 Frank Kienast
Frank Kienast Sergei P. Davydov
Sergei P. Davydov
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Earth Sci. , 09 November 2021
Sec. Paleontology
Volume 9 - 2021 | https://doi.org/10.3389/feart.2021.741473
This article is part of the Research Topic Yedoma Permafrost Landscapes as Past Archives, Present and Future Change Areas View all 27 articles
The lower Kolyma region is known for rich Early Olyorian large mammal assemblages including plesiomorphic musk ox, reindeer, horse, broad-fronted moose, steppe mammoth and cave bear. Data on the vegetation in zonal habitats from the Olyorian period are, in contrast, scarce. Earlier palynological results from classical Olyorian sites indicated predominant grassland vegetation with scattered larch occurrences but are, due to limited taxonomical resolution, uncertain. Plant macrofossil data were, so far, only available from azonal, aquatic habitats. Here, we describe Olyorian palaeo-vegetation from an exposure near Cherskiy, Nizhnekolymsky District, Sakha Republic, Russia. The macrofossil-based reconstruction of palaeo-vegetation revealed the existence of an open forest steppe comprising a mosaic of sparse larch groves in a dry grassland composed of tundra steppes, degraded meadow steppes and saline meadows. In the larch groves, light demanding shrubs and dwarf shrubs such as shrub birch (Betula cf. fruticosa, B. nana ssp. exilis), raspberry (Rubus idaeus), cowberry (Vaccinium vitis-idaea), and crowberry (Empetrum nigrum) formed the understory. Ruderal pioneer plants point to open ground as result of disturbances, possibly due to the activity of large herbivores. The nitrophytic ruderal species Urtica dioica, in particular, suggests locally increased nutrient supply from animal excretions. Also, the abundant remains of Chenopodium cf. prostratum might be explained by disturbances and nutrient enrichment, but Chenopodium is also characteristic of salt accumulation in the soil owing to high evaporation under arid conditions, which are also indicated by alkali grass (Puccinellia sp.). The presence of trees and shrubs indicates interglacial-like temperature conditions but the dominance of dry grassland species and the occurrence of facultatively halophytic plants (Chenopodium prostratum, Puccinellia sp.) suggests aridity, which is more typical of cold stages. During the early Pleistocene, i.e., prior to the Early-Middle Pleistocene transition (EMPT) culminating around 900 ka ago, the duration of climate cycles was shorter and the amplitude of climate fluctuations was smaller. Ice-rich permafrost formed only after the EMPT during increasingly extreme cold stages, and, during warm stages, its thawing resulted in paludification of the active layer. Prior to the EMPT, the climate in West Beringia was constantly relatively dry, more or less moderate and more stable than thereafter. In contrast to modern tundra and northern taiga in the study region, dry habitats apparently prevailed during the time of deposition of the plant macro-remains.
Due to continental climate and the consequent lack of extensive glaciation, Beringia was a refugium for arctic biota during Pleistocene warm stages and center of dispersal during cold stages, when cold-adapted taxa spread over much of Eurasia far into the middle latitudes. Also, the intercontinental faunal exchange between Eurasia and the New World took place via the Beringian landmass during phases of global sea level lowering. As result of ice sheet formation in Northern Europe and North America and the fixation of tremendous water masses, the huge and shallow Laptev, East Siberian, Chukchi and Bering shelves in the North of Yakutia and in the East of Chukotka became, at such times, fully exposed and together formed the Beringian landmass.
Apart from being a refugium during the late Middle and Late Pleistocene, the western part of Beringia is furthermore regarded as center of development or center of origin of preadapted late Pleistocene mammoth faunal complex components, like Ovibos (resp. Praeovibos), Rangifer, Gulo, Alopex and advanced forms of Mammuthus, which evolved there during the late Early to the early Middle Pleistocene giving rise to define the Northeast Siberian land mammal age Olyorian (Sher, 1986; Kahlke, 1999).
The Olyorian comprises a long period of time for the evolution of the mammoth faunal complex spanning from the late Early to the early Middle Pleistocene (early Eopleistocene to early Neopleistocene according to the Russian nomenclature), i.e., 1.4–0.5 Ma (Sher, 1971; Sher, 1986), though without exactly defined lower and upper boundaries. The type locality of the Olyorian is situated at the Chukochya River in the Kolyma lowlands about 150 km NW of Cherskiy (Figure 1; Sher, 1986). Classical Olyorian sites in NE Siberia are furthermore located at the Krestovka River (Figure 1; Sher et al., 1979) as well as at the Adycha River (Kaplina et al., 1983) and in the surroundings of Cherskiy, where important Olyorian sites are known from Tretiy Ruchey between Zeleniy Mys and the town of Cherskiy (Sher et al., 2011) as well as the Cherskiy Ovrag (Davydov, 2007) from where plant remains are described in the present paper.
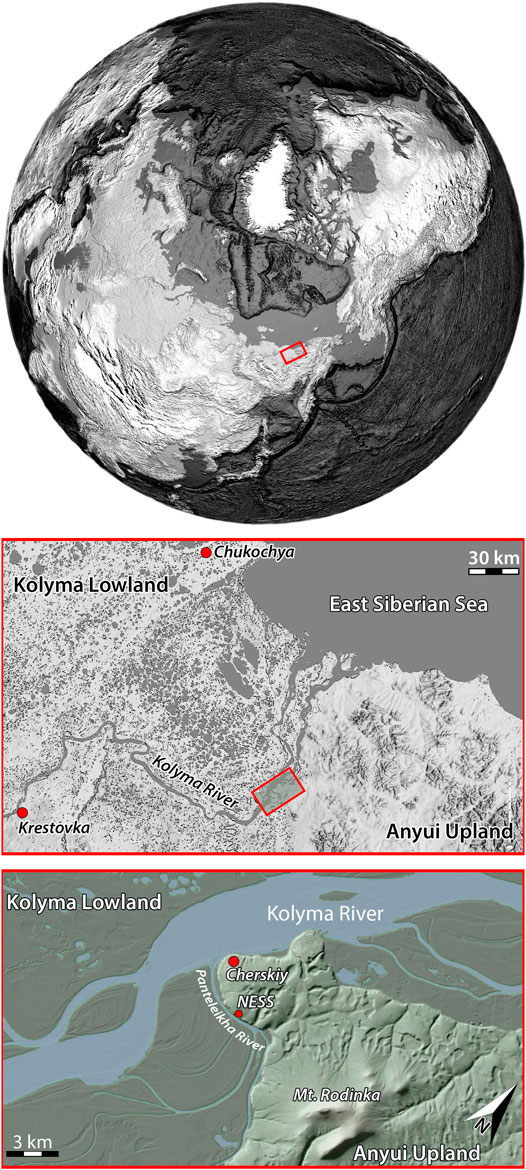
FIGURE 1. Location in the western part of Beringia (upper part), topography and regional context of the study site at the junction of the Northeast Siberian coastal lowlands and the Far Eastern Anyui uplands (central part) as well as location of the Chukochya and Krestovka sites. The study site is situated in close proximity to the North East Science Station (NESS) Cherskiy at the westernmost edge of the Anyui Upland (lower part). Upper part adapted from the NGDC, NOAA Satellite and Information Service, available at http://www.ngdc.noaa.gov/mgg/global/; central and lower part Digital Elevation Model adapted from the Arctic DEM Explorer available at https://livingatlas2.arcgis.com/arcticdemexplorer.
As part of the regional Northeast Siberian (West Beringian) Quaternary sequence, the Olyorian mammal age can be further separated into two faunal units on the base of the evolutionary succession of collared lemmings (Sher, 1997). The lower Olyorian (Chukochyan) is characterized by the presence of the predecessor of modern collared lemmings Predicrostonyx compitalis and the likewise archaic vole Allophaiomys pliocaenicus and is correlated with the late part of the reverse polarity Matuyama Chron in the palaeomagnetic record including the inserted magnetically normal Jaramillo event. The upper Olyorian (Akanian) is defined by the occurrence of the more advanced collared lemming Dicrostonyx renidens and corresponds with the earliest part of the normal polarity Brunhes Chron although also including the Matuyama/Brunhes reversal and the uppermost Matuyama Chron.
The Olyorian is characterized by a unique assemblage of large mammals, which appeared for the first time in this region and, in the case of Gulo, Rangifer and Mammuthus primigenius, for the first time worldwide. Remains of an “archaic northern elephant” in Chukochyan deposits and of a more advanced form in Akanian sediments (Sher, 1986) represent a distinct mammoth lineage. They are actually morphologically very advanced pre-empting an evolutionary level of mammoths that occurred outside Beringia only 300–400 ka later (Sher, 1986; Kahlke, 1999). Genomic analyses based on ancient DNA with an age of more than 1 million years prove the affiliation of lower Olyorian mammoth remains at the Krestovka and Adycha sites to the steppe-mammoth lineage (van der Valk et al., 2021). Genomic data from another, only 650,000-year-old, thus Akanian, mammoth found at the Chukochya site reveal that it was an early representative of the woolly mammoth (M. primigenius), which thus evolved in Western Beringia and descended from the Adycha lineage of the steppe-mammoth (van der Valk et al., 2021).
Numerous bones of a very large horse assigned to Equus (Plesippus) verae are characteristic of both units, the Chukochyan and the Akanian. According to recent cladistic analyses of morphometrical measurements of cranial and postcranial elements of 30 extinct and extant horse taxa, Plesippus must be placed within the genus Equus (Cirilli et al., 2021). Equus verae belongs to the stenonine clade, which comprises modern zebras and asses and descends from E. stenonis and, as all Old-World Equids, from its North American progenitor E. simplicidens. Equus suessenbornensis, occurring in Central Europe later during the Middle Pleistocene, coincides morphometrically with E. verae and is regarded successor of an Asian parent form (Forsten, 1986 referred to in; Kahlke, 1999). In addition to the large E. verae, the sympatric occurrence of a second, smaller, equid in Olyorian deposits is reported (Sher et al., 1979). From the upper (Akanian) unit, also caballine horses are described (Sher, 1986).
Remains of a giant moose (Cervalces sp.) and of Bison sp. were regularly found in both units. Both taxa were, at this time, widespread over the Eurasian middle and high latitudes as is documented by abundant findings, e.g., at the Epivillafranchian site of Untermassfeld in Germany (Kahlke et al., 2011). Also cave bears had an extensive distribution in middle latitudes at this time, but the discovery of a mandible determined as Ursus savini ssp. nordostensis near Cherskiy represents the northernmost and easternmost find of spelaeoid bears worldwide (Sher et al., 2011).
The Olyorian findings of Rangifer ex. gr. tarandus are, together with fossils from Cape Deceit, Kotzebue Sound, Western Alaska, the oldest known reindeer fossils worldwide suggesting the origin of reindeer in Beringia (Sher, 1986; Kahlke, 1999). An archaic form of another typically arctic faunal element is represented in Olyorian deposits as well: the oldest finds of Praeovibus beringiensis indicate the origin of musk ox in NE Siberia (Sher, 1971). Praeovibos can be considered as early morphotype of the extant Ovibos moschatus as is suggested by ancient DNA studies (Campos et al., 2010). Apart from Praeovibos, postcranial remains of another, smaller Ovibovine were also detected in Olyorian deposits and might possibly represent Soergelia (Sher et al., 1979), which is proven for the upper unit Akanian (Sher, 1986). Soergelia was widely distributed in NE Siberia (Sher, 1971; Boeskorov, 2019) and probably originated in the NE Siberian Arctic (Kahlke, 1999). Thus, it is regarded a Beringian bovid as well.
Even Saiga, undoubtedly an immigrant from the zonal steppe belt, was already present in West Beringia during Olyorian times. The Olyorian finds of Saiga are the worldwide oldest known fossils of that genus so far (Kahlke, 1999). As saiga is, more than any other Beringian faunal element, indicative of zonal steppe grasslands, it can be assumed that it evolved in the inner continental steppe belt.
Some large mammals expanded into the region from the inner continental steppe belt, as is the case of Saiga, Equus and Cervalces. Several of the listed taxa evolved, however, in Beringia during the Early Pleistocene developing adaptations to environmental conditions making them fit for huge range expansions across the Holarctic and far into the mid latitudes during later cold climate phases, which became, in the course of the Early-Mid Pleistocene Transition, progressively longer and colder. The Olyorian faunal complex is therefore regarded as an important predecessor of the late-Pleistocene Mammuthus-Coelodonta faunal complex (Kahlke, 1999).
Only limited data are available on the habitat conditions for large herbivores in Beringia during the Olyorian period. Palaeobotanical and entomological data from the Krestovka and the Chukochya sites suggest that climate and vegetation changed only slightly within the deposition period of the sequence (Giterman et al., 1982). According to palynological results from the Krestovka sequence, non-arboreal pollen with Poaceae, Artemisia, Chenopodiaceae and Caryophyllaceae were most abundant throughout the studied sequence indicating open grassland vegetation and aridity. The existence of such environments is confirmed by entomological data revealing the prevalence of grassland dwellers in Chukochyan deposits and the existence of few taiga species suggesting that groves of trees and shrubs persisted. The percentage of arboreal taxa in the pollen spectra, mainly Alnus (alnobetula ssp.) fruticosa, Betula, sp., Betula Sect. Nanae and Pinus pumila with rare Larix, declined from up to 40% at the base of the sequence to nearly 5% in the middle part and then slowly increased towards the top of the sequence. Available plant macrofossil data represent only azonal, aquatic vegetation presumably from small ponds (Giterman et al., 1982). Due to the limitations of pollen data in northern environments, like overrepresentation of long-distance-transported pollen in conjunction with low local pollen production and the lack of taxonomic differentiation (Birks and Birks, 2000), plant macrofossil data are, however, especially valuable for the reconstruction of vegetation and environmental conditions in particular when they represent zonal vegetation.
In the present paper, we use plant macrofossils sampled from Early Olyorian (Chukochyan) deposits in a gully near Cherskiy, to reconstruct zonal vegetation and environmental conditions existing at the life-time of the cold pre-adapted Beringian faunal complex.
The sampled outcrop “Cherskiy Ovrag” is situated near the North-East Science Station of the Pacific Geographical Institute and about 3 km east of Cherskiy at the right bank of the Panteleikha River, a tributary of the Kolyma River (Figure 1). The study site is located at the northeastern edge of Yakutia at the junction to the Anyui Upland macroslope. Starting from the left bank of the Panteleikha River, the Kolyma lowland extends over hundreds of km to the west and merges with the Yana-Indigirka lowland, together forming the vast Northeast Siberian coastal lowlands.
The bedrock exposed at the study site and at other sites along the right banks of the Kolyma and Panteleikha Rivers in the vicinity of Cherskiy is composed of Triassic sedimentary rocks cut through by small intrusions of Jurassic granitoids, and less often by Cretaceous effusive bodies. Subsequently, these bedrocks and the Neogene weathering crust were covered with permafrost sediments of various Pleistocene ages (Sher et al., 1979; Davydov, 2007; Sher et al., 2011). The major portion of rocks in the study area is overlain by thick ice-rich silty-sandy deposits of the late Pleistocene Yedoma Formation (Sher et al., 1979; Davydov et al., 2009) or by products of their Holocene transformation. Currently, these deposits are widely eroded and disturbed by industrial extraction of rock debris at the right banks of the Panteleikha and Kolyma Rivers. In particular, these disturbances exposed a formerly buried relief with fragments of ancient erosional channels cutting into the Neogene weathering crust. These channels resemble gullies, so-called ovrags (овраги) in Russian.
The entire territory of Yakutia is situated in the zone of continuous permafrost. Owing to the lack of inland glaciations, ice-bonded permafrost deposits (Ice Complex), penetrated by thick ice wedges and consisting of up to 90% of ice, formed on the Northeast Siberian coastal lowlands during cold phases of the Late Pleistocene (Tomirdiaro, 1980; Romanovskii et al., 2004). The adjacent shelves, which were subaerially exposed during Pleistocene cold stages, are still underlain by relict permafrost down to depths of 300–700 m (Romanovskii et al., 2004). The upper part of the Ice Complex largely eroded in the course of the Holocene resulting in characteristic thermokarst landforms. In recent years, Ice Complex deposits older than Late Pleistocene became exposed near Batagay in the Yana Highlands illustrating the continuity of cold continental climate in Northern Yakutia back until, at least, the Middle Pleistocene (Ashastina et al., 2017; Murton et al., 2017, 2021). Ice wedge casts in Olyorian deposits and in underlying Kutuyakh deposits at the Bolshaya Chukochya and Krestovka sites indicate that continuous permafrost existed at the time of their deposition, thus already during the Early Pleistocene (Giterman et al., 1982).
Climate in Yakutia is cold continental, i.e., the annual mean temperature is everywhere negative with very cold winters. The lowest temperature of the Northern hemisphere was measured in Verkhoyansk in the Yakutian Yana Highlands. The temperature during the growing season is the main factor controlling modern vegetation. During the summer, the temperature is relatively high in inland areas such as the Central Yakutian Plain and the Yana Highlands but considerably decreases towards the coast due to the cooling effect of the sea and increasing cloud cover. This maritime effect increases also eastwards at the Chukchi Peninsula. The region around Cherskiy is, thus, a transitional area, covering an E-W continentality gradient from inland Yakutia to Chukotka and, due to the proximity to the coast, a N-S gradient in terms of summer temperature and vegetation illustrated by the polar tree line, which is only 25 km to the Northwest. Climatic data from the weather station in Cherskiy (WMO 25123) indicate a mean annual air temperature of −9.7C, a mean temperature of the coldest month of −33.5C and a mean temperature of the warmest month (July) of 13.9C. The mean seasonal temperature gradient varies within 45.0–49.0C. The annual precipitation is very low with about 215 mm (Russia’s Weather Server, 2021) but, due to low evapotranspiration, the prevention of downward percolation by permafrost and additional moisture supply by thawing permafrost, the active layer is, on zonal sites, wet.
The zonal vegetation in the study area is sparse northern taiga and near-tundra woodland (forest tundra) composed of the extremely cold-tolerant deciduous larch (Larix gmelinii var. gmelinii; synonym with after GBIF https://www.gbif.org/species/2686192) with shrub alder (Alnus alnobetula ssp. fruticosa), shrub birches (Betula divaricata, B. fruticosa, B. exilis), more than a dozen species of shrub willows (Salix spp.) and Ericaceae (Vaccinium vitis-idaea, Rhododendron tomentosum, Arctous alpina) in the understorey (Troeva et al., 2010). The tree line is situated only about 25 km to the North where the floristically rich subarctic tundra of the Kolyma Lowland extends, a huge wetland that is covered with countless thermokarst lakes (Figure 1, middle part). The region is also known for the presence of extrazonal relict steppe patches considered as potential analogues of Pleistocene vegetation (Yurtsev, 1982). Detailed lists of the local flora were provided by Yurtsev, (1974), Kozhevnikov (1981), Zaslavskaya and Petrovskii (1994), Maksimovich (1998) and Davydov et al. (2020). In the lower Kolyma region, steppe patches are restricted to specific sites in southern exposures and to coarsely clastic substrate. One of the most extensive steppe patches in the whole region is immediately adjacent to the sampling site. An outline of the vegetation occurring in the study area is given by Kienast (2016).
In the course of quarrying rock debris from the Neogene weathering crust for airstrip reconstruction at the nearby Cherskiy Airport, ancient erosional channels were uncovered by bulldozers. These channels or gullies (ovrags) represent former erosive cuts into the bedrock that were subsequently filled with Pleistocene permafrost deposits. The extraction of rock gave access to Late Pleistocene Yedoma sediments and, in their lower parts, to fossiliferous Early Olyorian deposits rich in vertebrate fossils (Figure 2).
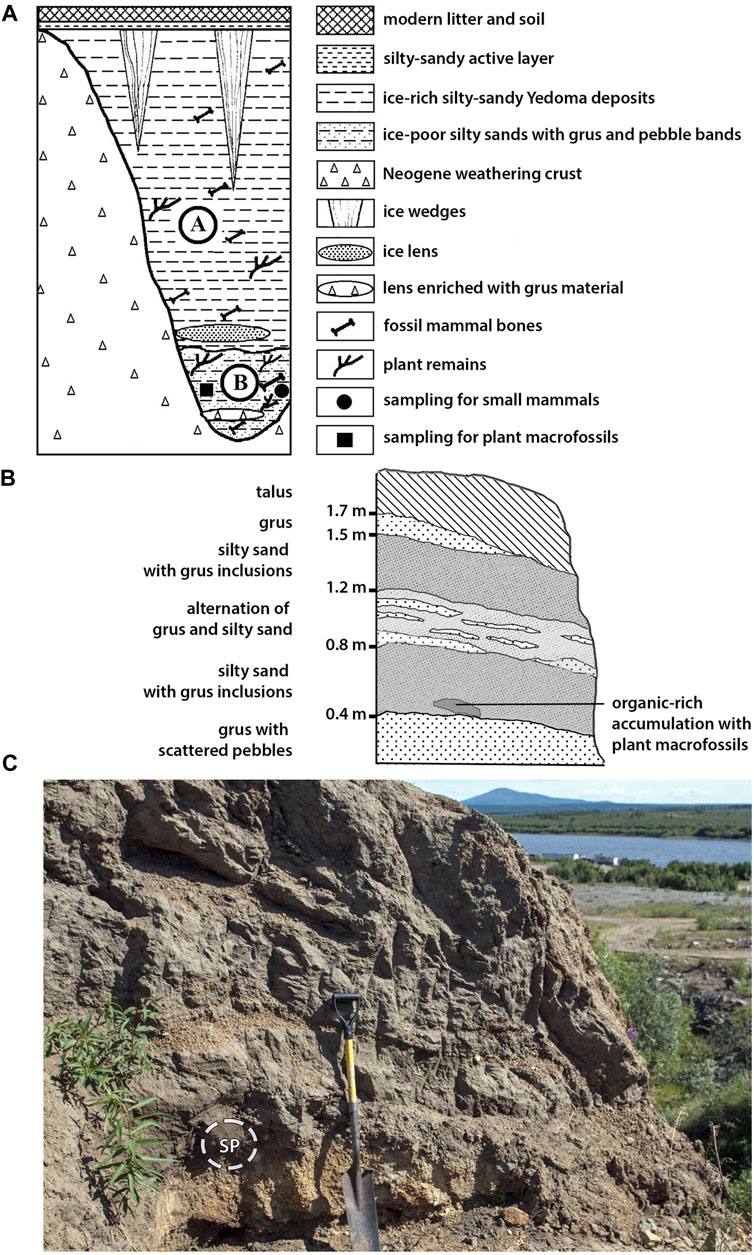
FIGURE 2. (A) Scheme of the original stratigraphic sequence at the Cherskiy Ovrag reconstructed on the basis of long-term monitoring by S.P. Davydov. The upper unit A–the late Pleistocene Yedoma Ice Complex—became eroded in the frame of industrial exposure and subsequent thawing, which gave access to the underlying unit B–the Olyorian sequence. (B) Sketch and description of the Olyorian section (unit B) exposed at the time of sampling. (C) Photo of the studied exposure and sampling point (SP) within the sequence. In the background, the Panteleikha River and Mt. Panteleikha are visible.
S.P. Davydov (2007) studied systematically the stratigraphical sequence and collected large mammal remains that became exposed as result of quarrying or of natural erosion in these gullies between the years 2000 and 2018. The above-lying Ice Complex deposits contained numerous bones of typical Late Pleistocene mammoth faunal representatives such as Mammuthus primigenius, Bison priscus, Rangifer tarandus and Equus sp.
The Olyorian deposits underlying the Ice Complex revealed a quite different faunal composition. 25 percent of all mammal bones recorded at the Cherskiy Ovrag originated from Ovibovini (Sher et al., 2011), which is in stark contrast to late Pleistocene assemblages in NE Siberia, where musk oxen play only a minor role in the large mammal composition in favor of the then much more abundant bison (Sher, 1971). Most of these Ovibovini remains can be assigned to the extinct Praeovibos beringiensis Sher et al. (2011), which is, according to Campos et al. (2010), considered as an archaic morphotype of the extant musk ox. Furthermore, bones of the large, archaic, stenonine horse Equus (subgen. Plesippus) verae, a large caballoid horse (Equus subgen. Equus), likewise large bodied Bison and mammoth forms, broad-fronted moose (Cervalces sp.), an ancient maral (Cervus sp.) and hare (Lepus sp.) were found at the site (Davydov, 2007). Finally, a mandible and a canine of Ursus savinii nordostensis, a small extinct cave bear, was found in the Cherskiy Ovrag indicating a much more extended range of cave bears than formerly expected (Sher et al., 2011).
From one of the gullies that were formerly sampled for large mammal bones, P. Nikolskyi screened fine sandy and silty sediments for the analysis of small mammals, which are crucial for biostratigraphical dating. The results, published by Sher et al. (2011), revealed, beside Lemmus sp. remains, the presence of the extinct vole Allophaiomys reservatus and a plesiomorphic collared lemming (Predicrostonyx compitalis) characteristic of the lower Olyor (Chukochyan). Two other teeth stem either from a more advanced form of P. compitalis or from its Late Olyorian successor Dicrostonyx renidens. Thus, both large and small mammal remains indicate an age of the studied sediments belonging to the Early Olyorian (Chukochyan).
From the layers earlier screened for small mammal remains, we took material for plant macrofossil analyses in 2015 (Figure 2). At the time of sampling, only a small section of the outcrop was accessible and we took the sample from the only observable organic-rich horizon, which apparently contained plant remains. The sampled sediments were wet-sieved using various mesh sizes from 250 µm up to 1 mm and then air-dried. The residue was then manually screened for identifiable plant remains using a zoom binocular (Olympus SZX 16) with variable magnification between 7 and 115 times. Identification of plant remains was conducted using modern plant material from the carpological collection of the Herbarium Senckenbergianum (IQW) as a reference. The residue contained many woody remains, mainly bark fragments, which probably originate from conifers (most likely larch), but they were not identified in further detail as we did not have modern material in the reference collection available and the taxa in question are represented by other, easily identifiable organs such as seeds, fruits and needles.
We recovered 750 plant macrofossils from altogether 33 plant taxa. The abundance of individual taxa was not considered in detail because the number of preserved seeds and fruits does not necessarily reflect the actual percentage of plant taxa in the former vegetation but depends on taphonomic circumstances and other coincidences. The studied assemblage represents plants from a wide spectrum of habitats co-occurring in distances of few dozens to hundreds of meters probably during a period of few years. As the plant remains are partly tiny and are well preserved, a redeposition over long distances can be excluded. Plant species only occur under environmental conditions that meet their requirements and together with other plants with similar requirements. Their find thus indicates the presence of these conditions, habitats and communities even when the original set of plant species is incomplete and fragmentary. The recovered taxa were assigned to discrete plant communities according to the phytosociological classification following the ecological requirements of their modern representatives. They are listed in Table 1 in the order of their probable occurrence in certain plant communities.

TABLE 1. List of the identified plant macrofossils and their classification into plant communities (syntaxa).
We found several boreal woody plants characteristic of the northern larch woodland that constitutes the zonal vegetation in the area today (Krestov et al., 2009). Several needles, fascicles and seeds indicate the presence of the Dahurian larch (Larix gmelinii s.l., Figure 3), which is competitive only under extreme cold continental climate in regions with continuous permafrost such as north-eastern Yakutia. We furthermore detected a pyrene of Rubus idaeus, which is characteristic for dry forest edges and glades, and frequently occurs after wildfires. The presence of birches is indicated by ancient fruits corresponding to those of Betula cf. fruticosa and B. nana ssp. exilis. Some wingless nutlets resemble those of tree birches (Figure 3), but it cannot be excluded that the remains stem from B. divaricata or even from Alnus alnobetula ssp. fruticosa, which have similar demands and often occur together. Shrub birches form the understorey of open larch forests and, as secondary woods, replace larches after disturbances such as wildfires and north of the tree line. Furthermore, Vaccinium vitis-idaea and Empetrum nigrum were detected as seeds. Both dwarf-shrubs are likewise typical for the understorey of open dry boreal woods and advance also beyond the tree line. Among forbs, we found seeds of Stellaria longifolia, which mainly occurs in the herb layer of boreal woodland. The shrubs and dwarf shrubs are light-demanding suggesting an open character of the groves.

FIGURE 3. Olyorian macrofossils of woodland plants. 1–Larix gmelinii s.l., needle fragment, 2–L. gmelinii s.l., spur branch fragment with young needles preserved, 3–L. gmelinii s.l., seed from two sides, wing is not preserved but the attachment scar of the former wing is distinct, 4–Betula cf. fruticosa, nutlet with partly preserved wings from both sides, 5–Betula cf. Sect Albae, nutlet, wings are not preserved but maximum width in upper third suggests assignment to tree birch, 6–Betula nana s.l., nutlet with completely preserved wings from both sides, 7–Empetrum nigrum, seed from two sides, 8–Vaccinium vitis-idaea, seed from both sides, 9–Stellaria longifolia, seed from both sides, 10–Rubus idaeus, pyrene from two sides.
The majority of identified plant remains originate from herbaceous taxa, which can be assigned to several grassland communities that together formed the ancient ecosystem or palaeo-biome mammoth steppe, steppe-tundra or tundra-steppe, in Beringia during Pleistocene cold stages (Yurtsev, 1972; Yurtsev, 1982; Yurtsev, 2001; Guthrie, 1982; Kienast, 2013). As the term tundra-steppe was only recently defined for the English description of the plant-sociological class Carici rupestris-Kobresietea bellardii occurring at dry places in the Arctic or in the alpine belt of mountains (Kucherov and Daniels, 2005), we will use, for description of the zonal vegetation during Pleistocene cold stages, the term mammoth steppe as proposed by Guthrie. As all biomes, the mammoth steppe can be regarded as a mosaic of different communities such as tundra steppe, meadow steppe and saline meadows occurring as a function of moisture, substrate, exposition, disturbances and other environmental factors. In the following, we will describe the grassland communities reconstructed on the base of phytosociology of modern vegetation.
As mentioned above, tundra steppes are characteristic for dry, exposed places in the Arctic such as pingos as well as for equivalent habitats in mountains above the tree line. Tundra steppe communities often merge into steppe communities and a differentiation is sometimes difficult. In the Olyorian plant macrofossil assemblage of the Cherskiy Ovrag, remains of Ranunculus pedatifidus var. affinis, Smelowskia sp., Saxifraga cernua, Silene involucrata, Phlox sibirica, Arnica frigida, Draba sp., Artemisia sp., Papaver Sect. Scapiflora, and Selaginella rupestris were detected (Figure 4). They indicate, for the time of deposition, cold and dry conditions and, in particular, a thin snow cover in winter. Some species within the Draba and Papaver Sect. Scapiflora clades occur, aside from tundra steppes, also in Arctic pioneer vegetation (Thaspithea rotundifolii). Tundra steppe communities were a constitutive component in the vegetation complex of the Yakutian coastal lowlands during the last cold stage and were, in earlier publications, designated as Kobresia-meadows or as dry arctic upland vegetation (Kienast et al., 2005; Kienast, 2013) and later defined as tundra-steppe by Kucherov and Daniels (2005).

FIGURE 4. Olyorian macrofossils of grassland and ruderal plants. 1–Arnica frigida, modern cypsela without pappus as reference, 1–A. frigida, fragment of cypsela, upper part with preserved pappus ring, 3–A. frigida, fragment of cypsela, basal part with abscission scar, 4–Artemisia sp., cypsela from two sides, 5–Saxifraga cernua, ancient seed, 6–Potentilla arenosa, nutlets, 7–Ranunculus pedatifidus ssp. affinis, nutlet fragment (carpel) from two sides, 8–Silene involucrata, seed from two sides, 9–Phlox sibirica, valve fragment of seed pod, 10–Smelowskia sp., seed from both sides, 11–Androsace septentrionalis, seed from two sides, 12–Rumex Subgen. acetosella, nutlet from two sides, 12a–R. Subgen. acetosella, detail of nutlet surface, 13–Selaginella rupestris, megaspore from two sides, 14–Draba sp., seed from both sides, 15–Papaver Sect. Scapiflora, seed, 16–Poa sp., caryopsis from two sides, note the circular hilum near the base at the left picture, 17–Festuca sp., caryopsis from two sides, note the linear hilum at the centre of the fruit, 18–Koeleria sp., caryopsis from two sides, 19–Puccinellia sp., caryopsis from two sides, 20–Chenopodium cf. prostratum, seed from both sides, 21–Polygonum aviculare, nutlet from two sides, 22–Carex duriuscula, ancient nutlet from both sides, 22a–detail of surface, 23–C. duriuscula, modern nutlet for comparison, 23a–detail of surface, 24–Sibbaldianthe bifurca, nutlet from two sides, 25–Urtica dioica, ancient seed from two sides.
During the cold stages of the Late Pleistocene, meadow steppes formed the primary vegetation in the extreme continental inner-Yakutian Yana Highlands, where the seasonal temperature gradient was higher and precipitation was likely even lower than in the Kolyma basin (Ashastina et al., 2018). The above-reported cryoxeric tundra steppe communities (Kobresietea) were largely absent there during the Late Pleistocene and completely replaced by meadow steppes (Ashastina et al., 2018). Remains of Androsace septentrionalis, Potentilla arenosa, Festuca sp., cf. Koeleria sp., Poa sp. and Rumex Subgen. acetosella are represented in the Olyorian palaeobotanical record from Cherskiy (Figure 4). Steppe vegetation reconstructed from plant macrofossil spectra in West Beringia was, in earlier papers (e.g., Kienast et al., 2005), subsumed under the European/West Asian steppes Festuco-Brometea. A revision of the phytosociological classification of Asian steppe vegetation followed only later (Ermakov et al., 2006) according to which the Eastern steppes of Mongolia, Transbaikalia and Yakutia form an own class (Cleistogenetea squarrosae) comprising two orders, true steppes (Stipetalia krylovii) and meadow steppes Festucetalia lenensis. True steppes have their northernmost occurrence currently in Central Yakutia, whereas meadow steppes primarily occur in less dry, more northern or mountainous areas often in a mosaic with herb-rich larch groves. In the study region at the lower Kolyma, appearances of thermophilic wormwood-grass-herb vegetation can be attributed to meadow steppes (Davydov et al., 2020). In the paleo-record of Northern Yakutia, they indicate warm and dry habitats such as south-exposed slopes. Some of the meadow steppe taxa found in the Cherskiy Ovrag occur in tundra-steppe communities as well.
Several of the detected species, particularly Potentilla cf. bifurca, Carex duriuscula, Chenopodium cf. prostratum, Polygonum aviculare, and Puccinellia sp., (Figure 4) indicate either degradation of grassland vegetation or salt influence or both. Sibbaldianthe bifurca is regarded, together with Carex duriuscula, Chenopodium prostratum, and Polygonum aviculare, as indicator of overgrazing in severely degraded steppes of Mongolia and Russia (Hilbig, 1995; Abaturov et al., 2005). The plant community occurs today on disturbed, nutrient-rich, heavily compacted ground near yurts or resting places for livestock, often in the range of seasonally dry drainage channels. Such sites are, due to arid climate conditions, often slightly saline. The presence of alkali grass (Puccinellia sp.) in the Olyorian palaeo-record suggests salt accumulation in the soil due to high evaporation and resulting capillary rise of solutes. Puccinellia sp. was one of the most abundant taxa in Late Quaternary cold stage plant macrofossil assemblages from the Arctic coastal lowlands of Yakutia, which were situated, during the time of deposition, far inland and were affected by more continental, arid climate (Kienast et al., 2005; Kienast et al., 2011). Salt accumulation in the soil is due to the combined effect of aridity, a lack of drainage in depressions and permafrost preventing percolation of solutes (Yelovskaya et al., 1966). Also, the other listed plant taxa are, to a certain degree, halotolerant. Chenopodium prostratum is together with Puccinellia described from saline meadows on solonchak (salty soils) along the shore lines of shrinking lakes and ponds in relict steppe landscapes at the middle course of the Indigirka River often disturbed by horses seeking salt (Yurtsev, 1982). The species occurs, in the steppe zone, also in nitrophilous forb communities together with Sibbaldianthe bifurca and Urtica dioica. Urtica dioica occurs in various floodplain, woodland and forb communities as well as in moist eutrophic ruderal vegetation (Ellenberg, 1996). The occurrence of the species is controlled by the access to solved nitrogen and phosphorus (Šrůtek and Teckelmann, 1998) – nutrients that are, in modern taiga and tundra soils, deficient (Guthrie, 1982; Chapin et al., 1995) but might have been released by Pleistocene herbivores via urine and faeces (Weber, 1914; Johnson, 2009; Mania et al., 2010). The detection of an ancient Urtica dioica seed in the Olyorian assemblage (Figure 3) therefore indicates zoogenic nutrient enrichment of the ground suggesting that disturbances and compaction is likewise the result of the work of megaherbivores, which were abundant in the study area during the Olyorian period.
The remaining taxa in the plant macrofossil assemblage of the Cherskiy Ovrag, Juncus biglumis, Luzula kjellmaniana, Equisetum palustre, Characeae, and Carex sp., can be assigned to wetland vegetation as it is typical for modern arctic tundra. The plants might have occurred near or, in the case of Characeae, in ponds or adjacent to snowbeds. As the fruits of Carex were only poorly preserved, we could not identify the remains down to the species level. Sedges occur in a variety of habitats in the high latitudes ranging from steppes like C. pediformis, C. obtusata or C. duriuscula, over tundra steppes like C. rupestris or C. argunensis to wetlands like C. aquatilis ssp. stans or C. chodorrhiza. The assignment of Carex remains to wetland vegetation is thus to be regarded tentative. Wetland plants are, in the studied assemblage, underrepresented suggesting that permanently wet habitats were, during the time of deposition, less common than today.
The composition of the studied Olyorian plant macrofossil assemblage of the Cherskiy Ovrag reflects predominant grassland vegetation consisting of meadow steppes and tundra steppes interspersed with wooded patches composed of cold-resistant deciduous trees and shrubs (Larix gmelinii s.l. and Betula). The results give the impression of a mosaic-like open landscape with sparse larch groves and are consistent with pollen and entomological data from the Krestovka site, about 160 km to the southwest of Cherskiy (Figure 1; Giterman et al., 1982), for the Olyorian mammal age. Like the Cherskiy Ovrag, the Krestovka site is situated at the junction of the Kolyma lowland and an upland, in this case the Yukagir Plateau, and the Chukochyan deposits at the Krestovka site display a similar lithology mainly comprising alluvial sand interspersed with sandy silt beds. In the pollen data, low levels of shrub taxa and larch are consistent with our data. Also, the abundance of Poaceae, Artemisia, Chenopodiaceae and Caryophyllaceae in the pollen spectra confirm steppe-like vegetation as reconstructed with our macrofossil data from the Cherskiy Ovrag. As described in the introduction, plant macrofossils from the Krestovka site mainly originate from aquatic plants such as Sparganium, Menyanthes and several Potamogeton species (Sher et al., 1979). The few terrestrial plant remains stem from Larix and Rubus; both taxa are preserved also in the studied deposits of the Cherskiy Ovrag.
In our results, the zonal vegetation, i.e., the vegetation that correlates with the macroclimate and that attains dominance under this climate, is represented by dry grassland, i.e., tundra steppe and meadow steppe. The majority of identified plant remains in both abundance and diversity belong to dry grassland taxa, which cannot tolerate shading, as occurring in closed woodlands, or surplus moisture in the active layer, as in tundra wetlands.
Among the mammals preserved in Olyorian deposits, there are several grassland taxa, which indicate an open landscape and steppe-like vegetation as well. Saiga, an immigrant from the zonal steppe belt, indicates dry climate, a thin snow cover, a firm ground or, respectively, a dry active layer and widespread grassland vegetation. Chenopodiaceae, detected in both the pollen and macrofossil data (Chenopodium prostratum), are, besides other steppe forbs, considered the basic food of saiga (Kahlke, 1999; Abaturov et al., 2005). The presence of the giant moose Cervalces with its iconic huge antlers suggests vegetation was open rather than consisting of closed taiga forests. Remains of steppe mammoth (M. trongontheri), and bones of archaic giant horses (E. verae) are also clue for open grassland and steppe like vegetation existing during the time of deposition. Steppe mammoths had already evolved abrasion-resistant teeth as an effective adaptation to relatively hard grass-rich diet and are thus indicators of steppe-like vegetation.
In the presented Early Olyorian (Early Pleistocene) plant macrofossil assemblage, the presence of trees and shrubs indicates interglacial-like temperature conditions but the dominance of dry grassland species and the occurrence of halotolerant plants suggests aridity, which is actually rather characteristic of cold stages. The palaeontological data suggest that the described grass and forb dominated open forest steppe persisted over a long period of time, i.e., during the entire Olyorian age as is indicated by pollen data from both the Chukochyan and the Akanian units at the Krestovka site continuously dominated by herb pollen, mainly from Poaceae, Artemisia and Chenopodiaceae and with only a small amount of larch and birch over all of the sequence (Giterman et al., 1982).
The Olyorian mammal age comprised a long period of time spanning from the late Early to the early Middle Pleistocene, i.e., 1.4–0.5 Ma (Sher, 1986) and, thus, included the Early-Middle Pleistocene transition (EMPT), which occurred about 1.25–0.7 Ma ago (between MIS 37, and MIS 18). The EMPT is described as a shift from relatively low-amplitude 41 kyr lasting climate cycles with a more or less symmetrical curve of warming and cooling to high-amplitude 100 kyr cycles with a more sawtooth-like pattern as result of abrupt warming followed by a slow descent of temperature (Pisias and Moore, 1981; Clark et al., 2006; Cohen and Gibbard, 2019). Prior to the EMPT, the amplitude of climate fluctuations was smaller and the duration of climate cycles was shorter, which possibly resulted, in West Beringia, in relatively dry, cool but more moderate climate that became progressively colder in the course of the EMPT but remained more stable than in other parts of the Holarctic. The palynological record from the composite sediment core of Lake El’gygytgyn, ICPD Site 5,011-1, about 467 km SE of the Cherskiy Ovrag provides valuable information on the regional history of vegetation and climate in West Beringia over the EMPT interval, i.e., between 1,091 and 715 kyr (Zhao et al., 2018). The pollen data indicate shrub tundra and cold steppe communities alternatingly dominating over the major part of the studied Early Olyorian period until MIS 24 (from 1,091 until 930 kyr) with Poaceae, Cyperaceae and Artemisia pollen and high contents of Selaginella rupestris spores prevailing during cold phases and increased amounts of Betula and Alnus (shrub birch and shrub alder) with sporadic occurrences of Larix during warm stages. The data suggest a gradual opening of the vegetation cover during this time period with a continuous but decreasing presence of larch in scattered woodland stands. The (supra-) regional pollen data thus indicate that the local vegetation we reconstructed for the Cherskiy Ovrag, consisting of grasslands in a mosaic with scattered woodlands, was widespread and existed over a long period of time. A hiatus in the pollen record comprised MIS 23.
MIS 22 around 890 kyr ago was a turning point in the West Beringian vegetation history as recorded in the El’gygytgyn core. Cold steppe biome scores in the pollen record significantly increased from this point in time onwards indicating accelerated aridification. The long-term cooling and aridification trend observed in the El’gygytgyn record corresponds with global ice-sheet expansion, sea-level lowering, coast-line shifts, shelf exposure and an intensified Siberian High and is considered additionally strengthened by the Tibetan Plateau uplift (Zhao et al., 2018).
The boundary between the lower and upper Olyorian, i.e., between the Chukochyan and Akanian was not yet exactly defined and is not preserved in a continuous geological record. But MIS 22 is a good candidate for this boundary as it represents the first high-amplitude glaciation that followed the 100 kyr periodicity. The switch from 41 to 100 kyr periodicity entailed long-lasting environmental changes resulting in the strengthening of aridification and the expansion of cold steppe vegetation and drove the evolution of cold and drought-resistant herbivores. The Chukochyan period therefore likely corresponds to the time prior to MIS 22. This phase was, according to the El’gygytgyn pollen record, characterized by a gradual expansion of grassland vegetation and the persistence of more or less scattered larch and shrub birch woodlands reflecting rather slow cooling and relative climatic stability. This relative stability of cold and dry climate over such a long period of time possibly resulted in the evolution of cold-adapted grazers and, eventually, of the mammoth steppe ecosystem.
The openness of vegetation is consequently not necessarily exclusively a function of climate in West Beringia but was probably also the result of megafaunal influence. The impact of the large herbivores on vegetation is documented in the macrofossil assemblage by plant taxa that are considered ruderal in modern vegetation, i.e., that occur at severely disturbed sites on nutrient-rich, compacted ground. Such taxa as Carex duriuscula, Chenopodium prostratum, Sibbaldianthe bifurca and Polygonum aviculare are resistant to overgrazing and trampling and are hardly competitive without regular disturbances of the plant cover, e.g., by herbivores. Steppe vegetation is fostered by the removal of woody plants due to decreasing shading and increasing insolation and evaporation. Steppes therefore replace woodlands in extremely continental areas at the middle courses of the Yana and Indigirka Rivers in Yakutia after wildfires (Yurtsev, 1982). In the Olyorian assemblage, the absence of any charcoal precludes that wildfire was the reason for the low percentage of trees and shrubs. Instead, the presence of nitrophilous pioneer plants such as Sibbaldianthe bifurca and Urtica dioica is rather an indication for zoogenic impact. Megaherbivores, especially proboscideans, are considered ecosystem engineers, which maintain vegetation openness, and in wooded landscapes, create mosaics of different structural types of vegetation with high habitat and species diversity (Johnson, 2009). Due to consumption of woody plants in winter and increased nutrient turnover, grasses and forbs are fostered by the presence of megaherbivores. The continuous existence of grassland vegetation and megaherbivores such as the steppe mammoth and, later, the evolution of the woolly mammoth in the study region demonstrates the importance of the unglaciated Arctic of Beringia as a cold laboratory for the evolution of the mammoth steppe biome – the ice-age ecosystem that, during the Middle and Late Pleistocene, spread over large proportions of the Northern hemisphere. Therefore, West Beringia can be regarded as the cradle of the mammoth steppe biome.
• A plant macrofossil assemblage from the Cherskiy Ovrag reveals the predominance of grassland vegetation composed of tundra steppes, meadow steppes and saline meadows in West Beringia during the Early Olyorian.
• The presence of larch and (shrub) birch as well as of dwarf shrubs, characteristic of the understorey of woodlands, indicates a mosaic-like interspersion of shrublands and larch groves in an altogether open landscape.
• Halotolerant plants indicate salt accumulation in the soil due to seasonally high evaporation.
• Nitrophytic ruderal plants suggest zoogenic impact on vegetation by nutrient enrichment, disturbances and compaction of the ground.
The original contributions presented in the study are included in the article/Supplementary Materials, further inquiries can be directed to the corresponding author.
SPD, as employee of the North East Science Station Cherskiy, continuously had access to the sections of the Cherskiy Ovrag over a long period of time, i.e., since 2001. He regularly studied the lithology and sedimentology of the sections, which were only temporarily and each time partially exposed and collected, over the years, numerous fossil mammal bones from both exposed units, the lower Olyorian (early Pleistocene) and the upper Yedoma (late Pleistocene). In the present paper, the description of topography, geology, lithology, geocryology and mammal remains recovered in the Cherskiy Ovrag base on the observations made by SPD in the course of his long-time monitoring. Sampling of organic-rich material was conducted by both authors, SPD and FK in 2015. FK conducted sieving of the sediment, screening of identifiable plant remains, identification of plant remains, their ecological interpretation, reconstruction of palaeo-vegetation and discussion of the results. FK wrote the article with contributions of SPD Figure 1, 3, 4 are made by FK Figure 2 bases on earlier publications of SPD and was adapted by both authors for the present paper.
The fieldwork was conducted with means from the Deutsche Forschungsgemeinschaft (DFG) in the frame of the research project TUNDRA-STEPPE (DFG project number 247453756) and was also partially conducted in the frame of the research project AAAA-A19-119030790003-1, Russia.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank Elena Troeva, Kseniia Ashastina, North Eastern Federal University Yakutsk (NEFU) and the Alfred Wegener Institute for Polar and Marine Resarch Potsdam (AWI) for help in preparing, conducting and logistics of the fieldworks as well as Nikita and Sergey Zimov for hospitality and inspiring discussions. We are grateful to the anonymous reviewers who helped us to improve the article.
Abaturov, B. D., Larionov, K. O., Kolesnikov, M. K., and Nikonova, O. A. (2005). State and Food Provision of Saigas on Pastures with Vegetation of Various Types. Zoologichesky Zhurnal 84 (3), 377–390.
Ashastina, K., Kuzmina, S., Rudaya, N., Troeva, E., Schoch, W. H., Römermann, C., et al. (2018). Woodlands and Steppes: Pleistocene Vegetation in Yakutia's Most continental Part Recorded in the Batagay Permafrost Sequence. Quat. Sci. Rev. 196, 38–61. doi:10.1016/j.quascirev.2018.07.032
Ashastina, K., Schirrmeister, L., Fuchs, M., and Kienast, F. (2017). Palaeoclimate Characteristics in interior Siberia of MIS 6-2: First Insights from the Batagay Permafrost Mega-Thaw Slump in the Yana Highlands. Clim. Past 13 (7), 795–818. doi:10.5194/cp-13-795-2017
Birks, H. H., and Birks, H. J. B. (2000). Future Uses of Pollen Analysis Must Include Plant Macrofossils. J. Biogeogr. 27 (1), 31–35. doi:10.1046/j.1365-2699.2000.00375.x
Boeskorov, G. G. (2019). To the Distribution and Taxonomy of a Fossil Soergelia, Soergelia Sp. (Caprinae, Bovidae, Artiodactyla, Mammalia), in Yakutia. Zoologichesky Zhurnal 98 (10), 1148–1155.
Campos, P. F., Sher, A., Mead, J. I., Tikhonov, A., Buckley, M., Collins, M., et al. (2010). Clarification of the Taxonomic Relationship of the Extant and Extinct Ovibovids, Ovibos, Praeovibos, Euceratherium and Bootherium. Quat. Sci. Rev. 29, 2123–2130. doi:10.1016/j.quascirev.2010.05.006
Chapin, F. S., Shaver, G. R., Giblin, A. E., Nadelhoffer, K. J., and Laundre, J. A. (1995). Responses of Arctic Tundra to Experimental and Observed Changes in Climate. Ecology 76, 694–711. doi:10.2307/1939337
Cirilli, O., Pandolfi, L., Rook, L., and Bernor, R. L. (2021). Evolution of Old World Equus and Origin of the Zebra-Ass Clade. Sci. Rep. 11 (1), 10156. doi:10.1038/s41598-021-89440-9
Clark, P. U., Archer, D., Pollard, D., Blum, J. D., Rial, J. A., Brovkin, V., et al. (2006). The Middle Pleistocene Transition: Characteristics, Mechanisms, and Implications for Long-Term Changes in Atmospheric pCO2. Quat. Sci. Rev. 25, 3150–3184. doi:10.1016/j.quascirev.2006.07.008
Cohen, K. M., and Gibbard, P. L. (2019). Global Chronostratigraphical Correlation Table for the Last 2.7 Million Years, Version 2019 QI-500. Quat. Int. 500, 20–31. doi:10.1016/j.quaint.2019.03.009
Davydov, S., Davydova, A., Schelchkova, M., Makarevich, R., Fyodorov-Davydov, D., Loranty, M., et al. (2020). Essential mineral Nutrients of the High-Latitude Steppe Vegetation and the Herbivores of mammoth Fauna. Quat. Sci. Rev. 228, 106073. doi:10.1016/j.quascirev.2019.106073
Davydov, S. P. (2007). “Features of Buried Fossil Fauna of mammoth Theriological Complex in Valleys of Low-Order Streams of the North East of Kolyma Lowland,” in Proceedings of the IV International Mammoth Conference, Yakutsk, June 18–22.
Davydov, S. P., Boeskorov, G. G., Sher, A. V., Bakulina, N. T., Davydova, A. I., and Schelchkova, M. V. (2009). “Mammoth Fauna Burial Places of the Northeast Kolyma Lowland Submontane Zone,” in Рroceedings of the International Conference Environmental Development of East Asia during the Pleistocene – Holocene (Boundaries, Factors, Stages of Human Mastering), Editor P.Y. Baklanov (Dal’nauka, Vladivostok, Russia), 49–51.
E.I. Troeva, A.P. Isaev, M.M. Cherosov, and N.S. Karpov (Editors) (2010). The Far North: Plant Biodiversity and Ecology of Yakutia (Dordrecht: Springer), 389.
Ellenberg, H. (1996). Vegetation Mitteleuropas mit den Alpen in ökologischer, dynamischer und historischer Sicht. 5th ed. Stuttgart: Eugen Ulmer, 1096.
Ermakov, N., Chytrý, M., and Valachovič, M. (2006). Vegetation of the Rock Outcrops and Screes in the forest-steppe and Steppe Belts of the Altai and Western Sayan Mts., Southern Siberia. phyto 36, 509–545. doi:10.1127/0340-269x/2006/0036-0509
Forstén, A. (1986). A Review of the Süssenborn Horses and the Origin of Equus Hydruntinus, Quartärpaläontologie, 6. Berlin, 43–52.
Giterman, R. E., Sher, A. V., and Matthews, J. V. (1982). “Comparison of the Development of Tundra-Steppe Environments in West and East Beringia: Pollen and Macrofossil Evidence from Key Sections,” in Paleoecology of Beringia. Editors D.M. Hopkins, J.V. Matthews-Jr., C.E. Schweger, and S.B. Young (New York: Academic Press), 43–73. doi:10.1016/b978-0-12-355860-2.50011-9
Guthrie, R. D. (1982). “Mammals of the mammoth Steppe as Paleoenvironmental Indicators,” in Paleoecology of Beringia. Editors D.M. Hopkins, J.V. Matthews-Jr., C.E. Schweger, and S.B. Young (New York: Academic Press), 307–326. doi:10.1016/b978-0-12-355860-2.50030-2
Johnson, C. N. (2009). Ecological Consequences of Late Quaternary Extinctions of Megafauna. Proc. R. Soc. B. 276, 2509–2519. doi:10.1098/rspb.2008.1921
Kahlke, R.-D. (1999). The History of the Origin, Evolution and Dispersal of the Late Pleistocene Mammuthus-Coelodonta Faunal Complex in Eurasia (Large Mammals). Hot Springs, South Dakota: Mammoth Site, 219.
Kahlke, R.-D., García, N., Kostopoulos, D. S., Lacombat, F., Lister, A. M., Mazza, P. P. A., et al. (2011). Western Palaearctic Palaeoenvironmental Conditions during the Early and Early Middle Pleistocene Inferred from Large Mammal Communities, and Implications for Hominin Dispersal in Europe. Quat. Sci. Rev. 30, 1368–1395. doi:10.1016/j.quascirev.2010.07.020
Kaplina, T. N., Kartashova, G. G., Nikitin, V. P., and Shilova, G. N. (1983). New Data on the Sand Unite of the Tuostakh Depression. Bulleten Komissii po izucheniyu chetvertichnogo perioda (Bulletin Quat. Commission) 52, 107–122. in Russian.
Kienast, F. (2016). Studies of Modern Vegetation and Sampling of Permafrost Deposits for Palaeobotanical Studies at the Lower Kolyma. Rep. Polar Mar. Res. 697, 87–160.
Kienast, F. (2013). “PLANT MACROFOSSIL RECORDS | Arctic Eurasia,” in The Encyclopedia of Quaternary Science. Editor S.A. Elias (Amsterdam: Elsevier), Vol. 3, 733–745. doi:10.1016/b978-0-444-53643-3.00213-2
Kienast, F., Schirrmeister, L., Siegert, C., and Tarasov, P. (2005). Palaeobotanical Evidence for Warm Summers in the East Siberian Arctic during the Last Cold Stage. Quat. Res. 63 (3), 283–300. doi:10.1016/j.yqres.2005.01.003
Kienast, F., Wetterich, S., Kuzmina, S., Schirrmeister, L., Andreev, A. A., Tarasov, P., et al. (2011). Palaeontological Records Indicate the Occurrence of Open Woodlands in a Dry Inland Climate at the Present-Day Arctic Coast in Western Beringia During the Last Interglacial. Quat. Sci. Rev. 30, 2134–2159.
Kozhevnikov, Y. P. (1981). “Botanical and Environmental Observations in the Kolyma Region at the Middle Course of the Berezovka River and the Cherskiy Settlement,” in Biology and Ecology of Plants of the Kolyma basin (Vladivostok: Far East Branch Acad. Sci. USSR), 99–117. in Russian.
Krestov, P. V., Ermakov, N. B., Osipov, S. V., and Nakamura, Y. (2009). Classification and Phytogeography of Larch Forests of Northeast Asia. Folia Geobot 44, 323–363. doi:10.1007/s12224-009-9049-6
Kucherov, I. B., and Daniëls, F. J. A. (2005). Vegetation of the Classes Carici-Kobresietea and Cleistogenetea Squarrosae in Central Chukotka. phyto 35 (4), 1019–1066. doi:10.1127/0340-269x/2005/0035-1019
Maksimovich, S. V. (1998). Ecology of Steppe Soils and Vegetation in Far Northeastern Yakutia (Tundra and forest-tundra Zone). Earth's Сryosphere 2 (2), 26–32. in Russian.
Mania, D., Mai, D. H., Seifert-Eulen, M., Thomae, M., and Altermann, M. (2010). The Special Environmental and Climatic Character of the Late Middle Pleistocene Interglacial of Neumark Nord (Geisel Valley). Hercynia N. F. 43, 203–256.
Murton, J. B., Edwards, M. E., Lozhkin, A. V., Anderson, P. M., Savvinov, G. N., Bakulina, N., et al. (2017). Preliminary Paleoenvironmental Analysis of Permafrost Deposits at Batagaika Megaslump, Yana Uplands, Northeast Siberia. Quat. Res. 87 (2), 314–330. doi:10.1017/qua.2016.15
Murton, J. B., Opel, T., Toms, P., Blinov, A., Fuchs, M., Wood, J., et al. (2021). A Multimethod Dating Study of Ancient Permafrost, Batagay Megaslump, East Siberia. Quat. Res., 1–22. doi:10.1017/qua.2021.27
Pisias, N. G., and Moore, T. C. (1981). The Evolution of Pleistocene Climate: A Time Series Approach. Earth Planet. Sci. Lett. 52, 450–458. doi:10.1016/0012-821x(81)90197-7
Romanovskii, N., Hubberten, H.-W., Gavrilov, A. V., Tumskoy, V. E., and Kholodov, A. L. (2004). Permafrost of the East Siberian Arctic Shelf and Coastal Lowlands. Quat. Sci. Rev. 23, 1359–1369. doi:10.1016/j.quascirev.2003.12.014
Russia’s Weather Server (2021). Weather of Russia. Available at: meteo.infospace.ru.
Sher, A. V. (1971). Mammals and Stratigraphy of the Pleistocene of the Far North-East of the USSR and North America. Moscow: Nauka. in Russian). In English: Pleistocene mammals and stratigraphy of the Far Northeast USSR and North America. International Geology Review 16 (7-10), 1-284.
Sher, A. V. (1986). Olyorian Land Mammal Age of Northeastern Siberia. Palaeontographia Italica 74, 97–112.
Sher, A. V., Kaplina, T. N., Giterman, R. E., Lozhkin, A. V., Arkhangelov, A. A., Kiselyov, S. V., et al. (1979). “Scientific Excursion on Problem “Late Cenozoic of the Kolyma Lowland”,” in XIV Pacific Science Congress (Moscow: Academy of Sciences USSR), 115.
Sher, A. V. (1997). “Late-Quaternary Extinction of Large Mammals in Northern Eurasia:A New Look at the Siberian Contribution,” Past and Future Rapid Environmental Changes. Editors B. Huntley, W. Cramer, AV Morgan, H C Prentice, and JRM Allen (New York, United States: Springer), 319–339. doi:10.1007/978-3-642-60599-4_25
Sher, A. V., Weinstock, J., Baryshnikov, G. F., Davydov, S. P., Boeskorov, G. G., Zazhigin, V. S., et al. (2011). The First Record of “Spelaeoid” Bears in Arctic Siberia. Quat. Sci. Rev. 30 (17-18), 2238–2249. doi:10.1016/j.quascirev.2010.10.016
Šrůtek, M., and Teckelmann, M. (1998). Review of Biology and Ecology of Urtica Dioica. Preslia, Praha 70, 1–19.
Tomirdiaro, S. V. (1980). Loess-ice Formation in Eastern Siberia during the Late Pleistocene and Holocene. Moscow: Nauka, 184.
van der Valk, T., Pečnerová, P., Díez-del-Molino, D., Bergström, A., Oppenheimer, J., Hartmann, S., et al. (2021). Million-year-old DNA Sheds Light on the Genomic History of Mammoths. Nature 591, 265–269. doi:10.1038/s41586-021-03224-9
Weber, C. A. (1914). Die Mammutflora von Borna. – Abhandlungen des Naturwissenschaftlichen Vereins zu Bremen 23, 1–69.
Yelovskaya, L. G., Konorovsky, A. K., and Savinov, D. D. (1966). Salt-Rich Soils above Permafrost (Kryosols) in Central Yakutia. Moscow: Nauka. In Russian.
Yurtsev, B. A. (1972). “Phytogeography of Northeastern Asia and the Problem of Transberingian Floristic Interrelations,” in Floristics and Paleofloristics and Asia and Eastern North America. Editor A. Graham (Amsterdam: Elsevier), 19–54.
Yurtsev, B. A. (1974). Steppe Communities of Chukotka Tundra and the Pleistocene “Tundra-steppe”. Botanicheskii Zhurnal 59 (4), 484–501. in Russian.
Yurtsev, B. A. (1982). “Relics of the Xerophyte Vegetation of Beringia in Northeastern Asia,” in Paleoecology of Beringia. Editors D.M. Hopkins, J.V. Matthews-Jr., C.E. Schweger, and S.B. Young (New York: Academic Press), 157–177. doi:10.1016/b978-0-12-355860-2.50018-1
Yurtsev, B. A. (2001). The Pleistocene "Tundra-Steppe" and the Productivity Paradox: the Landscape Approach. Quat. Sci. Rev. 20, 165–174. doi:10.1016/s0277-3791(00)00125-6
Zaslavskaya, T. M., and Petrovskii, V. V. (1994). Vascular Plants in the Environs of the Settlement of Cherskiy (North Yakutia). Botanicheskii Zhurnal 79 (2), 65–79. in Russian.
Zhao, W., Tarasov, P. E., Lozhkin, A. V., Anderson, P. M., Andreev, A. A., Korzun, J. A., et al. (2018). High-latitude Vegetation and Climate Changes during the Mid-pleistocene Transition Inferred from a Palynological Record from Lake El'gygytgyn, NE Russian Arctic. Boreas 47, 137–149. doi:10.1111/bor.12262
Keywords: olyorian, early-middle pleistocene transition, paleobotany, plant macrofossils, mammoth-steppe, megaherbivores, beringia, centre of origin
Citation: Kienast F and Davydov SP (2021) Forest Steppe-Like Vegetation Near Cherskiy (West Beringia) During the Early Pleistocene Olyorian Period Reconstructed Using Plant Macrofossils. Front. Earth Sci. 9:741473. doi: 10.3389/feart.2021.741473
Received: 14 July 2021; Accepted: 18 October 2021;
Published: 09 November 2021.
Edited by:
Alexandra Veremeeva, Institute of Physical-Chemical and Biological Problems in Soil Science (RAS), RussiaReviewed by:
Subir Bera, University of Calcutta, IndiaCopyright © 2021 Kienast and Davydov. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frank Kienast, ZnJhbmsua2llbmFzdEBzZW5ja2VuYmVyZy5kZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.