 Luiz Drude de Lacerda
Luiz Drude de Lacerda Rozane Valente Marins
Rozane Valente Marins Francisco José da Silva Dias
Francisco José da Silva Dias- 1Laboratório de Biogeoquímica Costeira, Instituto de Ciências do Mar (LABOMAR), Universidade Federal do Ceará, Fortaleza, Brazil
- 2Laboratório de Hidrodinâmica Costeira Estuarina e de Águas Interiores, Instituto de Ciências do Mar, Universidade Federal do Maranhão, Cidade Universitária, São Luís, Brazil
Global climate change interacts with regional alterations of land use in coastal basins and promotes rapid changes in the biogeochemistry of the coastal zone. These changes are generally stronger in extreme environments such as polar and semiarid regions. In these environments, biogeochemical changes result in a greater transfer of pollutants from the continent to the sea with resultant contamination of marine biota. In semiarid regions, there is a reduction in continental runoff and bulk pollutant transport, but paradoxically, there is an increase in the export of more bioavailable pollutants, resulting in higher contamination of the marine biota. In tropical latitudes, mangroves dominate coastal and estuarine environments and respond rapidly to environmental changes. The semiarid coast of Brazil is strongly affected by climate change. A major environmental response is altered hydrodynamics, generally involving decreasing continental runoff to the ocean, caused by reducing annual rainfall and damming of rivers. Also, strengthening of marine forcing due to heat accumulation in the South Atlantic Ocean and rising sea level pushes shelf and coastal waters into estuaries. Basin and oceanic processes undergo positive feedback, which causes an increase in the water residence time in estuaries, an extension of saline intrusion landward, accumulation of sediments in the inner reaches of estuaries, and the expansion of mangrove areas, particularly over the past 50 years. Expansion of mangroves means an expansion of sulfate reduction metabolism, which produces large amounts of dissolved organic carbon, characterized by a high capacity for forming organo-metallic complexes of high environmental significance. Dissolved and particulate Hg concentrations and fluxes are greater from the river to the estuary than from the estuary to the sea, producing an accumulation of particulate Hg in the estuary. Particulate Hg export may occur only during extremely rainy periods, and Hg is eventually deposited is shelf sediments, displaying low bioavailability. Export of dissolved, reactive, and DOC-bound Hg is practically nonexistent during rainy periods but increases by two orders of magnitude during the progressively longer dry periods. These highly bioavailable forms of Hg will enter food webs in the lower estuary and coastal areas, increasing contamination of the biota and human exposure to Hg.
Introduction
Mercury is a very toxic and persistent pollutant affecting the environment at a global scale through extensive accumulation in biota and enhancing human exposure through food consumption. Anthropogenic emissions of Hg are aggravated by significant natural emissions and remobilization processes of the deposited Hg in soils and sediments, and these are magnified by regional and global changes. As a result, widespread environmental and health issues have been largely associated with Hg pollution, resulting in international recognition of the relevance of understanding Hg remobilization processes and their response to global environmental changes (UNEP, 2019).
Mercury export from continents to the ocean depends on various drivers of natural and anthropogenic origin that are located along river basins and affect the river flows and watershed runoff. The final outflow to the sea also depends on the biogeochemical behavior of Hg along estuarine gradients (Leitch et al., 2007) and on processes taking place at the continental shelf–open ocean boundary (Fitzgerald et al., 2007). Apart from the relatively large natural environmental variability typical of the continent–ocean interface, changes in land use and global climate have been increasing or decreasing the Hg fluxes from continents to the sea dependent on location (Kopprio et al., 2015). However, there is a general consensus that extreme environments are more prone to be affected by such drivers, and the respective environmental responses are stronger and more conspicuous (Seneviratne et al., 2012; IPCC, 2019). For example, while average global temperature has increased by between 0° and 0.3° degrees per decade, the Arctic region has experienced an augmentation in average temperature between 0.5° to 0.8° degrees per decade (Macdonald et al., 2005). Similarly, drought conditions are becoming more extreme under already semiarid conditions, such as in northeastern Brazil, where an intensification in the number and duration of extended droughts from 1 to 3 per century during the 1,600 and 1,700 s to more than 14 in the 2,000 s have been recorded (Marengo et al., 2018). Therefore, more profound changes in Hg fluxes to the ocean are also expected to occur in such extreme environments.
During the past 25 years, a wide range of publications have noted an intensification of continent–ocean pollutant transfer to the Arctic due to environmental changes associated with global warming, almost twice as fast in the Arctic compared to other areas of the world. The major processes involved are the melting of glaciers and continental ice, causing increasing basin runoff (Dai et al., 2009) extended periods with soil temperatures >0°C and increasing Hg export from tundra, since the melting of permafrost increases the availability of organically bound Hg (Obrist et al., 2017), and increasing atmospheric deposition of Hg including atmospheric Hg depletion events (AMDE), which results in higher deposition, increasing the concentrations of total Hg in surface snow (Kirk et al., 2006; Carignan and Sone, 2010). Also of significance is the changing pattern of atmospheric Hg circulation over the Arctic, mostly associated with accelerating Hg oxidation in a warmer atmosphere (Stern et al., 2012). These processes are responsible for increasing pollutant fluxes into the ocean and have been associated with enhanced bioaccumulation in coastal food webs, whereas warming temperatures and reduced ice cover also trigger increased primary productivity, microbial activity and, as a consequence, methylation of Hg in sediments (Emmerton et al., 2013). This results in higher exposure to Hg, at least in some upper trophic-level species in the Arctic (Braune et al., 2005; Carrie et al., 2010; Schartup et al., 2015).
Along the semiarid coast of Northeastern Brazil, the already meager continental runoff of water is drastically decreasing. Formerly, this diminution was due to river damming for water consumption for a growing urban population and for irrigated agriculture (Marins et al., 2002a) followed by land use change, in particular conversion of gallery forests to pasture and agriculture; more recently, a steady decrease in annual rainfall has added its effect (Marengo et al., 2018; Alvalá et al., 2019). Offshore, increasing ocean forcing due to heat accumulation over the continental shelf and further into estuaries has been strongly affecting the hydrodynamics of these aquatic environments (Dias et al., 2011, 2013a, 2016, 2018). Taking the Arctic scenario, where increasing fluvial discharges drive elevated Hg concentrations in the Arctic biota (Schartup et al., 2015), as a template, one would expect a decrease of continent-derived pollutant transfer to the ocean due to decreasing continental runoff in semiarid NE Brazil and therefore, decreasing Hg concentrations in the regional aquatic biota. Paradoxically, however, the observed response of food webs in the NE Brazilian estuaries and continental shelf is increasing contamination by Hg, similar to what is being reported in Arctic biota, suggesting increasing fluxes and/or bioavailability of this toxic trace element. Therefore, decreasing continental runoff is causing an increase in Hg bioavailability, and in this study, we propose the mechanisms triggering higher Hg bioaccumulation. In the Arctic, as well as in the semiarid region of NE Brazil, the regional and global drivers responsible for increasing continental runoff to the Arctic Ocean and diminishing fluvial discharges to the Western Equatorial South Atlantic Ocean are strengthening due to global warming. Unfortunately, however, semiarid regions of the world are receiving much less attention from the scientific community than Arctic environments, notwithstanding the much larger human population inhabiting and using biological resources in semiarid regions. The present study reviews the results obtained in a 15-year-long research program in the Jaguaribe Estuary, in the semiarid NE Brazilian region. It discusses the biogeochemical processes involved with this apparent paradox, aiming to support mitigation and adaptation measurements to avoid an increase to unacceptable levels of the exposure risk to Hg of a population still largely dependent on a diet based on natural aquatic resources for its maintenance.
Climate and Hydrological Changes in Semiarid Basins of Brazil
Decreasing Annual Rainfall Due to Climate Change, Including Extended Drought
Whereas in higher latitudes continental runoff has increased in the past 50 years, in particular along the Canadian and Siberian basins draining into the Arctic ocean (Dai et al., 2009), in tropical and equatorial latitudes the reverse seems to occur, in particular under dry climates such as in NE Australia (Li et al., 2012) and in semiarid NE of Brazil. Castellanos et al. (2017) showed correlations between the increasing heat transfers from the Agulhas Current with increasing rainfall along a narrow stretch of the northeastern Brazilian coast. However, a few kilometers from the coast, annual rainfall drops to about 50% of the annual rainfall, and this reduction in precipitation has been observed over the semiarid northeastern region of Brazil by several studies. Moncunill (2006) reported a decrease in annual rainfall over Ceará State of about 6% per decade (5.3 mM year–1) between 1961 and 2003, with a few exceptions for areas where rainfall is dependent on topography. Salati et al. (2007) estimated a total annual rainfall decrease in the same region of about 11.6% between 1991 and 2004. In the southern portion of the semiarid region, Andrade et al. (2018) reported a decrease in annual rainfall in the past 35 years of 4.9 mM year–1, quite consistent with previous estimates. Godoy and Lacerda (2013) showed that out of 60 municipalities in the Jaguaribe River basin in Ceará State with a long historical annual rainfall record, 48 showed a negative trend in the past 50 years Supplementary Figure S1.
Based on historical registers of extended droughts (at least three consecutive years with annual rainfall below the historical mean) in the semiarid NE region of Brazil in the past 400 years, Marengo et al. (2018) observed a relationship with El Niño–Southern Oscillation (ENSO) through influencing the northward migration of the Inter-Tropical Convergence Zone (ITCZ), due also to the warming of the North Atlantic. Both large-scale phenomena that ultimately control rainfall and wind speed are strongly affected by global climate change (Maia et al., 2005). Their historical analysis suggests intensification in the frequency and duration of extended drought periods in the region, varying from 1 to 3 per century in the 1,600 and 1,700 s to over 14 in the 2,000 s (Marengo et al., 2018).
Decreasing Freshwater Fluxes and Fluvial Sediment Discharges to the Estuary
The impact of observed reductions in annual rainfall is maximized by regional environmental changes related to the water resource management of semiarid regions. At the Jaguaribe River basin proper, high water deficits are typical throughout most of the year, with annual rainfall ranging from 400 to 1,000 mM or lower, whereas annual potential evaporation can reach 2,500 mM. A major response to this water deficit is the construction of thousands of dams and artificial reservoirs, from innumerable small ponds in private farmlands to the three huge public reservoirs of 2 to 7 × 109 m3 of storage capacity. This storage capacity accumulates water during the short (3–4 months) rainy seasons to supply human demands during the long (8–9 months) dry periods (Medeiros et al., 2014). Apart from reservoirs, damming also diverts, through waterways and canals, a significant portion of the Jaguaribe river flow to the metropolitan basin of the State’s capital city of Fortaleza, reducing even further the freshwater discharge to the estuary. Prior to major dam construction and basin diversion, there was a covariance between rainfall and river flow. Afterward, there was a decoupling of these variables, and high river flow occurs only in very rainy years and generally depends on the opening of the dam’s gates (Godoy and Lacerda, 2013). The accumulated response to these multiple stressors on water resource availability is a decrease in the average freshwater discharge to the ocean through the Jaguaribe River, which varied from 60 to 130 m3.s–1 prior to the construction of major dams and diversion systems, to about 20 m3.s–1, mostly regulated and artificially controlled by the operation of major dams (Dias et al., 2013b).
Following the decreasing freshwater flow, sediment transport capacity has also been sharply affected by the annual precipitation decreases and the modification of watersheds. As a result, sediment trapping inside the middle estuary increased, enlarging river beaches and existing islands and sand banks and creating new fluvial islands (Lacerda et al., 2007). The lack of a significant continental sediment supply to the lower estuary resulted in the erosion of river margins and of littoral deposits, which helps in accelerating sediment accumulation inside the estuary (Godoy and Lacerda, 2013). In this scenario, the sedimentation-erosion equilibrium along the estuarine zone in most of the NE coast of Brazil is strongly affected, in particular under semiarid conditions (Jennerjahn et al., 2010; Medeiros et al., 2018).
Increased Marine Intrusion Into Estuaries
Increasing heat content in the Atlantic Ocean has been deduced from instrumental records that report this ocean to have warmed more intensively than any other ocean during the past 70 years (Sutton and Hodson, 2005; Lee et al., 2011). There is a consensus that the increase in the Meridional Heat Transport derives from an increase in advective heat that converges into the South Atlantic, mostly from the Indian Ocean, which contributes ∼70% of the total heat entering the South Atlantic through the Agulhas Current, suggesting that the warm water from the Indian Ocean plays a fundamental role in the northward water flow in the upper branch of the Atlantic Meridional Overturning Circulation (AMOC) (Dong et al., 2012).
The significance of the Agulhas leakage was elegantly demonstrated by modeling a scenario of a shutdown of this warm waterway, which resulted in an overall decrease in temperature and salinity in the South Atlantic, with anomalies in these variables propagating and finally reaching the equatorial belt within a decadal scale (Haarsma et al., 2011). As a consequence of the heat accumulation, climate change stressors, and sea surface temperature increase in particular, have resulted in a significant increase in the cumulative anthropogenic impact on the region’s ecosystems during this 70-years period that, apparently, has been accelerating in the past decade (Halpern et al., 2015).
More recently, Castellanos et al. (2017) concluded from a model considering an increase in the Agulhas leakage that the augmented transport may be related to higher latent heat flux observed along the northeastern coastline of Brazil since 2003. The increasing heat content in the South Atlantic has also been associated with ice melting in polar regions (Joughin et al., 2012), and since deep-ocean forcing alters momentum and buoyancy by currents at the shelf break (Weisberg and He, 2003), it also strengthens the forcing of central oceanic waters upon the continental shelf. This results in the choking of coastal and estuarine waters along the oriental Equatorial Atlantic as well as along the northeastern coast of Brazil (Dias et al., 2013a). Since continental shelves are buffer regions where estuarine and ocean waters mix, altering the quality and quantity of materials transported through this zone changes the chemical and physical properties of shelf waters in this region (Dias et al., 2016; Carvalho et al., 2017).
Dias et al. (2018) observed, through continuous monitoring of current and state properties by mooring instrumentation, that with the exception of unusual strong wet seasons when precipitation may be well above the historical average, ocean waters influenced by the direction of the velocity vectors between the middle and the outer shelf invade the continental shelf, moving coastal waters into estuaries and further inland, as observed in the Jaguaribe River estuary. In this area, under these typical conditions, water exchange between the estuary and the continental shelf involves <5% freshwater (Dias et al., 2011, 2013a) since the majority of the freshwater is blocked in the higher estuary. As a result, continental runoff from the river basin accumulates in the estuarine region rather than being exported to the continental shelf. Also, groundwater salinity is migrating further inland, as observed by radar mapping by Maia and Lima (2004). This extensive saline intrusion hampers the colonization of rivers banks by freshwater macrophytes, favoring their substitution by mangrove species.
In summary, taking into consideration the modeling exercises, which suggest a strengthening of the South Atlantic Ocean heat transfer patterns, and the observed results from local and regional studies, it seems reasonable to expect an increased choking of the continental runoff into estuarine areas, resulting in longer residence time of the fluvial inputs in the estuary prior to export to the continental shelf. Conditions under this scenario are highly maximized by decreasing annual rainfall and river fluxes, as discussed above.
Response of Estuarine Hydrodynamics and Sedimentation
Lacerda et al. (2013) summarized the hydrodynamics of the semiarid Jaguaribe River estuary in NE Brazil between 2005 and 2009, and Cavalcante (2019) expanded this database to 2012. The fluvial contribution reaching the upper estuary varies according to season and results in large variations in continental discharge to the coast. The volume of freshwater present in a given instant in the estuary varied seasonally from <5% during extreme drought periods to about 95% in extreme rainy years. There was also a varying proportion of the freshwater volume retained in the estuary (Dias et al., 2011). As a consequence, the residence time of waters from the basin’s runoff in the middle estuary also varied significantly between seasons. Under high freshwater inflow, shorter residence times of <1 day were typical at the upper and middle estuary, and freshwater fluxes to the ocean were maximal and mostly positive, even during high tide, as in the extreme 2009 rainy season. During rainy seasons, the proportion of freshwater in the total estuarine volume present in the middle estuary varied from 70% in normal precipitation years to 95% in exceptional extreme rainy years.
Water residence times in the middle estuary varied from 0.5 days in extreme rainfall seasons (>300 mM) to a maximum of 3 days in low-rainfall rainy seasons. For all rainy seasons monitored, the average residence time was 0.8 days. On the other hand, the water residence time in the middle estuary during the dry seasons was much longer and varied from 0.2 days in normal years to 13 days during extreme drought periods. For all dry seasons monitored during this study, the average residence time of the continental runoff waters at the middle estuary was 3.1 days. For comparison, Table 1 shows the average total fluxes and resulting water balance during dry periods in the Jaguaribe Estuary and a typical and similarly sized catchment in humid eastern Brazil, the Contas River, Bahia State (Dias et al., 2011).
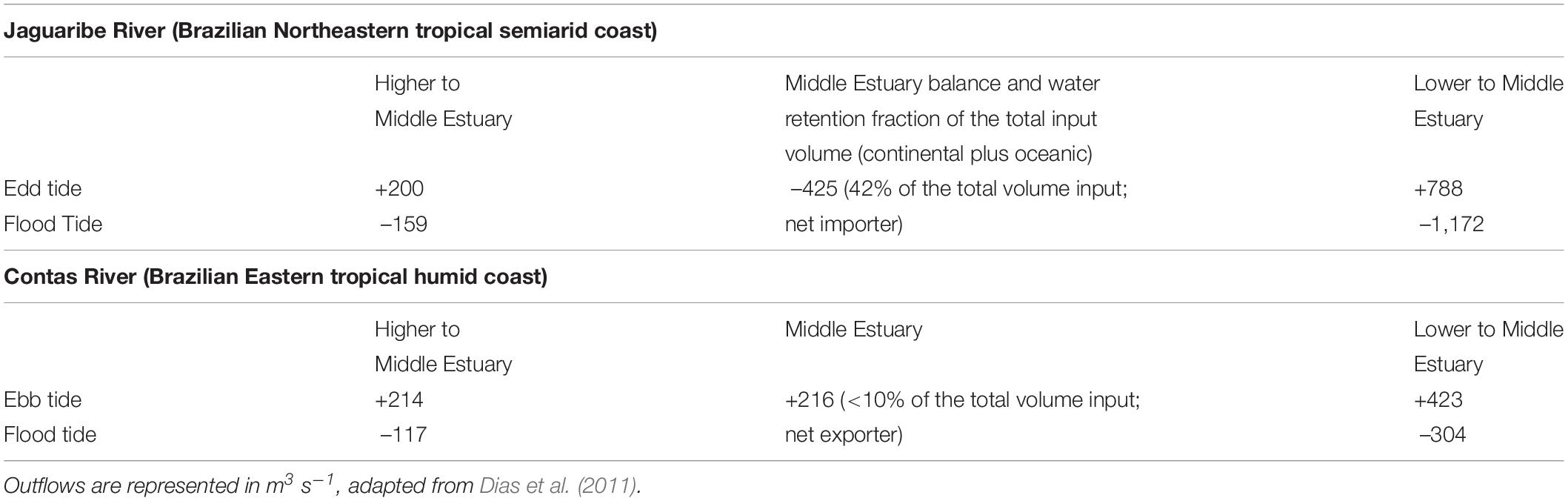
Table 1. Net water balance in the Jaguaribe and Contas Rivers during ebb and flood tides in dry periods from ADP data.
The net balance, meaning the difference between inputs and outputs of water volumes to an estuarine system, infers the system’s capacity for exporting or accumulating materials at the continent–ocean interface. The observed averages shown in Table 1 confirms a negative balance of water in the Jaguaribe River, where the middle estuary retains 42% of the water volume entering the system from the fluvial flux and the tides, acting as an importer of water.
This water retention is likely caused by the major influence of high tidal volumes and of a smaller input of freshwater from the upper estuary and the river proper. In contrast, the observed average inputs and outputs to the Contas River estuary characterize a positive balance, with 90% the water volume transferred to the middle estuary and then to the lower estuary, typically exporting water and materials to the sea. This exporting nature, which occurs both in ebb and flood periods, is caused by the larger freshwater inputs relative to the weaker influence of the smaller tidal contribution from a mesotidal regime.
Expansion of Mangroves: Increasing Mangrove Forest Cover
Increasing Mangrove Forest Cover
Recent reviews on mangrove responses to climate change highlight the main drivers of change in mangrove ecosystems to be changes in salinity, wave regime, increase in the frequency of extreme climatic events, and the quantity and quality of the sediment loading (Giri et al., 2011; Ward et al., 2016; Jennerjahn et al., 2017). Landward migration and increase in the area covered by these forests are the major responses of mangroves to these drivers, and the extension of such responses is also strongly influenced by the intensity of anthropogenic drivers, in particular river damming and water diversion. Mangroves displaying sediment accretion rates in pace with sea-level rise can adapt by migrating inland following saline intrusion, as shown in semi-arid coasts as in Brazil (Godoy and Lacerda, 2015), Arabian countries (Saderne et al., 2018), Mexico (López-Medellín et al., 2011), and Oceania (Alongi, 2015). Sediment input and sedimentation rates are frequently higher than current rates of sea-level rise, facilitating migration landward and allowing the mangrove forests to adjust to sea-level rise (Alongi, 2008; Lovelock et al., 2015). When sediment accumulation is similar to rising rates of sea level, forest distribution remains unaltered, and when the rate of sea-level rise exceeds the sediment accumulation, mangroves are eroded by mean high tides that are higher than the substrate elevation, in particular in low-lying limestone islands, such as in the Caribbean, where negligible continental runoff brings extremely small loads of allochthonous sediment (Ellison and Stoddart, 1991). Notwithstanding the potential capacity for adaptation, there may be, however, constraints to the extent of mangrove adaptation to climate change. Destruction of mangroves occurs in subsiding coasts, where deltaic islands are eroded by sea-level rise, such as in the Sundarbans (Alongi, 2015). Intensification of sand dune displacement, encroaching mangroves, has already been recorded in some arid and semiarid environments; this was related to climate changes (Maia et al., 2005; Lacerda, 2018) and poses a threat to mangrove resilience. Evidence from past Holocene sea-level rise due to post-glacial rebound, however, suggests that mangrove expansion can still occur under much higher rates of sea-level rise than those resulting from present climate change.
Along the semiarid coast of NE Brazil, mangroves are expanding over recently developed river bars and islands, a response to a decrease in rainfall and rising sea level, maximized by river damming and water diversion (Lacerda et al., 2007; Godoy and Lacerda, 2013, 2014, 2015; Godoy et al., 2018) Supplementary Figure S2. Maia et al. (2006) compared remote sensing images with map-based radar and aerial photographs from 1978 (Herz, 1991) and showed a mangrove area increase from 444 to 610 km2. Even considering the different methodologies and instrumentation, which probably overestimated this expansion by 30%, more recent comparisons using modern remote sensing techniques (Godoy et al., 2018) have confirmed this trend in mangrove expansion in northeastern Brazil (Nascimento et al., 2013) as well as for other areas, particularly under semiarid conditions (Doile et al., 2010) Supplementary Figure S4.
The expansion of mangroves is not a vegetation process only but is a creation of different biogeochemical realms, based on sulfate reduction over previous oxic areas. Under the new chemical state, preservation of carbon is favored, as are high production rates of dissolved organic matter (DOC) as a by-product of the incomplete oxidation of sedimentary organic matter by sulfate-reducing microorganisms (Kristensen et al., 2008). Sulfate-reducing bacteria and other prokaryotes are responsible for elevated Hg methylation rates in mangrove sediments, also favored by the high concentrations of ferrous ions in porewaters, intense burrowing crab activity and the unique architecture of the mangrove root system (Correia and Guimarães, 2016, 2017). This acts as a conveyor of highly bioavailable trace elements complexed with DOC or as organo-metallic species, such as methylmercury, from mangroves to the open estuarine areas and eventually to the continental shelf.
Biocheochemical Response and HG Availability
Increasing the Production of Young Organic Matter by Mangroves and Augmenting Dissolved Hg Concentrations and Fluxes
The fluorescence of the dissolved organic carbon (DOC) along the Jaguaribe Estuary shows striking seasonal differences. Fluorescence is highest in the wet months, the period with lower concentrations of DOC. During this season, fluorescence decreases only when it reaches the marine-influenced area Supplementary Figure S3. In contrast, in the dry season, fluorescence values are weaker along the entire estuary, and there is an increase in the fluorescence of peak A, which represents young organic matter, which occurs in the mangrove-influenced area of the estuary. This is clearly shown by the sharp increase in the Ia/Ic ratio from station 4 to 1 (Figure 1B). This suggests a higher production of young organic matter at the end of the fluvial end-member of the estuary, where mangroves abound, and its eventual flushing by tidal waters to the marine-influenced area (Mounier et al., 2018). The “younger” nature of the organic matter produced increases the metal complexing capacity of the DOC, as shown by the Hg speciation in mangrove pore waters (Figure 1A). This speciation supports the link between young DOC production from mangrove-dominated areas and increasing dissolved organic-bound Hg in pore waters in the upper layers of the sediment. This strongly suggests DOC as the major carrier of Hg out of the sediments to creeks and thence to the lower estuary and finally to the sea (Mounier et al., 2001).
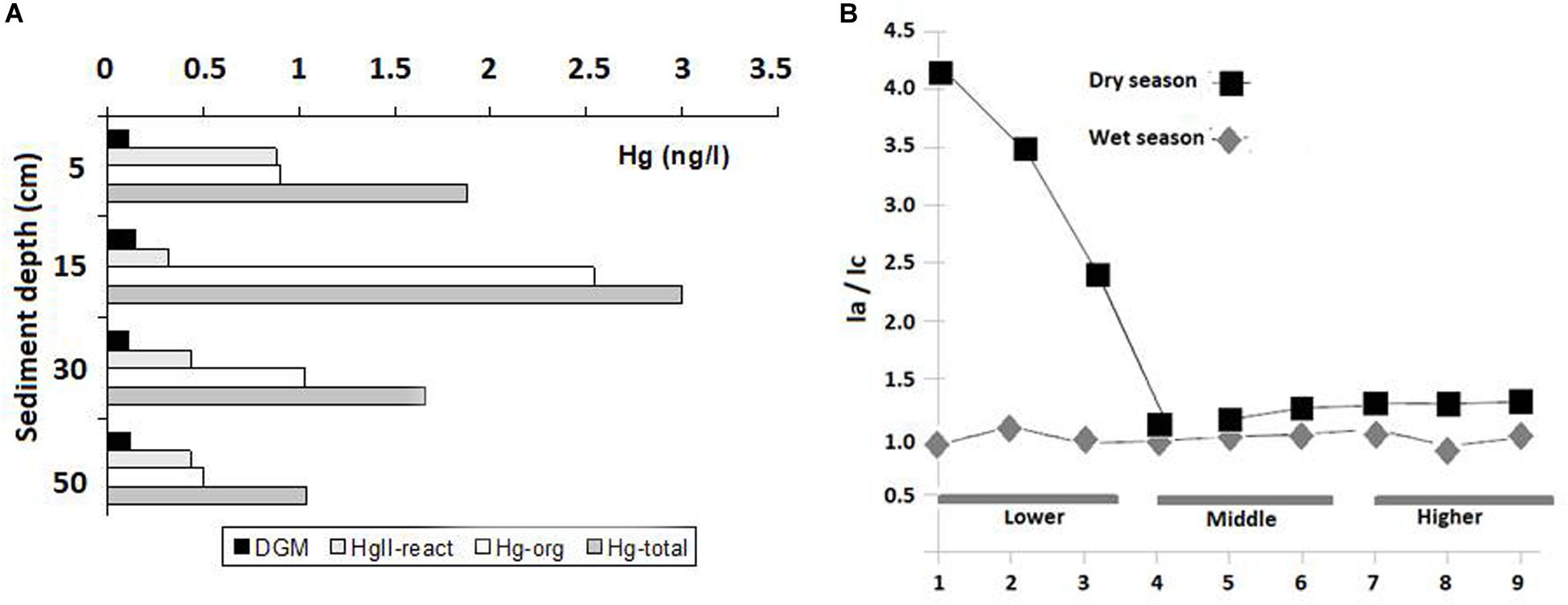
Figure 1. Changes in DOC fluorescence along the estuarine gradient of the Jaguaribe River. (A) Hg speciation in pore waters in a mangrove-dominated area showing increasing organic matter-Hg complex production, adapted from Mounier et al. (2001) and Marins et al. (1997). (B) Ia/Ic ratio matrices according to Coble (1996) and adapted from Mounier et al. (2018).
The seasonal enrichment in young organic matter produced in the mangrove-influenced area of the estuary supports a higher complexing capacity of DOC and increases Hg solubility. Dissolved Hg concentrations found in the lower estuary of the Jaguaribe River are higher during dry periods and dominate the Hg fluxes, whereas, in the wet season, particulate Hg is the major Hg fraction (Cavalcante, 2019). Concentrations measured during the past decade through 12 tidal cycles in the dry season varied from 1.2 to 12.3 ng.l–1, representing an average of 82% of the total Hg present in water. During the wet season, dissolved Hg concentrations measured during the same period through 16 tidal cycles are much lower, varying from <0.18 to 1.4 ng.l–1, representing an average of only 13% of the total Hg present in water. As a result, instantaneous dissolved Hg fluxes are highest in the dry season (average 0.57 mg.s–1; 89% of the total instantaneous Hg flux), whereas, during the wet season, dissolved Hg instantaneous fluxes averaged 0.03 mg.s–1, 2% of the total instantaneous Hg flux (Figures 2A,B).
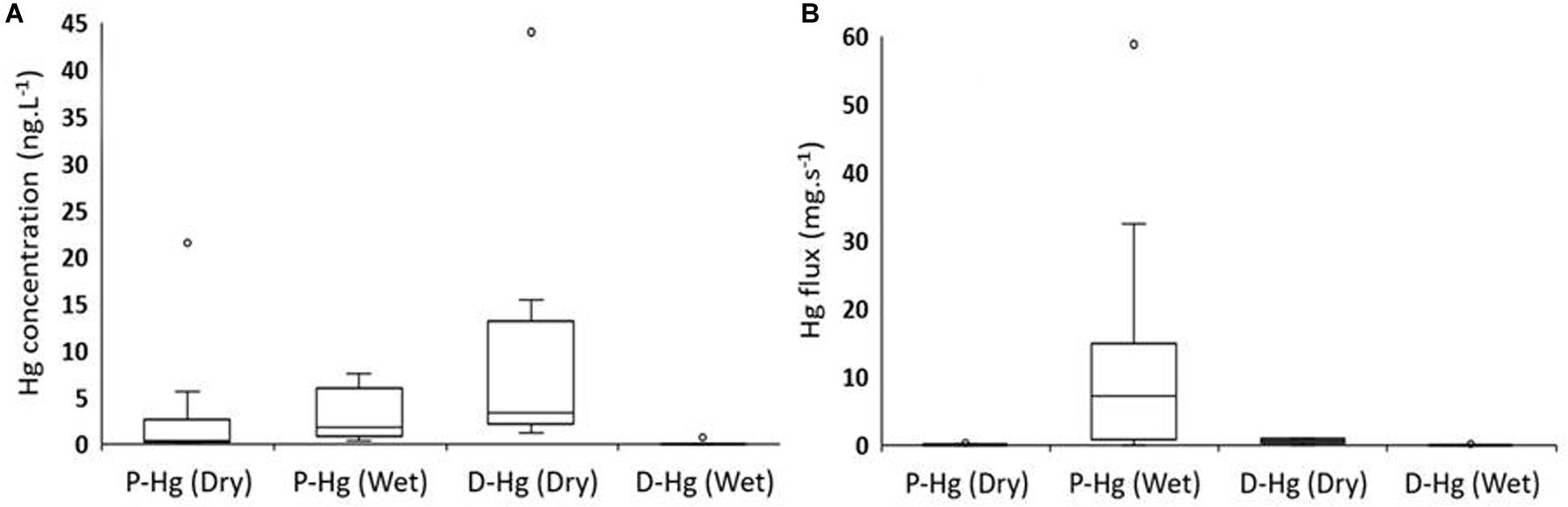
Figure 2. Seasonal variation of dissolved (D-Hg) and particulate (P-Hg) Hg concentrations (A) and instantaneous fluxes (B) in the Jaguaribe River estuary, NE Brazil, based on original data in Lacerda et al. (2013).
Other previous studies have already noted the association between mangrove-derived DOC and Hg export from mangroves. Bergamaschi et al. (2012) showed that a large production of dissolved Hg and Methyl-Hg occurs in the lower, mangrove-dominated portion of estuaries, which results in a tidally driven export of these dissolved Hg species. They have also shown that the variability in the total Hg load reaching the upper estuary has a small impact on the export of dissolved Hg species to the sea and that the dissolved Hg flux and the DOC fluxes to the sea are strongly associated. Lacerda et al. (2001) observed larger transport of total Hg entering mangroves during high tide and lower total Hg flux out of the mangrove but observed a larger export of dissolved, reactive Hg out of the system during low tide, suggesting in-situ production of reactive Hg species under the biogeochemical conditions present in mangroves. Marins et al. (2002b) measured a significant decrease in particulate Hg in the fluvial flow after passing mangrove areas but a simultaneous increase in the dissolved Hg fraction.
The production of Hg-DOC complexes in the mangrove environment in the lower Jaguaribe River estuary results in greater Hg export to the continental shelf, particularly during the dry season (Lacerda et al., 2013). Total Hg distribution in continental shelf surface water off the semiarid coast of Ceará State obtained from cruises during the dry seasons of 2010 and 2012 shows hot spots of Hg concentrations that are clearly associated with the Jaguaribe River Estuary, notwithstanding the absence of any significant anthropogenic Hg source (Lacerda et al., 2017; Lacerda and Marcovecchio, 2018). Concentrations in this area are similar, and sometimes even higher, than those observed in another enriched area off Fortaleza City, a major contributor of anthropogenic Hg from urban and industrial sources (Figure 3A). This spatial distribution pattern corroborated the greater dissolved Hg flux during the dry season.
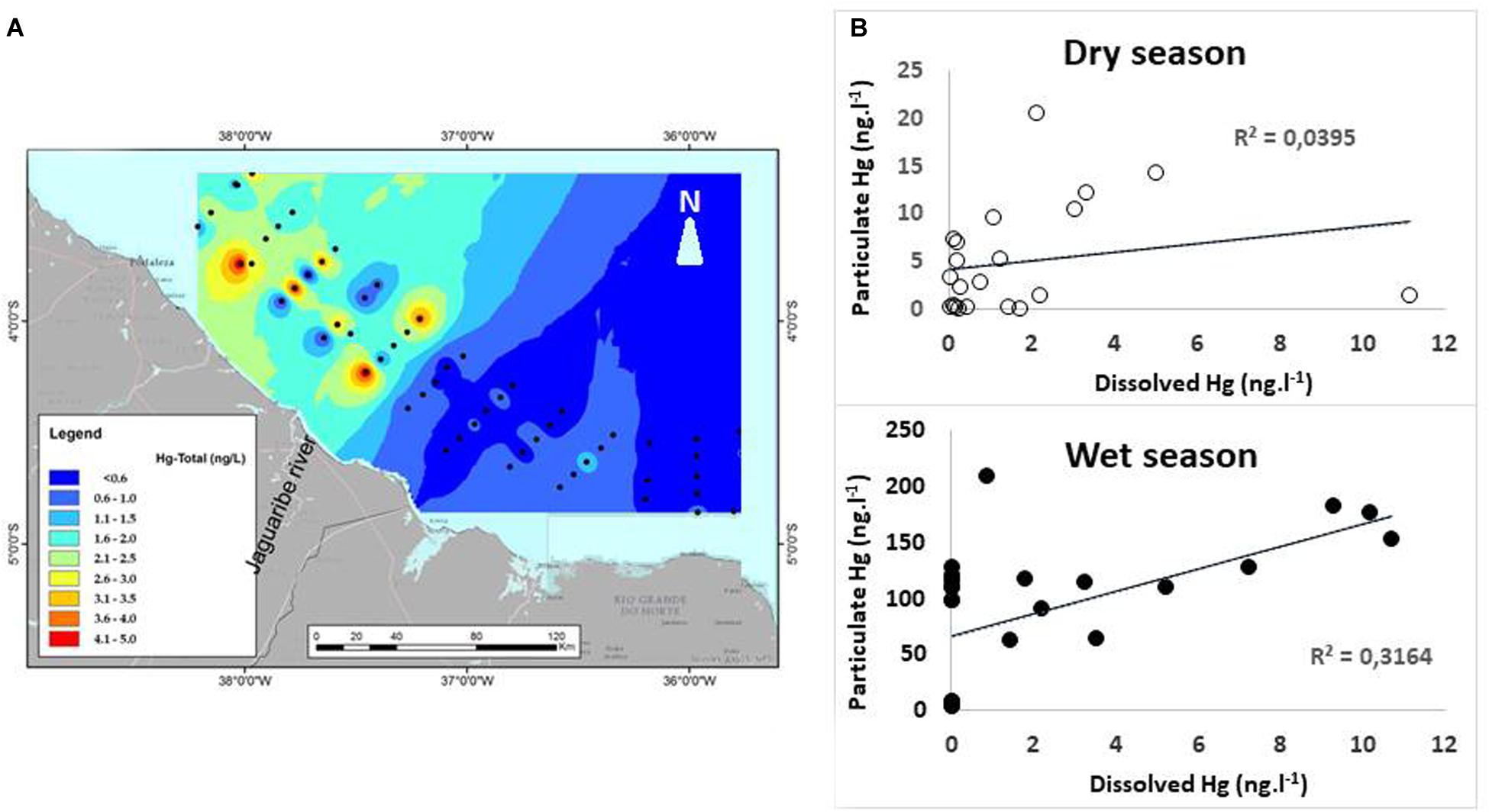
Figure 3. (A) Total Hg distribution in continental shelf surface water off the semiarid coast of NE Brazil. Hot spots of Hg concentrations are clearly shown off the Jaguaribe River Estuary, similar to the hot spot off Fortaleza City, a major contributor of anthropogenic Hg. (B) Relationship between dissolved and particulate Hg in waters off the Jaguaribe River in the dry and wet seasons.
The distribution of Hg in continental shelf waters off the Jaguaribe estuary obtained in the cruises in the dry and wet seasons shows a clear domain of particulate Hg in the wet season, similar to the pattern verified in the estuary, and dissolved Hg concentrations significantly correlated with the particulate Hg concentrations, suggesting that this fraction originates from desorption of Hg from the particulate flux. In the dry season, on the other hand, dissolved Hg concentration doubles and is independent of the particulate flux, suggesting direct export of dissolved species from the estuary (Figure 3B). If dissolved Hg concentrations are normalized by the particulate Hg concentrations, they are 15 times higher in the dry season than in the wet season.
These higher fluxes and concentrations of dissolved Hg in the dry season are a result of the longer residence time of water in the estuary during this season and of the output of the mangrove biogeochemistry based on sulfate reduction. Since residence time is inversely related to fluvial fluxes and Hg production is directly related to mangrove area, the observed response of these two drivers to global changes and watershed use strongly suggests an intensification of Hg bioavailability. Therefore, one may assume that there will be increasing Hg content in the biota and eventually human exposure to this pollutant.
Increasing Hg Availability Due to Complexation With DOC in the Lower Estuary Relative to Upstream Sections and Consequent Increase in Human Exposure
If the considerations above hold true, Hg concentrations in aquatic animals would be higher in individuals at the lower estuary than upstream in the watershed since they would respond to a higher bioavailability of dissolved Hg in water. Higher concentrations of dissolved Hg are observed in the lower estuary relative to the upper estuary, and this pattern is more extreme during the dry season. Many organisms of different taxa, trophic level, and food preferences occurring through the estuarine gradient respond to this high bioavailability. White shrimps, Litopanaeus vannamei, the blue crab Callinectes danae, and the fish Eugerres brasilianus, which respond to variations in dissolved Hg in water and Hg content in their diet, are present in higher concentrations in the lower estuary compared to the upper reaches, where fluvial influence is strongest (Figure 4A). This agrees with the dissolved Hg distribution along the estuary, confirming higher Hg bioavailability in the lower portion of the estuary associated with the large export of Hg-DOC complexes from mangrove areas. Additionally, an analysis of the Hg incorporation relative to the size of individual shrimps (L. vannamei) also showed an uptake rate of Hg that was 10 times faster relative to the weight gain of the animals in the lower estuary population compared to the population sampled at the fluvially dominated area (Moura and Lacerda, 2018), confirming higher rates of Hg bioaccumulation in the lower estuary relative to upstream regions.
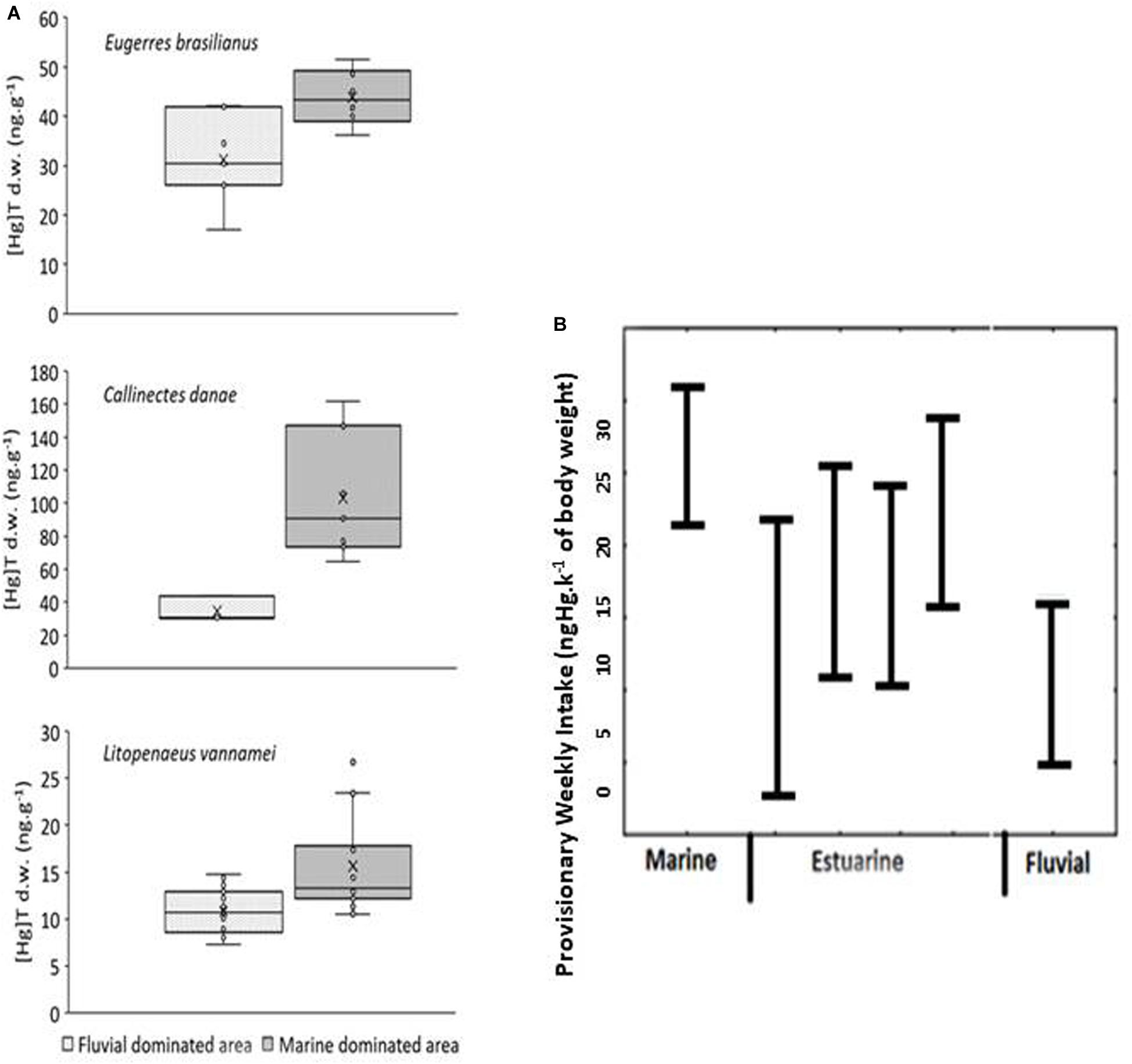
Figure 4. (A) Mercury distribution in aquatic organisms along the Jaguaribe River estuary, after Moura and Lacerda (2018). (B) Provisionary Hg Weekly Intake (ngHg.k– 1 of body weight) estimated for human communities resident along the Jaguaribe River estuary, after Costa and Lacerda (2014).
A recent study also reported an increase in Hg bioavailability and concentrations in aquatic organisms in the Paraíba do Sul estuary, a dry northern sector of the Rio de Janeiro coast in Southeast Brazil (Azevedo et al., 2018). The authors concluded that extended drought in recent years has promoted increases in total mercury and methylmercury concentrations in fish from the estuary and continental shelf.
A direct result of the observed increase in Hg availability and therefore Hg concentration in the aquatic biota is the increasing human exposure to this pollutant in populations consuming estuarine resources rather than fluvial ones (Figure 4B). Costa and Lacerda (2014) surveyed fish ingestion rates and major species consumed, simultaneously quantifying their Hg content along the Jaguaribe estuary. Their study reported 2- and 3-fold increases in the provisional weekly Hg intake in populations located along the marine-influenced area of the estuary relative to the middle estuary and the fluvially dominated region.
The high Hg bioavailability thus occurs under a scenario of reduced fluvial discharges, which promotes, by sedimentation, a rapid expansion of mangrove cover, increases the water residence time in the estuary, accumulates total Hg within the estuary and produces large amounts of Hg organic complexes. Therefore, decreased continental discharge increases rather than decreases Hg contamination, an apparent paradox when compared to what would be expected based on the observed results in the Arctic Ocean, where increased fluvial discharges result in increasing Hg fluxes to the ocean and bioaccumulation in the Arctic Ocean biota.
Conclusion
As in the Arctic Ocean, where increasing Hg content in the biota is linked to climate change linked to increasing continental runoff of Hg, in the Brazilian semiarid area, global climate change also results in increased Hg content in aquatic biota. However, the mechanisms are quite different, as it results from decreases in continental runoff through a sequence of events involving hydrodynamics and mangrove biogeochemistry. Figure 5 summarizes the major findings from the study of the Jaguaribe river estuary located in the semiarid coast of Northeastern Brazil. The main impacts of decreasing annual rainfall and strengthening oceanic forcing due to climate change and of diminishing fluvial flows resulting from basin alterations are increases in water residence times within the estuary, increasing saltwater intrusion and sedimentation. These hydrodynamic changes induce mangrove growth over new areas, expanding metabolism-based dissimilatory sulfate reduction to a larger portion of the estuary. There is an increase in DOC export as a result of the incomplete respiration of organic matter by anaerobes. DOC shows high complexing capacity for metals, in particular Hg, which forms strongly bound complexes with sulfidril groups.
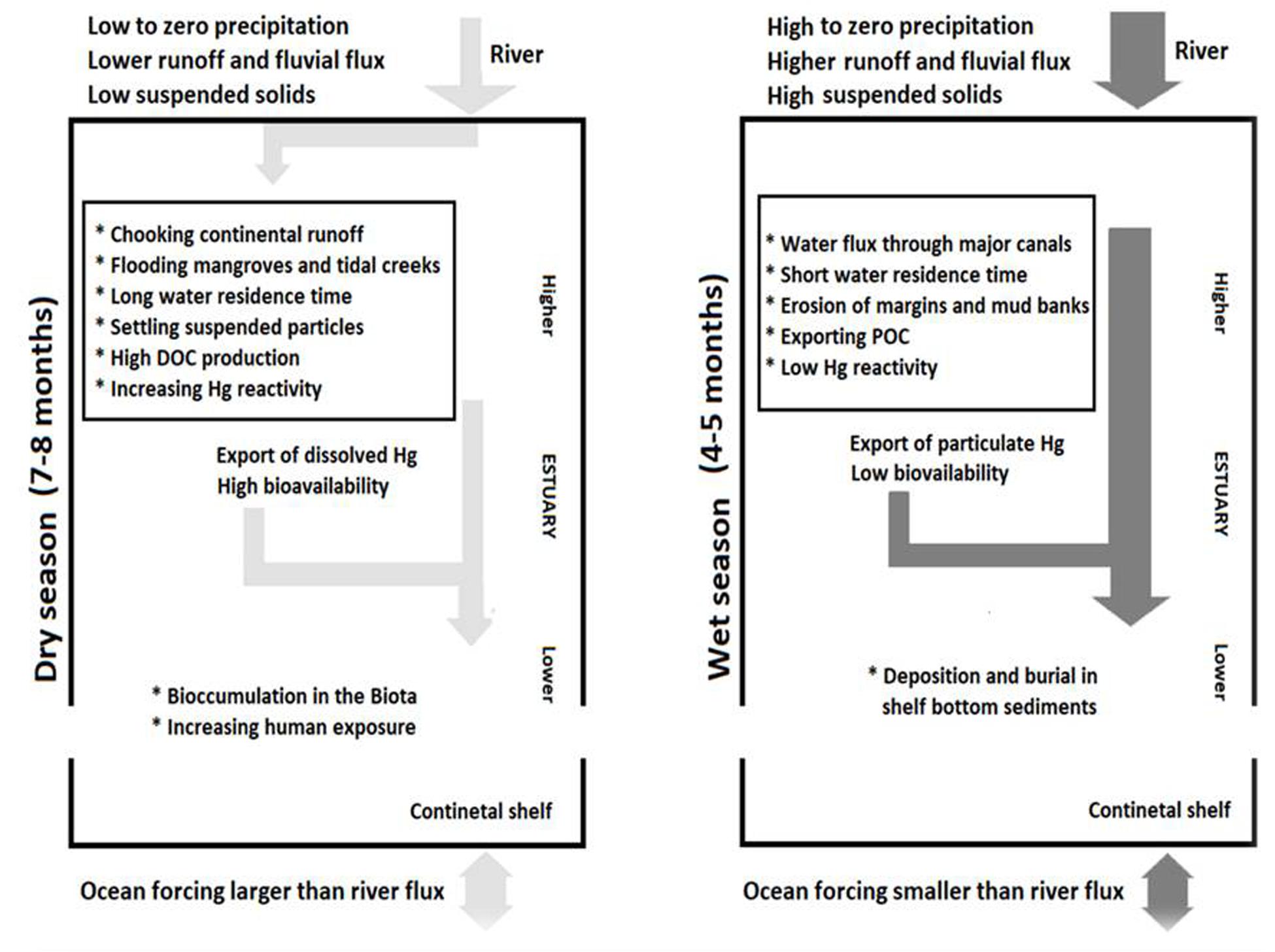
Figure 5. Hydrodynamics and biogeochemical model of the fate of Hg in the Jaguaribe River Estuary, NE Brazil.
Deposited Hg from continental origins accumulates and undergoes chemical changes instead of being rapidly exported in the particulate matter flux to the continental shelf. These biogeochemical changes mainly involve the formation of Hg-organic complexes with a higher bioavailability to aquatic food chains, including human populations exploiting them. The scenario described is most likely worsening, as global climate change will continue to cause negative anomalies in annual rainfall and thus reduce continental runoff even further into the semiarid NE region of Brazil. In addition, extended droughts are lasting longer, and heat accumulation is increasing in the South Atlantic. Human water uses will also expand, including fast-growing irrigated agriculture as well as urbanization, requiring more damming and diversion within the watersheds of the semiarid region. Therefore, Hg reactivity will possibly increase to higher levels in the estuary and eventually be exported as highly bioavailable Hg species to the adjacent estuarine waters and the coastal sea, increasing the Hg load of the local biota and consequently the Hg exposure risk of the local population.
Unfortunately, a link between global change and environmental Hg concentrations and human exposure, which is typically driven by multiple synergistic and antagonistic drivers operating simultaneously, remains uncertain and threatens the potential effectiveness of any eventual regulatory activities. However, taking into consideration the Precautionary Principle, as described by the Wingspread Consensus Statement of 1998, the observed processes composing this Arctic Paradox raise real and present threats of harm to human and environmental health. Therefore, precautionary measures should be taken even if some cause and effect relationships are not entirely established.
Data Availability Statement
All datasets generated for this study are included in article/Supplementary Material.
Author Contributions
LL conceived the ideas supporting this theory and wrote the major version of the manuscript. Most of the results discussed here are based on original research developed by RM on biogeochemical aspects of the Jaguaribe River estuary. FD provided the hydrological measurements and discussion on hydrodynamics. RM and FD reviewed and added to earlier versions of the manuscript.
Funding
This research was funded by Fundação Cearense de Apoio ao Desenvolvimento Científico e Tecnológico (FUNCAP) and Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank all members of the Coastal Biogeochemistry Laboratory (UFC/LABOMAR) for helping with the sampling and analysis. We thank Dr. Edmo Campos for discussion on the changes in the South Atlantic Ocean. We are especially thankful to Dr. Raymond Ward, U. Brighton, who kindly reviewed the final version of this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2020.00093/full#supplementary-material
References
Alongi, D. M. (2008). Mangrove forests: resilience, protection from tsunamis, and responses to global climate change. Estuar. Coast. Shelf Sci. 76, 1–13. doi: 10.1016/j.ecss.2007.08.024
Alongi, D. M. (2015). The impact of climate change on mangrove forests. Curr. Clim. Change Rep. 1, 30–39. doi: 10.1007/s40641-015-0002-x
Alvalá, R. C. S., Cunha, A. P., Brito, S. S. B., Seluchi, M. E., Marengo, J. A., and Moraes, O. L. L. (2019). Drought monitoring in the Brazilian Semiarid region. An. Acad. Bras. Ciênc. 91:e20170209. doi: 10.1590/0001-3765201720170209
Andrade, A. R. S., Godoy Neto, A. H., Cruz, A. F. S., Andrade, E. K. P., Santos, V. F., and Silva, T. N. P. (2018). Geostatistics applied to spatial variability and patterns in the temporal series of precipitation in Agreste of Pernambuco. J. Environ. Anal. Progr. 3, 126–145. doi: 10.24221/jeap.3.1.2018.1668.126-145
Azevedo, L. S., Pestana, I. A., Rocha, A. R. M., Meneguelli-Souza, A. C., Lima, C. A. I., Almeida, M. G., et al. (2018). Drought promotes increases in total mercury and methylmercury concentrations in fish from the lower Paraíba do Sul river, southeastern Brazil. Chemosphere 202, 483–490. doi: 10.1016/j.chemosphere.2018.03.059
Bergamaschi, B. A., Krabbenhoft, D. P., Aiken, G. R., Patino, E., Rumbold, D. G., and Orem, W. H. (2012). Tidally driven export of dissolved organic carbon, total mercury, and methylmercury from a mangrove-dominated estuary. Environ. Sci. Technol. 2012, 1371–1378. doi: 10.1021/es2029137
Braune, B. M., Outridge, P. M., Fisk, A. T., Muir, D. C. G., Helm, P. A., Hobbs, K., et al. (2005). Persistent organic pollutants and mercury in marine biota of the Canadian Arctic: an overview of spatial and temporal trends. Sci. Tot. Environ. 35, 4–56. doi: 10.1016/j.scitotenv.2004.10.034
Carignan, J., and Sone, J. (2010). The effect of atmospheric mercury depletion events on the net deposition flux around Hudson Bay, Canada. Atmos. Environ. 44, 4372–4379. doi: 10.1016/j.atmosenv.2010.07.052
Carrie, J., Wang, F., Sanei, H., Macdonald, R. W., Outridge, P. M., and Stern, G. A. (2010). Increasing contaminant burdens in an arctic fish, Burbot (Lota lota), in a warming climate. Environ. Sci. Technol. 44, 316–322. doi: 10.1021/es902582y
Carvalho, A. C. O., Marins, R. V., Dias, F. J. S., Rezende, C. E., Lefèvre, N., Cavalcante, M. S., et al. (2017). Air-sea CO2 fluxes for the Brazilian northeast continental shelf in a climatic transition region. J. Mar. Syst. 173, 70–80. doi: 10.1016/j.jmarsys.2017.04.009
Castellanos, P., Campos, E. J. D., Piera, J., Sato, O. T., and Silva Dias, M. A. F. (2017). Impacts of Agulhas leakage on the tropical Atlantic western boundary systems. J. Clim. 30, 6645–6659. doi: 10.1175/JCLI-D-15-0878.1
Cavalcante, M. S. (2019). Caracterização da Matéria Orgânica Natural no Estuário do Rio Jaguaribe em Diferentes Estações Climáticas em Anos de Seca Prolongada. Ph.D. thesis, Universidade Federal do Ceará, Fortaleza. Available online at: http://www.repositorio.ufc.br/handle/riufc/49622 (accessed December 12, 2018).
Coble, P. G. (1996). Characterization of marine and terrestrial DOM in seawater using excitation-emission matrix spectroscopy. Mar. Chem. 51, 325–346. doi: 10.1016/0304-4203(95)00062-3
Correia, R. R. S., and Guimarães, J. R. D. (2017). Mercury methylation and sulfate reduction rates in mangrove sediments, Rio de Janeiro, Brazil: the role of different microorganism consortia. Chemosphere 167, 438–443. doi: 10.1016/j.chemosphere.2016.09.153
Correia, R. S. S., and Guimarães, J. R. D. (2016). Impacts of crab bioturbation and local pollution on sulfate reduction, Hg distribution and methylation in mangrove sediments, Rio de Janeiro, Brazil. Mar. Pollut. Bull. 109, 453–460. doi: 10.1016/j.marpolbul.2016.05.028
Costa, B. G. B., and Lacerda, L. D. (2014). Mercury (Hg) in fish consumed by the local population of the Jaguaribe River Lower Basin, Northeast Brazil. Environ. Sci. Pollut. Res. 21, 13335–13341. doi: 10.1007/s11356-014-3297-6
Dai, A., Qian, T., and Trenberth, K. E. (2009). Changes in continental freshwater discharge from 1948 to 2004. J. Clim. 22, 2273–2792. doi: 10.1175/2008JCLI2592.1
Dias, F. J. S., Castro, B. M., and Lacerda, L. D. (2013a). Continental shelf water masses off the Jaguaribe River (4S), northeastern Brazil. Contin. Shelf Res. 66, 123–135. doi: 10.1016/j.csr.2013.06.005
Dias, F. J. S., Marins, R. V., and Maia, L. P. (2013b). Impact of drainage basin changes on suspended matter and particulate copper and zinc discharges to the ocean from the Jaguaribe River in the semiarid NE Brazilian coast. J. Coast. Res. 29, 1137–1145. doi: 10.2112/JCOASTRES-D-12-00115.1
Dias, F. J. S., Castro, B. M., and Lacerda, L. D. (2018). Tidal and low-frequency currents off the Jaguaribe River estuary (4oS, 37o4’W), northeastern Brazil. Ocean Dyn. 68, 967–985. doi: 10.1007/s10236-018-1172-6
Dias, F. J. S., Castro, B. M., Lacerda, L. D., Miranda, L. B., and Marins, R. V. (2016). Physical characteristics and discharges of suspended particulate matter at the continent-ocean interface in an estuary located in a semiarid region in northeastern Brazil. Estuar. Coast. Shelf Sci. 180, 258–274. doi: 10.1016/j.ecss.2016.08.006
Dias, F. J. S., Lacerda, L. D., Marins, R. V., and de Paula, F. C. F. (2011). Comparative analysis of rating curve and ADP estimates of instantaneous water discharge through estuaries in two contrasting Brazilian rivers. Hydrol. Proc. 25, 2188–2201. doi: 10.1002/hyp.7972
Doile, T. W., Krauss, K. W., Conner, W. H., and From, A. S. (2010). Predicting the retreat and migration of tidal forests along the northern Gulf of Mexico under sea-level rise. Forest Ecol. Manag. 259, 770–777. doi: 10.1016/j.foreco.2009.10.023
Dong, S., Garzol, S., and Baringer, M. (2012). The role of interocean exchanges on decadal variations of the meridional heat transport in the South Atlantic. J. Phys. Oceanogr. 41, 1498–1511. doi: 10.1175/2011JPO4549.1
Ellison, J. C., and Stoddart, D. R. (1991). Mangrove ecosystem collapse during predicted sea-level rise: holocene analogues and implications. J. Coast. Res. 7, 151–165.
Emmerton, C. A., Graydon, J. A., Gareis, J. A. L., St. Louis, V. L., Lesack, L. F. W., Banack, J. K. A., et al. (2013). Mercury export to the Arctic Ocean from the Mackenzie River, Canada. Environ. Sci. Technol. 47, 7644–7654. doi: 10.1021/es400715r
Fitzgerald, W. F., Lamborg, C. H., and Hammerschmidt, C. R. (2007). Marine biogeochemical cycling of mercury. Chem. Rev. 107, 641–662. doi: 10.1021/cr050353m
Giri, C., Ochieng, E., Tieszen, L. L., Zhu, Z., Singh, A., Loveland, T., et al. (2011). Status and distribution of mangrove forests of the world using earth observation satellite data. Global Ecol. Biogeogr. 20, 154–159. doi: 10.1111/j.1466-8238.2010.00584.x
Godoy, M. D. P., and Lacerda, L. D. (2013). Changes of estuarine islands and rainfall tendencies in the Jaguaribe river watershed – CE, Brazil. Arq. Ciênc. Mar. 46, 47–54. doi: 10.32360/acmar.v46i2.902
Godoy, M. D. P., and Lacerda, L. D. (2014). River-island response to land-use change within the Jaguaribe River, Brazil. J. Coast. Res. 30, 399–410. doi: 10.2112/JCOASTRES-D-13-00059.1
Godoy, M. D. P., and Lacerda, L. D. (2015). Mangroves response to climate change: a review of recent findings on mangrove extension and distribution. An. Acad. Brasil. Ciênc. 87, 651–667. doi: 10.1590/0001-3765201520150055
Godoy, M. D. P., Meireles, A. J. A., and Lacerda, L. D. (2018). Mangrove response to land use change in estuaries along the semiarid coast of ceará, Brazil. J. Coast. Res. 34, 524–533. doi: 10.2112/JCOASTRES-D-16-00138.1
Haarsma, R. J., Campos, E. J. D., Drijfhout, S., Hazeleger, W., and Severijns, C. (2011). Impacts of interruption of the Agulhas leakage on the tropical Atlantic in coupled ocean–atmosphere simulations. Clim. Dyn. 36, 989–1003. doi: 10.1007/s00382-009-0692-7
Halpern, B. S., Frazier, M., Potapenko, J., Casey, K. S., Koenig, K., Longo, C., et al. (2015). Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nat. Comm. 6:7615. doi: 10.1038/ncomms8615
Herz, R. (1991). Manguezais do Brasil. São Paulo: São Paulo. Inst. Oceanográfico, Universidade de São Paulo.
IPCC (2019). Summary for Policymakers, eds H.-O. Pörtner, D. C. Roberts, V. Masson-Delmotte, P. Zhai, M. Tignor, E. Poloczanska, et al. Geneva: IPCC.
Jennerjahn, T. C., Gilman, E., Krauss, K. W., Lacerda, L. D., Nordhaus, I., and Wolanski, E. (2017). “Mangrove ecosystems under climate change,” in Mangrove Ecosystems: A Global Biogeographic Perspective, eds V. H. Rivera-Monroy, S. Y. Lee, E. Kristensen, and R. R. Twilley (Berlin: Springer), 211–244. doi: 10.1111/gcb.12930
Jennerjahn, T. C., Knoppers, B. A., Cervalho, C. E. V., Mollenhauer, G., Hübner, M., and Ittekkot, V. (2010). “The tropical brazilian continental margin,” in Carbon and Nutrient Fluxes in Continental Margins: A Global Synthesis, eds K.-K. Liu, L. Atkinson, R. Quinones, and L. Talaue-McManus (Berlin: Springer), 427–442.
Joughin, I., Alley, R. B., and Holland, D. M. (2012). Ice-sheet response to oceanic forcing. Science 338, 1172–1176. doi: 10.1126/science.1226481
Kirk, J. L., St. Louis, V. L., and Sharp, M. J. (2006). Rapid reduction and reemission of mercury deposited into snowpacks during atmospheric mercury depletion events at Churchil, Manitoba, Canada. Environ. Sci. Technol. 40, 7790–7796. doi: 10.1021/es061299
Kopprio, G. A., Biancalana, F., Fricker, A., Cardona, J. E. G., Martines, A., and Lara, R. J. (2015). Global chage effects on biogeochemical process of Argentinian estuaries: an overlook of vulnerability and ecohydrological adaptative outlooks. Mar. Pollut. Bull. 91, 554–562. doi: 10.1016/j.marpollbul.2014.08.021
Kristensen, E., Bouillon, S., Dittmar, T., and Marchand, C. (2008). Organic carbon dynamics in mangrove ecosystems: a review. Aquat. Bot. 89, 201–219. doi: 10.1016/j.aquabot.2007.12.005
Lacerda, L. D. (2018). Burial of mangroves by mobile dunes: a climate change threat in semiarid coasts. ISME/GLOMIS Electr. J. 16, 6–10.
Lacerda, L. D., Dias, F. J. S., Marins, R. V., Soares, T. M., Godoy, J. M., and Godoy, M. L. D. P. (2013). Pluriannual watershed discharges of Hg into tropical semi-arid estuary of the Jaguaribe River, NE Brazil. J. Braz. Chem. Soc. 24, 1719–1731. doi: 10.5935/0103-5053.20130216
Lacerda, L. D., and Marcovecchio, J. E. (2018). “Continentderived metal pollutionthrough time: challenges of the global ocean,” in Marine Pollution and Climate Change, eds H. Andres, Arias, E. Jorge, and Marcovecchio (Boca Raton, FL: CRCPress), 99–117.
Lacerda, L. D., Marins, R. V., and Cavalcante, M. (2017). “Mercury mobilization due to global climate and regional land use changes in the Jaguaribe River Estuary, NE Brazil,” in Vulnerabilidad de las zonas Costeras de Latinoamérica al Cambio Climático, eds A. V. Botello, S. Villanueva, J. Gutiérrez, and J. L. Rojas Galaviz (Mexico: UJAT, UNAM, UAC), 333–344.
Lacerda, L. D., Menezes, M. O. T., and Molisani, M. M. (2007). Changes in mangrove extension at the Pacoti River estuary, CE, NE Brazil due to regional environmental changes between 1958 and 2004. Biota Neotropica 7, 1–6.
Lacerda, L. D., Silva, L. F. F., Marins, R. V., Mounier, S., Paraquetti, H. H. M., and Benaim, J. (2001). Dissolved mercury concentrations and reactivity in mangrove waters from the Itacurussá Experimental Forest, Sepetiba Bay, SE Brazil. Wet. Ecol. Manage. 9, 323–331. doi: 10.1023/A:1011868803439
Lee, S.-K., Park, W., van Sebille, E., Baringer, M. O., Wang, C., Enfield, D. B., et al. (2011). What caused the significant increase in Atlantic Ocean heat content since the mid-20th century? Geophys. Res. Let. 38:L17607. doi: 10.1029/2011GL048856
Leitch, D. R., Carrie, J., Lean, D., Macdonald, R. W., Stern, G. A., and Wang, F. (2007). The delivery of mercuryto the beaufort sea of the arctic ocean by the mackenzie river. Sci. Tot. Environ. 373, 178–195. doi: 10.1016/j.scitotenv.2006.10.041
Li, J., Fenga, J., and LicInt, Y. (2012). A possible cause of decreasing summer rainfall in northeast Australia. J. Climatol. 32, 995–1005. doi: 10.1002/joc.2328
López-Medellín, X., Ezcurra, E., González-Abraham, C., Hak, J., Santiago, L. S., and Sickman, J. A. (2011). Oceanographic anomalies and sea-level rise drive mangroves inland in the Pacific coast of Mexico. J. Veg. Sci. 22, 143–151. doi: 10.1111/j.1654-1103.2010.01232.x
Lovelock, C. E., Cahoon, D. R., Friess, D. A., Guntenspergen, G. R., Krauss, K. W., Reef, R., et al. (2015). The vulnerability of Indo-Pacific mangrove forests to sea-level rise. Nature 526, 559–563. doi: 10.1038/nature15538
Macdonald, R. W., Harner, T., and Fyfe, J. (2005). Recent climate change in the Canadian Arctic and its impact on contaminant pathways and interpretation of temporal trend data. Sci. Tot. Environ. 342, 5–86. doi: 10.1016/j.scitotenv.2004.12.059
Maia, L. P., Freire, G. S. S., and Lacerda, L. D. (2005). Accelerated dune migration and sand transport during El Niño events along the NE Brazilian coast. J. Coast. Res. 21, 1121–1126. doi: 10.2112/03-702A.1
Maia, L. P., Lacerda, L. D., Monteiro, L. H. U., and Souza, G. M. (2006). Atlas dos Manguezais do Nordeste do Brasil: Avaliação das Áreas de Manguezais dos Estados do Piauí, Ceará, Rio Grande do Norte, Paraíba e Pernambuco. Fortaleza: SEMACE.
Maia, L. P., and Lima, J. P. R. (2004). “Hydrogeochemical characterization of groundwater saline intrusion in the Western shore of Ceará, North-Eastern Brazil,” in Environmental Geochemistry in Tropical and Subtropical Environments, eds L. D. Lacerda, R. E. Santelli, E. K. Duursma, and J. J. Abrão (Heidelber: Springer), 277–291.
Marengo, J. A., Alves, L. M., Alvala, R. C. S., Cunha, A. P., Brito, S., and Moraes, O. L. L. (2018). Climatic characteristics of the 2010-2016 drought in the semiarid Northeast Brazil region. An. Acad. Bras. Ciênc. 90, 1973–1985. doi: 10.1590/0001-3765201720170206
Marins, R. V., Lacerda, L. D., Gonçalves, G. O., and Paiva, E. C. (1997). Effects of root metabolism on the post-depositional mobilization of mercury in salt marsh soils. Bull. Environ. Contam. Toxicol. 58, 733–738. doi: 10.1007/s001289900394
Marins, R. V., Lacerda, L. D., Mounier, S., Paraquetti, H. H. M., and Marques, W. S. (2002a). Caracterização hidroquímica, distribuição e especiação de mercúrio nos estuários dos Rios Ceará e Pacotí, Região Metropolitana de Fortaleza, Ceará, Brasil. Geochim. Brasil. 16, 37–48. doi: 10.21715/Gb.v16.i1.497
Marins, R. V., Freire, G. S. S., Maia, L. P., Lima, J. P. R., and Lacerda, L. D. (2002b). “Impacts of land based activities on the Ceará coast, NE Brazil,” in South American Basins: LOICZ Global Change Assessment and Synthesis of River Catchment – Coastal Sea Interaction and Human Dimensions, eds L. D. Lacerda, H. H. Kremer, B. Kjerfve, W. Salomons, J. I. Marshall-Crossland, and J. C. Crossland (Texel: LOICZ), 92–98.
Medeiros, P. H. A., Araújo, J. C., Mamede, G. L., Creutzfeldt, B., Güntner, A., and Bronstert, A. (2014). Connectivity of sediment transport in a semiarid environment: a synthesis for the Upper Jaguaribe Basin, Brazil. J. Soils Sed. 14, 1938–1948. doi: 10.1007/s11368-014-0988-z
Medeiros, P. R. P., Cavalcante, G. H., Melo, E. R., and Brandini, N. (2018). The São Francisco River (NE): review on the interannual loading of particulate matter suspended to the ocean and impacts of dams. Int. J. Hydrol. 2, 190–193. doi: 10.15406/ijh.2018.02.00067
Moncunill, D. F. (2006). “The rainfall trend over Ceará and its implications,” in Proceedings of 8th ICSHMO (Foz do Iguaçu: INPE), 315–323.
Mounier, S., Lacerda, L. D., Marins, R. V., and Benaim, J. (2001). Copper and mercury complexing capacity of organic matter from a mangrove mud flat environment (Sepetiba Bay, Brazil). Bull. Environ. Contam. Toxicol. 67, 519–525. doi: 10.1007/s001280154
Mounier, S. J., Lacerda, L. D., and Marins, R. V. (2018). “Determining the influence of urbanization onmangrove zones of Northeastern Brazil,” in Threats to Mangrove Forests, ed. C. Makowski (Berlin: Springer), 199–222.
Moura, V. L., and Lacerda, L. D. (2018). Contrasting mercury bioavailability in the marine and fluvial dominated areas of the Jaguaribe River, Ceará, Brazil. Bull. Environ. Contamin. Toxicol. 101, 49–54. doi: 10.1007/s00128-018-2368-7
Nascimento, W. R., Souza-Filho, P. W. M., Proisy, C., Lucas, R. M., and Rosenqvist, A. (2013). Mapping changes in the largest continuous Amazonian mangrove belt using object-based classification of multisensor satellite imagery. Estuar. Coast. Shelf Sci. 117, 83–93. doi: 10.1016/j.ecss.2012.10.005
Obrist, D., Agnan, Y., Jiskra, M., Olson, C. L., Colegrove, D., and Hueber, et al. (2017). Tundra uptake of atmospheric elemental mercury drives Arctic mercury pollution. Nature 547, 201–204. doi: 10.1038/nature22997
Saderne, V., Cusack, M., Almahasheer, H., Serrano, O., Masqué, P., Arias-Ortiz, A., et al. (2018). Accumulation of carbonates contributes to coastal vegetated ecosystems keeping pace with sea level rise in an arid region (Arabian Peninsula). J. Geophys. Res. Biogeosci. 123, 1498–1510. doi: 10.1029/2017JG004288
Salati, E., Campanhol, T., and Villa Nova, N. (eds).(2007). “Caracterização do clima atual e definição das alterações climáticas para o território brasileiro ao longo do Século XXI. Relatório 4,” in Tendências de Variações Climáticas para o Brasil no Século XX e Balanços Hídricos para Cenários Climáticos para o Século XXI (Brasília: Ministério do Meio Ambiente).
Schartup, A. T., Balcom, P. H., Soerensen, A. L., Gosnell, K. J., Calder, R. S. D., Mason, R. P., et al. (2015). Freshwater discharges drive high levels of methylmercury in Arctic marine biota. Proc. Nat. Acad. Sci. U.S.A. 112, 11789–11794. doi: 10.1073/pnas.1505541112
Seneviratne, S. I., Nicholls, N., Easterling, D., Goodess, C. M., Kanae, S., Kossin, J., et al. (2012). “Changes in climate extremes and their impacts on the natural physical environment,” in Managing the Risks of Extreme Events and Disasters to Advance Climate Change Adaptation. A Special Report of Working Groups I and II of the Intergovernmental Panel on Climate Change (IPCC), eds C. B. Field, V. Barros, T. F. Stocker, D. Qin, D. J. Dokken, and K. L. Ebi (Cambridge: Cambridge University Press), 109–230.
Stern, G. A., Macdonald, R. W., Outridge, P. M., Wilson, S., Chételat, J., Cole, A., et al. (2012). How does climate change influence arctic mercury? Sci. Total Environ. 414, 22–42. doi: 10.1016/j.scitotenv.2011.10.039
Sutton, R. T., and Hodson, D. L. R. (2005). Atlantic ocean forcing of north American and European summer. Science 309, 115–117. doi: 10.1126/science.1109496
UNEP (2019). Global Mercury Assessment. Geneva: United Nations Environmental Programme (Chemical and Health Branch).
Ward, R. D., Friess, D., Day, R., and Mackenzie, R. (2016). Impacts of climate change on global mangrove ecosystems: a regional comparison. Ecosyst. Health Sustain. 2, 1–25. doi: 10.1002/ehs2.1211
Keywords: climate change, semiarid, mangroves, mercury, human exposure
Citation: Lacerda LD, Marins RV and Dias FJS (2020) An Arctic Paradox: Response of Fluvial Hg Inputs and Bioavailability to Global Climate Change in an Extreme Coastal Environment. Front. Earth Sci. 8:93. doi: 10.3389/feart.2020.00093
Received: 17 April 2019; Accepted: 17 March 2020;
Published: 08 May 2020.
Edited by:
Alexandra V. Turchyn, University of Cambridge, United KingdomReviewed by:
Fernando Manuel Raposo Morgado, University of Aveiro, PortugalWim Salomons, Vrije Universiteit Amsterdam, Netherlands
Copyright © 2020 Lacerda, Marins and Dias. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luiz Drude de Lacerda, bGRydWRlQHBxLmNucHEuYnI=