 Qian Feng
Qian Feng Bing Pan
Bing Pan Aihua Yang1
Aihua Yang1- 1Centre for Research and Education on Biological Evolution and Environment, Nanjing University, Nanjing, China
- 2State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China
Small shelly fossils (SSFs) are a useful and critical tool for subdivision and correlation of the Cambrian Terreneuvian stratigraphy. The Maidiping Formation in southern Sichuan is a well-known Terreneuvian lithologic unit with rich SSFs. Here, the upper Maidiping Formation at the Fandian section, an important supplementary section of the classic Maidiping section, is studied for understanding its SSF assemblages and stratigraphic implication. There are 26 genera, 24 species, and 3 undetermined species recovered from the Fandian section, and Watsonella crosbyi was discovered from this section for the first time. Two SSF biozones of the Yangtze Platform, i.e., the possible Paragloborilus subglobosus–Purella squamulosa Assemblage Zone (? Zone II) and the W. crosbyi Assemblage Zone (Zone III), can be recognized. Comparing with the Maidiping section, the Fandian section has a relatively successive SSF biostratigraphy and carbon isotope stratigraphy in the upper Maidiping Formation. The SSF Zone II (?) and Zone III from the Fandian section can be, respectively, correlated to the same zones from the upper Maidiping Formation of the nearby Maidiping section (only Zone III), the upper Zhongyicun and Dahai members of the Zhujiaqing Formation in eastern Yunnan, the top Kuanchuanpu Formation in southern Shaanxi and northern Sichuan, and the beds 4 and 5 (or units 3 and 4) of the Yanjiahe Formation in western Hubei. The almost cosmopolitan distribution and nearly synchronous earliest appearance of W. crosbyi strengthen the applicability of its first appearance data (FAD) to define the base of Cambrian Stage 2 in South China (Xiaotanian) as well as in other continents. The FAD of W. crosbyi just around the onset of the ZHUCE carbon isotope excursion at the Fandian section also further reinforces the utility of the onset of ZHUCE as an auxiliary maker for defining the base of the Xiaotanian in South China.
Introduction
The current Cambrian chronostratigraphy consists of four series and ten stages (Babcock et al., 2005; International Stratigraphic Chart 2022). However, the criteria for the subdivision of the traditional Lower Cambrian, including Stage 2, 3 (Series 2) and 4, are still in debate. The Small Shelly Fossils (SSFs) are a group of micro-sized skeletons and sclerites belonging to multiple phyla. Considering their rapid evolution and worldwide distribution, the SSFs have been taken as a critical tool for the subdivision and correlation of the traditional Lower Cambrian, particularly for the Stage 2. The first appearance data (FADs) of two micro-molluscan taxa, i.e., Watsonella crosbyi Grabau, 1900 and Aldanella attleborensis Shaler and Foerste, 1888, have been proposed as the most potential candidates for the index fossils to define the global Cambrian Stage 2 (Zhu et al., 2006, 2008, 2019; Rozanov et al., 2008; Li et al., 2011; Parkhaev et al., 2011, 2012; Devaere et al., 2013; Jacquet et al., 2017; Kouchinsky et al., 2017). In addition to the SSFs, the acritarch Skiagia ornata (Volkova, 1968) has also been suggested to be another potential biostratigraphic maker to define the base of Stage 2 (Moczydłowska & Yin, 2012). On the other hand, the prominent global carbon isotope (δ13Ccarb) excursion ZHUCE (Zhujiaqing Carbon isotope Excursion) has also been proposed to be an important auxiliary maker for the Cambrian Stage 2 stratigraphic correlation (Zhu et al., 2006; Maloof et al., 2010; Landing et al., 2013). However, some researchers argue that carbon isotope curves can vary a lot among different local depositional environments (Steiner et al., 2020), and the utilization of interregional chemostratigraphic correlations in the Cambrian should be based on robust biostratigraphic constraint (Kouchinsky et al., 2017; Betts et al., 2018; Steiner et al., 2020; Guo et al., 2021), which is preferred herein.
For the well and relatively successive record of carbonate rocks, the Yangtze Platform is one of the best regions to study the early Cambrian (especially Terreneuvian) SSF biostratigraphy and the carbon isotope stratigraphy (Brasier et al., 1990; Zhou et al., 1997; Ishikawa et al., 2008; Li et al., 2009, 2013). The eastern Yunnan area yields the most complete Terreneuvian strata (the Zhujiaqing and Shiyantou formations) and many well exposed sections (Meishucun, Xianfeng, Laolin, Lishuping, Xiaotan, etc., in Figure 1A). The stratigraphy (litho-, bio- and chemo-) therein was well studied, and four Terreneuvian SSF biozones were established: the Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone (Zone Ⅰ), the Paragloborilus subglobosus–Purella squamulosa Assemblage Zone (Zone Ⅱ), the Watsonella crosbyi Assemblage Zone (Zone Ⅲ) and the Sinosachites flabelliformis–Tannuolina zhangwentangi Assemblage Zone (Zone IV), which have been widely used as the standard Terreneuvian biostratigraphic framework for the Yangtze Platform (Qian et al., 1999; Steiner et al., 2004, 2007, 2020; Guo et al., 2014, 2021; Yang et al., 2014, 2016; Zhu et al., 2019). However, due to the variations in sedimentary records and palaeogeographic conditions, the early Cambrian SSF assemblages also vary to some extent among different regions. Steiner et al. (2004, 2007, 2020; Yang & Steiner, 2021) argued that the SSF Zone Ⅱ is hard to recognize or even missing in many sections on the Yangtze Platform, such as in western Hubei, southern Shaanxi, and southern Sichuan, due to the hiatus of sedimentation, the poor preservation of fossils, and the rare occurrence of the index fossil, Purella squamulosa. Unlike Zone Ⅱ, Zone Ⅲ has been well recognized in all the main regions of the Yangtze Platform (Steiner et al., 2004, 2007, 2020; Li et al., 2011; Guo et al., 2014; Guo et al., 2021; 2022; Yang et al., 2014; Yang & Steiner, 2021). Therefore, Zhu et al. (2019) formally used the FAD of W. crosbyi at the base of the Dahai Member of the Zhujiaqing Formation in the Xiaotan section (Yongshan County, Yunnan; see Li et al., 2001) to define the newly named Xiaotanian Stage, replacing the Meishucunian as the Cambrian Stage 2 in China. In this stratotype section, the FAD of W. crosbyi is stratigraphically at the onset of the ZHUCE positive δ13C excursion, which can also be well recognized at the Laolin and Lishuping sections (Zhou et al., 1997; Li D. et al., 2009, 2013; Li G. et al., 2011; Yang et al., 2014; Steiner et al., 2020). But it is still noteworthy that Zone Ⅲ can only be recognized from one single bed (1–2 m vs. ca. 80 m in the Xiaotan section) from a few sections in western Hubei and southern Shaanxi (Xing et al., 1984; Steiner et al., 2004, 2007, 2020; Guo et al., 2014, 2021), indicating that the strata are severely condensed and/or bear hiatuses of different durations in these areas. Accordingly, the correlation of the ZHUCE positive δ13C excursion is problematic in western Hubei considering the SSF biostratigraphy (Ishikawa et al., 2008; Jiang et al., 2012; Guo et al., 2014, 2021; Steiner et al., 2020). While, in southern Shaanxi the ZHUCE is even yet to be recognized (Steiner et al., 2020). Yang & Steiner (2021) drew the Cambrian Terreneuvian framework on SSF biostratigraphy and carbon isotope chemostratigraphy in the classical Maidiping section, southern Sichuan. But due to a hiatus in the upper part of the Maidiping Formation, the Paragloborilus subglobosus–Purella squamulosa Assemblage Zone (Zone II), the base of the W. crosbyi Assemblage Zone (Zone Ⅲ), and the onset of ZHUCE positive δ13C excursion cannot be recognized perfectly. More studies of the sections with relatively continuous strata are required to verify the detailed correlations between the ZHUCE excursion and the FAD of W. crosbyi outside the classical Maidiping section in southern Sichuan. Fortunately, the Fandian section was relatively continuous and rich in Terreneuvian SSFs in the Maidiping Formation (Yin et al., 1980; He et al., 1984). But these fossils have not been described or illustrated. Thus, in 2014 and 2019, we systematically collected the SSF and carbon isotope samples from the exposed part of the Maidiping Formation at the Fandian section for detailed biostratigraphic and chemostratigraphic research. In this article, we will document the research results: the Zone II (?), Zone Ⅲ, and ZHUCE positive δ13C excursion are recognized in the upper Maidiping Formation, Fandian section, which provides a valuable supplement to the classical Maidiping section.
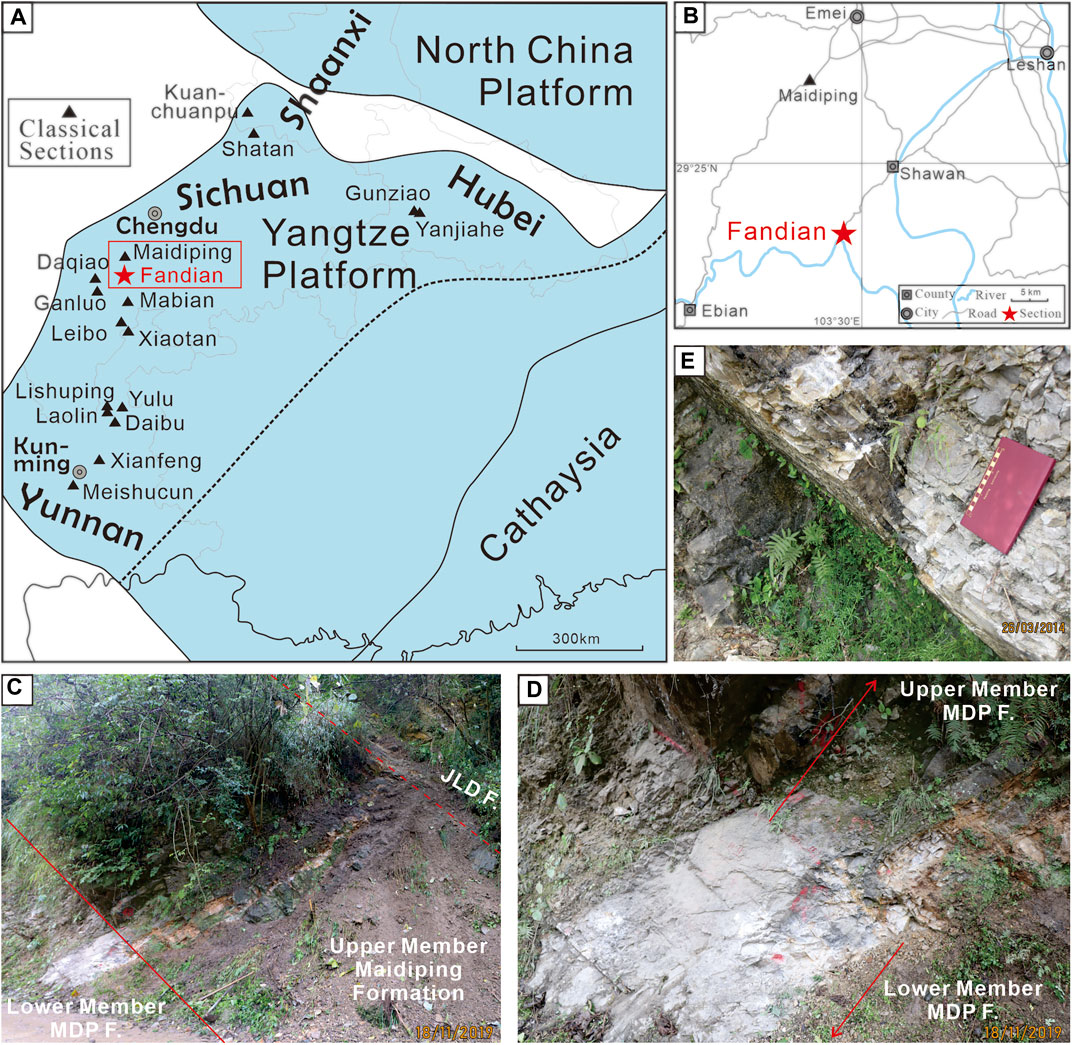
FIGURE 1. Location map and field photographs of the Fandian section. (A) General location of the Fandian section and other classical sections with diverse small shelly fossils on the western Yangtze Platform (modified from Steiner et al., 2007). (B) Traffic map showing the detailed location of the Fandian section. (C) Overview of the upper part of the Fandian section. (D) Boundary of the lower and upper members of the Maidiping Formation (MDP F.). (E) Gray dolostone with a few thin chert layers of the Lower Member of the Maidiping Formation. Abbreviation: JLD F., Jiulaodong Formation.
Materials and Methods
The samples for the small shelly fossils study are all collected from the Maidiping Formation at the Fandian section (103°28′1″ E, 29°19′2″ N), which outcrops along the west side of the river nearby the center of the previous Fandian Town (now belonging to the Shawan Town) of Leshan City, 30 km away from the classical Maidiping section at Gaoqiao Town, Emei City (Figure 1). In the previous description, the whole Maidiping Formation at the Fandian section was well exposed (He et al., 1984). However, during our investigation, the basal part of the Maidiping Formation was covered by the weathering residues and vegetations, and only the upper 11.2 m of the Lower Member is well exposed. Thus, the boundary between the Maidiping Formation and its underlying Dengying Formation is unknown. The uncovered Lower Member of the Maidiping Formation mainly consists of the medium-thin gray dolostone interlayered with a few thin chert layers. The well-exposed Upper Member of the Maidiping Formation (about 9 m-thick) conformably contacts with the Lower Member and is mainly composed of the gray-yellow dolostone containing abundant bioclasts and collophanite sands. The shale-dominated Jiulaodong Formation disconformably overlies the Maidiping Formation with a 20 cm-thick weathered yellow clay bed at its base.
The rock samples (each weighing ca. 3–5 kg) for the SSF study were processed with 5% acetic acid to get the undissolved residues. The small shelly fossil specimens were manually picked from the residues under a binocular microscope. The fossil specimens were bonded to aluminum sample stubs by using double-sided carbon glue for gold coating and scanning electron microscopy (SEM) photography. The SEM photos were all taken through HITACHI SU3500 at the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (NIGPAS). All specimens illustrated in this article are catalogued and housed in the storage facilities at NIGPAS.
All samples for carbon and oxygen isotopic analyses are collected from fresh, relatively unweathered carbonate rocks. Each sample was broken into small fragments first, and then used a diamond drill bit to drill on the broken surface and get c. 80 mg of fine powder. The micro-areas for drilling should be carefully selected under the microscope to avoid the calcite veins, recrystallized areas, and weathered rims. Twenty milligrams of carbonate powder from each sample were used for analysis. Limestone powders were reacted with 100% H3PO4 at 25°C for more than 12 h, and dolomite powders were reacted with 100% H3PO4 at 50°C for more than 24 h. Prepared gas samples were analyzed for δ13C and δ18O by a Finnegan MAT 253 mass spectrometer using the Chinese national standard, an Ordovician carbonate from a site near Beijing (reference number GBW 04405: δ13C = 0.57 ± 0.03‰ VPDB; δ18O = −8.49 ± 0.13‰ VPDB). All the above experiments and analysis processes were conducted in the laboratory of the NIGPAS. The results of δ13C and δ18O of the samples collected from the Maidiping Formation of the Fandian section (47 samples) and the Maidiping and Dengying formations of the Daqiao section (110 samples) are listed herein in the Supplementary Materials with linear-regression analysis and can also be acquired from Xiong et al. (2022, Table 1).
SSF Assemblages at the Fandian Section
The exposed upper part (0–11.2 m herein) of the Lower Member of the Maidiping Formation at the Fandian section consists of a monotonous dolostone intercalated by some thin chert layers (Figure 1E). After the acetic acid (5–10%) processing, there is no fossil found from the dolostone samples (Figure 2) from this interval. In contrast, many small shelly fossil specimens were obtained from the samples in the ca. 8 m-thick interval (12.1–20.2 m) of the Upper Member (11.2–20.2 m) of the Maidiping Formation. Based on the systematic study, we distinguished 26 genera, 24 species, and 3 unidentified species (Figures 3, 4), mainly including the hyolithids: Conotheca subcurvata Yu, 1974, Paragloborilus subglobosus He in Qian, 1977, Annelitellus yangtzensis Qian, 1989, Cupitheca mira He in Qian, 1977; the molluscs and cap-like fossils: Obtusoconus rostriptuetus Qian, 1978, Oelandiella korobkovi Vostokova, 1962, Igorella maidipingensis Yu, 1974, Bemella simplex Yu, 1979, Helcionella explanata Feng et al., 2000, Watsonella crosbyi Grabau, 1900, Lathamella caeca Liu, 1979, Aegitellus emeishanensis He in Yin et al., 1980, Securiconus simus Jiang, 1980; cf. Maikhanella sp., Dorispira lauta Yu, 1979, Asiapatella sinuata Parkhaev et al., 2011, Ocruranus finial Liu, 1979, Emarginoconus mirus Yu, 1979; the siphogonuchitids: Siphogonuchites triangularis Qian, 1977, Lopochites latazonalis Qian, 1977; the chancelloriids: Chancelloria eros Walcott, 1920; the zhijinitiids: Zhijinites longistriatus Qian, 1978, the paracarinachitids: Paracarinachites sinensis Qian and Jiang in Luo et al., 1982; the halkieriids: Halkieria sacciformis Meshkova, 1969; the protoconodonts: Protohertzina sp.; and the sclerites of problematic lineages: Kuanchuanella sp., Brushenodus prionodes Luo et al., 1982. Their stratigraphic distributions are shown in Figure 2.
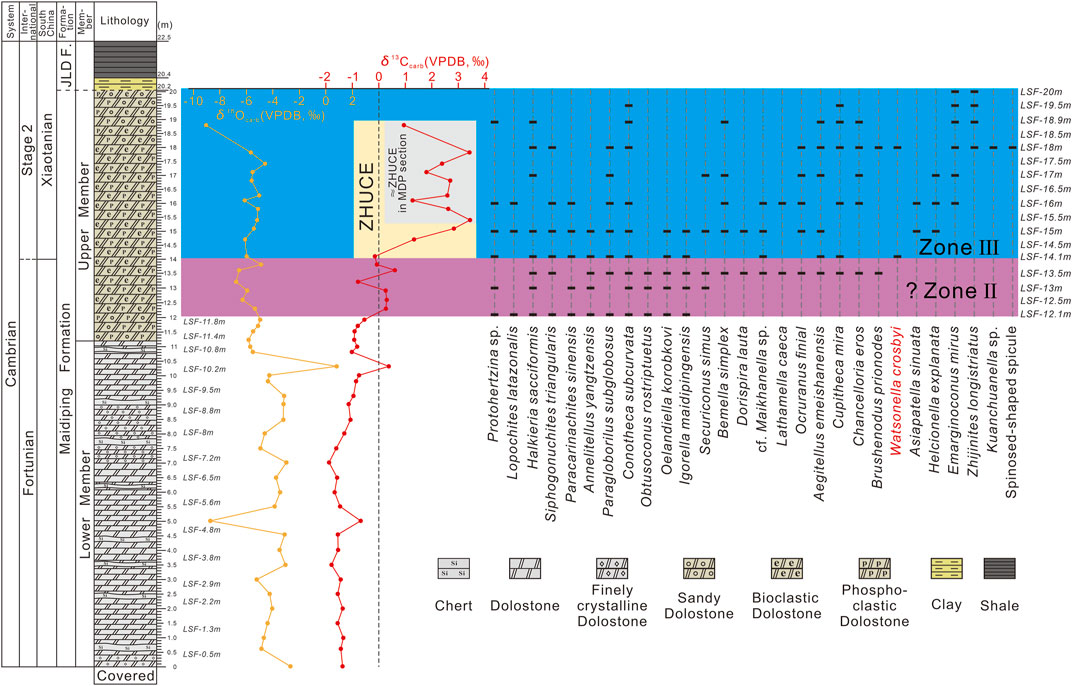
FIGURE 2. Stratigraphic column of the Fandian section with the distribution of small shelly fossils and the profile of δ13C and δ18O values of carbonate in the Maidiping Formation. Abbreviation: JLD F., Jiulaodong Formation; MDP, Maidiping; LSF, samples collected for the study of small shelly fossil at the Fandian section in Leshan City (Chronostratigraphy based on Steiner et al., 2007, 2020; Zhu et al., 2019).
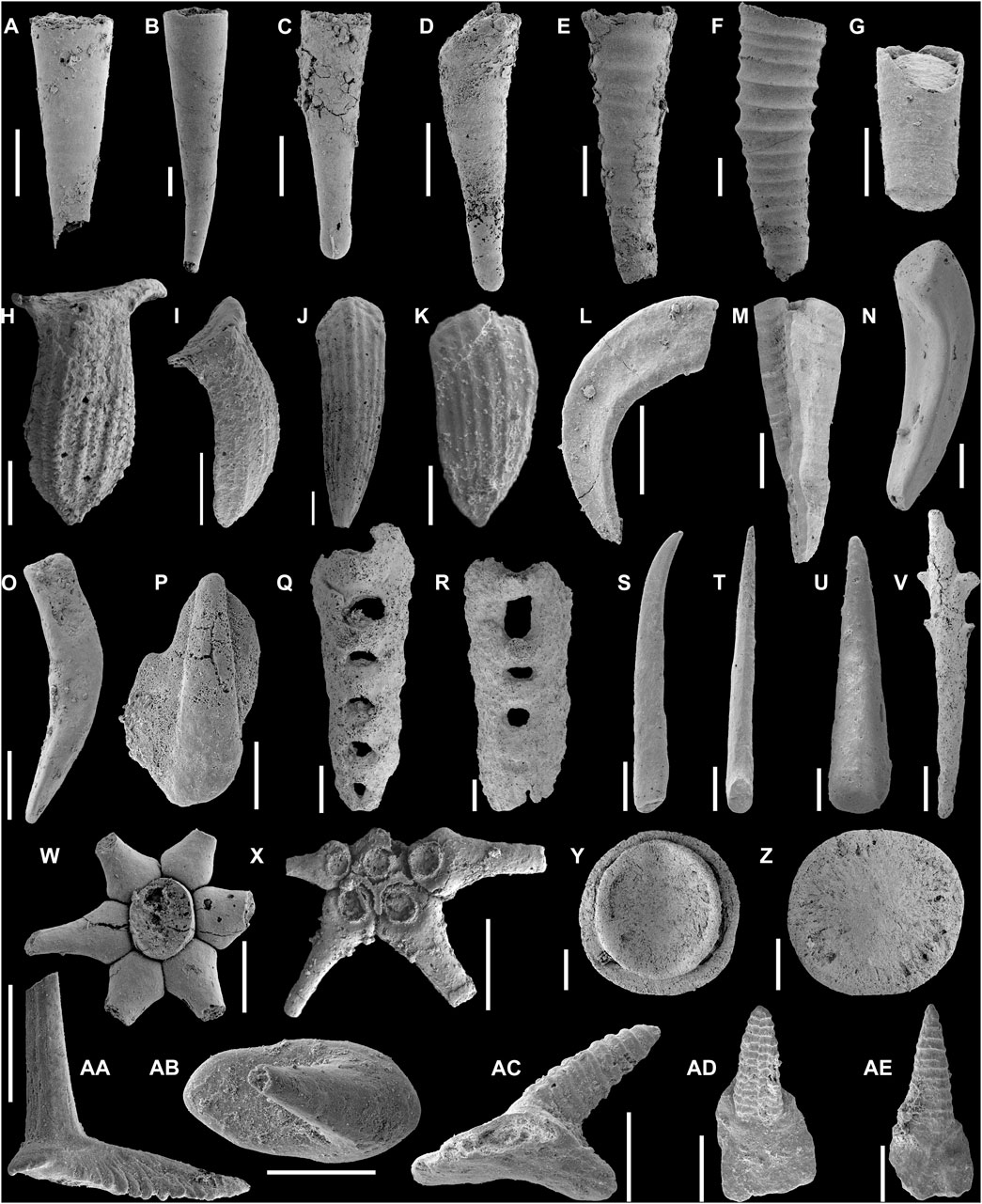
FIGURE 3. Small shelly fossils from the upper Maidiping Formation at the Fandian section. (A,B) Conotheca subcurvata (Yu, 1974); (A) NIGPAS 180248, LSF-13.5m, lateral view; (B) NIGPAS 180249, LSF-13.5m, lateral view. (C,D) Paragloborilus subglobosus Qian, 1977; (C) NIGPAS 180250, LSF-13.5m, lateral view; (D) NIGPAS 180251, LSF-17m, lateral view. (E,F) Annelitellus yangtzensis Qian, 1989; (E) NIGPAS 180252, LSF-13m, lateral view; (F) NIGPAS 180253, LSF-13m, lateral view. (G) Cupitheca mira He in Qian, 1977, NIGPAS 180254, LSF-18m, lateral view. (H–K) Halkieria sacciformis Meshkova, 1969; (H), NIGPAS 180255, LSF-12.1m, upper surface; (I).NIGPAS 180256, LSF-18m, lateral upper side; (J) NIGPAS 180257, LSF-13.5m, upper surface; (K) NIGPAS 180258, LSF-13m, upper surface. (L–N) Siphogonuchites triangularis Qian, 1977; (L) NIGPAS 180259 and LSF-18m, lower surface; (M) NIGPAS 180260, LSF-15m, upper surface; (N) NIGPAS 180261, LSF-16m, upper surface. (O) Lopochites latazonalis Qian, 1977, NIGPAS 180262, LSF-12.1m, lateral view. (P) Kuanchuanella sp., NIGPAS 180263, LSF-18m, distal view. (Q,R) Paracarinachites sinensis Qian and Jiang in Luo et al., 1982; (Q) NIGPAS 180264, LSF-12.1m, distal view; (R) NIGPAS 180265, LSF-12.1m, distal view. (S,T) Protohertzina sp.; (S) NIGPAS 180266, LSF-15m, lateral view; (T) NIGPAS 180267, LSF-15m, proximal view. (U) Spinose-shaped fossils, NIGPAS 180268, LSF-18m, lateral view. (V) Brushenodus prionodes (Luo et al., 1982), NIGPAS 180269, LSF-13.5m, lateral view. (W,X) Chancelloria eros Walcott, 1920; (W) NIGPAS 180270, LSF-13.5m, distal view; (X) NIGPAS 180271, LSF-13.5m, basal view. (Y,Z) Emarginoconus mirus Yu, 1979; (Y) NIGPAS 180272, LSF-18m, inner view; (Z) NIGPAS 180273, LSF-18m, outer view. (AA–AE),Zhijinites longistriatus Qian, 1978. (AA) NIGPAS 180274, LSF-18m, lateral view; (AB) NIGPAS 180275, LSF-18m, apical view; (AC) NIGPAS 180276, LSF-18m, lateral view; (AD) NIGPAS 180277, LSF-18m, distal view; (AE) NIGPAS 180278, LSF-18m, distal view. Scale bars: 300 μm.

FIGURE 4. Small shelly fossils from the upper Maidiping Formation at the Fandian section. (A,B) Igorella maidipingensis (Yu, 1974), NIGPAS 180279, LSF-12.1m; (A) lateral view; (B) dorsal view. (C,D) Oelandiella korobkovi Vostokova, 1962; (C) NIGPAS 180280, LSF-12.1m, lateral view; (D) NIGPAS 180281, LSF-13m, lateral-apertural view. (E) Aegitellus emeishanensis (He in Yin et al., 1980), NIGPAS 180282, LSF-13.5m, apical view. (F–I) Watsonella crosbyi Grabau, 1900; (E) NIGPAS 180283, LSF-18m, lateral view; (F) NIGPAS 180284, LSF-18m, lateral view; (G) NIGPAS 180285, LSF-18m, dorsal view; (H) NIGPAS 180286, LSF-18m, apertural view. (J) Lathamella caeca Liu, 1979, NIGPAS 180287, LSF-16m, dorsal view. (K), Asiapatella sinuata Parkhaev et al., 2011, NIGPAS 180288, LSF-16m, apical view. (L, M) Obtusoconus rostriptuetus (Qian, 1978); (L) NIGPAS 180289, LSF-12.1m, lateral view; (M) NIGPAS 180290, LSF-13.5m, apical view. (N) Securiconus simus Jiang, 1980, NIGPAS 180291, LSF-13.5m, lateral view. (O) cf. Maikhanella sp., NIGPAS 180292, LSF-16m, fragment of the shell. (P) Bemella simplex Yu, 1979, NIGPAS 180293, LSF-13.5m, dorsal view. (Q) Helcionella explanata (Feng et al., 2000), NIGPAS 180294, LSF-16m, dorsal view. (R) Ocruranus finial Liu, 1979, NIGPAS 180295, LSF-13.5m, dorsal view. (S) Dorispira lauta (Yu, 1979), NIGPAS 180296, LSF-13.5m, dorsal view. Scale bars: 300 μm.
Herein, the SSF assemblages from the Upper Member of the Maidiping Formation at Fandian section are temporarily assigned to the SSF Zone Ⅱ (?) and Zone Ⅲ of the Yangtze Platform (Figures 2, 5).
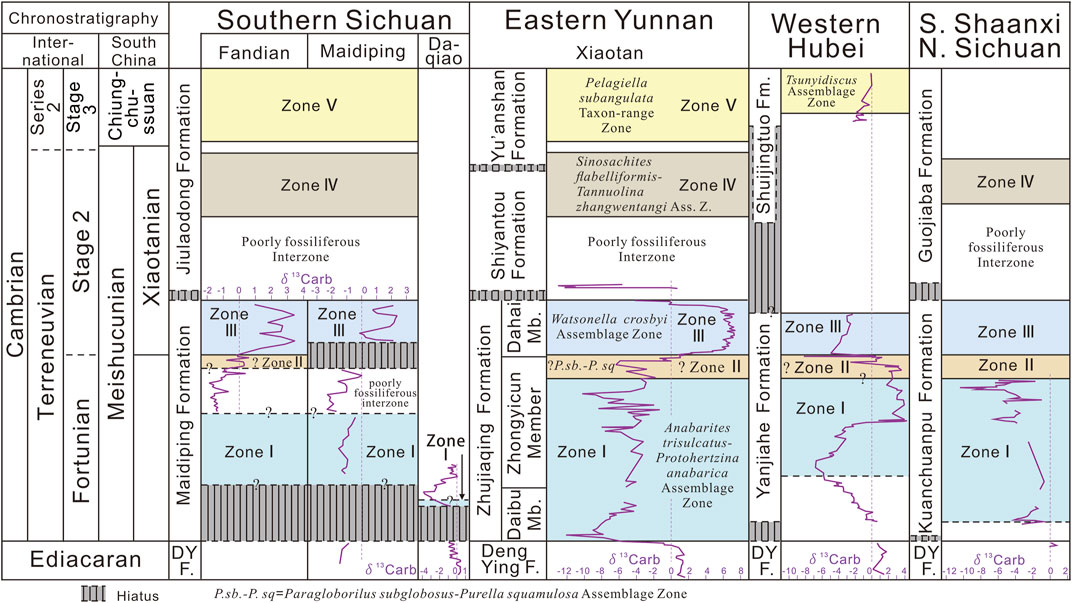
FIGURE 5. Terreneuvian SSF biozonation of the Fandian section compared with the Maidiping and Daqiao sections in southern Sichuan and other critical regions on the Yangtze Platform (modified from Steiner et al., 2020; chronostratigraphy of South China referring to Zhu et al., 2019; Steiner et al., 2020). SSF Zone II in western Hubei is according to Guo et al. (2014, 2021). The isotope curve of δ13Ccarb from each section is calibrated based on the SSF biozones. Isotope data of Fandian and Daqiao sections from Xiong et al. (2022, in press; Supplementary file herein), of Maidiping section from Yang & Steiner (2021), of Xiaotan section from Li et al. (2013), of western Hubei from Ishikawa et al. (2008, the Aijiahe/Gunziao section), and of the southern Shaanxi and northern Sichuan from Steiner et al. (2020).
? Paragloborilus subglobosus–Purella squamulosa Assemblage Zone
Luo et al. (1980) first proposed the Paragloborilus–Siphogonuchites Zone as the second Cambrian SSF Zone in the eastern Yunnan area. However, the significant problem of this zone is that all the index fossils can range into other biozones. In particular, S. triangularis has an extremely long range from Zone Ⅰ to Ⅲ. Thus, Steiner et al. (2007) established the Paragloborilus subglobosus–Purella squamulosa Assemblage Zone using the lowermost occurrence of P. subglobosus and Zhijinites longistriatus to define the lower boundary and the highest occurrence of P. squamulosa to define the upper boundary. Yang et al. (2014) excluded Z. longistriatus as the index fossil for its occurrence only in Zone Ⅲ and reassigned Oelandiella korobkovi (index fossil of Zone Ⅲ in Steiner et al., 2007) as the index fossil of Zone II for its most common occurrence in Zone II and only a few occurrences in Zone Ⅲ. It’s worth noting that all the index fossils except P. squamulosa (or Purella) can range into Zone Ⅲ. In addition, on the Yangtze Platform, P. squamulosa is not a wide-occurring species, and is only reported from a few sections in eastern Yunnan (Xiaotan, Xianfeng, and only P. elegans at Laolin) and northern Sichuan (Shatan) (Steiner et al., 2004, 2007; Yang et al., 2014). In western Hubei, only another species, P. antiqua, had been reported by Guo et al. (2014), but it was reinterpreted as maikhanellid sclerites by Steiner et al. (2020). All the above facts have made it difficult and problematical to recognize Zone II in the shallow facies on the Yangtze Platform, especially in southern Sichuan and western Hubei (Steiner et al., 2004, 2007, 2020; Yang et al., 2014; Yang and Steiner, 2021). Anyway, it’s still practicable to recognize Zone II based on the fossil combinations and their occurrences below the FAD of W. crosbyi in some sections with relatively successive strata (Meishucun, Lishuping, and Laolin section, Yang et al., 2014).
At the Fandian section, the SSF assemblage recovered from the 12.1–14 m interval of the basal Upper Member of the Maidiping Formation is temporarily assigned to the problematic Zone II (?) for the lack of Purella. Firstly, all the fossil elements in this interval are common in Zone II in eastern Yunnan, such as P. subglobosus, O. korobkovi, A. yangtzensis, O. rostriptuetus, cf. Maikhanella sp, B. simplex, C. subcurvata, I. maidipingensis, S. triangularis, P. sinensis, and Halkieria (Figure 2). Secondly, none of the three index fossils of Zone Ⅲ (FAD of W. crosbyi at 14.1 m; FAD of Z. longistriatus at 18 m; and no discovery of Aldanella attleborensis) occurs in this interval. Thirdly, the results of our continuous high-resolution carbon isotope and field observation show that there is no obvious hiatus or disconformity in the Upper Member of the Maidiping Formation (11.2–20.2 m) and this Zone (12.1–14 m) is below the onset of the ZHUCE excursion (Figure 2), which is consistent with the typical sections in eastern Yunnan (Li et al., 2009; 2013; Yang et al., 2014; Steiner et al., 2020; Figure 5). However, considering the much lower thickness of this zone compared with that in the sections in eastern Yunnan (Yang et al., 2014) and the lack of fossils below 12.1 m at the Fandian section, this problematic Zone II (?) at the Fandian section probably is not complete and lacks its lower interval of unknown thickness.
Watsonella crosbyi Assemblage Zone
This fossil assemblage was first proposed by Qian et al. (1996) as the Heraultipegma yunnanensis Zone, but it was not defined in much detail. Later, Steiner et al. (2007) considered that H. yunnanensis was a junior synonym of W. crosbyi and therefore renamed this zone as the W. crosbyi Assemblage Zone, with the lower boundary defined by the first appearance of W. crosbyi and the upper boundary drawn by the highest occurrence of Aldanella yanjiahensis (a junior synonym of Aldanella attleborensis) and Oelandiella korobkovi. Yang et al. (2014) added Zhijinites longistriatus as a new index fossil of Zone Ⅲ for its FAD almost the same or slightly later than W. crosbyi and excluded O. korobkovi for its common occurrence in Zone II rather than Zone Ⅲ. In addition, Yang et al. (2014) also pointed out that the operculum-like fossil, Emarginoconus mirus, occurred only in Zone Ⅲ and may be another potential auxiliary index fossil, consistent with its stratigraphic distribution observed herein.
In the previous studies, W. crosbyi had not been reported from the Fandian section (Yin et al., 1980; He et al., 1984). Herein, W. crosbyi was recovered from the two horizons of the upper member of the Maidiping Formation (8 specimens in total from horizons 14.1 m and 18 m in Figure 2; Figures 4F–I). In addition, the lowest occurrences of Z. longistriatus (Figures 3AA–AE) and E. mirus (Figures 3Y,X) are respectively at 18 m and 16 m and are higher than that of W. crosbyi, being consistent with their stratigraphic sequences in eastern Yunnan. It’s noteworthy to mention that samples from horizon 13.5 m yield many newly occurring species compared with the underlying samples. But most of these species are still typical elements of Zone II in eastern Yunnan. The few specimens of W. crosbyi found in the Fandian section (only one at 14.1 m and seven at 18 m) may suggest that W. crosbyi could also exist below its lowest occurrence at 14.1 m, where it is hard to find for its rarity. Considering no hiatus was observed in the interval between 12.1 and 14.1 m, in the future more sample collection with much higher resolution for SSF extraction in this interval is a necessary and reliable way to check this assumption. At present, we temporarily prefer to place the lower boundary of the SSF Zone Ⅲ at the lowermost occurrence horizon of W. crosbyi (14.1 m) at the Fandian section.
Yang and Steiner, (2021) mentioned that many SSFs, such as Annelitellus, Obtusoconus, Ocruranus, and Paracarinachites, previously discovered only in Zone II in eastern Yunnan, can range upwards to Zone Ⅲ in the adjacent Maidiping section. This situation is similar to that in the Fandian section (Figure 2). Yang and Steiner (2021) gave two possible explanations for this fact: either all these fossils indeed have a higher stratigraphic range than previously discovered in eastern Yunnan; or these fossils were redeposited by the reworking of the sediment from the older strata. Herein, we prefer the former explanation because there is no distinct difference in fossil preservation between SSF specimens in Zone II (?) and Zone Ⅲ in the Fandian section. In addition, our own unpublished data from the Xiaotan section in eastern Yunnan also shows that the basal Zone Ⅲ contains such fossil elements that previously only occurred in Zone II. It's necessary to point out that the specimens preserved as steinkerns and generally named as Protohertzina sp. nearly occurred in all the samples regardless of their rare number in Zone II and Ⅲ (Figure 2; Figures 3S,T). This is quite different from other sections in the Yunnan and Sichuan regions. Herein, we prefer to assign them to some other younger species of Protohertzina rather than to the older species like Protohertzina unguliformis.
Possible Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone
Although the basal part of the Maidiping section at the Fandian section is now absolutely covered by weathering residues and vegetations, but in the description of He et al. (1984, p. 73), the Maidiping Formation was fully exposed with the ca. 29.5m-thick Lower Member, and the basal ca. 6 m interval contains a few fossils, such as Anabarites trisulcatus, Conotheca, and Archaeooides, which are typical elements of the SSF Zone Ⅰ on the Yangtze Platform. Thus, we infer that the basal part of the Maidiping Formation at the Fandian section yields the Anabarites trisulcatus–Protohertzina anabarica Assemblage Zone.
Stratigraphic Correlation Within the Yangtze Platform and With Other Continents
With the Adjacent Maidiping and Daqiao Sections
As one of the most classical sections in southern Sichuan for the study of early Cambrian stratigraphy of the Yangtze Platform, the bio- and chemo-stratigraphy from the Maidiping section had got quite detailed studies by the pioneer researchers (Qian, 1977, 1989; Yin et al., 1980; He et al., 1984; Brasier et al., 1990; Qian et al., 1999). In the review of the SSF biostratigraphy of China, Qian et al. (1999, p.79-80, Figures 3, 4) subdivided the Maidiping Formation into 3 SSF biozones (Zones Ⅰ, II, Ⅲ) at the Maidiping section. However, recently, Yang and Steiner (2021) consider that the top 8 m (bed 36–39) of the Maidiping Formation, corresponding to the Zones II and Ⅲ in Qian et al. (1999), should only belong to the Zone Ⅲ, because below the lowest occurrence of Watsonella crosbyi, there is no Purella squamulosa in the less than 1m-thick interval of Zone II in Qian et al. (1999), and the fossil composition of this interval is similar to the above strata (Yang & Steiner, 2021). In addition, a disconformity has been observed below the base of the SSF Zone Ⅲ, and it can also be confirmed by the abrupt excursion of the δ13C carbon isotope (from –1.34‰ to +2.12‰ in Figure 2 of Yang & Steiner, 2021). This disconformity and the below 12m-thick dolostone with barren fossils (bed 35) might correlate to Zone II based on the restriction of the well-defined Zone Ⅰ and Ⅲ (Yang & Steiner, 2021). While in the Fandian section, no such hiatus or disconformity has been found in the Maidiping Formation. The δ13C value of the Fandian section (Figure 2, original data see Supplementary Material and Table 1 in Xiong et al., Forthcoming 2022) is relatively stable among –2‰ to –1‰ in the Lower Member (0–11 m) of the Maidiping Formation; gradually increases to about 0‰ (11–12.3 m) at the basal part of the Upper Member and keeps steady around 0‰ in a short interval (12.3–14 m, Zone II) below the lowest occurrence of Watsonella crosbyi; then from the base of the Zone Ⅲ the δ13C value increases fast but gradually from –0.141‰ to +3.432‰ (14.1–15.4 m) and oscillates between +0.941‰ and +3.432‰ in the rest part of the Upper Member (15.4–18.8 m). The δ13C value of the Fandian section records the relatively complete ZHUCE, while the ZHUCE in the Maidiping section, recognized by Yang & Steiner (2021, Figure 2), is probably equal to the top oscillating interval (14.1–15.4 m) of the Fandian section (Figures 2, 5). If the correlation of this positive excursion interval between the Fandian and Maidiping sections is the real case, the earliest/lowest occurrence of Watsonella crosbyi in the Maidiping section could be slightly later/higher than that in the Fandian section herein. It’s worthy to mention that both the maximum of this positive excursion at Fandian (+3.432‰) and Maidiping (+2.4‰) sections are much smaller than that of the Xiaotan section (+7.2‰ in Li et al., 2013), the referential section in eastern Yunnan, but close to that of the Lishuping (+2.69‰ in Steiner et al., 2020) and Laolin (+3.5‰ in Li et al., 2009) sections (Yang & Steiner, 2021). This may be due to the discrepancies in the local water environment as emphasized by Steiner et al. (2020). At least such positive excursion trends in these sections are comparable by the restriction of the SSF assemblage. The basal 11.2m-thick gray dolostone with no discovery of fossils at the Fandian section herein (same as He et al., 1984, p. 73) is comparable to the 12m-thick monotonous dolostone (bed 35) (containing rare indeterminate fossil fragments) at the Maidiping section described by Yang & Steiner (2021). For the lack of useful fossils, it is difficult to determine the age of this interval, which may correlate to either the lower part of Zone II or the upper part of Zone Ⅰ. In the description of the Fandian section by He et al. (1984, p. 73), the basal ca. 6m-thick interval of the fully exposed Maidiping Formation bears some typical fossils of the SSF Zone Ⅰ, such as Anabarites trisulcatus, Conotheca and Archaeooides. Thus, the basal Maidiping Formation at the Fandian section may also contain the SSF Zone Ⅰ and correlate to the beds 31 to 32 (or 34) of the Maidiping section (Figure 5). Rugatotheca and Ganloudina, together with both index fossils of the SSF Zone Ⅰ (P. anabarica and A. trisulcatus), have been reported from the basal Maidiping Formation at the Maidiping section (Yang and Steiner, 2021, Figures 2, 3). In eastern Yunnan, both P. anabarica and A. trisulcatus occurred not lower than the base of the Zhongyicun Member of the Zhujiaqing Formation and above the BACE (Basal Cambrian Carbon isotope Excursion) (Li et al., 2001; Steiner et al., 2007; Li et al., 2009, 2013; Yang et al., 2014). While the δ13C curve above the base of the Maidiping Formation from the Maidiping section is relatively steady and does not show the clear BACE (Yang & Steiner, 2021, Figure 2). Thus, we consider the base of the Maidiping Formation of the Maidiping section is probably above the base of the Zhongyicun Member at the Xiaotan section (reference Terreneuvian section in eastern Yunnan). However, in the adjacent Daqiao section, the basal Maidiping Formation (about 1 m) bears some common elements (Rugatotheca, Ganloudina, Olivooides) of the fossil assemblage at the base of the Daibu Member of the Zhujiaqing Formation at the Xiaotan section (Yang et al., 2016, Figure 1), together with Paracanthodus, Quadrosiphogonuchites, and Aetholicopalla and the possible P. anabarica and A. trisulcatus (Yang and Steiner, 2021, Figure 3). Furthermore, the δ13C results from the Daqiao section show a prominent negative excursion at the basal part of the Maidiping Formation, which may be correlated with the BACE at the Xiaotan section (Figure 5; Wusihe-Daqiao section in Xiong et al., Forthcoming 2022). Although the absolute value of the nadirs between Daqiao (−3.861‰) and Xiaotan (−12‰ in Li et al., 2013) sections shows quite a large discrepancy, this may be caused by the difference in regional aquafacies (see detailed discussion in Steiner et al., 2020). However, it’s also possible that this negative excursion of the Daqiao section could be compared one interval with the less negative δ13C curve of the upper Zone Ⅰ at the Xiaotan section, because Yang and Steiner (2021) mentioned there is an obvious hiatus (karst surface) at the boundary between the Maidiping and Dengying formations at the Daqiao section. Combining the above bio- and chemo-stratigraphic evidence, we tentatively infer that the base of the Maidiping Formation at the Daqiao section may roughly correlate to the upper Daibu Member of the Zhujiaqing Formation at the Xiaotan section (Figure 5).
With the Eastern Yunnan, the Western Hubei, and the Northern Sichuan and Southern Shaanxi areas
Three SSF zones (Zone Ⅰ to Ⅲ), the BACE and ZHUCE carbon isotope excursions can be well recognized from the Zhujiaqing Formation in eastern Yunnan (Qian, 1989; Qian & Bengtson, 1989; Zhou et al., 1997; Li et al., 2001, 2011; Steiner et al., 2007; Li et al., 2009, 2013; Yang et al., 2014; 2016). In the Xiaotan and Laolin sections with the most continuous and well-studied Terreneuvian sequence, the lowermost occurrence of Watsonella crosbyi is around the onset of the ZHUCE (Li et al., 2001; 2004; Li et al., 2009; 2013; Yang et al., 2016). Herein, we consider the SSF Zone II “(?)”, Ⅲ and the ZHUCE in the Upper Member of the Maidiping Formation at the Fandian section can correlate to that from the upper Zhongyicun Member and the Dahai Member of the Zhujiaqing Formation in eastern Yunnan (Figure 5), while the basal Maidiping Formation bearing the typical SSF Zone Ⅰ elements (He et al., 1984) probably correlates to the basal Zhongyicun Member in eastern Yunnan (Figure 5). In western Hubei, the Yanjiahe Formation at the Gunziao (or Aijiahe) section records the most continuous and complete Terreneuvian succession. Guo et al. (2014, 2021) recognized 3 SSF zones (Ⅰ to Ⅲ) in the Yanjiahe Formation. However, Steiner et al. (2020) noted that the Purella antiqua recognized by Guo et al. (2014) should be maikhanellid sclerites and considered that SSF Zone II in Guo et al. (2014) belongs to the upper part of SSF Zone Ⅰ, which means that SSF Zone II is also lacking in the Yanjiahe Formation. To resolve this controversy, the more careful systematic study of the three-dimensional SSFs retrieved by the acid process from this interval is the only reliable and practicable way (Guo et al., 2020). There is no doubt that the SSF Zone Ⅲ, including its index fossils, Watsonella crosbyi and Aldanella attleborensis, is merely restricted in the top 2.2m-thick Bed 5 (Guo et al., 2014; 2020; 2021) or 1.1m-thick Unit 4 (Steiner et al., 2020) of the Yanjiahe Formation. However, Steiner et al. (2020) point out that there was a mixture of the reworked older sediments and fossils together with the younger ones and that there existed a long-time hiatus in this short unit 4. Thus, the SSF Zone Ⅲ in western Hubei may be incomplete and lack the basal and/or top parts (Steiner et al., 2020, fig. 11; Figure 5). In contrast, Guo et al. (2020) suggest an alternative explanation that this interval consists of a thin background deposit with abundant fossils and a slight thick storm-formed deposit containing the broken sediments and fossils from the underlying background deposit. And, some fossils previously known in the older strata occurring in this bed may be due to their longer stratigraphic ranges (Guo et al., 2020). Guo et al. (2020) also indicate that it’s suitable to temporarily set the lower boundary of SSF Zone Ⅲ at the base of Bed 5 of the Yanjiahe Formation, but it’s yet to be confirmed whether Watsonella crosbyi and Aldanella attleborensis can exist in the underlying Bed 4 with scarce fossils. More detailed work is needed in the future. When it comes to the carbon isotope stratigraphy of the Yanjiahe Formation, based on the SSF biostratigraphy, it’s possible that the prominent positive excursion interval (P2 in Ishikawa et al., 2008) below the lowest/first occurrence of W. crosbyi and A. attleborensis (Guo et al., 2014, 2021; Steiner et al., 2020) from the middle and upper Yanjiahe Formation may correlate to one interval of the δ13C curve in the Zhongyicun Member belonging to SSF Zone Ⅰ or II (Li et al., 2013; Steiner et al., 2020, Figure 11) rather than the ZHUCE in the Dahai Member of the Zhujiaqing Formation in eastern Yunnan. Steiner et al. (2020) reassigned the basal part of the Shuijingtuo Formation in Ishikawa et al. (2008, p. 197, 77.91–78.95m) as unit 4 of the Yanjiahe Formation. Both their results of the δ13C of conglomeratic carbonate in the lower part of unit 4 show a clear positive excursion (with few repeated oscillations in Ishikawa et al., 2008, Figure 3; Figure 5), which may be caused by the redeposition of the sediments with a high positive δ13C value from the underlying unit 3 (Steiner et al., 2020). While the δ13C of the limestone matrix in the upper unit 4 are relatively steady in negative (–4 to –2‰ in Ishikawa et al., 2008, Figure 3; about –1‰ in Steiner et al., 2020, Figure 10), which more likely represents the original sedimental signal and correlates to the lower part of ZHUCE (Steiner et al., 2020). However, the topmost ZHUCE in the Xiaotan section also shows such a short but relatively steady and negative curve (Li et al., 2013). Anyway, herein, we think it’s uncertain to correlate this extremely short and problematic δ13C in the top Yanjiahe Formation at the Gunziao (or Aijiahe) section to other sections on the Yangtze Platform.
In the northern Sichuan and southern Shaanxi, the SSFs from the Kuanchuanpu Formation have also been widely studied, with 3 SSF zones recognized (Xing et al., 1984; Qian et al., 1999; Steiner et al., 2004). Recently, Steiner et al. (2020, Figure 11) marked all the three zones (Zones Ⅰ to Ⅲ) with an incomplete carbon isotope curve in the Kuanchuanpu Formation. It's worth mentioning that the occurrences of the key index fossils of SSF Zone II and Ⅲ in this area are quite rare compared with those of Zone Ⅰ. W. crosbyi and A. attleborensis were merely reported in a 1m-thick bed in the upper Kuanchuanpu Formation at the Yuanjiaping section (Xing et al., 1984, p. 118, pls. 26, 27). In the nearby Xuanjiangping section, they may have relatively long stratigraphic ranges (ca. 7 m in Qian et al., 1999, Figure 3-11), but none of them had been discovered in the detailed description of this section by Qian (1989, p. 18). Purella squamulosa only occurred in the top 16-cm-thick bed of the Kuanchuanpu Formation (5.75-m-thick) at the Shatan section (Steiner et al., 2004). Thus, the distribution and time range of the SSF Zone II and Ⅲ in different sections of northern Sichuan and southern Shaanxi may also be quite variable or even missing. In addition, no such positive δ13C excursions like the ZHUCE in eastern Yunnan and the Fandian section herein have been reported in the associated strata in this area. Herein, based on the occurrences of W. crosbyi, A. attleborensis, and P. squamulosa reported from the Kuanchuanpu Formation (Xing et al., 1984; Qian et al., 1999, Figure 3-11; Steiner et al., 2004; and references therein), we tentatively correlate the SSF Zones II (?) and Ⅲ from the Maidiping Formation of the Fandian section to that from the upper/top Kuanchuanpu Formation.
In general, the key index fossils of SSF Zone Ⅰ to Ⅲ have wide distributions along the western margin of the Yangtze Platform and almost have a synchronous FAD in different areas, which makes them more reliable and useful for the regional stratigraphic subdivision and correlation compared to the variable δ13C curve. At present, W. crosbyi has been discovered in most of the classical Terreneuvian sections on the Yangtze Platform, while the nearly coeval A. attleborensis has relatively restricted distribution and yet to be extensively searched in southern Sichuan. Thus, the FAD of W. crosbyi is more suitable and useful for the definition of the base of Cambrian Stage 2 (Xiaotanian) in South China. And the onset of the ZHUCE carbon isotope excursion almost corresponds to the FAD of W. crosbyi in the Xiaotan and Laolin sections in eastern Yunnan and the Fandian section in southern Sichuan, indicating that ZHUCE can be an auxiliary maker to define the lower boundary of the Xiaotanian in South China under the control of SSF biostratigraphy.
Correlation of Watsonella crosbyi Zone with other continents
Watsonella crosbyi was almost globally distributed in the upper Terreneuvian, such as, Siberia, Mongolia, South China, Australia, France, and Avalonia (see detailed list in Jacquet et al., 2017). It has been proposed to be a suitable candidate for the index fossil to define the base of the Cambrian Stage 2 (Li et al., 2011; Peng & Babcock, 2011; Zhu et al., 2019). However, W. crosbyi has been skeptical about the utility of global correlation due to its supposed long stratigraphic range (Parkhaev et al., 2012; Landing et al., 2013). The new bio, chemo, and chrono-stratigraphic progress of the Terreneuvian suggest that W. crosbyi had almost synchronous FAD in the areas mentioned above and its occurrence is in the lower to middle Cambrian Stage 2 (Brasier et al., 1996; Landing et al., 1989; Esakova & Zhegallo, 1996; Kouchinsky et al., 2007, 2017; Li et al., 2009, 2013; Maloof et al., 2010; Li et al., 2011; Devaere et al., 2013; Landing & Kouchinsky, 2016; Smith et al., 2016; see detailed discussion in Jacquet et al., 2017). However, as referred by Jacquet et al. (2017), the poorly preserved specimens on the bedding surfaces from the possible trilobite bearing zone in Avalonia (Bengtson & Fletcher, 1983; Landing et al., 2013) need to be reinvestigated. It’s noteworthy that the FAD of W. crosbyi all occurred below the prominent δ13C positive excursions in South China (ZHUCE in Zhu et al., 2006; Li et al., 2009, 2013), Siberia (I′ in Kouchinsky et al., 2007; Landing & Kouchinsky, 2016; Kouchinsky et al., 2017), Mongolia (peak “F” in Brasier et al., 1996; 5p in Smith et al., 2016) and Avalonia (Landing & Kouchinsky, 2016), which also proves that the ZHUCE δ13C positive excursion can be a useful auxiliary maker to define the base of Cambrian Stage 2 under well-constrained biostratigraphy.
Herein, based on the SSF biostratigraphic and carbon isotope stratigraphic correlations (ZHUCE), we consider that the Watsonella crosbyi Zone in the Yangtze Platform can be correlated to the Watsonella crosbyi Zone in Siberia (Kouchinsky et al., 2007; 2017), the Watsonella crosbyi Zone at the upper Bayangol Formation in the Western Mongolia (Esakova and Zhegallo, 1996; Smith et al., 2016), the Watsonella crosbyi–Oelandiella korobkovi Interval Zone in Montagne Noire of France (Devaere et al., 2013), the Watsonella crosbyi and Aldanella sp. cf. golubevi cooccurring strata in South Australia (Jacquet et al., 2017) and the Watsonella crosbyi Zone in the Avalon region (Landing et al., 1989).
Conclusion
The Terreneuvian of the Yangtze Platform in the South China Block yields four successive small shelly fossil assemblages (Zone I to IV) and a good record of carbonate isotope chemostratigraphy. Although at some sections in southern Sichuan, the SSF Zone II is missing and the Zone Ⅲ is incomplete, as well as the ZHUCE δ13C positive excursion from the Maidiping Formation (Steiner et al., 2020; Yang and Steiner, 2021), the Fandian section studied herein is relatively special. We recovered abundant small shelly fossils from the Upper Member of the Maidiping Formation at the Fandian section with 26 genera, 24 species, and 3 undetermined species. Two Terreneuvian SSF biozones on the Yangtze Platform (? Zone II and Zone Ⅲ) have been recognized from the Upper Member of the Maidiping Formation herein. Watsonella crosbyi, its lowermost occurrence (horizon 14.1 m) marking the base of Watsonella crosbyi Assemblage Zone (Ⅲ), was discovered from the Maidiping Formation at the Fandian section for the first time. Below the lowest occurrence of W. crosbyi, typical SSFs of the Paragloborilus subglobosus–Purella squamulosa Assemblage Zone (II) have been distinguished. Meanwhile, the lowermost occurrence of Watsonella crosbyi is rightly at onset of the ZHUCE carbon isotope excursion in the upper Maidiping Formation at the Fandian section, which can well correlate with that in the Xiaotan and Laolin sections in eastern Yunnan (Li et al., 2001, 2004, 2011; Li et al., 2009; 2013; Yang et al., 2014). The FAD of Watsonella crosbyi is a useful and practicable index fossil to define the base of Cambrian Stage 2 in South China (Xiaotanian) as well as in other continents. The ZHUCE δ13C positive excursion can be a useful auxiliary maker.
Data Availability Statement
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
Author Contributions
Conceptualization: BP, AY, and GL; sample collection: QF, BP, ML, and AY; funding acquisition: BP and AY; writing-original draft: QF and BP; writing-review and editing: QF, BP, AY, ML, and GL.
Funding
This work was supported by grants from the National Key Research and Development Program of China (2021YFA0718100), the National Natural Science Foundation of China (41872011; 42002024; 41890844), and the Strategic Priority Research Program of the Chinese Academy of Sciences (XDB260000000).
Acknowledgments
We thank Professor Yi Qian (NIGPAS) and Professor Tinggui He (Chengdu University of Technology) for their field assistance. We also thank Michael Steiner (Freie Universität Berlin, Germany), Pavel Parkhaev (Russian Academy of Sciences, Russia) and another reviewer for their constructive comments.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2022.922439/full#supplementary-material
References
Babcock, L. E., Peng, S. C., Geyer, G., and Shergold, J. H. (2005). Changing Perspectives on Cambrian Chronostratigraphy and Progress toward Subdivision of the Cambrian System. Geosciences J. 9, 101–106. doi:10.1007/BF02910572
Bengtson, S., and Fletcher, T. P. (1983). The Oldest Sequence of Skeletal Fossils in the Lower Cambrian of Southeastern Newfoundland. Can. J. Earth Sci. 20, 525–536. doi:10.1139/e83-050
Betts, M. J., Paterson, J. R., Jacquet, S. M., Andrew, A. S., Hall, P. A., Jago, J. B., et al. (2018). Early Cambrian Chronostratigraphy and Geochronology of South Australia. Earth-Science Rev. 185, 498–543. doi:10.1016/j.earscirev.2018.06.005
Brasier, M. D., Magaritz, M., Corfield, R., Huilin, L., Xiche, W., Lin, O., et al. (1990). The Carbon- and Oxygen-Isotope Record of the Precambrian-Cambrian Boundary Interval in China and Iran and Their Correlation. Geol. Mag. 127, 319–332.
Brasier, M. D., Shields, G., Kuleshov, V. N., and Zhegallo, E. A. (1996). Integrated Chemo-And Biostratigraphic Calibration of Early Animal Evolution: Neoproterozoic-Early Cambrian of Southwest Mongolia. Geol. Mag. 133, 445–485. doi:10.1017/s0016756800014886
Devaere, L., Clausen, S., Steiner, M., Álvaro, J. J., and Vachard, D. (2013). Chronostratigraphic and Palaeogeographic Significance of an Early Cambrian Microfauna from the Heraultia Limestone, Northern Montagne Noire, France. Palaeontol. Electron. 16 (17A), 1–91. doi:10.26879/366
Esakova, N. V., and Zhegallo, E. A. (1996). “Biostratigraphy and Fauna of the Lower Cambrian of Mongolia,” in Trudy Sovmestnoj Rossijsko-Mongol’skoj Paleontologičeskoj Ekspeditsii. Editor A. Û. Rozanov (Moscow: Science), 46, 1–216. (in Russian).
Feng, W. M., Sun, W. G., and Qian, Y. (2000). Early Cambrian Meishucunian Monoplacophora in Northeastern Yunnan with Some New Genera and Species. Acta Micropalaeontologica Sin. 17 (4), 362–377. (in Chinese with English summary).
Grabau, A. W. (1900). Palaeontology of the Cambrian Terranes of the Boston Basin. Occas. Pap. Boston Soc. Nat. Hist. 4, 601–694.
Guo, J. F., Li, G. X., Qiang, Y. Q., Song, Z. C., Zhang, Z. F., Han, J., et al. (2021). Watsonella crosbyi from the Lower Cambrian (Terreneuvian, Stage 2) Yanjiahe Formation in Three Gorges Area, South China. Palaeoworld 30 (1), 1–19. doi:10.1016/j.palwor.2020.04.006
Guo, J. F., Qiang, Y. Q., Han, J., Song, Z. C., Wang, W. Z., and Zhang, Z. F. (2020). Recent Research Progress on Small Shelly Fossils from the Cambrian (Terreneuvian) Yanjiahe Formation in the Three Gorges Area. Earth Sci. Front. 27 (6), 104–115. (in Chinese). doi:10.13745/j.esf.sf.2020.6.5
Guo, J. F., Li, Y., and Li, G. X. (2014). Small Shelly Fossils from the Early Cambrian Yanjiahe Formation, Yichang, Hubei, China. Gondwana Res. 25 (3), 999–1007. doi:10.1016/j.gr.2013.03.007
He, T. G., Ding, L. F., Zeng, Y. F., Wang, B., Li, S. L., Yin, J. C., et al. (1984). “The Sinian–Cambrian Boundary in Southwestern Part of Sichuan,” in The Sinian–Cambrian Boundary of China. Editors Y. S. Xing, Q. X. Ding, H. L. Luo, T. G. He, and Y. G. Wang (Beijing: Bulletin of the Institute of Geology, Chinese Academy of Geological Sciences), 64–90. (in Chinese with English summary).
Ishikawa, T., Ueno, Y., Komiya, T., Sawaki, Y., Han, J., Shu, D. G., et al. (2008). Carbon Isotope Chemostratigraphy of a Precambrian/Cambrian Boundary Section in the Three Gorge Area, South China: Prominent Global-Scale Isotope Excursions Just before the Cambrian Explosion. Gondwana Res. 14 (1-2), 193–208. doi:10.1016/j.gr.2007.10.008
Jacquet, S. M., Brougham, T., Skovsted, C. B., Jago, J. B., Laurie, J. R., Betts, M. J., et al. (2017). Watsonella crosbyi from the Lower Cambrian (Terreneuvian, Stage 2) Normanville Group in South Australia. Geol. Mag. 154 (5), 1088–1104. doi:10.1017/s0016756816000704
Jiang, G., Wang, X., Shi, X., Xiao, S., Zhang, S., and Dong, J. (2012). The Origin of Decoupled Carbonate and Organic Carbon Isotope Signatures in the Early Cambrian (Ca. 542-520Ma) Yangtze Platform. Earth Planet. Sci. Lett. 317-318, 96–110. doi:10.1016/j.epsl.2011.11.018
Jiang, Z. W. (1980). Monoplacophorans and Gastropods Fauna of the Meishucun Stage from the Meishucun Section, Yunnan. Acta Geol. Sin. 54, 112–123. + 168–170 (in Chinese).
Kouchinsky, A., Bengtson, S., Landing, E., Steiner, M., Vendrasco, M., and Ziegler, K. (2017). Terreneuvian Stratigraphy and Faunas from the Anabar Uplift, Siberia. Acta Palaeontol. Pol. 62, 311–440. doi:10.4202/app.00289.2016
Kouchinsky, A., Bengtson, S., Pavlov, V., Runnegar, B., Torssander, P., Young, E., et al. (2007). Carbon Isotope Stratigraphy of the Precambrian-Cambrian Sukharikha River Section, Northwestern Siberian Platform. Geol. Mag. 144, 609–618. doi:10.1017/s0016756807003354
Landing, E., Geyer, G., Brasier, M. D., and Bowring, S. A. (2013). Cambrian Evolutionary Radiation: Context, Correlation, and Chronostratigraphy-Overcoming Deficiencies of the First Appearance Datum (FAD) Concept. Earth-Science Rev. 123, 133–172. doi:10.1016/j.earscirev.2013.03.008
Landing, E., and Kouchinsky, A. (2016). Correlation of the Cambrian Evolutionary Radiation: Geochronology, Evolutionary Stasis of Earliest Cambrian (Terreneuvian) Small Shelly Fossil (SSF) Taxa, and Chronostratigraphic Significance. Geol. Mag. 153, 750–756. doi:10.1017/s0016756815001089
Landing, E., Myrow, P., Benus, A. P., and Narbonne, G. M. (1989). The Placentian Series: Appearance of the Oldest Skeletalized Faunas in Southeastern Newfoundland. J. Paleontol. 63, 739–769. doi:10.1017/s0022336000036465
Li, D., Ling, H. F., Jiang, S. Y., Pan, J. Y., Chen, Y. Q., Cai, Y. F., et al. (2009). New Carbon Isotope Stratigraphy of the Ediacaran-Cambrian Boundary Interval from SW China: Implications for Global Correlation. Geol. Mag. 146 (4), 465–484. doi:10.1017/s0016756809006268
Li, D., Ling, H. F., Shields-Zhou, G. A., Chen, X., Cremonese, L., Och, L., et al. (2013). Carbon and Strontium Isotope Evolution of Seawater across the Ediacaran-Cambrian Transition: Evidence from the Xiaotan Section, NE Yunnan, South China. Precambrian Res. 225, 128–147. doi:10.1016/j.precamres.2012.01.002
Li, G. X., Zhang, J. M., and Zhu, M. Y. (2001). Litho- and Biostratigraphy of the Lower Cambrian Meishucunian Stage in the Xiaotan Section, Eastern Yunnan. Acta Palaeontol. Sin. 40 (Suppl. l), 40–53.
Li, G., Zhao, X., Gubanov, A., Zhu, M., and Na, L. (2011). Early Cambrian Mollusc Watsonella crosbyi: a Potential GSSP Index Fossil for the Base of the Cambrian Stage 2. Acta Geol. Sinica‐English Ed. 85 (2), 309–319. doi:10.1111/j.1755-6724.2011.00400.x
Liu, D. Y. (1979). The Earliest Brachiopod Fossils of Early Cambrian in Southwest China. Acta Palaeontol. Sin. 18 (5), 93–100. + 113–114 (in Chinese).
Luo, H. L., Jiang, Z. W., Wu, X. C., Song, X. L., and Ouyang, L. (1982). The Sinian–Cambrian Boundary in Eastern Yunnan. Kunming: Yunnan People’s Publishing House, 265.
Luo, H. L., Jiang, Z. W., Xu, Z. J., Song, X. L., and Xue, X. F. (1980). On the Sinian–Cambrian Boundary of Meishucun and Wangjiawan, Jinning County, Yunnan. Acta Geol. Sin. 54 (2), 95–111. + 167 (in Chinese).
Maloof, A. C., Porter, S. M., Moore, J. L., Dudás, F. Ö., Bowring, S. A., Higgins, J. A., et al. (2010). The Earliest Cambrian Record of Animals and Ocean Geochemical Change. Bull. Geol. Soc. Am. 122 (11-12), 1731–1774. doi:10.1130/b30346.1
Meshkova, N. P. (1969). “On the Paleontological Characterization of the Lower Cambrian Deposits of the Siberian Platform,” in Biostratigraphy and Paleontology of the Lower Cambrian of Siberia and the Russian Far East. Editor I. T. Zhuravleva (Moscow: Nauka), 158–174. (In Russian).
Moczydłowska, M., and Yin, L. M. (2012). The Lower Cambrian Phytoplankton Record in China and Global Perspective for Stage Subdivision. J. Guizhou Univ. - Nat. Sci. 29 (Suppl. 1), 176–177.
Parkhaev, P. Y., Karlova, G. A., and Rozanov, A. Y. (2012). Stratigraphic Distribution of Two Potential Species for the GSSP of Cambrian Stage 2–Aldanella Attleborensis and Watsonella crosbyi. J. Guizhou Univ. - Nat. Sci. 29 (Suppl. 1), 179–180.
Parkhaev, P. Y., Karlova, G. A., and Rozanov, A. Y. (2011). Taxonomy, Stratigraphy and Biogeography of Aldanella Attleborensis–a Possible Candidate for Defining the Base of Cambrian Stage 2. Mus. North. Ariz. Bull. 67, 298–300.
Peng, S. C., and Babcock, L. E. (2011). Continuing Progress on Chronostratigraphic Subdivision of the Cambrian System. Bull. Geosci. 86, 391–396. doi:10.3140/bull.geosci.1273
Qian, Y., and Bengtson, S. (1989). Palaeontology and Biostratigraphy of the Early Cambrian Meishucunian Stage in Yunnan Province, South China. Fossils Strata 24, 1–156.
Qian, Y., Chen, M. E., He, T. G., Zhu, M. Y., Yin, G. Z., Feng, W. M., et al. (1999). Taxonomy and Biostratigraphy of Small Shelly Fossils in China. Beijing: Science Press, 247. (in Chinese).
Qian, Y. (1989). Early Cambrian Small Shelly Fossils of China with Special Reference to the Precambrian–Cambrian Boundary. Nanjing: Nanjing University Publishing House, 341.
Qian, Y. (1977). Hyolitha and Some Problematica from the Lower Cambrian Meishucun Stage in Central and S. W. China. Acta Palaeontol. Sin. 16, 107–130. (in Chinese).
Qian, Y. (1978). Study on Early Cambrian Hyolitha Fossils in Southwest Central China and the Stratigraphic Significance. J. Nanjing Inst. Geol. paleontology, Chin. Acad. Sci. 11, 1–50.
Qian, Y., Zhu, M. Y., He, T. G., and Jiang, Z. W. (1996). New Investigation of Precambrian–Cambrian Boundary Sections in Eastern Yunnan. Acta Micropalaeontologica Sin. 13 (3), 225–240. (in Chinese).
Rozanov, A. Y., Khomentovsky, V. V., Shabanov, Y. Y., Karlova, G. A., Varlamov, A. I., Luchinina, V. A., et al. (2008). To the Problem of Stage Subdivision of the Lower Cambrian. Stratigr. Geol. Correl. 16 (1), 1–19. doi:10.1134/S0869593808010012
Shaler, N. S., and Foerste, A. F. (1888). Preliminary Description of North Attleborough Fossils. Bull. Mus. Comp. Zoology 16, 27–41.
Smith, E. F., Macdonald, F. A., Petach, T. A., Bold, U., and Schrag, D. P. (2016). Integrated Stratigraphic, Geochemical, and Paleontological Late Ediacaran to Early Cambrian Records from Southwestern Mongolia. Geol. Soc. Am. Bull. 128 (3-4), 442–468. doi:10.1130/b31248.1
Steiner, M., Li, G., Qian, Y., Zhu, M., and Erdtmann, B.-D. (2007). Neoproterozoic to Early Cambrian Small Shelly Fossil Assemblages and a Revised Biostratigraphic Correlation of the Yangtze Platform (China). Palaeogeogr. Palaeoclimatol. Palaeoecol. 254, 67–99. doi:10.1016/j.palaeo.2007.03.046
Steiner, M., Li, G., Qian, Y., and Zhu, M. (2004). Lower Cambrian Small Shelly Fossils of Northern Sichuan and Southern Shaanxi (China), and Their Biostratigraphic Importance. Geobios 37 (2), 259–275. doi:10.1016/j.geobios.2003.08.001
Steiner, M., Yang, B., Hohl, S., Zhang, L., and Chang, S. (2020). Cambrian Small Skeletal Fossil and Carbon Isotope Records of the Southern Huangling Anticline, Hubei (China) and Implications for Chemostratigraphy of the Yangtze Platform. Palaeogeogr. Palaeoclimatol. Palaeoecol. 554, 659–676. doi:10.1016/j.palaeo.2020.109817
Volkova, N. A. (1968). “Acritarchs from Precambrian and Lower Cambrian Deposits in Estonia,” in Problematica of Riphean and Cambrian Layers of the Russian Platform, Urals and Kazakhstan. Editor B. M. Keller (Moscow: Nauka), 8–36.
Vostokova, V. A. (1962). Cambrian Gastropods from Siberian Platform and Taimyr. Statei po Paleontol. i Biostratigr. 28, 51–74.
Xing, Y. S., Ding, Q. X., Luo, H. L., He, T. G., and Wang, Y. G. (1984). The Sinian–Cambrian Boundary of China. Beijing: Bulletin of the Institute of Geology, Chinese Academy of Geological Sciences, 262. pp. (in Chinese with English summary).
Xiong, Y. W., Pan, B., Lu, M., Feng, Q., Yang, A. H., and Zhu, M. Y. (Forthcoming 2022). A Restudy of Carbon Isotope Chemostratigraphy of the Ediacaran - Cambrian Transition in Southern Sichuan Province. J. Stratigr. doi:10.19839/j.cnki.dcxzz.2022.0015
Yang, B., Steiner, M., Li, G., and Keupp, H. (2014). Terreneuvian Small Shelly Faunas of East Yunnan (South China) and Their Biostratigraphic Implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 398, 28–58. doi:10.1016/j.palaeo.2013.07.003
Yang, B., and Steiner, M. (2021). Terreneuvian Bio- and Chemostratigraphy of the South Sichuan Region (South China). J. Geol. Soc. 178, jgs2020–167. doi:10.1144/jgs2020-167
Yang, B., Steiner, M., Zhu, M., Li, G., Liu, J., and Liu, P. (2016). Transitional Ediacaran-Cambrian Small Skeletal Fossil Assemblages from South China and Kazakhstan: Implications for Chronostratigraphy and Metazoan Evolution. Precambrian Res. 285, 202–215. doi:10.1016/j.precamres.2016.09.016
Yin, J. C., Ding, L. F., He, T. G., Li, S. L., and Shen, L. J. (1980). The Palaeontology and Sedimentary Environment of the Sinian System in Emei–Ganluo Area, Sichuan. Chengdu: Sichuan People's Publishing House, 231. (in Chinese).
Yu, W. (1974). “Cambrian Hyolitha & Cambrian Gastropoda,” in Handbook of Stratigraphic Paleontology in Southwest China (Beijing: Science Press), 111–113. (in Chinese).
Yu, W. (1979). The Earliest Early Cambrian Aplacophora and Gastropods in Western Hubei and Their Biostratigraphic Significance. Acta Palaeontol. Sin. 18 (3), 234–270. (in Chinese).
Zhou, C. M., Zhang, J. M., Li, G. X., and Yu, Z. Y. (1997). Carbon and Oxygen Isotopic Record of the Early Cambrian from the Xiaotan Section, Yunnan, South China. Sci. Geol. Sin. 32, 201–211. (in Chinese with English abstract).
Zhu, M., Yang, A., Yuan, J., Li, G., Zhang, J., Zhao, F., et al. (2019). Cambrian Integrative Stratigraphy and Timescale of China. Sci. China Earth Sci. 62 (1), 25–60. doi:10.1007/s11430-017-9291-0
Zhu, M. Y., Babcock, L. E., and Peng, S. C. (2006). Advances in Cambrian Stratigraphy and Paleontology: Integrating Correlation Techniques, Paleobiology, Taphonomy and Paleoenvironmental Reconstruction. Palaeoworld 15 (3-4), 217–222. doi:10.1016/j.palwor.2006.10.016
Zhu, M. Y., Yang, A. H., Li, G. X., and Yuan, J. L. (2008). “A Working Model for Subdivision of the Lower Half Cambrian,” in 13th International Field Conference of the Cambrian Stage Subdivision Working Group. The Siberian Platform, Western Yakutia. Editor T. A. Voronin (Novosibirsk: SNIIGGIMS), 88–90.
Keywords: small shelly fossils, biostratigraphy, Watsonella crosbyi, Maidiping Formation, Fandian section, Cambrian stage 2, South China
Citation: Feng Q, Pan B, Yang A, Lu M and Li G (2022) Biostratigraphy of the Small Shelly Fossils From the Upper Maidiping Formation (Terreneuvian) at the Fandian Section, Sichuan Province, South China. Front. Earth Sci. 10:922439. doi: 10.3389/feart.2022.922439
Received: 18 April 2022; Accepted: 15 June 2022;
Published: 21 July 2022.
Edited by:
Ben Yang, Chinese Academy of Geologi-cal Sciences (CAGS), ChinaReviewed by:
Pavel Parkhaev, Russian Academy of Sciences, RussiaMichael Steiner, Freie Universität Berlin, Germany
Copyright © 2022 Feng, Pan, Yang, Lu and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bing Pan, YnBhbkBuaWdwYXMuYWMuY24=