 Nuno Bicho
Nuno Bicho Eduardo Esteves
Eduardo Esteves
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Earth Sci. , 25 August 2022
Sec. Quaternary Science, Geomorphology and Paleoenvironment
Volume 10 - 2022 | https://doi.org/10.3389/feart.2022.957214
This article is part of the Research Topic Living on the Edge - Interdisciplinary Perspectives on Coastal and Marine Ecosystems in Human Prehistory View all 12 articles
Coastal prehistoric hunter-gatherers in Atlantic Iberia were particularly important to understanding Paleolithic human innovation and resilience. This study will focus on Middle and Upper Paleolithic adaptations to the Iberian Atlantic border. Elements such as intensity and diversity of marine foods, site location, distance to shore, submerged platform, and bathymetry are discussed for the region between Gibraltar and the Gulf of Biscay.
Charles Darwin, some 170 years ago, wrote in his Journal and Remarks, describing the Tierra del Fuego coastal hunter-gatherers, “To knock a limpet from the rocks does not require even cunning, that lowest power of mind. Their skill in some respects may be compared to the instinct of animals…” (Darwin 1839:296–297). This plainly negative and biased perspective of coastal adapted human societies set the pace for the scientific notion of the importance of coastal landscapes and resources for human evolution for most of the 19th and 20th centuries. This same view was still deeply rooted in Man the Hunter, edited in 1968 by Lee and DeVore: hunter-gatherers hunt, gatherer, live in small groups, and move around a lot (Lee and De Vore 1968).
Only in the last 2 decades, authors such as Bailey (e.g., Bailey and Milner 2002; Bailey 2004; Bailey and Flemming 2008) or Erlandson (Erlandson 2001; Erlandson and Scott 2006; Erlandson 2008), among others, have published a great deal trying to reverse the generalized misconception of archaeologists towards marine and aquatic environments in early Prehistory. Marine foods are not perceived anymore as fallback resources in a time of demographic pressure and low resource availability (Bailey and Milner 2002; Bailey 2004; Erlandson and Scott 2006; Bicho and Haws 2008; Erlandson 2008; Bicho, Haws, and Davis 2011; Marean 2011; Marean 2014). On the contrary, they likely stimulated reduced mobility, increased human demographics and population packing (Fitzhugh 1972; Yesner et al., 1980; Keeley 1988; Renouf 1988; Ames and Maschener 1999; Bailey and Milner 2002; Marean 2014), increased complex technology, and economic and social inequality, as seen in the archaeological and ethnographic record (e.g. and examples within Marean 2014; Price 1985; Price and Feinman 2010, 1995). According to Binford (2001:216), the ethnological data show that c. 40% of the mobile human groups in the equatorial and tropical regions use aquatic resources. In the boreal zone, however, 97% of the sedentary hunter-gatherers base their economy and diet on coastal resources.
The time and spatially predictable, and in many cases, easily accessible and in great quantities, coastal resources have provided in the long timeline of the human career the prolonged use of specific areas. These were a perfect set up for the development of human societies that based their social system on social inequality, group identity, territoriality, and respective territorial land markers. All these are characteristics of complex hunter-gatherer groups (Ames 1994; Arnold 1996; Prentiss and Kuijt 2004) also named affluent when applied to coastal hunter-gatherers (e.g., Price, 1985; Koyama and Uchiyama, 2006; Ames, 2007). These aspects are now well accepted by the archaeological community that deals with late Pleistocene and early Holocene human populations in many areas of the world. It was only, however, in the last 15 years that coastal resources become important in the discussion of the emergence of human modernity (e.g., McBrearty and Brooks 2000; Erlandson 2001; Parkington 2001; Bailey and Milner 2002; Bailey et al., 2008; Bicho and Haws 2008; Fa 2008; Finlayson 2008; Marean 2010, 2014; Cortes-Sanchez et al., 2011; Marean 2011; Jerardino 2016; Will, Kandel, and Conard 2019) in both Africa and Europe. Perhaps the first and main reason for the acknowledgment of the importance of marine resources in the diet of early modern humans is that there are nutritional elements fundamental to human health, including the development of the nervous system as well as the pregnancy and early childhood, and that are not produced in the human body (Broadhurst et al., 2002; Jensen 2006; Milligan and Bazinet 2008). The argument has been, thus, within the evolutionary context of early human adaptation, that marine resources may have played a very important role in the adaptation of archaic modern humans due to the changes in the brain and general nervous system (Henshilwood and Marean 2003; Hockett and Haws 2005; Bicho, Haws, and Davis 2011; Marean 2011; Hockett 2012; Marean 2014). In fact, according to Marean (2011), the use of marine resources in South Africa implies a necessary complex cognitive jump shown by the relation between the availability and accessibility of resources and the astronomical calendar through the lunar and tidal cycles.
Marean (2014) has also offered another very interesting hypothesis stemmed on the idea of systematic use of marine resources with direct and important implications for human origins: the use of marine resources, specifically shellfish, provided a perfect context with predictable, both in time and space, coastal resources that allowed reduced mobility, increase human packing due to higher numbers of people. In turn, the sense of territoriality increased as did inter-group conflict, resulting in developed pro-social behavior, based on the presence of predictable high rank dietary resources and social boundary defenses.
In the two perspectives regarding the importance of marine resources for the emergence of human modernity (i.e., human health and brain development and prosocial behavior), the basis is the recognition in the archaeological record of the prolonged use of marine resources also known as coastal adaptations (Erlandson and Scott 2006; Marean 2014). Two problems stem from these concepts. In the archaeological literature, not all authors have used the same definitions or even the same scale for sporadic and systematic use of coastal resources and for coastal adaptations. Naturally, the differences in scale in the use of marine resources likely had different repercussions in human evolution. To solve that problem, Marean (2014), proposed that the only unequivocal proxy was the presence of a shellmidden context. Shellmiddens are contexts where the sediment is shell supported, shells are inter-fingered, and the fine sedimentary matrix fills in the spaces among the shells. The second issue is that a coastal adaptation can be more than just the use of shellfish for dietary reasons seen in a shellmidden context.
In this paper, we will present a survey of coastal or near-coastal Middle and Upper Paleolithic sites in the 2000 km-long coastal region of the Iberian Atlantic coastal margin, from the Gulf of Biscay, in the north, to the Strait of Gibraltar, in the south. In addition, we will also discuss aspects of Paleolithic coastal adaptations in Iberia based on a statistical model, and the importance in the general context of human evolution and the consolidation of our species in the region.
Human evolution is closely linked with the occasional dispersal and migration of various hominin species across time. Many, if not all, major migration episodes in our past are the result of sea and coastal settings. This is the case of Homo erectus in Indonesia at ∼800 ka (Morwood et al., 1998) as well H. floresiensis on the island of Flores (Brown et al., 2004). They offer tantalizing evidence for very early sea crossing capability in East Asia. Petraglia and Rose have also argued for a Red Sea crossing from Eastern Africa (Kenya, Somalia, Djibouti, and Eritrea) to the Arabian Peninsula (Oman and Yemen), for the dispersion of modern humans out of Africa and into Asia (Petraglia and Rose 2009; Rose et al., 2011; Rose and Marks 2014) (Petraglia and Rose 2009; Rose et al., 2011). Findings in the United Arab Emirates seem to confirm this perspective (Armitage et al., 2011). More recently, the sea was a fundamental factor on the dispersal of modern humans to many archipelagos in the Indian and the Pacific oceans, including Sri Lanka (>45 ka), New Guinea/Papua (>35 ka), Australia (>40 ka), New Ireland (30 ka) and the Japanese islands (26 ka) (Allen, Gosden, and White 1989; Gillespie 2002; O'Connell and Allen 2004; Roberts, Jones, and Smith 1990; Storm et al., 2013; Thorne et al., 1999; Wedage et al., 2019; Wedage et al., 2020). The peopling of the New World seems to be pretty much accepted as a coastal migration out of Asia (e.g., Erlandson et al., 2007; Erlandson, Braje, and Graham 2008; Davis 2011) probably sometime during the Last Glacial Maximum (Bennett et al., 2021).
Coastal settings are highly rich in biomass and offered, both in the past and in the present, numerous easily accessible and predictable dietary resources for hunter-gatherers on the move. Not surprisingly, the earliest evidence for human settlement at each continent and region frequently included the use of coastal resources.
In 1979, Thomas and Koyama (1979), applied the term Affluent Foragers to complex coastal temperate forager societies. This term was used in a conflicting perspective to that of Sahlins’ 1968 argument of simple, small-scale highly mobile hunter-gatherers with an egalitarian socioeconomic structure, minimal food storage technology and no sense of territoriality. Although the discussion on the meaning of hunter-gathering complexity is very important, it is not the focus of this article. It is, however, rather fundamental the notion that Affluent societies were (commonly?) characterized by the presence of large group sizes and sedentary lifestyles. Binford (1978, 1982) pointed out that high levels of demographic packing are conditioned by local and temporal carrying capacity and only achieved if resources are available in more productive ecosystems. The technological and economic systems are then marked by the presence of both resource intensification procurement and increasing storage capacity and technology. In his seminal work, Constructing Frames of Reference, Binford (2001) argued based on a huge ethnographical data set that when a landscape is highly packed with hunter-gatherers, these become less mobile. The result is that resource intensification pressure tends to occur, frequently based on territorial marking and defense, favoring the development of exchange of food and non-food items, both locally and regionally. The likely consequence is the transformation of social and territorial boundary arrangements, as well as their economic and technological structure with the presence of storage equipment and, ultimately, with production of food surplus. Testart (1982), for instance, believed that social stratification and inequality may be a consequence of intensification and diversification of resource acquisition, storage and sedentism. In many coastal hunter-gatherer cases there seems to be an association between abundance of marine resources and reduced mobility in highly packed populations with storage facilities (e.g., Ames and Maschener 1999; Bailey and Milner 2002; Prentiss and Kuijt 2004).
Many seafoods are potentially storable (e.g., Yesner 1987, 1998), and seasonal species (such as salmon) and various mollusks can be easily salted, smoked and dried for later consumption. Though in same cases there must be an important investment in storing, these have very little processing costs, in contrast to meat from large mammal prey species that have high to very high processing costs, more so in temperate and low latitudes. Still, in many cases, it seems that highly productive marine environments, more so in continental shelf ecosystems (Erlandson and Scott 2006), are associated to more limited terrestrial resources (Bailey and Milner 2002).
In areas where upwelling systems are present, such as Southern Africa and Southwestern Iberia, the biomass greatly increases seasonally during the windy months. This is the case in the summertime in coastal Portugal, producing today great amounts of fish and shellfish near the shore. At least in southern Iberia, it is clear that during the Late Pleistocene the biomass was many times higher than today (Abrantes 1991, 2000), creating an extremely rich coastal environment for human exploitation. These areas, thus, would be potentially good systems for increasing human population packing and decrease in mobility, but certainly for the exploitation of marine resources and coastal lifeways.
Coastal populations are frequently characterized by high population packing (Binford 2001; Yesner et al., 1980; Kelly 1995; Pálsson 1991, 1988) and reduced residential mobility (Binford 1980; Kelly 1995) due to the high biomass productivity and availability. They also tend to present the highest levels of social complexity (Pálsson 1991; Binford 2001; Prentiss and Kuijt 2004; Koyama and Uchiyama 2006), likely resulting in high levels of social conflict, both at the intra- and inter-group level (Lambert and Walker 1991; Moss, 1993; Lambert 1997; Marean 2014), as predicted by Dyson-Hudson and Smith (1978), probably based on passive territorial boundaries (Peterson 1975).
According to Marean, (2014), coastal hunter-gatherers have the highest levels of territoriality and inter-group conflict recorded among hunter-gathering societies. Marean has recently argued for the origin of prosocial behavior in early human societies in Southern Africa. The argument is based on a logical consideration, based on the facts stated above: when humans included in their diet marine resources, these provided a highly stable, very predictable, and dense high ranked staple food—mobility decreased and population packing increased with an improvement in complex technologies, resulting in economic and social differentiation and a more intensive and extensive gifting and exchange. The consequence was that the use of coastal ecosystems triggered the sense and need for territoriality and in a second phase the development of inter-group conflict. Mechanisms of adaptation and selection provided the appearance of prosocial behavior, a late addition to the modern human cognitive package.
As stated above, it is now believed by some that the importance of marine resources for early modern humans reside at least on the fact there are crucial nutritional elements for human health, specifically for the development of the brain and the retinal organs. These elements are omega 3 and omega 6, long-chain polyunsaturated fatty acids, such as the Docosahexaenoic acid (DHA) and Arachidonic acid (AA) (Crawford et al., 1999; Parkington 2001; Broadhurst et al., 2002; Langdon 2006; Uauy and Dangour 2006; Carlson and Kingston 2007; Cunnane et al., 2007). These fatty acids, also invaluable during pregnancy and early childhood, are not produced in the human body (Broadhurst et al., 2002; Jensen 2006; Milligan and Bazinet 2008) but occur naturally in aquatic plants and animals, if not exclusively, at least frequently in higher concentrations than in terrestrial food elements. The natural outcome, thus, is that marine resources most likely played a fundamental role in the development of the human brain during the evolutionary process of early human adaptation (McBrearty and Brooks 2000; Henshilwood and Marean 2003; Hockett and Haws 2005; Bicho, Haws, and Davis 2011; Marean 2011; Hockett 2012; Marean 2014).
Existing data suggest the use of coastal settings and even resources back to the Middle Pleistocene in the European continent. This was a time when a significant leap in the encephalization quotient occurred with the appearance in Europe of H. heidelbergensis (Ruff, Trinkaus, and Holliday 1997). There might be an important cause-effect connection between the two, so far, unrelated facts since no data are available to even suggest the possibility. If in fact, marine resources were a positive factor in human brain development and expansion related to the emergence of anatomical modern humans or even prior human species, then determining the antiquity of coastal adaptations and their scale is a critical research question.
Following Darwin’s perspective, most researchers up to the end of the millennium, saw coastal resources as low-ranking dietary elements, second quality food assets. They were perceived as limited and unreliable, costly to process or to harvest and, in general, as poor sources of nutrition (Osborn 1977), basically, as starvation foods or fallback resources that people used only in moments of terrestrial scarcity or land resource pressure (for a general discussion on the topic, see Bailey and Milner 2002; Erlandson 2001; Erlandson and Scott 2006). In addition to this view, or perhaps because of it, water and coastal settings were also perceived as a major barrier for human development. In the words of Washburn and Lancaster (1968:294), “During most of human history, water must have been a major physical and psychological barrier and the inability to cope with water is shown in the archaeological record by the absence of remains of fish, shellfish, or any other object going deeply into water or using boats. There is no evidence that resources of river and sea were utilized until late pre-agricultural period. (…) For early man, water was a barrier and a danger, not a resource.” Binford in his 2001 seminal work, Constructing frames of references, shows to a certain degree the type of bias: “So far my discussion of habitats has focused on the properties of terrestrial biotypes because human beings are terrestrial animals who are biologically incapable of living in aquatic environments without technological help. (…) Although human actors are capable of direct participation, ecologically speaking, in a terrestrial setting, they may be thought of as outsiders in aquatic biomes; they intrude at times, but always at very restricted locations and under rather specific conditions. To exploit aquatic resources, humans must be positioned on aquatic-terrestrial ecotones. This positioning, together with the fact that successful human exploitation of an aquatic biome is severely limited (…).” (Binford 2001:166–167). In fact, many of the propositions and generalizations related to the “aquatic biome” offered by Binford (2001) are based more on riverine and lacustrine environments than on coastal settings. Still, and unlike what was argued by Washburn and Lancaster in the 1968 Man the Hunter volume, there is a fairly wide range of early sites with marine and other aquatic elements (e.g., Erlandson 2001; Bailey and Milner 2002; Erlandson and Scott 2006; Bicho and Haws 2008; Finlayson 2008; Bicho, Haws, and Davis 2011; Villa et al., 2020). In the last 3 decades of literature focusing on the Paleolithic times, it is possible to find multiple references to the use of marine resources covering the time span from c. 400,000 years ago to the end of the Paleolithic and from the North Atlantic, in England, to South Africa, from the Mediterranean, the Atlantic and the Indian coasts of Africa. The countries with Mediterranean coast are also represented in this early evidence for the use of marine resources (fish, mollusks, and mammals). Many of these references (more so in the case of the earlier cases) are simple lists of species without any critical review of either how they got there or their importance in terms of hunter-gatherer adaptation. It is important, however, to note that coastal resources are not limited to mollusks. There are other resources groups present in coastal settings that were frequently used in coastal human adaptations: cephalopods, crustaceans, fish, marine mammals, and other, lesser important groups. Some of these are present in archaeological contexts from very early on, across the world (Erlandson 2001; Erlandson and Scott 2006). Some of the more relevant aspects will be described below on each of those prey groups, fundamental for many coastal hunter-gatherers across time and space, including the Iberian Atlantic margin (e.g., Finlayson 2008; Finlayson 2013).
In some areas of the globe, the easiest marine foods to collect are mollusks, both bivalves and gastropods. Erlandson (2001:293–294) argued that what shellfish lack in size make up in quantity and accessibility. On the other hand, Marean (Jerardino and Marean 2010; Marean 2011) has made a convincing argument that due to strong and fast oscillation of tides, mollusk shellfishing could have been dangerous, at least in the Indian waters of coastal South Africa. Nevertheless, another factor seems to have been an important and, perhaps, decisive element for the exploitation of these coastal resources—the fact that, unlike what was previously believed (e.g., Cohen 1977; Osborn 1977), mollusks fall within the group of high rank resources due to its reliability and productivity and efficient protein source (Erlandson 1988; Jones and Richman 1995; Hockett and Haws 2003) at both regional and temporal scales (Stiner, Munro, and Surovell 2000; Parkington 2001; Bailey and Milner 2002; Mannino and Thomas 2002), but also due to the fact that their exploitation can be fully integrated in a system based on gender and age division of labor, resulting in high gathering productivity (Meehan 1977; Glassow and Wilcoxon 1988; Moss, 1993; Bird and Bird 1997; Claassen 1998; Bird and Bliege Bird 2000; Erlandson 2001).
Among the edible species across the world, limpets and mussels seem to have been frequently exploited, at least in early times; although with different intensities, it is the case of Pinnacle Point in South Africa (Jerardino 2010; Marean 2011) and Bajondillo in Southern Spain (Cortes-Sanchez et al., 2011). The interesting aspect is that these are the species that produce the highest levels of AA and DHA, at least during the spawning phases (Brazão et al., 2003; Jensen 2006; Milligan and Bazinet 2008), and they may have been important and the reason why they are present in various early coastal sites both in Africa and Europe
Mollusks, in general, are rich in protein and many essential minerals, but they are low in fat and carbohydrate. Proteins can be metabolized to produce energy, although theoretically a diet high in protein and low in fat and carbohydrates could have serious health consequences, producing the so-called protein poisoning (Noli and Avery 1988; Hockett 2012). Nevertheless, Buchanan (1988) surveyed a range of hunter-gatherer groups with high protein diets and found no evidence of any health effects due to the diet.
Iberian coastal mollusks are marked by a great diversity of species, many of which are edible. Humans, however, do not use all the edible available species, pretty much the same way as land hunter-gatherers do not rely upon all the terrestrial resources available to them (Lee 1968). Coastal people tend to focus, and even specialize, in a certain group of available species. The range of species gathered might be wider in same areas, depending on the local productivity as well as on the fact that both sandy and rocky shores are exploited. Looking at the available archaeological data from early sites (e.g., Waechter 1951; Clark and Straus 1983; Straus and Clark 1986; Antunes 1991; Zilhão et al., 2010; Brown et al., 2011; Colonese et al., 2011; Manne and Bicho 2011; Steele and Álvarez-Fernández 2011; Cortés-Sánchez et al., 2019; Zilhão et al., 2020) there are sometimes a fairly large group of shellfish species. Some of those are likely to have been accidently brought to the site as they frequently live either within the colonies of the edible species (e.g., mussels) or are attached to them. Another possibility is that, independently of the fact that they can be used as food they were also used for ornamental reasons (e.g., Álvarez Fernández and Jöris 2008, Álvarez-Fernández, 2011; Tátá et al., 2014).
Cephalopods include three main groups of edible organisms, squids, cuttlefish, and octopus. While squid does rarely appear close to the intertidal shore, both cuttlefish and octopus are frequent users of the intertidal coastal band. Both species are very easy to fish, but they are present in different ecological settings: octopus is caught in the natural pools in the rocky shores during the low tide and is easily caught by hand without any specialized tools; cuttlefish come with the high tide into shallow coastal water lines to spawn during most of the year and while it is possible to catch by hand it is much easier to collect them with a sack of some kind. Today, at least in southern Portugal, it is very common to catch either species while shellfishing. From the archaeological point of view, remains are extremely rare, since the only hard part is the cuttlebone (in cuttlefish) and the gladius (in squid), as well as the rostrum present in all groups. These are all made mostly of chitin that, unfortunately, is prone to rapid erosion. Still, they were an easily gathered and available resource while looking for shellfish and likely exploited from very early. In addition to their dietary qualities, they are the origin for the so-called India-ink, potentially used for both human and clothing decoration.
The crustacean subphylum (part of the arthropods) is composed by more than 67000 species, covering an incredible array of morphological diversity. For the matter at hand, the important organisms used by hunter-gatherers includes crabs, lobsters and crawfish, shrimp, and goose barnacles. The latter are exclusively marine, and tend to live in shallow and tidal waters, usually in erosive settings. During spring tides, they are easy to gather and appear in large colonies. They can be pulled out directly from the rock, although they tend to come out as single individuals. With the help of a wedge, they come out as a set with dozens of specimens. During neap tides, they may not be so easy to gather since they tend to live in an erosive wavy action zone of the lower intertidal zone, particularly on those areas where they have been exploited systematically. They are small but the fleshy stalks are edible and thought to be very tasty. Perhaps due to both of those facts (low productivity and considered to be very good), goose barnacles tend to be today rare as well as one of the most expensive shellfish types at least in southern Iberia.
Crabs, lobsters, and shrimp are also composed of many species, many of which are edible. These are frequently on the shore and live in the intertidal zone, both in rocky and sandy bottom contexts. They are very easy to gather by hand and they do not need any specialized tool, though the gathering productivity is usually fairly low. In many cases, this will rise fast if baited and or a net is used. Some species tend to live just below the intertidal zone and, thus, shallow diving is required. Crab and lobster exoskeletons are made of chitin and crystalline magnesium calcite. While some body regions (e.g., the claws) are strongly mineralized and very hard and dense, the shell of the body is less mineralized and thus more elastic (Boßelmann et al., 2007). These differences seem to be related to the mechanical requirements of each organ. The differences of hardness in the shell among various species seem to be related to the biological escape behavior of the animals (Boßelmann et al., 2007). This is very important because it might explain the differential preservation found in archaeological contexts, where claws are more common than the rest of the crab shell. Nevertheless, crab shells are very rare in Paleolithic sites, although the cave site of Figueira Brava has evidence of very early exploitation of crabs by Neanderthals south of Lisbon (Zilhão et al., 2020).
Echinoderms includes various main classes, including starfishes, sea urchins and sea cucumbers. Only two of those classes are edible, the sea urchins and the sea cucumbers, but only the urchins have non-edible hard matter, the carapace, and the spines, to survive in the archaeological record. Still, these are very rarely present in Paleolithic sites because both parts are very delicate and break very easily if any force or weight is applied upon them.
Sea cucumbers can be prepared as food from either from raw or dried specimens, and everything is eaten. The sea urchin needs to be prepared, at least with the removal of the spines. The shell is then open from the ventral surface and the only the gonads are eaten, either raw or after the sea urchin has been boiled or grilled. This is perceived as a delicacy in many parts of the world, including Asia, South America and, of course, Iberia. There are over 950 sea urchin species, but in coastal Iberia the most common species is the common sea urchin (Paracentrotus lividus) found at least in the Middle Paleolithic site of Figueira Brava (Zilhão et al., 2020) as well as in the Gibraltar caves (Brown et al., 2011), as well as in Northern Spain the site of El Cuco (Gutiérrez-Zugasti et al., 2013, Gutiérrez-Zugasti et al., 2018).
There is an incredible range of fish species, from the tiniest specimen to extremely large animals (such as the tropical open sea 20 metric ton whale shark), of which many are edible. Fish schools are highly common and represent many species around the world, while some species follow a solitary type of behavior which is rarer.
Fish biomass and productivity can vary greatly, from ecosystem to ecosystem within a small region due to coastal morphology, geology, currents and winds, and the availability of each species’ food. This fact is probably the basis for Kelly’s statement of “… fish cannot be tracked—this is a particular problem in exploiting oceanic fish. The forager can only go to a likely place to find fish, then begin searching randomly. If there are no fish there, the forager could waste quite a bit of time before accepting this is likely” (Kelly 1995:209). Although this is largely true, there are times when certain fish species can be easily exploited in large numbers with the help of simple technology at the shore. It is possible to catch fish by hand, and it is quite easily to do so in the cases of low tide pools where fish get trapped. This type of fishing, of course, can occur casually as a forager looks for shellfish or simply roams the shore for raw materials, including minerals and previously floating wood, more durable and harder for making utensils of all kinds. With technology, fish gathering productivity will rapidly and exponentially increase. Simple technology such as fish gorges, hooks, and nets can improve the fishing results tremendously, more so because in many instances the predictability of the results will also increase. These include longlining, nets and fishtraps. These are expensive gear, and its production is time-consuming, but they have high efficiency, not only in terms of the numbers of captured specimens but also because they do not need the constant presence of humans: they can go there at intervals from one to more days, removing and collecting whatever the gear caught. Both nets and fishtraps do not need any bait (but the latter will work better with some bait), but longlining needs a daily replenishment of bait for each hook. The great advantage of both the traps and the longlining is that fish stay alive while with the nets the fish tend to die after a few hours. Certain species such as the anadromous types, come to the river every year and are easily caught with or without the use of gear while others tend to spawn on the shore or estuary settings. They are, thus, highly predictable both in terms of location and season of the year and they can be mass harvested during those times.
Processing of the catch for storage and later consumption is easy, mainly at coastal environments: salt is naturally available and due to the frequent coastal wind regimes, drying is also an easy task. Although, in both cases the fish in many cases needs to be processed, that is, scaled and eviscerated. In addition, smoking is also a possibility that usually does not require any direct fish processing.
Very much like mollusks, fish also tends to be rich in protein, vitamins, and minerals and less so in fat and carbohydrates. Nevertheless, there are many fish species, from coastal environments that are fatty fish, such as the salmon or the tuna. Fish is in general easily digestible and metabolized by the human body (Erlandson 2001)
Archaeologically preservation of fish is rare in Paleolithic contexts, although is present in some sites during the Middle Paleolithic in Europe (Guillaud et al., 2021). Both vertebra and otoliths tend to preserve better than the rest of the skeleton, but in many cases the recovery of fish remains at an early site depends greatly on the excavation strategy used by the archaeologists. Small mesh size (no bigger than 3 mm and preferably 1 mm) and water screening are the main elements for the recovery of fish remains (Bicho et al., 2000; Bicho, Haws, and Hockett 2006).
Although not nearly as diverse as shellfish, fish or crustaceans, marine mammals present a diverse array of species. These include cetaceans (whales and dolphins), pinnipeds (seals, sea lions, walrus), and sea cows (dugong and manatees), as well as smaller animals such as sea otters. On the other hand, they tend to be much larger than any of the other coastal resources, common weights are above the half metric ton and in the case of the biggest whales, as large as 100 tons. They are frequent biannual migrants, following pre-determined paths, easy to access in many cases. Due to their size (and in certain cases also due to their tusks and teeth) are dangerous preys and pursuit and encounter is frequently seasonal. Still, the amount of meat as well as subcutaneous blubber, other body parts (skin, bones, teeth, ivory) makes them a highly valuable asset after it is hunted and killed. Frequently, dead animals get stranded and end up dying on the shore. They can be then easily scavenged, and many parts of the body can be used if the animal has not died long before. The meat and organs of marine mammals are usually very rich in proteins, vitamins, and minerals, and, depending on the organ and on the species, also in fat. The blubber can also render oils and be stored for later use as fuel for both heating and lighting. The hard skeletal parts can be used for making fishing gears (as important as harpoons) as well as decorative and ornamental elements. The skin can be used not only for clothing as well as for constructing housing and boats.
Hunting of marine mammals can be costly, due to procurement of single individuals both on land and on sea. Technology, such as harpoons and boats, however, may greatly help to improve efficiency in many cases, although there are large, permanent, colonies of seals and sea lions that can be easily hunted down with little effort. In addition, there are also smaller colonies that are seasonal, usually with breeding or birthing locations, but tend to group always in the same spots across time—thus, at least pinnipeds offer a predictable and abundant resource that is usually on land (Muñoz 2011). Still, hunting gear as well as boating technology is frequently associated to pinniped and cetacean hunting. This type of technology is usually very costly in terms of production and maintenance, representing a significant investment of energy (Erlandson 2001).
Although apparently simple to understand the concept of human coastal adaptations, the reality is a bit more complex. This stems from the fact that there are many ways and many scales of adaptation to coastal environments and the use of its resources: from a very small scale, where, for example, one small group of foragers can accidently reach the shore and scavenge a dead cetacean laying on the sand, to a large-scale adaptation where a large society bases all its economic, dietary, social structure as well as its full daily lives on marine resources. If in fact, marine resources were decisive for human evolution and complex cognition (either/or for brain and social development), the main point of contention is to identify the scale in the use of coastal resources that might have had an impact in human evolution. Thus, as Marean (2014) stated, it is fundamental to clearly and unambiguously define and use concepts such as “coastal adaptation,” “maritime adaptation” and “systematic use of coastal resources” and examine what archaeological sites truly fall in each category.
It seems reasonably, then, to argue that if the exploitation of coastal resources is one of the hallmarks of modern cognition, then the use of coastal resources had to have been recurrent and/or intensive enough to impact continuously the human diet. In turn, this had to have a direct outcome on the overall human health of which possibly the brain and retinal development promoted by DHA and AA were the most important for human evolution and the emergence of human cognition. In addition, a new human adaptation may have been borne out of the use of coastal resources: according to Marean (2014), the exploitation and harvesting of coastal resources, at least in certain regions such as South Africa, require a deep understanding of the relation between the tidal cycles and the astronomical calendar and the diverse impact that lunar phases have on resource accessibility and, thus, on its economical return. To know and to understand the correlation between astronomical events and the best time of spring tides to safely exploit the exposed rich intertidal zone reflects necessarily a context of complex developed human cognitive system.
The underlying assumption is that to be able to recognize the economic potential of marine resources it is necessary to have a stable and continuously use of the coastal environment. Thus, for Marean, the origin of a true coastal adaptation in South Africa represents a turning point in the hunter-gatherer hominin adjustment from the previous highly mobile, low density and non-territorial system to a denser and more packed demographic situation and also more permanent and longer settlement.
It is thus necessary to define the applicability of each term to archaeological contexts. While it seems that in the scientific literature the definition of Maritime adaptations is fairly consensual, that is not the case for Coastal adaptations and Systematic use of coastal resources. The former includes seafaring with boat technology and the regular use of boats for both travel and subsistence. In this case, travelling frequently placed hunter-gatherers-fishers away from the shores, while coastal and marine subsistence are a significant part of their diet (e.g., Erlandson and Scott, 2006; Marean 2014; Will, Kandel, and Conard 2019).
The definition of Coastal adaptations and Systematic use of coastal resources, however, is not nearly as consensual, mostly because both terms are frequently used very loosely in the literature. Meanings of Coastal adaptations range from the presence in a coastal area (without necessarily the use of any marine dietary resources) to those contexts where human life-way was transformed by the exploitation of the marine available resources (e.g., Beaton 1995; Bicho and Haws 2008; Marean 2014). It is, thus, necessary to define its meaning better and narrowly, to improve its application.
A few years back, in the inaugural paper of the Journal of Island and Coastal Archaeology, Erlandson and Fitzpatrick (2006: 8) defined coastal adaptations “as any subsistence lifestyle based along the margins of a large body of water that includes the regular use of foods from aquatic habitats.” This definition seems clear enough, but nevertheless issues stem from the grainy definition of “regular use of resources”.
Marean (2014) has proposed a three-level approach to mend the lack of scale and intensity of the “regular use of resources”:
1) Sporadic use of coastal resources is when only a small part of the diet is derived from coastal resources and their exploitation is not regular and cyclic that is, daily or monthly. The result is that the mobility system is not organized around a local or regional mapping or schedule based on the cyclic nature and location of coastal environments.
2) Systematic use of coastal resources reflects a context where coastal resources were regularly and recurrently used, but economically they did not alter the regional adaptation, nor did they change the human settlement system.
3) Coastal adaptation implies an import transformation in the human adaptation directly related to the sea and its resources. Here, the main staple food is from animals that live along the shoreline, mostly from the intertidal zone, including marine mammals, fish, mollusks, crustaceans, birds, etc. Coastal foods, thus, are so important that the mobility strategy deliberate crosses the shore niche as part of the annual plan of landscape use, and in the limit becomes a year-round coastal settlement.
These definitions are straightforward, but still the main problem is their application to the archaeological record, that is how to identify and separate a systematic coastal resource use from its higher-level sister, the coastal adaptation. Marean proposed a simple and highly pragmatic approach to resolve the problem: the presence/absence of shellmiddens and shell remains. The presence of coastal adaptations is confirmed by the occurrence of a shellmiddens (defined as a context where sediment is shell supported, that is where shells are inter-fingered and the fine sedimentary matrix fills in the spaces among the shells); a systematic use of a coastal niche is established by the presence of large amounts of shells, although not making up a midden, but present with regularity and recurrently through the sediments.
Recently, Will et al. (2019) have proposed a different approach, based on an evolutionary perspective: coastal adaptations are a multifaceted set of traits in a group that increases the reproductive fitness of that population through the incorporation of the consumption and use of marine resources during the occupation of coastal landscapes. In practical terms, this includes the expansion of settlements to coastal or near-coastal settings while regularly exploiting and consuming marine resources, such as mollusks, mammals, birds, or fish, independently of forming true shellmiddens.
The “shellmidden” variable suggested by Marean is without a doubt a valuable proxy for the definition of coastal adaptations and systematic use of coastal resources. Nevertheless, it is limited by various factors including shellmidden taphonomical alterations. Perhaps more importantly, is the use of marine coastal resources such as fish that may not be fully represented in the archaeological record due to poor preservation of the ichthyologic remains - their bone density is very low and destruction is usually fast, even if they were abundant at the time of occupation and deposition.
It is unequivocal that edible marine shell species are the main element considered representative of coastal economies. Those remains are highly visible in the landscape and because of that they help to pinpoint and find archaeological sites representative of coastal adaptations. They also accumulate fairly rapidly and, thus, create large middens easily seen in almost any landscape—it is the case of the Mesolithic Atlantic shellmidden, such as those from the Muge in central Portugal (Bicho et al., 2010; Bicho, Cascalheira, et al., 2013), the 3,000 year old “mega” shellmiddens from coastal South Africa (Jerardino 2010), the so called Sambaquis from Brazil (Gaspar 1998; DeBlasis et al., 2007; Villagran et al., 2010), or the present-day accumulations seen in both sides of the African continent, such as in Senegal (Hardy et al., 2016; Camara et al., 2017) or on the Indian side, as seen by one of us in the field work near Inhambane, central Mozambique. The visibility and the volume would suffice to consider them one of the most reliable indicators of coastal use.
Still, the low number of known early sites with marine remains is due to the low data visibility—not all have shells, and in some cases these have not preserved well. As pointed above, this visibility has been partially a result of the bias in the theoretical paradigms of authors: the appearance of marine resources was just a “glitch” in the normal diet and those early humans as was site location. Site location, however, is likely a very important element in the issue of visibility. Many of these early sites with marine resources (or with indirect evidence for their use—Haws et al., 2011) tend to be located in very specific areas on or near the modern coast line. In general, and without any transportation technology such as horse riding, hunter-gatherers have a distance limit of 10 km for inland transportation of edible shellfish (Bigalke 1973; Meehan 1975), even if they process the shellfish and remove the shells (Bird and Bird 1997). If the distance is more than that between logistical and residential base camps, then the skeletons and shells of coastal dietary resources (shellfish, fish, and sea mammals) are left behind. The general consequence of that fact is that coastal resources will be found only within a 10 km radius from the ancient paleoshore. In any case, a study on zebra mussel (Dreissena
polymorpha) and quagga mussel (D. bugensis) coming from the Great Lakes region seems to indicate that at least the adult specimens of these two species may survive up to between 3 and 5 days of transportation (Ricciardi, Serrouya, and Whoriskey 1995), as long as they are not detached from the rock, when the survival to aerial exposure is limited to 24 h (Tucker et al., 1997).
For at least the last c. 100,000 years sea was lower that today, with the lowest peak some 20 ka ago during the Last Glacial Maximum (-120 m), and thus the continental shelf extended considerably the terrestrial landscape of Middle and Upper Paleolithic times. The present modern high sea-stand covered that platform used by Paleolithic hunter-gatherers, with continuous rise since the beginning of the Holocene, and thus submerged almost entirely the coastal archaeological record between the end of the Marine Isotope Stage 5e (MIS5e) and today (Flemming et al., 2017; Flemming, Harff, and Moura 2017). The same phenomenon took place before the previous sea highstand, the MIS5e. From a timeframe perspective, it is very clear that probabilistically the number of sites that should be underwater is incredibly high, since the amount of time is very long compared to the period when highstand settings took place (Bailey and Flemming 2008). Also, the submerged region is a very large area exceeding an estimate of 16 million km2, or about 10% of the total current habitable land surface. However, since those moments were marked by a large ice cap cover and the Americas and Oceania were not occupied yet, that means than the coastal shelf of Africa, Asia, and Europe, now submerged, was even more important for early human adaptation, since apparently it was a well-watered land (Bailey 2004; Finlayson 2013) less arid than the inland regions. That is clear in the Iberian Atlantic coastal region, with physical massive transformation after MIS 5 (Moura, Gomes, and Horta 2017; Bicho, Infantini, and Marreiros 2020).
The Atlantic Iberian coast is close to 2000 km long, starting on the French-Spanish border on the western edge of the Pyrenees, at the extreme north, and running westerly, south, and then easterly again to reach the Rock of Gibraltar, the divide between the open Atlantic Ocean and the Mediterranean (Figure 1).
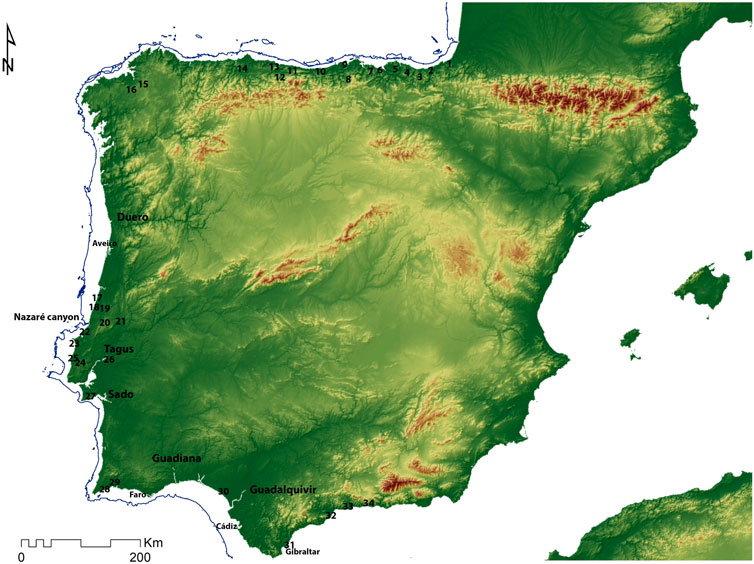
FIGURE 1. Map of Iberia with the main sites discussed in the text. 1. Jaizkibel, 2. Altxerri, 3. Amalda, 4. Santimamine, 5. Toralete, 6. El Cuco, 7. El Pindal, 8. El Perro and La Garma, 9. El Juyo and Morín, 10. Altamira and El Salín, 11. La Riera, 12. El Castillo, 13. Tito Bustillo, 14. Las Caldas, 15. Pena Lliboi, 16. Xestido III, 17. Mira Nascente, 18. Praia do Pedrogão, 19. Lagar Velho, 20. Coelhos, Picareiro, and Anecrial, 21. Caldeirão, 22. Praia Rei Cortiço, 23. Furninha, 24. Suão, 25. Toledo, 26. Cabeço da Amoreira and Cabeço da Arruda, 27. Figueira Brava, 28. Vale Boi, 29. Companheira and Ibn Ammar, 30 Matalascañas, 31, Gorham’s and Vangard; 32. Bajondillo, 33. Humo, 34. Nerja.
From the geomorphological point of view, this coast is highly diverse with long stretches of flat sandy beaches, some as long as 100 km, high consolidated dunes dating from the Miocene to the early Holocene, and medium to high vertical rocky cliffs, standing more than 100 m tall. All these settings are cut through by rivers, from small streams to vary large-sized rivers such as the Tagus, occasionally related to underwater canyons (e.g., off Nazaré and Portimão). Another coastal feature of Atlantic Iberia is the presence of large barrier-islands, such as those present today in Aveiro, Faro, and Cadiz, all extremely rich in biomass and ecological diversity.
Another key element in the region is the presence of the upwelling system, that increases the biomass productivity, mostly in southern Portugal (Loureiro, Newton, and Icely 2005), and occurs at least since MIS5 times (Abrantes 1991, 2000). This phenomenon made the region particularly reach in coastal resources during the Late Pleistocene, even more than today.
Paleoenvironmental proxies seem to indicate that climate was fairly mild in the coastal regions of Portugal and southern Spain even during periods traditionally seen as more rigorous (Carvalho and Bicho 2021; Carvalho et al., 2021). Although terrestrial resources likely were never truly lacking, coastal resources due to their easy access and abundance were frequently exploited from very early on (Bicho and Haws 2008).
Another relevant aspect of the Atlantic coastal region in Iberia is the extent of the present submerged platform. This corresponds to the area above the -125 m bathymetric line, the likely maximum extent of the sea level during the Last Glacial Maximum in Iberia (Flemming et al., 2017; Moura, Gomes, and Horta 2017; Bailey et al., 2020). While this submerged landscape may be as much as 30 km away from the present coastline, in many places is much closer than that. This is due to a very steep bathymetry, mostly in areas where high cliffs are present such as at the Finisterra Cape, the Nazaré, Lisbon and Portimão Canyons, and the Espichel, Sines, Sagres, and Santa Maria capes (Moura, Gomes, and Horta 2017). Thus, it is very likely that this band was intensively used during the Paleolithic, and while many sites may have been destroyed, there should be submerged sites still today as suggested by a number of sites dating to the Late Pleistocence and early Holocene in Atlantic coastal Iberia (Arias Cabal, 2020; Bicho, Infantini, and Marreiros 2020).
The earliest evidence for coastal Atlantic human occupation is dated to the MIS5, seen in a couple of sites in central Portugal. Two of those sites are securely dated, the cave of Figueira Brava (Zilhão et al., 2020) and the open-air site of Praia Rei Cortiço (Haws et al., 2011; Minckley et al., 2015). Praia Rei Cortiço is located just above the intertidal zone, although erosion is frequent due to strong wave action during winter storms. The site was located near a coastal lagoon, during a time of sea high-stand phase, and other than pollen there are no other organic remains. The pollen indicates a typical mixture of temperate and Mediterranean forests, with a series of shifts between cool-wet and warm-dry phase between c. 110–90 thousand years ago. The lithic assemblage is made mostly of quartzite and quartz with a few pieces in chert. From a technological point of view, the assemblage is marked by the presence of typical Levallois, with a few points and flakes, as well as cores (Minckley et al., 2015).
The site of Figueira Brava is located right on the present coast, on a cave open in the limestone cliff between the Tagus and the Sado rivers, just a couple of meters above present sea level. This is an area where the bathymetry is very steep and as a consequence Zilhão and colleagues (Zilhão et al., 2020) argue for a close distance to the contemporaneous shore, no more than 2 km away. Based on the abundant presence of limpets (Patella sp., including P. vulgata, ullyssiponensis, depressa, and rustica), mussels (Mytillus galloprovincialis), clams (Calista chione, Ruditapes decussatus, and Scrobicularia plana) and two types of large crabs (Cancer pagarus and Maja squinado), Zilhão suggests that there was a routine collection of shellfish. This intensive exploitation of coastal resources is also confirmed by the presence of a wide range of fish that includes European eels (Anguilla anguilla), congers (Conger conger), morays (Muraena helena), sharks (Squatina squatina), sea bream (Diplodus vulgaris and Sparus aurata, among others), mackerel (Scomber scombrus), and sea bass (Dicentrarchus labrax). In addition, there are also common dolphin (Delphinus delphis) and ringed seals (Pusa hispida), as well as various types of marine birds (auks, gannets, shags, and cormorants—see Zilhão et al., 2020 for the complete list). The birds are likely to be natural depositions, but the presence of mallards and geese suggests human exploitation. The marine diet was complemented by a wide range of terrestrial sources that include not only mammalians (lagomorphs, red deer, horse, ibex, and aurochs), but also the intensive use of pine nuts. The lithic assemblage follows the normal standard for Mousterian technology in Portugal with the presence of centripetal technology as well as some Levallois. There are rare, retouched pieces, mostly denticulates and sidescrapers. Most of the assemblage is made on local quartz, but there are a few pieces of chert.
Although not dated, the open-air site of Praia do Pedrogão, futher north, is in a very similar geomorphological setting than that of Praia Rei Cortiço and it might have a similar chronology. While there are no preserved organic materials, the lithic assemblage is also similar: quartzite and quartz, with a very low number of formal retouched tools and the presence of Levallois (Aubry, Ribeiro, and Angelucci 2005; Benedetti et al., 2009).
There are other cave sites found on the coast of Portugal, namely, Furninha, (Portuguese Estremadura), Iban Ammar, and Companheira, both in Algarve in the estuary of the Arade river, near the city of Portimão. Unfortunately, there are no dates but some similarities with the assemblages and settings of the site described above, suggests that they are earlier rather than late Mousterian.
Furninha was excavated at the end of the 19th century by Nery Delgado (Delgado and Filipe, 1884; Cardoso and Bicho 2021). The cave, located on a low limestone coastal cliff, had a 4 m sequence, where Middle and Upper Paleolithic horizons were present (Bicho et al., 2010), overlaid by Neolithic and Chalcolithic deposits (Cardoso and Carvalho 2011). There are references to some shells in the sequence, but unfortunately, there is no unequivocal confirmation that those aquatic resources were associated with the Levallois and discoidal technology.
Ibn Ammar and Companheira caves are located just a few km from each other, on either side of the estuary of the Arade river, near the city of Portimão. They are both part of complex cave systems, and are both characterized by small collections, composed of stone tools, terrestrial and marine fauna, specifically different shellfish species (Bicho 2004; Rüther et al., 2022). Two human bones were found in Companheira, but no species has been yet assigned to either fossil (Rüther et al., 2022). The lithic assemblages are characterized by the presence of Levallois and discoidal technologies, made on quartzite, quartz, chert, and limestone. Terrestrial faunas include red deer, equids, aurochs, and rabbit.
Mira Nascente is an open-air location, dated to c. 41 ka ago. It can be found some 30 m above sea level, half-way upslope in a coastal dune (Haws et al., 2010; Haws et al. 2011; Haws et al. 2020), north of the town of Nazaré. This is a high-quality chert-rich country, and thus the lithic assemblage is composed mostly of a regional red chert with just a few pieces of quartzite. The technology includes classic Levallois for both flakes and points. Like in other Middle Paleolithic sites, formal tools are rare. Other than charcoal, no organics were preserved, but use-wear indicates the likelihood of fish consumption (Haws et al., 2011).
The Atlantic Spanish coast provided much elusive evidence for the use of coastal resources during the Middle Paleolithic. While no traditional archaeological sites are known from the southern section, the only northern examples are found close to the coast, on the narrow flat band between the sea and the hilly inland terrain of the majestic Cantabrian mountains. The main site is the cave site of El Cuco (Gutiérrez-Zugasti et al., 2013, Gutiérrez-Zugasti et al., 2018), where levels X through XIII have high numbers of individuals and a wide diversity of shellfish species as well as echinoderms and crustaceans associated with Levallois and discoidal technology. These are dated to c. 40,000 years ago. In addition to El Cuco, there are also a few other sites (Morín, El Castillo, or Amalda) with a few shells in Middle Paleolithic horizons (Gutiérrez-Zugasti et al., 2011). Perhaps the most interesting Middle Paleolithic location is that of Matalascañas, near Cadiz in the Doñana National Park. There, an important set of Neanderthal footprint track was documented, showing that our ancestors were using the coastal dune field over 100 thousand years ago (Mayoral et al., 2021), perhaps at the same time as other Neanderthal groups were visiting the Praia Rei Cortiço in central Portugal and eating shellfish in Figueira Brava.
At the far southern end lies the Rock of Gibraltar with its Middle Paleolithic caves (e.g., Finlayson et al., 2008; Stringer et al., 2008). Gorham’s and Vangard caves lie today literally on the edge of the water at high tide. Even during past times of Neanderthal occupation, the shore was very close. Perhaps, that is the reason why those two sites offer such a diversity of shellfish, marine mammals, and marine birds dating back at least 60 ka (Stringer et al., 2008; Brown et al., 2011).
The Upper Paleolithic evidence is very different from the previous phase since its evidence in Vasco-Cantabrian Spain is extremely well documented. There are innumerous long sequences, generally with good organic preservation, including bones and shells, with art evidence, both in cave and mobile art examples (e.g., Straus and Clark 1986; Arias 2009; Arias et al., 2011; Straus 2015; Straus et al., 2015). The latter in sites such as El Pendo, La Garma, El Juyo, Jaizkibel, Santimamine, and Toralete, attest the connection to the coast and to the presence of marine resources available, and most likely exploited by those hunter-gatherers: there are there are pictorial and engraved references to marine elements both in cave and mobile art—it is the case of the caves of El Pindal and Altxerri, with the representation of pleuronectiform fish (Arias Cabal, 2020) or the image of a whale on a sperm whale tooth dating to the Magdalenian from the Las Caldas cave (Rivero 2015). Also, the presence of frequent ornamental marine shell beads is very common in most sites in the region (Álvarez Fernández and Jöris 2008).
The intensive use of coastal resources clearly started in the Cantabrian Gravettian, although there is some evidence that the exploitation of marine resources took place in the earlier phases of the Chatelperronian and the Aurignacian in the region. Mollusks, echinoderms, crustaceans (both crabs and barnacles), fish and even cetaceans and seals were consumed starting 30,000 years ago, from the Gravettian to the Azilian (Straus and Clark 1986; Straus 2005; Gutiérrez-Zugasti et al., 2011; Álvarez-Fernández, 2011; Straus 2018).
Mollusks are very abundant in many sites, mostly in caves, in the Vasco-Cantabrian region (Álvarez-Fernández, 2011) and include a wide range of edible species. Cuenca Solana was also able to identify their use as tools (Cuenca Solana 2012). Although the frequency and the number of shells is very high, there are no true shell middens during the Upper Paleolithic, despite the presence of thousands of shells at some of those archaeological horizons (Gutiérrez-Zugasti et al., 2011; Álvarez-Fernández, 2011). Their growing importance through time is the basis for the argument that during this period there was a continuous increase in human population and as a result, there was a change in the economy, based on intensification and diversification in the exploitation of the natural resources, including marine resources (Straus and Clark 1986). On northwestern Iberia, there are only a few sites known, of which the most important are Pena Lliboi and Xestido III (Villar Quinteiro, 1997). However, no fauna has been found at these sites, and only their location seems to be of importance since they are within a few kms of the present shore.
In the Portuguese coast, the scenario seems different from that seen in northern Iberia. While the coast in the west section of Iberia is much longer than in the north, it offers a much diverse range of settings and is not limited by a single, closed mountain range such as those found in the Vasco-Cantabrian Spain. Consequently, the landscape is much more open and it offered a wide range of ecological niches and topographical situations that allowed a more diverse human settlement—also a more difficult context to identity preserved Pleistocene archaeological sites, since caves are not as common as in Northern Spain.
The result is that there are only a handful of sites with marine resources found in Atlantic Portugal, that increases slightly if one includes ornamental shells. All sites are located between 10 and 50 km as the crow flies, from the present coastline to those cave sites where marine elements are present. These sites are Suão, Lagar Velho, Coelhos, Picareiro, Anecrial, and Caldeirão (Zilhão 1997; Almeida 2000; Zilhão and Trinkaus 2002; Almeida et al., 2004; Bicho, Haws, and Hockett 2006; Haws, 2012), all in Portuguese Estremadura, and Vale Boi in the Southern coast, in Algarve (Bicho, Stiner, and Lindly 2004; Bicho, Cascalheira, and Marreiros 2012; Bicho, Manne, et al., 2013). Most of these sites present long Upper Paleolithic stratigraphies, sometimes even with Mousterian at the base, and overlaid by Holocene deposits.
Marine resources include fish (more common during the Magdalenian), marine mammals (dolphin and seal), crustaceans (barnacles and crabs), all usually in very residual numbers, and shellfish (limpets, mussels, cockles, clams, scallops), that in the case of Vale Boi are present in high numbers forming thin midden layers, mostly during the Gravettian. The ornaments, that is, perforated shells (Figure 2) include a wide range of species (Chauviere 2002; Vanhaeren and D'Errico 2002; Tátá et al., 2014), more diverse during the Gravettian (Tátá et al., 2014) than during later phases. The main species used for bead production, however, are Littorina obtusata/mariae, Trivia monacha/arctica, Tritia reticulata, and Antalis (or the extinct Dentalium), all coming from the Paleolithic shores.

FIGURE 2. Examples of ornamental perforated shells in the Portuguese Upper Paleolithic. Top to bottom: Littorina obtusata from Vale Boi; Trivia sp from Vale Boi; Theodoxus fluviatilis from Vale Boi; Antalis from Vale Boi.
The main issue that remains is to understand when both coastal settings and marine resources were used in a systematic way and become part of the settlement system as well as of the diet and economy of those hunter-gatherers.
As discussed above, Marean (2014) suggested that the presence of shellmiddens was the only form that one could ascertain the presence of coastal adaptations in the archaeological record. Unfortunately, while this seems to be true, a large part of the Iberian sites (and those from many parts of the world) do now have enough information due to erosion and lack of preservation (both depositional structures and organic materials) to be integrated in this equation. Also, in many cases, publications, mostly old ones, do not offer full numeric data in terms of NISP or MNI on species, particularly on shellfish and fish. Thus, it is impossible to ascertain the quantity of marine resources that were effectively excavated and found at each site or layer. Only partial and proxy information is available in many cases because of those factors and this, in turn, makes it impossible to make comparisons and to ascertain the type of coastal adaptation that took place in each site. To surpass this issue, we have applied a simple statistical method (Supplementary information) to archaeological data coming from a range of Iberian sites dating to the Middle and Upper Paleolithic and the Mesolithic (Table 1) to verify if some variables are good proxies to infer the presence of coastal adaptations (sensu Marean). The variables we used are distance to present shoreline, the number of species of fish, non-edible mollusks, edible mollusks, crustaceans, birds, marine mammals, and perforated shells, as well as other potential proxies (e.g., use wear results in the case of the Middle Paleolithic site of Mira Nascente). The data were analyzed with principal component analysis (PCA). PCA is used in exploratory data analysis and for analyzing the structure of multivariate data. Commonly it is used for dimensionality reduction. This representation of data in a reduced number of dimensions as well as small sample size facilitates the observation of clustering in archeological data that might indicate distinct artifact types (Carlson 2017) or variables such as in the present case . The interest of implementing the PCA on these data was assessed using Bartlett’s sphericity test and Keiser-Mayer-Olkin measure of sampling adequacy (KMO MSA). Components with eigenvalues >1 were retained. Analyses were performed in R version 4.1.3 (RCoreTeam 2022) using functionalities from the add-on packages FactoMineR (Lê, Josse, and Husson 2008), psych (Revelle 2015), and factoextra (Kassambara and Mundt 2020).

TABLE 1. List of sites used for the PCA analyses (Serrano Lozano et al., 1995).
Our data set includes a total of 32 layers from 11 Middle Paleolithic, seven Upper Paleolithic, and three Mesolithic sites. The geographic provenance of the sites includes mostly the Atlantic coast, but we also have site from north Africa (Benzu) as well as relevant sites from the Mediterranean coast (Bajondillo and Humo from the Bay of Malaga, both Middle Paleolithic) and the Upper Paleolithic Nerja cave. Some of these have very little data, limited just to the location near the extant shore, others have full numbers of NISP or MNE per species. We used a set of nine variables: number of fish, edible mollusks, non-edible mollusks, crustacean, bird, marine mammals, and perforated shell species; number of the potential proxies (presence of art with marine elements, echinoderms, cephalopods, use wear showing fish use); and distance in km to the present shoreline.
The Mesolithic shellmiddens, Toledo, on the coast (Araújo 2012), and Cabeço da Arruda and Cabeço da Amoreira, in the Muge valley (Bicho et al., 2010) are clearly the result of coastal adaptations, confirmed by isotopic data on human skeletons where marine resources contributed to the diet up to 70% (Bicho 2012; Peyroteo Stjerna 2016; Bicho et al., 2017), as well as by the size of mounds (40x60 m and 3–5 m in height in the case of Muge), mostly composed of shells. These sites can be used as anchors or proxies for the determination of both which sites are resulting from coastal adaptations and which variables can be used for determining that status.
The results of the PCA for the variables and sites are shown in the plots in Figure 3 and Table 2 (and compiled in the Supplementary Material S1). PCA fairly represented the data (Bartlett’s, p<10–4 with 36 d.f., and KMO MSA=0.603). Distance to the shoreline is not a significant variable. Perhaps, if we had the true distance to the contemporaneous shoreline, the results were different but, unfortunately, it would be impossible to ascertain such distance with the present-day data and an estimate could potentially bring more errors than to use the extant shore. Nevertheless, we decided to include it in the study. There are four variables that are clearly significant in dimension 1 (46.6% of the variance): the number of species of crustaceans, fish, edible mollusks, and mollusks, with birds showing a lower contribution to the system. In dimension 2 (17.5%) there are three variables with strong contributions: the number of species of perforated shells and of birds, and the variable of other proxies.
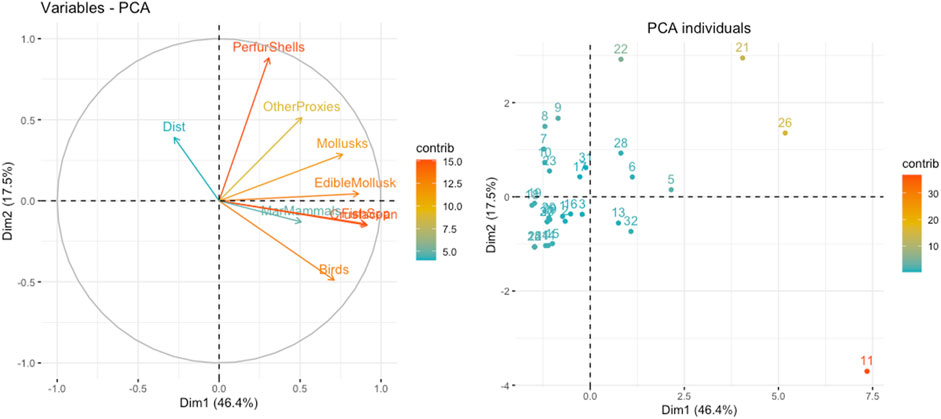
FIGURE 3. PCA Biplot of Dimensions 1 and 2. For site code, see Table 1.
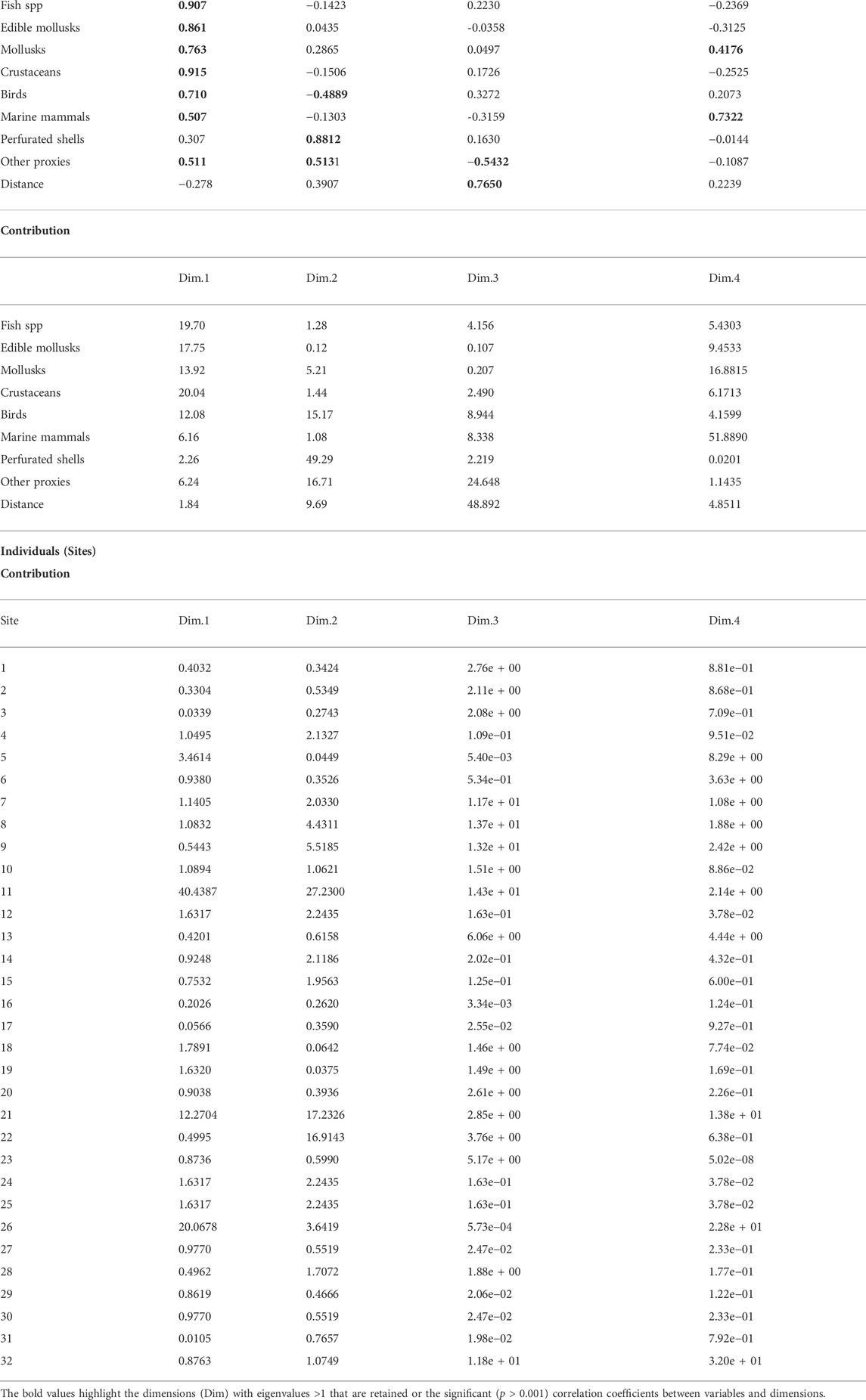
TABLE 2. Summary of results of PCA: Dimensions eigenvalues; variables correlations and contributions to dimensions; and individuals (Sites) contributions to dimensions.
Sites on the positive side of Dimension 1 are those that are likely representing coastal adaptations, and include in addition to the three Mesolithic shellmiddens, three Upper Paleolithic assemblages, two from Nerja cave in the Spanish Mediterranean, and the Gravettian occupation in Vale Boi; and three Middle Paleolithic sites (Vangard, Gorham’s and Figueira Brava). However, Dimension 2 separates the Mesolithic and the Upper Paleolithic sites from the Middle Paleolithic based on the presence of ornamental shells and the lack of bird species (in the Upper Paleolithic sites).
The presence of coastal sites is heavily influenced by the fact that the Atlantic coast in Iberia has important submerged landscapes. In fact, the Pleistocene shore may rest, depending on the chronology, some 30–50 km away from the extant shore, corresponding to the -125 bathymetric line dating to the Last Glacial Maximum (Flemming, Harff, and Moura 2017; Harff and Flemming, 2017; Moura, Gomes, and Horta 2017; Bailey et al., 2020). Nevertheless, most areas present a very steep bathymetry, so the submerged zone is much narrow than that and the distance to present shoreline is within 10 km as the crow flies. Unfortunately, wave action and depth, as well as intertidal range are key elements that have had a strong impact on underwater archaeological preservation (Fa 2008; Arias Cabal, 2020; Bicho, Infantini, and Marreiros 2020) as well as that extent of underwater archaeology field work. The result is that underwater evidence is non-existent in Iberia and, thus, marine and coastal Pleistocene archaeology is coming from terrestrial settings, some very close to the intertidal zone.
The special upwelling conditions, particularly in the South, during Pleistocene times, may have provided a particular ecological coastal setting with high coastal marine productivity (Abrantes 1991, 2000) and because of that, coastal richness may have brought people to live and exploit the Iberian Atlantic shores since Middle Paleolithic times, as suggested by the presence of various sites located on the edge of the modern intertidal zone as well as by the presence of marine faunas where organic preservation exists.
The presence of edible marine species such as fish, marine mammals, and shellfish, many km inland in cave sites namely in Coelhos, Picareiro, or Lagar Velho, raises some interesting aspects. The distance likely precluded that inland populations exploited themselves coastal resources. This is due to the travelling time coupled with the fact that marine resources tend to have a short living time and spoil fast and are not able to endure a trip of a few of days as it would be the case for inland hunter-gatherers. However, there could have been a developed and systematic exchange network, where coastal populations would bring the marine resources halfway, and exchanged those for inland raw materials, including chert, with the inland groups. Otherwise, it would be hard to conceive a mobility system that integrated a round-trip of 100 km, that included both a whole day for shellfish gathering and fishing, followed by a rapid return to the residential campsite transporting those resources in a day, so they would not spoil. Of course, to this, one would have to add the knowledge of the tides, which, as we know inland people tend to ignore completely. Of course, travelling time would not be a concern in the case of ornamental shells, since spoiling was not a concerning factor. And that is likely why there are more inland sites with ornamental shells than with edible resources.
In the case of Vale Boi, there is a clear indication that distance to the contemporaneous shore was a key factor on the frequency of marine resources: while during the Gravettian, shells are present in high numbers, they decrease severely during the LGM, that is, when the coast line is furthest away (Manne and Bicho 2011; Bicho, Manne, et al., 2013).
Archaeological data from the Atlantic Iberian coast clearly shows that the seascape was used starting more than 100,000 years ago, possibly as long as in the Mediterranean world (Stiner 1994; Stringer et al., 2008; Cortes-Sanchez et al., 2011; Villa et al., 2020). For this chronology, site location seems to be the most important proxy, but in some cases (e.g., Figueira Brava) it is unequivocal that marine resources were key elements on the diet of those populations, perhaps in certain times and locations as important as the terrestrial resources. There also seems to be an increase through time on the importance of coastal settings and resources on the life of the Iberian Pleistocene hunter-gatherers.
Our PCA results show two relevant, but very different elements: the variables that can be used to potentially ascertain the sites/layers corresponding to systematic use of the coastal settings (the number of species of crustaceans, fish, edible mollusks, and non-edible mollusks); and the fact that this took place already during the Middle Paleolithic, but likely different from the Upper Paleolithic adaptations.
The intensity in the use of crustaceans, fish, edible mollusks, and mollusks are present in the Middle Paleolithic sites of Figueira Brava and the two-cave site in Gibraltar, as well as in the Upper Paleolithic site of Nerja and Vale Boi, although not all the layers show that. In the case of Vale Boi, only the Gravettian occupation is likely a systematic based coastal adaptation, while those in the latter periods are not—this suggests that the distance to the shoreline is the key variable here that changed the type of diet and economy of that human group. The main differences between the Middle and Upper Paleolithic seems to be the presence of birds in the Middle Paleolithic sites and ornamental shells in the Upper Paleolithic. This difference indicates an identical use of the natural marine resources, and thus a similar diet of those populations. On the other hand, the differences in the birds and perforated shells, suggest a different way of dealing with body ornamentation, with shells in the case of Upper Paleolithic, and feathers and potentially bird bones in the case of the Neanderthals populations, as seen in other areas of Spain and Gibraltar (Finlayson et al., 2012; Rodríguez-Hidalgo et al., 2019).
This study shows that the Atlantic Iberian margins were used in a diverse form during the Pleistocene. Aspects such as the orography of the Vasco-Cantabrian world were an important variable in the human settlement during the Paleolithic, while the flat Portuguese coast, marked by a strong upwelling system provided a highly rich coastal environment that was frequently exploited from very early on, adding to the available terrestrial resources. It is also likely that coastal resources travelled inland, possibly as a part of a complex, but systematic mechanism of network exchanges, sending marine resources inland and bringing lithic resources to the coast. Potentially, this had a seasonal character, but no archaeological evidence can today confirm this hypothesis, neither it can confirm the likelihood that those regional networks were also a key factor on the exchange of people between groups increasing the genetic diversity at each group.
Also relevant is that a wide range of marine resources were exploited from very early on, including crustaceans, fish, mollusks, and marine mammals, some of which were also used for body ornaments. The latter apparently circulated a lot more than the edible resources, probably due to the spoiling effect of travelling inland of fresh marine resources.
Our study also suggests that there are a series of variables, that together, can be used to ascertain or as proxies for the presence of coastal adaptations (sensuMarean 2014). These are the number of species of crustaceans, fish, edible mollusks, and non-edible mollusks. Also, based on this work, it is possible that while coastal adaptations existed since the Middle Paleolithic in Iberia, the lack of perforated shells and the abundance of marine birds characterizing the Middle Paleolithic sites separate them from the Upper Paleolithic occupations. While the presence of ornamental shells in the Upper Paleolithic is a direct consequence of cultural activities related to social behavior, and the presence of birds in the Middle Paleolithic may just be a consequence of a semi-symbiotic relation between Neanderthals and birds as cavers, avoiding the presence of other larger predators. The evidence from various sites in Iberia seem to indicate that those birds were used by Neanderthals for both feathers and talons for body decoration. While the diet and economy were possibly similar between the two populations, apparently the human decorative form and, thus, the social and visual representation of both groups, were distinct.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
NB prepared the archaeological data and wrote the manuscript. EE prepared the statistics and wrote the manuscript
This study received Portuguese national funds from FCT—Foundation for Science and Technology through projects UIDB/04326/2020, UIDP/04326/2020, UIDB/ARQ/04211/2020, UIDP/ARQ/04211/2020, PTDC/HAR-ARQ/27833/2017 (Fundos FEDER through Programa Operacional Regional do Algarve - CRESC Algarve2020 for the project ALG-01-0145-FEDER-027833), and LA/P/0101/2020.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2022.957214/full#supplementary-material
Abrantes, F (2000). 200 000 yr diatom records from Atlantic upwelling sites reveal maximum productivity during LGM and a shift in phytoplankton community structure at 185 000 yr. Earth Planet. Sci. Lett. 176 (1), 7–16. doi:10.1016/S0012-821X(99)00312-X
Abrantes, F (1991). Increased upwelling off Portugal during the last glaciation: Diatom evidence. Mar. Micropaleontol. 17 (3-4), 285–310. doi:10.1016/0377-8398(91)90017-z
Allen, J, Gosden, C, and Peter White, J. (1989). Human Pleistocene adaptations in the tropical island pacific: Recent evidence from new Ireland, a greater Australian outlier. Antiquity 63 (240), 548–561. doi:10.1017/S0003598X00076547
Almeida, F., angelucci, D., Gameiro, C., Correia, J., and Pereira, T. (2004). A ocupação da camada 3 da lapa dos Coelhos (casais martanes, torres novas): Novos elementos sobre a produção de suportes lamelares durante o magdalenense final da Estremadura portuguesa. Promontoria (2), 193–238.
Almeida, F (2000). The terminal gravettian of Portuguese estremadura: Technological variability of the lithic industries. Dallas: S. M. U.
Álvarez-Fernández, E. (2011). Humans and marine resource interaction reappraised: Archaeofauna remains during the late Pleistocene and Holocene in Cantabrian Spain. J. Anthropol. Archaeol. 30 (3), 327–343. doi:10.1016/j.jaa.2011.05.005
Ames, K M. (1994). The Northwest Coast: Complex hunter-gatherers, ecology, and social evolution. Annu. Rev. Anthropol. 23 (1), 209–229. doi:10.1146/annurev.an.23.100194.001233
Ames, K., and Maschener, H. (1999). Peoples of the northwest coast: Their archaeology and prehistory. London: Thames and Hudson.
Ames, K. (2007). Whither affluently complex hunter-forager-gatherers: A review of:‘Beyond affluent foragers: Rethinking hunter-gatherer complexity. In Before farming: The anthropology and archaeology of hunter-gatherers.
Antunes, M. (1991). O Homem da gruta da Figueira Brava (ca. 30.000 BP). Memórias da Academia das Ciências de Lisboa. Cl. Ciências 31, 487–536.
Arias Cabal, P. (2020). “Spain: Underwater exploration on a narrow continental shelf,” in The archaeology of europe's drowned landscapes. Editors G. Bailey, N. Galanidou, H. Peters, H. Jons, and M. Mennenga (Cham: Springer).
Arias, P, Laval, E, Menu, M, González Sainz, C, and Ontañón, R (2011). Les colorants dans l’art pariétal et mobilier paléolithique de La Garma (Cantabrie, Espagne). L'Anthropologie. 115 (3-4), 425–445. doi:10.1016/j.anthro.2011.05.005
Arias, P. (2009). Rites in the dark? An evaluation of the current evidence for ritual areas at magdalenian cave sites. World Archaeol. 41 (2), 262–294. doi:10.1080/00438240902843964
Armitage, S., Jasim, S., Marks, A., Parker, A., Usik, V., Uerpmann, H.-P., et al. (2011). The southern route “out of Africa”: Evidence for an early expansion of modern humans into arabia. Science 331, 453–456. doi:10.1126/science.1199113
Arnold, J. E. (1996). The archaeology of complex hunter-gatherers. J. Archaeol. Method Theory 3 (1), 77–126. doi:10.1007/bf02228931
Aubry, T., Ribeiro, J., and Angelucci, D. (2005). “Testemunhos da ocupação pelo homem de Neanderthal: O sítio da Praia do Pedrogão,” in Habitantes e habitats: Pré e proto-História na Bacia do Lis. Editor S. Carvalho (Leiria: Câmara Municipal de Leiria), 56–66.
Bailey, G., Carrion, J., Fa, D., Finlayson, C., Finlayson, G., Rodriguezvidal, J., et al. (2008). The coastal shelf of the Mediterranean and beyond: Corridor and refugium for human populations in the Pleistocene. Quat. Sci. Rev. 27 (23-24), 2095–2099. doi:10.1016/j.quascirev.2008.08.005
Bailey, G, Galanidou, N, Peeters, H, Jöns, H, and Mennenga, M (2020). “The archaeology of europe's drowned landscapes: Introduction and overview,” in The archaeology of europe's drowned LandscapesGeoff Bailey, nena galanidou, hans peeters, hauke jöns and moritz mennenga (Cham: Springer), 1–23.
Bailey, G., and Flemming, N. (2008). Archaeology of the continental shelf: Marine resources, submerged landscapes and underwater archaeology. Quat. Sci. Rev. 27 (23-24), 2153–2165. doi:10.1016/j.quascirev.2008.08.012
Bailey, G., and Milner, N. (2002). Coastal hunter-gatherers and social evolution: Marginal or central? Before Farming 3/4 (1), 1–22. doi:10.3828/bfarm.2002.3-4.1
Bailey, G. (2004). World prehistory from the margins: The role of coastlines in human evolution. J. Interdiscip. Stud. Hist. Archaeol. 1 (1), 39–50.
Beaton, J M. (1995). The transition on the coastal fringe of Greater Australia. Antiquity 69 (265), 798–806. doi:10.1017/s0003598x0008234x
Benedetti, M M., Haws, J A., Funk, C. L., Michael Daniels, J., Hesp, P. A., Bicho, N. F., et al. (2009). Late Pleistocene raised beaches of coastal Estremadura, central Portugal. Quat. Sci. Rev. 28 (27-28), 3428–3447. doi:10.1016/j.quascirev.2009.09.029
Bennett, M. R., Bustos, D., Pigati, J., Springer, K., Urban, T., Holliday, V., et al. (2021). Evidence of humans in North America during the last glacial maximum. Science 373 (6562), 1528–1531. doi:10.1126/science.abg7586
Bicho, N (2004). The middle Paleolithic occupation of southern Portugal. Settlement Dynamics in Middle Paleolithic and Middle Stone Age 2, (ed.) N. Conard, Kerns Verlag, Tübingen 513–531.
Bicho, N (2012). The chronology of the Mesolithic occupation of the Muge valley, central Portugal. Quat. Int. 279-280, 51. doi:10.1016/j.quaint.2012.07.223
Bicho, N (2015). Ecological niches of the iberian Peninsula: A comparative analysis of European coastal adaptations. In Human origin sites and the world heritage convention in eurasia. UNESCO–World Heritage Papers. (Paris: United Nations Educational, Scientific and Cultural Organization).
Bicho, N, and Cardoso, J (2010). Paleolithic occupations and lithic assemblages from Furninha cave, peniche (Portugal). Revista de Prehistoria y Arqueologia, 17–38.
Bicho, N., Cascalheira, J, and Marreiros, J (2012). “On the (L)edge: The case of Vale Boi rockshelter (Algarve, southern Portugal),” in Caves in context - the cultural significance of caves and rockshelters in EuropeKnut andreas bergsvik and robin skeates (Oxford: Oxbow Books).
Bicho, N, Cascalheira, J, Gonçalves, C, Umbelino, C, Rivero, D. G, André, L, et al. (2017). Resilience, replacement and acculturation in the Mesolithic/Neolithic transition: The case of Muge, central Portugal. Quat. Int. 446, 31–42. doi:10.1016/j.quaint.2016.09.049
Bicho, N, Cascalheira, J, Marreiros, J, Gonçalves, C, Pereira, T, Dias, R, et al. (2013). Chronology of the Mesolithic occupation of the Muge valley, central Portugal: The case of Cabeço da Amoreira. Quat. Int. 308-309, 130–139. doi:10.1016/j.quaint.2012.10.049
Bicho, N., and Haws, J. (2008). At the land's end: Marine resources and the importance of fluctuations in the coastline in the prehistoric hunter–gatherer economy of Portugal. Quat. Sci. Rev. 27 (23-24), 2166–2175. doi:10.1016/j.quascirev.2008.08.011
Bicho, N, Haws, J. A., and Davis, L. G. (2011). Trekking the shore: Changing coastlines and the antiquity of coastal settlement. Springer Science & Business Media.
Bicho, N, Haws, J, and Hockett, B (2006). Two sides of the same coin—Rocks, bones and site function of Picareiro cave, central Portugal. J. Anthropol. Archaeol. 25 (4), 485–499. doi:10.1016/j.jaa.2006.03.009
Bicho, N., Hockett, B, Haws, J, and Belcher, W. (2000). Hunter-gatherer subsistence at the end of the Pleistocene: Preliminary results from Picareiro cave, central Portugal. Antiquity 74 (285), 500–506. doi:10.1017/s0003598x00059834
Bicho, N., Infantini, L., and Marreiros, J. (2020). “Portugal: Intertidal archaeology and submerged coastal landscapes,” in The archaeology of europe’s drowned landscapes. Editors G. Bailey, N. Galanidou, H. Peeters, H. Jöns, and M. Mennenga (Springer International Publishing), 295–300.
Bicho, N, Manne, T, Marreiros, J, Cascalheira, J, Pereira, T, Tátá, F, et al. (2013). The ecodynamics of the first modern humans in Southwestern Iberia: The case of Vale Boi, Portugal. Quat. Int. 318, 102–116. doi:10.1016/j.quaint.2013.06.029
Bicho, N, Stiner, M. C., and Lindly, J. (2004). “Shell ornaments, bone tools and long distance connections in the Upper Paleolithic of Southern Portugal,” in La Spiritualité - actes du colloque International de Liège (Liege: Marcel Otte), 71–80.
Bicho, N, Umbelino, C, Detry, C, and Pereira, T (2010). The emergence of Muge mesolithic shell middens in central Portugal and the 8200 cal yr BP cold event. J. Isl. Coast. Archaeol. 5 (1), 86–104. doi:10.1080/15564891003638184
Bigalke, E. H. (1973). The exploitation of shellfish by coastal tribesmen of the Transkei. Ann. Cape Prov. Mus. Nat. Hist. 9 (9), 159–175.
Binford, Lewis R. (2001). Constructing frames of reference: An analytical method for archaeological theory building using ethnographic and environmental data sets. University of California Press.
Binford, Lewis R. (1982). The archaeology of place. J. Anthropol. Archaeol. 1 (1), 5–31. doi:10.1016/0278-4165(82)90006-x
Binford, Lewis R. (1980). Willow smoke and dogs’ tails: Hunter-gatherer settlement systems and archaeological site formation. Am. Antiq. 45 (1), 4–20. doi:10.2307/279653
Bird, D. W., and Bird, R. L. B (1997). Contemporary shellfish gathering strategies among the meriam of the torres strait islands, Australia: Testing predictions of a central place foraging model. J. Archaeol. Sci. 24 (1), 39–63. doi:10.1006/jasc.1995.0095
Bird, D. W., and Bliege Bird, R (2000). The ethnoarchaeology of juvenile foragers: Shellfishing strategies among meriam children. J. Anthropol. Archaeol. 19 (4), 461–476. doi:10.1006/jaar.2000.0367
Boßelmann, F., Romano, P., Fabritius, H., Raabe, D., and Epple, M. J. T. A. (2007). The composition of the exoskeleton of two crustacea: The American lobster Homarus americanus and the edible crab Cancer pagurus. Thermochim. Acta 463 (1-2), 65–68. doi:10.1016/j.tca.2007.07.018
Brazão, S, Morais, S, Boaventura, D, Pedro, R, Narciso, L, Hawkins, S. J., et al. (2003). Spatial and temporal variation of the fatty acid composition of Patella spp.(Gastropoda: Prosobranchia) soft bodies and gonads. Comp. Biochem. Physiology Part B Biochem. Mol. Biol. 136 (3), 425–441. doi:10.1016/s1096-4959(03)00243-4
Broadhurst, C. L, Wang, Y, Crawford, M. A., Cunnane, S. C., Parkington, J. E., Schmidt, W. F., et al. (2002). Brain-specific lipids from marine, lacustrine, or terrestrial food resources: Potential impact on early african Homo sapiens. Comp. Biochem. Physiology Part B Biochem. Mol. Biol. 131 (4), 653–673. doi:10.1016/s1096-4959(02)00002-7
Brown, K., Darren, A. Fa, Finlayson, G, and Finlayson, C (2011). “Small game and marine resource exploitation by Neanderthals: The evidence from Gibraltar,” in Trekking the shore. Editors N. Bicho, J. Haws, and S. Davis (Springer), 247–272.
Brown, P, Sutikna, T, Morwood, M. J., Raden P Soejono, E., Awe Due, Rokus, Wayhu Saptomo, E., et al. (2004). A new small-bodied hominin from the Late Pleistocene of Flores, Indonesia. Nature 431 (7012), 1055–1061. doi:10.1038/nature02999
Buchanan, W. F. (1988). Shellfish in prehistoric diet: Elands Bay." SW cape coast, South Africa. Oxford: British Archaeological Reports.
Camara, A, Hardy, K, Dioh, E, Gueye, M, Piqué, R, Carré, M, et al. (2017). Shellfish and shell midden sites in the saloum delta (Senegal): Past and present. L'Anthropologie. 121, 204–214. doi:10.1016/j.anthro.2017.03.018
Cardoso, J L, and Bicho, N (2021). Nery Delgado, Pioneer of Archaeological Excavation Methods at the Casa da Moura Cave (Portugal) in 1879–1880. Eur. J. Archaeol. 24 (2), 249–265. doi:10.1017/eaa.2020.55
Cardoso, João Luís, and Faustino Carvalho, António (2011). A gruta da Furninha (peniche): Estudo dos espólios das necrópoles neolíticas. Estud. Arqueol. Oeiras 18, 333–392.
Carlson, B. A., and Kingston, J. D. (2007). Docosahexaenoic acid, the aquatic diet, and hominin encephalization: Difficulties in establishing evolutionary links. Am. J. Hum. Biol. 19 (1), 132–141. doi:10.1002/ajhb.20579
Carvalho, M, and Bicho, N (2021). Complexity in the middle to upper paleolithic transition in peninsular southern Europe and application of refugium concepts. J. Quat. Sci. 37, 380–393. doi:10.1002/jqs.3350
Carvalho, M, Emily Lena Jones, M. G. E., Cascalheira, João, Bicho, Nuno, Meiggs, David, Benedetti, Michael, et al. (2021). Neanderthal palaeoecology in the late middle palaeolithic of Western Iberia: A stable isotope analysis of ungulate teeth from lapa do Picareiro (Portugal). J. Quat. Sci. 37, 300–319. doi:10.1002/jqs.3363
Chauviere, F-X. (2002). Industries et parures sur matières dures animales du Paléolithique supérieur de la grotte de Caldeirão. Rev. Port. Arqueol. 5 (1), 5–28.
Clark, G. A., and Straus, L. G. (1983). “Late pleistocene/early Holocene hunter-gatherer adaptations in cantabrian Spain,” in Hunter-gatherer economy in prehistory. Editor G. Bailey, 131–147.
Cohen, M. N. (1977). The food crisis in prehistory: Overpopulation in the origins of agriculture. New Haven: Yale University Press.
Colonese, A. C., Mannino, M. A., Bar-Yosef Mayer, D. E., Fa, D. A., Finlayson, J. C., Lubell, D., et al. (2011). Marine mollusc exploitation in Mediterranean prehistory: An overview. Quat. Int. 239 (1-2), 86–103. doi:10.1016/j.quaint.2010.09.001
Cortes-Sanchez, M., Morales-Muniz, A., Simon-Vallejo, M. D., Lozano-Francisco, M. C., Vera-Pelaez, J. L., Finlayson, C., et al. (2011). Earliest known use of marine resources by Neanderthals. PLoS One 6 (9), e24026. doi:10.1371/journal.pone.0024026
Cortés-SánchezSimón-Vallejo, D., Jiménez-Espejo, Francisco J., Luis Vera-Peláez, José, Maestro González, Adolfo, and Morales-Muñiz, Arturo (2019). Shellfish collection on the westernmost mediterranean, Bajondillo cave (∼ 160-35 cal kyr BP): A case of behavioral convergence? Quat. Sci. Rev. 217, 284–296. doi:10.1016/j.quascirev.2019.02.007
Crawford, M. A., Bloom, M., Broadhurst, C. L., Schmidt, W. F., Cunnane, S. C., Galli, C., et al. (1999). Evidence for the unique function of docosahexaenoic acid during the evolution of the modern hominid brain. Lipids 34 (1), S39–S47. doi:10.1007/bf02562227
Cuenca Solana, D (2012). Utilización de instrumentos de concha para la realización de actividades productivas en las formaciones económico sociales de los cazadores-recolectores-pescadores y primeras sociedades tribales de la fachada atlántica europea. Santander: Ph.D., Universidad de Cantabria.
Cunnane, S. C., Plourde, M., Stewart, K., and Crawford, M. A. (2007). Docosahexaenoic acid and shore-based diets in hominin encephalization: A rebuttal. Am. J. Hum. Biol. 19 (4), 578–581. doi:10.1002/ajhb.20673
Davis, L G. (2011). “The North American paleocoastal concept reconsidered,” in Trekking the shore. Editors N. Bicho, J. Haws, and S. Davis (Springer), 3–26.
DeBlasis, P, Kneip, A, Scheel-Ybert, R, Giannini, P C, and Gaspar, M. Du (2007). Sambaquis e paisagem: Dinâmica natural e arqueologia regional no litoral do sul do brasil. Arqueol. suramericana 3 (1), 29–61.
Delgado, J, and Filipe, N (1884). “La grotte de Furninha a Peniche,” in Compte-Rendu de la IX éme Session à Lisbonne (1880) (Lisbon: Typographie de l'Academie Royale des Sciences).
Detry, Cleia (2007). Paleoecologia e paleoeconomia do baixo tejo no mesolítico final: O contributo do estudo dos mamíferos dos concheiros de Muge. Salamanca: Universidad de Salamanca.
Dias, R., Detry, C., and Bicho, N. (2016). Changes in the exploitation dynamics of small terrestrial vertebrates and fish during the pleistocene-holocene transition in the SW iberian Peninsula: A review. Holocene 26, 964–984. doi:10.1177/0959683615622547
Dyson‐Hudson, Rada, and Alden Smith, Eric (1978). Human territoriality: An ecological reassessment. Am. Anthropol. 80 (1), 21–41. doi:10.1525/aa.1978.80.1.02a00020
Erlandson, J., Braje, T. J., and Graham, M H. (2008). How old is MVII?—seaweeds, shorelines, and the pre-clovis chronology at monte verde, Chile. J. Isl. Coast. Archaeol. 3 (2), 277–281. doi:10.1080/15564890802412098
Erlandson, J., Graham, M H., Debra Corbett, B. J. B, Estes, J. A., and Steneck, R. S. (2007). The kelp highway hypothesis: Marine ecology, the coastal migration theory, and the peopling of the Americas. J. Isl. Coast. Archaeol. 2 (2), 161–174. doi:10.1080/15564890701628612
Erlandson, J. (2008). Racing a rising tide: Global warming, rising seas, and the erosion of human history. J. Isl. Coast. Archaeol. 3 (2), 167–169. doi:10.1080/15564890802436766
Erlandson, J. (2001). The archaeology of aquatic adaptations: Paradigms for a new millennium. J. Archaeol. Res. 9 (4), 287–350. doi:10.1023/a:1013062712695
Erlandson, J. M. (1988). The role of shellfish in prehistoric economies: A protein perspective. Am. Antiq. 53 (1), 102–109. doi:10.2307/281156
Erlandson, J, and Scott, F (2006). Oceans, islands, and coasts: Current perspectives on the role of the sea in human prehistory. J. Isl. Coast. Archaeol. 1 (1), 5–32. doi:10.1080/15564890600639504
Fa, D. (2008). Effects of tidal amplitude on intertidal resource availability and dispersal pressure in prehistoric human coastal populations: The mediterranean–atlantic transition. Quat. Sci. Rev. 27 (23-24), 2194–2209. doi:10.1016/j.quascirev.2008.07.015
Fernández, Alvarez, E., and Jöris, Olaf (2008). Personal ornaments in the early upper paleolithic of western eurasia: An evaluation of the record. Eur. Prehistory 5 (2), 29–42.
Fernández, J R, Douka, K, Alistair, W. G. P, Louise, T, Peter van, C, and Zilhão, J (2011). Dating of the middle to upper paleolithic transition at the abrigo 3 del Humo (málaga, Spain): Cronología de la transición del paleolítico medio al Superior en el abrigo 3 del Humo (málaga, españa). Mainake 33, 275–284.
Finlayson, C, Brown, K, Blasco, R, Rosell, J, José Negro, J, Bortolotti, G R., et al. (2012). Birds of a feather: Neanderthal exploitation of raptors and corvids.
Finlayson, C (2013). The Water Optimisation Hypothesis and the human occupation of the mid-latitude belt in the Pleistocene. Quat. Int. 300, 22–31. doi:10.1016/j.quaint.2013.03.040
Finlayson, C. (2008). On the importance of coastal areas in the survival of Neanderthal populations during the Late Pleistocene. Quat. Sci. Rev. 27 (23-24), 2246–2252. doi:10.1016/j.quascirev.2008.08.033
Finlayson, G., Finlayson, C., Giles Pacheco, F., Rodriguez Vidal, J., Carrión, J. S., Recio Espejo, J. M., et al. (2008). Caves as archives of ecological and climatic changes in the Pleistocene—the case of gorham's cave, Gibraltar. Quat. Int. 181 (1), 55–63. doi:10.1016/j.quaint.2007.01.009
Fitzhugh, W. (1972). Environmental archaeology and cultural systems in Hamilton inlet, labrador. A survey of the central labrador coast from 3000 B.C. To the present. Washington: Smithsonian.
Flemming, N C., Jan, H, Moura, D, Burgess, A, and Bailey, G. N. (2017). “Introduction: Prehistoric remains on the continental shelf—why do sites and landscapes survive inundation?,” in Submerged landscapes of the European continental shelf (Quaternary Paleoenvironments) (Oxford: Wiley Blackwell), 1–10.
Flemming, N C., Jan, Harff, and Moura, Delminda (2017). “Non-cultural processes of site formation, preservation and destruction,” in Submerged landscapes of the European continental shelf. Editors N. Flemming, J. Harff, D. Moura, A. Burgess, and G. Bailey (Oxford: Wiley Blackwell), 51–82.
Gaspar, M D (1998). Considerations of the sambaquis of the Brazilian coast. Antiquity 72 (277), 592–615. doi:10.1017/s0003598x00087020
Gillespie, R (2002). Dating the first Australians. Radiocarbon 44 (2), 455–472. doi:10.1017/s0033822200031830
Glassow, M A., and Wilcoxon, L R. (1988). Coastal adaptations near Point Conception, California, with particular regard to shellfish exploitation. Am. Antiq. 53 (1), 36–51. doi:10.2307/281153
Guillaud, E, Béarez, P, Daujeard, C, R Defleur, A, Desclaux, E, Roselló-Izquierdo, E, et al. (2021). Neanderthal foraging in freshwater ecosystems: A reappraisal of the middle paleolithic archaeological fish record from continental western Europe. Quat. Sci. Rev. 252, 106731. doi:10.1016/j.quascirev.2020.106731
Gutiérrez-Zugasti, I., Cuenca-Solana, D., Rasines del Rio, P., Muñoz, E., Santamaría, S, Morlote, J., et al. (2013). The role of shellfish in hunter–gatherer societies during the early upper palaeolithic: A view from el Cuco rockshelter, northern Spain. J. Anthropol. Archaeol. 32 (2), 242–256. doi:10.1016/j.jaa.2013.03.001
Gutiérrez-Zugasti, I, S., Araújo, Ana C., Dupont, C, Milner, N, AntonioMonge-Soares, M., and Monge-Soares, A. M. (2011). Shell midden research in Atlantic Europe: State of the art, research problems and perspectives for the future. Quat. Int. 239 (1-2), 70–85. doi:10.1016/j.quaint.2011.02.031
Hardy, K, Camara, A, Piqué, R, Dioh, E, Guèye, M, Diaw Diadhiou, H, et al. (2016). Shellfishing and shell midden construction in the Saloum Delta, Senegal. J. Anthropol. Archaeol. 41, 19–32. doi:10.1016/j.jaa.2015.11.001
Harff, J, and Flemming, N C. (2017). “Andreas groh, birgit hünicke, gilles lericolais, martin meschede, alar rosentau, dimitris sakellariou, szymon uścinowicz, and wenyan ZhangSea level and climate,” in Submerged landscapes of the European continental shelf. Editors N. Flemming, J. Harff, D. Moura, A. Burgess, and G. Bailey (Oxford: Willey Blackwell), 11–49.
Haws, J. A., Funk, C L., Benedetti, M. M., Bicho, N. F., Daniels, J. M, Minckley, T. A., et al. (2011). “Paleolithic landscapes and seascapes of the west coast of Portugal,” in Trekking the shore. Editors N. Bicho, J. Haws, and S. Davis (Springer), 203–246.
Haws, J. (2012). Paleolithic socionatural relationships during MIS 3 and 2 in central Portugal. Quat. Int. 264, 61–77. doi:10.1016/j.quaint.2011.10.003
Haws, J A., Benedetti, M M., Funk, C L., Bicho, N F., Michael Daniels, J., Hesp, P A., et al. (2010). Coastal wetlands and the neanderthal settlement of Portuguese Estremadura. Geoarchaeology 25 (6), 709–744. doi:10.1002/gea.20330
Haws, J A., Benedetti, M M., Funk, C L., Bicho, N F., Pereira, T, Marreiros, J, et al. (2020). Late Pleistocene landscape and settlement dynamics of Portuguese Estremadura. J. Field Archaeol. 45 (4), 222–248. doi:10.1080/00934690.2020.1733780
Henshilwood, C. S., and Marean, C. (2003). The origin of modern human behavior-Critique of the models and their test implications. Curr. Anthropol. 44 (1), 627–651. doi:10.1086/377665
Hockett, B, and Haws, J A. (2005). Nutritional ecology and the human demography of Neandertal extinction. Quat. Int. 137 (1), 21–34. doi:10.1016/j.quaint.2004.11.017
Hockett, B, and Haws, J (2003). Nutritional ecology and diachronic trends in Paleolithic diet and health. Evol. Anthropol. 12 (5), 211–216. doi:10.1002/evan.10116
Hockett, B (2012). The consequences of Middle Paleolithic diets on pregnant Neanderthal women. Quat. Int. 264, 78–82. doi:10.1016/j.quaint.2011.07.002
Igor, G-Z, Marín-Arroyo, A B., Rasines del Río, P, Maroto, J, JenniferJones, R., Bailey, G N., et al. (2018). A chrono-cultural reassessment of the levels VI–xiv from el Cuco rock-shelter: A new sequence for the late middle paleolithic in the cantabrian region (northern Iberia). Quat. Int. 474, 44–55. doi:10.1016/j.quaint.2017.06.059
Izquierdo, R, EufrasiaMuñiz, A. M, and Cañas Díaz, J. M (1995). “Estudio ictioarqueológico de la Cueva de Nerja (Málaga). Resultados de las campañas de 1980 y 1982. Salas de la Mina e da la Torca,ȝ in Fauna de la Cueva de Nerja. Salas de la Mina e de la Torca. Campañas de 1980-82 (Nerja: Patronato de la Cueva de Nerja), 161–218.
Jensen, Craig L. (2006). Effects of n− 3 fatty acids during pregnancy and lactation. Am. J. Clin. Nutr. 83 (6), S1452–1457S. doi:10.1093/ajcn/83.6.1452s
Jerardino, A., and Marean, C. W. (2010). Shellfish gathering, marine paleoecology and modern human behavior: Perspectives from cave PP13B, Pinnacle point, south Africa. J. Hum. Evol. 59 (3-4), 412–424. doi:10.1016/j.jhevol.2010.07.003
Jerardino, A (2010). Large shell middens in lamberts Bay, south Africa: A case of hunter–gatherer resource intensification. J. Archaeol. Sci. 37 (9), 2291–2302. doi:10.1016/j.jas.2010.04.002
Jerardino, A (2016). On the origins and significance of Pleistocene coastal resource use in southern Africa with particular reference to shellfish gathering. J. Anthropol. Archaeol. 41, 213–230. doi:10.1016/j.jaa.2016.01.001
Jones, T L., and JenniferRichman, R. (1995). On mussels: Mytilus californianus as a prehistoric resource. North Am. Archaeol. 16 (1), 33–58. doi:10.2190/g5tt-yfhp-je6a-p2tx
Kassambara, A, and Mundt, F (2020). factoextra: Extrac and visualize the results of multivariate data analyses. R Package Version 1.0.7Available at: https://CRAN.R-project.org/package=factoextra.
Keeley, L H. (1988). Hunter-gatherer economic complexity and “population pressure”: A cross-cultural analysis. J. Anthropol. Archaeol. 7 (4), 373–411. doi:10.1016/0278-4165(88)90003-7
Koyama, S, and Uchiyama, J (2006). “Why "beyond affluent foragers"? Looking back at the original affluent foragers concept,” in Beyond affluent foragers. Rethinking hunter-gatherer complexity. Editors C. Grier, J. Kim, and Junzō Uchiyama (Oxford: Oxbow Books), 1–4.
Lambert, P M., and Walker, P L. (1991). Physical anthropological evidence for the evolution of social complexity in coastal southern California. Antiquity 65 (249), 963–973. doi:10.1017/s0003598x00080765
Lambert, P. M. (1997). “Patterns of violence in prehistoric hunter gatherer societis of coastal southern California,” in Troubled times: Violence and warfare in the past (war and society). Editors D. Frayer, and D. Martin (London: Routledge), 77–109.
Langdon, J. H. (2006). Has an aquatic diet been necessary for hominin brain evolution and functional development? Br. J. Nutr. 96 (1), 7. doi:10.1079/bjn20061805
Lê, S, Josse, J, and Husson, F (2008). FactoMineR: an R package for multivariate analysis. J. Stat. Softw. 25, 1–18. doi:10.18637/jss.v025.i01
Lee, Richard B. (1968). “What hunters do for a living, or, how to make out on scarce resources,” in Man the hunter (London: Routledge), 30–48.
Loureiro, S, Newton, A, and Icely, J D. (2005). Microplankton composition, production and upwelling dynamics in Sagres (SW Portugal) during summer of 2001. Sci. Mar. 69 (3), 323–341. doi:10.3989/scimar.2005.69n3323
Manne, T, and Bicho, N. F. (2011). “Prying new meaning from limpet harvesting at Vale Boi during the upper paleolithic,” in Trekking the shore. Editors N. Bicho, J. Haws, and L. Davis (Springer), 273–289.
Mannino, M. A., and Thomas, K. D. (2002). Depletion of a resource? The impact of prehistoric human foraging on intertidal mollusc communities and its significance for human settlement, mobility and dispersal. World Archaeol. 33 (3), 452–474. doi:10.1080/00438240120107477
Marean, C. W. (2011). “Coastal South Africa and the coevolution of the modern human lineage and the coastal adaptation,” in Trekking the shore. Editors N. Bicho, J. Haws, and Loren G. Davis (Springer), 421–440.
Marean, C. W. (2010). Pinnacle point cave 13B (western cape province, south Africa) in context: The cape floral kingdom, shellfish, and modern human origins. J. Hum. Evol. 59 (3-4), 425–443. doi:10.1016/j.jhevol.2010.07.011
Marean, C. W. (2014). The origins and significance of coastal resource use in Africa and Western Eurasia. J. Hum. Evol. 77, 17–40. doi:10.1016/j.jhevol.2014.02.025
Mayoral, E, Díaz-Martínez, I, Duveau, J, Santos, A, Rodríguez Ramírez, A, Morales, J A., et al. (2021). Tracking late Pleistocene neandertals on the iberian coast. Sci. Rep. 11 (1), 4103. doi:10.1038/s41598-021-83413-8
McBrearty, S., and Brooks, A. S. (2000). The revolution that wasn't: A new interpretation of the origin of modern human behavior. J. Hum. Evol. 39 (5), 453–563. doi:10.1006/jhev.2000.0435
Meehan, B F (1975). Shell bed to shell midden. PhD dissertation. Australia: The Australian National University.
Meehan, Betty (1977). “Man does not live by calories alone: The role of shellfish in coastal cuisine,“ in Sunda and Sahul. Editors Allen, J., Golson, J., and Jones, R. (New York, NY: Academic Press), 493–531.
Milligan, L A., and Bazinet, R P. (2008). Evolutionary modifications of human milk composition: Evidence from long-chain polyunsaturated fatty acid composition of anthropoid milks. J. Hum. Evol. 55 (6), 1086–1095. doi:10.1016/j.jhevol.2008.07.010
Minckley, T. A., Haws, J. A., Benedetti, M M., Brewer, S. C., and Forman, S. L. (2015). Last interglacial vegetation and climate history from the Portuguese coast. J. Quat. Sci. 30 (1), 59–69. doi:10.1002/jqs.2758
Morales, A. (1998). Late upper paleolithic subsistence strategies in southern Iberia: Tardiglacial faunas from cueva de Nerja (Malaga, Spain). Eur. J. Archaeol. 1 (1), 9–50. doi:10.1177/146195719800100102
Morwood, M J., PaulO'Sullivan, B., Aziz, F, and Raza, A (1998). Fission-track ages of stone tools and fossils on the east Indonesian island of Flores. Nature 392 (6672), 173–176. doi:10.1038/32401
Moss, M., and Erlandson, J. M. (1992). Forts, refuge rocks, and defensive sites: The antiquity of warfare along the north pacific coast of north America. Arct. Anthropol., 73–90.
Moss, M. (1993). Shellfish, gender, and status on the Northwest Coast: Reconciling archeological, ethnographic, and ethnohistorical records of the Tlingit. Am. Anthropol. 95 (3), 631–652. doi:10.1525/aa.1993.95.3.02a00050
Moura, D, Gomes, A, and Horta, J (2017). “The iberian atlantic margin,” in Submerged landscapes of the European continental shelf. Quaternary paleoenvironments. Editors N. Flemming, J. Harff, D. Moura, A. Burgess, and G. Bailey (OxfordOxford: Wiley-BlackwellWilley Blackwell), 281–300.
Muñoz, A. S (2011). “Pinniped zooarchaeological studies in southern patagonia: Current issues and future research agenda,” in Trekking the shore. Editors N. Bicho, J. Haws, and L. Davis (Springer), 305–331.
Noli, D, and Avery, G (1988). Protein poisoning and coastal subsistence. J. Archaeol. Sci. 15 (4), 395–401. doi:10.1016/0305-4403(88)90037-4
O'Connell, J., and Allen, J (2004). Dating the colonization of sahul (Pleistocene Australia–new Guinea): A review of recent research. J. Archaeol. Sci. 31 (6), 835–853. doi:10.1016/j.jas.2003.11.005
Pálsson, G (1991). Coastal economies, cultural accounts: Human ecology and Icelandic discourse. Manchester: Manchester University Press.
Parkington, J (2001). Milestones: The impact of the systematic exploitation of marine foods on human evolution. Humanity from African naissance to coming millennia, 327–336.
Peterson, N (1975). Hunter-gatherer territoriality: The perspective from Australia. Am. Anthropol. 77 (1), 53–68. doi:10.1525/aa.1975.77.1.02a00040
Peyroteo Stjerna, R. (2016). Occasional papers in archaeology, 60. Uppsala: Department of Archaeology and Ancient History, Uppsala University.On death in the mesolithic. Or the mortuary practices of the last hunter-gatherers of the south-western iberian Peninsula, 7th–6th millennium BCE
Prentiss, W C., and Ian Kuijt, (2004). Complex hunter gatherers: Evolution organization of prehistoric communities plateau of northwestern NA. Salt Lake City: University of Utah Press.
Price, T. D (1985). “Affluent foragers of mesolithic southern scandinavia,” in Prehistoric hunters-gatherers (Elsevier), 341–363.
Price, T. D, and Feinman, G M. (1995). Foundations of social inequality, 1. Springer Science & Business Media.
Price, T. D, and Feinman, G M. (2010). “Social inequality and the evolution of human social organization,” in Pathways to power (Springer), 1–14.
Villar Quinteiro, R (1997). El Paleolítico Superior y Epipaleolítico en Galicia. Santiago de Compustela: Ph.D., Universidad de Santiago de Compustela.
Ramos, M, Francisco, J, Bernal Casasola, D, Fernández, V C, Arribas-Herrera, A, Bastir, M, et al. (2003). El abrigo y la cueva de Benzú en la Prehistoria de Ceuta. Aproximación al estudio de las sociedades cazadoras-recolectoras y tribales comunitarias en el ámbito norteafricano del Estrecho de Gibraltar. Cadiz: Servicio de Publicaciones de la Universidad de Cádiz.
RCoreTeam (2022). R: A language and environment for statistical computing. https://www.R-project.org/.
Renouf, M. A. P. (1988). Sedentary coastal hunter-Fishers: An example from the younger stone age of northern Norway. Archaeol. Prehist. Coastlines, 102–115.
Revelle, W (2015). Procedures for personality and psychological research. https://CRAN.R-project.org/package=psych Version = 2.2.5.
Ricciardi, A, Serrouya, R, and Whoriskey, F G. (1995). Aerial exposure tolerance off zebra and quagga mussels (Bivalvia: Dreissenidae): Implications for overland dispersal. Can. J. Fish. Aquat. Sci. 52 (3), 470–477. doi:10.1139/f95-048
Rivero, O (2015). Art mobilier des chasseurs magdaléniens de la façade atlantique: ERAUL, études et recherches archéologiques de l'Université de Liège.
Roberts, Richard G., Jones, Rhys, and Smith, Michael A. (1990). Thermoluminescence dating of a 50, 000-year-old human occupation site in northern Australia. Nature 345 (6271), 153–156. doi:10.1038/345153a0
Rodríguez-Hidalgo, A., Morales, J. I., Cebrià, A., Courtenay, L. A., Fernández-Marchena, J. L., García-Argudo, G., et al. (2019). The Châtelperronian Neanderthals of Cova Foradada (Calafell, Spain) used imperial eagle phalanges for symbolic purposes. Sci. Adv. 5 (11), eaax1984. doi:10.1126/sciadv.aax1984
Rose, J I., and Marks, A E. (2014). Out of arabia” and the middle-upper palaeolithic transition in the southern levant. Quartär 61, 49–85.
Rose, Jeffrey I., Usik, Vitaly I., Marks, Anthony E., Yamandu, H. Hilbert, Galletti, Christopher S., Parton, Ash, et al. (2011). The nubian complex of dhofar, Oman: An african middle stone age industry in southern arabia. PloS one 6 (11), e28239. doi:10.1371/journal.pone.0028239
Ruff, C. B., Trinkaus, E., and Holliday, T. W. (1997). Body mass and encephalization in Pleistocene Homo. Nature 387 (6629), 173–176. doi:10.1038/387173a0
Rüther, P L, Mirnes Husic, I, Bangsgaard, P, Murphy Gregersen, K, Pantmann, P, Carvalho, M, et al. (2022). Marie louise schjellerup jørkov, michael M. Benedetti, jonathan Haws, nuno Bicho, frido welker, enrico cappellini, and jesper velgaard OlsenSPIN enables high throughput species identification of archaeological bone by proteomics. Nat. Commun. 13 (1), 2458. doi:10.1038/s41467-022-30097-x
Sahlins, M (1968). “Notes on the original affluent society,” in Man the hunter. Editors Richard B. Lee, and Irven DeVore (New York: Aldine), 85–89.
Serrano Lozano, F., Lozano Francisco, M., Vera-Peláez, J., and Guerra Merchán, A. (1995). “Malacofauna en yacimientos prehistóricos de la Cueva de Nerja,” in Fauna de la Cueva de Nerja I (Nerja: Salas de la Mina y de la Torca, campañas).
Steele, T E., and Álvarez-Fernández, E (2011). “Initial investigations into the exploitation of coastal resources in North Africa during the Late Pleistocene at Grotte des Contrebandiers, Morocco,” in Trekking the shore. Editors N. Bicho, J. Haws, and L. Davis (Springer), 383–403.
Stiner, M C. (1994). Honor among thieves: A zooarchaeological study of neandertal ecology. Princeton: Princeton University Press.
Stiner, M. C., Munro, N. D., and Surovell, T. A. (2000). The tortoise and the hare. Curr. Anthropol. 41 (1), 39–79. doi:10.1086/300102
Storm, P, Wood, R, Stringer, C, Bartsiokas, A, de Vos, J., Aubert, M, et al. (2013). U-series and radiocarbon analyses of human and faunal remains from Wajak, Indonesia. J. Hum. Evol. 64 (5), 356–365. doi:10.1016/j.jhevol.2012.11.002
Straus, L, and Clark, G. A. (Editors) (1986). “La riera cave: Stone age hunter-gatherer adaptations in northern Spain,” Anthropological research papers No. 36 (Tempe: Arizona State University).
StrausGuy, L (2018). Environmental and cultural changes across the Pleistocene-Holocene transition in Cantabrian Spain. Quat. Int. 465, 222–233. doi:10.1016/j.quaint.2016.10.005
StrausGuy, L, ManuelMorales, R. G, and Cuenca-Solana, D (2015). The magdalenian human burial of el mirón cave (ramales de la Victoria, cantabria, Spain): Introduction, background, discovery and context. J. Archaeol. Sci. 60, 1–9. doi:10.1016/j.jas.2015.02.018
StrausGuy, Lawrence (2015). Recent developments in the study of the upper paleolithic of vasco-cantabrian Spain. Quat. Int. 364, 255–271. doi:10.1016/j.quaint.2014.05.008
StrausGuy, L (2005). The upper paleolithic of cantabrian Spain. Evol. Anthropol. 14 (4), 145–158. doi:10.1002/evan.20067
Stringer, C B., Clive Finlayson, J. R, Fernández-Jalvo, Yolanda, Cáceres, Isabel, Sabin, Richard C., Rhodes, Edward J., et al. (2008). Neanderthal exploitation of marine mammals in Gibraltar. Proc. Natl. Acad. Sci. U. S. A. 105 (38), 14319–14324. doi:10.1073/pnas.0805474105
Tátá, F, Cascalheira, J, Marreiros, J, Pereira, T, and Bicho, N (2014). Shell bead production in the upper paleolithic of Vale Boi (SW Portugal): An experimental perspective. J. Archaeol. Sci. 42, 29–41. doi:10.1016/j.jas.2013.10.029
Testart, Alain, Forbis, R. G., Hayden, B., Ingold, T., Perlman, S. M., Pokotylo, D. L., et al. (1982). The significance of food storage among hunter-gatherers: Residence patterns, population densities, and social inequalities [and comments and reply]. Curr. Anthropol. 23 (5), 523–537. doi:10.1086/202894
Thomas, D. H, and Koyama, S (1979). Affluent foragers: Pacific coasts East and West. Osaka: National Museum of Ethnology.
Thorne, A, Grün, R, Mortimer, G, Spooner, N. A., Simpson, J J., McCulloch, M, et al. (1999). Australia's oldest human remains: Age of the lake mungo 3 skeleton. J. Hum. Evol. 36 (6), 591–612. doi:10.1006/jhev.1999.0305
Tucker, J. K., Theiling, C. H., Janzen, F. J., and Paukstis, G. L. (1997). Sensitivity to aerial exposure: Potential of system-wide drawdowns to manage zebra mussels in the Mississippi river. Regul. Rivers Res. Mgmt. 13 (6), 479–487. doi:10.1002/(SICI)1099-1646(199711/12)13:6<479::AID-RRR469>3.0.CO;2-B
Uauy, R., and Dangour, A. D. (2006). Nutrition in brain development and aging: Role of essential fatty acids. Nutr. Rev. 64 (2), S24–S33. doi:10.1111/j.1753-4887.2006.tb00242.x
Vanhaeren, M, and D'Errico, F (2002). “The body ornaments associated with the burial,” in Portrait of the artist as a child. The gravettian human skeleton from the abrigo do lagar Velho and its archeological contex. Editors J. Zilhão E,, and E. Trinkaus (Lisbon: Portuguese Institute of Archaeology), 154–186.
Villa, P, Soriano, S, Pollarolo, L, Smriglio, C, Gaeta, M, D’orazio, M, et al. (2020). Neandertals on the beach: Use of marine resources at Grotta dei Moscerini (Latium, Italy). PloS one 15 (1), e0226690. doi:10.1371/journal.pone.0226690
Villagran, X., Klokler, D., Nishida, P., Gaspar, M., and Deblasis, P. (2010). Lecturas estratigráficas: Arquitectura funeraria y depositación de residuos en el sambaquí jabuticabeira II. Lat. Am. Antiq. 21, 195–216. doi:10.7183/1045-6635.21.2.195
Waechter, J. d'A. (1951). Excavations at gorham's cave, Gibraltar. Proc. Prehist. Soc. 17 (1), 83–92. doi:10.1017/S0079497X00018806
Washburn, S L., and Lancaster, G. S. (1968). “The evolution of hunting,” in Man the hunter. Editors d. Lee, and Irven DeVore (London: Routledge), 293–303.
Wedage, O, Picin, A, James, B, Douka, K, Deraniyagala, S, Kourampas, N, et al. (2019). Microliths in the south asian rainforest ∼45-4 ka: New insights from fa-hien lena cave, Sri Lanka. PLOS ONE 14 (10), e0222606. doi:10.1371/journal.pone.0222606
Wedage, O, Roberts, P, Faulkner, P, Crowther, A, Douka, K, Picin, A, et al. (2020). Late Pleistocene to early-holocene rainforest foraging in Sri Lanka: Multidisciplinary analysis at kitulgala beli-lena. Quat. Sci. Rev. 231, 106200. doi:10.1016/j.quascirev.2020.106200
Will, Ml, Kandel, A W., and Conard, N J. (2019). Midden or molehill: The role of coastal adaptations in human evolution and dispersal. J. World Prehist. 32 (1), 33–72. doi:10.1007/s10963-018-09127-4
Yesner, D R., Ayres, W S., Carlson, D L, Dewar, R, Morales, M, Hassan, Fekri A., et al. (1980). Maritime hunter-gatherers: Ecology and prehistory [and comments and reply]. Curr. Anthropol. 21 (6), 727–750. doi:10.1086/202568
Yesner, D. R. (1987). Food and evolution: Toward a theory of human food habits, 285–310.Life in the Garden of Eden: Causes and consequences of the adoption of marine diets by human societies
Yesner, D. R. (1998). Origins and development of maritime adaptations in the northwest pacific region of north America: A zooarchaeological perspective. Arct. Anthropol. 35 (1), 204–222.
Zilhão, J., Angelucci, D. E., Badal-Garcia, E., d'Errico, F., Daniel, F., Dayet, L., et al. (2010). Symbolic use of marine shells and mineral pigments by Iberian Neandertals. Proc. Natl. Acad. Sci. U. S. A. 107 (3), 1023–1028. doi:10.1073/pnas.0914088107
Keywords: middle paleolithic, upper paleolithic, hunter-gatherers, coastal adaptations, Iberia
Citation: Bicho N and Esteves E (2022) Pleistocene hunter-gatherer coastal adaptations in Atlantic Iberia. Front. Earth Sci. 10:957214. doi: 10.3389/feart.2022.957214
Received: 30 May 2022; Accepted: 11 July 2022;
Published: 25 August 2022.
Edited by:
Manuel Will, University of Tübingen, GermanyReviewed by:
Andrea Picin, Friedrich Schiller University Jena, GermanyCopyright © 2022 Bicho and Esteves. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nuno Bicho, bnVub2ZiaWNob0BnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.