 Zhikun Ma
Zhikun Ma Shu Liu
Shu Liu Zhao Li
Zhao Li Maolin Ye3
Maolin Ye3 Xiujia Huan
Xiujia Huan- 1Key Laboratory of Cultural Heritage Research and Conservation Technology (Ministry of Education), School of Cultural Heritage, Northwest University, Xi’an, China
- 2The Department of History, Jinan University, Guangzhou, China
- 3Institute of Archaeology, Chinese Academy of Social Sciences, Beijing, China
- 4Key Laboratory of Cenozoic Geology and Environment, Institute of Geology and Geophysics, Chinese Academy of Sciences, Beijing, China
The diet of prehistoric humans in the Qijia period (4,400–3,500 BP) was significantly changed by the advent of dry agriculture and food globalization. However, it is yet to be proven whether wild plants were exploited despite the cultivation of millet, and whether wheat crops, cattle, and sheep originating from southwestern Asia were adopted into the regional human diet. This study presents stable isotope, starch grain, and phytolith analyses of 24 human teeth from the Lajia site in Qinghai, which is a representative Qijia culture settlement site. The carbon and nitrogen isotope results show that the subjects primarily ate C4 plants and had a high protein diet. Starch grain and phytolith results showed that the inhabitants consumed broomcorn millet (Panicum miliaceum), foxtail millet (Setaria italica), Triticeae, tubers and roots, along with other Pooideae and Poaceae plants. This data proves that although broomcorn and foxtail millet formed the mainstay of the Qijia diet, it also included a wide range of plants, such as the tribe Triticeae, tubers and roots, which would have been foraged rather than grown. Compared with the other three contemporaneous sites in Northern China, the proportion of millet starches was highest at the Lajia site, while the type and proportion of foraged plants were the lowest. This was probably because of the arid environment in the region, which could not have supported sufficient plant resources for foraging, which in turn might have led to enhanced millet cultivation and/or a greater reliance on hunting. No wheat or barley traces were found in human teeth in Lajia, and the high proportion of nitrogen was possibly related to the consumption of sheep because sheep bones were found in a zooarchaeological study. This study enhances our understanding of the subsistence strategies present in Qijia culture and of prehistoric food globalization, which is of pivotal significance for a deeper understanding of interactions between east and west Asia during the Neolithic and Bronze ages.
1 Introduction
Marking a major evolutionary episode in human history, the transition from hunter/gatherer to agricultural economies spanned several millennia and occurred independently in 10 or more different world regions, including western and eastern Asia (Harris, 1972; Rindos, 1986; Lev-Yadun et al., 2000; Yang et al., 2012a). In these independent regions, a number of indigenous wild progenitor species from wheat and millet plants were domesticated in the Fertile Crescent of western Asia and northern China in east Asia, respectively, commencing approximately 10,000 BP (Lev-Yadun et al., 2000; Riehl et al., 2013; Lu et al., 2009a; Yang et al., 2012a; Wang et al., 2016). Thereafter, a new era in the use and creation of food resources by humans saw the rise of intensive activities around plant cultivation and animal husbandry. This led to a gradual decline in the proportion of hunter/gatherer economies and a reduction in the diversity of the human diet. This then significantly changed the subsistence economic pattern of ancient humans (Yan, 1982; Crawford et al., 2005; Crawford, 2006; Gignoux et al., 2011; Lu, 2017). Over time, wheat (Triticum aestivum) and barley (Hordeum vulgare), which were first domesticated in western Asia, spread to Europe and western Central Asia by 8,000 BP, reaching eastern Central Asia and northwestern China between 4,500 and 4,000 BP (Zhao, 2015; Liu et al., 2017; Liu et al., 2019). Broomcorn (Panicum miliaceum) and foxtail millet (Setaria italica), domesticated in eastern Asia, spread into eastern central Asia between 4,500 and 4,000 BP and into western Asia and Europe prior to 3,500 BP (Frachetti et al., 2010; Liu et al., 2016a; Stevens et al., 2016; Dong et al., 2017; Lu, 2017). In other words, transcontinental cultural exchange at this time was correlated; wheat and millet crops as well as sheep, cattle, bronze and gold vessels, lapis lazuli, bead decorations, and jade ware were used mainly in eastern central Asia and Northwest China between 4,500 and 3,500 BP with the development of different technologies and the emergence of modes of transport, including domesticated horses and vehicles (Kuz’mina, 2008; Anthony, 2010; Spengler and Willcox, 2013; Spengler et al., 2014). The exchange of food globalization in prehistory between 4,500 and 3,500 BP (Figure 1A) brought important changes to the structure and diversity of the human diet in eastern Central Asia and Northwest China (Frachetti et al., 2010; Jones et al., 2011; Spengler and Willcox, 2013; Spengler et al., 2014; Jones et al., 2016; Dong et al., 2017). In this case, hunter/gatherers were gradually marginalized in the system of the human food acquisition between 4,500 and 3,500 BP, due to the emergence and development of agriculture and transcontinental cultural and technological exchanges in Eurasia. However, hunter/gatherers have always been associated with different human economic systems; for example, the economy of hunter/gatherers is still an important part of the local economic system in some ethnic groups, which survives to this day (Du and Chen, 2016).
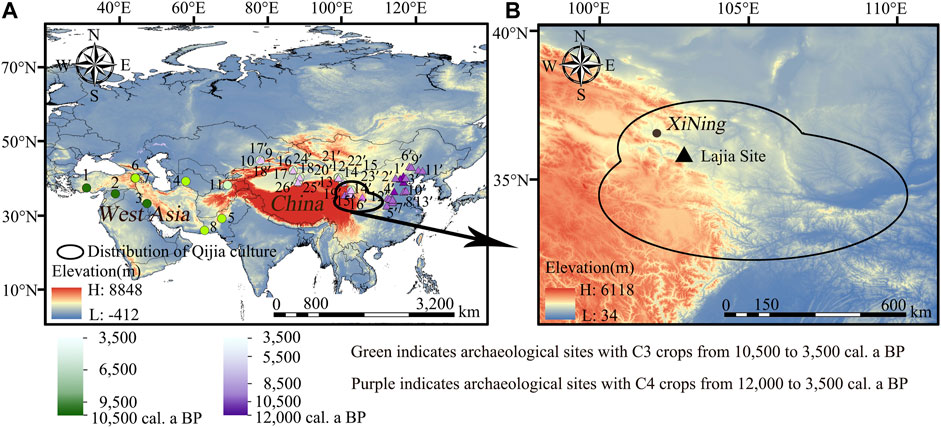
FIGURE 1. (A) Location of archaeological sites with C3 and C4 crops from 12,000 to 3,500 cal a BP (Modified from Dong et al., 2017), (B) Location of the Lajia site.
Qijia culture (4,400–3,500 BP) is in the ecotone of agriculture and animal husbandry between the Qinghai Tibet Plateau and Loess Plateau in northwest China (Figure 1B) and plays an important role in cultural communication between the eastern and western parts of Eurasia (Han, 2019). Previous studies have suggested that millet cultivation became well established during the Qijia cultural period (Zhang, 2013; Ma et al., 2014; Wang et al., 2015; Zhang and Ye, 2016; Wang and Zhen, 2021), however, only a few carbonized wheat traces were recorded for this time period. For example, carbonized wheat remains represent less than three percent of the whole plants, while the proportion of wheat remains in the Lajia and Changning sites is less than 0.01% (Zhang, 2013; Wang and Zhen, 2021). Although the carbonized plant remains from the cultural layers of the archaeological sites can reflect the agricultural structure of the Qijia culture, these results cannot supply direct evidence of the human diet. However, isotopic analysis of human skeletal remains provides a rough outline of the likely food sources and serves as a reflection of the human diet at that time. Values of δ13C from human bones dated to between 4,500 and 4,000 BP from northwestern China were dominated by C4 signals (Figure 1A), suggesting that the extent of mixing between western and eastern crops was very limited, but the C3 signals (Figure 1A) increased between 4,000 and 3,500 BP (Liu et al., 2014; Zhang and Ye., 2016; Dong et al., 2017). However, these results do not allow for the identification of specific plant species present in human diet during the Qijia period.
Recently, human teeth and dental calculus are being increasingly used as target sample types for the analysis of human diet (Beaumont et al., 2012; Henry et al., 2012; Chen et al., 2021). Stable carbon and nitrogen isotopic (δ13C and δ15N) analyses of bone collagen from human teeth remains is a conventional method of exploring the subsistence strategy of ancient humans (Beaumont et al., 2012). Dental calculus, a biomineral that entraps food particles, including some plant micro-remains as it forms, provides a direct record of plant consumption (Henry et al., 2012; Chen et al., 2021). Earlier research indicates that plant micro-remains, including starch grains and phytoliths, are well preserved in the calcification of dental calculus (Lieverse, 1999), and provide direct evidence of the human diet (Henry et al., 2012; Chen et al., 2021).
In the present study, human teeth and dental calculus unearthed from the Lajia site were selected and investigated using isotopic analysis, starch grain, and phytolith analyses to build a more comprehensive view of the human diet and subsistence strategy, in addition to transcontinental cultural and technological exchanges during the Qijia period. As a part of starch grain analysis, we also explored the extent to which different processing methods may have led to morphological changes in Poaceae food plants, to understand if and how the foods consumed by people in the Qijia culture were produced.
2 Materials and Methods
2.1 Materials
The Lajia site (Figure 1B; 102°49′40″E, 35°51′15″N) covers an area of 200,000 m2 and is located on a terrace on the north bank of the Yellow River in the Guanting Basin near Xialajia Village, Minhe County, Qinghai Province. The Lajia site is a large Qijia culture settlement situated in the transition zone between the eastern foot of the Qinghai Tibet Plateau and western Loess Plateau. Three excavation seasons in 2000, 2002–2003, and 2014–2017 uncovered house foundations and tombs in addition to ceramics, stone tools, jade objects, bone artifacts, and human bones (Ye, 2002; Du et al., 2019). Radiocarbon dating of human bones collected from house foundations in F3 and F4 (Nos. 2000QMLF3Ⅰ and 2000QMLF4IV) revealed that the site was occupied around 3,565 ± 25 and 3,580 ± 25 cal a BP (Zhang et al., 2014). When corrected by IntCal 20 (Ramsey, 2009; Reimer et al., 2020), the dates were adjusted to 3,860 ± 48 cal a BP and 3,882 ± 44 cal a BP.
Location of archaeological sites with C3 crops from 10,000 to 3,500 cal a BP: 1) Nevali Cori site (10,557–10,259 BP; Lösch et al., 2006), 2) Abu Hureyra site (10,117–9,021 BP; Moors, 2016), 3) Chogha Golan site (9,800 BP; Riehl et al., 2013), 4) Jietun site (8,328–7,712 BP; Harris et al., 1993), 5) Mehrgarh site (8,000–7,000 BP; Costantini, 2008), 6) Aratashen site (6,948–6,660 BP; Hovsepyan and Willcox, 2008), 7) Aknashen site (7,035–6,350 BP; Hovsepyan and Willcox, 2008), 8) Miri Qalat site (6,000–4,000 BP; Tengberg, 1999), 9) Tasbas site Phase 1 (4,655–4,423 BP; Spengler et al., 2014), 10) Begash site Phase 1a (4,407–4,148 BP; Spengler et al., 2014), 11) Sarazm site (5,000–4,000 BP; Spengler and Willcox, 2013), 12) Huoshiliang site (3,979–3,819 BP; Sheng et al., 2020), 13) Ganggangwa site (3,974–3,710 BP; Dodson et al., 2013), 14) Xichengyi site (3,972–3,848 BP; Zhang X. L. et al., 2015), 15) Jinchankou site (3,976–3,846 BP; Chen et al., 2015), 16) Xintala site (3,921–3,580 BP; Zhao et al., 2013), 17) Gumugou cemetery (3,834–3,695 BP; Zhang G. L. et al., 2015), 18) Xiaohe cemetery (3,707–3,478 BP; Flad et al., 2010).
Location of archaeological sites with C4 crops from 12,000 to 3,500 cal a BP: 1’) Nanzhuangtou site (12,408–11,000 BP; Yang et al., 2012a), 2’) Zhuannian site (11,000–9,700 BP; Yang et al., 2014), 3’) Donghulin site (11,000–9,500 BP; Yang et al., 2012a), 4’) Cishan site (10,300–7,000 BP; Lu et al., 2009a), 5’) Baligang site (8,700–5,000 BP; Deng et al., 2015), 6’) Baiyinchanghan site (8,500–7,400 BP; Tao et al., 2011), 7’) Egou site (8,500–7,000 BP; Liu et al., 2015), 8’) Shawoli site (8,500–7,000 BP; Zhang et al., 2011), 9’) Xinglonggou Loc.1 (8,000–7,500 BP; Zhao, 2014), 10’) Yuezhuang site (8,000–7,800 BP; Wu et al., 2011), 11’) Chahai site (8,000–7,500 BP; Wu, 2015), 12’) Bangou site (8,000–7,000 BP; Liu et al., 2013), 13’) Xihe site (7,660–7,000 BP; Wu, 2014), 14’) Heibiya site (4,760–4,330 BP; Jia, 2012), 15’) Hurere site (5,290–4,620 BP; Jia, 2012), 16’) Hongtujiaozi site (4,960–4520 BP; Jia, 2012), 17’) Tasbas site Phase 1 (4,655–4,423 BP; Spengler et al., 2014), 18’) Begash site Phase 1a (4,407–4,148 BP; Spengler et al., 2014), 19’) Gongshijia site (4,210–3,670 BP; Jia, 2012), 20’) Huoshiliang site (3,979–3,819 BP; Sheng et al., 2020), 21’) Ganggangwa site (3,974–3,710 BP; Dodson et al., 2013), 22’) Xichengyi site (3,972–3,848 BP; Zhang X. L. et al., 2015), 23’) Jinchankou site (3,976–3,846 BP; Chen et al., 2015), 24’) Xintala site (3,921–3,580 BP; Zhao et al., 2013), 25’) Gumugou cemetery (3,834–3,695 BP; Zhang G. L. et al., 2015), 26’) Xiaohe cemetery (3,707–3,478 BP; Flad et al., 2010).
Twenty-four teeth belonging to seven individuals (numbered F4Ⅱ, F4Ⅲ, F4Ⅴ, F4Ⅹ, F7Ⅰ, M8, and M11) from two house foundations and two tombs at the Lajia site were selected as samples (Table 1 and Figure 2), seven human teeth from each individual were chosen for carbon stable isotope measurements, twenty-four dental calculi were analyzed using starch grain and phytolith analyses. The human teeth from the Lajia site used in this study were dated to between 3,900 and 3,800 BP (Zhang et al., 2014).
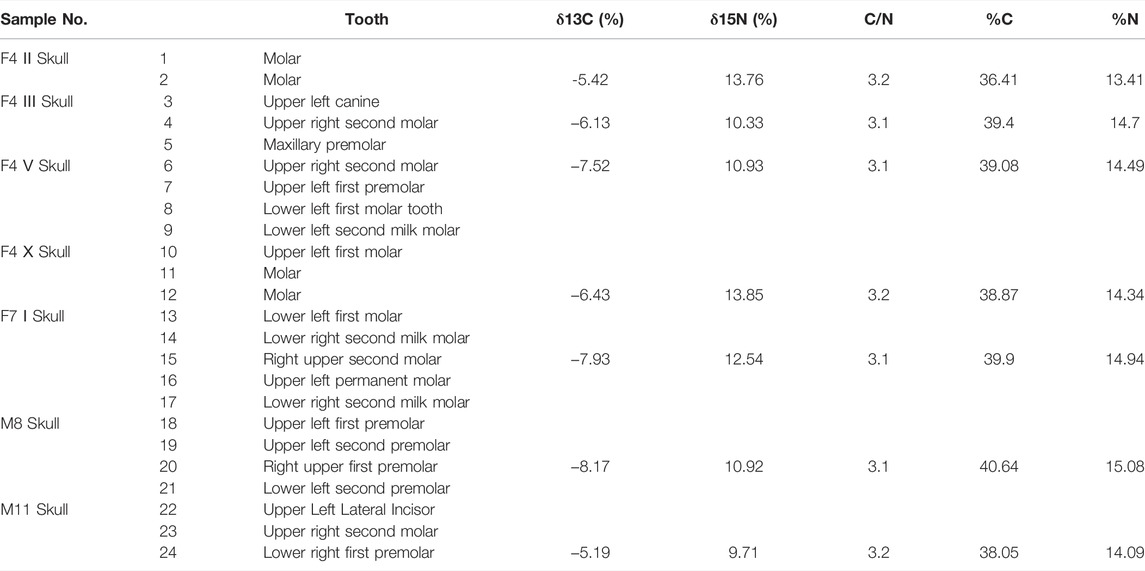
TABLE 1. Information on human tooth samples at the Lajia site and results of carbon and nitrogen isotopic testing.

FIGURE 2. Twenty-four human teeth from seven individuals from the house foundations and tombs of the Lajia site in Qinghai Province. (1,2) belong to the F4 II human skull; (3-5) belong to the F4 III human skull; (6-9) belong to the F4 V human skull; (10–12) belong to the F4 X human skull; (13–17) belong to the F7 I human skull; (18–21), belong to the M8 human skull; (22–24) belong to the M11 human skull. (1–24) lateral and frontal views of each tooth. Black arrows show the sampling locations. (Scale bar: 5 mm).
2.2 Methods
2.2.1 Analysis of Carbon and Nitrogen Stable Isotope
Seven of the 24 human teeth were selected for carbon and nitrogen stable isotope measurements. The δ13C and δ15N values were measured at Beta Analytic laboratories in Miami, United States, and the analytical precisions were ±0.3% and ±0.5%, respectively. The test results are listed in Table 1. The data were analyzed and processed using Origin 9.1 software.
2.2.2 Extraction Experiments of Starch Grains and Phytoliths From Dental Calculus
A dental pick was used to detach the visible calculus from all the teeth. Calculus fragments from each individual sample were collected in a piece of aluminum foil and immediately transferred into 1.5-ml centrifuge tubes. Dental calculus from each tooth was then treated using an ultrasonic cleaner according to the following steps (Lu et al., 2009a; Lu et al., 2009b; Yang et al., 2012a; Yang et al., 2012b; Chen et al., 2021):
Dental calculus was first cleaned gently with ultrapure water to remove adhesive particles. After washing, the samples were placed in a centrifuge tube and 20 ml of ultrapure water was added before the tube was placed in an ultrasonic cleaner for 10 min. A solution of 6% H2O2 was added to release starch grains that were potentially trapped within the dental calculus and let it stand for 1 h. Ultrapure water was then added, and the sample was centrifuged and cleaned at 2,500 rpm for 5 min. Excess liquid was poured off and only the residue was retained. This step was repeated three times. Then, 10% HCl was added to remove calcium impurities and the mixture was allowed to stand for 20 min. The solution was washed until a neutral pH was reached. A CsCl heavy liquid with a specific gravity of 1.8 g/cm3 was added and the solution was centrifuged. The supernatant containing the starches was decanted into a fresh tube. Ultrapure water was then used to clean the floating substances four times for the preparation of starch granule slides. The lower residual liquid after a flotation selection of starch to extract phytoliths was then added to the 2.35 g/cm3 ZnBr2 heavy liquid for another floating selection. Starch granule slides were prepared by mounting the recovered residue onto a slide in a solution of 25% glycerin and 75% ultrapure water. The slides were then sealed with neutral gum. Phytolith slides were prepared by mounting the recovered residue in a neutral resin and fixing them using a cover glass.
2.2.3 Modern Simulation Experiments
Prehistoric foods required preparation prior to human consumption, and the main methods included physical methods, such as shelling and grinding (Wright, 1994), as well as heat treatment, such as boiling (Barton and Torrence, 2015). Therefore, the starch grains and phytoliths were manually prepared and/or boiled prior to the formation of dental calculus. Earlier research has shown that morphological changes in ancient starches taken from dental calculus were evident, but had little effect on phytoliths (Wang and Lu, 1993; Wu and Wang, 2012). To accurately identify ancient starches, we performed a simulation experiment of grinding and boiling starch grains from millet and wheat crops from dry farming. The experimental procedure was as follows:
The seeds of modern foxtail and broomcorn millets, and common wheat were selected as samples. The seeds were then cleaned and dried to remove any impurities. To observe the morphological change after grinding the starches, each sample (0.5 g) was placed in a mortar and ground for 2–5 min to obtain three samples for the grinding simulation experiment. The ground samples were then added to a centrifuge tube containing 5 ml of ultrapure water and placed in a low-temperature environment for 1 h. To observe the morphological changes after boiling the starches, each sample (0.5 g) was added to a centrifuge tube with 5 ml of ultrapure water and boiled for 30 min in a water bath. A timer was started after boiling the water. Samples were taken at 5-min intervals, and 18 samples (six groups) were obtained for the cooking simulation experiment. After heating, the samples were cooled for 1 h. The starch suspension generated during the grinding and cooking simulation experiments was extracted using a disposable pipette, a few drops were then added to a glass slide before 25% glycerin was added, and the slide was then sealed with neutral gum.
After being dried at ≤40°C, the slide was examined using a Leica DM 750 microscope to observe starch grains and phytoliths at ×200 magnification, once identified they were photographed at ×400 magnification. The size and morphological characteristics of the starch grains were measured and recorded. The analysis was performed at the Environmental Archaeology Laboratory at Northwest University. The morphological identification and classification of starch grains were based on the modern starch grain database and related research results, which were accumulated in this laboratory (Yang et al., 2010; Wan et al., 2012; Yang and Perry, 2013; Ma et al., 2014). A comparison with published pictures and materials has been made for the identification and description of phytoliths (Wang and Lu, 1993; Lu and Liu, 2003; Piperno, 2006; Lu et al., 2009a; Lu et al., 2009b; ICPT, 2019; Ge et al., 2020). We counted and identified the starch grains and phytoliths found on the glass slide from each ancient tooth sample, and 200 grains from each sample from the simulation experiment were analyzed to obtain data on the morphological characteristics and length of the starch grains.
All the utensils used in the above experiments were cleaned in ultrapure water, and then subjected to high-temperature steaming-boiling and ultrasonic cleaning before use. In addition, all utensils for each sample were treated in the same way before extraction of starch grains to prevent external pollution or cross-contamination between samples. All toothpicks, pipettes, centrifuge tubes, and powder-free gloves used in this study were disposable, to prevent cross-contamination.
3 Results
3.1 Results of Carbon and Nitrogen Stable Isotope Analysis
According to the results from Beta Laboratory, United States, the mole ratios of the seven human teeth from Lajia site range from 3.1 to 3.2, and the C and N content was 36.41–40.64% and 13.41–15.08% (Table 1), respectively, which is consistent with the values of modern bone collagen. The range of δ13C was −5.42–−8.17, and the average value was −6.68 ± 1.11 (n = 7); the average value of δ15N was 11.72 ± 1.54 (n = 7; Table 1 and Figure 3). Previous studies have shown that collagen with a C and N content of bone collagen falls in the range of 15.3–47.0% and 5.5–17.3%, respectively, and a C/N ratio range of 2.9%–3.6 can be considered as non-contaminated (DeNiro, 1985). Therefore, seven samples from the Lajia site were valid and could be used for stable isotope analysis (Table 1).
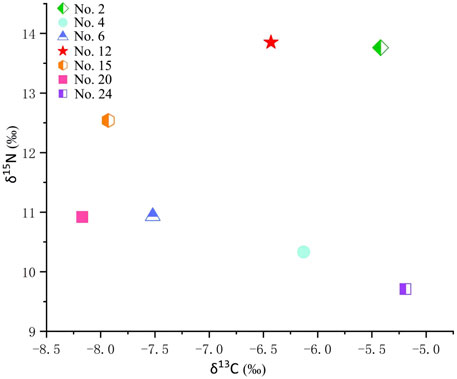
FIGURE 3. Scatter diagram for δ13C and δ15N values of the seven human tooth samples from the Lajia site.
3.2 Results of Starch Grain Analysis From Dental Calculus
A total of 611 starch grains were extracted from 24 teeth taken from seven ancient human individuals and divided into Types I–IV (Figure 4; Table 2) according to morphology and granule size.
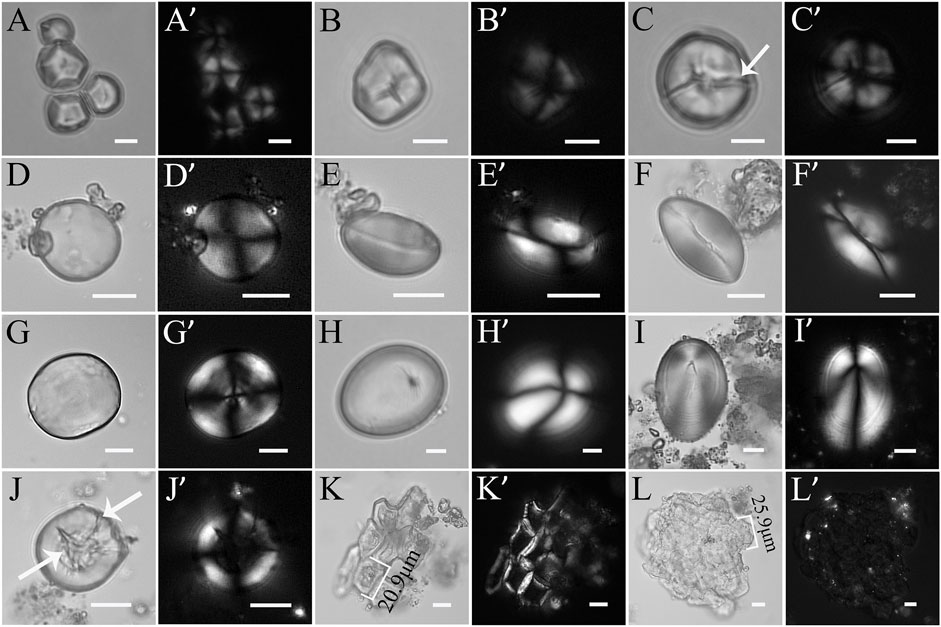
FIGURE 4. Types of starch grains extracted from 24 human dental calculus at the Lajia site. (A-C′), Type Ⅰ: (A-B′), Type Ⅰ intact starch grains; (C,C′), Type Ⅰ damaged starch grains. (D-G′), Type Ⅱ: (D-F′), Type Ⅱ-A intact starch grains; (G,G′), Type Ⅱ-B damaged starch grains. (H-I′), Type Ⅲ; (J-L′), Type Ⅳ: (J,J′), Type Ⅳ-A; (K,K′), Type Ⅳ-B; (L,L′), Type Ⅳ-C. (A, A′-L, L′), Photos were taken under bright field (left) and polarized light (right). (Arrow indicates damage in starch grains. Scale bar: 10 μm).
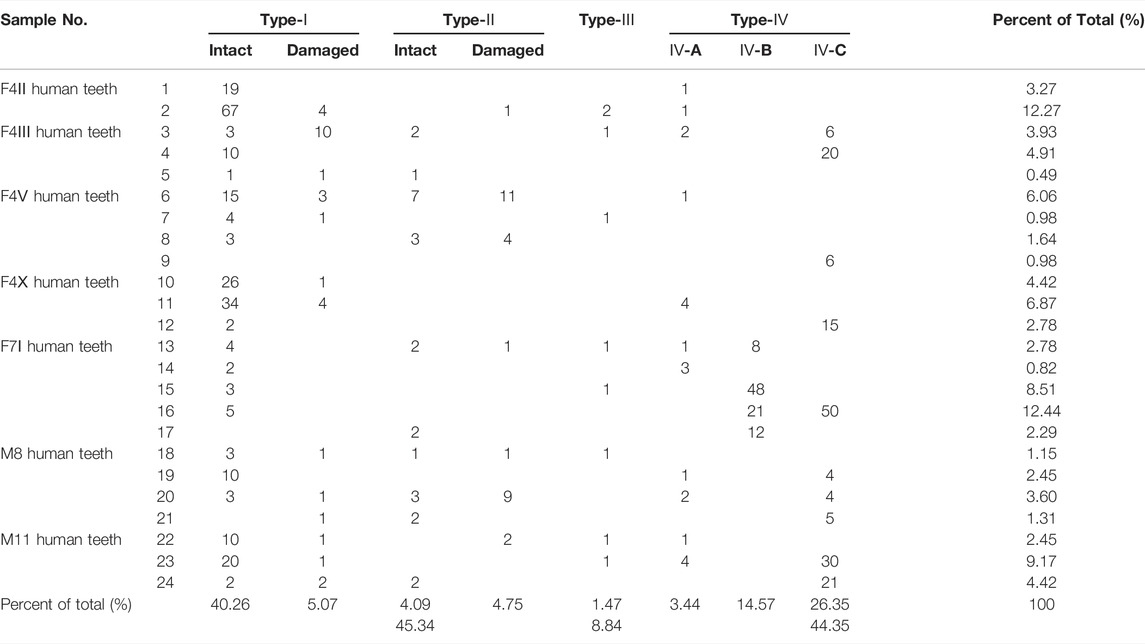
TABLE 2. Classification, quantity and percentage statistics of starch grains from human dental calculus at the Lajia site.
Type I: 277 granules were observed. They were polyhedral, free of layered texture with centric hila, and most had Y- and star-shaped fissures through the hila (Figures 4A–C′). This class of starch grains can be further divided into Type I-A intact starch grains (n = 226) with the long-axis particle size ranges from 8.0 to 27.0 μm and an average particle size of 14.9 ± 3.8 μm and Type I–B damaged starch grains (n = 31; Figures 4A–B′) with long-axis particle size ranging from 11.7 to 30.2 μm and an average particle size of 21.3 ± 5.2 μm. These are characterized by surface collapse, blurred extinction cross under polarized light, and cracks on some of the surfaces (Figures 4C, C′). Such starch grains were most likely from dry crops of foxtail and broomcorn millet.
Type II: 54 granules were observed. These granules have a biconvex shape and are mostly almost circular with a centric hilum. The extinction cross is seen to be X-shaped under polarized light, and after a slight knock to twist it around, the starch grains were oval shaped and possessed a longitudinal dent (Figures 4D,F′). This class of starch grains can be further divided into Type II-A (n = 25) intact starch grains with a long-axis particle size range of 12.9–37.7 μm and an average particle size of 26.3 ± 6.5 μm; and Type II-B (n = 29) damaged starch grains with a long-axis particle size range of 18.8–48.6 μm and an average particle size of 32.4 ± 8.2 μm. These starch molecules appeared transparent, and their layered texture is clear, with some having lost their extinction cross or on which the extinction cross is blurred under polarized light (Figures 4G, G′). These starch grains are likely derived from the Triticeae tribe.
Type III: A total of nine granules were extracted. They were oval, with an extremely unusual hilum, and no cracks were identified. After rotation, they were still oval, and the arms of the extinction crosses were slightly bent (Figures 4H–I′). Their long-axis particle size ranges from 19.2 to 55.6 μm, and their average particle size is 38.5 ± 11.1 μm. Type III starch grains originate from roots and tubers.
Type IV: A total of 271 granules are classified as unidentified, but can be further subdivided into three types: IV-A, IV-B, and IV-C. Type Ⅳ-A (n = 21) are mainly the independent starch grains that are damaged and unidentified, with the middle part sunken or hollow, the extinction blurred or lost, and the basic outline identifiable. Their long-axis particle size ranged from 13.6 to 49.6 μm, and their average particle size is 25.9 ± 9.8 μm (Figures 4J, J′). Type Ⅳ-B (n = 89) starch grains were in an aggregated state, with a residual edge outline and collapsed internal structure. Their long-axis particle size ranged from 13.1 to 31.0 μm and their average particle size is 21.3 ± 3.4 μm (Figures 4K, K′). Type Ⅳ-C (n = 161) starch grains, overlapped with each other, had completely lost their independent outline, with no extinction characteristics under polarized light. Their long-axis particle size ranged from 22.8 to 37.6 μm and their average particle size is 31.1 ± 3.4 μm (Figures 4L, L′).
3.3 Results of Phytolith Analysis of Dental Calculus
A total of 1,085 phytoliths were extracted from 24 ancient human teeth, of which 1,019 were identifiable and included 13 types. It was observed that INTERDIGITATING η-type phytoliths from the husks of broomcorn millet (n = 7, 0.65%; Figure 5B), INTERDIGITATING Ω-type phytoliths from the husks of foxtail millet (n = 2, 0.18%; Figure 5A), and INTERDIGITATING phytoliths from the husks of the family Poaceae (n = 5, 0.46%; Figures 5C, D). Besides, a total of 495 ACUTE BULBOSUS phytoliths were found, which accounted for the highest proportion (45.62%; Figure 5K); there were also BLOCKY phytoliths (25.44%; Figure 5M), ELONGATE ENTIRE (8.29%; Figure 5I), RONDEL phytoliths (5.90%; Figure 5E), ELONGATE DENTATE (3.50%; Figure 5J), BILOBATE phytoliths of Panicoideae (1.84%; Figure 5H), BULLIFORM FLABELLATE phytoliths (1.20%; Figure 5M), CRENATE phytoliths (0.37%; Figure 5F), ELONGATE DENDRITIC of Pooideae (0.37%; Figure 5G), and TRACHEARY phytoliths (0.09%; Figure 5N).
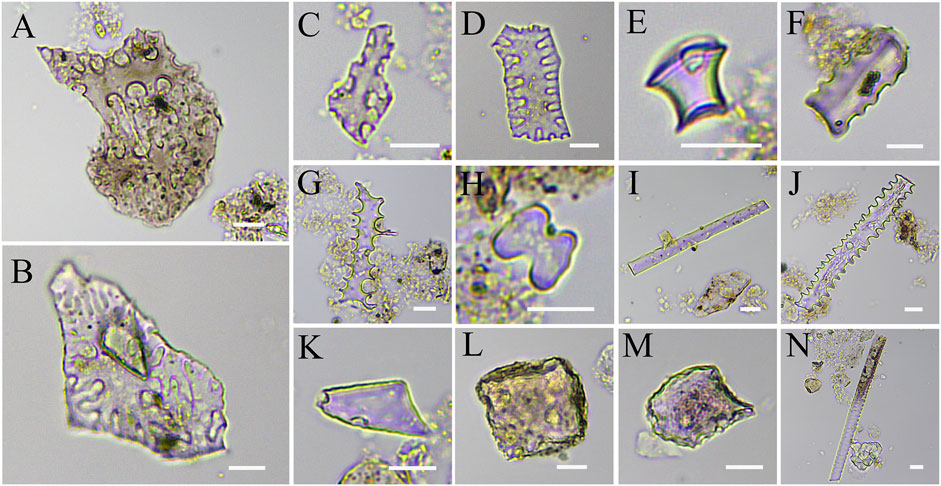
FIGURE 5. Types of phytoliths extracted from 24 human dental calculus at the Lajia site. (A)–(D), Epidermal silica layer: (A) INTERDIGITATING Ω-type (B) INTERDIGITATING η-type, (C,D) INTERDIGITATING; (E) RONDEL, (F) CRENATE, (G) ELONGATE DENDRITIC, (H) BILOBATE, (I) ELONGATE ENTIRE, (J) ELONGATE DENTATE, (K) ACUTE BULBOSUS, (L) BLOCKY, (M) BULLIFORM FLABELLATE, (N) TRACHEARY (Scale bar: 10 μm).
3.4 Results of Starch Grain Analysis in Grinding and Cooking Experiments
The results of the grinding simulation experiments for modern foxtail and broomcorn millets, and common wheat are consistent with those of our earlier study (Ma et al., 2019). The observations and statistical results suggest that: 1) grinding causes physical damage to some starch grains and results in changes in their appearance and shape, which is mainly reflected by the roughened surface and the edges becoming cracked and/or broken. 2) A positive correlation between the grinding strength and the content of damaged starch grains was observed (Figures 7V–X’). 3) Grinding causes deformation of the extinction cross of starch grains under polarized light, but the position of the hilum is generally not influenced.
The starch grain analysis results in the cooking simulation experiments of modern foxtail and common millets, and common wheat were consistent with those of earlier studies (Ge et al., 2010; Hong et al., 2013; Lu et al., 2014). It is suggested that: 1) crystal structure of starch grains becomes damaged during heating, and their central parts become sunken. Starch molecules became transparent with gradual expansion and deformation. With increased heating duration, fewer small grains were visible, and the content of large grains increased. Upon determination, the long-axis particle size of modern foxtail millet starch grains increased to 14.1, 21.3, and 31.1 μm after being heated for 5, 15, and 30 min, respectively (Figures 7D, G, J, M, P, S). This indicates that the grain size of both foxtail and broomcorn millet doubled throughout the cooking procedure (Figures 7E, H, K, N, Q, T). The starch grain size of common wheat increased to 25.44 μm and 38.91–59.36 μm after being heated for 5 and 30 min, respectively (Figures 7F, I, L, O, R, U). 2) Upon observation, the starch grains became transparent from the center of the hilum, and the extinction crosses were unclear after heating for 5 min, but remained mostly as independent scattered individual grains (Figures 7D, D’–F, F′). After 5–15 min of heating, the starch grains started to aggregate with increased transparency, and the extinction cross had completely disappeared, but the basic outline was still intact (Figures 7G, G’–L, L′). After heating for 15–30 min, the independent outline was lost, and the starch grains were almost completely transparent without extinction characteristics (Figures 7M, M’–U, U′). 3) Grinding damage and cracks were observed in a small number of starch grains (Figures 7: Y, Y’–AA, AA’).
4 Discussion
4.1 Human Diet at the Lajia Site
Using carbon and nitrogen isotope analysis of bone collagen, it is possible to trace the absorption of food by humans or animals and back trace the ratios to the food types. Earlier studies have shown that the carbon isotope ratios of bone collagen shifted by approximately 6‰ between dietary protein and consumer bone collagen (Zhang et al., 2003). Generally, the average δ13C values of C3 and C4 plants are −26% and −13% (Zhang et al., 2003), respectively. The enrichment of δ13C values between trophic levels is very low (approximately 1%–1.5%), which is often negligible. Therefore, the average δ13C values of C3 and C4 types of food are preserved in bone collagen with an approximate 6% offset, namely −20% and 7%, respectively (Merwe, 1982). The δ13C values of seven samples from the Lajia site were relatively similar (Table 1). The range of δ13C was −5.42 to −8.17, and the average value is −6.68 ± 1.11% (n = 7). According to a formula that converts the results of δ13C determination into the percentage of C4 plants in the diet (Cai and Qiu, 1984; Zhang et al., 2003; Zhang et al., 2017), the proportion of C4 plants was calculated to be 91%. This suggests that the people of the Lajia site mainly consumed C4-based foods.
Unlike the carbon stable isotope value, there is a 3–5% enrichment between consumers for the nitrogen isotope value. It is possible to use nitrogen isotopes to determine the trophic levels of the main dietary sources (Bocherens et al., 1994). Under the condition of a lack of δ15N value of each trophic level in archaeological sites, the δ15N value of herbivores is usually around 3–7%, that of omnivores is often between 7 and 9%, and that of carnivores is over 9% (Ambrose and Katzenberg, 2000). Accordingly, the δ15N values of human samples can be assessed and the relative proportions of plants and animals can be analyzed. The average δ15N above 9.7% in the Lajia site falls in the δ15N range of carnivores, showing that animal food resources were dominant. Faunal remains, including fragmented bones from pigs, sheep, and deer, were also found at the Lajia site (Ye, 2015), indicating that humans probably consumed meat from these animals, with the influence of food globalization needing to be recognized too.
In addition, compared to the stable isotopes of four human bones from the Lajia site (range of δ13C value between −7.57% and −8.4%, average δ13C value of −7.90 ± 0.35%; range of δ15N value between 9.77 and 10.25%, average δ15N value of 9.99 ± 0.17%) (Zhang and Ye, 2016), the range and average δ13C and δ15N values of this study obtained from human teeth were almost the same. As mentioned above, the δ13C and δ15N values of seven human teeth in Lajia suggest that human diets mainly included C4-based food resources supplemented with large quantities of meat.
4.2 Type and Proportion of Plant-Based Diet at the Lajia Site
The δ13C and δ15N values of human teeth at the Lajia site suggest that human diets mainly included C4-based food resources, but they do not inform us of the type and proportion of plant-based diet. Fortunately, plant residues remain embedded in dental calculus, and studies on human teeth have provided valuable data for reconstructing a comprehensive view of human diets in the past (Henry and Piperno, 2008). More than 1,600 plant remains were recovered from 24 human teeth in this study, including an assemblage of 611 starch grains (Table 2), representing grains from foxtail and broomcorn millets; the tribe Triticeae, roots and tubers; and an assemblage of 1,085 phytoliths (Figures 5, 6), the majority of which were from the husks of the foxtail and broomcorn millets; and stems and leaves of Pooideae and Panicoideae. Of these plant micro-remains, ∼54% (n = 331) of the total starches showed damage and were large in size (Table 2), which is different from the characteristics of over 200 Asian species housed in modern reference collections (Yang et al., 2010; Wan et al., 2012; Yang and Perry, 2013). The combination of the damage condition of extinction characteristics and contour decoration of the ancient starches suggests that the starches extracted from human dental calculus have undergone a process of alteration.
FIGURE 6. Changes in percentages of phytolith types extracted from the 24 human dental calculus at the Lajia site.
Based on the morphological characteristics identified as resulting from grinding and boiling grass grains (foxtail and broomcorn millets, and wheat) (Figure 7), the grains from Lajia appear to have been processed by cooking, and there is limited evidence of grinding. First, owing to the influence of factors such as the evenness of heating and water absorption, the starch grains of the same samples showed different gelatinization during boiling. Some of the starch grains only exhibited mild gelatinization, indicating that their identification features were still present (Figures 4C, C′, G, G’), which were identified as being from foxtail and broomcorn millets, and the tribe Triticeae. Those starch grains which exhibited serious gelatinization (Figures 4K, K′, I, I′), were found upon comparison of size and shape, to be closest to those of the foxtail and broomcorn millets from the cooking simulation specimens. Additionally, in the cooking simulation experiments without grinding, a handful of starch grains exhibited signs of mechanical damage (Figures 4J, J′), which may have been caused by stirring and squeezing during cooking, from smashing grains during hulling, or from chewing.
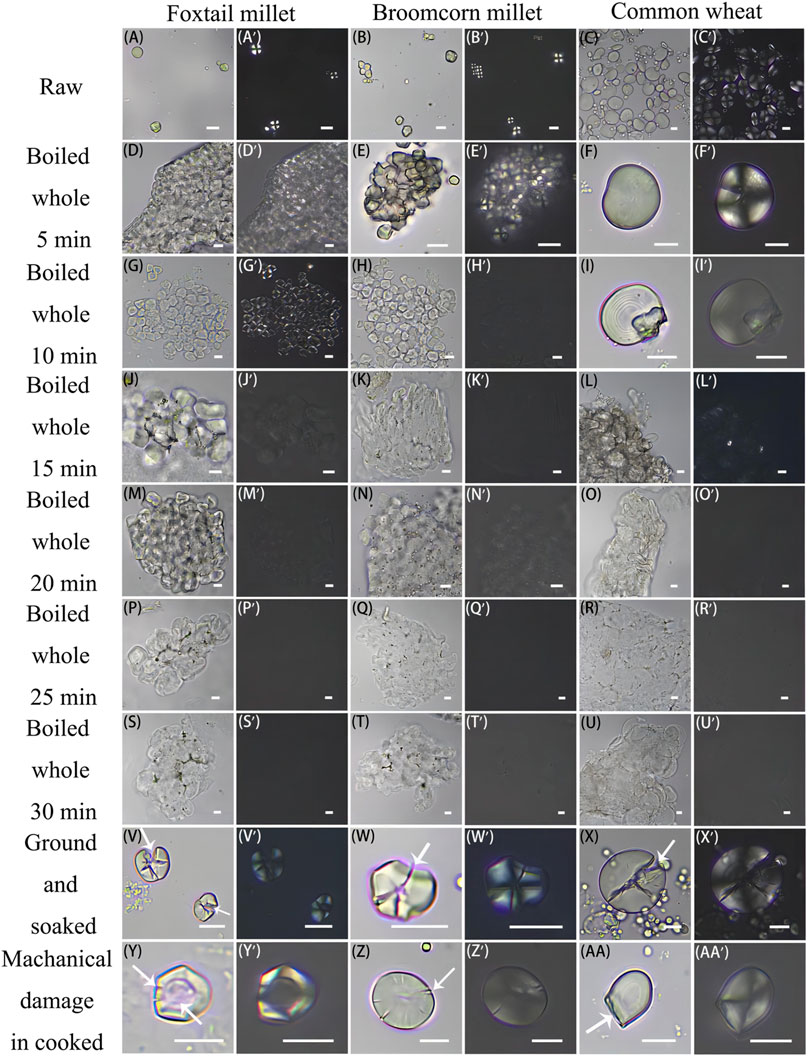
FIGURE 7. Starch grain specimens for simulation experiments of cooking and grinding. Different characteristics of starch grains from three Poaceae plants, both raw and after processing. (A, A’ - AA, AA′) Photos were taken under bright field (left) and polarized light (right); Arrow indicates the damage characteristics of starch grains; Scale bar: 10 μm.
Except for the phytoliths from the husks of foxtail and broomcorn millets and those from the husks of Gramineae, the phytoliths of the stems and leaves of Poaceae (98.71% of the total) extracted from the dental calculus are quite likely to represent the limited grain processing techniques of the Qijia period. The stems, leaves, and husks of foxtail and broomcorn millet, and other food plants require separation before grinding and cooking (Wang and Yang, 2015). An earlier simulation study indicated that using mortars, pestles, slabs, and mullers as dehusking and grinding tools does not completely hull the grains; the highest dehusking rates achieved in the experiments were 99 and 97.9%, respectively, whereas the husked grains were not 100% intact (Wang et al., 2013). Stone knives in the Lajia site were mainly used to harvest millet crops, but they also cut the stems or leaves of plants such as Panicoideae and related taxa during use (Ma et al., 2014). During the process of harvesting and dehusking, husks, stems, or leaves of millet and other plants were combined by ancient humans; therefore, these phytoliths were commonly found in the dental calculus of the Lajia site.
During the data analysis of plant micro-remains extracted from 24 human teeth at the Lajia site, the analytical results of starch grains and phytoliths were relatively consistent in foxtail and broomcorn millets. In addition, starch grains and phytoliths from the Poaceae subfamily were similar. The tribe Triticeae belongs to the Pooideae of the Poaceae subfamily. Furthermore, the phytoliths included BILOBATE phytoliths from the leaves of Panicoideae, Elongate Dendritic, Crenate, and Rondel phytoliths from Pooideae leaves, and Elongate Dentate phytoliths from the family Poaceae (Ge et al., 2020). In summary, the plant resources of ancient people in the Qijia cultural period included foxtail and broomcorn millets, plants from the tribe Triticeae, roots, tubers, Pooideae, and Panicoideae.
With respect to the absolute quantities and appearance frequencies of the identified plant micro-remains extracted from the dental calculus, the starch grains were mainly millet belonging to C4 plants (n = 277; 81.4%); phytoliths from the husks of millet crops were also recovered from the dental calculus (n = 9, 0.82%). These results are consistent with those of carbon isotopes (91% C4-based food resources) in this study and plant macro-remains in previous research, suggesting that the dry farming structure during the Qijia cultural period was also dominated by foxtail and broomcorn millets. However, no wheat or barley was recovered from the dental calculus of the Lajia site, which is consistent with the phytolith results of earlier research on the cultural sediments of the Lajia site (Wang et al., 2015). However, two carbonized wheat remains (<0.01% of the total) were recovered from the 193 flotation assemblages of Lajia (Zhang, 2013), but were not dated using AMS (radiocarbon dating). On the one hand, these wheat remains have to be treated with caution because the quantities are small. On the other hand, we do not deny the existence of wheat crops in Lajia, but this needs to be confirmed by future research. Nevertheless, evidence from plant micro and macro-remains, and carbon isotopes suggests that wheat crops played a considerably lesser role in the human diet patterns of the inhabitants of the Lajia site during the Qijia period; that is, the degree of influence of wheat crop consumption during the spread of foods around the globe is quite limited. One of the reasons why ancient humans did not consume wheat crops is probably that the cultivation and use of wheat crops was limited by environmental factors around the Lajia site. The climate of northwest China around 4,000 years ago was different to that of the wheat origin area (Yang et al., 2021), which possibly affected its growth and cultivation. Another possible reason is due to the context of culinary choices; in West and Central Asia, there was a grinding and bread-baking tradition, while in East Asia steaming and boiling of whole grains was practiced (Liu et al., 2016b). Therefore, the adoption rate of wheat crops with hard seed coats might have been slower in this region. Wheat crops were widely used with the appearance of flour grinding technology in Northern China until the Han Dynasty (Zeng, 2005).
Notably, 9% of C3-based food resources were found in the food structures during the carbon isotope analysis of human teeth in Lajia. Meanwhile, the tribe Triticeae (n = 54; 15.9%), roots and tubers (n = 9; 2.6%) belonging to C3 plants was common in the dental calculus from human teeth (Table 2). Phytoliths from Panicoideae and Pooideae, belonging to C3 plants, were also found. Based on the results of the current analysis, we can confirm that the ancient inhabitants of the Lajia site ate foxtail and broomcorn millets belonging to the C4 plants of the Panicoideae subfamily. However, we cannot completely rule out the possibility that ancient people of the Lajia site ate C3 plants belonging to Panicoideae, although it is still uncertain which C3 plants of Panicoideae they may have eaten. The results show that the human diet at the Lajia site is dominated by C4-based food resources (broomcorn and foxtail millets), but they were not the only plant resources consumed. The presence of a few C3 plants from the tribe Triticeae, roots and tubers, Panicoideae, and Pooideae in this study, along with some other C3 species of Poa and Oats in an earlier flotation study (Zhang, 2013; Ye, 2015) suggest that gathering was still an important supplementary resource for the occupants of the Lajia site.
Finally, millet starch grains represented 81.4% (n = 277) of the total identified starches taken from the teeth of the human remains at this site. Four plant species were identified in the present study. The results were compared with those from the Xinzhang (4,500–4,000 cal a BP, >50%, 6), Mogou (4,000 cal a BP, 14.3%, 7), and Nancheng sites (4,000–3,600 cal a BP, 50.6%, 4) (Li et al., 2010; Ren et al., 2017; Chen et al., 2021), and the proportion of millet starch grains in Lajia was significantly higher than that at the other sites; simultaneously, the number and proportion of plant species was lower. The composition of the plant resources may have been driven by a relatively extreme local environment. The Lajia site is located on the eastern edge of the Qinghai-Tibet Plateau climatic zone, with a low annual average temperature and fragile agricultural ecological environment. In addition, the climate tended to be dry and cold after 4,500 cal. BP in this region (Gong et al., 2011). This means that the number of plant resources that were collected in the surrounding areas and animal resources available for hunting would be further reduced from their currently sparse modern-day levels. In addition, the Lajia site was a settlement society, observed from the rich remains including house foundations, ash pits, altars, and community squares excavated from the site (Ye, 2002), which restricted the scope of activities of the ancient humans. Therefore, this region did not have sufficient plant resources to allow a community to survive through foraging alone. This might explain the enhanced millet cultivation and/or a greater reliance on hunting and sheep rearing.
5 Conclusion
The results of δ13C and δ15N values in the seven human teeth and plant micro-remains from the 24 dental calculus at the Lajia site indicate that: 1) human diets mainly consisted of C4-based food with a few C3-based foods originating from the combination of millet and plants gathered, and these ancient humans had a high-protein diet possible due to their hunting activities. Based on other archaeological evidence at the site, it is probable that pigs, sheep, and deer were the main sources of this protein. 2) Based on the analysis of ancient starch grains and phytoliths, the diet of the occupants of the Lajia site included foxtail and broomcorn millet, the tribe Triticeae, roots and tubers, and plants from Pooideae of the Poaceae subfamily. Combined with data from human teeth and dental calculus, a predominantly millet-based lifestyle occurred at the Lajia site, but gathering factors are still evident. Regarding the impact of food globalization, the dietary patterns of ancient humans at the Lajia site were probably affected by the introduction of sheep, but no direct evidence of wheat crops or their consumption was observed. Finally, the results of our simulation experiment highlight the need to consider the enlargement of size when identifying different species of cooked or processed starch grains.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
ZM and XH designed the study and prepared the manuscript. ZM, ZL, and SL collected the samples and conducted the experiments. ZM, XH, and SL analyzed the data. ZM, XH, ZL, and MY contributed to the discussion and approved the final manuscript.
Funding
This study was supported by the National Natural Science Foundation of China (Grant Nos. 41807436 and 41907372).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2022.884856/full#supplementary-material
References
Ambrose, S. H., and Katzenberg, M. A. (2000). Biogeochemical Approaches to Paleodietary Analysis. New York: Kluwer Academic/Plenum Publisher.
Anthony, D. W. (2010). The Horse, the Wheel, and Language: How Bronze Age Riders from the Eurasian Steppes Shaped the Modern World. Princeton: Princeton University Press.
Barton, H., and Torrence, R. (2015). Cooking up Recipes for Ancient Starch: Assessing Current Methodologies and Looking to the Future. J. Archaeol. Sci. 56, 194–201. doi:10.1016/j.jas.2015.02.031
Beaumont, J., Gledhill, A., Lee-Thorp, J., and Montgomery, J. (2012). Childhood Diet: a Closer Examination of the Evidence from Dental Tissues Using Stable Isotope Analysis of Incremental Human Dentine*. Archaeometry 55 (2), 277–295. doi:10.1111/j.1475-4754.2012.00682.x
Bocherens, H., Fizet, M., and Mariotti, A. (1994). Diet, Physiology and Ecology of Fossil Mammals as Inferred from Stable Carbon and Nitrogen Isotope Biogeochemistry: Implications for Pleistocene Bears. Palaeogeogrn Palaeocl 107 (3-4), 213–225. doi:10.1016/0031-0182(94)90095-7
Bronk Ramsey, C. (2009). Bayesian Analysis of Radiocarbon Dates. Radiocarbon 51 (1), 337–360. doi:10.1017/S0033822200033865
Cai, L. Z., and Qiu, S. L. (1984). Determination of Carbon-13 and Study of Ancient Recipes. Archaeol 10, 949
Chen, F. H., Dong, G. H., Zhang, D. J., Liu, X. Y., Jia, X., An, C. B., et al. (2015). Agriculture Facilitated Permanent Human Occupation of the Tibetan Plateau after 3600 B.P. Science 347 (6219), 248–250. doi:10.1126/science.1259172
Chen, T., Hou, L., Jiang, H., Wu, Y., and Henry, A. G. (2021). Starch Grains from Human Teeth Reveal the Plant Consumption of Proto-Shang People (C. 2000-1600 BC) from Nancheng Site, Hebei, China. Archaeol. Anthropol. Sci. 13, 153. doi:10.1007/s12520-021-01416-y
Costantini, L. (2008). The First Farmers in Western Pakistan: The Evidence of the Neolithic Agro-Pastoral Settlement of Mehrgarh. Pragdhara 18, 167
Crawford, G., Underhill, A., Zhao, Z., Lee, G. A., Feinman, G., Nicholas, L., et al. (2005). Late Neolithic Plant Remains from Northern China: Preliminary Results from Liangchengzhen, Shandong. Curr. Anthropol. 46 (2), 309–317. doi:10.1086/428788
Crawford, G. W. (2006). “East Asian Plant Domestication,” in In Archaeology of Asia. Editor Y. Miriam (Malden: Blackwell publishing).
Deng, Z., Qin, L., Gao, Y., Weisskopf, A. R., Zhang, C., and Fuller, D. Q. (2015). From Early Domesticated Rice of the Middle Yangtze Basin to Millet, Rice and Wheat Agriculture: Archaeobotanical Macro-Remains from Baligang, Nanyang Basin, Central China (6700-5000 BC). PloS ONE 10 (10), e0139885. doi:10.1371/journal.pone.0139885
DeNiro, M. J. (1985). Postmortem Preservation and Alteration of In Vivo Bone Collagen Isotope Ratios in Relation to Palaeodietary Reconstruction. Nature 317 (6040), 806–809. doi:10.1038/317806a0
Dodson, J. R., Li, X., Zhou, X., Zhao, K., Sun, N., and Atahan, P. (2013). Origin and Spread of Wheat in China. Quat. Sci. Rev. 72, 108–111. doi:10.1016/j.quascirev.2013.04.021
Dong, G., Yang, Y., Han, J., Wang, H., and Chen, F. (2017). Exploring the History of Cultural Exchange in Prehistoric Eurasia from the Perspectives of Crop Diffusion and Consumption. Sci. China Earth Sci. 60 (5), 1110–1123. doi:10.1007/s11430-016-9037-x
Du, X. M., and Chen, Q. D. (2016). The Gathering Economies from the Perspective of Economic Anthropology——Investigation and Analysis from the Dulong River Canyon. ethno-national Stud. 1, 75
Du, Z. W., Wang, Q. Q., Yu, M. Z., Du, W., Zhen, Q., Ma, Q., et al. (2019). A preliminary report on the excavation of Xindian culture remains from 2015 to 2016 at the Lajia site in Minhe County, Qinghai. Archaeol 6, 3
Flad, R., Li Shuicheng, S. C., Wu Xiaohong, X. H., and Zhao Zhijun, Z. J. (2010). Early wheat in China: Results from new studies at Donghuishan in the Hexi Corridor. Holocene 20 (6), 955–965. doi:10.1177/0959683609358914
Frachetti, M. D., Spengler, R. N., Fritz, G. J., and Mar'yashev, A. N. (2010). Earliest direct evidence for broomcorn millet and wheat in the central eurasian steppe region. Antiquity 84 (326), 993–1010. doi:10.1017/S0003598X0006703X
Ge, W., Liu, L., Chen, X. C., and Jin, Z. Y. (2010). An experimental study on starch grain damage during food processing and the application in archaeology. Archaelogy 7, 77
Ge, Y., Lu, H., Zhang, J., Wang, C., and Gao, X. (2020). Phytoliths in Inflorescence Bracts: Preliminary Results of an Investigation on Common Panicoideae Plants in China. Front. Plant Sci. 10, 1736. doi:10.3389/fpls.2019.01736
Gignoux, C. R., Henn, B. M., and Mountain, J. L. (2011). Rapid, global demographic expansions after the origins of agriculture. Proc. Natl. Acad. Sci. U.S.A. 108, 6044–6049. doi:10.1073/pnas.0914274108
Gong, Y., Yang, Y., Ferguson, D. K., Tao, D., Li, W., Wang, C., et al. (2011). Investigation of ancient noodles, cakes, and millet at the Subeixi Site, Xinjiang, China. J. Archaeol. Sci. 38 (2), 470–479. doi:10.1016/j.jas.2010.10.006
Han, J. Y. (2019). The Development and Evolution of Qijia Culture: Cultural Interaction and Eurasian Context. Cult. Relics 7, 60–65. (in Chinese). doi:10.13619/j.cnki.cn11-1532/k.2019.07.004
Harris, D. R., Masson, V. M., Berezkin, Y. E., Charles, M. P., Gosden, C., Hillman, G. C., et al. (1993). Investigating early agriculture in Central Asia: new research at Jeitun, Turkmenistan. Antiquity 67 (225), 324–338. doi:10.1017/S0003598X00045385
Henry, A. G., and Piperno, D. R. (2008). Using plant microfossils from dental calculus to recover human diet: a case study from Tell al-Raqā'i, Syria. J. Archaeol. Sci. 35 (7), 1943–1950. doi:10.1016/j.jas.2007.12.005
Henry, A. G., Ungar, P. S., Passey, B. H., Sponheimer, M., Rossouw, L., Bamford, M., et al. (2012). The diet of australopithecus sediba. Nature 487 (7405), 90–93. doi:10.1038/nature11185
Hong, J., Zheng, X. L., and Liu, C. (2013). Study on particle and gel properties of total,A-type,B-type starch with different thermal damage degrees. Cereals & Oils 26 (8), 8
Hovsepyan, R., and Willcox, G. (2008). The earliest finds of cultivated plants in Armenia: evidence from charred remains and crop processing residues in pisé from the Neolithic settlements of Aratashen and Aknashen. Veget Hist. Archaeobot 17, 63–71. doi:10.1007/s00334-008-0158-6
Jia, X. (2012). Cultural Evolution Process and Plant Remains during Neolithic-Bronze Age in Northeast Qinghai Province. Lanzhou (Chinese): Lanzhou University. [Ph.D. dissertation].
Jones, M., Hunt, H., Kneale, C., Lightfoot, E., Lister, D., Liu, X., et al. (2016). Food globalisation in prehistory: The agrarian foundations of an interconnected continent. Jba 4, 73–87. doi:10.5871/jba/004.073
Jones, M., Hunt, H., Lightfoot, E., Lister, D., Liu, X., and Motuzaite-Matuzeviciute, G. (2011). Food globalization in prehistory. World Archaeol. 43 (3), 665–675. doi:10.1080/00438243.2011.624764
Kuzmina, E. E. (2008). The Prehistory of the Silk Road. Philadelphia: University of Pennsylvania Press.
Lev-Yadun, S., Gopher, A., and Abbo, S. (2000). The Cradle of Agriculture. Science 288 (5471), 1602–1603. doi:10.1126/science.288.5471.1602
Li, M., Yang, X., Wang, H., Wang, Q., Jia, X., and Ge, Q. (2010). Starch grains from dental calculus reveal ancient plant foodstuffs at Chenqimogou site, Gansu Province. Sci. China Earth Sci. 53 (5), 694–699. doi:10.1007/s11430-010-0052-9
Lieverse, A. R. (1999). Diet and the aetiology of dental calculus Int. J. Osteoarchaeol. 9, 2192–2232. doi:10.1002/(SICI)1099-1212(199907/08)9:4%3C219::AID-OA475%3E3.0
Liu, L., Chen, X. C., and Zhao, H. (2013). Analysis on the function of the grinding stone unearthed from the Zhaigen and Bangou sites during the late Peiligang culture period in Mengjin, Henan. Cult. Relics. Cent. China. 5, 76
Liu, L., Field, J., Fullagar, R., Bestel, S., Chen, X., and Ma, X. (2015). What did grinding stones grind? New light on Early Neolithic subsistence economy in the Middle Yellow River Valley, China. Antiquity 84 (325), 816–833. doi:10.1017/S0003598X00100249
Liu, X., Jones, P. J., Motuzaite Matuzeviciute, G., Hunt, H. V., Lister, D. L., An, T., et al. (2019). From ecological opportunism to multi-cropping: Mapping food globalisation in prehistory. Quat. Sci. Rev. 206, 21–28. doi:10.1016/j.quascirev.2018.12.017
Liu, X., Lightfoot, E., O’Connell, T. C., Wang, H., Li, S., Zhou, L., et al. (2014). From necessity to choice: dietary revolutions in west China in the second millennium BC. World Archaeol. 46 (5), 661–680. doi:10.1080/00438243.2014.953706
Liu, X., Lister, D. L., Zhao, Z., Petrie, C. A., Zeng, X., Jones, P. J., et al. (2017). Journey to the east: Diverse routes and variable flowering times for wheat and barley en route to prehistoric China. PLoS ONE 12 (11), e0187405. doi:10.1371/journal.pone.0187405
Liu, X., Lister, D. L., Zhao, Z., Staff, R. A., Jones, P. J., Zhou, L., et al. (2016a). The virtues of small grain size: Potential pathways to a distinguishing feature of Asian wheats. Quat. Int. 426, 107–119. doi:10.1016/j.quaint.2016.02.059
Liu, X., Reid, R. E., Lightfoot, E., Matuzeviciute, G. M., and Jones, M. K. (2016b). Radical change and dietary conservatism: Mixing model estimates of human diets along the Inner Asia and China's mountain corridors. Holocene 26, 1556–1565. doi:10.1177/0959683616646842
Lösch, S., Grupe, G., and Peters, J. (2006). Stable isotopes and dietary adaptations in humans and animals at pre-pottery Neolithic Nevallı Çori, southeast Anatolia. Am. J. Phys. Anthropol. 131 (2), 181–193. doi:10.1002/ajpa.20395
Lü, H., Li, Y., Zhang, J., Yang, X., Ye, M., Li, Q., et al. (2014). Component and simulation of the 4,000-year-old noodles excavated from the archaeological site of Lajia in Qinghai, China. Chin. Sci. Bull. 59 (35), 5136–5152. doi:10.1007/s11434-014-0663-1
Lu, H., and Liu, K.-B. (2003). Morphological variations of lobate phytoliths from grasses in China and the south-eastern United States. Divers Distrib. 9 (1), 73–87. doi:10.1046/j.1472-4642.2003.00166.x
Lu, H. (2017). New methods and progress in research on the origins and evolution of prehistoric agriculture in China. Sci. China Earth Sci. 60, 2141–2159. doi:10.1007/s11430-017-9145-2
Lu, H., Zhang, J., Liu, K.-b., Wu, N., Li, Y., Zhou, K., et al. (2009a). Earliest domestication of common millet ( Panicum miliaceum ) in East Asia extended to 10,000 years ago. Proc. Natl. Acad. Sci. U.S.A. 106 (18), 7367–7372. doi:10.1073pnas.090015810610.1073/pnas.0900158106
Lu, H., Zhang, J., Wu, N., Liu, K.-b., Xu, D., and Li, Q. (2009b). Phytoliths analysis for the discrimination of foxtail millet (Setaria italica) and common millet (Panicum miliaceum). PLoS ONE 4 (2), e4448. doi:10.1371/journal.pone.0004448
Ma, Z., Li, Q., Huan, X., Yang, X., Zheng, J., and Ye, M. (2014). Plant microremains provide direct evidence for the functions of stone knives from the Lajia site, northwestern China. Chin. Sci. Bull. 59, 1151–1158. doi:10.1007/s11434-014-0174-0
Ma, Z., Perry, L., Li, Q., and Yang, X. (2019). Morphological changes in starch grains after dehusking and grinding with stone tools. Sci. Rep. 9 (1), 2355. doi:10.1038/s41598-019-38758-6
Merwe, N. J. (1982). Carbon Isotopes, Photosynthesis, and Archaeology: Different pathways of photosynthesis cause characteristic changes in carbon isotope ratios that make possible the study of prehistoric human diets. Am. Sci. 70 (6), 596–606. doi:10.2307/27851731
Moors, A. M. T. (2016). The impact of accelerator dating at the early village of Abu Hureyra on the Euphrates. Radiocarbon 34 (3), 850
International Committee for Phytolith Taxonomy (ICPT) Neumann, K., Neumann, K., Strömberg, C. A. E., Ball, T., Albert, R. M., Vrydaghs, L., et al. (2019). International Code for Phytolith Nomenclature (ICPN) 2.0. Ann. Bot. 124, 189–199. doi:10.1093/aob/mcz064
Piperno, D. R. (2006). Phytolith: A Comprehensive Guide for Archaeologist and Paleoecologists. Lanham MD: Altamira Press.
Reimer, P. J., Austin, W. E. N., Bard, E., Bayliss, A., Blackwell, P. G., Bronk Ramsey, C., et al. (2020). The IntCal20 Northern Hemisphere Radiocarbon Age Calibration Curve (0-55 cal kBP). Radiocarbon 62 (4), 725–757. doi:10.1017/RDC.2020.41
Ren, G. Y., Zhao, J., and Hou, L. L. (2017). Analysis of starch granules in human dental calculus at the Xinzhang site in Yuanping. World. Antiq. 2, 20
Riehl, S., Zeidi, M., and Conard, N. J. (2013). Emergence of agriculture in the foothills of the Zagros Mountains of Iran. Science 341 (6141), 65–67. doi:10.1126/science.1236743
Sheng, P. F., Shang, X., and Zhang, P. C. (2020). A preliminary study on crop selection of ancestors from late Longshan to early Xia Dynasty in Yulin area. Archaeol. Cult. Relic. 2, 114
Spengler, R., Frachetti, M., Doumani, P., Rouse, L., Cerasetti, B., Bullion, E., et al. (2014). Early agriculture and crop transmission among Bronze Age mobile pastoralists of Central Eurasia. Proc. R. Soc. B 281 (1783), 20133382. doi:10.1098/rspb.2013.3382
Spengler, R. N., and Willcox, G. (2013). Archaeobotanical results from Sarazm, Tajikistan, an Early Bronze Age Settlement on the edge: Agriculture and exchange. Environ. Archaeol. 18 (3), 211–221. doi:10.1179/1749631413Y.0000000008
Stevens, C. J., Murphy, C., Roberts, R., Lucas, L., Silva, F., and Fuller, D. Q. (2016). Between China and South Asia: A middle Asian corridor of crop dispersal and agricultural innovation in the Bronze Age. Holocene 26 (10), 1541–1555. doi:10.1177/0959683616650268
Tao, D., Wu, Y., Guo, Z., Hill, D. V., and Wang, C. (2011). Starch grain analysis for groundstone tools from Neolithic Baiyinchanghan site: implications for their function in Northeast China. J. Archaeol. Sci. 38 (12), 3577–3583. doi:10.1016/j.jas.2011.08.028
Tengberg, M. (1999). Crop husbandry at Miri Qalat Makran, SW Pakistan (4000-2000 B.C.). Veget Hist. Archaebot 8 (1), 3–12. doi:10.1007/BF02042836
Wan, Z. W., Yang, X. Y., Li, M. Q., Ma, Z. K., and Ge, Q. S. (2012). China modern starch grain database. Quat. Sci. 32 (2), 371–372. doi:10.3969/j.issn.1001-7410.2012.02.24
Wang, C., Lu, H. Y., Zhang, J. P., Ye, M. L., and Cai, L. H. (2015). Phytolith evidence of millet agriculture in the late Neolithic archaeological site of Lajia, Northwestern China. Quat. Sci. 35 (1), 1–9. doi:10.3969/j.issn.1001-7410.2015.01
Wang, C., Lu, H., Zhang, J., He, K., and Huan, X. (2016). Macro-process of past plant subsistence from the Upper Paleolithic to Middle Neolithic in China: a quantitative analysis of multi-archaeobotanical data. PLoS ONE 11 (2), e0148136. doi:10.1371/journal.pone.0148136
Wang, Q., Kamijyou, O., and Luan, F. S. (2013). The Study of Stone Saddle Querns and Rollers at Beiqian Site, Jimo, Shandong Province. Dongfang Archaeol. 1, 154
Wang, Q. Q., and Zhen, Q. (2021). Analysis on subsistence composition and structure of Qijia culture in Qinghai province, China. J. Qinghai Norm. Univ. Soc. Sci. 43 (2), 77
Wang, Q., and Yang, H. Y. (2015). Several problems in the study of grinding stone tools. Wenwu Chunqiu 2, 3
Wang, Y. J., and Lu, H. Y. (1993). The Study of Phytolith and its Application. Beijing: China Ocean Press
Wright, K. I. (1994). Ground-Stone Tools and Hunter-Gatherer Subsistence in Southwest Asia: Implications for the Transition to Farming. Am. Antiq. 59 (2), 238–263. doi:10.2307/281929
Wu, W. W. (2015). Preliminary analysis of the economics of production at the Chahai site in Fuxin, Liaoning: Indications from the results of the analysis of stone tools starch granules. Agricul Archaeol. 3, 1
Wu, W. W. (2014). Research on the Subsistence of Peiligang Period in North China. Jinan (Chinese): Shandong University. [Ph.D. dissertation].
Wu, W. W., Yang, X. Y., and Jin, G. Y. (2011). The application of starch analysis in archaeology: Case studies on Yuezhuang and other sites. Dongfang Archaeol. 1, 330
Wu, Y., Wang, C., and Hill, D. V. (2012). The transformation of phytolith morphology as the result of their exposure to high temperature. Microsc. Res. Tech. 75 (7), 852–855. doi:10.1002/jemt.22004
Yang, B., Qin, C., Bräuning, A., Osborn, T. J., Trouet, V., Ljungqvist, F. C., et al. (2021). Long-Term Decrease in Asian Monsoon Rainfall and Abrupt Climate Change Events Over the Past 6,700 Years. Proc. Natl. Acad. Sci. U.S.A. 118 (30), e2102007118. doi:10.1073/pnas.2102007118
Yang, X., Ma, Z., Wang, T., Perry, L., Li, Q., Huan, X., et al. (2014). Starch grain evidence reveals early pottery function cooking plant foods in North China. Chin. Sci. Bull. 59 (32), 4352–4358. doi:10.1007/s11434-014-0500-6
Yang, X., and Perry, L. (2013). Identification of ancient starch grains from the tribe Triticeae in the North China Plain. J. Archaeol. Sci. 40 (8), 3170–3177. doi:10.1016/j.jas.2013.04.004
Yang, X., Wan, Z., Perry, L., Lu, H., Wang, Q., Zhao, C., et al. (2012a). Early millet use in northern China. Proc. Natl. Acad. Sci. U.S.A. 109 (10), 3726–3730. doi:10.1073/pnas.1115430109
Yang, X. Y., Kong, Z. C., Liu, C. J., and Ge, Q. S. (2010). Morphological characteristics of starch grains of millets and their wild relatives in North China. Quat. Sci. 30 (2), 364–371. doi:10.3969/j.issn.1001-7410.2010.02.13
Yang, X., Zhang, J., Perry, L., Ma, Z., Wan, Z., Li, M., et al. (2012b). From the modern to the archaeological: Starch grains from millets and their wild relatives in China. J. Archaeol. Sci. 39 (2), 247–254. doi:10.1016/j.jas.2011.09.001
Ye, M. L. (2015). Ecological adaptation of agricultural development in Qijia culture: A preliminary study on primitive grass farming—Taking the Lajia site in Qinghai as an example. Agricul Archaeol. 6, 19
Zhang, C. (2013). Floating Selection Carbonized Plants Result and Analysis at the Lajia Site in Minhe County, Qinghai Province. Xi’an (Chinese): Northwest University. [Master's thesis].
Zhang, G. L., Wang, S. Z., Ferguson, D. K., Yang, Y. M., Liu, X. Y., and Jiang, H. E. (2015a). Ancient plant use and palaeoenvironmental analysis at the Gumugou Cemetery, Xinjiang, China: Implication from desiccated plantremains. Archaeol. Anthropol. Sci. 9 (2), 145–152. doi:10.1007/s12520-015-0246-3
Zhang, X. L., Qiu, S. H., Zhong, J., Huang, Y., Lu, X. F., and Du, H. (2015b). A report on the dating of radiocarbon (41). Archaeol 7, 107.
Zhang, X. L., Wang, J. X., Xian, Z. Q., and Qiu, S. H. (2003). Studies on ancient human diet. Archaeol 2, 62
Zhang, X. L., Xu, D. G., He, Y. L., and Qiu, S. H. (2017). Analysis of Carbon and Nitrogen Stable Isotopes of Human Bones unearthed from M54 at Yinxu Site. Archaeol 3, 100
Zhang, X. L., and Ye, M. L. (2016). Preliminary discussion on the diet of ancestors at the Lajia site: carbon and nitrogen stable isotope analysis of human bones unearthed from the disaster site of the Lajia site. Cult Relics South. China 4, 197–202. doi:10.3969/j.issn.1004-6275.2016.04.027
Zhang, X. L., Ye, M. L., Qiu, S. L., and Zhong, J. (2014). 14C dating and Preliminary Analysis of Lajia Site in Minhe Country. Archaeol 11, 91–104. (in Chinese).
Zhang, Y. H., Weng, Y., Yao, L., Zhang, J. Z., Zhou, Y. J., Fang, F., et al. (2011). Identification and analysis of starch granules on the surface of the slabs from peiligang site. Quat. Sci. 31 (5), 891–899. doi:10.3969/j.issn.1001-7410.2011.05.14
Zhao, K., Li, X., Zhou, X., Dodson, J., and Ji, M. (2013). Impact of agriculture on an oasis landscape during the late Holocene: Palynological evidence from the Xintala site in Xinjiang, NW China. Quat. Int. 311, 81–86. doi:10.1016/j.quaint.2013.06.035
Zhao, Z. J. (2015). Research on the introduction of wheat into China--archaeobotany data. Cult Relics South. China 3, 44
Keywords: prehistoric diet, the lajia site, dental calculus, carbon and nitrogen isotopes, starch grain, phytolith
Citation: Ma Z, Liu S, Li Z, Ye M and Huan X (2022) Human Diet Patterns During the Qijia Cultural Period: Integrated Evidence of Stable Isotopes and Plant Micro-remains From the Lajia Site, Northwest China. Front. Earth Sci. 10:884856. doi: 10.3389/feart.2022.884856
Received: 27 February 2022; Accepted: 08 June 2022;
Published: 08 July 2022.
Edited by:
Ying Guan, Institute of Vertebrate Paleontology and Paleoanthropology (CAS), ChinaCopyright © 2022 Ma, Liu, Li, Ye and Huan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiujia Huan, aHVhbnhpdWppYUBtYWlsLmlnZ2Nhcy5hYy5jbg==