 Cleber Q. C. Diniz
Cleber Q. C. Diniz Juliana de Moraes Leme
Juliana de Moraes Leme Paulo C. Boggiani
Paulo C. Boggiani- 1Institute of Geosciences, University of São Paulo, São Paulo, Brazil
- 2Department of Sedimentary and Environmental Geology, Institute of Geosciences, University of São Paulo, São Paulo, Brazil
The Tamengo Formation (Corumbá Group) is an important Ediacaran stratigraphic unit in South America due to the presence of metazoan fossils and geochemistry data of carbonate rocks, with excellent geochronological delimitation (between 555–541 Ma) obtained by U–Pb dating on volcanic zircons. The present work shows three new species of macroalgae found as carbonaceous compressions and studied for their morphology and taxonomy. All new taxa are characterized as centric macroalgae; Tamengophyton espinosa sp. nov. is a fan-shaped alga with striated thalli, dichotomous branches, trichomes with perpendicular growth, and a connecting membrane. Ladariella hidria sp. nov. is formed by a set of striated and branched thalli in a cylindrical form with almond-shaped structures in the top. Ladariophyton veinosa sp. nov. is characterized by the main growth thallus and an enlarged longitudinal structure at the center. These new occurrences of macroalgae add to the largest life assemblages in the Neoproterozoic of South America, which contributes to documentation of the evolutionary history of macroalgae and the paleoecological settings of the Late Ediacaran.
Introduction
The evolution of eukaryotic organisms changed the dynamic of the Earth. Even though the timing of their appearance is debated, through the paleontological data and molecular clock analysis, we have an estimative age for the last eukaryotic ancestor appearance between 1.8 and 2.3 Ga in the Paleoproterozoic Era (Hedges et al., 2004; Knoll et al., 2006; Parfrey et al., 2011). These evolutions brought new cellular behaviors, which were capable of sensing and reacting to environmental change (Cohen and Mcdonald, 2015; Wan and Jekély, 2021). Reaching a higher level of multicellular organization, they could expand and differentiate the cells to adapt to new opportunities.
During the Ediacaran, such new life forms were mainly represented by macroalgae (Bykova et al., 2020). They still are defined as eukaryotic multicellular organisms capable of doing photosynthesis, megascopic with a size larger than 1 mm, and possible to see with the naked eye, whose oldest fossil record dates to the Neoproterozoic (Xiao and Dong, 2006; Tang et al., 2020). Their complexity and diversity have records in China (Xiao et al., 1998), India (Srivastava, 2012), the United States (Rowland and Rodriquez, 2014), Russia (Nagovitsin et al., 2015), Mongolia (Dornbos et al., 2016), and Canada (Maloney et al., 2021). Interacting with other organisms of their time, the macroalgae form biotas, which serve as protection sites or creating oxygen niches (Xiao et al., 2002; Zhao et al., 2004). Here, we present three new species of macroalgae from Mato Grosso do Sul Brazil and discuss their significance to the Ediacaran assemblage from Tamengo Formation and the influence of the depositional process from the Corumbá Group in their preservation.
Geological Context
The Corumbá Group exposes in Serra da Bodoquena and Corumbá region (Figure 1A) (Mato Grosso do Sul State, SW Brazil), close at Urucum Massif. The fossiliferous sites are located in the cities of Corumbá, Ladário, in mines and at the margin of the Paraguay River, in the Pantanal plain. The lowest units in the Corumbá Group (Figure 1B) are the Cadieus and Cerradinho formations representing a terrigenous sedimentation. Follow-up by the Bocaina Formation containing stromatolitic dolomites and a volcanic ash at the top dating 555.18 ± 0.30 Ma. The Tamengo Formation is the best-known unit from the Corumbá Group, around 200 m thick, characterized by black limestones, grainstones, organic siltstones, where the macroalgae occur, and shales. Follow-up by the the Guaicurus Formation containing laminated siltstones and macroalgae named Eoholynia corumbensis-up (Boggiani, 1998; Boggiani et al., 2010; Parry et al., 2017).
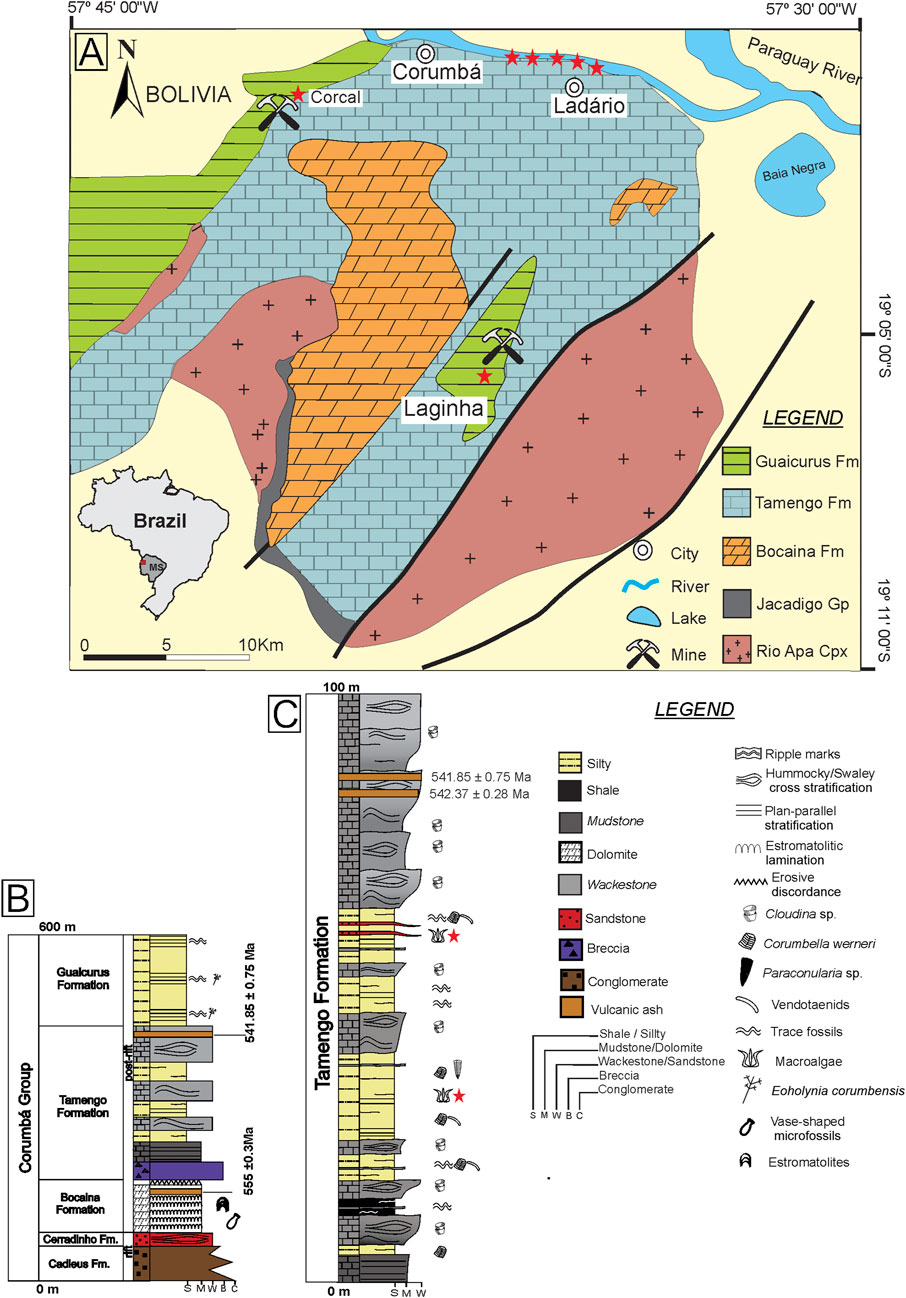
FIGURE 1. Geological setting of the study area (modified from Amorim et al., 2020). (A) Stars represent the sites where the outcrop is exposed and studied. (B) Stratigraphic column of the Corumbá Group (modified from Parry et al., 2017). (C) Condensed section of the Tamengo Formation, in Ladário, Porto Sobramil, and Corcal Mine.
The Tamengo Formation (Figure 1C) is marked at the base by carbonatic breccia, overlain by carbonate facies of grainstones and mudstones with sedimentary structures, such as hummocky cross-stratification and wave marks with a volcanic ash at the top, in which zircons are dated at 541.85 ± 0.75 Ma. Packages of shales, siltstones, and fine sandstones with parallel plane lamination are interspersed with the carbonate facies (Boggiani et al., 2010; Parry et al., 2017; Amorim et al., 2020).
The Ediacaran Tamengo Formation exhibits paleontological remains with metazoan fossils such as Corumbella werneri (Pacheco et al., 2015; Walde et al., 2019), Cloudina sp. (Adorno et al., 2017; Becker-Kerber et al., 2017), Paraconularia sp. (Van Iten et al., 2014), and trace fossils such as Multina minima (Parry et al., 2017). Based on geological and paleontological evidence, the Tamengo Formation represents a carbonatic ramp in a marine environment, between the fair-weather wave base level and storm wave base level during a transgression phase. The facies characterized by siliciclastic rocks, metazoans, trace fossils, and plane parallel structures suggest an environment of low energy, associated with the offshore zone. The facies with limestone, hummocky cross-stratification, wave ripples, and Cloudina sp., is associated with the shoreface zone (Boggiani et al., 2010; Oliveira et al., 2019; Amorim et al., 2020).
Materials and Methods
The fossils described here were collected in the siltstones of Tamengo Formation near Ladário city from Mato Grosso do Sul state. Each species has one specimen deposited in the Geoscience Institution, University of São Paulo. The samples were analyzed and photographed under a Zeiss stereomicroscope, and the fossil measurement data from each specimen were made with digital images using software Zen™ (ZEISS), where we catalog and measure the entire dimension containing their morphological features.
The samples were subjected to analysis in a scanning electron microscopy (SEM) to observe main morphological features in high resolution and energy dispersive spectroscopy (EDS) to make elemental mapping, allowing for the characterization of the chemical composition of the macroalgae.
Systematic Paleontology
Usually, the morphological features of the fossils are fundamental to the taxonomy, since other characteristics such as biochemistry or the cellular structure is more difficult and rarely. The use of morphological characters to propose a new taxon needs to be approached with caution because the convergent evolution process does not make it a conclusive tool to identify phylogenetic affinity (Xiao and Dong, 2006). Also, the morphological features can be altered or lost through taphonomic processes, which strongly affects fossil algae. Their soft bodies are often recorded as broken, flattened, or distorted, and their most fragile structures can be dissolved and lost. But despite taphonomic processes, information can also be interpreted based on the fossil record.
The systematics of the fossils presented here is based only on their morphology, taking into account the taphonomic bias. The characteristics and description of the structures are based on Xiao et al. (2002) and Ye et al. (2019) who proposed the distinction of each genus considering the general morphology, branching pattern, the width of the thallus, the change in width along the length, the internode length flexibility, and the presence of microstructures. The phylogenetic affinities of the macroalgae are still uncertain because it needs a cellular information that the fossil record cannot provide. The systematic description is based on the genus and species levels. The descriptions of the new taxa follow the international code for plant, fungi, and algae nomenclature (IAPT global).
Genus Tamengophyton C.Q.C. Diniz & J.M. Leme sp. nov.
Type species: Tamengophyton espinosa C.Q.C. Diniz & J.M. Leme sp. nov.
Material: One specimen preserved.
Diagnosis: As for type species, by monotype.
Etymology: Tamengo Formation, where it was found.
Type locality: Ladário city located in the west of Mato Grosso do Sul state, Brazil.
Type horizon and stratigraphy: Ediacaran, Tamengo Formation, Brazil.
Tamengophyton espinosa C.Q.C. Diniz & J.M. Leme sp. nov.
Holotype: GP/1E - 10,990 (Figure 2), from the Tamengo Formation, Ediacaran, Corumbá Group, Ladário, Brazil (Repository: IGc-USP).
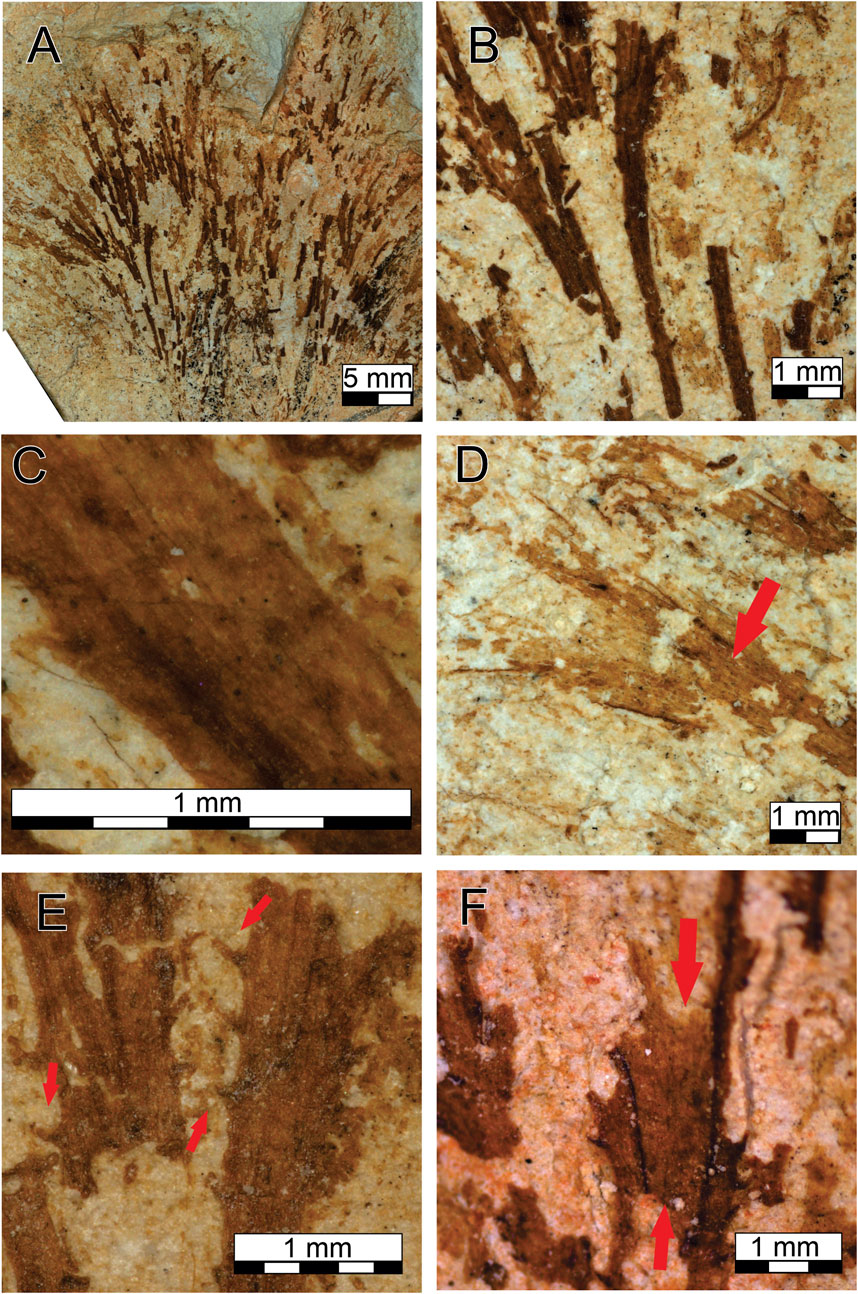
FIGURE 2. Tamengophyton espinosa. (A) Complete fossil. (B) Widening in width through the length of thalli. (C) Details of the striation. (D) branching (arrow). (E) Details of the trichomes (arrows). (F) Details of the membrane (between the arrows).
Diagnosis: A centimeter-scale fan-shaped macroalgal fossil. Possess thalli with heterogeneous thickness (Figure 2B), striated, and with dichotomous ramifications (Figures 2C,D). Along the thalli, microstructures projects in the edges (Figure 2E). In the apical part, a membrane is preserved that connects the stems (Figure 2F).
Description: The sample preserved in two-dimensional carbonaceous compression, with a size of 2.9 cm and a minimum width of 0.5 cm at base and a maximum width of 3.5 cm, formed by a set of approximately 27 striated thalli that branch per segment, widening at the top. The thalli do not overlap each other and are arranged in a straight pattern. The thickness of each thallus varies from 0.2 mm to 0.8 mm along the fossil species, with the base thinner than the top; this heterogeneity is repeated along the entire fossil. Parts preceding the branches tend to be thicker, appearing from the base through the top but can be observed only once per thallus. The preserved striation pattern follows the same longitudinal direction of the thalli. Microstructures are projecting throughout the thalli; there is no pattern for their positioning, and some are quadrilateral grouped, while others are arranged horizontally or vertically. They are preserved in different sizes, their length varies from 0.1 to 0.25 mm, and their width varies from 0.06 to 0.09 mm. The membrane is present in the distal part of the fossil, where a connection between the thalli is visible, and has the same composition as the thallus but is not striated.
Etymology: The fossil species name is determined by the presence of the microstructures that resemble thorns.
Remarks: Tamengophyton gen. nov. is characterized by the overall fan shape caused by the spreading of the stalks due the branching at the top; the straight thallus indicates little flexibility, the micro-triangle–shaped structures that grow perpendicularly throughout the thalli here is known as trichomes. No cellular structure has been preserved, but the presence of longitudinal striations implies a more complex internal structure resembling a pseudoparenchyma.
T. espinosa is similar to other species of macroalgae from Ediacaran around the world. The phylogeny of higher grades is uncertain, but the specimen fits into the family of Eoholyneacea (Hoffman 1994) and could be attributed to macroalgae of the Neoproterozoic due to the presence of branching. In addition, similar morphological features such as compressed cylindrical bifurcated thalli and a width < than 2 mm can also be observed with the morphogroup denominated delicately dichotomously branched, as proposed by LoDuca et al. (2017) for specimens of early Paleozoic seaweeds.
T. espinosa can be compared with genera of Doushantuophyton (Steiner, 1994), Konglingiphyton (Chen and Xiao, 1992) emend. Xiao et al. (2002) and Maxiphyton Ye et al. (2019) because it presents similar characteristics such as straight and rigid branches, regular dichotomy, and the capacity to widen the thallus toward the distal part. But their size does not combine; Doushantuophyton is 0.04–0.2 mm wide per thallus and Konglingiphyton can reach 1.4 mm, while the size of T. espinosa stays between the two genera. However, the principal diagnosis for distinguishing them is that the other taxa have smooth branches, while T. espinosa possesses the trichome projections and the connecting membrane to assign a new genus and new species.
Genus Ladariella C.Q.C. Diniz & J.M. Leme gen. nov.
Type species: Ladariella hidria C.Q.C. Diniz & J.M. Leme sp. nov.
Material: One specimen preserved.
Diagnosis: As for type species, by monotype.
Etymology: After the city of Ladário, located close to the type section where the fossil genus was found.
Type locality: Ladário city located in the west of Mato Grosso do Sul state, Brazil.
Type horizon and stratigraphy: Ediacaran, Tamengo Formation.
Ladariella hidria C.Q.C. Diniz & J.M. Leme sp. nov.
Holotype: GP/1E - 10,989 (Figure 3), from the Tamengo Formation, Ediacaran, Corumbá Group, Ladário, Brazil (Repository: IGc-USP).

FIGURE 3. Ladariella hidria. (A,B) Complete fossil with the arrow pointed out to the rhizoidal holdfasts in (A). (C) Details of the rhizoidal holdfasts. (D) Branching (arrow). (E) Details of the striation in the thalli. (F,G) Details of the almond structure (arrows).
Diagnosis: Centimeter-sized fossil preserved as a set of irregular curved thalli (Figures 3A,B). Presents rhizoidal holdfasts (Figure 3C) with a central axis and secondary elements at the base of the specimens. Possesses dichotomous striated branches (Figures 3D,E) and almond-shaped structures formed at the top (Figures 3F,G).
Description: The sample consists of a two-dimensional carbonaceous compression. With an elongate rhizoidal holdfast structure that grows perpendicular from the base of the thallus with 0.13 to 0.20 mm in width. It has a set of 20 thalli, with 1.5 cm maximum length. Each thallus is arranged obliquely and overlap each other from varied angles and difficult to measure their length, but the overall size varies from 5 to 15 mm. The width also varies between 0.15 and 0.9 mm, getting thinner in the top. The branching occurs once per thallus, and the striation follows the same longitudinal direction. The almond-shaped structures at the end ranges in size from 0.08 to 0.2 mm and are positioned obliquely with no apparent direction.
Etymology: Reference to the Greek mythological creature named hydra based on the similarity of the structure of the thalli arranged as one set and the multiple heads of the hydra in mythology.
Remarks: In the basal part, one structure at each side grows a central axis perpendicular to the thallus, most likely attributed to rhizoidal holdfasts. The branching of the thallus only occurs in the basal part of the fossil so the thickness in that area can be caused due to the enlargement of the branching process. The curvature state of the thalli indicates a flexibility that could help in a better response to hydrodynamics, and the almond-shaped structures has more accumulation of organic matter.
The phylogeny of higher grades remains uncertain, and the specimen fits into the Eoholyneacea family proposed by Hoffman (1994) because of the presence of branching and also shares the same features of T. spinosa, such as compressed cylindrical bifurcated thalli and a width < 2 mm that fits in the morphogroup proposed by LoDuca et al. (2017) based on delicately dichotomously branches.
L. hidria and Miaohephyton (Xiao et al. 1998) share a characteristic of a possible apical meristem. Nevertheless, considering the entire morphology, branching pattern, the heterogeneity, and especially the flexibility of the thallus as well as the almond structures, a new genus needs to be established.
Genus Ladariophyton C.Q.C. Diniz & J.M. Leme gen. nov.
Type species: Ladariophyton veinosa C.Q.C. Diniz & J.M. Leme sp. nov
Material: One specimen preserved.
Diagnosis: As for type species, by monotype.
Etymology: Named after the city of Ladário, in where it was found.
Type locality: Ladário city located in the west of Mato Grosso do Sul state, Brazil.
Type horizon and stratigraphy: Ediacaran, Tamengo Formation, Brazil.
Ladariophyton veinosa C.Q.C. Diniz & J.M. Leme sp. nov.
Holotype: GP/1E - 11,187 (Figure 4), from the Tamengo Formation, Ediacaran, Corumbá Group, Ladário, Brazil (Repository: IGc-USP).

FIGURE 4. Ladariophyton veinosa. (A,B) The complete fossil. (C) Details of the branching. (D) Details of the edge. (E,F) Details of the thickness of the longitudinal structure.
Diagnosis: Fossil organism of centimeter-scale with pseudomonopodial irregular branches (Figure 4B). It presents a defined border and has an enlarged longitudinal structure on the center of each thallus (Figure 4D–F).
Description: The sample has a set of thalli, which are arranged in a specific direction without overlapping each other. They vary in length, with the longer thallus being 1.1 cm. This does not represent the full length of the organism because none of the thallus is complete without a basal and an apical part. The thalli are homogeneous in width, measuring between 0.2 and 0.35 mm. Branching can be observed more than once, but one thallus represents the principal growth of the alga where the branching always occurs. The fossil has well-demarcated edges with a corrugated shape. The characteristic, which differentiates and defines this fossil, is a thickening in the longitudinal structure present at the center of the thalli of approximately 0.1 mm.
Etymology: Reference for veins, based on the central structure along the thalli.
Remarks: This specimen fits into the Eoholyniaceae family proposed by Hoffman (1994) for macroalgae of the Neoproterozoic because of the presence of branches and the presence of characteristics of the morphogroup of delicately dichotomous branches as proposed by LoDuca et. al. (2017) for specimens of early Paleozoic seaweeds.
L. veinosa sp. nov. is a very simple macroalga difficult to attribute a major group, can be compared to Chinggiskhaania bifurcata (Dornbos et al. 2016) due to its a thin filament with an average width of 0.45 mm, does not densely assemble and without distal tapering in width, but lacks a longitudinal striation. Macroalgae from Dolores Creeks Formation, Canada (Maloney et al., 2021), presents a longitudinal division but have septa along the fossil, structure that our specimen does not have; the aspects differentiating L. veinosa are the corrugated edges and the thick longitudinal structure in the center of the thallus.
Discussion
The different records of Ediacaran fossils have various modes of exceptional preservation, such as phosphatization (e.g., Xiao et al., 2004), permineralization (e.g., Zhang et al., 1998), pyritization (e.g., Cai et al., 2010), and silicification (e.g., Liu, 2016). Analysis using energy-dispersive X-ray spectroscopy (EDS) of two samples was performed (Figures 5, 6), which indicated a high carbon and a low aluminum and silica content in the fossil in relation to the matrix.

FIGURE 5. Analysis of SEM and EDS of T. espinosa. (A) The complete fossil, where the red square highlights the zoom made in image (B). (B) Analysis of SEM. (C) Concentration of carbon. (D) Concentration of calcium. (E) Concentration of silica. (F) Concentration of aluminum.

FIGURE 6. Analysis of SEM and EDS of L. hidria. (A) The complete fossil, where the red square highlights the zoom made in image (B). (B) Analysis of SEM, where the red rectangle highlights the zoom made in the other images. (C) Concentration of carbon. (D) Concentration of calcium. (E) Concentration of silica. (F) Concentration of aluminum.
The macroalgal fossils are preserved as carbonaceous compressions in pelite, from the information provided by SEM and EDS, which can be compared to Cambrian Burgess Shale–type preservation (Butterfield, 1995; Xiao et al., 2002; Butterfield, 2005). In addition, the preservation occurred because of a transformation of the non-mineralized soft tissues (organic matter) to a stable kerogen is a process named kerogenization (Cai et al., 2012).
Amorim et al. (2020) propose a facies association in the deposits from the Tamengo Fm. where the macroalgae were found. The silty shale facies have a parallel lamination with wide paleodiversity, such as fossils of Corumbella werneri, Paraconularia sp., and vendotaenids. It is intercalated with carbonated facies such as massive to poorly laminated calcimudstone, laminated wackestone, and hummocky/swaley cross-stratified packstone. The alternation between fossiliferous pelites with tempestites (carbonates) suggested the preservation of the fragile macroalgae during the interval of the storms, during the suspension fallout of fine sediments (Labaj and Pratt, 2016; Amorim et al., 2020).
The absence of hard structures in the macroalgae makes these fossils difficult to be preserved, especially their delicate characteristics as presented in the two samples (T. espinosa and L. hidria). The fossil T. espinosa have a basal part incomplete, where the distal part is more preserved, which has been difficult to measure the full length of individual thallus. The absence of structures such as trichomes and membranes at the base of the fossil could be explained by an influence of taphonomic processes shaping the preserved morphology of the algae. The flattening of the fossil is visible, creating a two-dimensional form, affecting the trichomes forming small circles, and causing rupture in the thalli.
In Ladariella hidria, taphonomic processes influenced the preservation and shape of the fossil organism, including flattening throughout the structure. Better preservation occurs in the almond structures, where a thicker and darker layer was reported due to a larger accumulation of organic matter. This might indicate an apical meristem, where greater cell division for algae growth is expected.
In the case of Ladariophyton veinosa, a connection between the thalli is missing, together to an indication of a basal and top parts. Therefore, it is hard to acknowledge the total length of the fossil. The corrugated shape of the edges could also have been formed by a taphonomic influence; when body parts start to lose humidity, they tend to wrinkle.
Even though the macroalgae are not preserved in life position, a fossil of higher fragility could not be transported over long distances or remain in the active high-energy zone for a long period of time without significant destruction. So, the presence of the macroalgae could indicate a depth when they would receive enough sunlight to enable photosynthesis and with energy sufficient to partially destroy their body.
Other fossils of metazoan are preserved in the same stratigraphic level such as Corumbella werneri; they also miss the basal part but present an overall well-preserved level in their carapace (Amorim et al., 2020). Corumbella werneri possess a thin organic and delicate wall (Pacheco et al., 2015; Walde et al., 2019), so the well-preserved fossil associated with the silty shale facies can indicate a minimal transportation or rework at a low-energy environment (Warren et al., 2012; Amorim et al., 2020).
The preservation of macroalgae together with carapaces of metazoan in the siltstone bed in the Tamengo Formation can be explained by environmental variations in higher sedimentation rates, probably a sedimentary deposit under offshore condition in a depth below storm wave base, with a low-energy environment representing an outer to distal mid-ramp.
Conclusion
Three specimens of macroalgae are described for the Brazilian Ediacaran, Tamengo Formation in Mato Grosso do Sul, representing new taxonomic species and genera. The new descriptions contribute to the local paleobiodiversity along with metazoans, ichnofossils, and microfossils previously documented in this formation. This further reinforces the contribution of Brazil to the knowledge of the Ediacaran fossils. These three genera and species add new evidence to evolutionary issues of macroalgae during the Ediacaran, especially T. espinosa with its more complex features such as differentiated trichome and an interconnecting membrane. This also emphasizes more complex organisms already existed before the Cambrian but remain extremely rare in the fossil record due to their fragile structures.
Data Availability Statement
The original contributions presented in the study are included in the article/supplementary materials; further inquiries can be directed to the corresponding author.
Author Contributions
CD, JL, and PB collected the fossils. CD performed laboratory work and morphological taxon identifications. JL assisted with the interpretation. PB collaborated with hypothesis in the discussion. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – CAPES (167787/2017-9) and Fundanção de Amparo à Pesquisa – FAPESP (Proc. 13/17835-8; 16/06114-6; 17/25433-8; 17/25433-8).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank you the engineer Isaac Jamil Sayeg for providing SEM and EDS data. We are grateful to Andrea K. Kern from the Department of Sedimentary and Environmental Geology (GSA) of the University of São Paulo for their language help. Our gratitude goes to the editor and reviewers for their work and suggestions.
References
Adorno, R. R., do Carmo, D. A., Germs, G., Walde, D. H. G., Denezine, M., Boggiani, P. C., et al. (2017). Cloudina Lucianoi (Beurlen & Sommer, 1957), Tamengo Formation, Ediacaran, Brazil: Taxonomy, Analysis of Stratigraphic Distribution and Biostratigraphy. Precambrian Res. 301, 19–35. doi:10.1016/j.precamres.2017.08.023
Amorim, K. B., Afonso, J. W. L., Leme, J. M., Diniz, C. Q. C., Rivera, L. C. M., Gómez‐Gutiérrez, J. C., et al. (2020). Sedimentary Facies, Fossil Distribution and Depositional Setting of the Late Ediacaran Tamengo Formation (Brazil). Sedimentology 67, 3422–3450. doi:10.1111/sed.12749
Becker-Kerber, B., Pacheco, M. L. A. F., Rudnitzki, I. D., Galante, D., Rodrigues, F., and Leme, J. M. (2017). Ecological Interactions in Cloudina From the Ediacaran Of Brazil: Implications for the Rise of Animal Biomineralization. Sci. Rep. 7, 5482–5487. doi:10.1038/s41598-017-05753-8
Boggiani, P. C. (1998). Análise estratigráfica da Bacia Corumbá (Neoproterozoico) - Mato Grosso do Sul. Ph.D. Thesis, Brasil: Universidade de São Paulo, 181.
Boggiani, P. C., Gaucher, C., Sial, A. N., Babinski, M., Simon, C. M., Riccomini, C., et al. (2010). Chemostratigraphy of the Tamengo Formation (Corumbá Group, Brazil): A Contribution to the Calibration of the Ediacaran Carbon-Isotope Curve. Precambrian Res. 182, 382–401. doi:10.1016/j.precamres.2010.06.003
Butterfield, N. J. (1995). Secular Distribution of Burgess-shale-type Preservation. Lethaia 28, 1–13. doi:10.1111/j.1502-3931.1995.tb01587.x
Butterfield, N. (2005). Reconstructing a Complex Early Neoproterozoic Eukaryote, Wynniatt Formation, Arctic Canada. Lethaia 38, 155–169. doi:10.1080/00241160510013231
Bykova, N., LoDuca, S. T., Ye, Q., Marusin, V., Grazhdankin, D., and Xiao, S. (2020). Seaweeds Through Time: Morphological And Ecological Analysis of Proterozoic And Early Paleozoic Benthic Macroalgae. Precambrian Res. 350, 105875. doi:10.1016/j.precamres.2020.105875
Cai, Y., Hua, H., Xiao, S., Schiffbauer, J. D., and Li, P. (2010). Biostratinomy of the Late Ediacaran Pyritized Gaojiashan Lagerstatte from Southern Shaanxi, South China: Importance of Event Deposits. Palaios 25, 487–506. doi:10.2110/palo.2009.p09-133r
Cai, Y., Schiffbauer, J. D., Hua, H., and Xiao, S. (2012). Preservational Modes in the Ediacaran Gaojiashan Lagerstätte: Pyritization, Aluminosilicification, and Carbonaceous Compression. Palaeogeogr. Palaeoclimatol. Palaeoecol. 326-328, 109–117. doi:10.1016/j.palaeo.2012.02.009
Chen, M., and Xiao, S. (1992). Macrofossil Biota from Upper Doushantuo Formation In Eastern Yangtze Gorges, China. Acta Palaeontologica Sinica 31, 513–529.
Cohen, P. A., and Macdonald, F. A. (2015). The Proterozoic Record Of Eukaryotes. Paleobiology 41, 610–632. doi:10.1017/pab.2015.25
Dornbos, S. Q., Oji, T., Kanayama, A., and Gonchigdorj, S. (2016). A New Burgess Shale-type deposit from the Ediacaran Of Western Mongolia. Sci. Rep. 6, 23438. doi:10.1038/srep23438
Hedges, S., Blair, J. E., Venturi, M. L., and Shoe, J. L. (2004). A Molecular Timescale of Eukaryote Evolution and the Rise of Complex Multicellular Life. BMC Evol. Biol. 4, 2. doi:10.1186/1471-2148-4-2
Hofmann, H. J. (1994). “Proterozoic Carbonaceous Compressions (‘metaphytes’ and ‘worms’),” in Early Life on Earth. Editor S. Bengtson (New York: Columbia University Press), 342–357.
Knoll, A. H., Javaux, E. J., Hewitt, D., and Cohen, P. (2006). Eukaryotic Organisms in Proterozoic Oceans. Phil. Trans. R. Soc. B 361, 1023–1038. doi:10.1098/rstb.2006.1843
Labaj, M. A., and Pratt, B. R. (2016). Depositional Dynamics in a Mixed Carbonate-Siliciclastic System: Middle-Upper Cambrian Abrigo Formation, southeastern Arizona. U.S.A. J. Sed. Res. 86, 11–37.
Liu, A. G. (2016). Framboidal Pyrite Shroud Confirms the 'Death Mask' Model for Moldic Preservation of Ediacaran Soft-Bodied Organisms. Palaios 31, 259–274. doi:10.2110/palo.2015.095
LoDuca, S. T., Bykova, N., Wu, M., Xiao, S., and Zhao, Y. (2017). Seaweed Morphology and Ecology during the Great Animal Diversification Events of the Early Paleozoic: a Tale of Two Floras. Geobiology 15, 588–616. doi:10.1111/gbi.12244
Maloney, K. M., Halverson, G. P., Schiffbauer, J. D., Xiao, S., Gibson, T. M., Gibson, T. M., Lechte, M. A., et al. (2021). New Multicellular marine Macroalgae from the Early Tonian Of Northwestern Canada. Geology 49, 6. doi:10.1130/G48508.1
Nagovitsin, K. E., Rogov, V. I., Marusin, V. V., Karlova, G. A., Kolesnikov, A. V., Bykova, N. V., et al. (2015). Revised Neoproterozoic And Terreneuvian Stratigraphy of the Lena-Anabar Basin and north-western Slope of the Olenek Uplift, Siberian Platform. Precambrian Res. 270, 226–245. doi:10.1016/j.precamres.2015.09.012
Oliveira, R. S. d., Nogueira, A. C. R., Romero, G. R., Truckenbrodt, W., and da Silva Bandeira, J. C. (2019). Ediacaran Ramp Depositional Model of the Tamengo Formation, Brazil. J. South Am. Earth Sci. 96, 102348. doi:10.1016/j.jsames.2019.102348
Pacheco, M. L., Galante, D., Rodrigues, F., Leme, J. M., Bidola, P., Hagadorn, W., et al. (2015). Insights Into the Skeletonization, Lifestyle, and Affinity of the Unusual Ediacaran Fossil Corumbella. PLoS ONE 10, e0114219–19. doi:10.1371/journal.pone.0114219
Parfrey, L. W., Lahr, D. J. G., Knoll, A. H., and Katz, L. A. (2011). Estimating The Timing of Early Eukaryotic Diversification with Multigene Molecular Clocks. Proc. Natl. Acad. Sci. 108, 13624–13629. doi:10.1073/pnas.1110633108
Parry, L. A., Boggiani, P. C., Condon, D. J., Garwood, R. J., Leme, J. M., McIlroy, D., et al. (2017). Ichnological Evidence for Meiofaunal Bilaterians from the Terminal Ediacaran And Earliest Cambrian Of Brazil. Nat. Ecol. Evol. 1, 1455–1464. doi:10.1038/s41559-017-0301-9
Steiner, R. (1994). Einführung. Berliner geowissenschaftliche Abhandlungen 15, 1–4. doi:10.1007/978-3-322-84265-7_1
Rowland, S. M., and Rodriguez, M. G. (2014). A Multicellular Alga with Exceptional Preservation from the Ediacaran Of Nevada. J. Paleontol. 88, 263–268. doi:10.1666/13-075
Srivastava, P. (2012). “Morphodiversity, Complexity and Macroevolution: Revealed by the Megascopic Life of the Palaeo-Neoproterozoic Vindhyan Supergroup, India, 365,” In Palaeoproterozoic Of India, eds. R. Mazumder, and D. Saha London, Geological Society, Special Publications, 247–262. doi:10.1144/sp365.13
Tang, Q., Pang, K., Yuan, X., and Xiao, S. (2017). Electron Microscopy Reveals Evidence for Simple Multicellularity in the Proterozoic fossilChuaria. Geology 45, 75–78. doi:10.1130/G38680.1
Van Iten, H., Marques, A. C., Leme, J. M., Pacheco, M. L. A. F., and Simões, M. G. (2014). Origin and Early Diversification of the Phylum Cnidaria Verrill: Major Developments in the Analysis of the Taxon's Proterozoic-Cambrian History. Palaeontology 57, 677–690. doi:10.1111/pala.12116
Walde, D. H.-G., Weber, B., Erdtmann, B. D., and Steiner, M. (2019). Taphonomy of Corumbella Wernerifrom the Ediacaran of Brazil: Sinotubulitid Tube or Conulariid Test. Alcheringa: Australas. J. Palaeontology 43, 335–350. doi:10.1080/03115518.2019.1615551
Wan, K. Y., and Jékely, G. (2021). Origins Of Eukaryotic Excitability. Phil. Trans. R. Soc. B 376, 20190758. doi:10.1098/rstb.2019.0758
Warren, L. V., Pacheco, M. L. A. F., Fairchild, T. R., Simões, M. G., Riccomini, C., and Boggiani, P. C. (2012). The Dawn of Animal Skeletogenesis: Ultrastructural Analysis of the Ediacaran Metazoan Corumbella Werneri. Geology 40, 691–694.
Xiao, S., and Dong, L. (2006). “On The Morphological and Ecological History of Proterozoic Macroalgae,” in Neoproterozoic Geobiology And Paleobiology, eds. S. Xiao, and A. J. Kaufman (Dordrecht, Springer), 57–90.
Xiao, S., Knoll, A. H., and Yuan, X. (1998). Morphological Reconstruction of Miaohephyton Bifurcatum, a Possible Brown Alga from the Neoproterozoic Doushantuo Formation, South China. J. Paleontol. 72, 1072–1086. doi:10.1017/s0022336000027414
Xiao, S., Knoll, A. H., Yuan, X., and Pueschel, C. M. (2004). Phosphatized Multicellular Algae in the Neoproterozoic Doushantuo Formation, China, and the Early Evolution of Florideophyte Red Algae. Am. J. Bot. 91, 214–227. doi:10.3732/ajb.91.2.214
Xiao, S., Yuan, X., Steiner, M., and Knoll, A. H. (2002). Macroscopic Carbonaceous Compressions in a Terminal Proterozoic Shale: a Systematic Reassessment of the Miaohe Biota, South China. J. Paleontol. 76, 347–376. doi:10.1017/s0022336000041743
Ye, Q., Tong, J., An, Z., Hu, J., Tian, L., Guan, K., et al. (2019). A Systematic Description of New Macrofossil Material from the Upper Ediacaran Miaohe Member In South China. J. Syst. Palaeontology 17, 183–238. doi:10.1080/14772019.2017.1404499
Zhang, Y., Yin, L., Xiao, S., and Knoll, A. H. (1998). Permineralized Fossils from the Terminal Proterozoic Doushantuo Formation, South China, Memoir Paleontological Society, 72, 1–52. doi:10.1017/s0022336000059977
Keywords: Macroalgae, taxonomy, Ediacaran, Tamengo, Brazil
Citation: Diniz CQC, Leme JM and Boggiani PC (2021) New Species of Macroalgae from Tamengo Formation, Ediacaran, Brazil. Front. Earth Sci. 9:748876. doi: 10.3389/feart.2021.748876
Received: 28 July 2021; Accepted: 14 September 2021;
Published: 29 October 2021.
Edited by:
Haijun Song, China University of Geosciences, ChinaReviewed by:
Katie Maloney, University of Toronto Mississauga, CanadaQin Ye, China University of Geosciences, China
Copyright © 2021 Diniz, Leme and Boggiani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Cleber Q. C. Diniz, Y2xlYmVyLmRpbml6QGhvdG1haWwuY29tLmJy