 Janina Schoen1,2‡
Janina Schoen1,2‡ Marco Muñoz-Becerra1,2
Marco Muñoz-Becerra1,2 Jasmin Knopf3Favour Ndukwe1,2
Jasmin Knopf3Favour Ndukwe1,2 Moritz Leppkes2,4Dominik Roth4Anne Zeitler1,2
Moritz Leppkes2,4Dominik Roth4Anne Zeitler1,2 Verena Gerlinde Frings5
Verena Gerlinde Frings5 Bettina Hohberger6Victoria Zeisberg6
Bettina Hohberger6Victoria Zeisberg6 Luis E. Muñoz1,2
Luis E. Muñoz1,2 Georg Schett1,2
Georg Schett1,2 Martin Herrmann1,2,3*†
Martin Herrmann1,2,3*† Christine Schauer1,2†
Christine Schauer1,2†- 1Department of Internal Medicine 3, Rheumatology and Immunology, Friedrich-Alexander-Universität Erlangen-Nürnberg (FAU) and Universitätsklinikum Erlangen, Erlangen, Germany
- 2Deutsches Zentrum für Immuntherapie (DZI), Friedrich-Alexander-Universität Erlangen-Nürnberg (FAU) and Universitätsklinikum Erlangen, Erlangen, Germany
- 3Department of Pediatric Surgery, University Medical Center Mannheim, University of Heidelberg, Mannheim, Germany
- 4Department of Internal Medicine 1 - Gastroenterologie, Pneumologie und Endokrinologie, Friedrich-Alexander-Universität Erlangen-Nürnberg (FAU) and Universitätsklinikum Erlangen, Erlangen, Germany
- 5Department of Dermatology, Venereology and Allergology, University Hospital Würzburg, Würzburg, Germany
- 6Department of Ophthalmology, Universitätsklinikum Erlangen and Friedrich-Alexander-University of Erlangen-Nürnberg (FAU), Erlangen, Germany
Aims: Neutrophils and neutrophil extracellular traps (NETs) play multifaceted roles in inflammatory diseases. If the balance of NET formation and clearance is disturbed, they contribute to the development and pathogenesis of a plethora of inflammatory diseases. They promote inflammation and tissue degradation, and occlude vessels and ducts. This study focused on the presence of NET remnants generated during the clearance by nucleases and phagocytes.
Methods: NET associated parameters in serum and plasma samples from various pathological conditions were investigated. We performed fluorescence-based assays to analyze the concentration of cell free DNA and the activity of neutrophil elastase. The presence of citrullinated histone H3, as well as neutrophil elastase- or myeloperoxidase-DNA complexes were examined employing enzyme-linked immunosorbent assays.
Results: We analyzed samples from a variety of inflammatory conditions: (I) the rheumatic autoimmune diseases systemic lupus erythematosus, rheumatoid arthritis, and primary Sjögren’s syndrome (II) the inflammatory bowel diseases ulcerative colitis and Crohn’s disease (III) hidradenitits suppurativa and (IV) the viral-induced pathologies Coronavirus disease 2019 (COVID-19), and Post COVID Syndrome (PCS). While most NET associated parameters were detected in all inflammatory conditions, certain markers displayed disease-specific patterns. We compared the markers in terms of the concentration, correlations with each other and to disease activity, and their impact on sample variability. Systemic lupus erythematosus and rheumatoid arthritis were associated with increased levels of cell free DNA, and citrullinated histone H3 as well as neutrophil elastase-activity, respectively. Samples from patients with COVID-19 were characterized by elevated levels of neutrophil elastase- and myeloperoxidase-DNA complexes.
Conclusion: Different diseases are linked to characteristic patterns of NET associated parameters. These patterns offer insights into aberrant NET formation and clearance in different pathologies and may represent key targets for treatment development.
Introduction
Neutrophils play a crucial role in our immune system, acting as the first line of defense against invading pathogens (Boeltz et al., 2019; Schoen et al., 2022; Mayadas et al., 2014). Despite their undoubted beneficial role in host defense, neutrophils can become double-edged swords if dysregulated. Then, they contribute to onset and pathology of a plethora of inflammatory diseases (Chatfield et al., 2018; Papayannopoulos, 2018; Wang et al., 2018b; Singh et al., 2023). Neutrophils accomplish their defensive mission employing phagocytosis, degranulation, the production of reactive oxygen species (ROS) and inflammatory cytokines, and the release of neutrophil extracellular traps (NETs) (Brinkmann et al., 2004; Segal, 2005; Tecchio et al., 2014; Winterbourn et al., 2016; Burgener and Schroder, 2020; Burn et al., 2021; Wang et al., 2023a). NETs are composed of mesh-like DNA-bundles decorated with granular and cytoplasmic proteins, forming a trap to intercept and neutralize invaders. Additionally, they serve as a protective shield against inflammatory mediators (Brinkmann et al., 2004; Schoen et al., 2022). Nucleases and phagocytes are responsible for the cleavage and removal of these DNA-peptide complexes from the bloodstream and tissues (Figure 1). DNases play a central role in the degradation of NETs. They hydrolyze the phosphodiester bonds of the chromatin. This enzymatic reaction results in the breakdown of NETs, allowing phagocytes to engulf and subsequently degrade the remnants (Demkow, 2023). Extracellular degradation is achieved by nucleases of the DNase family I, which are present in body fluids like blood (Laukova et al., 2020). DNase1 and DNase1L3 are particular important in this context (Hakkim et al., 2010; Jimenez-Alcazar et al., 2015). Enzymes of the 3′-exonucleases (TREX) family also contribute to NET degradation. They can dismantle oxidized DNA, that is resistant to canonical DNases (Lazzaretto and Fadeel, 2019; Laukova et al., 2020). After cleavage of the chromatin fibers, the smaller remnants become accessible for digestion by phagocytes - mainly macrophages, but also dendritic cells (Lazzaretto and Fadeel, 2019; Haider et al., 2020). These residues are broken down in the lysosomes without triggering inflammatory reactions under non-physiological conditions (Farrera and Fadeel, 2013; Hu et al., 2017). This process is facilitated by opsonization of NETs with complement factor 1q (C1q) (Farrera and Fadeel, 2013).
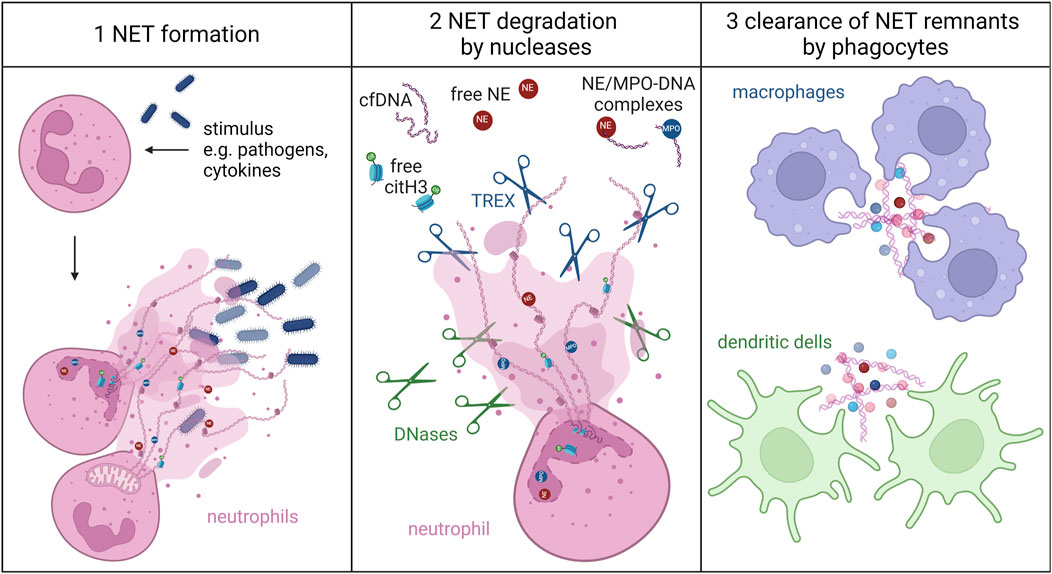
Figure 1. The clearance of NETs. Upon stimulation by pathogens or endogenous inflammatory mediators, neutrophils release NETs, composed of decondensed chromatin bundles decorated with cytoplasmic and granular proteins. These NETs immobilize and eliminate invaders. The chromatin-peptide complexes are degraded by nucleases like DNases and TREX, releasing NET degradation products such as cell free DNA (cfDNA), citH3, NE, and NE- or MPO-DNA complexes. NETs remnants are eventually engulfed and degraded by phagocytes, like macrophages and dendritic cells. Created with BioRender.com.
If nucleases and phagocytes fail to rapidly degrade and clear NETs, the NET-borne citrullinated Histone H3 (citH3), and the enzymes neutrophil elastase (NE), and myeloperoxidase (MPO) can precipitate damage and inflammation in the “battlefield” (Fuchs et al., 2007; Kaplan and Radic, 2012; Klebanoff et al., 2013; Nakazawa et al., 2017; Okeke et al., 2020). Extended NETs accumulations, referred to as aggregated NETs (aggNETs) can clog vessels, but can also contribute to the resolution of inflammation by scavenging and degrading inflammatory cytokines (Hahn et al., 2019; Vorobjeva, 2020; Yaykasli et al., 2021). This multifaceted nature of NETs renders neutrophils and NETs important players in the development and pathogenesis of various inflammatory diseases. In addition, they are not only formed in response to invading pathogens but also to endogenous inflammatory signals. The delicate balance between NET production and clearance is crucial for tissue homeostasis (Figure 2) (Papayannopoulos, 2018; Mulay and Anders, 2020; Knopf et al., 2022). If this balance is disturbed and NETs, together with their associated peptides, accumulate, they may trigger the development of autoantibodies and immune complexes (Hakkim et al., 2010). The binding of these autoantibodies to the DNA of the chromatin bundles sterically hinders the degradation by DNases and further stabilizes the NETs (Al-Mayouf et al., 2011; Carmona-Rivera et al., 2015; Zuo et al., 2021).
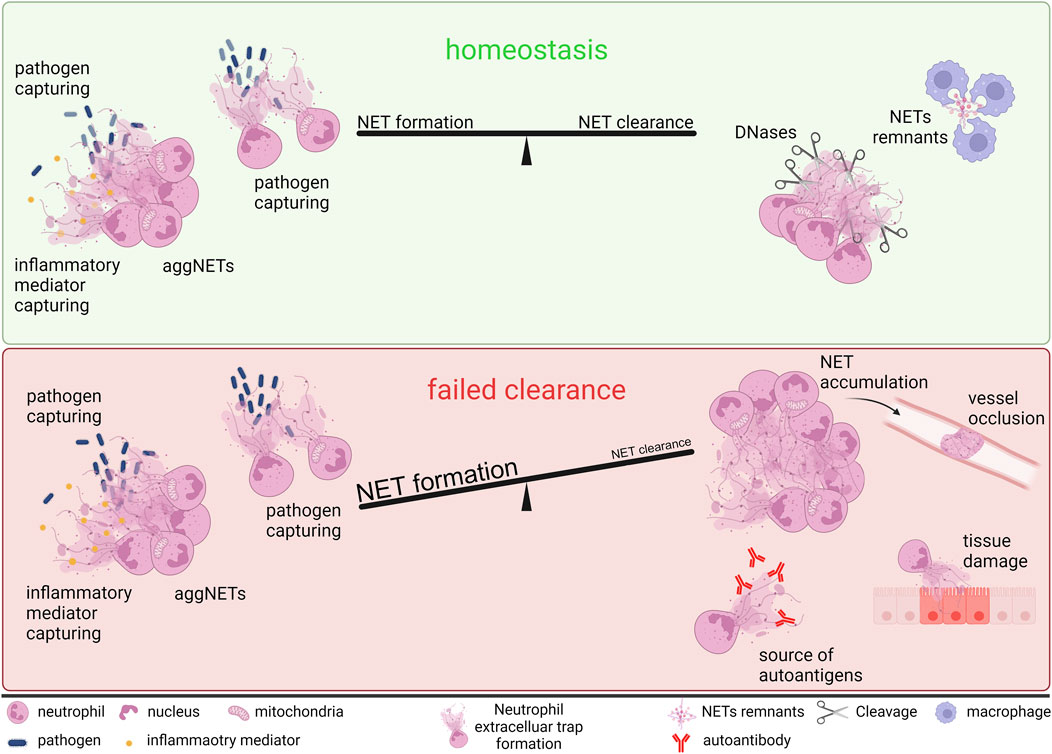
Figure 2. Neutrophils and NETs in tissue homeostasis and disease development. To maintain tissue homeostasis, a delicate balance between NET formation and clearance is mandatory. NETs and aggNETs are formed to trap and kill invaders, as well as to control inflammation by capturing inflammatory mediators. If the released chromatin is quickly degraded and cleared by DNases and phagocytes, tissue homeostasis is preserved. If this balance shifts in favor of NET formation, extracellular chromatin accumulates and promotes inflammation and tissue damage. In larger aggregates, NETs can clog vessels and ducts, serve as autoantigens, leading to the production of autoantibodies, and toxify the surrounding tissue. Created with BioRender.com.
In patients with systemic lupus erythematosus (SLE), NET-IgG complexes cause endothelial damage (Villanueva et al., 2011). The complexes promote the release of type I interferon by mononuclear phagocytes. They also induce NF-kB activation in endothelial cells (Grayson and Kaplan, 2016; Lou et al., 2020). Consequently, these complexes enhance inflammatory immune reactions and thus contribute to disease progression. In a similar way, NETs contribute to the pathology of rheumatoid arthritis (RA). A hallmark of RA is the formation of autoantibodies against citrullinated antigens (ACPA) (Makrygiannakis et al., 2006; Khudhair, 2023). Citrullination is a crucial step of the formation of NETs. NET-bound citrullinated histones can be detected in the extracellular matrix, which situates them as an antigen source for the development of ACPAs (Li et al., 2010; Khandpur et al., 2013). In addition, NETs contribute to joint damage by enhancing proinflammatory responses, including an increased stimulation of proinflammatory cytokine production (Khandpur et al., 2013). Due to enhanced type I IFN, ROS production and associated inflammation and endothelial damage, neutrophils contribute to the pathogenesis of primary Sjögren’s syndrome (pSS). Elevated levels of neutrophils and NETs in lungs and salivary glands are linked to disease severity (Fu et al., 2021; Peng et al., 2022). In inflammatory bowel diseases, enhanced NET formation is also associated with disease severity (Dos Santos Ramos et al., 2021). They stimulate surrounding immune cells to release higher levels of pro-inflammatory mediators and play a role in the break down the extracellular matrix. This results in the impairment of the intestinal epithelial barrier’s function. Here we examine the two main forms of this group of diseases, ulcerative colitis (UC) (Bennike et al., 2015; Dos Santos Ramos et al., 2021; Wang et al., 2023b), and Crohn’s disease (CD) (Dos Santos Ramos et al., 2021; Fu et al., 2021; Schroder et al., 2022). Neutrophils migrate towards the abscess-like nodules of patients with hidradenitits suppurativa (HS), where they abundantly form NETs. Impaired NET clearance is associated with prolonged inflammation and disease severity (Ogawa et al., 2021; Oliveira et al., 2023). Neutrophils and NET can also contribute to the development and pathogenesis of viral-induced diseases. In Coronavirus disease 2019 (COVID-19), caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) neutrophils and an increased NET formation promotes clogging of vessels. The occlusion of the microvasculature contributes to tissue damage and eventually organ dysfunction (Leppkes et al., 2020; Knopf et al., 2022; Singh et al., 2022). In patients with PCS, NETs are linked to the prolonged inflammation, cardiovascular irregularities, and malfunction of the neurological system (Shafqat et al., 2023).
In general, neutrophils and NETs are implicated in the development and pathogenesis of various inflammatory diseases. They cause inflammatory reactions, increase tissue damage, and vascular occlusions. With this study, we aimed to find specific evidence of the different roles of neutrophils, especially NETs, in various pathologies. It shall serve as a guide to identify which aspects of NET formation, and their impact on inflammatory processes, should be prioritized for future therapeutic development. Therefore, we analyzed the NET degradation products NE-DNA and MPO-DNA complexes; and the NET-associated parameters citrullinated histone 3 (citH3) and NE activity in samples from patients and healthy controls (HCs).
Methods
Patients and healthy controls
Healthy controls (n = 96), and patients with SLE (n = 94), RA (n = 80), pSS (n = 21), UC (n = 12), CD (n = 37), HS (n = 30), COVID-19 (n = 67), and PCS (n = 30) were recruited in Bavaria, Germany. Studies were performed in accordance with the ethical approval, with serum and plasma samples obtained from blood samples of the patients. Samples from patients with HS were collected at the university hospital of Würzburg, Germany. Samples from patients with all other listed diseases were collected at the university hospital of Erlangen, Germany. Patients with pSS, SLE, and RA met the criteria of classification of the American College of Rheumatology/European League Against Rheumatism. Disease severity of patients with SLE was identified by the European Consensus Lupus Activity Measurement (ECLAM). Patients with RA were further classified using the Disease Activity Score 28 (DAS28).
Ethical approval
All experiments with human material were performed in accordance with the Declaration of Helsinki in Ethical Principles for Medical Research Involving Human Subjects, and with the approval of the Ethical Committee of the University Hospital Erlangen (permit #193_13B and # 174_20B). For the HS serum samples, an approval of the Ethics Committee (107/20) of the University of Würzburg and a signed informed consent were obtained from all patients prior to inclusion.
Detection of cfDNA in serum and plasma samples
Cell-free DNA (cfDNA) circulating in samples from patients with inflammatory diseases was quantified and compared to samples from normal healthy donors (Supplementary Figure S1). For patients with COVID-19, plasma samples were used, for patients with all other listed diseases, the assay was performed with serum samples. For serum collection we used Sarstedt S-monovette tubes. We allowed the blood to clot by leaving the Sarstedt tube undisturbed at room temperature for 1 h. After clotting, we centrifuged the Sarstedt tube at 3,000 × g for 15 min. For the collection of plasma samples we centrifuged EDTA blood twice at 3,000 g for 15 min.
The Quant-iT™ PicoGreen™ dsDNA Assay-Kit (ThermoScientific, P11496) was used following the manufacturer’s instructions. The samples were diluted 1:20 with the provided TE-buffer and mixed in a 1:1 ratio with Quant-iT™ PicoGreen™ dsDNA reagent in a 96-well flat bottom plate (Greiner, 655180). After a 5-min incubation in the dark at room temperature (RT), fluorescence was measured using an Infinite® 200 PRO plate reader (Tecan; ex. 485 nm, em. 535 nm). The concentration of cfDNA was determined using Excel (Microsoft Office Professional Plus 2019) with the standard provided by the kit; all data were normalized for comparability.
citH3 ELISA of serum and plasma samples
Citrullinated Histone H3 served as marker to assess the presence of neutrophil extracellular traps (NETs) in serum samples from patients with inflammatory diseases and from normal healthy donors. The concentration was determined using a citrullinated Histone H3 ELISA-Kit (Cayman Chemical, Clone 11D3, 501620). The assay was performed according to the manufacturer’s instructions. The OD values of the samples – 1:4 diluted with the provided assay-buffer – were measured in a SUNRISE microplate reader (Tecan, 450 nm, reference 620 nm). The concentration of citrullinated Histone H3 in the samples was calculated in Excel (Microsoft Office Professional Plus 2019) using the provided standard; all data were normalized for comparability.
Evaluation of NE-activity in serum and plasma samples
The enzymatic activity of neutrophil elastase (NE) of both, patients with inflammatory diseases and healthy controls, was determined using the fluorogenic substrate MeOSuc-SSPV-AMC (Santa Cruz Biotechnology, sc-201163) at the final concentration of 100 µM. For patients with COVID-19, plasma samples were used, for patients with all other listed diseases, the assay was performed with serum samples. Samples were diluted 1:10 in PBS in a 96-well flat bottom plate (Greiner, 655180) and incubated with the fluorogenic substrate in a final volume of 100 µL for 24 h at 37°C. Pure Enzyme (NE, Sigma, E8140-1UN) at various dilutions served as controls. Fluorescent readings were recorded at 10 min intervals using an Infinite® 200 PRO plate reader (Tecan; ex. 360 nm, em. 465 nm). The NE-activity of the serum samples compared to the controls was evaluated using Excel (Microsoft Office Professional Plus 2019); all data were normalized for comparability.
Quantification of NET degradation products by NE-DNA/MPO-DNA ELISA
Circulating neutrophil elastase (NE)-DNA and myeloperoxidase (MPO)-DNA complexes, serving as indicators of NET presence, were quantified using ELISAs. For patients with COVID-19, plasma samples were used, for patients with all other listed diseases, the assay was performed with serum samples. 96-well maxisorp immune-plates (ThermoScientific, 442404) were coated overnight at 4°C with an anti-NE (rabbit anti-human NE, R&D, MAB91673) or an anti-MPO antibody (rabbit anti-human MPO, Merck, 07-496-I) in coating buffer (0.1 M Na2CO3, 0.1 M NaHCO3, pH 9.6). Following the coating, the plates were washed with 0.05% Tween-20 in PBS three times and were subsequently blocked with 3% BSA in PBS for 2 h at RT on a horizontal shaker. After each of the following steps, the washing procedure was repeated, and all incubation steps were performed at RT with continuous shaking. Serum samples (40 µL per patient or normal healthy donor) were incubated undiluted for 2 h on the pre-coated plates. Enzyme-DNA complexes were then detected using the peroxidase-conjugated anti-DNA POD from the Cell Death Detection ELISA plus kit (Sigma/Roche, 11774425001), diluted 1:100 with the provided lysis-buffer, by incubation for 90 min. The wells were then incubated with TMB substrate (BioLegend, #421101) for 60 min in the dark. The reaction was stopped with 25% H2SO4, and OD values were measured using a SUNRISE microplate reader (Tecan, 45 nm, reference 620 nm) and evaluated with Excel (Microsoft Office Professional Plus 2019); all data were normalized for comparability.
Statistical analyses
For comparison between three of more groups, the non-parametric analysis of variance (ANOVA) Kruskal-Wallis-test with Dunns multiple comparisons correction was performed. For comparisons between two groups, a Mann-Whitney test was conducted. Correlation analyzes regarding the connection of NET associated parameters were performed using the Pearson correlation test. Correlation coefficients are shown as heatmaps with hierarchical clustering. Regression lines are included in the respective plots. Values of p < 0.05 were considered statistically significant. All tests were performed using the GraphPad Prism 9 Software (by Dotmatics). In order to visualize the five NET associated parameters from the data set, dimensionality reduction was performed by principal component analysis (PCA) and FFT-accelerated Interpolation-based t-Stochastic Neighbor Embedding (FIt-SNE). All numeric variables were exported into an R data table object containing also the categorical label of the patient group. For PCA a correlation matrix was calculated using the function “cor()” of the “corrr” package in R. The function “prncomp()” computes the PCA and the contribution of each variable or patient was visualized using the function “fviz_pca_var.” FIt-SNE was carried out through the “run.Fitsne ()” function. The values of each variable were represented in a color scale created with the function “make.colour.plot()” available in the Spectre R package with detailed instructions and source code accessible https://github.com/ImmuneDynamics/spectre.
Results
Inflammatory conditions display disease-specific patterns of NET degradation products
Neutrophils and NETs play a multifaceted role in inflammatory diseases. To understand their involvement in disease development, pathogenesis, and healing, we investigated the presence and potential variations of NET associated parameters in various pathological conditions (Figure 3). NETs are composed of extended chromatin bundles. During the degradation process these are cleaved by DNases and consequently appear in the circulation as soluble fragments, which are measurable as cfDNA. The release of neutrophil proteins and enzymes during NET formation promotes inflammation of surrounding tissues. We analyzed the presence of two key NET-associated molecules, citH3 and NE, focusing on the potential toxicity of the latter as indicated by its activity. NET proteins also appear chromatin-bound, leading to the release of peptide-DNA complexes during NET degradation. We quantified NE-DNA and MPO-DNA complexes.
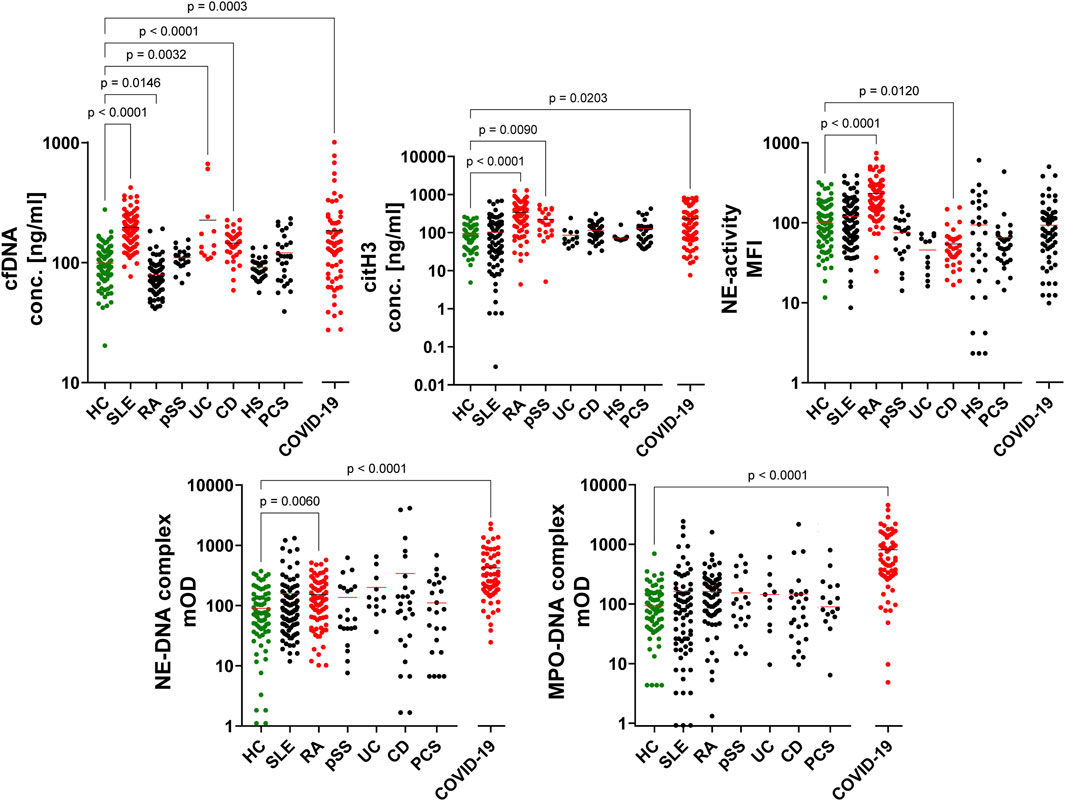
Figure 3. NET associated parameters are serum markers for inflammatory diseases. Samples from patients with the inflammatory diseases SLE, RA, pSS, UC, CD, HS, COVID-19, or PCS are represented by dots. These samples were tested for the presence of NET associated parameters, in comparison to a cohort of healthy controls (HC; green). Significant changes compared to the HC cohort are highlighted in red (Kruskal–Wallis p < 0.05). n(HC) = 96, n(SLE) = 94, n(RA) = 80, n(pSS) = 21, n(UC) = 12, n(CD) = 37, n(HS) = 30, n(COVID-19) = 67, n(PCS) = 30. Note: COVID-19 are plasma samples.
Patients with SLE exhibited elevated levels of cfDNA in their serum. Whereas in RA patients, an increase of the citH3 concentration, NE-activity, and appearance of NE-DNA complexes was to be observed. The pSS cohort only displayed higher concentrations of citH3. Patients with inflammatory bowel diseases, UC or CD, showed elevated cfDNA concentrations, with CD patients also exhibiting a significant decrease in NE-activity. COVID-19 patients displayed elevated levels of all tested NET associated parameters, except for NE-activity. The HS and PCS cohorts did not reveal any significant differences compared to the HC group, although the former showed a broad distribution of NE-activity. These analyses offer a first insight into the differences in NET associated parameters as inflammatory patterns in various pathological conditions.
Correlation analyzes reveal disease-specific patterns of NET degradation products
We then performed a principal component analysis (PCA) to reveal the relationship between the relative contributions of NET associated parameters and their importance for to the variability in among patients. The PCA plot of variables (Figure 4A) shows the relationships between the NET associated parameters analyzed. NE-activity, and the concentration of citH3 contribute similarly to the variance, as do NE- and MPO-DNA complexes but in opposite direction. The cfDNA is located in a different quadrant from other markers, indicating an independent contribution. The square cosine values (cos2) denotes the grade of contribution of each variable to the whole variance. cfDNA emerges with the highest cos2 value as the most important variable among the patient groups studied, followed by NE-DNA complexes, MPO-DNA complexes, NE-activity, and finally citH3. The score PCA plot of patients (Figure 4B) illustrates a division the clustering of the patients in two major groups depending on component 1 (Dim1), which explains 56.6% of the variation in the data. One group predominantly comprises dots representing patients with COVID-19 (purple). On the opposite side, a large cluster of dots represents patients with SLE (blue) and patients with RA (red). This cluster subdivides based on component 2 (Dim2), which explains 30% of the variance.

Figure 4. Dimensionality reduction of NET associated parameters in inflammatory diseases. (A) Principal component analysis (PCA) of the NET associated parameters. Markers with co-variability are clustered in the same quadrant. The square cosine value (cos2) indicates the variable representation. High cos2 values are depicted in green (0.17), low values in black (0.12), and intermediate values by a spectrum from yellow to brown (0.13–0.16). (B) PCA of NET associated parameters. Each patient cohort is assigned to a color to identify representing dots. Component 1 (Dim1) explains 56.6%, component 2 (Dim2) 30% of the variability. (C) t-distributed stochastic neighbor embedding (tSNE) tSNE plots of the patient cohorts and NET associated parameters visualize disease clusters. Each dot represents a patient, different colors the diseases. In tSNE plots that visualize the distribution of the NET associated parameters and their measured levels, high and low values are displayed in red and dark blue, respectively. Note: COVID-19 are plasma samples (*).
To gain further insight into the role of each marker in the examined pathological conditions, t-distributed stochastic neighbor embedding (tSNE) plots were used for the visual representation of the data (Figure 4C). The first graph shows the distribution of the diseases. Three major clusters can be observed – SLE, RA, and COVID-19. All other cohorts and the HC group are evenly spread in the center with only a few dots shifted towards the edges. The visualization of the NET associated parameters illustrates the sample distribution with varying levels, depicted by a color gradient from red to dark blue (indicating higher to lower values). The marker with the highest impact on variability, cfDNA, shows a scattered color map, but forms a distinct cluster within the SLE cohort. NE-activity and citH3, which correlated in the PCA, exhibit a similar distribution pattern with accumulation of high values in the area representing the majority of RA patients. A comparable association can be observed between NE-, and MPO-DNA complexes with the COVID-19 group. These data highlight the variability of the markers and their distinct distribution among the various pathologies.
COVID-19 is associated with high levels of NE- and MPO-DNA complexes
As COVID-19 is the only acute and non-chronic disease in our study, and the correlation analyzes revealed distinct differences between this cohort and the other groups of diseases, we decided to perform a separate, in-depth analysis. We compared patients with COVID-19 to those with the chronic variant, PCS, and the HC group. The distribution of the groups in the tSNE plots emphasize the accumulation of dots representing COVID-19 patients within the left area of the graph (Figure 5A). This distinctive pattern was not observed in the plot highlighting the distribution of patients with PCS, which demonstrated a closer similarity to the HC cohort.
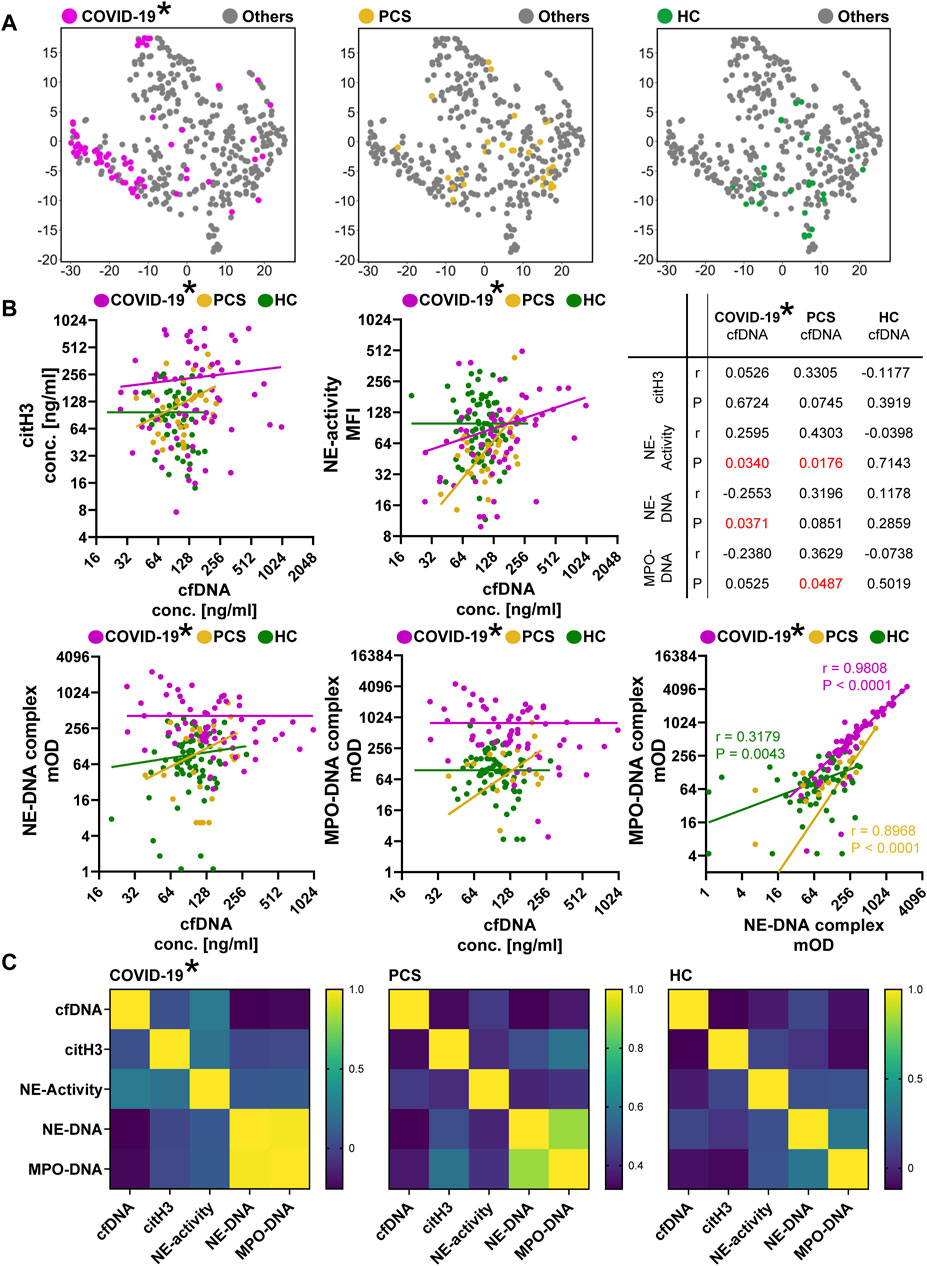
Figure 5. Differential analysis of patients with COVID-19 and PCS. (A) tSNE plots highlighting the differences in the distribution of patients with COVID-19 (purple), PCS (yellow), and the HC group (green). (B) Correlation analyzes between cfDNA and the other NET-degradation markers citH3, NE-activity, NE-complexes and MPO-complexes. Each dot represents a patient–COVID-19 in purple, PCS in yellow, and HCs in green. Information about the statistical parameter r and the significance P are presented in the provided table. An additional graph shows the correlation of NE- and MPO-DNA complexes. Statistical information is included in the graph. (C) Heatmaps of the correlation analyzes. High values are depicted in yellow (1.0), low values in dark blue (0), and intermediate values in a color gradient from light green to blue (high to low values, respectively). For all correlation analyzes shown in this figure, a Two-tailed Pearson correlation was performed (p < 0.05). Note: COVID-19 are plasma samples (*).
We then conducted detailed correlation analyzes of the NET degradation markers and NET associated parameter of the three groups (Figure 5B). Recognizing cfDNA as the variable with the highest impact on variability, we focused on its correlation with all other markers. Patients with COVID-19 exhibit higher values for citH3, but without correlation to cfDNA. The activity of NE is similar for the three groups, but a positive correlation to cfDNA is detectable for patients with COVID-19 (p = 0.0340) and even more pronounced for PCS (p = 0.0176). High levels of NE- and MPO-DNA complexes are visible in the COVID-19 cohort. Slightly significant correlations exist for NE-DNA complexes and COVID-19 (p = 0.0371), or MPO-DNA complexes and PCS patients (p = 0.0487). Given the apparent similarity in distribution patterns of NE- and MPO-DNA complexes, we additionally correlated these two markers. There was a highly significant correlation between the two complexes (p < 0.0001); the values for patients with COVID-19 were almost identical, as shown by the r value of 0.9808. These correlations are visualized as heatmaps in Figure 5C, that highlight the strong connection between NE- and MPO-DNA complexes. Taken together these data show a clear association of the neutrophil-derived enzyme-DNA complexes and COVID-19, which is also detectable to a lesser extent in patients with PCS.
NE-DNA and MPO-DNA complexes are of minor importance for chronic rheumatic diseases
To gain a more comprehensive understanding of the patterns of NET degradation markers in chronic rheumatic diseases, the PCA was repeated, excluding viral-induced pathologies such as COVID-19 and PCS. The PCA plot of variables (Figure 6A) displays the same co-variabilities of the markers, as seen in Figure 4A – NE-activity and citH3, as well as NE- and MPO-DNA complexes. However, the complexes now show a opposite influence in the variance than NE and citH3. With the highest cos2 value, cfDNA remains the marker that contributes the most to variability. The order of the other markers changes in this context to citH3, NE-activity, NE-DNA, and finally MPO-DNA complexes. Two larger clusters appear in the PCA plots of patients (Figure 6B), divided by both components. Component 1 explains 45.8%, component 2 37.7% of the variance. One of these clusters consists mainly of dots representing patients with SLE (blue), the other of dots representing patients with RA (red).
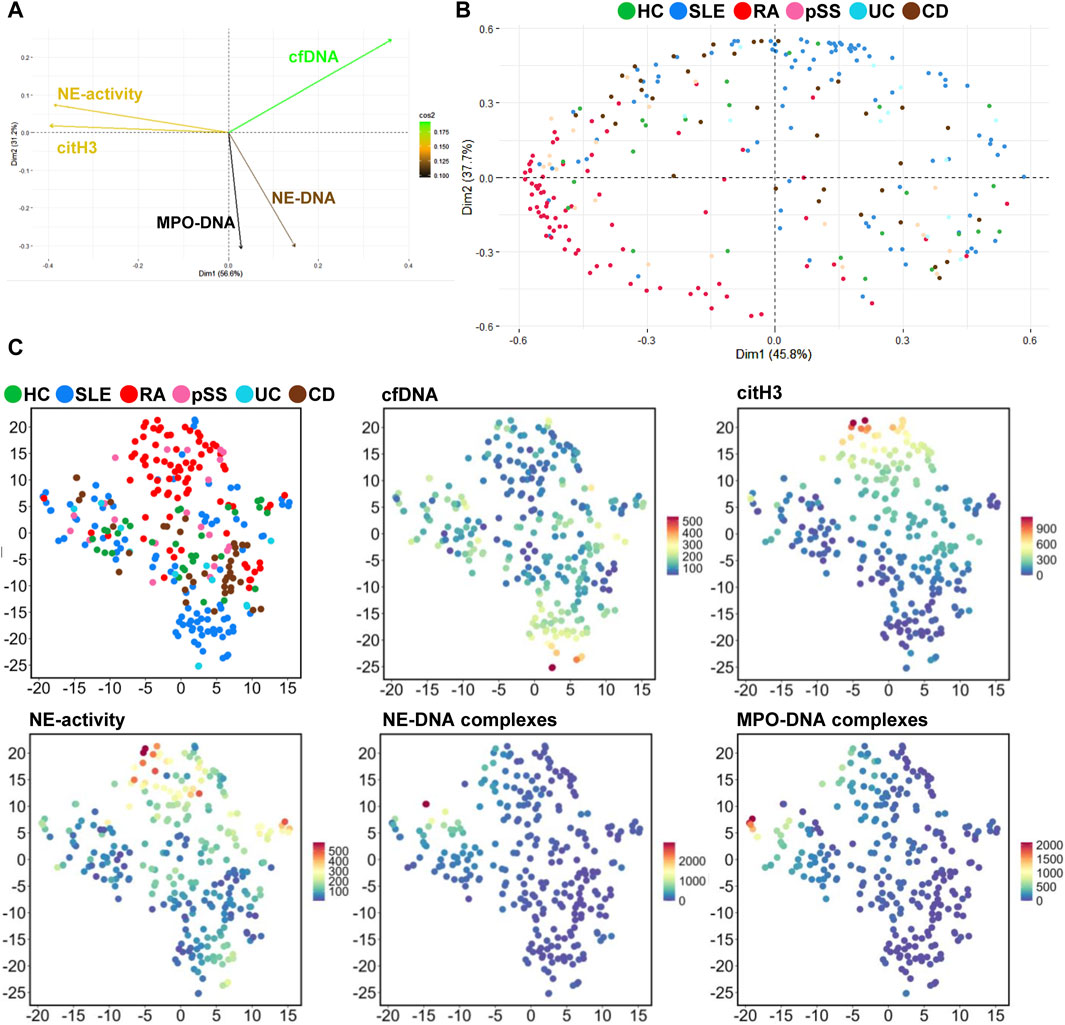
Figure 6. Dimensionality reduction of NET associated parameters in chronic rheumatic diseases. (A) PCA plot of the NET associated parameters. Markers with co-variability are clustered in the same quadrant. High cos2 values are depicted in green (0.175), low values in black (0.100), and intermediate values by a spectrum from yellow to brown (0.253-0.150). (B) PCA plot of NET associated parameters. Each patient cohort is assigned to a color to identify representing dots. Component 1 (Dim1) explains 45.8%, component 2 (Dim2) 37.7% of the variability. The square cosine value (cos2) indicates the variable representation. (C) A tSNE plot of the patient cohorts and NET associated parameters visualizes disease clusters. Each dot represents a patient, different colors the diseases. Further tSNE plots visualize the distribution of the NET associated parameters and their measured levels. High values are shown in red, low values in dark blue, and intermediate values in a spectrum from orange to light blue (representing higher to lower values).
The tSNE plot which shows the distribution of diseases, highlights the clustering into two main groups containing either patients with RA (red) or SLE (blue) (Figure 6C). A small cluster of patients with CD is located near the cluster of SLE (dark brown). A similar distribution pattern is visible for patients with UC (bright blue). The dots representing patients with pSS (pink) are well distributed in the plot, with a small accumulation in near the cluster of RA. The tSNE plot of the distribution and concentration of cfDNA now shows a clear accumulation of high values in the area of the SLE cluster. The markers citH3 and NE-activity are elevated in the area of the cluster of patients with RA. Few dots that represent high levels of NE- or MPO-DNA complexes can be identified. These data indicate that NE- and MPO-DNA complexes are primarily characteristic of patients with the acute COVID-19 and play a minor role in the chronic rheumatic diseases SLE and RA.
SLE is associated with high levels of cfDNA, while RA is mainly associated with elevated levels of citH3 and high NE-activity
Considering that SLE and RA represent the two main clusters in the group of chronic and rheumatic diseases, we performed detailed correlation analyzes with these cohorts compared to the HC group. The distinct clustering with predominantly SLE or RA patients is illustrated in tSNE plots, each highlighting the distribution of one group at a time (Figure 7A). The correlation analyzes were again performed compared to cfDNA, the marker with the highest impact on the data’s variability (Figure 7B). Given that NE- and MPO-DNA complexes play a minor role in SLE and RA, we focused on citH3 and NE-activity. Both graphs depict a clear distribution of dots representing SLE and RA. While patients with SLE exhibit high values for cfDNA, patients with RA show elevated levels of citH3 and NE-activity. Both markers significantly correlate with cfDNA (p < 0.0001).
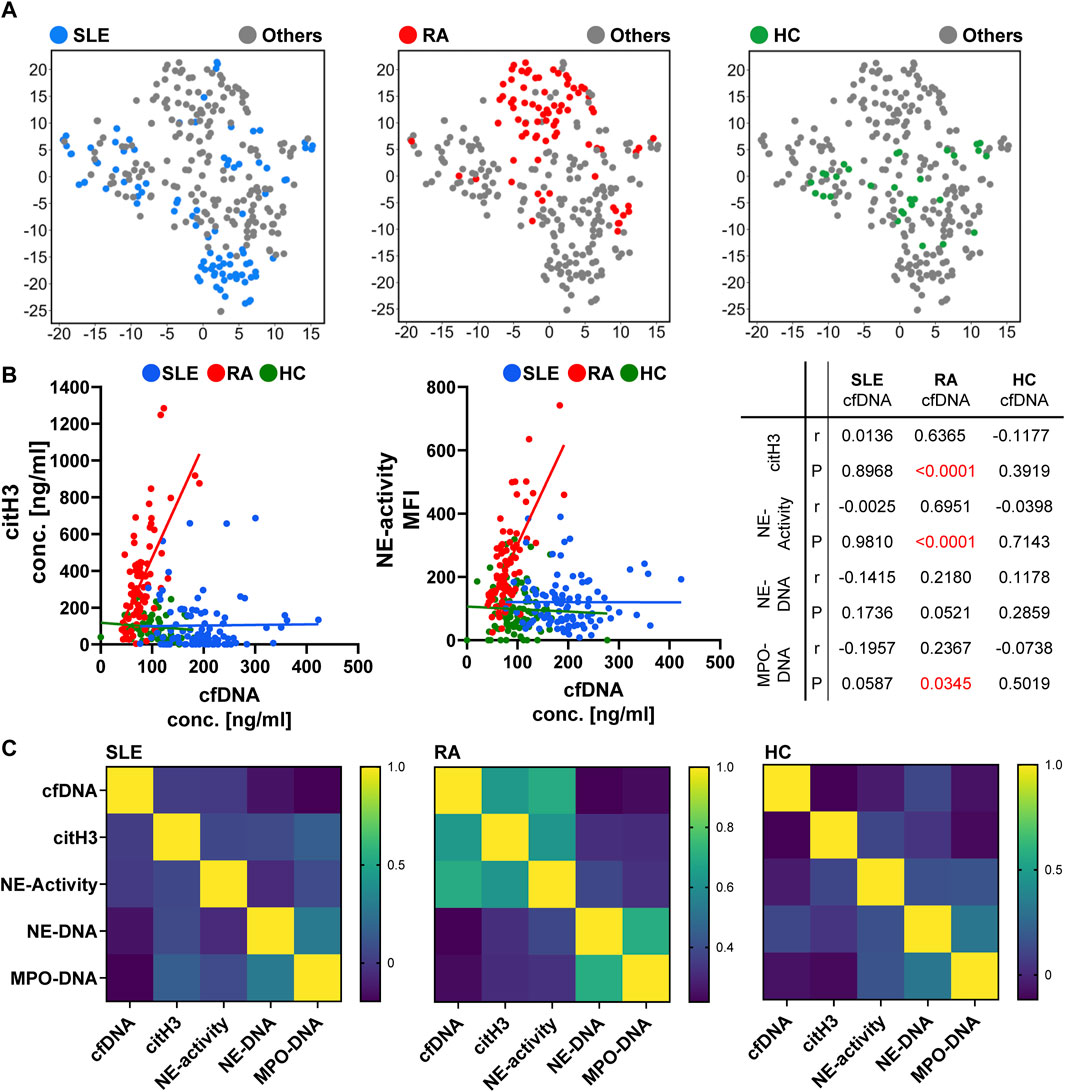
Figure 7. Detailed analyzes of patients with SLE and RA. (A) tSNE plots highlighting the differences in the distribution of patients with SLE (blue), RA (red), and the HC group (green). (B) Correlation analyzes between cfDNA and the NET-degradation markers citH3, and NE-activity. Each dot represents a patient–SLE in blue, RA in red, and HCs in green. Information about the statistical parameter r and the significance P are presented in table. (C) Heatmaps of the correlation analyzes. High values are depicted in yellow (1.0), low values in dark blue (0), and intermediate values in a color gradient from light green to blue (high to low values, respectively). For all correlation analyzes shown in this figure, a Two-tailed Pearson correlation was performed (p < 0.05).
The correlations are illustrated as heatmaps (Figure 7C) to visualize distinct patterns of NET degradation markers and NET associated parameters of SLE compared to RA patients. Patients with SLE only show high levels of cfDNA without correlation to other markers. In contrast, patients with RA exhibit high citH3 concentrations and NE-activity, both correlating with cfDNA, and peptide-DNA complexes, only correlating with each other. Together, these data indicate a characteristic pattern of NET degradation markers for the investigated inflammatory diseases.
The NET degradation markers in SLE and RA are independent of disease activity
To investigate whether the characteristic patterns observed for SLE and RA are associated with disease severity, we analyzed the data accordingly. Patients with SLE were classified employing their ECLAM scores (Figure 8A). Compared to the HC group, only the values of the cfDNA concentration in the serum of SLE patients were significantly increased. No significant differences between the SLE groups with low or high activity are seen, defined as ECLAM 1-5 and 6-14, respectively, although there was a trend for the DNA-enzyme complexes. The distribution in the characteristic pattern in the tSNE plot appears similar between patients with low and higher disease activities. Similar trends can be seen for patients with RA (Figure 8B). The NET markers citH3, NE-activity, and NE-DNA complexes depicted significantly increased values. There was only a trend that distinguished the RA groups of low and high activity, defined as DAS28 < 2.6 or DAS28 > 4, respectively. The distribution in the tSNE plot is comparable for both groups. Correlation analyses of the NET degradation data and the markers for disease activity did not reveal any significant relationships (not shown). Taken together, this suggests that the characteristically patterns of NET degradation markers for SLE and RA are independent of the disease activity.
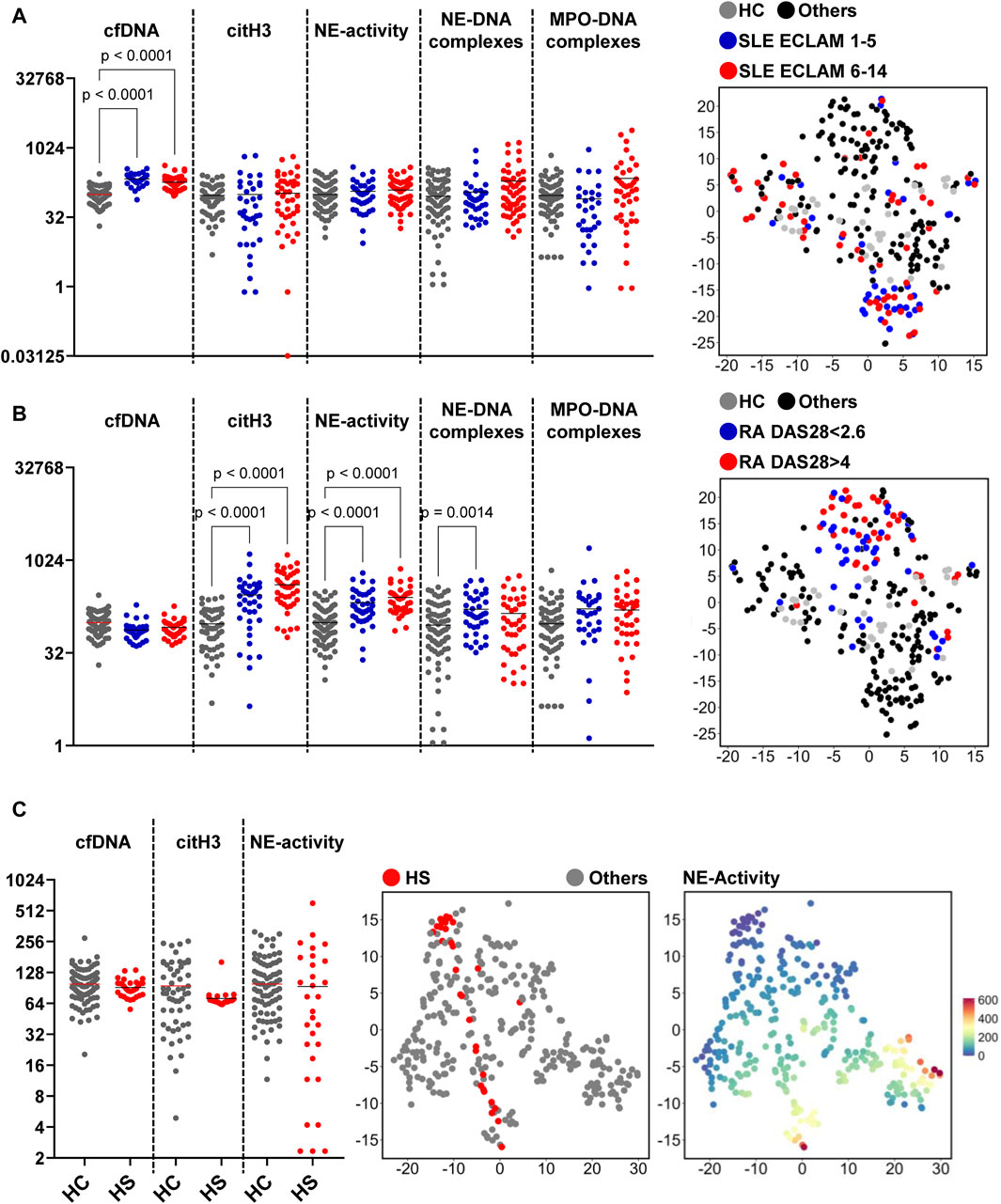
Figure 8. NET associated parameters in the context of disease activity. (A, B) Patients with low activity (SLE ECLAM values 1-5; RA DAS28 < 2.6) and high activity (SLE ECLAM values 6-13.5; RA DAS28 > 4) are represented in dot plots, highlighting differences in NET associated parameters and a tSNE plot to visualize the distribution. (Kruskal–Wallis p < 0.05) (C) NET associated parameters of patients with HS compared to HCs are shown in a dot plot. The distribution of the patients depending on the markers, and the distribution of high and low values of NE-activity are illustrated as tSNE plots. Patients with HS are depicted in red, HCs in grey. High values of NE-activity shown in red, low values in dark blue, and intermediate values in a spectrum from orange to light blue (representing higher to lower values). (Mann-Whitney test p < 0.05). Note all data are displayed using logarithmic scaling.
An additionally tested cohort of patients with HS, however, showed a wide distribution of the values of NE-activity (Figure 8C). The tSNE plot that highlights the HS cohort revealed a noticeable straight line of high to low NE-activity. The activity of NE correlated with levels of triglyceride and glucose (Supplementary Figure S2). This indicates that the NET degradation patterns may depend on activity and inflammation in the context of some other diseases. Non-significant correlations (AST, ALT, thrombocytes, albumin, cholesterol, HDL, LDL, GGT, HbA1c, age, weght, BMI, waist circ.) were not shown.
Discussion
Neutrophils and NETs impact the development, pathogenesis, and healing of inflammatory diseases. Their role in these processes is multifaceted ranging from inflammation and tissue degradation, to vessel and ducts occlusion, and eventually the resolution of inflammation (Mayadas et al., 2014; Boeltz et al., 2019). It is thus of great importance to explore detailed mechanisms and processes of NETs in diseases. This study focused on NET degradation products in inflammatory pathologies. NETs are cleared by nucleases and phagocytes (Lazzaretto and Fadeel, 2019; Demkow, 2023). Incomplete clearance of NETs leads to the accumulation of remnants, potentially exacerbating pathological conditions. NETs consist of chromatin bundles, decorated with proteins (Tecchio et al., 2014). Degradation leads to the release of cfDNA, toxic proteins like citH3, enzymes like NE, and NE- or MPO-DNA complexes. Although several other mechanisms than NETs have been described as source of cfDNA, leukocytes are demonstrably its main source (Oberhofer et al., 2022). We examined the concentrations or activity of these factors in samples from patients with various diseases. The different NET degradation markers were detectable to varying degrees in patients with SLE, RA, pSS, UC, MC, COVID-19, and PCS. We found that NET degradation markers exhibit distinct patterns rather than universal presence across these diseases.
In patients with COVID-19, we found a pronounced enrichment of NE- and MPO-DNA complexes. This acute inflammatory immune disease is associated with extensive vascular occlusions, facilitated by the formation and aggregation of NETs in the vessels, especially those of the microvasculature. These aggregates contribute to vascular damage and organ malfunction in COVID-19 (Leppkes et al., 2020; Singh et al., 2022). The increased concentration of enzyme-DNA complexes may stand in direct connection to the excessive formation of aggNETs. Within the aggregates, some of the chromatin bundles are not accessible and escape cleavage by nucleases. This may lead to the release of larger DNA fragments, still associated with the proteins.
Patients with SLE exhibit predominantly elevated concentrations of cfDNA in their serum, without an association with peptides. SLE is characterized not only by increased NET formation but also by compromised NET clearance. In the germinal centers of the lymph nodes, extracellular chromatin serves as selecting antigen for the stimulation of auto-reactive B-cells that produce autoantibodies directed against the body’s own DNA (Baumann et al., 2002; Gomez-Banuelos et al., 2023). Antibodies bound to the chromatin structure, stabilize NET constructs and hinder their degradation (Zuo et al., 2021). The reportedly reduced DNase activity in patients with SLE delays the clearance of NETs (Chitrabamrung et al., 1981; Al-Mayouf et al., 2011; Leffler et al., 2013). It is fair to argue that the reduced DNase activity, in combination with the stabilized NET-IgG adducts, interfere with proper degradation into smaller chromatin fragments, leaving behind an increased concentration of aggregated cfDNA (Hakkim et al., 2010). Steric hindrance may also explain the low enzyme-DNA complexes due to a low penetrance of the detection antibodies into these aggregates.
In our study, samples form patients with RA were mainly associated with increased concentrations of citH3 and higher NE-activity. Hypercitrullination is a key element in the pathogenesis of RA (Darrah and Andrade, 2018). The elevated enzymatic activity of PAD4 in tissues of RA patients has been associated with the appearance and progression of RA since it promotes the formation of neoantigens by hypercitrullination of vimentin, fibrinogen, type II collagen, and α-enolase (10.1186/ar1845). The activity of the enzyme in serum has been only recently reported elevated in a subgroup of patients with RA (10.3390/cimb44090293). This is in line with the increased amount of citH3 observed in the circulation of our patients. Increased activity of NE is associated with joint inflammation in RA (Muley et al., 2016), reflected by the high NE-activity detectable in serum samples.
Patients with the inflammatory bowel diseases UC and CD showed a tendency towards the SLE cluster in the tSNE plots and had elevated cfDNA levels. In addition, a notable decrease in NE-activity was observed, along with a widespread distribution of enzyme-DNE complexes present. The activity of NE varies depending on whether it is bound to DNA or free (Podolska et al., 2019). Together, this suggests the presence of NET aggregates, which, similar to the situation in SLE, may hinder the effective and thorough clearance of NETs. More studies with larger groups of patients are required to verify this theory.
Patients with pSS showed a broad distribution in the tSNE plots and only a significant increase in citH3 in our study. Previous studies reported an increase in cfDNA compared to HCs (Peng et al., 2022). These results emphasize the heterogeneity within this patient group and highlight the necessity for more extensive studies to draw distinct conclusions.
Our results revealed characteristic patterns of NET degradation products in inflammatory diseases. All examined conditions of disease were associated with increased neutrophil activity and NET formation in general. However, certain markers were disease-specifically increased, reflecting different mechanisms in NET formation and clearance. These distinctions may be attributed to hallmark pathologies characteristic of each course of disease. No significant differences were to be observed for patients with higher or lower activities of SLE or RA; the characteristic patterns of NET degradation products were related to the disease pathology and not to its severity.
Patients with HS showed a wide distribution of NE-activity. Correlation analyzes with clinical parameters revealed a statistically significant connection to triglycerides and glucose levels. Patients with HS show a prevalence of the metabolic syndrome (Sabat et al., 2012). The connection of NE with these metabolic markers requires further examination.
In summary, each phase of NET formation, NET degradation and the components related to NETs and their degradation are intertwined with the pathologies of the diseases under investigation. All markers - cfDNA, citH3, NE activity, NE and MPO DNA complexes - are found in the circulation of patients with inflammatory diseases (Wang et al., 2018a; Xu et al., 2018; Ng et al., 2021; Reshetnyak et al., 2023). However, the significant increase in specific markers for NET degradation in these diseases provides valuable insights into potential key targets for the development of differential therapies.
Conclusion
We investigated NET degradation products as distinctive patterns characteristic of inflammatory diseases. Different selective patterns were identified for three diseases - COVID-19, SLE and RA. COVID-19, SLE, and RA were primarily associated with increased levels of enzyme-DNA complexes, cfDNA, and citH3, respectively. RA was furthermore characterized by higher NE-activity. These different patterns offer potential for understanding the role of NETs in inflammatory diseases and may promote the development of treatments that target the disease-specific NET degradation products.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by Ethical Committee of the University Hospital Erlangen (permit #193_13B and # 174_20B); Ethics Committee (107/20) of the University of Würzburg. The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from primarily isolated as part of your previous study for which ethical approval was obtained. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and institutional requirements.
Author contributions
JS: Writing–original draft, Writing–review and editing, Data curation, Formal Analysis, Investigation, Conceptualization. MM-B: Writing–review and editing, Data curation, Formal Analysis, Validation. JK: Writing–review and editing, Investigation, Validation. FN: Writing–review and editing, Investigation. ML: Writing–review and editing, Resources. DR: Writing–review and editing, Resources. AZ: Writing–review and editing, Resources. VF: Writing–review and editing, Resources. BH: Writing–review and editing, Resources. VZ: Writing–review and editing, Resources. LM: Writing–review and editing, Data curation, Formal Analysis, Resources. GS: Writing–review and editing, Funding acquisition, Resources. MH: Writing–original draft, Writing–review and editing, Project administration, Supervision. CS: Writing–original draft, Writing–review and editing, Funding acquisition, Project administration, Supervision.
Funding
The authors declare that financial support was received for the research, authorship, and/or publication of this article. The authors acknowledge the support by Deutsche Forschungsgemeinschaft (DFG), and Friedrich-Alexander-Universität Erlangen-Nürnberg (FAU). This work was partially supported by the Deutsche Forschungsgemeinschaft (DFG) SCHA 2040/1-1 to CS, by the Deutsche Forschungsgemeinschaft (DFG) 2886 PANDORA Project-No. B3; CRC1181- 261193037 (C03); SFB/TRR 241 (B04); the Volkswagen-Stiftung (Grant 97744) to MH, and by DFG MU 4240/2-1 (Project Nr. 470134687). This work is supported by the ELAN Fonds of the Universitätsklinikum Erlangen (grant number P130) to CS. This work was partially supported by the Deutsche Forschungsgemeinschaft (DFG; GRK2504/1, Project Nr. 401821119, subproject C6) to BH and VZ, the Federal Ministry of Education and Research (BMBF), Advanced Clinician Scientists (ACS, Grant 01EO2105) to BH and the Bavarian Health and Food Safety Authority (LGL, disCOVer 2.0, Gesamt-2490-PC-2021-V14) to MH and BH.
Acknowledgments
Figures 1, 2 of the introduction were created with BioRender.com and are licensed for publication.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fddsv.2024.1404103/full#supplementary-material
SUPPLEMENTARY FIGURE S1 | Quantitative comparison of cfDNA concentrations in paired serum and plasma samples of healthy controls. A Student’s t-test for paired samples was applied (n = 40).
SUPPLEMENTARY FIGURE S2 | Clinical parameters and NE-activity in HS. Correlation analyzes of measured activity of NE in serum samples from patients with HS and clinical parameters. Each dot represents one patient. Triglyceride values are shown in red, Glucose levels in purple. For all correlation analyzes shown in this figure, a Two-tailed Pearson correlation was performed (P < 0.05).
Abbreviations
ACPA, autoantibodies against citrullinated antigens; aggNETs, aggregated NETs; citH3, citrullinated Histone H3; cos2, square cosine values; C1q, complement factor 1q; DAS28, Disease Activity Score 28; ECLAM, European Consensus Lupus Activity Measurement; MPO, myeloperoxidase; NE, neutrophil elastase; NET, neutrophil extracellular trap; PCA, principal component analysis; RA, rheumatoid arthritis; ROS, reactive oxygen species; RT, room temperature; SLE, systemic lupus erythematosus; TREX, 3′-exonucleases; t-SNE, t-Stochastic Neighbor Embedding.
References
AL-Mayouf, S. M., Sunker, A., Abdwani, R., Abrawi, S. A., Almurshedi, F., Alhashmi, N., et al. (2011). Loss-of-function variant in DNASE1L3 causes a familial form of systemic lupus erythematosus. Nat. Genet. 43, 1186–1188. doi:10.1038/ng.975
Baumann, I., Kolowos, W., Voll, R. E., Manger, B., Gaipl, U., Neuhuber, W. L., et al. (2002). Impaired uptake of apoptotic cells into tingible body macrophages in germinal centers of patients with systemic lupus erythematosus. Arthritis Rheum. 46, 191–201. doi:10.1002/1529-0131(200201)46:1<191::AID-ART10027>3.0.CO;2-K
Bennike, T. B., Carlsen, T. G., Ellingsen, T., Bonderup, O. K., Glerup, H., Bogsted, M., et al. (2015). Neutrophil extracellular traps in ulcerative colitis: a proteome analysis of intestinal biopsies. Inflamm. Bowel Dis. 21, 2052–2067. doi:10.1097/MIB.0000000000000460
Boeltz, S., Amini, P., Anders, H. J., Andrade, F., Bilyy, R., Chatfield, S., et al. (2019). To NET or not to NET:current opinions and state of the science regarding the formation of neutrophil extracellular traps. Cell Death Differ. 26, 395–408. doi:10.1038/s41418-018-0261-x
Brinkmann, V., Reichard, U., Goosmann, C., Fauler, B., Uhlemann, Y., Weiss, D. S., et al. (2004). Neutrophil extracellular traps kill bacteria. Science 303, 1532–1535. doi:10.1126/science.1092385
Burgener, S. S., and Schroder, K. (2020). Neutrophil extracellular traps in host defense. Cold Spring Harb. Perspect. Biol. 12, a037028. doi:10.1101/cshperspect.a037028
Burn, G. L., Foti, A., Marsman, G., Patel, D. F., and Zychlinsky, A. (2021). The neutrophil. Immunity 54, 1377–1391. doi:10.1016/j.immuni.2021.06.006
Carmona-Rivera, C., Zhao, W., Yalavarthi, S., and Kaplan, M. J. (2015). Neutrophil extracellular traps induce endothelial dysfunction in systemic lupus erythematosus through the activation of matrix metalloproteinase-2. Ann. Rheum. Dis. 74, 1417–1424. doi:10.1136/annrheumdis-2013-204837
Chatfield, S. M., Thieblemont, N., and Witko-Sarsat, V. (2018). Expanding neutrophil horizons: new concepts in inflammation. J. Innate Immun. 10, 422–431. doi:10.1159/000493101
Chitrabamrung, S., Rubin, R. L., and Tan, E. M. (1981). Serum deoxyribonuclease I and clinical activity in systemic lupus erythematosus. Rheumatol. Int. 1, 55–60. doi:10.1007/BF00541153
Darrah, E., and Andrade, F. (2018). Rheumatoid arthritis and citrullination. Curr. Opin. Rheumatol. 30, 72–78. doi:10.1097/BOR.0000000000000452
Demkow, U. (2023). Molecular mechanisms of neutrophil extracellular trap (NETs) degradation. Int. J. Mol. Sci. 24, 4896. doi:10.3390/ijms24054896
Dos Santos Ramos, A., Viana, G. C. S., DE Macedo Brigido, M., and Almeida, J. F. (2021). Neutrophil extracellular traps in inflammatory bowel diseases: implications in pathogenesis and therapeutic targets. Pharmacol. Res. 171, 105779. doi:10.1016/j.phrs.2021.105779
Farrera, C., and Fadeel, B. (2013). Macrophage clearance of neutrophil extracellular traps is a silent process. J. Immunol. 191, 2647–2656. doi:10.4049/jimmunol.1300436
Fuchs, T. A., Abed, U., Goosmann, C., Hurwitz, R., Schulze, I., Wahn, V., et al. (2007). Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 176, 231–241. doi:10.1083/jcb.200606027
Fu, X., Liu, H., Huang, G., and Dai, S. S. (2021). The emerging role of neutrophils in autoimmune-associated disorders: effector, predictor, and therapeutic targets. MedComm (2020) 2, 402–413. doi:10.1002/mco2.69
Gomez-Banuelos, E., Fava, A., and Andrade, F. (2023). An update on autoantibodies in systemic lupus erythematosus. Curr. Opin. Rheumatol. 35, 61–67. doi:10.1097/BOR.0000000000000922
Grayson, P. C., and Kaplan, M. J. (2016). At the Bench: neutrophil extracellular traps (NETs) highlight novel aspects of innate immune system involvement in autoimmune diseases. J. Leukoc. Biol. 99, 253–264. doi:10.1189/jlb.5BT0615-247R
Hahn, J., Schauer, C., Czegley, C., Kling, L., Petru, L., Schmid, B., et al. (2019). Aggregated neutrophil extracellular traps resolve inflammation by proteolysis of cytokines and chemokines and protection from antiproteases. FASEB J. 33, 1401–1414. doi:10.1096/fj.201800752R
Haider, P., Kral-Pointner, J. B., Mayer, J., Richter, M., Kaun, C., Brostjan, C., et al. (2020). Neutrophil extracellular trap degradation by differently polarized macrophage subsets. Arterioscler. Thromb. Vasc. Biol. 40, 2265–2278. doi:10.1161/ATVBAHA.120.314883
Hakkim, A., Furnrohr, B. G., Amann, K., Laube, B., Abed, U. A., Brinkmann, V., et al. (2010). Impairment of neutrophil extracellular trap degradation is associated with lupus nephritis. Proc. Natl. Acad. Sci. U. S. A. 107, 9813–9818. doi:10.1073/pnas.0909927107
Hu, Z., Murakami, T., Tamura, H., Reich, J., Kuwahara-Arai, K., Iba, T., et al. (2017). Neutrophil extracellular traps induce IL-1β production by macrophages in combination with lipopolysaccharide. Int. J. Mol. Med. 39, 549–558. doi:10.3892/ijmm.2017.2870
Jimenez-Alcazar, M., Napirei, M., Panda, R., Kohler, E. C., Kremer Hovinga, J. A., Mannherz, H. G., et al. (2015). Impaired DNase1-mediated degradation of neutrophil extracellular traps is associated with acute thrombotic microangiopathies. J. Thromb. Haemost. 13, 732–742. doi:10.1111/jth.12796
Kaplan, M. J., and Radic, M. (2012). Neutrophil extracellular traps: double-edged swords of innate immunity. J. Immunol. 189, 2689–2695. doi:10.4049/jimmunol.1201719
Khandpur, R., Carmona-Rivera, C., Vivekanandan-Giri, A., Gizinski, A., Yalavarthi, S., Knight, J. S., et al. (2013). NETs are a source of citrullinated autoantigens and stimulate inflammatory responses in rheumatoid arthritis. Sci. Transl. Med. 5, 178ra40. doi:10.1126/scitranslmed.3005580
Khudhair, H. A. A. (2023). A study of the roles of some immunological biomarkers in the diagnosis of rheumatoid arthritis. J. Med. Life 16, 1194–1200. doi:10.25122/jml-2023-0158
Klebanoff, S. J., Kettle, A. J., Rosen, H., Winterbourn, C. C., and Nauseef, W. M. (2013). Myeloperoxidase: a front-line defender against phagocytosed microorganisms. J. Leukoc. Biol. 93, 185–198. doi:10.1189/jlb.0712349
Knopf, J., Mahajan, A., Munoz, L. E., and Herrmann, M. (2022). Formation and clearance of NETs in Health and disease. Cells 11, 4022. doi:10.3390/cells11244022
Laukova, L., Konecna, B., Janovicova, L., Vlkova, B., and Celec, P. (2020). Deoxyribonucleases and their applications in biomedicine. Biomolecules 10, 1036. doi:10.3390/biom10071036
Lazzaretto, B., and Fadeel, B. (2019). Intra- and extracellular degradation of neutrophil extracellular traps by macrophages and dendritic cells. J. Immunol. 203, 2276–2290. doi:10.4049/jimmunol.1800159
Leffler, J., Gullstrand, B., Jonsen, A., Nilsson, J. A., Martin, M., Blom, A. M., et al. (2013). Degradation of neutrophil extracellular traps co-varies with disease activity in patients with systemic lupus erythematosus. Arthritis Res. Ther. 15, R84. doi:10.1186/ar4264
Leppkes, M., Knopf, J., Naschberger, E., Lindemann, A., Singh, J., Herrmann, I., et al. (2020). Vascular occlusion by neutrophil extracellular traps in COVID-19. EBioMedicine 58, 102925. doi:10.1016/j.ebiom.2020.102925
Li, P., Li, M., Lindberg, M. R., Kennett, M. J., Xiong, N., and Wang, Y. (2010). PAD4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. J. Exp. Med. 207, 1853–1862. doi:10.1084/jem.20100239
Lou, H., Wojciak-Stothard, B., Ruseva, M. M., Cook, H. T., Kelleher, P., Pickering, M. C., et al. (2020). Autoantibody-dependent amplification of inflammation in SLE. Cell Death Dis. 11, 729. doi:10.1038/s41419-020-02928-6
Makrygiannakis, D., Af Klint, E., Lundberg, I. E., Lofberg, R., Ulfgren, A. K., Klareskog, L., et al. (2006). Citrullination is an inflammation-dependent process. Ann. Rheum. Dis. 65, 1219–1222. doi:10.1136/ard.2005.049403
Mayadas, T. N., Cullere, X., and Lowell, C. A. (2014). The multifaceted functions of neutrophils. Annu. Rev. Pathol. 9, 181–218. doi:10.1146/annurev-pathol-020712-164023
Mulay, S. R., and Anders, H. J. (2020). Neutrophils and neutrophil extracellular traps regulate immune responses in Health and disease. Cells 9, 2130. doi:10.3390/cells9092130
Muley, M. M., Reid, A. R., Botz, B., Bolcskei, K., Helyes, Z., and Mcdougall, J. J. (2016). Neutrophil elastase induces inflammation and pain in mouse knee joints via activation of proteinase-activated receptor-2. Br. J. Pharmacol. 173, 766–777. doi:10.1111/bph.13237
Nakazawa, D., Kumar, S. V., Marschner, J., Desai, J., Holderied, A., Rath, L., et al. (2017). Histones and neutrophil extracellular traps enhance tubular necrosis and remote organ injury in ischemic AKI. J. Am. Soc. Nephrol. 28, 1753–1768. doi:10.1681/ASN.2016080925
Ng, H., Havervall, S., Rosell, A., Aguilera, K., Parv, K., Von Meijenfeldt, F. A., et al. (2021). Circulating markers of neutrophil extracellular traps are of prognostic value in patients with COVID-19. Arterioscler. Thromb. Vasc. Biol. 41, 988–994. doi:10.1161/ATVBAHA.120.315267
Oberhofer, A., Bronkhorst, A. J., Uhlig, C., Ungerer, V., and Holdenrieder, S. (2022). Tracing the origin of cell-free DNA molecules through tissue-specific epigenetic signatures. Diagn. (Basel) 12, 1834. doi:10.3390/diagnostics12081834
Ogawa, Y., Muto, Y., Kinoshita, M., Shimada, S., and Kawamura, T. (2021). Neutrophil extracellular traps in skin diseases. Biomedicines 9, 1888. doi:10.3390/biomedicines9121888
Okeke, E. B., Louttit, C., Fry, C., Najafabadi, A. H., Han, K., Nemzek, J., et al. (2020). Inhibition of neutrophil elastase prevents neutrophil extracellular trap formation and rescues mice from endotoxic shock. Biomaterials 238, 119836. doi:10.1016/j.biomaterials.2020.119836
Oliveira, C. B., Byrd, A. S., Okoye, G. A., Kaplan, M. J., and Carmona-Rivera, C. (2023). Neutralizing anti‒DNase 1 and ‒DNase 1L3 antibodies impair neutrophil extracellular traps degradation in hidradenitis suppurativa. J. Invest. Dermatol 143, 57–66. doi:10.1016/j.jid.2022.06.024
Papayannopoulos, V. (2018). Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 18, 134–147. doi:10.1038/nri.2017.105
Peng, Y., Wu, X., Zhang, S., Deng, C., Zhao, L., Wang, M., et al. (2022). The potential roles of type I interferon activated neutrophils and neutrophil extracellular traps (NETs) in the pathogenesis of primary Sjogren's syndrome. Arthritis Res. Ther. 24, 170. doi:10.1186/s13075-022-02860-4
Podolska, M. J., Mahajan, A., Hahn, J., Knopf, J., Maueroder, C., Petru, L., et al. (2019). Treatment with DNases rescues hidden neutrophil elastase from aggregated NETs. J. Leukoc. Biol. 106, 1359–1366. doi:10.1002/JLB.3AB0918-370R
Reshetnyak, T., Nurbaeva, K., Ptashnik, I., Kudriaeva, A., Belogurov, A., Lila, A., et al. (2023). Markers of NETosis in patients with systemic lupus erythematosus and antiphospholipid syndrome. Int. J. Mol. Sci. 24, 9210. doi:10.3390/ijms24119210
Sabat, R., Chanwangpong, A., Schneider-Burrus, S., Metternich, D., Kokolakis, G., Kurek, A., et al. (2012). Increased prevalence of metabolic syndrome in patients with acne inversa. PLoS One 7, e31810. doi:10.1371/journal.pone.0031810
Schoen, J., Euler, M., Schauer, C., Schett, G., Herrmann, M., Knopf, J., et al. (2022). Neutrophils' extracellular trap mechanisms: from physiology to pathology. Int. J. Mol. Sci. 23, 12855. doi:10.3390/ijms232112855
Schroder, A. L., Chami, B., Liu, Y., Doyle, C. M., El Kazzi, M., Ahlenstiel, G., et al. (2022). Neutrophil extracellular trap density increases with increasing histopathological severity of crohn’s disease. Inflamm. Bowel Dis. 28, 586–598. doi:10.1093/ibd/izab239
Segal, A. W. (2005). How neutrophils kill microbes. Annu. Rev. Immunol. 23, 197–223. doi:10.1146/annurev.immunol.23.021704.115653
Shafqat, A., Omer, M. H., Albalkhi, I., Alabdul Razzak, G., Abdulkader, H., Abdul Rab, S., et al. (2023). Neutrophil extracellular traps and long COVID. Front. Immunol. 14, 1254310. doi:10.3389/fimmu.2023.1254310
Singh, J., Boettcher, M., Dolling, M., Heuer, A., Hohberger, B., Leppkes, M., et al. (2023). Moonlighting chromatin: when DNA escapes nuclear control. Cell Death Differ. 30, 861–875. doi:10.1038/s41418-023-01124-1
Singh, J., Herrmann, I., Mahajan, A., Schauer, C., Shan, X., Hartmann, A., et al. (2022). A pleomorphic puzzle: heterogeneous pulmonary vascular occlusions in patients with COVID-19. Int. J. Mol. Sci. 23, 15126. doi:10.3390/ijms232315126
Tecchio, C., Micheletti, A., and Cassatella, M. A. (2014). Neutrophil-derived cytokines: facts beyond expression. Front. Immunol. 5, 508. doi:10.3389/fimmu.2014.00508
Villanueva, E., Yalavarthi, S., Berthier, C. C., Hodgin, J. B., Khandpur, R., Lin, A. M., et al. (2011). Netting neutrophils induce endothelial damage, infiltrate tissues, and expose immunostimulatory molecules in systemic lupus erythematosus. J. Immunol. 187, 538–552. doi:10.4049/jimmunol.1100450
Vorobjeva, N. V. (2020). Neutrophil extracellular traps: new aspects. Mosc. Univ. Biol. Sci. Bull. 75, 173–188. doi:10.3103/S0096392520040112
Wang, H., Stehr, A. M., Singh, J., Zlatar, L., Hartmann, A., Evert, K., et al. (2023a). Anti-DNA-IgM favors the detection of NET-associated extracellular DNA. Int. J. Mol. Sci. 24, 4101. doi:10.3390/ijms24044101
Wang, P., Liu, D., Zhou, Z., Liu, F., Shen, Y., You, Q., et al. (2023b). The role of protein arginine deiminase 4-dependent neutrophil extracellular traps formation in ulcerative colitis. Front. Immunol. 14, 1144976. doi:10.3389/fimmu.2023.1144976
Wang, W., Peng, W., and Ning, X. (2018a). Increased levels of neutrophil extracellular trap remnants in the serum of patients with rheumatoid arthritis. Int. J. Rheum. Dis. 21, 415–421. doi:10.1111/1756-185X.13226
Wang, X., Qiu, L., Li, Z., Wang, X. Y., and Yi, H. (2018b). Understanding the multifaceted role of neutrophils in cancer and autoimmune diseases. Front. Immunol. 9, 2456. doi:10.3389/fimmu.2018.02456
Winterbourn, C. C., Kettle, A. J., and Hampton, M. B. (2016). Reactive oxygen species and neutrophil function. Annu. Rev. Biochem. 85, 765–792. doi:10.1146/annurev-biochem-060815-014442
Xu, Y., Song, Y., Chang, J., Zhou, X., Qi, Q., Tian, X., et al. (2018). High levels of circulating cell-free DNA are a biomarker of active SLE. Eur. J. Clin. Invest. 48, e13015. doi:10.1111/eci.13015
Yaykasli, K. O., Schauer, C., Munoz, L. E., Mahajan, A., Knopf, J., Schett, G., et al. (2021). Neutrophil extracellular trap-driven occlusive diseases. Cells 10, 2208. doi:10.3390/cells10092208
Keywords: degradation markers, cfDNA, MPO, neurophil elastase, inflammatory pattern
Citation: Schoen J, Muñoz-Becerra M, Knopf J, Ndukwe F, Leppkes M, Roth D, Zeitler A, Frings VG, Hohberger B, Zeisberg V, Muñoz LE, Schett G, Herrmann M and Schauer C (2024) The chronicles of inflammation: uncovering of distinct patterns of NET degradation products. Front. Drug Discov. 4:1404103. doi: 10.3389/fddsv.2024.1404103
Received: 20 March 2024; Accepted: 31 July 2024;
Published: 13 September 2024.
Edited by:
Ahmed Sheriff, Charité University Medicine Berlin, GermanyReviewed by:
Dongyi He, Shanghai Guanghua Rheumatology Hospital, ChinaChristian Maueröder, VIB-UGent Center for Inflammation Research (IRC), Belgium
Copyright © 2024 Schoen, Muñoz-Becerra, Knopf, Ndukwe, Leppkes, Roth, Zeitler, Frings, Hohberger, Zeisberg, Muñoz, Schett, Herrmann and Schauer. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martin Herrmann, bWFydGluLmhlcnJtYW5uQHVrLWVybGFuZ2VuLmRl
‡This work will be used as part of Janina Schoen’s doctoral thesis
†These authors have contributed equally to this work and share senior authorship