 Weiwei Wang1
Weiwei Wang1 Kim Dung Nguyen2Hai Dang Le3
Kim Dung Nguyen2Hai Dang Le3 Chunguang Zhao4Mike T. Carson5Xiaoyan Yang6,7,8
Chunguang Zhao4Mike T. Carson5Xiaoyan Yang6,7,8 Hsiao-chun Hung1*
Hsiao-chun Hung1*- 1Department of Archaeology and Natural History, The Australian National University, Canberra, ACT, Australia
- 2Vietnam Association of Archaeology, Hanoi, Vietnam
- 3Institute of Archaeology, Vietnam Academy of Social Science, Hanoi, Vietnam
- 4Department of Archaeology, Wuhan University, Wuhan, China
- 5Micronesian Area Research Center, University of Guam, Mangilao, GU, United States
- 6Key Laboratory of Western China’s Environmental Systems (Ministry of Education), College of Earth and Environmental Sciences, Lanzhou University, Lanzhou, China
- 7State Key Laboratory of Tibetan Plateau Earth System, Resources and Environment (TPESRE), Beijing, China
- 8Group of Alpine Paleoecology and Human Adaptation (ALPHA), Institute of Tibetan Plateau Research, Chinese Academy of Sciences, Beijing, China
Mainland Southeast Asia experienced a long, sustained period of foraging economy before rice and millet farming spread into this area prior to 4,000 years BP. Although hundreds of individuals from dense cemeteries are found in several hunter-gatherer sites in Guangxi, Southern China, and Northern Vietnam, dating from the early to middle Holocene (ca. 9,000–4,500 years BP), so far, little has been known about food sources in these pre-farming contexts. In particular, plant food resources have been unclear, although they likely were crucial to supporting rather large populations of hunter-gatherers in this region. To investigate this issue, micro plant remains, including starches and phytoliths, were recovered from stone tools excavated at the Cai Beo site in Ha Long Bay of coastal Northeastern Vietnam, and those findings revealed new understanding of the ancient diet. Examinations of those residues indicated that the hunter-gatherers at Cai Beo as early as 7,000–6,000 years BP exploited a broad spectrum of plants, such as taros, yams, acorns, palms, and more. This study exemplifies how maritime hunter-gatherers interfaced with the local plants and generated population growth from about 7,000 to 4,500 years BP. The results help us to conceptualize the early exploitation, management, and potential cultivation of subtropical and tropical plants over the broad geography of Asia and the Pacific before the arrival of rice and millet farming. In particular, the result validates the significance of roots and tubers in the ancient subsistence economy of Southeast Asia. Moreover, from the archaeological context of 4,500 to 4,000 years BP, the rice discovered in this study represents one of the earliest known in Mainland Southeast Asia.
1 Introduction
Mainland Southeast Asia (MSEA) experienced a long, sustained period of foraging economy before agriculture spread into this area more than 4,000 years ago (Bellwood et al., 2011; Castillo et al., 2018; Higham, 2021). Before rice in coastal Southern China and MSEA, several complex hunter-gatherer groups and affluent village settlements emerged around 7,000 years BP or even earlier. They are remarkable in terms of their population size and social development (Zhang and Hung, 2012; Hung, 2019). This phenomenon probably was related to an advanced subsistence economy, wherein people obtained marine resource supplies and exploited various starch-rich plants, which provided carbohydrates and produced energy to support the stable growth of coastal societies.
Historical records and modern observations have shown clearly about the importance of tropical, starch-rich plants, such as roots and tubers, palms, bananas, and others, among the traditional societies living in the subtropical and tropical areas of Asia and the Pacific islands (Barton, 2012; Matthews et al., 2012). Therefore, these same food sources may have supported the initial cultural development and population growth in these areas. However, until now, the archaeobotanical evidence has been limited across the region, other than in Papua New Guinea, which has been considered as the forerunner in exploiting, managing, and domesticating taros, yams, and bananas (Denham et al., 2003; Fullagar et al., 2006; Loy et al., 2015). Nonetheless, the hypothesis of the original domestication of taro (Colocasia esculenta, Araceae) in Papua New Guinea has been challenged by recent DNA analysis (Ahmed et al., 2020) that instead indicated an origin generally within Southeast Asia (Matthews, 1991; Yoshino, 2002; Matthews, 2014; Ahmed et al., 2020).
Through archaeobotanical studies at a few sites in coastal Southern China and Island Southeast Asia (ISEA), the discovery of a broad spectrum of plants, including taros and yams, sago palms, bananas, tree nuts, and others (Barton and White, 1993; Barker et al., 2007; Oliveira, 2008; Yang et al., 2013; Yang et al., 2017) has provided new insights into the exploitation of subtropical and tropical plants from the late Pleistocene to middle Holocene periods.
In the previous studies in MSEA, most efforts concentrated on the emergence of rice and millet agriculture (Weber et al., 2010; Nguyen, 2013; Barron et al., 2017; Castillo et al., 2018). Therefore, the knowledge about ancient plant usage beyond rice was limited to a few woody plants, palms, bamboos, and wild fruits (Pyramarn, 1989; Bowdery, 1999; Nguyen, 2008). In such a research background, systematic studies of the pre-farming period (i.e., prior to 4,500–4,100 years BP) in MSEA can fill the knowledge gap in the Asia-Pacific archaeobotanical record.
Cai Beo, a representative ancient coastal settlement in Northeastern Vietnam dating to ca. 7,000 to 4,000 years BP, is ideal for the archaeobotanical studies that can address such a research question. Furthermore, the integrated findings of ancient starches and phytolith analyses could clarify the subsistence strategies and human-environment relationships of the ancient coastal people in Southeast Asia (SEA).
2 Materials and Methods
2.1 Site Description and Sample Collection
The Cai Beo site (N 20°43′8″, E 107°3′2″), nearly 4 m above current sea level, is located on Cat Ba Island, the largest island in Ha Long Bay in the Hai Phong Municipality (Figure 1). Cai Beo was reported first by French archaeologist Madeleine Colani who then conducted a small-scale test excavation in 1938 (Colani, 1938). Later, in 1972, 1973, 1981, 1986, and 2006, the site was excavated five times by Vietnamese archaeologists, opening a total excavated area of 449 m2 (Nguyen, 2005; Nguyen, 2009). The known distribution area of Cai Beo is around 18,000 square meters.
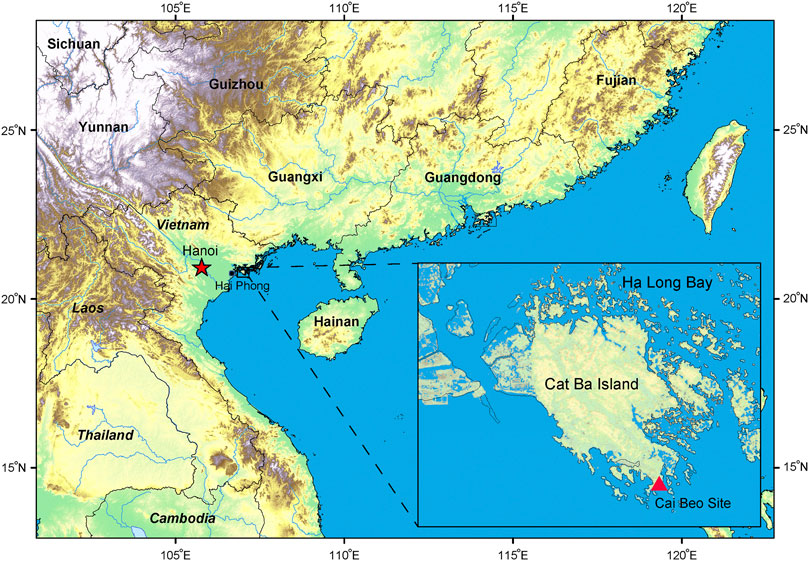
FIGURE 1. Location of the Cai Beo site. The site is shown by a red triangle.
Several hundreds of kg of large fish bones and marine shells were reported from the excavations at Cai Beo, indicating a sea-oriented economy (Nguyen, 2019). Additionally, rich terrestrial animal remains, such as masked civet, brown bear, deer, elk, wild boar, elephant and monkey, were reported from the site (Li, 2019), indicating an ancient forest environment at that time.
The deposit of Cai Beo contains three cultural layers, named as Pre-Halongian, Proto-Halongian, and Ha Long Culture during the 1973 excavation. However, since 1981 excavation, it has been generally suggested that the deposit of the site involves a local “Cai Beo phase” (subdivided into Cai Beo phases 1 and 2) and the later “Ha Long phase” (Figure 2). The lower two cultural layers of Cai Beo, phases 1 and 2, span about 7,000 through 5,000 years BP. This estimate is based on a radiocarbon date (ZK-328-0, see below) and comparisons with similar findings at another five Cai Beo cultural sites in Ha Long Bay (Nguyen, 2005; Nguyen, 2009).
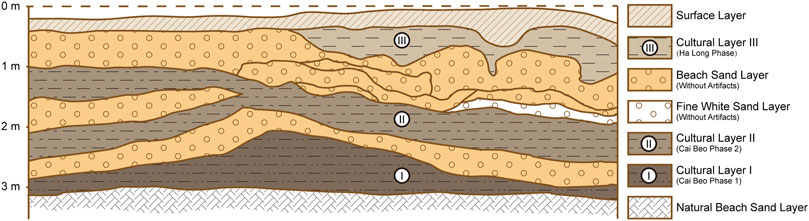
FIGURE 2. Trench profile of south wall of H3, Cai Beo site (1973 excavation, after Nguyen, 2009: 62).
The lowest layer (Cai Beo phase 1, see Figure 2: Cultural Layer I) represents the earliest occupation of the site. This layer is 2.4–3.2 m deep, consisting of sand deposits mixed with round gravels, debris, marine fish bones, oysters, and other seashells. Large numbers of stone tools were discovered, including many grinding tools (Figure 3A, all of which exhibited visible usage wear. Most stone tools are flaked chopping tools produced by the direct strike method. Additional items were disc-shaped tools, pointed tools, ¼ round pebble tools, stone hammers, stone anvils, and others. The so-called “polished stone tools” unearthed from this layer had shown sharpening only on the blade part. The discovered potsherds are small in number and size, characterized as thick and coarse. The middle cultural layer (see Figure 2: Cultural Layer II/Cai Beo Phase 2) is 1.2–2.4 m deep, and polished stone axes appeared in this layer.

FIGURE 3. Typical stone tools collected from the Cai Beo site (Scale bar: 5 cm). (A) Grinding stone, (B) Muller/Pounder, (C) Pitted stone, (D) Stone tool with “Ha Long Mark”, (E) Scraper, (F) Pointed tool.
The radiocarbon date (ZK-328-0) of 6,893–6,391 cal. years BP (5,810 ± 115 uncal. years BP, half-life: 5,730 years) (or 5,645 ± 115 uncal. years BP, half-life: 5,570 years) was obtained from fish bones (Laboratory of the Institute of Archaeology, 1977) excavated from the Cai Beo cultural layer. Other time indicators include the characteristics of human burial practice, craniometry, pottery and lithic remains (Nguyen, 2009). For instance, similar Haolizhuo (oyster picks) lithic tools were found at Cai Beo cultural layers widely distributed along the coastal southern China sites in Fujian, Guangdong, and Guangxi from 7,000 to 5,000 years BP (Hung, 2019). A flexed burial unearthed from the Cai Beo cultural phase has been identified as the Australo-Papuan affinity by Nguyen Lan Cuong (Nguyen, 2009). Both the flexed burial and the Australo-Papuan affinity are the key cultural indicators of hunter-gatherers in pre-Neolithic Southern China and Southeast Asia (Higham, 2013; Hung et al., 2017; Matsumura et al., 2019).
During the 1981 excavation, the lowest cultural layer was noticed in a position beneath and therefore pre-dating a marine transgression layer (about 20 centimeter thickness, a sterile gravel layer) that appeared in the depth of 2.4 meters under the square surface. Therefore, the earliest occupation at the site must have pre-dated this marine transgression (Nishimura, 2006) that had occurred at 6000 to 5500 years ago in the area of Ha Long Bay (Nguyen and Tran, 2009). This dating constraint further supports the proposed age of the Cai Beo cultural phase 1 as preceding the range of 6000–5500 cal. years BP.
The upper layer of the Cai Beo site represents the Ha Long cultural phase (Figure 2: Cultural Layer III), generally considered as starting around 4,500 years BP (Nishimura, 2006; Nguyen, 2009; Peng, 2018). At this site in particular, the associated layer is 0.2–1.2 m deep. New forms of wholly polished stone tools included well polished shouldered axes and quadrangular adzes. Other stone objects were chisels, hammers, sandstones with the so-called “Ha Long mark”, and stone rings. Compared with the findings from the preceding Cai Beo cultural layers, the pottery in the Ha Long cultural layer appeared in large quantities and in more varied forms and decorations, including the use of a wheel-thrown technique in some cases.
More than 38 sites around Ha Long Bay have been grouped into the Ha Long Culture in terms of the artifact association and time period (Nguyen, 2019). Among those Ha Long cultural sites, radiocarbon dates show an age post-dating the older “Cai Beo” context. The available C14 dates from several Ha Long cultural sites concentrated from ca. 4,500 through 3,500 cal. years BP (Chen, 2007; Nguyen, 2009; Nguyen and Tran, 2009; Nguyen and Clarkson, 2013; McColl et al., 2018; Peng, 2018). For instance, one of the C14 dates from Ba Vung site is 4,727–4,517 cal. years BP (4,100 ± 40 uncal. years BP, charcoal sample) (Chen, 2007), and another Ha Long cultural site of Hon Hai Co Tien is dated to 4,381–3,926 cal. years BP (3,755 ± 60 uncal. years BP, human bone sample) (McColl et al., 2018). At the Cai Beo site, the pottery and other artifacts from the Ha Long layer generally accord with the expectations of the early Ha Long Culture association.
During this project, we had submitted 12 samples from Cai Beo, including one human skull, one deer horn, one deer tooth, five mammal bones, and four marine fish bones, to Beta Analytic for AMS dating. Unfortunately, a reliable collagen fraction could not be isolated and purified from any of these samples. As a result, the bone material cannot be dated. However, the basic chronology and two major cultural phases of the site can be reconstructed through multiple lines of evidence as has been outlined here.
2.2 Micro Plant Extraction and Identification
In total, 22 stone tools from Cai Beo were examined in this study for micro-plant remains. The samples include grinding stones, mullers/pounders, pitted stones, stone tools with the so-called “Ha-Long Mark”, scrapers, and pointed tools (Figure 3; Table 1). In addition, four control samples were collected from the non-use surface of stone tools and the storage in Hai Phong and Hanoi to detect modern contamination. All of the stone tools analyzed in this study had been excavated by Kim Dung Nguyen and colleagues from 1972 to 1986 from the Cai Beo site, and they were stored at the Hai Phong Museum and Institute of Archaeology in Hanoi.
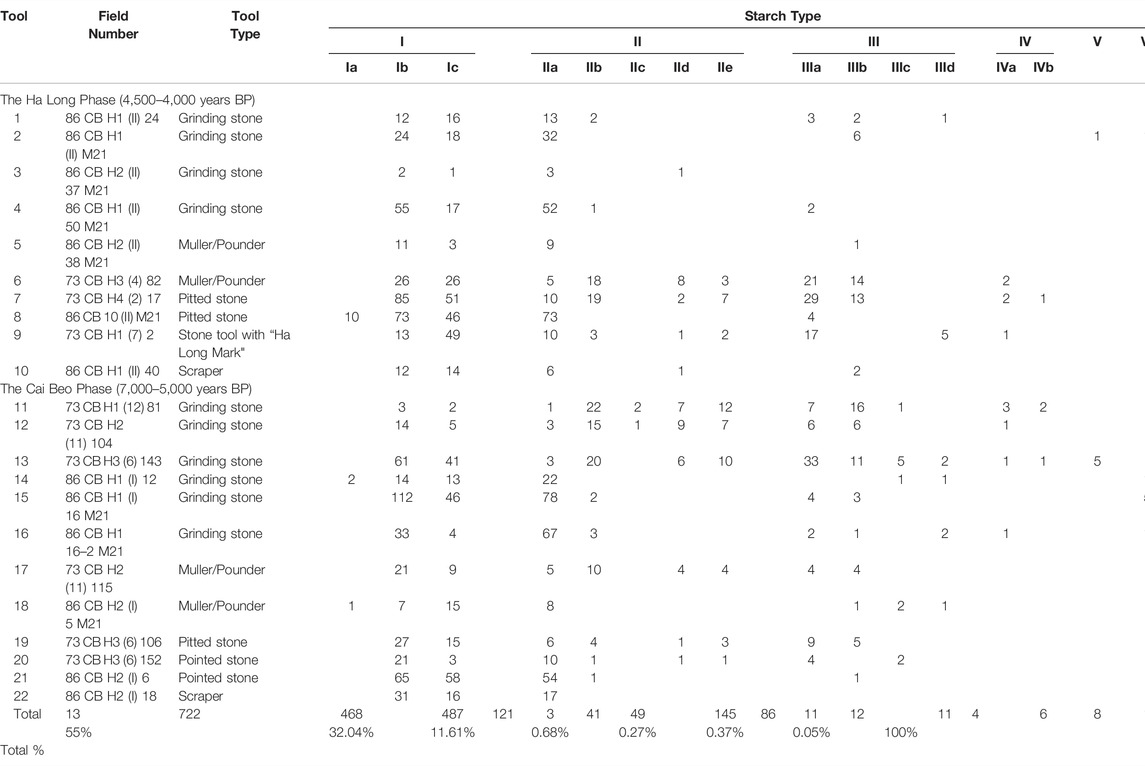
TABLE 1. Types and numbers of starches recovered from the study of stone tools from the Cai Beo site.
The sediments and dust on each tool’s surface were first rinsed with distilled water and then cleaned in an ultrasonic bath with distilled water for 5 minutes to recover the residues. Next, the ultrasound mixtures were transferred to test tubes, which were processed to recover the micro plant remains, including starches and phytoliths, in the Department of Archaeology and Natural History laboratory, The Australian National University.
The extraction process of starch and phytolith followed the process of previous studies (Lu et al., 2008; Yang et al., 2013; Pearsall, 2016; Deng et al., 2017). First, the starch samples were isolated with LST (lithium heteropolytungstate) heavy liquid (density 1.9), mounted on a slide in a solution of 10% glycerine and 90% distilled water, then sealed with nail polish. For the preparation of the phytolith samples, the residues were rinsed three times with distilled water, and then the samples were separated with heavy liquid (density 2.35). After the above steps, the samples were rinsed twice with distilled water and then one more time with 30% ethyl alcohol. Finally, the phytolith samples were mounted on the slide with Canada Balsam. The slides of starch and phytolith were observed, measured, and counted under the optical microscope (Machine model: Olympus BX-51), respectively.
The identification of ancient starches is based on the modern reference collections from Vietnam by the authors of this study, the database (http://cmsgd.igsnrr.ac.cn/) built by the Institute of Geographic Sciences and Natural Resources Research (IGSNRR) (Yang et al., 2018), and other related published studies from tropical and subtropical areas of Asia and the Pacific (Fullagar et al., 2006; Lentfer, 2009). Furthermore, the identification of phytolith types was in accordance with the International Code for Phytolith Nomenclature and published documents (Wang and Lu, 1993; Katharina et al., 2019).
3 Results
3.1 Starch Grains
In total, 2,364 starch grains were recovered from the 22 stone tools (Table 1). Only very few starches were found in the control samples of non-use contexts, indicating that the starches recovered from the use surfaces of stone tools were indeed related with the ancient use. One hundred seventy-six starch grains could not be categorized due to the lack of diagnosable characteristics. In comparison, the other 2,188 starch grains can be classified into seven types and 17 subtypes based on their morphological features and biological attributes after comparison with modern references (Figures 4–6). The details of each type are described below.
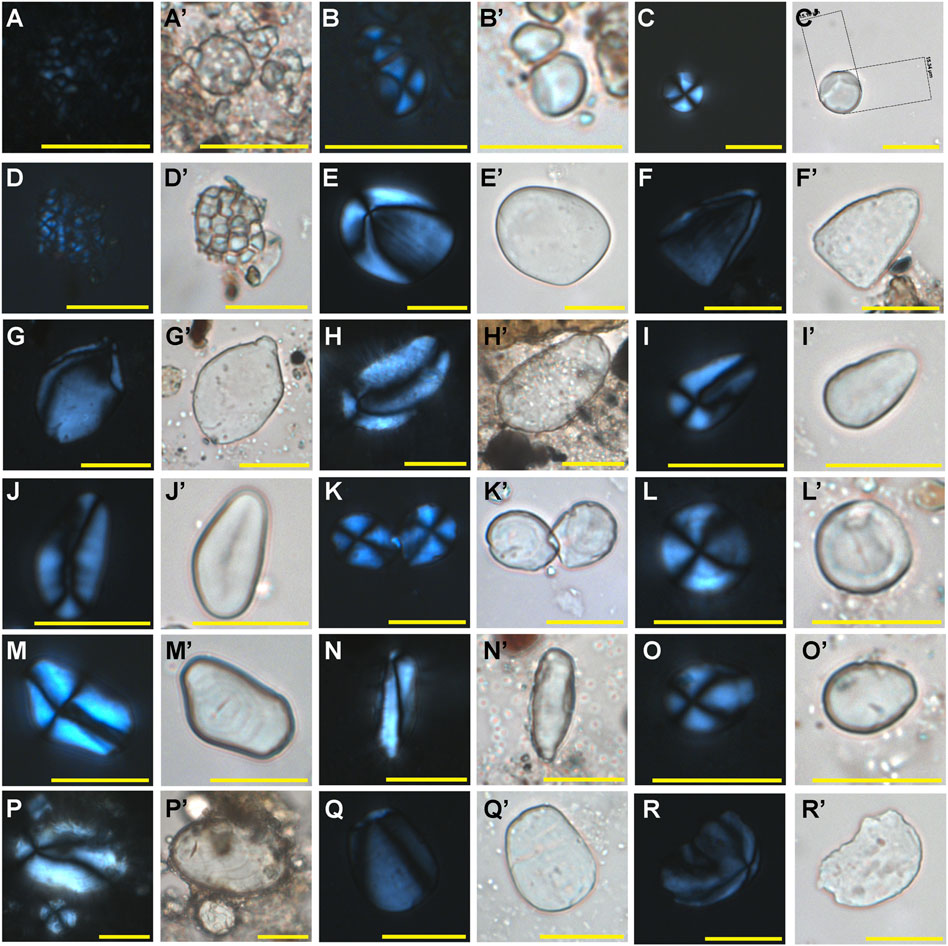
FIGURE 4. Ancient starches recovered from residues on the stone tools (under polarized and brightfield light). (A-A′) Type Ia, Colocasia esculenta, (B-B′) Type Ib, Colocasia spp., (C-C′) Type Ic, Alocasia spp., (D-D′) Type IIa, Dioscorea esculenta, (E-E′) Type IIb, Dioscorea alata, (F-F′) Type IIc, Dioscorea bulbifera, (G-G′) Type IId, Dioscorea sp., (H-H′) Type IIe, Dioscorea sp., (I-J’) Type IIIa, Quercus sp., (K-K′) Type IIIb, Cyclobalanopsis sp., (L-L′)Type IIIc, Lithocarpus sp., (M-M′) Type IIId, Castanopsis sp., (N-N′) Type IVa, Arenga pinnata, (O-O′) Type IVb, Caryota urens, (P-P′) Type V, Angiopteris sp., (Q-Q′) Type VI, Kaempferia sp., (R-R′) Type VII, Musa sp. Scale bar, 20 μm.
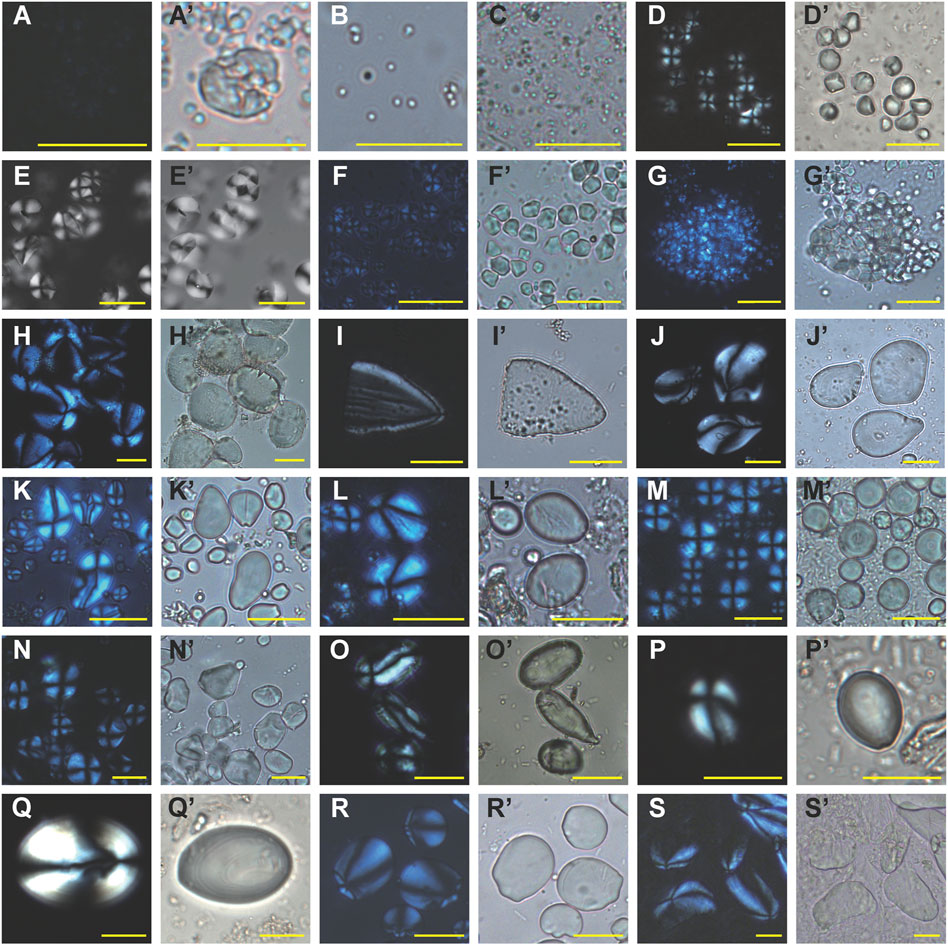
FIGURE 5. Modern starch references relevant to this study (under polarized and brightfield light). (A-A′) Colocasia esculenta, (B) Colocasia antiquorum, (C) Colocasia sp., (D-D′) Colocasia konishii, (E-E′) Alocasia macrorrhizos (picture is sourced from (Fullagar et al., 2006)), (F-G′) Dioscorea esculenta, (H-H′) Dioscorea alata, (I-I′) Dioscorea bulbifera, (J-J′) Dioscorea sp., (K-K′) Quercus franchetii, (L-L′) Cyclobalanopsis phanera, (M-M′) Lithocarpus litseifolius, (N-N′) Castanopsis hystrix, (O-O′) Arenga pinnata, (P-P′) Caryota urens, (Q-Q′) Angiopteris yunnanensis, (R-R′) Kaempferia galanga, (S-S′) Musa sp. Scale bar, 20 μm.
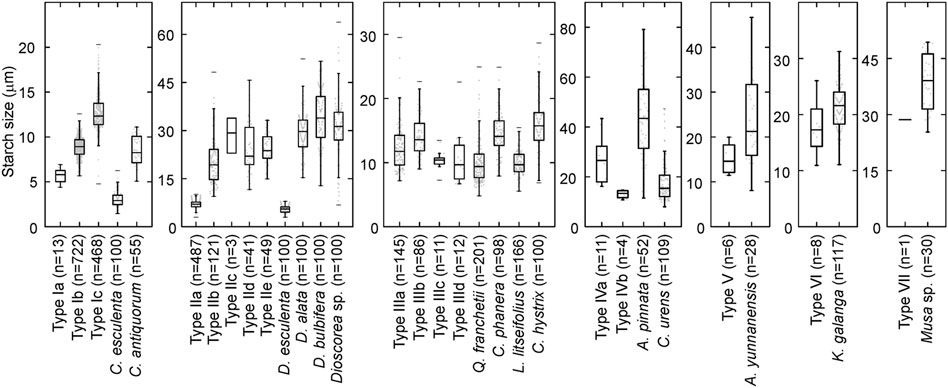
FIGURE 6. Starch sizes from Cai Beo compared with modern reference samples.
3.1.1 Type I Starch
Type I starch grains (N=1,203, 55% of the total) are identified as Aroids. They are classified into three subtypes, mainly according to their sizes.
Type Ia starch grains (N=13) are usually smaller than 5 μm in dimension, and their shapes are difficult to distinguish. As a result, we could not identify all of them under the microscope. However, if the grains appeared in sheets or as clusters (compound grains), then they were easier to find (Figures 4A–A’). Type Ia starch grains best matched the features of taros (Colocasia esculenta) after comparing them to our reference material (Figures 5A–A’). Notably, large quantities of small granules were extracted from 21 stone tools. Although the small granule sizes created difficulty in observing their cross-arms under the polarized light, they nonetheless resembled the starch grains of wild taro (Colocasia antiquorum) or Alocasia sp. (Figures 5B,C).
Type Ib starch grains (N=722) are spherical, sub-rounded, or rounded polygonal with multiple facets (Figures 4B–B’). Their sizes range from 5.67 to 12.59 μm, with the mean size being 8.87 ± 1.19 μm. These features are most consistent with the Colocasia spp. (Figures 5D–D’).
Type Ic starch grains (N=468) exhibit shapes similar to Ib starch grains (Figures 4C–C’), except that their sizes are much larger, with the mean size of 12.73 ± 1.92 μm. About 96% of starch grains are larger than 10 μm. Many of the underground storage organs from the Araceae plants produce this kind of starch. However, this type best matches the features of the Alocasia spp. (Figures 5E–E’).
3.1.2 Type II Starch
Type II starch grains (N=701, 32.04% of the total) exhibit the distinct morphology that characterizes them into the Dioscorea genus, including five subtypes.
Type IIa starch grains (N=487) are characterized by their small size, polygonal shape, and centric hilum (Figures 4D–D’). Only a few species of Dioscorea produce this kind of starch (Fullagar et al., 2006; Hang et al., 2006). Type IIa starch grains are most comparable with tubers of the lesser yam (Dioscorea esculenta) (Figures 5F–G’).
Type IIb starch grains (N=121) are generally triangular or elliptical in shape with eccentric hilum (Figures 4E–E’). The mean size is 20.26 ± 7.1 μm. They best match the characteristics of the purple yam (Dioscorea alata) (Figures 5H–H’).
Three starch granules are classified as Type IIc, resembling angular triangles with flat bases (Figures 4F–F’). This granule type was easy to distinguish from others due to its angular shape. This type could be identified as “air potatoes” (Dioscorea bulbifera) (Figures 5I–I’).
Type IId starch grains (N=41) are irregular ovate shapes, with highly eccentric hilum and bent cross-arms (Figures 4G–G’). This type closely resembles a modern reference belonging to the Dioscorea genus, collected from local markets in the Yunnan Province of China (Figures 5J–J’).
Type IIe starch grains (N=49) resemble elongated ovals with eccentric hilum (Figures 4H–H’). Their grain sizes range from 14.9 to 46.68 μm, with the mean size being 26.15 ± 6.59 μm. This type most likely comes from a kind of yam (Dioscorea sp.), but more comparable reference material will need to be collected in order to ascertain a species.
3.1.3 Type III Starch
Type III starch grains (N=254, 11.61% of the total) share the same features with acorns (Fagaceae). Therefore, they are classified into four subtypes.
Type IIIa starch grains (N=145) are irregularly oval or droplet-shaped (Figures 4I–J’), which are the typical features of the Quercus sp. (Figures 5K–K’).
Type IIIb starch grains (N=86) are oval or bell-shaped. The hilum is eccentric, and sometimes the lamellae are visible (Figures 4K–K’). Compared with our reference material, this type most closely resembles the species of the Cyclobalanopsis genus (Figures 5L–L’).
Type IIIc starch grains (N=11) are round with centric hilum. Some granules present linear fissures and visible lamellas (Figures 4L–L’), showing the same features as the Lithocarpus sp. (Figures 5M–M’).
Type IIId starch grains (N=12) are characterized by their polygonal shape, centric hilum, and visible lamellae (Figures 4M–M’). This type best matches the feature of the Castanopsis sp. (Figures 5N–N’).
3.1.4 Type IV Starch
Type IV starch grains (N=15, 0.68% of the total) can be sub-divided into two groups. Eleven of the 15 starch grains (Type IVa), ranging 16.2–40.93 μm in size, with an elongated, irregular oval shape and highly eccentric hilum (Figures 4N–N’), best match those from sugar palm (Arenga pinnata) (Figures 5O–O’) from our reference collection. The other four starch grains (Type IVb) are small ovate in shape with faint lamellae, similar to the starch from Kitul Palm (Caryota urens) (Figures 4O–O’, Figures 5P–P’).
3.1.5 Type V Starch
Type V starch grains (N=6, 0.27% of the total) have a large oval shape with well defined lamellae or irregularly compounded with complex extinction crosses (Figures 4P–P’). Based on the comparative collections, Type V starch grains share the typical features of the terrestrial fern (Angiopteris sp.) (Figures 5Q–Q’), from which the starches in rhizomes are known as substitutes for staple foods (Liu et al., 2012).
3.1.6 Type VI Starch
Eight starch grains are classified as Type VI starch. They are oval in shape with highly eccentric hilum, ranging 10.94–26.03 μm in size (Figures 4Q–Q’). The morphological characteristics of Type VI starch grains are typical in gingers (Zingiberaceae), and they most closely resemble the rhizomes of the Kaempferia sp. (Figures 5R–R’) from our reference material.
3.1.7 Type VII Starch
Only one starch grain belonging to Type VII. It is 28.61 μm in length, with an irregular ovoid shape and wrinkled texture (Figures 4R–R’) similar to the starch characteristics of bananas (Musa sp.) (Figures 5S–S’).
3.2 Phytoliths
Given the fact that most roots and tubers do not produce phytoliths, the phytoliths extracted from the surface of the stone tools are scarce. However, 80 phytoliths were recovered (Figure 7), the majority of which could be classified taxonomically only to the family level.
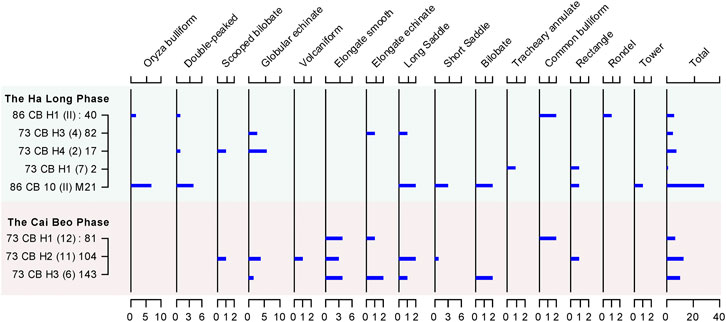
FIGURE 7. Phytolith assemblages recovered from surface residues on stone tools.
Fifteen globular echinate phytoliths are from palms (Aracaceae) (Figure 8A. Additionally, phytoliths from rice were discovered, including the scale decorated bulliform produced by rice leaf tissue, double-peaked phytolith produced by rice glumes, and parallel bilobate from stems (Figures 8B–E). The percentage of rice bulliform phytoliths with ≥9 fish-scale decorations can be used to differentiate wild rice from domesticated rice (Saxena et al., 2006; Huan et al., 2015). However, the numbers of rice bulliform phytoliths recovered in Cai Beo is small (n=9) and the decorations were obscure to observe, and therefore we could not ascertain whether they were wild or domesticated. One volcaniform phytolith was identified, originally from the leaf of banana (Musa sp.) (Figure 8F).
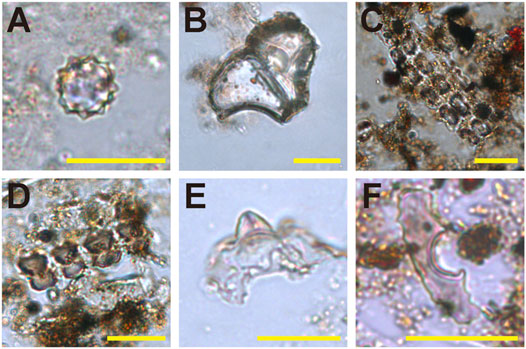
FIGURE 8. Examples of phytoliths extracted from the Cai Beo site. (A) Palm spheroid echinate, (B) Bulliform flabellate from Oryza, (C–D) Bilobates parallel, (E) Double-peaked glume cell, (F) Volcaniform phytolith. Scale bar, 20 μm.
4 Discussion
The microfossil studies of plant residues recovered from Cai Beo indicate that the coastal hunter-gatherers utilized a broad spectrum of plants, including taros, yams, acorns, palms, and others. More than 80% of the total starch grains are identified as yams and taros, reflecting the practical economic significance of roots and tubers in the ancient subsistence system of MSEA. This finding constitutes the first validation that taros and yams were essential plant food sources during the pre-farming ancient contexts in the tropical coastal area of MSEA.
4.1 Edible Aroids (55% of the Total Starches)
Edible aroids (family Araceae) comprise many underground food crops grown in tropical and sub-tropical regions. They are essential food crops in India, SEA, and the Pacific islands (Opara, 2003). At several sites in the ISEA and the Pacific, aroids often are discovered together with yams, in which Colocasia, Alocasia, and Cyrtosperma are reported (Barton and White, 1993; Barton, 2005; Fullagar et al., 2006; Barker et al., 2007; Loy et al., 2015). A study of ancient starch remains from the Niah cave in Borneo have identified grains of aroids (Alocasia spp.) from the upper layer of the Hell Trench sequence dating to less than 40,000 years BP (Barker et al., 2002; Barton, 2005).
Taro (Colocasia esculenta) is one of the oldest and most important cultigens in the Indo-Pacific region (Blench, 2012; Spriggs and Matthews, 2012). Nowadays, taro has persisted as one of the most significant staple diets and cultural elements in many parts of SEA. Still, the role of taro in Southeast Asian prehistory has been underestimated due to the lack of palaeobotanical study. Furthermore, the original domestication center of taro is still disputed (Oliveira, 2012). The greatest diversity of wild Colocasia species appears to extend from northeast India to southern China, within the Himalayan region of MSEA (Matthews, 1991). As mentioned, although some suggested independent domestication of taro occurred in Papua New Guinea during the early to middle Holocene (Matthews, 1991; Fullagar et al., 2006; Golson et al., 2017; Golson, 2020), a recent study of chloroplast DNA of taro does not support this hypothesis. Instead, it reveals that the cultivated taro was introduced into Papua New Guinea from SEA after an early or middle Holocene domestication (Ahmed et al., 2020).
Previously, a study in Guangxi of southwest China has recovered taro (Colocasia esculenta) starches from the Zengpiyan cave site (ca. 12,500 to 7,600 years BP). Starches were discovered on the excavated stone tools (Lu, 2003). Our new finding from Cai Beo confirms the early and long-term exploitation of taro in MSEA, which in accordance with archaeological, ethnobotanical, taxonomic, and genetic studies, helping to reconceptualize the management and domestication process of taro on a larger scale. A future archaeobotanical survey will concentrate on the remains found in older cave sites in Southern China and MSEA.
4.2 Yams (32.04% of the Total Starches)
SEA and its adjacent areas are presumed to be one central origin and diversification center for yams (Arnau et al., 2017). In addition, several species of Dioscorea, such as D. alata, D. esculenta, D. pentaphylla, etc. commonly are consumed as regular dietary foods or as famine foods, fodder, and ethnomedicines in tropical Asia and the Pacific region (Maneenoon et al., 2008; Dutta, 2015; Andres et al., 2017). For example, an ethnobotany study in Peninsular Thailand reported that 15 species of Dioscorea were found in the living areas of the hunter-gatherer Sakai tribe at Banthad Range, in which eight species are consumed as main food sources by the Sakai people there (Maneenoon et al., 2008).
Previously, many archaeobotanical findings of yams have been reported from highland New Guinea, where modern humans possibly exploited yams around 40,000 years ago and integrated them into cultivation before 6,000 years BP (Fullagar et al., 2006; Summerhayes et al., 2010). In ISEA, related archaeobotanical evidence had been sporadic (Paz, 2001; Mijares, 2007; Oliveira, 2008). The starch granules and charred parenchyma from Dioscorea sp. discovered in the Niah cave presumably could date back to 40,000 years ago (Barton, 2005; Barker et al., 2007). Compounded with the lack of sufficient approaches of archaeobotanical works in ISEA, the roots and tubers naturally were preserved quite poorly in the humid tropical environments. Until now, with our new research, the evidence has been missing about Dioscorea consumption dating back to pre-Neolithic or even Neolithic MSEA. The study at Cai Beo has recovered a variety of yams, such as Dioscorea alata, Dioscorea esculenta, and Dioscorea bulbifera. These Dioscorea spp. plants require a set of knowledge and skill in harvesting, grinding, roasting, and processing (Maneenoon et al., 2008; Sharma and Bastakoti, 2010).
4.3 Acorns (11.61% of the Total Starches)
At least four genera of Fagaceae were recovered from Cai Beo, identified as Quercus sp., Cyclobalanopsis sp., Lithocarpus sp., and Castanopsis sp. As seen in other hunter-gatherer societies (Nguyen, 2008; Nguyen, 2014), the exploitation of acorns was an essential practice among the ancient hunter-gatherers in Ha Long Bay.
Several types of tree nuts, such as Juglans, Quercus, Castanopsis, Canarium, are found frequently at Hoabinhian sites (ca. 20,000–9,000 years BP) in Northern Vietnam (Nguyen, 2008; Nguyen, 2013), as well as in other parts of Mainland Southeast Asia (Higham, 2014). Pollen studies have shown that these trees, including Fagaceae plants, were prevalent during the middle Holocene in Northern Vietnam (Nguyen, 2014). The Fagaceae plant resources in Vietnam, rich with 45 species of the genus Quercus, have been identified. Among these 45 species, 38 belong to the subgenus Cyclobalanopsis, and seven species belong to subgenus Quercus (Binh et al., 2018a; Binh et al., 2018b). In addition, Castanopsis sp. and Lithocarpus sp., which belong to the evergreen forest type, can be found on rocky slopes in Cat Ba National Park on Cat Ba Island today (Thin, 1998).
The processing tasks of acorns as foods are diverse and labor intensive. Many species of acorns were used as tree crops in ancient societies across the world (Cao et al., 2007). The seeds of acorn trees could be consumed after being ground into flour and leached (Mason, 1995). Furthermore, acorn trees often were used as timber for floors, furniture, and traditional remedies (Dolai et al., 2012; Wan Omar et al., 2019).
Ancient acorns were reported from nearby coastal hunter-gatherer sites in the Pearl River Delta in Guangdong, dating to a similar age as the Cai Beo phase (Li, 2020). According to the study by Li (2020), acorns and oak-chestnuts undoubtedly were the essential starch sources for the indigenous communities in the Pearl River Delta during 6,000 through 4,500 years BP, and the use of acorns reduced after 4,500 years BP, about the time when rice farming started to appear. Similarly, our discoveries from Cai Beo suggest that acorns were part of the standard diet of ancient foragers in Ha Long Bay.
4.4 Palms
Micro-remains from Arecaceae are commonly found in ancient sites in Mainland and Island SEA (Yen, 1977; Paz, 2001; Barton, 2005; Oliveira, 2008; Castillo et al., 2020). At Cai Beo, two types of starches from palms were recovered. The Sugar Palm (Arenga pinnata) is an endemic plant to Southeast Asian countries (Haryoso et al., 2019). In Vietnam, it is grown on the highlands in the central or northern parts of Vietnam (Nguyen et al., 2014). It is a versatile plant, and almost all of its physical and production parts can be utilized (Ishak et al., 2013; Azhar et al., 2019).
The Kitul Palm (Caryota urens) naturally inhabits the understory tree stratum in tropical Asia’s moist lowlands and submontane forests (Rangabhashiyam and Selvaraju, 2015). It is a multipurpose palm. The starch extracted from its pith is known as “Kithul flour”, and it is claimed to have health benefits according to folklore and Ayurveda (Wimalasiri et al., 2016).
Moreover, the discovery of palm phytoliths in Cai Beo indicates that the plant naturally grew near the site. A previous study at Xincun (ca. 5,300 through 4,420 years BP) of the Guangdong coast, about 700 km to the north from the Ha Long Bay, demonstrated that the sago-type palms were a primary plant food before the rice in south China (Yang et al., 2013). Although the finding of starches from Arecaceae at Cai Beo is limited (N=15, 0.68% of the total), we cannot exclude the possibility that people had produced and consumed sago in Ha Long Bay.
4.5 Edible Ferns
The starches of a terrestrial fern (Angiopteris sp.) have been identified on the grinding stones excavated from Cai Beo. Edible ferns are among the most common wild food plants used by people worldwide. The stems, rhizomes, leaves, young fronds, shoots, and some whole plants of ferns can be used as food (Mannan et al., 2008). The rhizomes and stems of the Angiopteris sp. are rich in starch, and they are served as food in India and in China (Chen et al., 2010; Liu et al., 2012). In particular, in China, ethnic minorities in Yunnan, Guangxi, Guizhou, and Sichuan provinces of the southwest region consume much more fern species than the other parts of China (Liu et al., 2012).
The border region between the South-eastern Yunnan Province and Northern Vietnam is one of the areas with the richest biological diversity, including that of Angiopteris (Wang et al., 2020). During the historical period of SEA, the ferns of Angiopteris evecta (G. Forst.) Hoffm., Cibotium barometz (L.) J. Smith, Cyathea spp., and Pteridium aquilinum (L.) Kuhn were served as supplementary food sources or used for producing alcohol. These traditions have mostly been abandoned since modern times (De Winter and Amoroso, 2003). The recovered ancient starches of Angiopteris sp. from Cai Beo could reflect an early case of extracting and processing fern starch food in SEA more than 5,000 years ago.
4.6 Rice
Today, the findings of rice remain in MSEA date back to about 4,100–4,000 years BP (Weber et al., 2010; Barron et al., 2017; Nguyen, 2017; Castillo et al., 2018). The evidence for rice farming in the southern coastal areas of China dates back to about 5,000–4,500 years BP (Zhang and Hung, 2010; Yang et al., 2017; Li, 2020). A recent study confirms that the earliest rice and millet appeared together in the Pearl River Delta of Guangdong at 4,800–4,600 cal. years BP (Deng et al., 2022).
Sixteen rice phytoliths (N=16) were identified from the Ha Long cultural layer at Cai Beo. Although the discovered rice phytoliths from the site were too few to conclude whether they were wild or domesticated, importantly the rice phytoliths were restricted solely to the upper layer of this site within the Ha Long Culture association (Figure 7). These findings likely relate to the general context of agriculture that started in the northeastern coast of Vietnam around 4,500 years BP.
These findings emphasize the many new elements of cultural material remains that first appeared in the Ha Long cultural layer of this site. Indeed, further research will be needed to investigate this topic through more radiocarbon dating and a detailed chronology of the sites in this region.
4.7 Kaempferia
The genus of Kaempferia is widespread and cultivated throughout SEA (Nopporncharoenkul and Jenjittikul, 2017). Nowadays, Rhizomes and leaves of Kaempferia galanga often are used as a flavoring in Vietnamese cuisine. However, because the discovered number of Type VI Starch in this study is small (N=8), the ancient use of these plants is unclear at this time.
4.8 Banana
Although only one starch granule and one phytolith of bananas (Musa sp.) were found in Cai Beo, this finding indicates the possibility that these coastal hunter-gatherers may have known about bananas and attempted to exploit them.
5 Cai Beo in a Wider Context
The coastal habitations in Northeastern Vietnam and Southern China experienced a rapid development by 7,000 years BP, reflected in larger settlements and dense population. Several cultural groups contemporary with Cai Beo flourished in inland and offshore Southern China and northern Vietnam, for example as seen in the Xiantouling cultural group in Guangdong coast, the Keqiutou group in Fujian, the Dingshishan group in Guangxi, and the Da But group in Thanh Hoa and Ninh Binh. Even though they presented distinct cultural characteristics, some of the similarities in burial practices, pottery vessels, or stone artifacts indicated the cultural contacts between these hunter-gatherer communities in southern China and northern Vietnam (Zhang and Hung, 2012; Hung, 2019).
These hunter-gatherers shared similar subsistence patterns and likely consumed or utilized specific types of plants and animals. Archaeobotanical research in Guangdong and Guangxi revealed the diverse plant resources exploited by these affluent hunter-gatherers. For example, at least since 9,000 years BP or even earlier, Canarium nuts had been in long-term use by hunter-gatherers in southern China and Southeast Asia (Deng et al., 2019). The ancient settlement of Xincun in coastal Guangdong utilized a wide range of starch-rich plant foods, particularly sago palms, their dominant exploited plant (Yang et al., 2013). In addition, macro and micro remains of Acorns (Quercus, Lithocarpus, Cyclobalanopsis, Castanopsis) have been recovered from several sites in the Pearl River Delta region (Yang et al., 2017; Li, 2020). Domesticated animals such as dogs and pigs served essential roles in farming economies in East Asia. Still, a few of these animals may have been managed or domesticated in older pre-farming contexts. One of the best representatives is the domesticated dog (Canis familiaris) that appeared at least 9,000–7,000 years BP in the contexts of hunter-gatherer sites in Guangxi (Lu, 2010), belonging to the Dingsishan pre-farming group (Lu, 2010; Zhang and Hung, 2012; Hung et al., 2017; Matsumura et al., 2019). Some of the domestic dogs probably arrived in northern Vietnam during the pre-farming context.
Although starches and unidentified charred tubers that may come from Dioscorea sp. or Colocasia sp. were found at the Zengpiyan cave site in Guangxi (Institute of Archaeology, Chinese Academy of Social Sciences, 2003; Lu, 2003), this hypothesis of southern China origin for taro was disputable for the lack of sufficient evidence (Denham et al., 2018). Through our study, the exploitation of a broad range of plant resources by the Cai Beo people, particularly starchy root and tuber crops, including taro, can be confirmed confidently. These geophytes provided the most critical support for the coastal hunter-gatherers before rice farming in Northeastern Vietnam. As mentioned, the large subtropical and tropical area around the Himalaya region, with the high diversity of plants, has been regarded as one of the plant domestication centers for geophytes, especially taro (Matthews, 1991; Zhao, 2011; Matthews, 2014). Under such a wider cultural context, we may reconsider whether geophyte cultivation had been practiced in some ways before the arrival of rice and millet detestation in MSEA.
6 Conclusion
In addition to the evidence of acorns and edible ferns, the Cai Beo research validates the intensive use of taros and yams in ancient MSEA for the first time. The findings provide solid evidence to highlight the significance of tuber foods in early SEA, wherein some root crops likely were managed artificially or formally cultivated in certain degrees before rice and millet agriculture dispersed into this area around 4,500 to 4,100 years ago. Recently, a similar conclusion was proposed for other nearby coastal sites of similar age, specifically where geophyte cultivation possibly had existed in the Pearl River Delta before the time of rice farming (Li, 2020).
The first discovery of rice phytoliths in the Ha Long cultural layer is significant. The result is consistent with the findings of a recent study of ancient DNA, concluding that the Ha Long population (4,381–3,926 cal. years BP from Hon Hai Co Tien) was the admixture between the local hunter-gatherers (the Hoabinhian) and the ancestors of East Asians (McColl et al., 2018) who came to MSEA with rice and/or millet agriculture.
Further systematic research in the chronology of the Cai Beo site, as well as the cultural relation and farming dispersal between coastal southern China and the Ha Long Bay area will contribute to a complete picture of understanding the dynamic and possibly diverse transformation of human dietary habits and the early management, domestication, or translocation of certain animals and plants.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Author Contributions
HH, WW, and KN conceived and designed the study. KN and HL provided the archaeological samples. HH, WW, KN, HL, and CZ collected the study samples. WW analyzed the data. WW and HH wrote the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study was funded in part by the Australian Research Council (Grant Number: DP190101839), the National Natural Science Foundation of China (Grant No. 41930323), and the Chinese Scholarship Council (Grant Number: E94962376).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We appreciate Director Doi Nguyen (Institute of Archaeology, Vietnam Academy of Social Science, Hanoi) and Deputy Director Do Xuan Trung (Hai Phong Museum) for their support of this study.
References
Ahmed, I., Lockhart, P. J., Agoo, E. M. G., Naing, K. W., Nguyen, D. V., Medhi, D. K., et al. (2020). Evolutionary Origins of Taro (Colocasia esculenta) in Southeast Asia. Ecol. Evol. 10 (23), 13530–13543. doi:10.1002/ece3.6958
Andres, C., AdeOluwa, O. O., and Bhullar, G. S. (2017). “Yam (Dioscorea spp.),” in Encyclopedia of Applied Plant Sciences. 2nd ed (San Diego: Academic Press), 435–441. doi:10.1016/b978-0-12-394807-6.00177-5
Arnau, G., Bhattacharjee, R., Mn, S., Chair, H., Malapa, R., Lebot, V., et al. (2017). Understanding the Genetic Diversity and Population Structure of Yam (Dioscorea Alata L.) Using Microsatellite Markers. PLoS One 12 (3), e0174150. doi:10.1371/journal.pone.0174150
Azhar, I., Risnasari, I., MuhdiSrena, M. F., and Riswan, M. F. (2019). The Utilization of Sugar Palm (Arenga Pinnata) by the People Around Batang Gadis Nasional Park Area. IOP Conf. Ser. Earth Environ. Sci. 305, 012016. doi:10.1088/1755-1315/305/1/012016
Barker, G., Barton, H., Beavitt, P., Bird, M., Daly, P., Doherty, C., et al. (2002). Prehistoric Foragers and Farmers in South-East Asia: Renewed Investigations at Niah Cave, Sarawak. Proc. Prehist. Soc. 68, 147–164. doi:10.1017/s0079497x00001481
Barker, G., Barton, H., Bird, M., Daly, P., Datan, I., Dykes, A., et al. (2007). The ‘human Revolution' in Lowland Tropical Southeast Asia: the Antiquity and Behavior of Anatomically Modern Humans at Niah Cave (Sarawak, Borneo). J. Hum. Evol. 52 (3), 243–261. doi:10.1016/j.jhevol.2006.08.011
Barron, A., Turner, M., Beeching, L., Bellwood, P., Piper, P., Grono, E., et al. (2017). MicroCT Reveals Domesticated Rice (Oryza Sativa) within Pottery Sherds from Early Neolithic Sites (4150-3265 Cal BP) in Southeast Asia. Sci. Rep. 7 (1), 7410. doi:10.1038/s41598-017-04338-9
Barton, H. (2005). The Case for Rainforest Foragers: the Starch Record at Niah Cave, Sarawak. Asian Perspect. 44 (1), 56–72. doi:10.1353/asi.2005.0005
Barton, H. (2012). The Reversed Fortunes of Sago and Rice, Oryza sativa, in the Rainforests of Sarawak, Borneo. Quat. Int. 249, 96–104. doi:10.1016/j.quaint.2011.03.037
Barton, H., and White, J. P. (1993). Use of Stone and Shell Artifacts at Balof 2, New Ireland, Papua New Guinea. Asian Perspect. 32 (2), 169–181.
Bellwood, P., Oxenham, M., Hoang, B. C., Dzung, N. K., Willis, A., Sarjeant, C., et al. (2011). An Son and the Neolithic of Southern Vietnam. Asian Perspect. 50, 144–175. doi:10.1353/asi.2011.0007
Binh, H. T., Ngoc, N. V., Bon, T. N., Tagane, S., Suyama, Y., and Yahara, T. (2018a). A New Species and Two New Records of Quercus (Fagaceae) from Northern Vietnam. PhytoKeys 92, 1–15. doi:10.3897/phytokeys.92.21831
Binh, H. T., Ngoc, N. V., Tagane, S., Toyama, H., Mase, K., Mitsuyuki, C., et al. (2018b). A Taxonomic Study of Quercus Langbianensis Complex Based on Morphology and DNA Barcodes of Classic and Next Generation Sequences. PhytoKeys 95, 37–70. doi:10.3897/phytokeys.95.21126
Blench, R. M. (2012). Vernacular Names for Taro in the Indo-Pacific Region: Implications for Centres of Diversification and Spread in Senri Ethnological Studies. Editors M. Spriggs, D. Addison, and P. J. Matthews (Osaka: National Museum of Ethnology) 78, 21–43. doi:10.15021/00002508
Bowdery, D. (1999). Phytoliths from Tropical Sediments: Reports from Southeast Asia and Papua New Guinea. Bull. Indo-Pacific Prehistory Assoc. 18, 159–168. doi:10.7152/bippa.v18i0.11711
Cao, M., Deng, M., and Zhang, D. X. (2007). Primary Research on Resources and Exploitation of Fagaceae in Guangxi. Guihaia 27 (2), 170–173. (In Chinese).
Castillo, C. C., Carter, A., Kingwell-Banham, E., Zhuang, Y., Weisskopf, A., Chhay, R., et al. (2020). The Khmer Did Not Live by Rice Alone: Archaeobotanical Investigations at Angkor Wat and Ta Prohm. Archaeol. Res. Asia 24, 100213. doi:10.1016/j.ara.2020.100213
Castillo, C. C., Fuller, D. Q., Piper, P. J., Bellwood, P., and Oxenham, M. (2018). Hunter-gatherer Specialization in the Late Neolithic of Southern Vietnam - the Case of Rach Nui. Quat. Int. 489, 63–79. doi:10.1016/j.quaint.2016.11.034
Chen, W.-c. (2007). The Late Neolithic Adaptation of the Northeastern Coast of Vietnam. Field Archaeology of Taiwan 11, 1–26. (In Chinese).
Chen, Y., Tao, Y., Lian, X., Wang, L., Zhao, Y., Jiang, J., et al. (2010). Chemical Constituents of Angiopteris esculenta Including Two New Natural Lactones. Food Chem. 122 (4), 1173–1175. doi:10.1016/j.foodchem.2010.03.111
Colani, M. (1938). Découvertes Préhistoriques dans les parages de la Baie d’Along. Hanoi: Dans Institut Indochinois pour l’Etude de l’Homme.
De Winter, W., and Amoroso, V. (2003). Plant Resources of South-East Asia No 15 (2) Cryptogams: Ferns and Fern Allies. Leiden: Backhuys Publishers.
Deng, Z., Huang, B., Zhang, Q., and Zhang, M. (2022). First Farmers in the South China Coast: New Evidence from the Gancaoling Site of Guangdong Province. Front. Earth Sci. 10, 858492. doi:10.3389/feart.2022.858492
Deng, Z., Hung, H.-c., Fan, X., Huang, Y., and Lu, H. (2017). The Ancient Dispersal of Millets in Southern China: New Archaeological Evidence. Holocene 28 (1), 34–43. doi:10.1177/0959683617714603
Deng, Z., Hung, H.-c., Li, Z., Carson, M. T., Huang, Q., Huang, Y., et al. (2019). Food and Ritual Resources in Hunter-Gatherer Societies: Canarium Nuts in Southern China and beyond. Antiquity 93 (372), 1460–1478. doi:10.15184/aqy.2019.173
Denham, T. P., Haberle, S. G., Lentfer, C., Fullagar, R., Field, J., Therin, M., et al. (2003). Origins of Agriculture at Kuk Swamp in the Highlands of New Guinea. Science 301 (5630), 189–193. doi:10.1126/science.1085255
Denham, T., Zhang, Y., and Barron, A. (2018). Is There a Centre of Early Agriculture and Plant Domestication in Southern China? Antiquity 92 (365), 1165–1179. doi:10.15184/aqy.2018.156
Dolai, N., Karmakar, I., Kumar, R. B., Bala, A., Mazumder, U. K., and Haldar, P. K. (2012). Antitumor Potential of Castanopsis Indica (Roxb. Ex Lindl.) A. DC. Leaf Extract against Ehrlich's Ascites Carcinoma Cell. Indian J. Exp. Biol. 50, 359–365.
Dutta, B. (2015). Food and Medicinal Values of Certain Species of Dioscorea with Special Reference to Assam. J. Pharmacogn. Phytochemistry 3 (5), 15–18.
Fullagar, R., Field, J., Denham, T., and Lentfer, C. (2006). Early and Mid Holocene Tool-Use and Processing of Taro (Colocasia Esculenta), Yam (Dioscorea sp.) and Other Plants at Kuk Swamp in the Highlands of Papua New Guinea. J. Archaeol. Sci. 33 (5), 595–614. doi:10.1016/j.jas.2005.07.020
Golson, J., Denham, T., Hughes, P., Swadling, P., and Muke, J. (2017). Ten Thousand Years of Cultivation at Kuk Swamp in the Highlands of Papua New Guinea. Canberra: ANU Press.
Golson, J. (2020). “The Origins and Development of New Guinea Agriculture,” in The Emergence of Agriculture. Editors D. R. Harris, and G. C. Hillman. 1st ed (London: Unwin Hyman), 175–186. doi:10.4324/9781003060765-9
Hang, Y., Xu, L., Shi, D., Qin, H., and Zhou, Y. (2006). Morphological Features and its Taxonomic Significance of Starch Grain in Subterranean Organ of Dioscorea L. In China. J. Plant Resour. Environ. 15 (4), 1.
Haryoso, A., Amzu, E., Hikmat, A., Sunkar, A., and Darusman, D. (2019). Ethnobotany of Sugar Palm (Arenga Pinnata) in the Sasak Community, Kekait Village, West Nusa Tenggara, Indonesia. Biodiversitas 21 (1). doi:10.13057/biodiv/d210116
Higham, C. (2013). Hunter-Gatherers in Southeast Asia: From Prehistory to the Present. Human Biol. 85 (1–3), 21–44. doi:10.3378/027.085.0302
Higham, C. (2014). Early Mainland Southeast Asia: From First Humans to Angkor. Bangkok: River Books Press.
Higham, C. F. W. (2021). “Chapter 2 The Neolithic Occupation of Southeast Asia,” in The Languages and Linguistics of Mainland Southeast Asia: A Comprehensive Guide, 21–32. 8. doi:10.1515/9783110558142-002
Huan, X., Lu, H., Wang, C., Tang, X., Zuo, X., Ge, Y., et al. (2015). Bulliform Phytolith Research in Wild and Domesticated Rice Paddy Soil in South China. PloS One 10 (10), e0141255. doi:10.1371/journal.pone.0141255
Hung, H.-c. (2019). Prosperity and Complexity without Farming: the South China Coast, c. 5000-3000 BC. Antiquity 93 (368), 325–341. doi:10.15184/aqy.2018.188
Hung, H-c., Zhang, C., Matsumura, H., Li, Z., and Shinoda, K. (2017). “Neolithic Transition in Guangxi: A Long Development of Hunting-gathering Society in Southern China,” in Bio-Anthropological studies of Early Holocene Hunter-gatherer Sites at Huiyaotian and Liyupo in Guangxi, China. Editors H. Matsumura, H-c. Hung, Z. Li, and K. Shinoda (Tokyo: National Museum of Nature and Science), 205–228.
Institute of Archaeology, Chinese Academy of Social Sciences (IA CASS) (2003). “Archaeological Team of Guangxi Province, Guilin Zengpiyan Site Museum, and Archaeological Team of Guilin City,” in Guilin Zengpiyan (Beijing: Wenwu). (In Chinese).
Ishak, M. R., Sapuan, S. M., Leman, Z., Rahman, M. Z. A., Anwar, U. M. K., and Siregar, J. P. (2013). Sugar Palm (Arenga pinnata): Its Fibres, Polymers and Composites. Carbohydr. Polym. 91 (2), 699–710. doi:10.1016/j.carbpol.2012.07.073
Katharina, N., Ae, C., Terry, B., Maria, A. R., Luc, V., and Scott, C. L. (2019). International Code for Phytolith Nomenclature (ICPN) 2.0. Ann. Bot. 124 (2), 189–199. doi:10.1093/aob/mcz064
Laboratory of the Institute of Archaeology (CASS) (1977). Report of Radiocarbon Dates (4). Archaeology (3). 200–204.
Lentfer, C. J. (2009). “Building a Comparative Starch Reference Collection for Indonesia and its Application to Palaeoenvironmental and Archaeological Research,” in Archaeological Science Under a Microscope : Studies in Residue and Ancient DNA Analysis in Honour of Thomas H. Loy. Editors M. Haslam, G. Robertson, A. Crowther, S. Nugent, and L. Kirkwood. 1st ed (Canberra, Australia: ANU E Press), 80–101. doi:10.22459/ta30.07.2009.07
Li, Z. (2019). “Early Maritime Subsistence and Adaptive Ocean Cultures along the Beibu Gulf Coast,” in Prehistoric Maritime Cultures and Seafaring in East Asia. Editors C. M. Wu, and B. V. Rolett (Singapore: Springer Nature Singapore Pte Ltd), 127–139. doi:10.1007/978-981-32-9256-7_7
Li, Z. (2020). Plant-related Subsistence in the Pearl River Delta, Southern China, from 6,000 BP to 3,000 BP. PhD Thesis. Leicester: University of Leicester.
Liu, Y., Wujisguleng, W., and Long, C. (2012). Food Uses of Ferns in China: A Review. Acta Soc. Bot. Pol. 81 (4), 263–270. doi:10.5586/asbp.2012.046
Loy, T. H., Spriggs, M., and Wickler, S. (2015). Direct Evidence for Human Use of Plants 28,000 Years Ago: Starch Residues on Stone Artefacts from the Northern Solomon Islands. Antiquity 66 (253), 898–912. doi:10.1017/s0003598x00044811
Lu, H., Liu, Z., Wu, N., Berné, S., Saito, Y., Liu, B., et al. (2008). Rice Domestication and Climatic Change: Phytolith Evidence from East China. Boreas 31 (4), 378–385. doi:10.1111/j.1502-3885.2002.tb01081.x
Lu, P. (2010). Zooarchaeological Study on the Shell Middens in the Yong Valley of Guangxi. PhD Thesis. Beijing: Chinese Academy of Social Sciences. (In Chinese).
Lu, T. L.-D. (2003). “A Preliminary Analysis for the Residues on the Surface of Unearthed Stone Tools in Zengpiyan Site,” in Zhengpiyan Site of Guilin. Editors: Institute of Archaeology (Chinese Academy of Social Science), Antiquities Task Force in Guangxi Zhuang Autonomous Region, Museum of Zengpiyan site in Guilin, and Antiquities Task Force in Guilin. 1st ed (Beijing: Cultural Relics Press), 646–651.
Maneenoon, K., Sirirugsa, P., and Sridith, K. (2008). Ethnobotany of Dioscorea L. (Dioscoreaceae), a Major Food Plant of the Sakai Tribe at Banthad Range, Peninsular Thailand. Ethnobot. Res. App. 6, 385–394. doi:10.17348/era.6.0.385-394
Mannan, M. M., Maridass, M., and Victor, B. (2008). A Review on the Potential Uses of Ferns. Ethnobot. Leafl. 12, 281–285.
Mason, S. (1995). “Acornutopia? Determining the Role of Acorns in Past Human Subsistence,” in Food in Antiquity. Editors J. Wilkins, M. Harvey, and M. Dobson. 1st ed (Exeter: University of Exeter Press), 12–24.
Matsumura, H., Hung, H.-c., Higham, C., Zhang, C., Yamagata, M., Nguyen, L. C., et al. (2019). Craniometrics Reveal “Two Layers” of Prehistoric Human Dispersal in Eastern Eurasia. Sci. Rep. 9 (1), 1451. doi:10.1038/s41598-018-35426-z
Matthews, P. J. (1991). A Possible Tropical Wildtype Taro: Colocasia esculenta Var. Aquatilis. Indo-Pacific Prehistory Assoc. Bull. 11, 69–81. doi:10.7152/bippa.v11i0.11374
Matthews, P. J., Agoo, E. M. G., Tandang, D. N., and Madulid, D. A. (2012). “Ethnobotany and Ecology of Wild Taro (Colocasia esculenta) in the Philippines: Implications for Domestication and Dispersal,” in Senri Ethnological Studies. Editors M. Spriggs, D. Addison, and P. J. Matthews (Osaka: National Museum of Ethnology) 78, 307–340. doi:10.15021/00002523
Matthews, P. J. (2014). On the Trail of Taro: An Exploration of Natural and Cultural History. Osaka: National Museum of Ethnology.
McColl, H., Racimo, F., Vinner, L., Demeter, F., Gakuhari, T., Moreno-Mayar, J. V., et al. (2018). The Prehistoric Peopling of Southeast Asia. Science 361 (6397), 88–92. doi:10.1126/science.aat3628
Mijares, A. S. B. (2007). Unearthing Prehistory: The Archaeology of Northeastern Luzon, Philippine Islands. Oxford: British Archaeological Reports.
Nguyen, D. T., and Clarkson, C. (2013). The Organisation of Drill Production at a Neolithic Lithic Workshop Site of Bai Ben, Cat Ba Island, Vietnam. Bull. Indo-Pacific Prehistory Assoc. 33, 24–39. doi:10.7152/bippa.v33i0.14508
Nguyen, H. T. M. (2013). Neolithic Vegetation in Northern Vietnam: an Indication of Early Agricultural Activities. J. Austronesian Stud. 4, 1.
Nguyen, K. D. (2019). “The Late Neolithic to Early Bronze Age on the Northeastern Coast of Vietnam,” in Prehistoric Maritime Cultures and Seafaring in East Asia. Editors C. M. Wu, and B. V. Rolett (Singapore: Springer Nature Singapore Pte Ltd), 141–157. doi:10.1007/978-981-32-9256-7_8
Nguyen, K. S. (2009). Prehistorical Site Cai Beo on Cat Ba Island. Hanoi: Social Sciences. (In Vietnamese).
Nguyen, Q. M., and Tran, T. H. (2009). Geoarchaeology of The Maritime Region in Northeast Vietnam. Bull. Indo-Pacific Prehistory Assoc. 29, 56–60. doi:10.7152/bippa.v29i0.9478
Nguyen, T. M. H. (2014). Holocene Vegetation and Human Activities in Northern Vietnam. PhD Thesis. Tokyo: University of Tokyo.
Nguyen, T. M. H. (2017). Burnt Rice from Four Archaeological Sites in Northern Vietnam. Vietnam Soc. Sci. 179, 64–77.
Nguyen, V. (2005). The Da but Culture: Evidence for Cultural Development in Vietnam during the Middle Holocene. Bull. Indo-Pacific Prehistory Assoc. 25, 89–94. doi:10.7152/bippa.v25i0.11918
Nguyen, V. (2008). Hoabinhian Macrobotanical Remains from Archaeological Sites in Vietnam: Indicators of Climate Changes from the Late Pleistocene to the Early Holocene. Bull. Indo-Pacific Prehistory Assoc. 28, 80–83. doi:10.7152/bippa.v28i0.12019
Nishimura, M. (2006). “Chronological Framework from the Palaeolithic to Iron Age in the Red River Plain and the Surrounding,” in Prehistoric Archaeology of South China and Southeast Asia. Editor Institute of Archaeology, Chinese Academy of Social Sciences (Beijing: Cultural Relics Publishing House), 347–373.
Nopporncharoenkul, N., and Jenjittikul, T. (2017). Kaempferia Noctiflora (Zingiberaceae), a New Species from Northern Thailand. Phytotaxa 316 (1), 67. doi:10.11646/phytotaxa.316.1.6
Oliveira, N. V. (2008). Subsistence Archaeobotany: Food Production and the Agricultural Transition in East Timor. PhD Thesis. Canberra: The Australian National University.
Oliveira, N. V. (2012). Recovering, Analysing and Identifying Colocasia Esculenta and Dioscorea spp. From Archaeological Contexts in Timor-Leste. Senri Ethnol. Stud. 78, 265–284. doi:10.15021/00002521
Opara, L. U. (2003). Edible Aroids: Post Harvest Operations. Rome: Food and Agriculture Organization of the United Nations.
Paz, V. J. (2001). Archaeobotany and Cultural Transformation : Patterns of Early Plant Utilisation in Northern Wallacea. J. Acoust. Soc. Am. 106 (4), 1538–1544. doi:10.17863/CAM.15977
Peng, C. L. (2018). The Research of Early Archaeological Cultures in Vietnam. Nanning: Guangxi Science & Technology Publishing House. (In Chinese).
Pyramarn, K. (1989). “New Evidence on Plant Exploitation and Environment during the Hoabinhian (Later Stone Age) from Ban Kao Caves, Thailand,” in Foraging and Farming: The Evolution of Plant Exploitation. Editors D. R. Harris, and G. C. Hillman. 1st ed (London: Routledge), 282–291.
Rangabhashiyam, S., and Selvaraju, N. (2015). Evaluation of the Biosorption Potential of a Novel Caryota Urens Inflorescence Waste Biomass for the Removal of Hexavalent Chromium from Aqueous Solutions. J. Taiwan Inst. Chem. Eng. 47, 59–70. doi:10.1016/j.jtice.2014.09.034
Saxena, A., Prasad, V., Singh, I. B., Chauhan, M. S., and Hasan, R. (2006). On the Holocene Record of Phytoliths of Wild and Cultivated Rice from Ganga Plain: Evidence for Rice-Based Agriculture. Curr. Sci. 90, 1547–1552.
Sharma, L. N., and Bastakoti, R. (2010). Ethnobotany of Dioscorea L. With Emphasis on Food Value in Chepang Communities in Dhading District, Central Nepal. Bot. Orient. 6, 12–17. doi:10.3126/botor.v6i0.2905
Spriggs, M., and Matthews, P. J. (2012). “Irrigated Taro in the Indo-Pacific: Multiple Perspectives,” in Senri Ethnological Studies. Editors M. Spriggs, D. Addison, and P. J. Matthews (Osaka: National Museum of Ethnology), 341–347.
Summerhayes, G. R., Leavesley, M., Fairbairn, A., Mandui, H., Field, J., Ford, A., et al. (2010). Human Adaptation and Plant Use in Highland New Guinea 49,000 to 44,000 Years Ago. Science 330 (6000), 78–81. doi:10.1126/science.1193130
Thin, N. N. (1998). The Vegetation of the Catba National Park in Vietnam. Korean J. Ecol. 21 (2), 139–149.
Wan Omar, W. B., Zakeri, H. A., Salim, J. M., Fisall, U. F. M., and Amran, M. A. (2019). “Genetic Diversity of Seed Storage Protein in Selected Melastomataceae and Fagaceae from Tasik Kenyir,” in Greater Kenyir Landscapes. Editor M. T. Abdullah, A. Mohammad, M. N. Mohammad, and M. S. Lola (Cham: Springer), 83–90. doi:10.1007/978-3-319-92264-5_8
Wang, T., Xiao, B., Liu, E.-D., Nguyen, K. S., Duan, J.-Q., Wang, K.-L., et al. (2020). Rediscovery of Angiopteris Tonkinensis (Marattiaceae) after 100 years, and its Revision. PhytoKeys 161, 1–9. doi:10.3897/phytokeys.161.54912
Wang, Y., and Lu, H. (1993). Research on Phytolith and its Application. Beijing: Ocean Press. (In Chinese).
Weber, S., Lehman, H., Barela, T., Hawks, S., and Harriman, D. (2010). Rice or Millets: Early Farming Strategies in Prehistoric Central Thailand. Archaeol. Anthropol. Sci. 2 (2), 79–88. doi:10.1007/s12520-010-0030-3
Wimalasiri, G. E. M., Ranasinghe, P., Gunaratne, D. M. A., and Arachchi, L. P. V. (2016). Antioxidant and Anti-diabetic Properties of Caryota urens (Kithul) Flour. Procedia Food Sci. 6, 181–185. doi:10.1016/j.profoo.2016.02.044
Yang, X., Barton, H. J., Wan, Z., Li, Q., Ma, Z., Li, M., et al. (2013). Sago-type Palms Were an Important Plant Food Prior to Rice in Southern Subtropical China. PLoS One 8 (5), e63148. doi:10.1371/journal.pone.0063148
Yang, X., Wang, W., Zhuang, Y., Li, Z., Ma, Z., Ma, Y., et al. (2017). New Radiocarbon Evidence on Early Rice Consumption and Farming in South China. Holocene 27 (7), 1045–1051. doi:10.1177/0959683616678465
Yang, X., Wu, W., Perry, L., Ma, Z., Bar-Yosef, O., Cohen, D. J., et al. (2018). Critical Role of Climate Change in Plant Selection and Millet Domestication in North China. Sci. Rep. 8 (1), 7855. doi:10.1038/s41598-018-26218-6
Yen, D. E. (1977). “Hoabinhian Horticulture? the Evidence and the Questions from Northwest Thailand,” in Sunda Sahul: Prehistoric Studies in Southeast Asia, Melanesia Australia. Editor J. Allen, J. Golson, and R. Jones (London: Academic Press), 567–599.
Yoshino, H. (2002). Morphological and Genetic Variation in Cultivated and Wild Taro. Vegeculture East. Asia Ocean. 16, 95–116.
Zhang, C., and Hung, H.-c. (2010). The Emergence of Agriculture in Southern China. Antiquity 84 (323), 11–25. doi:10.1017/s0003598x00099737
Zhang, C., and Hung, H.-c. (2012). Later Hunter-Gatherers in Southern China, 18 000–3000 BC. Antiquity 86 (331), 11–29. doi:10.1017/s0003598x00062438
Keywords: Hunter-gatherers, starch, phytolith, Vietnam, Southeast Asia, rice, Ha Long Bay
Citation: Wang W, Nguyen KD, Dang Le H, Zhao C, Carson MT, Yang X and Hung H-c (2022) Before Rice and the First Rice: Archaeobotanical Study in Ha Long Bay, Northern Vietnam. Front. Earth Sci. 10:881104. doi: 10.3389/feart.2022.881104
Received: 22 February 2022; Accepted: 02 June 2022;
Published: 30 June 2022.
Edited by:
Steven L. Forman, Baylor University, United StatesReviewed by:
Xueqin Zhao, Shanghai Ocean University, ChinaRajesh Agnihotri, Birbal Sahni Institute of Palaeobotany, India
Copyright © 2022 Wang, Nguyen, Dang Le, Zhao, Carson, Yang and Hung. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hsiao-chun Hung, SHNpYW8tY2h1bi5IdW5nQGFudS5lZHUuYXU=