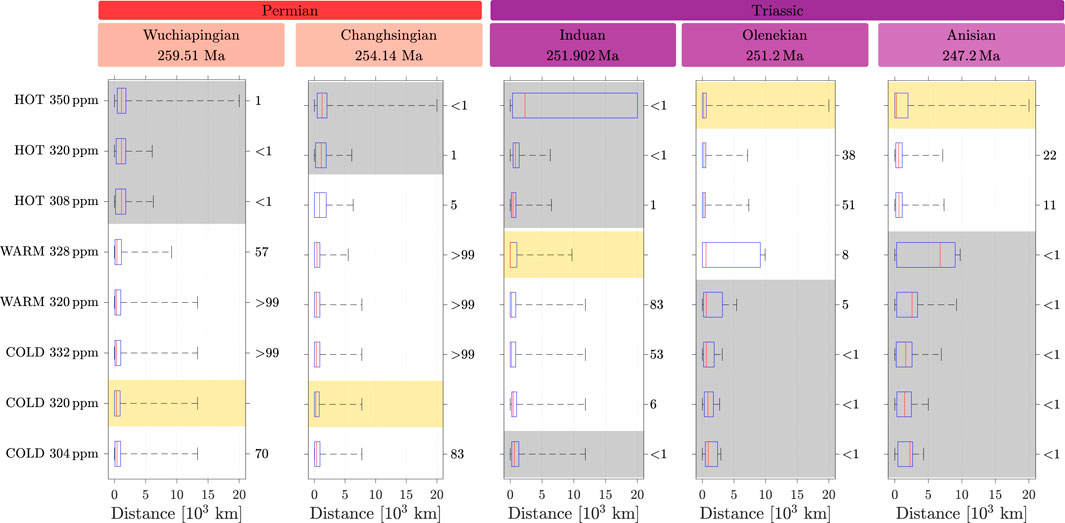
Figure 6. Whisker plot of the distances computed between each of the eight simulations and geological records for the five stages. For each stage, the yellow area indicates the simulation minimizing the median distance. The numbers on the right correspond to the
For the Induan, the median in the warm-state simulation with 328 ppm is lower than that for the other simulations and is statistically different from the whole hot state and the lower edge of the cold-state branch.
For the Olenekian and the Anisian, the hot state at 350 ppm minimizes the median but is not significantly different from the same attractor with other atmospheric content of
3.4 Robustness of the results against the classification of the temperate forbland biome
The classification of temperate forbland into cold temperate is questionable, as discussed in Section 2.3. In this study, we test the robustness of the results against the classification of this biome by including it in the desert major biome (case 2) instead of cold temperate (case 1).
3.4.1 Case 2: test on the mean values
The statistical analysis of the mean values is shown in Figure 7; Supplementary Table S3. The Wuchiapingian, Changhsingian, and Induan stages show better alignment between the geological records and cold and warm attractors. This tendency was already observed in the Lopingian, with the earlier classification of temperate forbland in the cold temperate major biome (case 1). However, the Induan did not display a clear tendency in favor of the hot or cold state in the previous analysis.
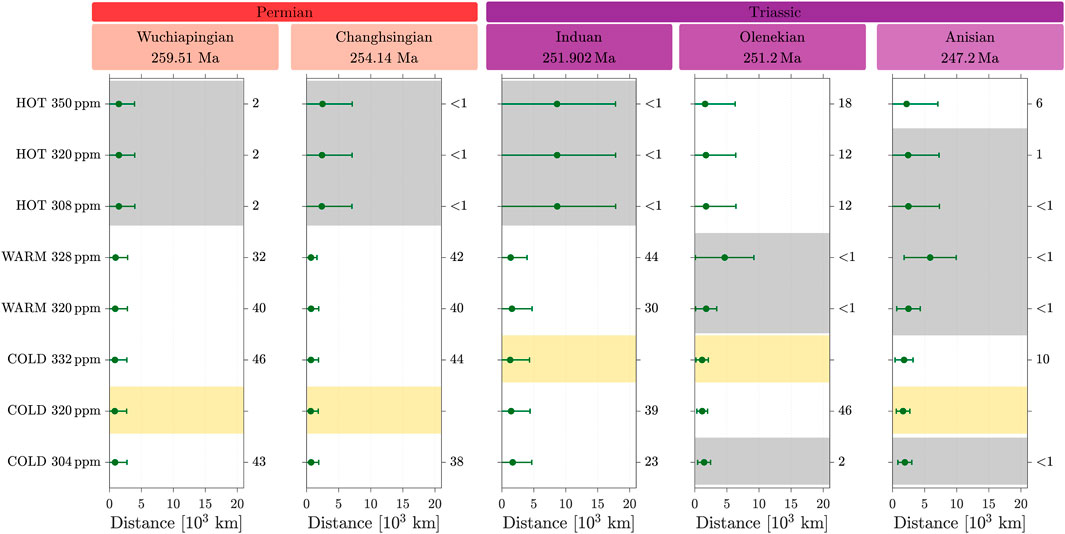
Figure 7. Same as Figure 5, except that the temperate forbland biome is included in the desert major biome (case 2).
In contrast, the results for both the Olenekian and Anisian stages do not differentiate the hot from the cold state. Although the minimal
Therefore, in the mean test, this alternative classification (case 2) is less effective in distinguishing between the attractors compared to the previous option (case 1).
3.4.2 Case 2: test on the median values
The
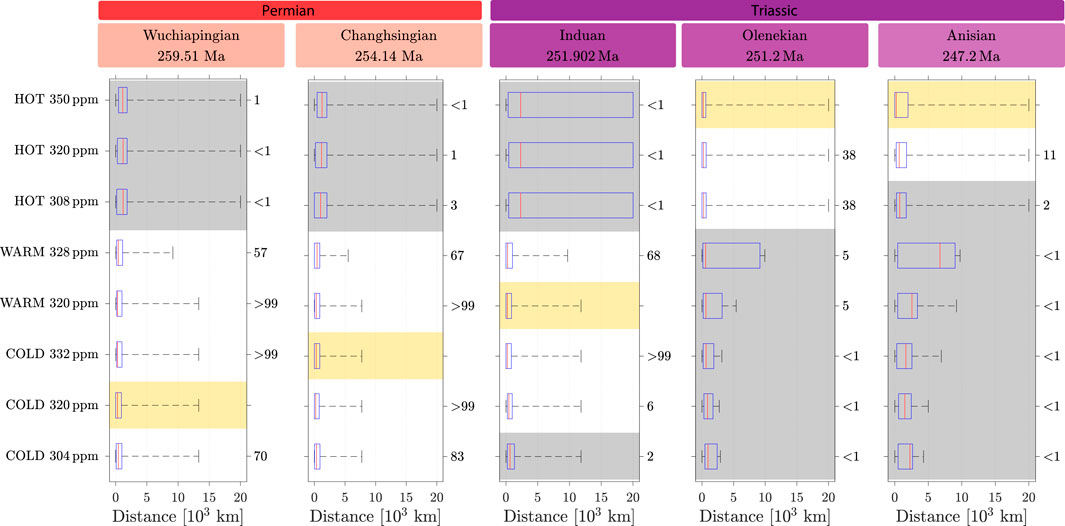
Figure 8. Same as Figure 6, except that the temperate forbland biome is included in the desert major biome (case 2).
For the Olenekian, the hot state with 350 ppm minimizes the median and is comparable to the other positions along the branch. In comparison with the case where temperate forbland is included in the cold temperate major biome, the hot state is favored here over the warm state at 328 ppm. For the Anisian, hot states at 320 ppm and 350 ppm match the vegetation pattern and are significantly different from the other simulations.
4 Summary and conclusion
The increasing temperatures observed between the Lopingian and Middle Triassic (Joachimski et al., 2012) are associated with a transition in the vegetation patterns reported by plant macrofossils (Nowak et al., 2020). We compared the changes over time in macrofossil assemblages to modeled biomes obtained from a series of climate simulations around the Permian–Triassic boundary, which revealed the existence of three alternative steady states with SATs differing by approximately 10
The classification of the temperate forbland biome from the BIOME 4 vegetation model is ambiguous; therefore, two cases have been tested by including it either in cold temperate or desert major biomes. Its classification does not change the results for the Lopingian but has a larger impact on the Early Triassic, for which the presence of the cold temperate major biome in hot-state simulations is fully dependent on the assignment of the temperate forbland biome. The outlier values resulting from the absence of the cold temperate major biome in the hot-state simulations highly impact the mean values. However, the median is less sensitive to the outliers and, thus, to the classification of a particular biome, making the test on median values more robust and conclusive than the test on mean values.
The overall results are summarized in Figure 9 and show that the Lopingian matches well with both the cold and warm states in all tests. The early Middle Triassic (Anisian) is marked by the stabilization of the climate, as reported by
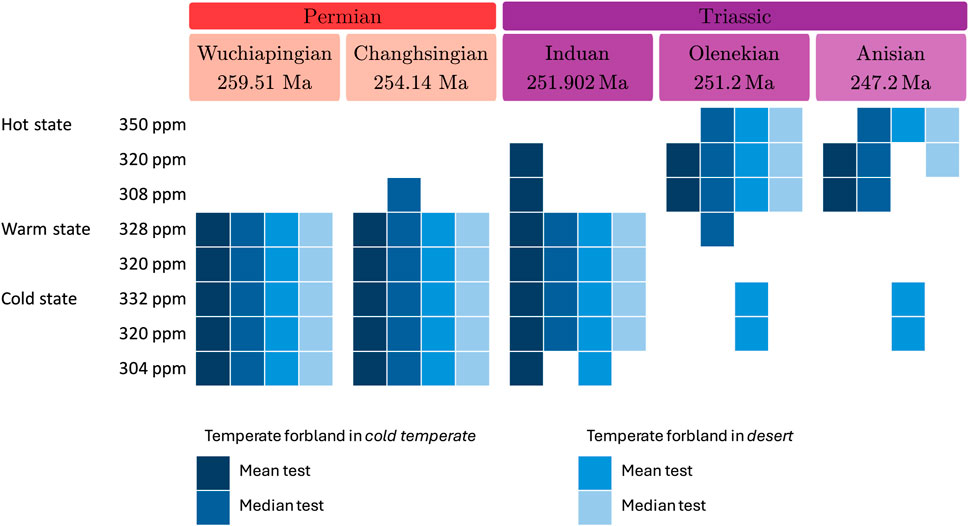
Figure 9. For each stage, the simulations that cannot be excluded from the data are highlighted in blue. The four statistical tests performed are shown from left to right (darker to lighter blue): test on mean and median values for temperate forbland included in the cold temperate major biome (case 1) and test on mean and median values for temperate forbland included in the desert major biome (case 2).
During the Induan, three tests out of four are in favor of a cold or warm state, whereas the remaining one is not conclusive (see Figure 9). Isotope ratios of
The warm state has a narrow, stable branch and cannot be reached through bifurcation-induced tipping from the other attractors (Ragon et al., 2024). Moreover, in all conditions, it is indistinguishable from at least one of the other attractors. Thus, it is relevant to focus on the two robust attractors, the hot and cold states, for the interpretation of the results.
In general, the results presented in this study on changes in the vegetation distribution over time suggest a transition from the cold attractor in the Lopingian to the hot attractor in the latest Early Triassic and early Middle Triassic, with a transient phase in the earliest Triassic (Induan) (see Figure 10). This study provides the most direct comparison to date between the Permian–Triassic plant fossil assemblages and climate simulations. The possibility of discriminating between attractors at the stage level highlights the relevance of using the multistability framework to describe the climatic variations recorded around the Permian–Triassic boundary.
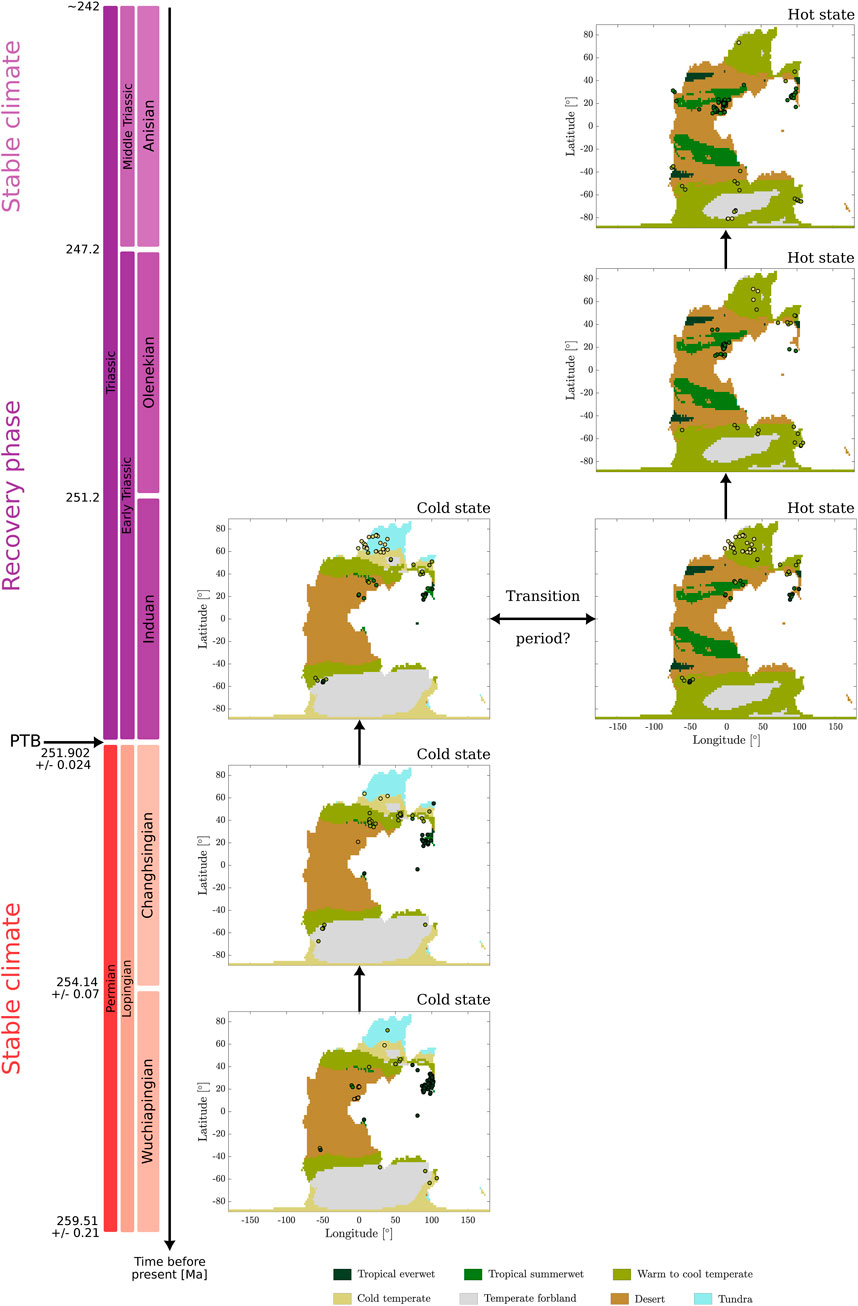
Figure 10. Summary of the comparison between the vegetation distribution associated with macrofossil records from the late Permian to the Early Triassic (Nowak et al., 2020) and the simulated climatic attractors, obtained using an offline coupling between the MITgcm and BIOME4 models, as described by Ragon et al. (2024).
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material; further inquiries can be directed to the corresponding author.
Author contributions
CR: methodology, writing–original draft, writing–review and editing, formal analysis, and visualization. CV: supervision, validation, and writing–review and editing. JK: supervision, validation, writing–review and editing, and methodology. HN: validation and writing–review and editing. EK: validation and writing–review and editing. MB: validation, writing–review and editing, conceptualization, funding acquisition, methodology, supervision, and writing–original draft.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. We acknowledge the financial support from the Swiss National Science Foundation (Sinergia Project No. CRSII5_180253).
Acknowledgments
The authors thank all the Sinergia project members (PaleoC4, https://www.unige.ch/paleoc4/) and Emmanuel Castella for useful discussions. The simulations were performed on the Baobab and Yggdrasil clusters at the University of Geneva.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2025.1520846/full#supplementary-material
References
Adcroft, A., Campin, J.-M., Hill, C., and Marshall, J. (2004). Implementation of an atmosphere ocean general circulation model on the expanded spherical cube. Mon. Weather Rev. 132, 2845–2863. doi:10.1175/MWR2823.1
Ashwin, P., Wieczorek, S., Vitolo, R., and Cox, P. (2012). Tipping points in open systems: bifurcation, noise-induced and rate-dependent examples in the climate system. Philosophical Trans. R. Soc. A Math. Phys. Eng. Sci. 370, 1166–1184. doi:10.1098/rsta.2011.0306
Brunetti, M., and Ragon, C. (2023). Attractors and bifurcation diagrams in complex climate models. Phys. Rev. E 107, 054214. doi:10.1103/PhysRevE.107.054214
Bucur, I. I., Rigaud, S., Del Piero, N., Fucelli, A., Heerwagen, E., Peybernes, C., et al. (2020). Upper triassic calcareous algae from the panthalassa ocean. Riv. Ital. Paleontol. Stratigr. 126. doi:10.13130/2039-4942/13681
Burgess, S. D., Muirhead, J. D., and Bowring, S. A. (2017). Initial pulse of Siberian Traps sills as the trigger of the end-Permian mass extinction. Nat. Commun. 8, 164. doi:10.1038/s41467-017-00083-9
Callegaro, S., Svensen, H. H., Neumann, E. R., Polozov, A., Jerram, D. A., Deegan, F., et al. (2021). Geochemistry of deep Tunguska Basin sills, Siberian Traps: correlations and potential implications for the end-Permian environmental crisis. Contributions Mineralogy Petrology 176, 49. doi:10.1007/s00410-021-01807-3
Chablais, J., Martini, R., Kobayashi, F., Stampfli, G. M., and Onoue, T. (2011). Upper Triassic foraminifers from Panthalassan carbonate buildups of Southwestern Japan and their paleobiogeographic implications. Micropaleontology 57, 93–124.
Chu, D., Grasby, S. E., Song, H., Corso, J. D., Wang, Y., Mather, T. A., et al. (2020). Ecological disturbance in tropical peatlands prior to marine Permian-Triassic mass extinction. Geology 48, 288–292. doi:10.1130/G46631.1
Cohen, K., Finney, S., Gibbard, P., and Fan, J. (2013). The ics international chronostratigraphic chart. Episodes 36, 199–204. doi:10.18814/epiiugs/2013/v36i3/002
Davydov, V. (2021). Tunguska сoals, siberian sills and the permian-triassic extinction. Earth-Science Rev. 212, 103438. doi:10.1016/j.earscirev.2020.103438
Feudel, U. (2023). Rate-induced tipping in ecosystems and climate: the role of unstable states, basin boundaries and transient dynamics. Nonlinear Process. Geophys. Discuss. 2023, 1–29. doi:10.5194/npg-2023-7
Fielding, C. R., Frank, T. D., McLoughlin, S., Vajda, V., Mays, C., Tevyaw, A. P., et al. (2019). Age and pattern of the southern high-latitude continental end-Permian extinction constrained by multiproxy analysis. Nat. Commun. 10, 385. doi:10.1038/s41467-018-07934-z
Galfetti, T., Bucher, H., Ovtcharova, M., Schaltegger, U., Brayard, A., Brühwiler, T., et al. (2007a). Timing of the Early Triassic carbon cycle perturbations inferred from new U–Pb ages and ammonoid biochronozones. Earth Planet. Sci. Lett. 258, 593–604. doi:10.1016/j.epsl.2007.04.023
Galfetti, T., Hochuli, P. A., Brayard, A., Bucher, H., Weissert, H., and Vigran, J. O. (2007b). Smithian-Spathian boundary event: evidence for global climatic change in the wake of the end-Permian biotic crisis. Geology 35, 291–294. doi:10.1130/G23117A.1
Ghil, M., and Lucarini, V. (2020). The physics of climate variability and climate change. Rev. Mod. Phys. 92, 035002. doi:10.1103/RevModPhys.92.035002
Goudemand, N., Romano, C., Leu, M., Bucher, H., Trotter, J. A., and Williams, I. S. (2019). Dynamic interplay between climate and marine biodiversity upheavals during the early Triassic Smithian -Spathian biotic crisis. Earth-Science Rev. 195, 169–178. doi:10.1016/j.earscirev.2019.01.013
Gradstein, S., and Kerp, H. (2012). “Chapter 12 - a brief history of plants on earth,” in The geologic time scale. Editors F. M. Gradstein, J. G. Ogg, M. D. Schmitz, and G. M. Ogg (Boston: Elsevier), 233–237. doi:10.1016/B978-0-444-59425-9.00012-3
Haxeltine, A., and Prentice, I. C. (1996). BIOME3: an equilibrium terrestrial biosphere model based on ecophysiological constraints, resource availability, and competition among plant functional types. Glob. Biogeochem. Cycles 10, 693–709. doi:10.1029/96GB02344
Hochuli, P. A., Hermann, E., Vigran, J. O., Bucher, H., and Weissert, H. (2010). Rapid demise and recovery of plant ecosystems across the end-permian extinction event. Glob. Planet. Change 74, 144–155. doi:10.1016/j.gloplacha.2010.10.004
Hochuli, P. A., Sanson-Barrera, A., Schneebeli-Hermann, E., and Bucher, H. (2016). Severest crisis overlooked—worst disruption of terrestrial environments postdates the Permian–Triassic mass extinction. Sci. Rep. 6, 28372. doi:10.1038/srep28372
Joachimski, M. M., Lai, X., Shen, S., Jiang, H., Luo, G., Chen, B., et al. (2012). Climate warming in the latest Permian and the Permian–Triassic mass extinction. Geology 40, 195–198. doi:10.1130/G32707.1
Kaplan, J. O., Bigelow, N. H., Prentice, I. C., Harrison, S. P., Bartlein, P. J., Christensen, T. R., et al. (2003). Climate change and arctic ecosystems: 2. modeling, paleodata-model comparisons, and future projections. J. Geophys. Res. Atmos. 108. doi:10.1029/2002JD002559
Le Houedec, S., Fucelli, A., Peyrotty, G., Vérard, C., and Martini, R. (2024). Does the panthalassa ocean circulation really rely on single hemispheric gyre? New insight from Nd isotopes from open ocean late triassic terranes. SSRN Prepr. doi:10.2139/ssrn.4939666
Looy, C. V., Twitchett, R. J., Dilcher, D. L., Cittert, J. H. A. V. K.-V., and Visscher, H. (2001). Life in the end-Permian dead zone. Proc. Natl. Acad. Sci. 98, 7879–7883. doi:10.1073/pnas.131218098
Margazoglou, G., Grafke, T., Laio, A., and Lucarini, V. (2021). Dynamical landscape and multistability of a climate model. Proc. R. Soc. A Math. Phys. Eng. Sci. 477, 20210019. doi:10.1098/rspa.2021.0019
Marshall, J., Adcroft, A., Campin, J.-M., Hill, C., and White, A. (2004). Atmosphere–ocean modeling exploiting fluid isomorphisms. Mon. Weather Rev. 132, 2882–2894. doi:10.1175/MWR2835.1
Marshall, J., Adcroft, A., Hill, C., Perelman, L., and Heisey, C. (1997a). A finite-volume, incompressible Navier Stokes model for studies of the ocean on parallel computers. J. Geophys. Res. 102, 5753–5766. doi:10.1029/96JC02775
Marshall, J., Hill, C., Perelman, L., and Adcroft, A. (1997b). Hydrostatic, quasi-hydrostatic, and nonhydrostatic ocean modeling. J. Geophys. Res. 102, 5733–5752. doi:10.1029/96JC02776
Mays, C., and McLoughlin, S. (2022). End-Permian burnout: the role of Permian-Triassic wildfires in extinction, carbon cycling, and environmental change in Eastern Gondwana. PALAIOS 37, 292–317. doi:10.2110/palo.2021.051
Mays, C., Vajda, V., Frank, T. D., Fielding, C. R., Nicoll, R. S., Tevyaw, A. P., et al. (2019). Refined Permian–Triassic floristic timeline reveals early collapse and delayed recovery of south polar terrestrial ecosystems. GSA Bull. 132, 1489–1513. doi:10.1130/B35355.1
Mays, C., Vajda, V., and McLoughlin, S. (2021). Permian–Triassic non-marine algae of Gondwana—distributions, natural affinities and ecological implications. Earth-Science Rev. 212, 103382. doi:10.1016/j.earscirev.2020.103382
Nowak, H., Schneebeli-Hermann, E., and Kustatscher, E. (2019). No mass extinction for land plants at the permian–triassic transition. Nat. Commun. 10, 384. doi:10.1038/s41467-018-07945-w
Nowak, H., Vérard, C., and Kustatscher, E. (2020). Palaeophytogeographical patterns across the permian–triassic boundary. Front. Earth Sci. 8. doi:10.3389/feart.2020.613350
Payne, J. L., Lehrmann, D. J., Wei, J., Orchard, M. J., Schrag, D. P., and Knoll, A. H. (2004). Large perturbations of the carbon cycle during recovery from the end-permian extinction. Science 305, 506–509. doi:10.1126/science.1097023
Peyrotty, G., Brigaud, B., and Martini, R. (2020). δ18O, δ13C, trace elements and REE in situ measurements coupled with U–Pb ages to reconstruct the diagenesis of upper triassic atoll-type carbonates from the Panthalassa Ocean. Mar. Petroleum Geol. 120, 104520. doi:10.1016/j.marpetgeo.2020.104520
Ragon, C., Vérard, C., Kasparian, J., and Brunetti, M. (2024). Alternative climatic steady states near the Permian-Triassic Boundary. Sci. Rep. 14, 26136. doi:10.1038/s41598-024-76432-8
Raup, D. M., and Sepkoski, J. J. (1982). Mass extinctions in the marine fossil record. Science 215, 1501–1503. doi:10.1126/science.215.4539.1501
Renne, P. R., and Basu, A. R. (1991). Rapid eruption of the siberian Traps flood basalts at the permo-triassic boundary. Science 253, 176–179. doi:10.1126/science.253.5016.176
Renne, P. R., Black, M. T., Zichao, Z., Richards, M. A., and Basu, A. R. (1995). Synchrony and causal relations between permian-triassic boundary crises and siberian flood volcanism. Science 269, 1413–1416. doi:10.1126/science.269.5229.1413
Retallack, G. J. (1999). Postapocalyptic greenhouse paleoclimate revealed by earliest Triassic paleosols in the Sydney Basin, Australia. GSA Bull. 111, 52–70. doi:10.1130/0016-7606(1999)111⟨0052:PGPRBE⟩2.3
Rogger, J., Judd, E. J., Mills, B. J. W., Goddéris, Y., Gerya, T. V., and Pellissier, L. (2024). Biogeographic climate sensitivity controls earth system response to large igneous province carbon degassing. Science 385, 661–666. doi:10.1126/science.adn3450
Romano, C., Goudemand, N., Vennemann, T. W., Ware, D., Schneebeli-Hermann, E., Hochuli, P. A., et al. (2013). Climatic and biotic upheavals following the end-Permian mass extinction. Nat. Geosci. 6, 57–60. doi:10.1038/ngeo1667
Schneebeli-Hermann, E. (2020). Regime shifts in an early triassic subtropical ecosystem. Front. Earth Sci. 8. doi:10.3389/feart.2020.588696
Sibik, S., Edmonds, M., Maclennan, J., and Svensen, H. (2015). Magmas erupted during the main pulse of siberian Traps volcanism were volatile-poor. J. Petrology 56, 2089–2116. doi:10.1093/petrology/egv064
Stanley, S. M. (2016). Estimates of the magnitudes of major marine mass extinctions in earth history, Proc. Natl. Acad. Sci. U. S. A. 113, E6325–E6334. doi:10.1073/pnas.1613094113
Svensen, H. H., Frolov, S., Akhmanov, G. G., Polozov, A. G., Jerram, D. A., Shiganova, O. V., et al. (2018). Sills and gas generation in the siberian Traps. Philosophical Trans. R. Soc. A Math. Phys. Eng. Sci. 376, 20170080. doi:10.1098/rsta.2017.0080
Vajda, V., McLoughlin, S., Mays, C., Frank, T. D., Fielding, C. R., Tevyaw, A., et al. (2020). End-Permian (252 Mya) deforestation, wildfires and flooding—an ancient biotic crisis with lessons for the present. Earth Planet. Sci. Lett. 529, 115875. doi:10.1016/j.epsl.2019.115875
Vérard, C. (2019). Panalesis: towards global synthetic palaeogeographies using integration and coupling of manifold models. Geol. Mag. 156, 320–330. doi:10.1017/s0016756817001042
Vérard, C. (2021). 888–444 Ma global plate tectonic reconstruction with emphasis on the formation of Gondwana. Front. Earth Sci. 9. doi:10.3389/feart.2021.666153
Walter, H. (2012). Vegetation of the earth and ecological systems of the geo-biosphere. Springer Science and Business Media.
Widmann, P., Bucher, H., Leu, M., Vennemann, T., Bagherpour, B., Schneebeli-Hermann, E., et al. (2020). Dynamics of the largest carbon isotope excursion during the early triassic biotic recovery. Front. Earth Sci. 8. doi:10.3389/feart.2020.00196
Keywords: Permian–Triassic, paleobiogeography, plant fossils, modeling, tipping, climatic shift
Citation: Ragon C, Vérard C, Kasparian J, Nowak H, Kustatscher E and Brunetti M (2025) Comparison between plant fossil assemblages and simulated biomes across the Permian-Triassic Boundary. Front. Earth Sci. 13:1520846. doi: 10.3389/feart.2025.1520846
Received: 31 October 2024; Accepted: 07 January 2025;
Published: 11 March 2025.
Edited by:
Folco Giomi, University of Rome Tor Vergata, ItalyReviewed by:
Deborah Woodcock, Clark University, United StatesJessie George, Natural History Museum of Los Angeles County, United States
Copyright © 2025 Ragon, Vérard, Kasparian, Nowak, Kustatscher and Brunetti. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Maura Brunetti, bWF1cmEuYnJ1bmV0dGlAdW5pZ2UuY2g=