 Calogero M. Santoro1
Calogero M. Santoro1 Eugenia M. Gayo2Chris Carter3Vivien G. Standen4
Eugenia M. Gayo2Chris Carter3Vivien G. Standen4 Victoria Castro5,6Daniela Valenzuela4Ricardo De Pol-Holz7
Victoria Castro5,6Daniela Valenzuela4Ricardo De Pol-Holz7 Pablo A. Marquet8,9,10
Pablo A. Marquet8,9,10 Claudio Latorre8,9,11*
Claudio Latorre8,9,11*- 1Laboratorio de Arqueología y Paleoambiente, Instituto de Alta Investigación, Universidad de Tarapacá, Arica, Chile
- 2Centro de Ciencia del Clima y la Resiliencia (CR)2 & Departamento de Oceanografía, Universidad de Concepción, Concepción, Chile
- 3School of Archaeology and Anthropology, Australian National University, Canberra, ACT, Australia
- 4Departamento de Antropología, Universidad de Tarapacá, Arica, Chile
- 5Departamento de Antropología, Universidad Alberto Hurtado, Santiago, Chile
- 6Departamento de Antropología, Universidad de Chile, Santiago, Chile
- 7GAIA Antártica, Universidad de Magallanes, Punta Arenas, Chile
- 8Departamento de Ecología, Facultad de Ciencias Biológicas, Pontificia Universidad Católica de Chile, Santiago, Chile
- 9Institute of Ecology and Biodiversity, Santiago, Chile
- 10Santa Fe Institute, Santa Fe, NM, United States
- 11Centro UC del Desierto de Atacama, Pontificia Universidad Católica de Chile, Santiago, Chile
The abundance of the southern Pacific mollusk loco (Concholepas concholepas), among other conspicuous marine supplies, are often cited as critical resources behind the long-term cultural and demographic fluctuations of prehistoric hunter-gatherers in the coastal Atacama Desert. These societies inhabited one of the world's most productive marine environments flanked by one the world's driest deserts. Both of these environments have witnessed significant ecological variation since people first colonized them at the end of the Pleistocene (c. 13,000 cal yr BP). Here, we examine the relationship between the relative abundance of shellfish (a staple resource) along a 9,500-year sequence of archeological shell midden accumulations at Caleta (a small inlet or cove) Vitor, with past demographic trends (established via summed probability distributions of radiocarbon ages) and technological innovations together with paleoceanographic data on past primary productivity. We find that shellfish extraction varied considerably from one cultural period to the next in terms of the number of species and their abundance, with diversity increasing during periods of regionally decreased productivity. Such shifts in consumption patterns are considered community based management decisions, and for the most part they were synchronous with large and unusual regional demographic fluctuations experienced by prehistoric coastal societies in northern Chile. When taken together with their technological innovations, our data illustrates how these human groups tailored their socio-cultural patterns to what were often abrupt and prolonged environmental changes throughout the Holocene.
Introduction
Sixteenth century written impressions of Spaniard conquerors state that the inhabitant of the coast of northern Chile's Atacama Desert were “barbarians,” “brutes,” or “retrograde” (Lozano Machuca, 1965[1581]). According to Lozano Machuca, they did not cultivate the land and maintained subordinate relationships with the “more advanced” groups of the interior. The Spaniards, however, admired the abundance of coastal marine resources (especially shellfish) and made detailed observations of the unique abilities these people had when taking advantage of their coastal environment through locally specialized technologies, despite the lack of fresh water (Polo de Ondegardo, 1916[1571]; Bibar, 1966[1558]; Bittmann, 1983; Lizárraga, 1987[1594-1608]). For instance, and cultural biases aside, written records describe the ingenious procedures used to make rafts from sea lion skins, crafts which persisted for centuries attracting comments from eighteenth-century naturalists (Frézier, 1713). Since these societal groups depended almost exclusively on coastal marine resources, these “rafts” can be viewed as part of a complex technological system, and were used until the nineteenth century in several northern Chilean ports for fishing, the transportation of goods and passengers, and to load saltpeter onto transoceanic clipper ships (Núñez, 1986a; Paez, 1986; Horta, 2015).
The former narrative has often led researchers to infer that these northern Chile prehistoric coastal peoples persisted throughout the Holocene in their well-established hunting-gathering mode of life (Arriaza et al., 2008; Marquet et al., 2012; Escobar, 2015; Flores et al., 2015; Salazar et al., 2015; Andrade et al., 2016; Castro et al., 2016). This agrees with analyses using stable carbon and nitrogen isotopes of human bones from Caleta Vitor and other coastal areas which have revealed a long-term pattern of food consumption based primarily on marine resources compared with terrestrial inputs (see also Olguín, 2014; Olguín et al., 2014; Pestle et al., 2015; Salazar et al., 2015 for the arheic coast of northern Chile), despite local environmental fluctuations and socio-cultural changes among inland groups that evolved into Formative (Neolithic) societies, with farming and pastoralism as key socio-economic factors (see Table 1 for regional cultural phases; Núñez and Santoro, 2011; McRostie, 2014; Muñoz et al., 2016; Núñez et al., 2017). A recent study also found that terrestrial animal resources were incorporated into other economic processes of production and consumption in different periods (Valenzuela et al., 2014). Yet, the evidence for such hypotheses of resilience or stability has yet to be evaluated in a systematic way, starting with the examination of the shell midden domestic remains often cited to back such claims.
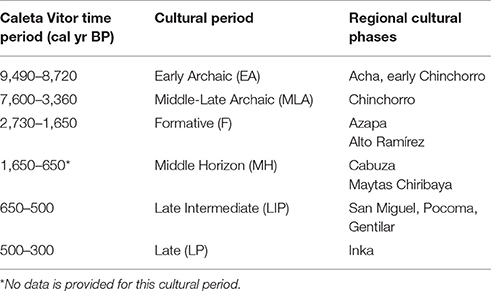
Table 1. Chronology, time period, and socio-cultural phases based on AMS dating from Caleta Vitor.
Stable carbon and nitrogen isotope analyses of human bones as well as other remains excavated at the Caleta Vitor archeological site show, as elsewhere along the Pacific coast of the Atacama Desert, that people maintained a marine diet that included shellfish, fish, sea birds, sea mammals, and seaweed since their arrival to the region (Petruzzelli et al., 2012; Poulson et al., 2013; Roberts et al., 2013; Pestle et al., 2015; Salazar et al., 2015; Bland et al., 2016; Standen et al., 2017). Ecofacts (or biofacts) found both in domestic and funerary contexts further support this assertion (Valenzuela et al., 2014). The exploitation of these resources was undertaken using a specialized tool-kit that included composite shell, bone, and cactus spine fishhooks, harpoons with lanceolate and rhomboidal lithic points, fishing nets, and prying tools among others. This marine resource extraction tool-kit improved notably over time since the late Pleistocene and throughout the Archaic (Figure 1A; Schiappacasse and Niemeyer, 1984; Standen, 2003; Standen et al., 2004; Arriaza et al., 2008; Flores et al., 2015; Reitz et al., 2015) as a consequence of the interaction between resource abundance, human population size, and technological innovations (Marquet et al., 2012; Derex and Boyd, 2015; Henrich et al., 2016; Acerbi et al., 2017).
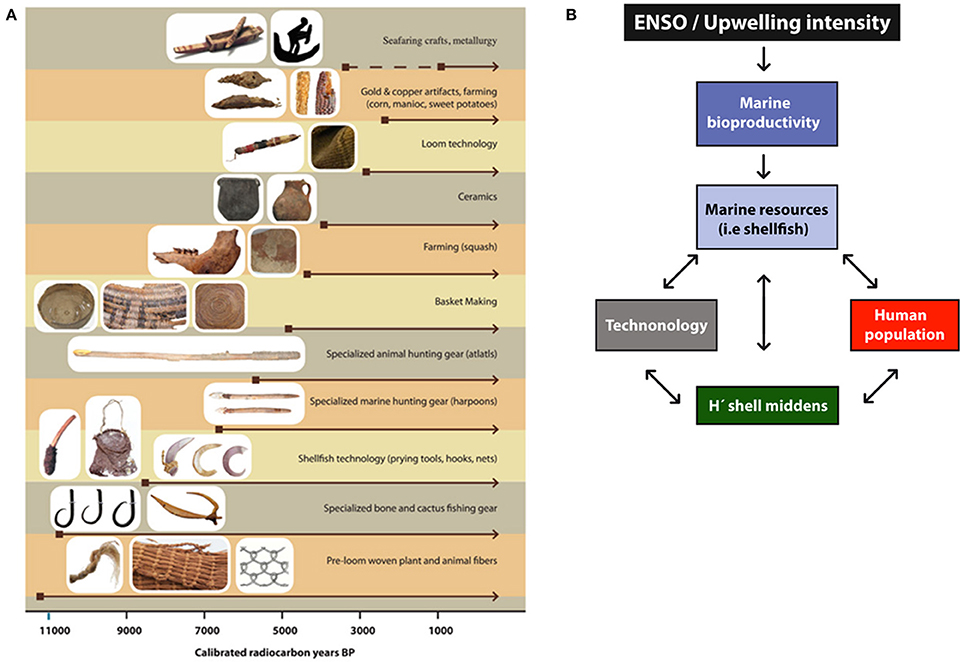
Figure 1. (A) Key technological innovations over the last 11,000 years associated with the extraction of coastal resources in northern Chile. (B) A “cascade effect” diagram showing the relationship between El Niño-Southern Oscillation (ENSO), coastal upwelling and marine bioproductivity and how these impact coastal human populations, technological innovations and resource extraction (measured using a diversity index such as H′).
Although shellfish alone have insufficient nutritional value for sustaining a population (Bailey, 1975; Schiappacasse and Niemeyer, 1984; Brown et al., 2011; Salazar et al., 2015), Concholepas concholepas (loco) the most preferred shellfish is a good source of nourishment (120 calories per 100 g) and as such is expected to be a fundamental part of the diet (Stephens and Krebs, 1986). Under certain cultural or environmental circumstances, such a protein-rich resource could play a key role in the sustainability of human populations (Erlandson, 1988). This is particularly true for the coastal Atacama Desert, where shellfish are abundant, easy to catch as they live in large aggregations either on tidally exposed rocks or sandy beaches, and are not subject to seasonal variations, although when affected by warm El Niño events, new species show up in replacement within a short period of time. This is in stark contrast with the Atacama Desert inland resources (plant and animals), which were scarce, highly unpredictable, and spread out over such large areas that people were almost completely circumscribed to coastal habitats until the late Holocene (Grosjean et al., 2007; Gayo et al., 2012, 2015; Santoro et al., 2017). Moreover, inland camps were maintained by seafood transported from the coast up to 20–30 km (Núñez, 1986b). Marine resources were consumed year-round and the mollusks consumed yielded tons of shells that today constitute the most visible ecofacts and make up the bulk component of shell middens, ubiquitous features along the coast of northern Chile, and elsewhere (Bird, 1943; Campbell, 2005; Croes, 2015; Salazar et al., 2015; Latorre et al., 2017). Shell diversity can thus be used as bio-indicators to explore continuities and changes of human preferences of seafood consumption over long periods of time.
External environmental factors as well as cultural ones can drive shellfish utilization diversity over time in coastal shell middens. El Niño—Southern Oscillation (ENSO) is an important factor for understanding marine bioproductivity and biogeochemical variations along the Pacific coast of South America (Chavez et al., 1999). La Niña (El Niño) events are characterized by increased (subdued) upwelling intensity of nutrient-rich-oxygen poor waters; conditions that enable augmented (diminished) rate of biomass production (autotrophs and heterotrophs) in the marine ecosystem (e.g., Thiel et al., 2007; Masotti et al., 2011). ENSO coupled with changes in coastal upwelling could play a major role in explaining past shell midden assemblage diversity, along with local human population size and extraction technologies (Figure 1B).
In this work, we explore the link between dietary preferences and marine bioproductivity by analyzing variations in shellfish assemblages present in Caleta Vitor shell middens in northernmost Chile. We examine long-term variations in the selection of mollusks consumed, by quantifying and comparing the diversity of taxa through time and compare these data to a proxy for changes in past ocean primary productivity (increased vs. subdued). We discuss the relationship between changes in the dietary preferences of human-collected shellfish, paleodemographic trends, and paleoclimate.
Materials and Methods
Caleta Vitor is a small inlet located at the mouth of Quebrada Chaca ~30 km south of the city of Arica (18°45′09.94″S, 70°20′08.65″W; Figure 2). Quebrada Chaca is one of the few major canyons that reach the Pacific coast of the Atacama Desert of northern Chile (Standen et al., 2004; Núñez and Santoro, 2011; Roberts et al., 2013; Santoro et al., in press). The canyon is narrow and very steep-sided, and presently has an intermittent stream that originates in the Andes to the east that becomes highly incised (>1 km) into the Coastal Cordillera as it reaches the sea (Hoke et al., 2004). Vegetation is limited to either the valley floor, the areas adjacent to the streambed, or to a phreatic wetland (presently dry) at the stream's outlet. The coastline is bounded by high cliffs (700–1,000 m a.s.l.) and access on foot along the shore from either the north or the south is very difficult.
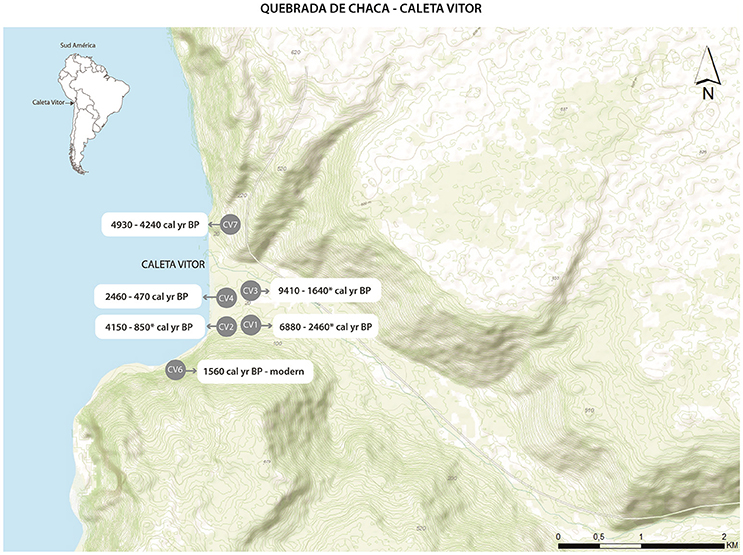
Figure 2. Map of the Caleta Vitor archeological area, showing the major shell midden deposits (in gray and numbered CV1, CV2, etc.) along with associated radiocarbon date age ranges. All ages in calibrated 14C years BP.
Caleta Vitor is an archeological area that comprises several sites of distinct functionality, characteristics, and extension, such as shell middens, rockshelters, rock paintings, tumulus, and burials. This area was selected for this study as it spans over 9,000 years of prehistoric occupation and offers well-dated archeological records (Table 2; Figure 2). Neither has it been affected by urban development, which makes it a superb study area for our ongoing interdisciplinary research into the prehistory of the Atacama Desert (Roberts et al., 2013; Swift et al., 2015; Bland et al., 2016; Disspain et al., 2016; Latorre et al., 2017). Archaeological sites at Caleta Vitor are extensive and cover an area of at least 525,000 m2. Surface surveys have outlined seven sectors delimited by activity zones and concentrations of homogeneous surface archeological materials. Specific topographic features were also considered when defining the boundaries of these sub-divisions, which are classified with the prefix “CV” and numbered from one to seven (Figure 2). The extensive shell middens at Caleta Vitor contain abundant shellfish and fish left behind by fishermen, hunters, and gatherers that span at least the last 9,500 cal yr BP until the Spanish Colonial and even modern era (Figure 2). CV5, a badly preserved cave site, was not considered in this study. A total of 70 radiocarbon dates on a variety of different elements (i.e., plant material, shell, human bone, charcoal), selected both in situ or from samples in the laboratory comprise the chronology of the six studied CV archeological sites that have sequential occupational histories (Table 2). These dates show that most of these middens accumulated episodically over time and post-depositional alteration was limited in extent (see Latorre et al., 2017 for a recent discussion of shell midden formation at Caleta Vitor).
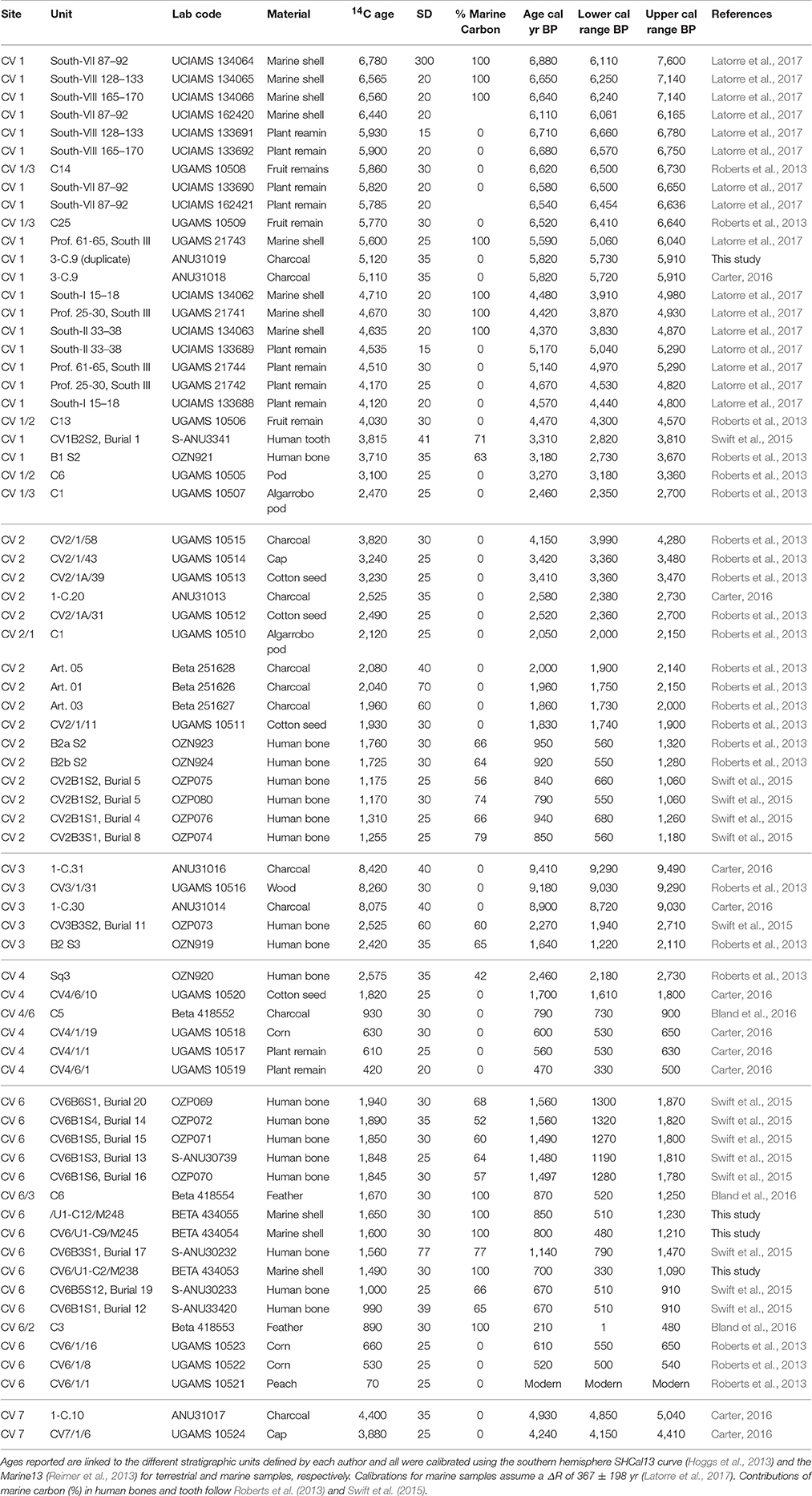
Table 2. Compilation of 70 AMS dates from six archeological areas defined at Caleta Vitor (CV).
The data for this study comes from a total of 12 trenches (5.5 m2 of logged sections) excavated in 2010. Excavations were undertaken by hand, following stratigraphic units in most cases. Such units were generally defined by changes in soil color, density, texture, and/or composition. Trenches CV4/2, CV4/3, CV4/6, and CV6/2 did not display clear stratigraphic divisions and were excavated in arbitrary 100 mm- or 50 mm-thick spits. Excavated material was sieved through a 1.7 mm screen. A full range of artifacts and ecofacts were sorted, including fish (vertebrae, otoliths from eight species, dentition, mixed undifferentiated bone), crabs (mixed shell remains), shells (17 species of mollusks), echinoderms, tunicates (Pyura chilensis), bones (mammals and birds), ceramics (fragments only), plant remains, stone artifacts, and dung (of both camelid and rodent origin). A total of 118.3 kg of shells were recovered from 181 (91.4%) of the 198 excavated stratigraphic units—those without shells consisted of large rocks, overburden, or sterile infill (Carter, 2016).
Much of the assemblage was highly fragmented due to both initial processing and post-depositional taphonomic processes. Shells were then weighed following Glassow (2000). Usage of both weights and MNIs has attracted criticism with discussion mostly focused on the validity of such approaches in providing unambiguous measures for the representation of taxa within a given assemblage (see Waselkov, 1987; Erlandson, 1994; Mason et al., 1998; Álvarez et al., 2011; Faulkner, 2011; Habu et al., 2011; Rick and Waselkov, 2015). Glassow, however, stated that “neither MNI nor weight proportions, is a universal unit of analysis for addressing the various questions about human predation on shellfish that archeologists frequently ask” (Glassow, 2000, p. 412). This is particularly relevant when comparing numbers of robust shells (such as C. concholepas—loco) with lighter shelled animals (such as Perumytilus purpuratus—chorito). Glassow (2000) has also argued that both can be used to complement each other in situations where results can be skewed, whereas Mason et al. (2000, p. 760) stated “MNI and weight yielded different results but did not demonstrate that one set of results was ‘better’ than the other.”
Most of the stratigraphic units contain highly fragmented shells, however, so the identification of non-repetitive elements (NREs) for most specimens was not possible. This implies that MNI-based measures could be overestimating the abundance of robust shells over more fragile shell species. Hence, we focused our analyses in gross weight per species (hereafter biomass in grams) to quantify variations in the abundance of shellfish through time. Taxonomic classification was possible even for tiny fragments of the different shells with conspicuous features facilitating identification to the genus or species level (e.g., fragile shelled sea urchins are easily recognizable). Two additional reasons support this methodological decision. First, the MNI method results are more informative for reconstructing gathering strategies, whereas weigh-based measures are more useful when comparing the relative importance of different shellfish items in dietary regimes, which was our purpose. Second, our framework relies on the link between dietary preferences and marine bioproductivity (Figure 1B), which is ultimately expressed as the amount (weight) of biomass (accumulated material) within an individual/assemblage for a given period. To test some of these assumptions, we obtained both measures for mussel species (Choromytilus chorus, Aulacomya ater, and P. purpuratus) from CV shell middens to evaluate discrepancies/convergences between these methods (see Supplementary Material 1). To standardize trends for changes in taxa representativeness within and among assemblages, we calculated the relative biomass abundance (percentage of total shellfish biomass) for species present in chronologically constrained stratigraphic units. The overall change in shellfish consumption among cultural periods was evaluated by summing the weight for every taxon over all strata that integrate the corresponding period. We then re-calculated the relative biomass parameter for each cultural period. The statistical analyses were applied to seven of the 17 taxa identified, as they are the most common throughout all of Caleta Vitor's trenches.
Variations in the assemblage structure were evaluated by calculating and comparing three diversity indices for each stratum and across different cultural periods. Biomass data were rounded into integer numbers to facilitate statistical analyses (i.e., bootstrap). The Shannon index (H′) measures diversity based on the relationship between richness (number of species) and relative abundances among species. The Simpson Dominance index (D) reflects the degree of dominance exerted by species within an assemblage. This ranges from 0 to 1, with lower values indicative of decreasing dominance. Conversely, the Evenness index of Buzas and Gibson (E) expresses how evenly individuals in the assemblage are distributed among the different species. If the E index trends toward 1, then all represented species within the assemblage have equivalent relative abundances. Diversity indices and subsequent statistical analyses were performed using PAST Paleontological Statistics Package Version 3.11 (Hammer et al., 2001). Corresponding confidence intervals at 95% for indices were generated via bootstrapping with 9,999 iterations. Diversity Permutation tests (9,999 random permutations) were used to assess significant differences in estimated diversity indices among strata and between selected cultural periods.
Complementary consumption of other marine resources was recorded using the number of identified specimens (NISP) of fish remains, which were grouped into four main categories based on the measurement of the diameter of the vertebrae: very small (1–3 mm), small (4–6 mm), medium to medium large (7–12 mm), and large (13–16 mm). Very large vertebrae correspond to two or three fish (CV1/3/30-31, L-XL = 17; CV2/1/27-58, L-XL = 67; CV3/1/30-31, L-XL = 40; Carter, 2016).
Based either on relative or available absolute dates for the stratigraphic units observed in our trenches, shellfish assemblages were sorted into main cultural periods for the Andes: Early Archaic, Middle-Late Archaic, Formative, Late Intermediate period, and Late period (see Table 1), complemented in some cases by the presence of diagnostic cultural material.
The role of ENSO in driving variations in shellfish assemblages was evaluated by comparing diversity indices obtained for each cultural period with a proxy record for past ocean primary productivity (Rein et al., 2005). This is a high-resolution record for photosynthetic pigment concentrations (carotenoids) in a marine core (SO147-106KL) collected offshore Peru (~12°S), where high (low) concentrations are interpreted as prevailing La Niña-like (Niño-like) conditions brought about by increased (reduced) primary productivity (Rein et al., 2005). Although ENSO-induced fluctuations in marine productivity could vary (especially on seasonal or annual timescales) in intensity along the eastern cold-tongue (Chavez et al., 1999), the carotenoid record is representative of long-term variations (increased vs. subdued productivity) at the regional scale. Indeed, the impacts of ENSO across the Pacific rely on large-scale oceanographic-atmospheric perturbations that are rapidly and consistently transmitted along the Pacific coast of South America at interannual timescales (Chavez et al., 1999).
A recent paleodemographic reconstruction for the coastal Atacama (16–25°S; Gayo et al., 2015) is used to examine the relationship between human-collected shell diversity, population dynamics, technology/innovations, and paleoclimate conditions. This reconstruction uses the summed probability distributions (SPD) of 579 archeological 14C-dates spanning the last 11,000 years contained within the “South Central Andes Radiocarbon Database” (SCAR; Gayo et al., 2015). Paleodemographic time-series based on SPDs assume that the accumulation of radiocarbon dates reflects the intensity of human activities in the landscape, and in turn in population levels (Rick, 1987; Buchanan et al., 2008; Chaput and Gajewski, 2016). Despite the wide popularity gained as a prehistoric demographic proxy, this method has received criticism regarding methodological biases imposed on the accumulation of data by research/sampling interests, taphonomy, and/or calibration procedures used to produce the SPD (Contreras and Meadows, 2014; Crombé and Robinson, 2014; Torfing, 2015; Hiscock and Attenbrow, 2016). Different methods have been proposed to overcome such biases (Michczynski and Michczynski, 2006; Williams, 2012; Timpson et al., 2014; Gayo et al., 2015) which show that these methodological biases in the long-term reconstruction are negligible for the paleodemographic history of coastal Atacama populations over the past 14,000 years. Hence, in this article we accept the reliability of the analyses presented and discussed by Gayo et al. (2015).
The statistical overall relationship, in terms of direction and magnitude, between assemblage structure, demographic trends and changes in bioproductivity trigged by ENSO was assessed using Pearson's correlation. The chronologies for our six assemblages (CV1, CV2, CV3, CV4, CV6, and CV7) show that these are discrete and discontinuous “snapshots” of each cultural period, so we delimited the chronological extensions of every index per assemblage by pooling minimum and maximum ranges of calendar ages (cal yr BP) using the 70 available radiocarbon dates from CV (Table 2). For that reason, correlations between temporal changes in marine productivity in the eastern Pacific and three diversity indices per assemblage were compared to the mean carotenoid concentration recorded during these interludes. The same procedure was adopted in evaluating the link between long-term regional demographic trends and ENSO variability.
Results
Shell remains are common throughout the trenches and strata of the six CV excavated shell midden sites (Figure 2), ranging from whole items of the more robust specimens (i.e., locos, C. concholepas) to very small fragmentary remains (i.e., sea urchins). The following list contains the species identified and itemized by prevalence (by mass). Despite elevated mollusk diversity along the Atacama Desert littoral (Rivadeneira et al., 2010), the people at Caleta Vitor preferred a narrow selection of just 18 taxa. As all these species are available or accessible in relatively shallow waters, they were easy to catch and the cost of extracting them was the same. From the 18 identified taxa, however, seven species were common throughout all trenches at the Caleta Vitor sites, and these quantitative variations over time are presented below. These include C. chorus (choro), A. ater (cholga), P. purpuratus (chorito), Fissurella spp. (lapa), C. concholepas (loco), Acanthopleura echinata (chitón), and Loxechinus albus (erizo or sea urchin).
Fluctuations in abundance frequencies were compared to ascertain if any pattern could be established. Because we are interested in the fluctuations in term of biomass of the major selected food items, 11 additional taxa with food and ornamental value that appear only irregularly and in very low frequencies were not considered in this analysis (i.e., Collisella d'orbigny, Thais chocolata, Protothaca thaca, Mesodesma donacium, Oliva peruviana, Argopecten purpuratus, barnacles (Cirripedia), Priene rude, Crepidula dilatata, Tegula spp., and Prisogaster niger).
Biomass distributions varied considerably among trenches and strata (Table 3). The highest shell abundance comes from CV6, where Late Intermediate period (CV6-1) and Late period (CV6-1 and CV6-2) units contribute with 30,478 g of shells. CV2 and CV4 trenches display comparable accumulated biomasses of 18,738 and 20,749 g, respectively. Minor total weights are recorded in CV1 (9,228 g) and CV3 (10,675 g) shell middens. Shell biomass in the Middle-Late Archaic CV7 trench is far lower compared to the other CV units and trenches (Table 3).
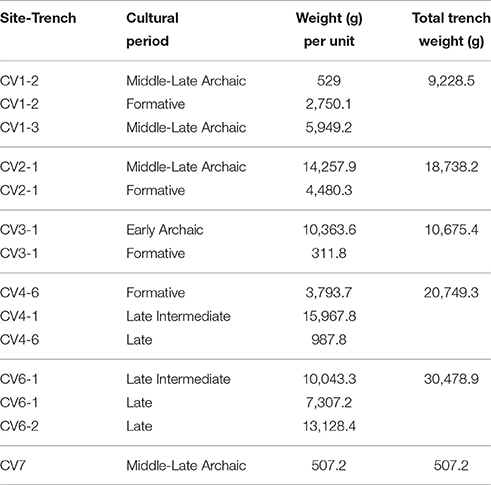
Table 3. Total biomass (g) per site and trench for eight taxa recorded in shellfish assemblages from Caleta Vitor.
Temporal Variations in the Structure of Shellfish and Fish Assemblages from Caleta Vitor
We defined the chronological framework for the socio-cultural periods known for northern Chile by pooling the 14C locally available dates from Caleta Vitor archeological sites. Thus, the three 14C dates for the early Archaic reveal a period between 9,490 and 8,720 cal yr BP (Figure 3). Similarly, the Middle and Late Archaic are represented practically in their entirety by dates spanning 7,600 – 3,360 cal yr BP. In contrast, the Formative, Late Intermediate and Late periods are more constrained between 2,730–1,610, 650–500, and 500–300 cal yr BP, respectively (see Table 1). The pattern of change in the composition of shellfish associations over time was inferred from an Early Archaic stratum in trench CV3-1, followed by four stratigraphic trench units for the Middle-Late Archaic (trenches CV1-2, CV1-3, CV2-1, and CV7) and the Formative (CV1-2, CV2-1, CV3-1, and CV4-6), two trench units for the Late Intermediate period (CV4-1 and CV6-1) and three strata for the Late period (trench units CV4-6, CV6-1, and CV6-2; Figure 3B).
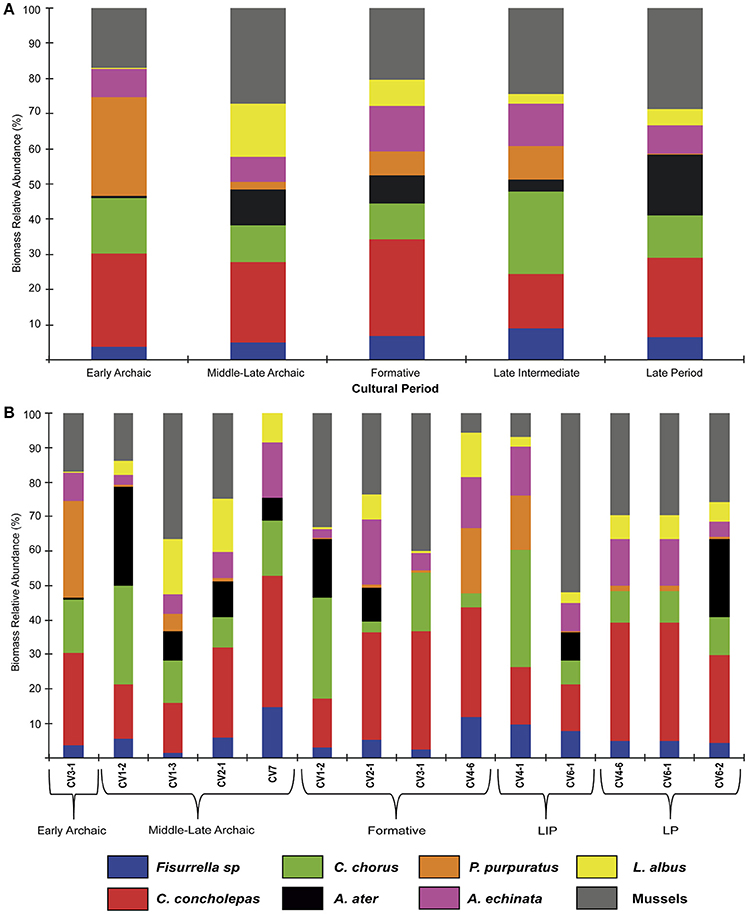
Figure 3. (A) Percent biomass relative abundance for each of the major archeological periods described for the Caleta Vitor archeological sites. Different colors pertain to different taxa after the legend at the bottom. (B) Same as (A), but for each of the different archaeological sites at Caleta Vitor.
In general, C. concholepas and mussels (Figure 3) are dominant in all cultural periods and strata, representing from 40 to 50.2% in all associations. These two taxa are also frequently observed in Chinchorro funerary contexts (Standen, 2003). The less abundant species were Fissurella spp., P. purpuratus and L. albus (Figure 3).
The mussel P. purpuratus is important during the Early Archaic, with 28% (Figure 3A, see Table in Supplementary Material 2). Less well-represented taxa include Fissurella spp. (3.6%), A. atra (0.48%) and L. albus (0.124%). Fish-remains reach the lowest value with respect to other cultural periods (2,022 individuals, Figure 3A).
An important faunal turnover occurred during the Middle-Late Archaic, marked by a significant increase in the abundance of L. albus and T. atra (Figure 3B). This was coeval with a dramatic increase in the incidence of fish remains, up to ~18,700 individuals (Figure 4A), with the highest value recorded in CV3-1 and CV2-1 (Figure 4B). The sea urchin L. albus is the third most abundant taxa (15.1%), during these periods, after mussels (27.3%) and C. concholepas (22.9%; Figure 3A). Middle-Late Archaic trenches (Figure 3B) show a significant decrease in P. purpuratus (2.1%, Figure 3B), which becomes a rare species along with the A. echinata (7.2%) and Fissurella spp. (4.7%; Figure 3A). This prominent pattern across all associations for P. purpuratus remained practically stable until the Late period (Figure 3A), except for trench CV4 during the Late Intermediate period (Figure 3B).
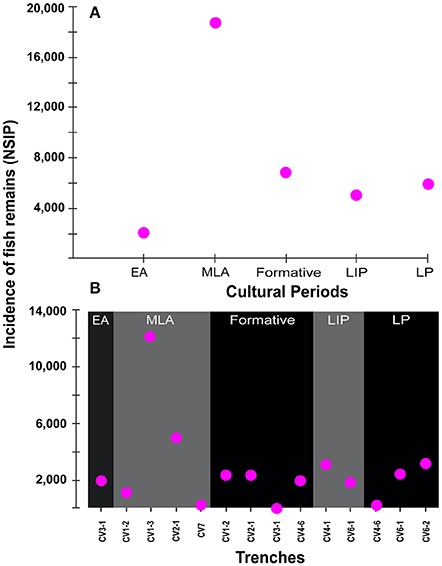
Figure 4. (A) Incidence of fish remains present in shell middens for each of the major cultural periods excavated in the Caleta Vitor archeological area. (B) Same as for (A), but for each of the archaeological sites at Caleta Vitor.
A dramatic decline is observed in the abundances of L. albus (<7.3%, Figure 3A) and fish remains in virtually all strata from the Formative period onwards (Figure 4). Fissurella spp. (6.8%), A. atra (7.9%) and P. purpuratus (6.8%) also remain poorly represented (Figure 3A). Compared to the Archaic, a significant increase in the abundance of A. echinata (13.2%, Figure 3A) occurs during the Formative, particularly in CV2-1, CV3-1, and CV4-6 (Figure 3B). Thus, this species co-dominates the spectrum of abundances with C. concholepas and mussels (Figure 3A).
The most abundant species of the Late Intermediate period were C. chorus (23.5%), mussels (24.4%), and C. concholepas (15.5%; Figure 3A). The less abundant species are similar for those recorded in the Early Archaic, although the proportions obtained in this latter range for L. albus (2.8%), A. atra (3.2%), and Fissurella spp. (8.9%) are comparatively higher. Fish abundances have a modest decrease to 5,065 individuals (Figure 4A).
The relative abundance of C. chorus decreases (Figure 3A) in the Late period whereas that of A. atra (17.1%) increases. This trend is especially visible at CV6-2 (Figure 3B). The representation of fish remains decreases slightly, particularly in trench CV4-6 (Figure 4). The less represented species are the same as those identified for the Formative, although values for L. albus (4.5%), Fissurella spp. (6.3%), and P. purpuratus (0.5%) are lower during the Late period.
Our assemblage structure analyses reveal statistically significant differences (p < 0.001) in Diversity (H′), Dominance (D), and Evenness (E) for shellfish species from the Early Archaic until the Late period (Table 4). D values obtained show that C. concholepas and mytilids (mussels) dominate all cultural and stratigraphic units. Together with a constant specific richness common to all of our analyses (n = 8), this dominance pattern indicates that variations in assemblage structure are explained by changes in relative abundance (i.e., exploitation) of C. chorus, A. ater, P. purpuratus, Fissurella spp., A. echinata, and L. albus.
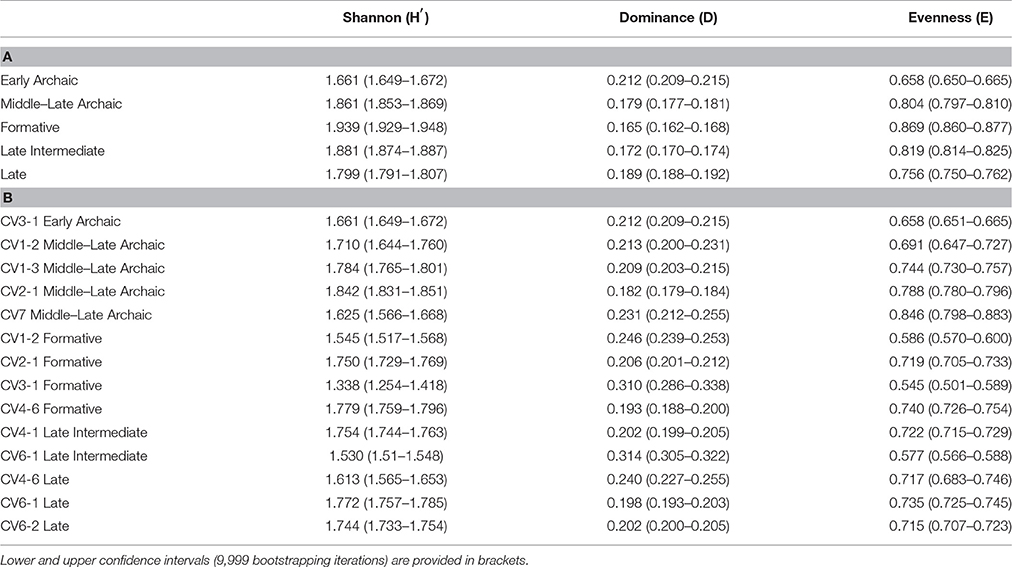
Table 4. Diversity indices for shellfish assemblages computed across different cultural periods (A) and for each site and trench (B).
The diversity (H′ = 1.66) and evenness (E = 0.66) of the Early Archaic assemblage are significantly lower than those recorded for later periods (Table 4). Indeed, dominance (D = 0.21) is comparatively higher due to the marked predominance in terms of relative biomass of three species (i.e., P. purpuratus, C. concholepas, and mytilids) and the marginal values shown by less well represented taxa (<3%, Figure 3A).
H′ and E increase to 1.86 and 0.8 respectively at the onset of the Middle-Late Archaic with maximum values during the Formative (H′ = 1.94, E = 0.87; Table 4A). The opposite pattern occurs for the Simpson Dominance Index. Permutation analyses show no statistically significant differences (p > 0.001) in Shannon, Dominance, and Evenness indices among strata from the Middle-Late Archaic as well as among Formative units. In fact, species with relatively low or intermediate abundances remain relatively constant throughout strata from both cultural periods (save during the Formative in Trench CV3-1; Figure 3B).
H′ and E indexes drop persistently from the Late Intermediate period (H′ = 1.88, E = 0.82) until the Late period (H′ = 1.79, E = 0.76) whereas D shows the opposite (Table 4A). Significantly different statistical differences (p < 0.001) exist for the Late Intermediate period for all three indexes. This is due to the dominance of mytilids in CV6-1 (51%, Figure 3B) in this assemblage, thus reducing (increasing) the respective E (D) index. No significant differences exist for changes in Evenness across the strata of the Late period (p > 0.001). In contrast, the Shannon and Dominance indexes are significantly different (p < 0.001) due in part to the absence of A. atra in CV4-6, a species that is particularly important in CV6-2 (Figure 3B).
Discussion
The time-series derived from the marine record of photosynthetic pigment concentrations argues for important variations in primary productivity at millennial and centennial scales over the past 10,000 years (Rein et al., 2005; Figure 5). We verified significant differences (Kruskal–Wallis test, p < 0.001) in down-core carotenoid concentrations across the five cultural periods examined at Caleta Vitor. Pigment concentrations were maximum (mean = 14.95 ± 6.81) during the Early Archaic and steadily decreased during the Middle-Late Archaic (mean = 10.58 ± 6.47) until the Formative (mean = 8.74 ± 5.25). Concentrations then increased during the Late Intermediate period (mean = 11.65 ± 7.82) and then fell to their lowest values during the Late period (mean = 5.23 ± 2.27).
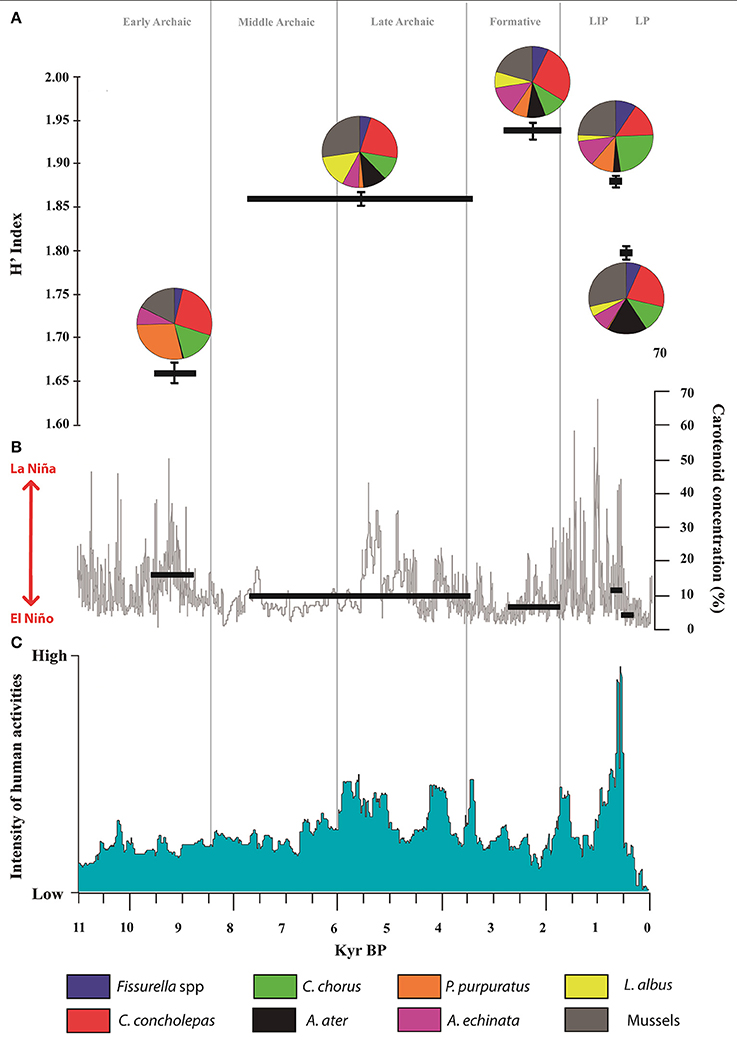
Figure 5. (A) H′ Diversity index and pie charts for biomass relative abundances expressed as percentages for each shellfish assemblage. Fissurella spp. proportion: blue, Concholepas concholepas proportion: red, Choromytilus chorus proportion: light green, Aulacomya ater proportion: dark, Perumytilus purpuratus proportion: cyan, Acanthopleura echinata proportion: magenta, Loxechinus albus proportion: yellow, Mussels proportion: olive. (B) ENSO-driven changes in bioproductivity. Magenta and gray curves show raw (gray) and 50-year running mean (magenta) for carotenoid concentrations within the SO147-106KL marine core (Rein et al., 2005). Horizontal green bars show mean values for photosynthetic pigments throughout the constrained chronology of each cultural period. (C) Reconstructed intensity of human activity in the coastal Atacama Desert over the past 10,000 years (after Gayo et al., 2015). Vertical light gray lines represent the real duration of cultural periods. Major regional-scale wet (light blue) and dry (orange bars) phases are provided at the top of the figure.
Analysis of the shellfish collected by people at Caleta Vitor (i.e., C. chorus, A. atra, and P. purpuratus, C. concholepas, Fissurella spp., L. albus, and A. echinata) reveals significant fluctuations in their frequency through time that has a clear correspondence with these variations in productivity. These variations seem to be associated with major changes in tropical sea surface temperatures (SST) driven by ENSO fluctuations. ENSO thus likely exerted an important role on the structuring of our shellfish assemblages present at Caleta Vitor. Indeed, the low H′- and E-values during the Early Archaic were coeval with an exceptionally prolonged productive phase given by the persistence of a cold phase of ENSO-like variations.
Save for the trend observed in the Late period (Figure 5), the majority of the cultural periods evince an inverse relationship with between Diversity H′ and pigment concentration (i.e., marine primary productivity). The same is true for Evenness, but the opposite occurs for Dominance. Indeed, Pearson correlations analyses show that either H′ or E parameters are negative and strongly correlated with marine bioproductivity from the Early Archaic to the Late Intermediate Period; both correlations yielding Pearson's r = −0.956 (n = 4, p < 0.05). In contrast, we verify a strong positive but marginally non-significant correlation for Dominance and down-core carotenoid concentrations (r = 0.934, n = 4, p = 0.06).
For example, as photosynthetic pigment concentrations decreased offshore Lima (Peru) beginning at 8,500 cal yr BP, diversity indices of collected shellfish increased during the Middle-Late Archaic at CV (Figures 3, 5). Large increases in marine productivity also occurred at 5,590 and 4,380 cal yr BP, possibly due to brief incursions of La Niña-like conditions (more favorable to strong upwelling conditions) despite overall El Niño-like conditions during the middle Holocene (Moy et al., 2002). This would explain the prominent decrease seen in the diversity indices (Table 4) at trench CV7 (5,040–4,150 cal yr BP, Table 2), which is coeval with the irruption of stronger La Niña-like conditions during the late Archaic (Rein et al., 2005).
Regionally, major socio-cultural innovative reorganizations and transformations occurred during the Middle-Late Archaic, including the diversification of artificial mummification practices, and the appearance of specialized technology for exploiting offshore and intertidal resources along the arheic coast (Marquet et al., 2012; Olguín et al., 2014; Castro et al., 2016; Standen et al., 2017). Coincidently, after c. 4,000 to 3,000 cal yr BP, the fishing, hunting gathering societies abandoned the long term traditional ritual mortuary system of artificial mummification and new ideologically base funerary patterns were introduced along the coast of northernmost Chile (Marquet et al., 2012). Similarly, an extreme reduction in marine productivity (i.e., due to sustained El Niño-like conditions) correlates with increases in the Evenness and Shannon indexes for the Formative shellfish assemblages (Figure 5). In contrast, these indices decreased during strong La Niña-like conditions at 650 and 500 cal yr BP, which correspond to periods of increased offshore primary productivity.
As gleaned from the shell middens, since the Early Archaic and throughout the Formative and in younger deposits (c. 9,500–1,650 cal yrs BP), people maintained a marine economy by arranging their behavioral patterns of consumption during periods of low marine productivity through reliance on a more diverse menu of mollusks. This, aside from confirming the key role of seafood for pre-Columbian coastal populations of western South America, shows that the everyday cuisine of people from Caleta Vitor underwent modifications over time following cultural preferences.
As for fish remains, their incidence closely follows the observed relationship between Diversity H′ and carotenoid concentrations (Figures 4, 5). Although the lowest values for fish incidence and H′ index by the Early Archaic coincide with maximum pigment concentrations, the strength for this co-variation is not linear. The highest fish abundance occurs during a period of moderately high Shannon index and subdued accumulation of photosynthetic pigments in the marine core. Increased diversity of shellfish species in the Formative was coeval with moderate increased fish consumption (Figure 4), and was also coeval with a period of reduced carotenoid concentrations compared to the Middle-Late Archaic (Figure 5).
The overall abundance of mussels—including unidentified specimens and those assigned to P. purpuratus, C. chorus and A. ater—suggests that, collectively, these items were an important part of the diet. In the case for P. purpuratus and C. chorus, it is evident that their representation into shellfish assemblages co-varies negatively either at cultural period or trench level (Figures 3, Supplementary Material 1). Indeed, P. purpuratus is the most dominant (28%) mussel species in the Early Archaic (Figure 3) which implies a certain cultural preference community base management decision (Tompkins and Adger, 2004) for specific food items by these coastal peoples at this time. In this regard, ENSO-induced changes in coastal SST could be responsible for such patterns, as C. chorus and P. purpuratus prefer cool and warmer waters, respectively (Diaz and Ortlieb, 1993; Avendaño and Cantillánez, 2011). Because increased upwelling implies decreased SSTs along the Pacific coast of South America, we would expect an increase in C. chorus abundances during cultural periods with amplified carotenoid concentrations in the SO147-106KL marine core. Yet, except for the Late Intermediate our data do not support this relationship. In fact, C. chorus prevails over other mussel species even during intervals of increased SSTs and reduced upwelling. This lends support for a culturally-driven hypothesis for why specific seafood taxa were consumed based on community management decisions (Figures 3, 5).
Whereas the basic economy remained without major changes, our long-term paleodemographic reconstructions show that coastal populations maintained a distinct but unusual saw-tooth oscillating population pattern over the last 10,000 years (Figure 5C) most likely coupled with “boom or bust” cycles linked to the frequency of ENSO events on centennial timescales, although other factors (such as regional aquifer recharge, see also Marquet et al., 2012) could be important. Furthermore, the positive impact of population size upon cumulative socio-cultural continuities and innovation dynamics likely increased the technological toolkit associated with resource use through time. Save for the Early Archaic, Pearson's r coefficients—ranging from 0.202 to 0.374—suggest moderate positive correlations between regional demographic trends and ENSO-driven changes in marine productivity. The resulting negative correlation for the Early Archaic period (Pearson = −0.306, p < 0.05) is related to low population levels albeit increased marine productivity as evinced by pigment concentrations at 9,490–8,720 cal yr BP (Figure 5). Conversely, since the Middle-Late Archaic onwards there is a systematic positive co-variation between both variables. Sustained population growth from 6,000 to 5,000 and then from 4,300 to 3,300 cal yr BP closely match positive incursions in pigment concentrations detected in the offshore marine core. Similarly, reduced demographic levels are found as carotenoid accumulations dropped from 7,600 to 6,000 and 5,000 to 4,300 cal yr BP. Low population levels over most of the Formative (2,700–1,610 cal yr BP) are consistent with reduced carotenoid values. The marked recovery at 2,100–1,400 cal yr BP coincides with high but variable concentrations in photosynthetic pigments. Coastal populations declined abruptly in the Late period, along with regional marine productivity (Figure 5), which coincided with a reduction in the consumption of C. chorus and fish and saw an increase in A. Atra. Thus, as previously mentioned C. chorus is a good example of the discordance between climatic conditions and cultural preferences: during the Late Intermediate people maintained its consumption despite the increasing of SSTs and reduced upwelling (meaning warm impropriated water for this taxon). This implies that cultural preference for specific food items persisted despite increased environmental stress.
Nevertheless, the adoption of a more generalist feeding behavior—represented by periods of increased H′ and E indices in shellfish associations form Caleta Vitor (Figure 3)—represents an adaptive strategy (Winterhalder and Smith, 2000) through the expansion of their diet over low-ranked species to cope with resource variability and unpredictability. These adaptation strategies of flexible consumption of shellfish were adopted, transmitted and reproduced during the Formative period when marine productivity was depleted. In general, such patterns of prey use consumption associated with a continuous consumption of the most energetically rich items (e.g., C. concholepas) together with the increased consumption of less energetically rewarding items under low productivity (and likely less abundant than the preferred prey) can be explained by optimal diet theory (Stephens and Krebs, 1986), which does a good job at predicting the consumption of prey of reduced mobility (Sih and Christensen, 2001), as is the case of coastal invertebrates.
Finally, the environmental variations described and associated impacts on the diversity of selected shellfish for consumption did not trigger, however, any major shifts toward a dependence on terrestrial dietary sources, which reveals that coastal peoples were highly flexible to their changing coastal environment. Thus farming products introduced into the interior valleys and oases of northern Chile during the Formative were not integrated into the economy of Caleta Vitor social groups. Instead, their economy remained close to its traditional methods of food extraction for thousands of years; which highlights the social or community-based management decisions to retain their traditional marine food adaptive strategy. The most important introductions of inland farming crops, particularly maize, and fruit tree dietary sources (such as Prosopis) occurred during the Late Intermediate period (McRostie et al., 2017), and consumption was intensified during the Late period linked to local production reorganized under the Inka State regime.
Conclusions
Along the Pacific coast of northern Chile, as elsewhere, our study shows that the peoples at Caleta Vitor subsisted on a rather narrow selection of a wide array of available marine and littoral species, which included a mixture of gastropods, bivalves, crustaceans, fish, sea birds, and sea mammals. This narrow selection persisted throughout most of the Holocene despite pronounced variations in human population size and marine coastal productivity.
Indeed, the most important changes most likely occurred at the technological and ideological level by the end of the Archaic period and throughout the Formative, through the introduction of new funerary behavior that included tombs marked with timber pots, and the construction of monumental tumulus as seen in CV3. This was coincidental with a regional decline in marine bioproductivity, local human population expansion and immigration from the adjacent inland areas (Rothhammer and Cocilovo, 2008; Rothhammer et al., 2009; Orellana et al., 2014). Although, certain Chinchorro features were maintained (i.e., crania filled with ash, soil, and plant matter), this change meant the abandonment of the long-term tradition of Chinchorro artificial mummification.
Significant fluctuations occurred in the frequency of Caleta Vitor shellfish assemblages associated with major changes in tropical SST. In other words, ENSO likely exerted a strong influence on the structuring of these assemblages. People, however, seem to have been highly selective as they centered their consumption on just seven species (lapa, loco, chitón, erizo, and three species of choro). Within this selection locos and choros were dominant in all cultural periods. This predilection for shellfish (easier to catch) over fish began in the Early Archaic, and increased dramatically during the Middle Archaic along with increased consumption of L. albus and T. atra. In contrast, consumption of P. purpuratus, A. echinata, and Fissurella spp. decreased. A dramatic decrease in fish remains, L. albus, Fissurella spp., A. atra, and P. purpuratus occurred from the Formative onwards. Meanwhile, A. echinata increased and joined the dominant taxa (choros y locos). The predilection for the two latter species continued even into the Late Intermediate period. In sum, the environmental variations that impacted the diversity of selected shellfish for consumption did not trigger any major shifts on their persistent marine coastal economy, revealing a high degree of resilience to constantly shifting coastal ecosystems. The Caleta Vitor sequence is thus a key example of human cultural socio-cultural dietary resilience in the context of large environmental changes over many millennia.
Author Contributions
CS, CC, RDP, and CL designed and funded the research and wrote the paper. EG, VS, VC, DV, and PM contributed data, additional analyses and wrote the paper.
Funding
Our research was funded by FONDECYT projects #1150763 (to CL, CS, and RDP), #1151046 (to DV, VC, and CS) and #11150210 (to EG). CL and PM also acknowledge funding from the IEB (grant PFB-23). EG and RDP acknowledge support from the Center for Climate and Resilience Research (CR)2 (FONDAP/CONICYT #15110009). CS, VC, DV, VS, EM, PM, and CL thank CONICYT's Programa de Investigación Asociativa (PIA), project Anillo SOC1405. VS thanks UTA Mayor Grant 3718-16.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Hérida Zamora for drafting Figure 1A, and Constanza Gayo for Figure 2. This study was conducted with archaeological excavation permit #3127/15 from the Consejo de Monumentos Nacionales (Chilean National Council on Cultural Heritage).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2017.00077/full#supplementary-material
References
Acerbi, A., Kendal, J., and Tehrani, J. J. (2017). Cultural complexity and demography: the case of folktales. Evol. Hum. Behav. 38, 474–480. doi: 10.1016/j.evolhumbehav.2017.03.005
Álvarez, M., Briz Godino, I., Balbo, A., and Madella, M. (2011). Shell middens as archives of past environments, human dispersal and specialized resource management. Q. Int. 239, 1–7. doi: 10.1016/j.quaint.2010.10.025
Andrade, P., Castro, V., and Aldunate, C. (2016). Reconstrucción del modo de vida de individuos del arcaico de la costa arreica del norte de Chile: una aproximación bioarqueológica desde el sitio Copaca 1. Chungara Rev. de Antropol. Chil. 48, 73–90. doi: 10.4067/S0717-73562016005000007
Arriaza, B. T., Standen, V. G., Cassman, V., and Santoro, C. M. (2008). “Chinchorro culture: pioneers of the coast of the atacama desert,” in Handbook of South American Archaeology, eds H. Silverman and W. H. Isbell (New York, NY: Springer), 45–58.
Avendaño, M., and Cantillánez, M. (2011). Reestablecimiento de Choromytilus chorus (Molina, 1782) (Bivalvia: Mytilidae) en el norte de Chile. Lat. Am. J. Aquat. Res. 39, 390–396. doi: 10.3856/vol39-issue2-fulltext-20
Bailey, G. N. (1975). The role of molluscs in coastal economies: the results of midden analysis in Australia. J. Archaeol. Sci. 2, 45–62. doi: 10.1016/0305-4403(75)90045-X
Bibar, G. D. (1966[1558]). Crónica y Relación Copiosa y Verdadera de los Reynos de Chile. Santiago: Fondo Histórico y Bibiográfico José Toribio Medina.
Bittmann, B. (1983). Cobija: panorama etnohistórico en relación a los informes del Dr. José Agustín de Arze. Chungara Rev. de Antropol. Chil. 10, 147–153.
Bland, C. A., Roberts, A. L., Popelka-Filcoff, R. S., Santoro, C. M., Carter, C., Bennett, J. W., et al. (2016). 1500 years of pottery: neutron activation analysis of northern chilean domestic ceramics from caleta vitor and clay samples from nearby valley, coast and highland contexts. Archaeometry 59, 815–833. doi: 10.1111/arcm.12277
Brown, K., Fa, D. A., Finlayson, G., and Finlayson, C. (2011). “Small game and marine resource exploitation by Neanderthals: the evidence from Gibraltar,” in Trekking the Shore, eds N. F. Bicho, J. A. Haws, and L. G. Davis (New York, NY: Springer), 247–272.
Buchanan, B., Collard, M., and Edinborough, K. (2008). Paleoindian demography and the extraterrestrial impact hypothesis. Proc. Nat. Acad. Sci. U.S.A. 105, 11651–11654. doi: 10.1073/pnas.0803762105
Campbell, S. K. (2005). “Shell middens,” in Encyclopedia of Coastal Science, ed M. L. Schwartz (Dordrecht: Springer), 870–873.
Carter, C. P. (2016). The Economy of Prehistoric Coastal Northern Chile: Case Study. Doctor of Philosophy Thesis, Canberra, The Australian National University.
Castro, V., Aldunate, C., Varela, V., Olguín, L., Andrade, P., García-Albarido, F., et al. (2016). Ocupaciones arcaicas y probables evidencias de navegación temprana en la costa arreica de Antofagasta, Chile. Chungara Rev. de Antropol. Chil. 48, 503–530. doi: 10.4067/S0717-73562016005000039
Chaput, M. A., and Gajewski, K. (2016). Radiocarbon dates as estimates of ancient human population size. Anthropocene 15, 3–12. doi: 10.1016/j.ancene.2015.10.002
Chavez, F. P., Strutton, P., Friederich, G. E., Feely, R., Feldman, G. C., Foley, D., et al. (1999). Biological and chemical response of the equatorial Pacific Ocean to the 1997–98 El Ni-o. Science 286, 2126–2131.
Contreras, D. A., and Meadows, J. (2014). Summed radiocarbon calibrations as a population proxy: a critical evaluation using a realistic simulation approach. J. Archaeol. Sci. 52, 591–608. doi: 10.1016/j.jas.2014.05.030
Croes, D. R. (2015). The undervalued black katy chitons (katharina tunicata) as a shellfish resource on the northwest coast of North America. J. Northwest Anthropol. 49, 13–25.
Crombé, P., and Robinson, E. (2014). 14C dates as demographic proxies in Neolithisation models of northwestern Europe: a critical assessment using Belgium and northeast France as a case-study. J. Archaeol. Sci. 52, 558–566. doi: 10.1016/j.jas.2014.02.001
Derex, M., and Boyd, R. (2015). The foundations of the human cultural niche. Nat. Commun. 6:8398. doi: 10.1038/ncomms9398
Diaz, A., and Ortlieb, L. (1993). El Fenómeno “El Niño” y los moluscos de la costa peruana. Bull. Inst. Fr. Etudes Andines 22, 159–177.
Disspain, M. C. F., Ulm, S., Santoro, C. M., Carter, C., and Gillanders, B. M. (2016). Pre-Columbian fishing on the coast of the Atacama Desert, northern Chile: an investigation of fish size and species distribution using otoliths from Camarones Punta Norte and Caleta Vitor. J. Island Coast. Archaeol. 23, 428–450. doi: 10.1080/15564894.2016.1204385
Erlandson, J. M. (1988). The role of shellfish in prehistoric economies: a protein perspective. Am. Antiq. 53, 102–109. doi: 10.2307/281156
Escobar, M. (2015). Para vivir en Cobija: prevalencia del modo de vida costero en Atacama. Chungara Rev. de Antropol. Chil. 45, 303–318. doi: 10.4067/S0717-73562015005000024
Faulkner, P. (2011). Quantifying shell weight loss in archaeological deposits. Archaeol. Ocean. 46, 118–129. doi: 10.1002/j.1834-4453.2011.tb00106.x
Flores, C., Figueroa, V., and Salazar, D. (2015). Middle Holocene production of mussel shell fishing artifacts on the coast of Taltal (25° Lat South), Atacama Desert, Chile. J. Isl. Coast. Archaeol. 11, 411–424. doi: 10.1080/15564894.2015.1105884
Frézier, A. (1713). Voyage to the South-Sea, and along the Coasts of Chili and Peru, in the Years 1712, 1713, and 1714: Particularly Describing the Inhabitants, as well Indians as Spaniards. London: Jonah Bowyer.
Gayo, E. M., Latorre, C., and Santoro, C. M. (2015). Timing of occupation and regional settlement patterns revealed by time-series analyses of an archaeological radiocarbon database for the South-Central Andes (16–25°S). Q. Int. 356, 4–14. doi: 10.1016/j.quaint.2014.09.076
Gayo, E. M., Latorre, C., Santoro, C. M., Maldonado, A., and De Pol-Holz, R. (2012). Hydroclimate variability on centennial timescales in the low-elevation Atacama Desert over the last 2,500 years. Climate Past 8, 287–306. doi: 10.5194/cp-8-287-2012
Glassow, M. A. (2000). Weighing vs. counting shellfish remains: a comment on Mason, Peterson, and Tiffany. Am. Antiq. 65, 407–414. doi: 10.2307/2694067
Grosjean, M., Santoro, C. M., Thompson, L., Núñez, L., and Standen, V. G. (2007). “Mid-Holocene climate and culture change in the south-central Andes,” in Climate Change and Cultural Dynamics: A global Perspective on Mid-Holocene Transitions, eds D. G. Anderson, K. A. Maasch, and D. H. Sandweiss (San Diego, CA: Elsevier), 51–115.
Habu, J., Matsui, A., Yamamoto, N., and Kanno, T. (2011). Shell midden archaeology in Japan: aquatic food acquisition and long-term change in the Jomon culture. Q. Int. 239, 19–27. doi: 10.1016/j.quaint.2011.03.014
Hammer, Ø., Harper, D. A. T., and Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 4, 1–9.
Henrich, J., Boyd, R., Derex, M., Kline, M. A., Mesoudi, A., Muthukrishna, M., et al. (2016). Understanding cumulative cultural evolution. Proc. Nat. Acad. Sci. U.S.A. 133, 6724–6725. doi: 10.1073/pnas.1610005113
Hiscock, P., and Attenbrow, V. (2016). Dates and demography? The need for caution in using radiometric dates as a robust proxy for prehistoric population change. Archaeol. Ocean. 51, 218–219. doi: 10.1002/arco.5096
Hoggs, A. G., Hua, Q., Blackwell, P. G., Niu, M., Buck, C. E., Guilderson, T. P., et al. (2013). SHCal13 southern hemisphere calibration, 0–50,000 years cal BP. Radiocarbon 55, 1889–1903. doi: 10.2458/azu_js_rc.55.16783
Hoke, G. D., Isacks, B. L., Jordan, T. E., and Yu, J. S. (2004). Groundwater-sapping origin for the giant quebradas of northern Chile. Geology 32, 605–608. doi: 10.1130/G20601.1
Horta, H. (2015). El Señorío Arica y los Reinos Altiplánicos. Santiago: Quillqa, Universidad Católica del Norte.
Latorre, C., De Pol-Holz, R., Carter, C., and Santoro, C. M. (2017). Using archaeological shell middens dating as a proxy for past local coastal upwelling in northern Chile. Q. Int. 427, 128–136. doi: 10.1016/j.quaint.2015.11.079
Lizárraga, R. D. (1987[1594–1608]). Descripción del Perú, Tucumán, Río de la Plata y Chile. Editado por I. Ballesteros. Madrid: Historia.
Lozano Machuca, J. (1965[1581]). Carta del Factor de Potosí Juan Lozano Machuca al Virrey del Perú, en Donde se Describe la Provincia de los Lipes. Madrid: Biblioteca de Autores Espa-Oles.
Marquet, P. A., Santoro, C. M., Latorre, C., Standen, V. G., Abades, S. R., Rivadeneira, M. M., et al. (2012). Emergence of social complexity among coastal hunter-gatherers in the Atacama Desert of northern Chile. Proc. Nat. Acad. Sci. U.S.A. 109, 14754–14760. doi: 10.1073/pnas.1116724109
Mason, R. D., Peterson, M. L., and Tiffany, J. A. (1998). Weighing vs. counting: measurement reliability and the California school of midden analysis. Am. Antiq. 63, 303–324.
Mason, R. D., Peterson, M. L., and Tiffany, J. A. (2000). Weighing and counting shell: a response to Glassow and Claassen. Am. Antiq. 65, 757–761. doi: 10.2307/2694427
Masotti, I., Moulin, C., Alvain, S., Bopp, L., Tagliabue, A., and Antoine, D. (2011). Large-scale shifts in phytoplankton groups in the Equatorial Pacific during ENSO cycles. Biogeosciences 8, 539–550. doi: 10.5194/bg-8-539-2011
McRostie, V. B. (2014). Arboricultura y silvopastoralismo en el período Formativo (1.400 a.c.–500 d.C.) de la cuenca del Salar de Atacama. Chungara Rev. de Antropol. Chil. 46, 543–557. doi: 10.4067/S0717-73562014000400002
McRostie, V. B., Gayo, E. M., Santoro, C. M., De Pol-Holz, R., and Latorre, C. (2017). The pre-Columbian introduction and dispersal of Algarrobo (Prosopis, section algarobia) in the Atacama Desert of northern Chile. PLoS ONE 12:e0181759. doi: 10.1371/journal.pone.0181759
Michczynski, A., and Michczynski, D. J. (2006). The effect of PDF peaks' height increase during calibration of radiocarbon date sets. Geochronometria 25, 1–4.
Moy, C. M., Seltzer, G. O., Rodbell, D. T., and Anderson, D. M. (2002). Variability of El Ni-o/Southern Oscillation activity at millennial timescales during the Holocene epoch. Nature 420, 162–165. doi: 10.1038/nature01194
Muñoz, I., Agüero, C., and Valenzuela, D. (2016). “Poblaciones prehispánicas de los valles occidentales del norte de Chile: desde el período Formativo al Intermedio Tardío (ca. 1.000 a.c.–1.400d.c.),” in Prehistoria en Chile desde sus Primeros Habitantes hasta los Incas, eds F. Falabella, M. Uribe, L. Sanhueza, C. Aldunate, and J. Hidalgo (Santiago: Editorial Universitaria y Sociedad Chilena de Arqueología), 181–237.
Núñez, L. (1986a). Balsas prehistóricas del litoral chileno: grupos, funciones y secuencia. Boletín del Museo Chileno de Arte Precolombino 1, 35–40.
Núñez, L. (1986b). Evidencias arcaicas de maíces y cuyes en Tiliviche: hacia el sedentarismo en el litoral fértil y quebradas del Norte de Chile. Chungara Rev. de Antropol. Chil. 16–17, 25–49.
Núñez, L., Cartajena, I., Carrasco, C., López, P., de Souza, P., Rivera, F., et al. (2017). Presencia de un centro ceremonial formativo en la circumpuna de Atacama. Chungara Rev. de Antropol. Chil. 49, 3–33. doi: 10.4067/S0717-73562017005000003
Núñez, L., and Santoro, C. M. (2011). El tránsito Arcaico-Formativo en la circunpuna y valles occidentales del Centro Sur Andino: hacia los cambios “neolíticos.” Chungara Rev. Antropol. Chil. 43, 487–531. doi: 10.4067/S0717-73562011000300010
Olguín, L. (2014). El Rol económico de los recursos malacológicos durante el Holoceno medio en la costa arreica del norte de Chile: sitio Agua Dulce. Rev. Chil. de Antropol. 29, 32–39. doi: 10.5354/0719-1472.2014.36203
Olguín, L., Salazar, D., and Jackson, D. (2014). Tempranas evidencias de navegación y caza de especies oceánicas en la costa pacífica de Sudamérica (Taltal, ~ 7.000 años cal. a.p.). Chungara Rev. de Antropol. Chil. 46, 177–192. doi: 10.4067/S0717-73562014000200002
Orellana, N., Fuentes-Guajardo, M., Capriles, J. M., and Rothhammer, F. (2014). En torno al poblamiento de los Andes sur-centrales y su viculación con la Amozonía. Interciencia 39, 586–590.
Paez, R. (1986). Balsas de cuero de lobo en la segunda mitad del siglo XIX: antecendentes cuantitativos para el norte de Chile. Chungara Rev. de Antropol. Chil. 16–17, 421–428.
Pestle, W. J., Torres-Rouff, C., Hubbe, M., Santana, F., Pimentel, G., Gallardo, F., et al. (2015). Explorando la diversidad dietética en la prehistoria del Desierto de Atacama: un acercamiento a los patrones regionales. Chungara Rev. de Antropol. Chil. 47, 201–209. doi: 10.4067/S0717-73562015005000013
Petruzzelli, B., Roberts, A., Pate, F. D., Santoro, C. M., Maddern, T., Carter, C., et al. (2012). Stable carbon and nitrogen isotopic analysis of skeletal remains from Azapa 71 and Pica-8, northern Chile: an assessment of human diet and landscape use in the late Holocene. J. Anthropol. Soc. South Aust. 35, 52–80.
Polo de Ondegardo, J. (1916[1571]). Informaciones acerca de la Religión y Gobierno de los Incas. Edición de Horacio Urteaga y Carlos. Romero; Lima: Colección de Libros y Documentos Referentes a la Historia del Perú, Primera Serie Tomo, III.
Poulson, S. R., Kuzminsky, S. C., Scott, G. R., Standen, V. G., Arriaza, B. T., Muñoz, I., et al. (2013). Paleodiet in northern Chile through the Holocene: extremely heavy δ15N values in dental calculus suggest a guano-derived signature? J. Archaeol. Sci. 40, 4576–4585. doi: 10.1016/j.jas.2013.07.009
Reimer, P. J., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G., Bronk Ramsey, C., et al. (2013). IntCal13 and Marine13 radiocarbon age calibration curves 0–50,000 years cal BP. Radiocarbon 55, 1869–1887. doi: 10.2458/azu_js_rc.55.16947
Rein, B., Lückge, A., Reinhardt, L., Sirocko, F., Wolf, A., and Dullo, W. C. (2005). El Ni-o variability off Peru during the last 20,000 years. Paleoceanography 20:PA4003. doi: 10.1029/2004PA001099
Reitz, E. J., deFrance, S. D., Sandweiss, D. H., and McInnis, H. E. (2015). Flexibility in southern Peru coastal economies: a vertebrate perspective on the terminal Pleistocene/Holocene transition. J. Isl. Coastal Archaeol. 10, 155–183. doi: 10.1080/15564894.2014.934492
Rick, J. W. (1987). Dates as data: an examination of the Peruvian preceramic radiocarbon record. Am. Antiq. 52, 55–73. doi: 10.2307/281060
Rick, T. C., and Waselkov, G. A. (2015). Shellfish gathering and shell midden archaeology revisited: chronology and taphonomy at White Oak point, potomac river estuary, Virginia. J. Isl. Coastal Archaeol. 10, 339–362. doi: 10.1080/15564894.2014.967896
Rivadeneira, M., Santoro, C. M., and Marquet, P. A. (2010). Reconstructing the history of human impacts on coastal biodiversity in Chile: constraints and opportunities. Aquat. Conserv. 20, 74–82. doi: 10.1002/aqc.1051
Roberts, A., Pate, F. D., Petruzzelli, B., Carter, C., Westaway, M. C., Santoro, C. M., et al. (2013). Retention of hunter–gatherer economies among maritime foragers from Caleta Vitor, northern Chile, during the late Holocene: evidence from stable carbon and nitrogen isotopic analysis of skeletal remains. J. Archaeol. Sci. 40, 2360–2372. doi: 10.1016/j.jas.2013.01.009
Rothhammer, F., and Cocilovo, J. A. (2008). Microdiferenciación craneana en la costa de Arica y valle de Azapa, norte de Chile. Resúmen de resultados y conclusiones. Chungara Rev. de Antropol. Chil. 40, 309–315. doi: 10.4067/S0717-73562008000300007
Rothhammer, F., Santoro, C. M., Poulin, E., Moraga, M., and Standen, V. G. (2009). Archeological and mtDNA evidence for tropical lowland migrations and cultural change during the Late Archaic/Formative in northern Chile. Rev. Chil. de Hist. Nat. 82, 543–552. doi: 10.4067/S0716-078X2009000400008
Salazar, D., Figueroa, V., Andrade, P., Salinas, H., Olguín, L., Power, X., et al. (2015). Cronología y organización económica de las poblaciones arcaicas de la costa de Taltal. Estud. Atacam. 50, 7–46. doi: 10.4067/S0718-10432015000100002
Santoro, C., Castro, V., Carter, C., and Valenzuela, D. (in press). “Maritime Communities in the Atacama Desert. Masters of the sub tropical pacific coast of South America,” in Maritime Communities of the Ancient Andes, eds G. Prieto D. H. Sandweiss (Gainesville, FL: University Press of Florida)
Santoro, C. M., Gayo, E. M., Capriles, J. M., de Porras, M. E., Maldonado, A., Standen, V. G., et al. (2017). Continuities and discontinuities in the socio-environmental systems of the Atacama Desert during the last 13,000 years. J. Anthropol. Archaeol. 46, 28–39. doi: 10.1016/j.jaa.2016.08.006
Schiappacasse, V., and Niemeyer, H. (1984). Descripción y Análisis Interpretativo de un Sitio Arcaico Temprano en la Quebrada de Camarones. Santiago: Publicación Ocasional del Museo Nacional de Historia Natural.
Sih, A., and Christensen, B. (2001). Optimal diet theory: when does it work, and when and why does it fail? Anim. Behav. 61, 379–390. doi: 10.1006/anbe.2000.1592
Standen, V. G. (2003). Bienes funerarios del cementerio Chinchorro Morro 1: descripción, análisis e interpretación. Chungara Rev. de Antropol. Chil. 35, 175–207. doi: 10.4067/S0717-73562003000200002
Standen, V. G., Santoro, C. M., Arriaza, B., and Coleman, D. (2017). Hunting, gathering, and fishing on the coast of the Atacama Desert: Chinchorro population mobility patterns inferred from strontium Isotopes. Geoarcaheol Int. J. doi: 10.1002/gea.21594. [Epub ahead of print].
Standen, V. G., Santoro, C. M., and Arriaza, B. T. (2004). Síntesis y propuesta para el período Arcaico en la costa del extremo norte de Chile. Chungara Rev. Antropol. Chil. 36, 201–212. doi: 10.4067/S0717-73562004000300023
Stephens, D. W., and Krebs, J. R. (1986). Foraging Theory. Princeton, NJ: Princeton University Press.
Swift, J., Cupper, M. L., Greig, A., Westaway, M. C., Carter, C., Santoro, C. M., et al. (2015). Skeletal arsenic of the Pre-Columbian population of Caleta Vitor, northern Chile. J. Archaeol. Sci. 58, 31–45. doi: 10.1016/j.jas.2015.03.024
Thiel, M., Macaya, E. C., Acuna, E., Arntz, W. E., Bastias, H., Brokordt, K., et al. (2007). The Humboldt current system of northern and central Chile: oceanographic processes, ecological interactions and socioeconomic feedback. Oceanogr. Mar. Biol. 45, 195–344. doi: 10.1201/9781420050943.ch6
Timpson, A., Colledge, S., Crema, E., Edinborough, K., Kerig, T., Manning, K., et al. (2014). Reconstructing regional population fluctuations in the European Neolithic using radiocarbon dates: a new case-study using an improved method. J. Archaeol. Sci. 52, 549–557. doi: 10.1016/j.jas.2014.08.011
Tompkins, E., and Adger, W. N. (2004). Does adaptive management of natural resources enhance resilience to climate change? Ecol. Soc. 9:10.
Torfing, T. (2015). Neolithic population and summed probability distribution of 14C-dates. J. Archaeol. Sci. 63, 193–198. doi: 10.1016/j.jas.2015.06.004
Valenzuela, D., Sepúlveda, M., Santoro, C. M., and Montt, I. (2014). Arte rupestre, estilo y cronología: la necesidad de un contexto histórico para las manifestaciones rupestres en costa y valles del extremo norte de Chile. Interciencia 39, 444–449.
Waselkov, G. A. (1987). Shellfish gathering and shell midden archaeology. Adv. Archaeol. Method Theor. 10, 93–210. doi: 10.1016/B978-0-12-003110-8.50006-2
Williams, A. N. (2012). The use of summed radiocarbon probability distributions in archaeology: a review of methods. J. Archaeol. Sci. 39, 578–589. doi: 10.1016/j.jas.2011.07.014
Keywords: marine diet, cultural resources management, prehistoric technology, Holocene climate, Atacama Desert, ENSO
Citation: Santoro CM, Gayo EM, Carter C, Standen VG, Castro V, Valenzuela D, De Pol-Holz R, Marquet PA and Latorre C (2017) Loco or no Loco? Holocene Climatic Fluctuations, Human Demography, and Community Based Management of Coastal Resources in Northern Chile. Front. Earth Sci. 5:77. doi: 10.3389/feart.2017.00077
Received: 29 June 2017; Accepted: 22 September 2017;
Published: 18 October 2017.
Edited by:
Gary E. Stinchcomb, Murray State University, United StatesReviewed by:
David K. Wright, Seoul National University, South KoreaSilviu Constantin, Emil Racovita Institute of Speleology, Romania
Copyright © 2017 Santoro, Gayo, Carter, Standen, Castro, Valenzuela, De Pol-Holz, Marquet and Latorre. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Claudio Latorre, Y2xhdG9ycmVAYmlvLnB1Yy5jbA==