
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Conserv. Sci., 14 March 2025
Sec. Animal Conservation
Volume 6 - 2025 | https://doi.org/10.3389/fcosc.2025.1537563
This article is part of the Research TopicNew Challenges and Perspectives in Conservation Breeding ProgramsView all 9 articles
 David Javier Galindo1,2*
David Javier Galindo1,2* Pedro Henrique de Faria Peres1
Pedro Henrique de Faria Peres1 Eveline dos Santos Zanetti1
Eveline dos Santos Zanetti1 Márcio Leite de Oliveira1,3
Márcio Leite de Oliveira1,3 Luciana Diniz Rola1
Luciana Diniz Rola1 José Maurício Barbanti Duarte1*
José Maurício Barbanti Duarte1*Introduction: The loss of genetic diversity is a critical factor in the extinction process, exacerbated by anthropogenic pressures and demographic stochasticity, particularly in small populations. Traditional population restoration methods, such as individual translocation and ecological corridors, present challenges, including high costs, adaptation difficulties, pathogen introduction, and outbreeding risks. Embryo reintroduction has emerged as a potential strategy for genetic rescue. This study aimed to evaluate the feasibility of embryo transfer as a genetic rescue tool in the marsh deer (Blastocerus dichotomus), a species classified as Vulnerable by the IUCN.
Methods: Following the construction of the Sérgio Motta Hydroelectric Power Plant (UHSM) in 1998, a population of over 1,000 marsh deer was impacted, leading to the capture of 93 individuals for ex situ (82) and in situ (11) conservation efforts. Between 1998 and 2001, an experimental reintroduction program established a new population in a 2,000-hectare wetland near the Jataí Ecological Station (EEJ) in Luis Antônio, São Paulo, Brazil. Over time, this population reached a carrying capacity of 25 individuals but experienced genetic diversity loss. To address this, we conducted an embryo transfer experiment using a female from the reintroduced population as a receipt for embryos from the captive population. The female, captured during late pregnancy, was subjected to estrous cycle synchronization for embryo transfer after giving birth and being apart from its fawn. Two embryos from a captive population were implanted, and the female was re-released after 10 days. Monitoring was conducted via radio transmitter collar (GPS–GSM) and helicopter tracking.
Results: The female did not give birth to the implanted embryos but was recaptured nine months later for an ultrasonographic evaluation, which indicated a six-month pregnancy. This suggests that the implanted embryos were lost early in gestation, but natural fertilization occurred approximately three months post-release.
Discussion: This study demonstrates the technical feasibility of embryo reintroduction as a genetic rescue strategy, even though pregnancy was not carried to term. The ability to capture, temporarily hold, and successfully reintroduce a free-ranging female suggests minimal disruption to natural behaviors. Future improvements in embryo quality, hormonal protocols, and pregnancy confirmation prior to release could enhance the success rate of this method. Embryo reintroduction presents a promising alternative to traditional reintroduction methods, offering a novel approach to mitigating genetic risks in small, isolated populations.
Reintroduction programs are crucial for conserving species at risk of extinction, but their success depends on several factors, with genetic concerns playing a central role. A major challenge in reintroductions is the potential loss of genetic diversity, often exacerbated by the small number of individuals typically involved. This can lead to inbreeding and the founder effect. Inbreeding may lead to inbreeding depression and, along with genetic drift, results in the loss of genetic variability, negatively affecting fertility, survival, and overall population viability (Falconer, 1989). The founder effect occurs when a small subset of individuals is separated from the larger population, carrying only a fraction of the genetic diversity, which over time leads to allele loss, reduced heterozygosity, and decreased population fitness (Shorrock, 1980). Genetic and demographic founder effects can have long-term consequences for the colonizing populations, compounding the risks of inbreeding depression and hindering the population’s recovery (Szűcs et al., 2017). These genetic challenges are particularly concerning for species like the marsh deer (Blastocerus dichotomus), given its restricted distribution and the risk of future genetic bottlenecks, that threaten its survival.
The marsh deer is the largest deer species in South America, reaching up to 100 kg for females and 130 kg for males (Piovezan et al., 2010). It inhabits floodplains and wetland areas, historically ranging from the southern Amazon Rainforest to northeastern Argentina (Piovezan et al., 2010). Several populations are currently at risk of extinction due to poaching, livestock disease exposure, floodplain drainage for agricultural expansion, and the construction of hydroelectric dams; therefore, the species is categorized as vulnerable by the IUCN (Szabó et al., 2003, 2007; Piovezan et al., 2010; Duarte et al., 2016). These factors may cause severe population declines and reduced genetic variability due to inbreeding and genetic drift, impairing reproductive performance and limiting adaptive potential (Roldan et al., 2006; Duarte et al., 2012).
In the early 1990s, following the construction of the Três Irmãos Hydroelectric Power Plant on the Tietê River by Companhia Energética de São Paulo (CESP), 45 marsh deer were rescued and placed in captivity. This led to the creation of the Marsh Deer Captive Management Program, currently managed by Tijoá Energia. The program was operated at the Marsh Deer Conservation Center (CCCP) in Promissão, São Paulo, and the Ilha Solteira Zoo in Ilha Solteira, São Paulo. Then within the same decade, the construction of the Porto Primavera Hydroelectric Power Plant (UHSM) on the Paraná River in 1998 brought the last significant population of marsh deer in São Paulo to the brink of extinction. In response, the Porto Primavera Marsh Deer Project was established to study and conserve the affected population through ex situ conservation efforts, including the reintroduction of animals into remnant floodplain areas in São Paulo (Zanetti and Duarte, 2008).
In late 1998, the Marsh Deer Conservation Program was initiated by researchers from the Deer Research and Conservation Center (NUPECCE) at São Paulo State University (UNESP-Jaboticabal) to study the impact of the UHSM on the local marsh deer population. The project aims to conserve the species through ex situ programs to preserve genetic diversity and provide animals for reintroduction efforts, such as those performed at the Jataí Ecological Station (EEJ) in São Paulo (Figueira et al., 2005; Zanetti and Duarte, 2008). Despite initial challenges, a reintroduced population at the EEJ was established from eight individuals (three males and five females) between 1998 and 2001. Then, between July 2004 and June 2007, individuals from this population were captured for transmitter ear-tag placement, biological material collection, and monitoring, estimating the population size between 20 and 25 individuals (Ferreira, 2011). Although the loss of genetic diversity and inbreeding were not yet concerns, simulations indicated that long-term genetic diversity could only be maintained by introducing new individuals every four generations (Ferreira, 2011).
Given the challenges of translocation efforts, including high mortality rates and diseases risks (Warne and Chaber, 2023), embryo translocation (ET) offers a promising alternative for genetic management and disease risk reduction in reintroduced populations (Rola et al., 2021, 2023). However, to fully utilize ET for species conservation, it is crucial to develop effective superovulation protocols for generating embryos from captive animals. Reproductive studies on marsh deer provide important insights into their breeding biology, which can help to develop and adapt these protocols. Female marsh deer reach sexual maturity at around two years of age, with an average estrous cycle of 21.3 ± 1.3 days. They are polystric and uniparous, with an average pregnancy length of 253 ± 4 days and no reproductive seasonality, as evidenced by postpartum estrus (Polegato et al., 2018). Males display individual variation in antler cycles, with minimal synchronization and no apparent photoperiod influence, indicating year-round fertility (Ramos, 2004; Pereira, 2010). These basic reproductive characteristics are crucial for optimizing ET techniques and enhancing genetic management in conservation programs (Comizzoli, 2020).
Despite progress, the program faces challenges in maintaining optimal population sizes and genetic diversity, particularly concerning male management and institution capacity (Ferreira, 2011). The original population of 82 animals distributed in 18 Brazilian institutions had decreased to 67 individuals distributed in 12 institutions by 2020 (considering only the UHSM population). These challenges highlight the need for a genomic embryo bank to enhance genetic management and ensure the long-term success of the conservation program (Comizzoli, 2020; Rola et al., 2021).
This study aimed to bridge captive populations with wild counterparts by collecting embryos from the Brazilian captive population to establish a cryogenic bank for the species and developing an embryo reintroduction technique to assist the wild EEJ population.
Our captivity experimental group to develop an embryo bank consisted of ten adult females and four fertile males of marsh deer from the Brazilian captive populations, both of the populations of Três Irmãos Hydroelectric Power Plant and the Porto Primavera Hydroelectric Power Plant (UHSM). This group was subdivided into three different treatments (n = 3, Table 1), and one free-living female captured for the embryo transfer. The animals were maintained in two installations. The first one was the Marsh Deer Conservation Center (Centro de Conservação do Cervo-do-Pantanal, CCCP), Promissão, São Paulo, Brazil. Animals were maintained individually in paddocks (40 m x 100 m) and were fed with a diet consisting of a pelleted ration (Essence Traditional – Presence®), approximately 500 g/deer/day of fresh abobra (Abobra tenuifolia), 250 g/deer/day of fresh roselle (Hibiscus sabdariffa) and water ad libitum. The second one was the Deer Research and Conservation Center (Núcleo de Pesquisa e Conservação de Cervídeos, NUPECCE), São Paulo State University (UNESP), Jaboticabal, São Paulo, Brazil. Animals were maintained individually in stalls (4 m x 4 m) and were placed in paddocks (20 m x 60 m) when estrus detection was performed. Animals were fed with a diet consisting of a pelleted ration (Equi Tech 12MA – Presence®), approximately 2 kg/deer/day of fresh perennial soybean (Neonotonia wightii), ramie (Boehmeria nívea), or mulberry branches (Morus alba), and water ad libitum. All animals maintained auditory and olfactory contact with conspecific males and females and were exposed to normal fluctuations in the photoperiod. All animal procedures were approved by the Ethics Committee on Animal Use (CEUA) of the School of Agricultural and Veterinarian Sciences from São Paulo State University (approval N° 13721/15), in accordance with the ethical principles adopted by the Brazilian College of Animal Experimentation (COBEA).
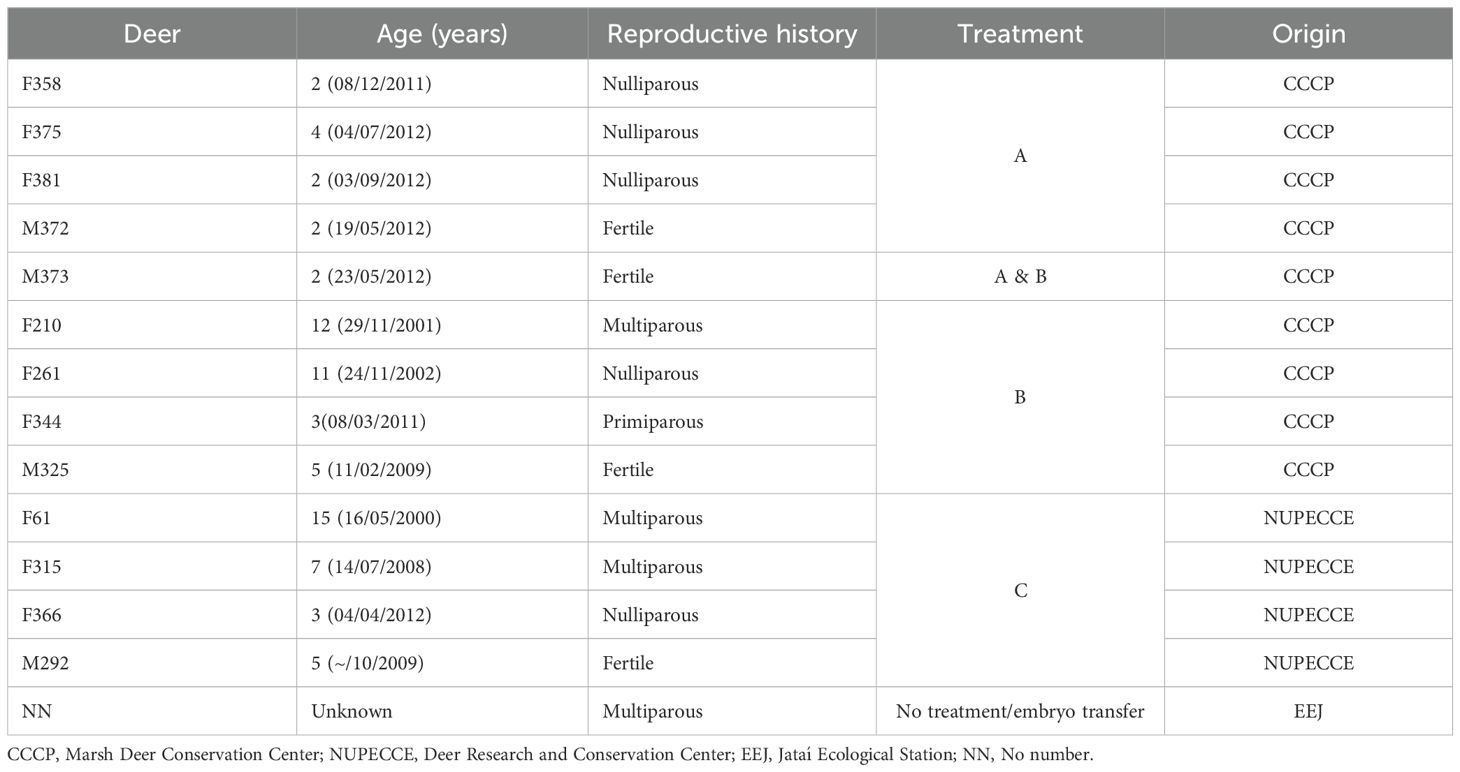
Table 1. Characteristics of 13 deer of the species Blastocerus dichotomus that comprised the experimental groups.
We conducted three equine chorionic gonadotropin (eCG-based) superovulation (SOV) protocols for B. dichotomus (Figure 1). An adaptative system was adopted, and adjustments were made after each treatment until a superovulatory response was obtained. The treatments did not take into consideration the seasons of the year, because marsh deer hinds are non-seasonal breeders (Polegato et al., 2018).
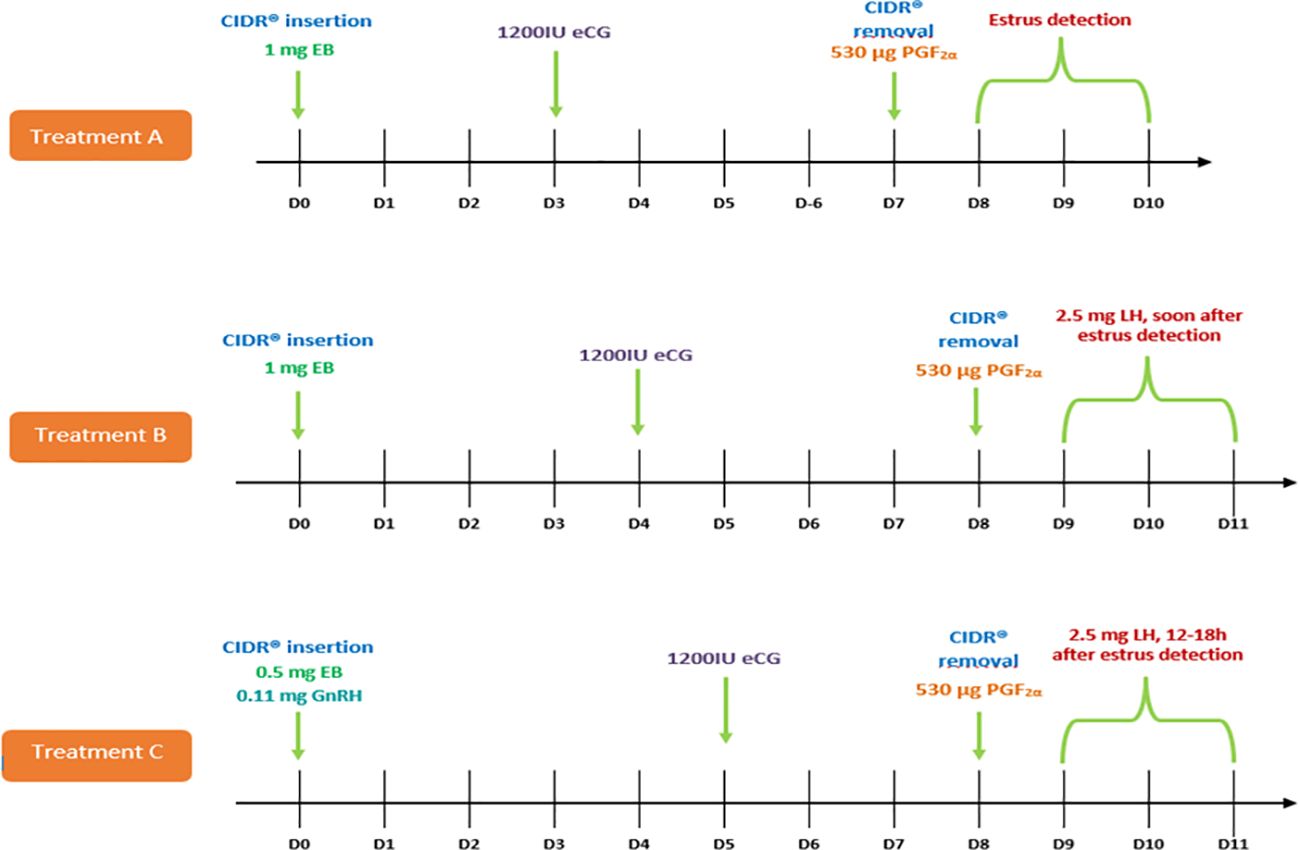
Figure 1. Treatments for superovulation and ovulation induction in Blastocerus dichotomus. Treatment A: CIDR® (7 days) + 1 mg EB (D0) + 1,200IU eCG (D3) + 530 µl PGF2α (D7); Treatment B: CIDR® (8 days) + 1 mg EB (D0) + 1,200IU eCG (D4) + 530 µl PGF2α (D8) + 2.5 mg LH soon after estrus detection; Treatment C: CIDR® (8 days) + 0.5 mg EB + 0.11 mg GnRH (D0) + 1,200IU eCG (D5) + 530 µl PGF2α (D8) + 2.5 mg LH between 12-18 h after estrus detection.
Treatment A. In June 2014, three adult females (n = 3) (weighing 70.0 to 80.0 kg) and two males (weighing 90.0 to 100.0 kg) (Table 1) were housed in the CCCP. On a random day of the estrous cycle (D0), females received a single intravaginal device containing 0.33 g of progesterone (CIDR®-type T; Pfizer®; USA) for seven days. Still on D0, they received 1 mg of estradiol benzoate (EB) (Sincrodiol®; Ourofino Saúde Animal Ltda., Brazil) by intramuscular (i.m.) injection. They also received 1, 200 IU eCG (Folligon®; Intervet® Schering-Plough Animal Health, Brazil) by i.m. injection on day 3 following CIDR® insertion, and 530 µg of cloprostenol (PGF2α; Ciosin®; Intervet® Schering-Plough Animal Health, Brazil) by i.m. injection the day CIDR® was removed (D7).
Treatment B. In July 2014, three adult females (n = 3) (weighing 80.0 to 95.0 kg) and two males (weighing 90.0 to 110.0 kg) (Table 1) were housed in the CCCP. On a random day of the estrous cycle (D0), females received a CIDR® for eight days. Still on D0, they received 1 mg of EB by i.m. injection. They also received 1,200 IU of eCG by i.m. injection on day 4 following CIDR® insertion and 530 µg of PGF2α by i.m. injection the day the device was removed (D8). Soon after estrus detection, they received 2.5 mg of luteinizing hormone swine pituitary (LH) (LUTROPIN®-V - Bioniche® - Canada) by i.m. injection.
Treatment C. Between May and June 2015, three adult females (n = 3) (weighing 70.0 to 95.0 kg) and one male (weighing 110.0 kg) (Table 1) were housed in NUPECCE. On a random day of the estrous cycle (D0), females received a CIDR® for eight days, 0.5 mg of EB and 0.11 mg of gonadorelin diacetate tetrahydrate (GnRH) (Cystorelin®; Merial®, USA) by i.m. injection. They also received 1,200 IU of eCG by i.m. injection on day 5 following CIDR® insertion and 530 µg of PGF2α by i.m. injection the day the device was removed (D8). Between 12 and 18 h after estrus detection, they also received 2.5 mg of LH by i.m. injection.
CIDR® insertion and removal, as well as EB and PGF2α applications were performed during chemical immobilization with an association between 1 mg/kg of azaperone (Stresnil®, Janssen Pharmaceutica, Bélgica) and 0.5 mg/kg of xilazine hydrochloride (Sedomin®, König Brazil Ltda., Brazil) (Carregaro et al., 2019) i.m. dart injection, that was applied at a distance by a blowgun. The eCG application was also performed by i.m. dart injection. In treatment C, the eCG and PGF2α applications, as well as CIDR® removal were carried out with the aid of a box with a small window to restrict and manipulate the animals (Duarte et al., 2010).
Behavioral estrus was defined as the period in which females permitted mating (Pereira et al., 2006) and was determined by grouping a female with a male in the paddock (10 – 20 minutes every 12 h) from one day after CIDR® removal, until the moment that the female no longer accepted copulation (end of estrus).
SOV response was evaluated by counting corpus luteum (CL) on both ovaries. Total follicular stimulus was evaluated by counting CL and persistent follicles (large anovulatory follicles) (Blanco et al., 2003). Counting was performed 8 days after the first mating by ventral midline laparotomy. Thus, females with two or more CL were classified as superovulated (Zanetti et al., 2014). For surgical procedure, females were submitted to food and water fasting for 24 hours. Chemical restraint was done with a pharmacological combination of 7.0 mg/kg of ketamine hydrochloride and 1 mg/kg of xylazine hydrochloride at a distance by a dart gun (JM-standard, Dan-Inject) (Favoretto et al., 2012). Then, intubation was performed with an endotracheal catheter n° 9 and maintained under inhalation anesthesia with isoflurane (Generic Isoflurane - Instituto Biochimico Ind. Farm, Ltda. - Brazil). Cannulation of the cephalic vein was performed for the administration of fluid therapy at a rate of 10 ml/kg/h with 0.9% NaCl solution. Embryo collection was performed by oviduct and uterine flushing (Duarte and Garcia, 1995). The flushing medium used for embryo recovery was phosphate buffered saline (PBS) solution (Nutricell Nutrientes Celulares® - Campinas - Brazil) associated with 1% fetal bovine serum (FBS) (Nutricell Nutrientes Celulares® - Campinas - Brazil), heated to 37°C. At the end of the procedure, the surgical wound was sutured with nylon thread 2.0. In the immediate postoperative period, hinds received 40,000 IU/Kg of benzathine penicillin i.m. and 500 mg of phenylbutazone, i.v. They also received an i.m. application of 530 μg of PGF2α to induce luteolysis of the multiple CL and, eventually, prevent the implantation of embryos that remained after collection.
For embryo evaluation, flushing medium collected was analyzed using a stereo microscope at 10x. The collected embryos were washed in PBS plus 10% FBS for removal of cell debris and classified (with 50x magnification) according to the developmental stage (number of cells and morphology) and structural integrity, following the standards used for bovine embryos (Stringfellow and Seidel, 1999). All embryos were vitrified using commercial medium (Bioclone, Jaboticabal, São Paulo), stored in labeled vitrification straws, and subsequently preserved in liquid nitrogen.
In order to perform the embryo transfer into the reintroduced population at the Jataí Ecological Station, Luis Antônio, São Paulo, one adult female (weight = 84 kg) was captured and brought into captivity during July 2015. The animal was captured using the ‘Bulldogging’ method (Duarte, 2008), with the assistance of a Robinson 44 helicopter. After physical restraint, chemical sedation was administered via i.v. injection with a pharmacological combination of 0.05 mg/kg acepromazine, 5 mg/kg ketamine, and 0.5 mg/kg midazolam. A transrectal ultrasound examination (linear transducer, 7.5 MHz) was then conducted to confirm pregnancy and estimate gestational age (Zanetti et al., 2010a) (Figure 2). Following the decision to remove the animal from its original location and transport it to NUPECCE for embryo transfer procedures, the animal was airlifted by helicopter to the outer edge of the floodplain, where a vehicle was stationed to transport it to NUPECCE. The female was kept at the NUPECCE facilities with minimal handling to avoid its adapting to captivity. As a result, human contact was highly restricted.

Figure 2. The animal was physically restrained after capture, and a transrectal ultrasound was performed, showing the fetus in an advanced stage of development.
One month after parturition, late October 2015, and considering the postpartum estrus in the species (Polegato et al., 2018), the wild female marsh deer was synchronized for embryo transfer. On a random day (D-8) of the estrous cycle the females received a CIDR® for eight days and an i.m. injection of 0.5 mg of EB. On the day of CIDR removal (D0), i.m injections of 530 µg of PGF2α and 450 IU of eCG were administered (Zanetti et al., 2009; modified).
Nine days after CIDR removal (D9), chemical restraint was administered, and the female underwent a laparoscopic procedure to assess the presence of the CL (Zanetti et al., 2010b; Zanetti and Duarte, 2012) (Figure 3A), followed by embryo transfer (Figure 3B). At the end of the procedure, the incisions were sutured, and the female received an i.m. injection of 40,000 IU/kg of benzathine penicillin. Before recovery from anesthesia, the animal was fitted with a Wild CellMG® radio transmitter collar (Lotek Wireless®, Canada - pre-programmed to send the animal’s geographic coordinates via GSM). The collar weighed 950 g, which corresponds to 1.13% of the animal’s body weight.
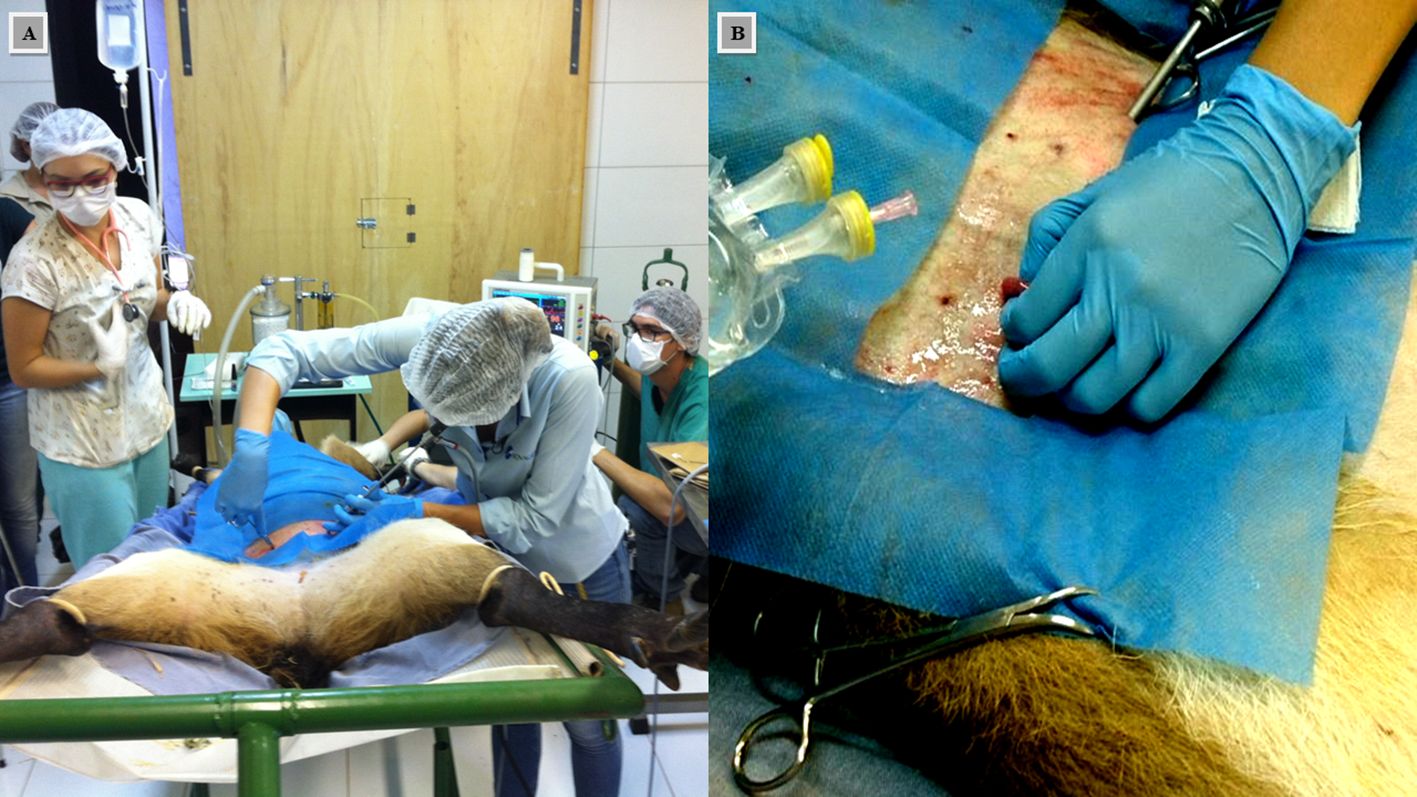
Figure 3. Embryo transfer surgery. (A) Exploratory laparoscopy to confirm the presence of a corpus luteum. (B) Exposure of the uterus for embryo inoculation.
Ten days after embryo transfer, in November 2015, the female was anesthetized and transported to its natural habitat, into the EEJ, in a transport box. Monitoring of the female after release was conducted using the GPS – GSM system of the radio transmitter collar. Two types of sampling were scheduled: (1) continuous location collection every hour to obtain information on the animal’s area use and survival, and (2) intensive sampling every 15 minutes for 3 days following release and for another 4 days throughout gestation (one day per week) to track the animal’s exact movements. We analyzed data from the first 2 months of monitoring to investigate the release process. During this period the GPS data resulted in 1514 localities collected from which 1427 (94%) were with high quality coordinates (DOP<10) and were used for area and movements estimations. Information was sent by the GSM-collar every 5 locations collected, thus resulting in a daily real-time monitoring, which was also used to recapture procedures. Nine months after the reintroduction of the female, a recapture was scheduled, in August 2016, with the expectation of finding female with a fawn at its side. The same capture protocol previously described was followed, with the help of the coordinates of its recent location transmitted via GSM.
Descriptive statistics, including the mean and standard deviation, were calculated for the number of CL, anovulatory follicles (AF), and total follicular stimulus (TFS) across treatments. Data normality was assessed using the Shapiro-Wilk test. For non-normal data (Treatment B), the Friedman rank test was applied, while data from Treatments A and C were analyzed using ANOVA. Post-hoc comparisons were conducted using Tukey’s HSD or equivalent pairwise tests, depending on data distribution. A significance level of p < 0.05 was set for all analyses.
Results for SOV response are presented in Table 2. For Treatment A, all females displayed behavioral signs of estrus, with subsequent mating with males after CIDR® removal, 61.33 ± 7.06 h (± SEM, ranged 48 – 72 h) after the end of treatment. Only females F375 and F381 ovulated, presenting 1 CL each, giving a mean rate of total follicle stimulus of 5.33 ± 1.33 and an ovulation rate of 12.5% (2/16). However, the mean rate of anovulatory follicles was 4.67 ± 1.2. Therefore, no SOV was obtained with this treatment.
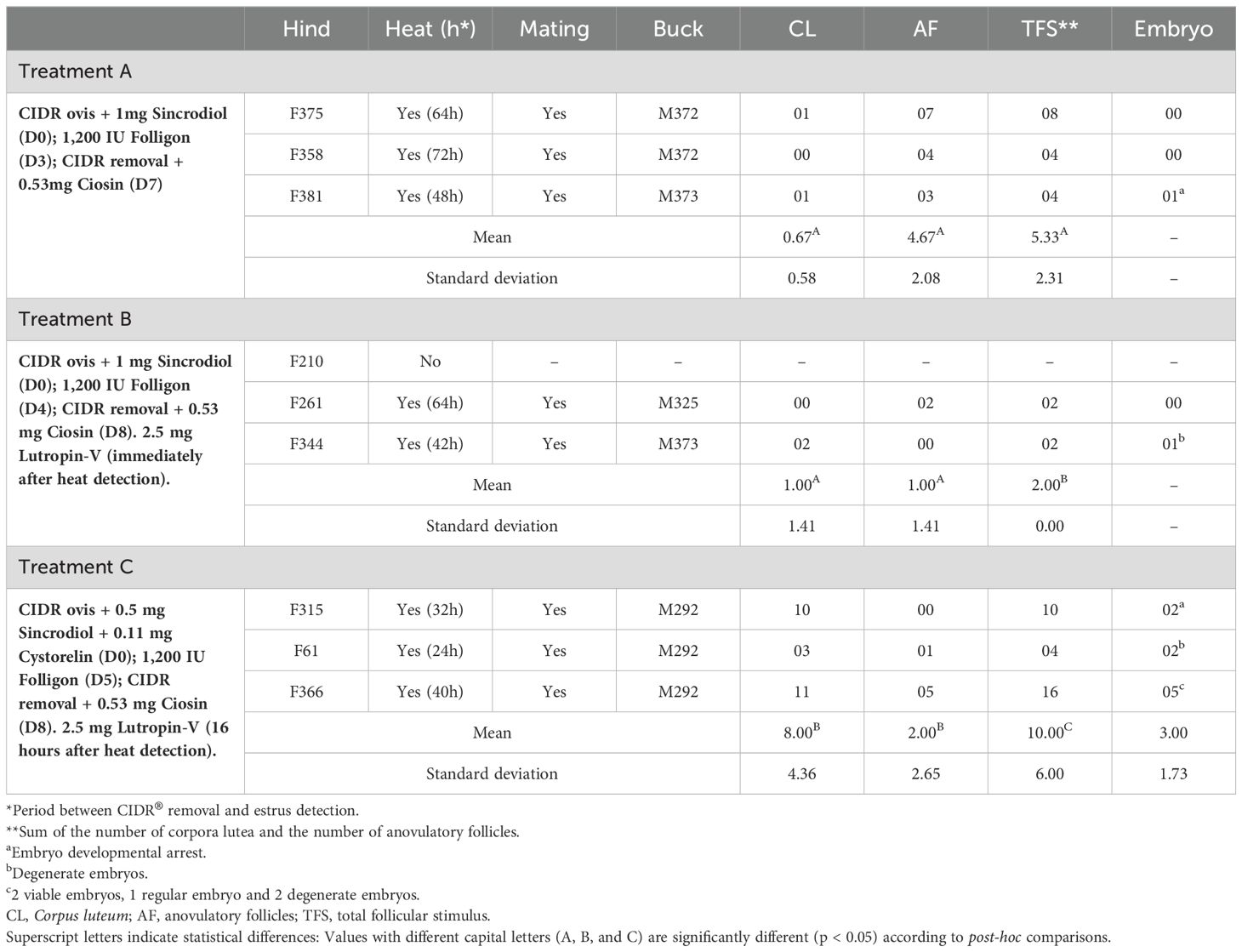
Table 2. Data of the estrus and ovarian response of nine hinds of the species Blastocerus dichotomus, submitted to three different superovulation treatments.
For Treatment B, estrous synchronization failed in female F210. Females F261 and F344 displayed behavioral signs of estrus, with subsequent mating with males after CIDR® removal, 53 ± 11 h (± SEM, ranged 42 – 64 h) after the end of treatment. Only female F344 presented 2 CLs, and the total follicle stimulus was 1.33 ± 0.67.
Treatment C was successful in generating SOV response and estrous synchronization in all females. Behavioral signs of estrus were displayed 32 ± 4.62 h (± SEM, ranged 24 – 40 h) after the end of treatment. The mean rate of anovulatory follicles was 2 ± 1.53 and the mean rate of CL was 8 ± 2.52 (± SEM, ranged 3 – 11). This treatment showed a total follicle stimulus of 10 ± 3.46 and an ovulation rate of 80% (24/30) (Figure 4).
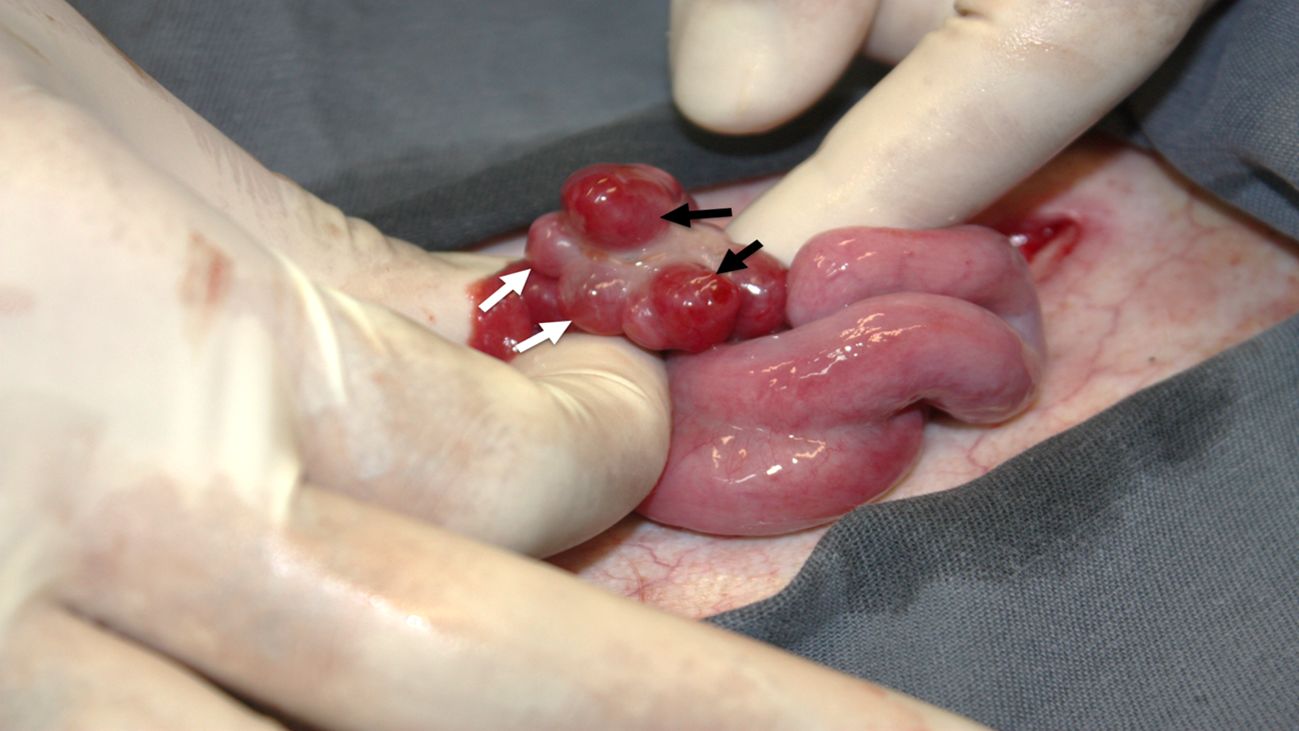
Figure 4. Horns, uterine body, and left ovary of the hind F366, of the species Blastocerus dichotomus after Treatment C: CIDR® (8 days) + 0.5 mg EB + 0.11 mg GnRH (D0) + 1,200UI eCG (D5) + 530 µl PGF2α (D8) + 2.5 mg LH, 12-18 h after detection of estrus. The presence of anovulatory follicles (white arrows) and corpora lutea (black arrows) are observed.
To evaluate the effectiveness of superovulation treatments, we analyzed the number of CL, anovulatory follicles (AF), and total follicular stimulus (TFS) across Treatments A, B, and C. The Shapiro-Wilk test indicated that Treatment B data were non-normal (p < 0.05), while Treatments A and C followed a normal distribution (p > 0.05). Consequently, we applied the Friedman rank test for non-normal data and ANOVA for normal data. The ANOVA revealed a statistically significant difference for Treatment C compared to the other treatments (p = 0.045). Post-hoc comparisons confirmed a significant difference between Treatments B and C (p = 0.0332), while no significant differences were observed between Treatments A and B or A and C (p > 0.05).
Embryo recovery was successful in all treatments. However, only one embryo was recovered in Treatments A and B, with the former exhibiting developmental arrest and the latter showing degeneration. As a result, the recovery rate for both treatments was 50% (1/2). In contrast, nine embryos were recovered in Treatment C, which also achieved an ovulation rate of 80% (24/30). Overall, the embryo recovery rate for B. dichotomus females was 37.5% (9/24). Of the nine embryos collected, only two were viable (Figure 5). Of the remaining embryos, one was of fair quality, four were degenerated, and two exhibited developmental arrest, following the standards used for bovine embryos (Stringfellow and Seidel, 1999).
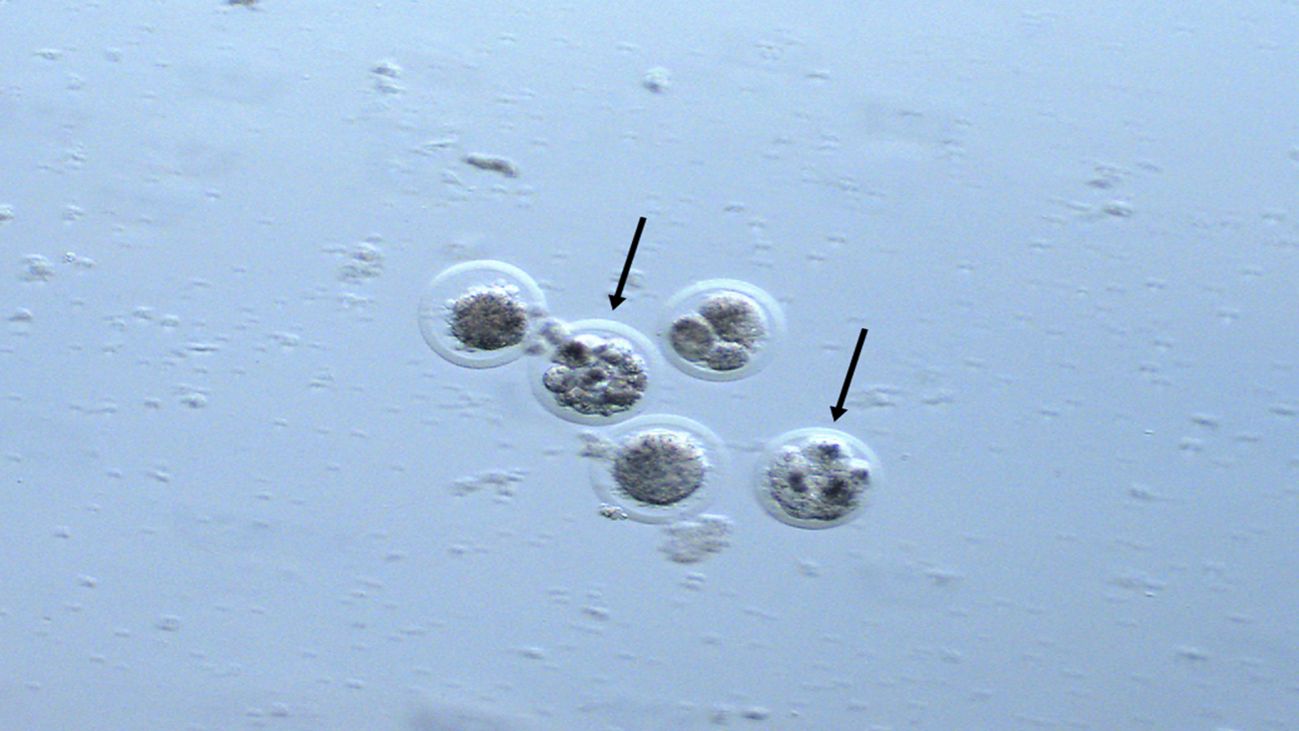
Figure 5. Structures recovered from hind F366, of the species Blastocerus dichotomus after Treatment C: CIDR® (8 days) + 0.5 mg EB + 0.11 mg GnRH (D0) + 1,200UI eCG (D5) + 530 µl PGF2α (D8) + 2.5 mg LH, 12-18 h after detection of estrus. The black arrows indicate two viable structures for embryo transfer.
Before embryo transfer, fawning occurred in October 2015 and the fawn was separated from its mother and transitioned to artificial feeding, subsequently becoming part of the ex situ Marsh Deer Conservation Program as a founding female. Then, nine days after the end of synchronization protocol, and upon confirming the presence of the CL through laparoscopic procedure, the uterine horn was exteriorized via median ventral laparotomy (Figure 3B), and two high-quality vitrified embryos were re-warmed and surgically transferred using a “tom cat” catheter.
Once the female was released, monitoring began using a GPS collar with data transmission via GSM on a daily basis (Figure 6). The data collection and transmission schedule functioned perfectly, enabling real-time monitoring of the animal. As expected for open monitoring, the majority (94%) of locations obtained were of high quality. In addition to the planned 9-month monitoring period, tracking continued for an additional 6-month period without any battery issues. During the first three days post-release, the female exhibited notably greater movement, averaging 124 meters per hour (approximately 3 km per day). In contrast, movement during the remaining fine-scale sampling (n=4) averaged 50 m/h (approximately 1.2 km/day), with increased movement observed in early January. Throughout the entire monitoring period, the female occupied an area of 2.4 km² (Figure 6).
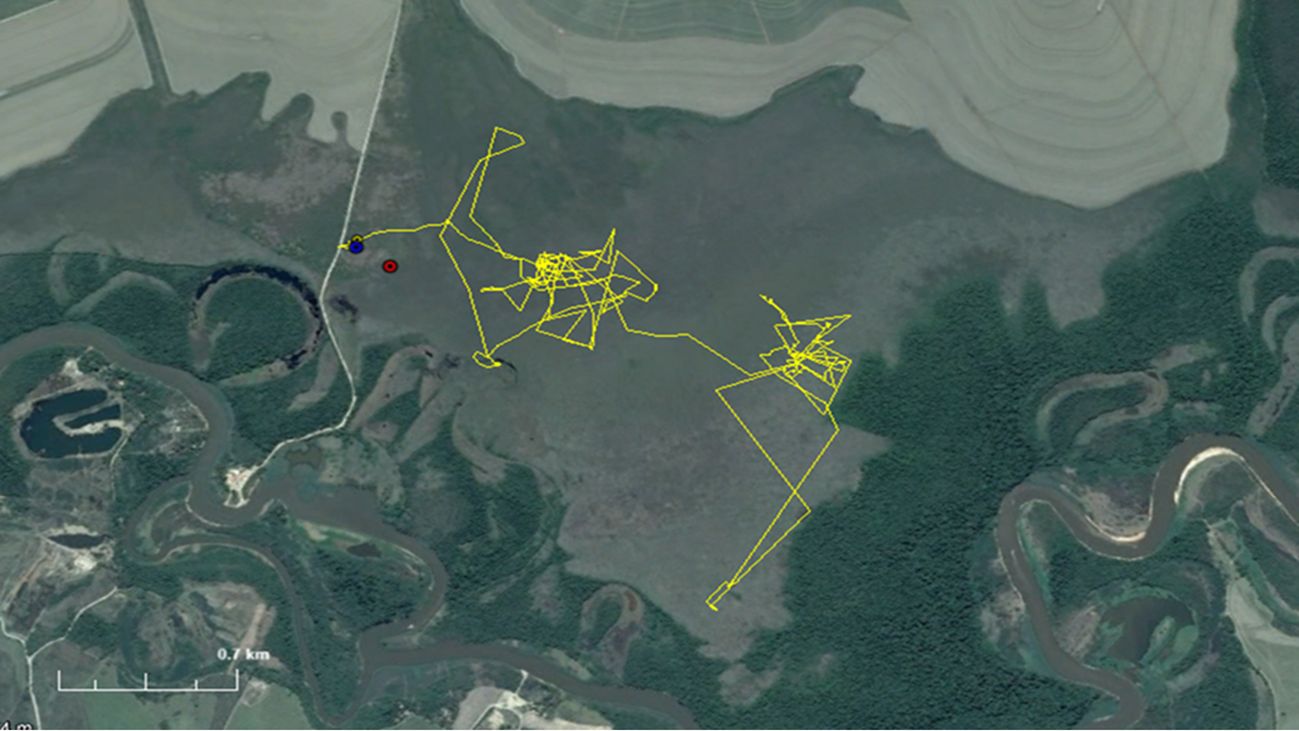
Figure 6. Paths taken by the monitored female marsh deer during the first 25 days of return to the wild (yellow), location of the female’s capture in July (red), and release in November (blue).
In August 2016, nine months after female reintroduction into EEJ, a second capture for parturition confirmation was performed. After physical restraint, the female deer underwent an ultrasound to confirm parturition, as female deer typically leave their fawns in secluded areas, hidden in the grass, while they feed to avoid predation. The ultrasonographic evaluation revealed a pregnancy of approximately six months, indicating the loss of the transferred embryos and suggesting natural fertilization three months after reintroduction. After the procedure the female was released at the same point of capture.
The establishment of the Marsh Deer Conservation Program represents a significant milestone in Brazil and South America’s conservation efforts, being one of the first fully local conservation programs. Over the course of two decades, the program has navigated challenges, setbacks, and successes, underscoring the complexities of implementing such a comprehensive initiative with the collaboration of all stakeholders. Despite notable advancements, the marsh deer remains at risk across its range, highlighting the critical importance of integrating reproductive management strategies and biotechnologies to support both at-risk wild and captive populations. While considerable progress has been made in animal reproductive sciences, continued research is essential to furthering our understanding of the reproductive physiology of Neotropical deer species. Based on previous studies of superovulation (SOV) in species such as the brown brocket deer (Subulo gouazoubira) (Duarte and Garcia, 1995; Zanetti and Duarte, 2012; Zanetti et al., 2014) and the red brocket deer (Mazama americana) (Cursino et al., 2014), this study aims to evaluate the application of SOV and embryo transfer techniques as potential tools for advancing the conservation of the marsh deer.
Due to the limited number of captive marsh deer, we could not use a standard experimental design as used with non-endangered species. Instead, we developed an adaptive system with sequential experiments, making small adjustments between trials based on earlier outcomes. In Treatment A, the inadequate superovulatory response may have resulted from several factors, including handling-induced stress, which affects oocyte maturation, suppress the preovulatory LH surge, and may influence adrenal progesterone production (Monfort et al., 1990; Brann and Mahesh, 1991; González et al., 2008; Zanetti et al., 2014). The absence of an ovulation inducer may have contributed to the lack of ovulation, as the proper timing and type of inducer are critical for follicle maturation and ovulation (Xu et al., 1995; D’Occhio et al., 1997; Liu and Sirois, 1998; D’Occhio et al., 1999; Kaim et al., 2003). Insufficient LH receptor development in ovarian follicles can lead to follicular luteinization without ovulation (Brann and Mahesh, 1991). Common inducers include LH, GnRH and human chorionic gonadotrophin (hCG). Zanetti et al. (2014) observed multiple anovulatory follicles following SOV protocols based on eCG and FSH in brown brocket deer (4.5 ± 1.7 and 7.6 ± 0.9, respectively). Elevated fecal glucocorticoid levels during the SOV period further supported this hypothesis. To address these issues in Treatment B, we added an additional day of exogenous progesterone, adjusted the timing of eCG administration, and included LH as an ovulation inducer after estrus detection.
Modifications made in Treatment B did not effectively induce a superovulatory response in marsh deer females, and female F210 showed no response at all. We suggest that F210’s lack of response may be related to stress from the management procedures during treatment, which could have disrupted her reproductive physiology (Monfort et al., 1990). However, glucocorticoid hormone levels were not measured to confirm this hypothesis. For the other females, we hypothesize that a delayed eCG application may have interfered with the timing of follicular development. By the time eCG was administered, a dominant follicle had already begun to suppress the development of subordinate follicles, which are still FSH-dependent at the recruitment stage (Castilho and Garcia, 2005). Although female F261 developed two ovulatory follicles, there was a lack of synchronicity between follicular maturation and the ovulation inducer. In contrast, female F344 exhibited two CL, which might be considered a superovulatory response, but the overall follicular response was suboptimal due to the limited number of follicles (Table 2).
After the unsatisfactory response to Treatment B, further adjustments were made to the SOV protocol in Treatment C, including lowering the EB dose to 0.5 mg, adding GnRH (0.11 mg), administering eCG on day 5, and delaying LH application until 12-18 hours after estrus detection. These modifications successfully induced an adequate superovulatory response in marsh deer females. Previous studies support the effectiveness of a lower EB dose during progesterone implant insertion (Kumar et al., 2003; Zanetti and Duarte, 2012). Additionally, GnRH improved dominant follicle ovulation, and combining EB with CIDR® promoted follicular atresia in subordinate follicles, aiding estrus synchronization (Bo et al., 1995; Diskin et al., 2002; McCorkell et al., 2008; Zanetti et al., 2014). The delayed eCG application was aimed at optimizing follicular growth, with studies indicating follicular wave emergence occurs 4-5 days after estradiol-17β and progesterone treatment (Burke et al., 2000; McCorkell et al., 2008), and selection occurs 3-4 days after GnRH (Twagiramungu et al., 1995). The delayed LH application allowed for proper follicular maturation, making them more responsive to the LH peak stimulus (D’Occhio et al., 1997, 1999). Despite being 15 years old, female F61 responded to Treatment C, showing 3 CL and 1 anovulatory follicle. Similar to Zebu cattle, older ruminants typically have a reduced ovarian follicle reserve (Peixoto et al., 2006), with a decrease in CL and recovered structures. This suggests that the treatment may be more effective in younger females, as seen with F315 (6 years) and F366 (3 years) (Table 2). The superior response in Treatment C was characterized by higher CL counts and an improved total follicular stimulus (mean CL = 8, TFS = 10) compared to Treatments A and B (mean CL = 0.67 and 1, TFS = 5.33 and 2, respectively). These results suggest that the protocol used in Treatment C significantly enhances follicular development and superovulation outcomes compared to the less effective protocols employed in Treatments A and B.
Embryo recovery was successfully performed in all treatments, with Treatment C yielding the highest recovery rate. The embryo recovery rate in Treatment C was 37.5% (9/24), which is consistent with rates reported in other deer species, such as Elaphurus davidianus (29.3%) (Argo et al., 1994); Cervus elaphus (43.1%) (Argo et al., 1994), (32.7%) (Asher et al., 1995); and Dama dama (30.6%) (Asher et al., 1995). Of the nine embryos recovered in Treatment C, four were degenerated, and two showed developmental arrest. Only two of the remaining embryos were of sufficient quality for transfer and were subsequently used in this study (Figure 5). Both viable embryos came from female F366, who exhibited the best response in terms of follicular development within Treatment C. This suggests that females of optimal reproductive age may respond more favorably to the SOV protocol used in Treatment C, further supporting the importance of age in reproductive success. The results presented here represent an opportunity to adjust the protocol for the implementation of a genetic resource bank for the species. This will be of vital importance for supporting both the captive population and the free-ranging population in the Marsh Deer Conservation Program.
The decision to transfer the only two viable embryos into the same female was a difficult one, made with the intention of maximizing the chances that at least one of them would implant. Although this species is considered uniparous and twin or multiple births have not been observed (Piovezan et al., 2010; Polegato et al., 2018), unfortunately, nine months later, during recapture, it was discovered that the female had lost both embryos. Suggesting that approximately three months after her reintroduction into the wild, it displayed estrus and had the opportunity to mate, eventually showing signs of pregnancy with approximately six months of gestation at the time of the ultrasound evaluation. While this was disappointing in terms of potential genetic diversity enhancement for the EEJ population, it nonetheless provided hope regarding the entire procedure conducted since the female’s capture from the wild. It indicates that the surgical procedures for embryo transfer did not negatively affect its physiological reproductive capacity, which is crucial for the viability of this procedure in other wild females.
This attempt demonstrated the feasibility of holding a free-ranging animal in captivity for four months and successfully reintroducing it to the wild, offering valuable insights into the potential of ex situ conservation techniques. It is important to note, however, that the female was reintroduced to an area very close (100 meters) to her original capture site, minimizing environmental variations and ensuring she was familiar with the surroundings. The successful reintroduction of this female marsh deer supports findings on the positive effects of longer acclimation periods on site fidelity (Mertes et al., 2019). Consistent with studies on elk (Ryckman et al., 2010; Bleisch et al., 2017), the marsh deer remained near the release site without signs of stress or displacement, suggesting that the extended captivity period facilitated proper acclimation. In contrast, shorter captive holding periods (4–11 days), as seen in elk, led to greater dispersal from the release site (22.6–26 km) (Ryckman et al., 2010). This case highlights the importance of controlled acclimation in familiar environments to improve reintroduction success and reduce risks such as post-release dispersal (Parker et al., 2008; Tuberville et al., 2008). Besides, it is important to consider that deer are prey species in the wild and are highly susceptible to stress stimuli. For this reason, their release must be carefully planned to ensure that the animal can adequately adapt and return to its natural habitat. Proper planning of reintroduction strategies, including minimizing handling and environmental stressors, is essential for the success of wildlife restoration efforts (Fischer and Lindenmayer, 2000; Seddon et al., 2007). Moreover, stress during reintroduction can negatively impact animal health and survival rates, further highlighting the need for a thorough, well-managed release process (Teixeira et al., 2007).
The use of GPS-GSM technology to monitor the reintroduced female marsh deer provided invaluable insights into her post-release behavior, survival, and adaptation to the wild. Successful reintroduction programs, as highlighted by previous studies, depend on tracking released animals to assess their integration into the environment (Parker et al., 2008; Mertes et al., 2019). In this study, the GPS collar with GSM data transmission was crucial for understanding the female’s movements and habitat use post-release. It enabled two types of data sampling: continuous hourly location tracking for broader area uses and survival, and intensive 15-minute intervals to monitor fine-scale movements during the immediate post-release period and some days of gestation. This dual approach provided comprehensive data on both daily behavior and long-term adaptation, aligning with findings from other reintroduction studies where detailed movement data is essential for evaluating habitat selection (Ryckman et al., 2010; Bleisch et al., 2017). Initially, the female showed higher mobility, moving an average of 124 meters per hour (3 km/day) during the first three days, typical of post-release behavior as animals re-establish home ranges (Mertes et al., 2019). Afterward, the female’s movement slowed to 50 meters per hour (1.2 km/day), indicating stabilization and adaptation. Therefore, GPS tracking proves a valuable tool for wildlife managers, supporting informed decisions on habitat suitability, release strategies, and the effectiveness of ex situ conservation efforts.
This study demonstrates the potential of embryo transfer as a viable strategy for enhancing genetic diversity in natural marsh deer populations, particularly those threatened by low genetic variability (Rola et al., 2021, 2023). Traditional reintroduction programs often face challenges related to inbreeding depression, which can lead to reduced fitness, lower reproductive success, and heightened disease susceptibility (Seddon et al., 2007). By introducing genetically diverse embryos sourced from ex situ conservation programs, this technique offers a controlled and scientifically grounded approach to mitigating these issues. The findings highlight key advancements, including improvements in hormonal protocols, optimized embryo collection techniques from the captive population, and the refinement of transfer methods. The success achieved in obtaining viable embryos and performing transfers indicates that these methods can significantly contribute to conservation programs to sustain healthy and diverse populations. Continued research and technological refinement are necessary to enhance the effectiveness of this approach and adapt it to the specific reproductive physiology of marsh deer and other threatened Neotropical deer species. By integrating embryo transfer into broader conservation strategies, this method holds promise as a valuable tool for addressing genetic bottlenecks and improving the long-term viability of wildlife populations facing environmental and genetic challenges.
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding authors.
The animal study was approved by Ethics Committee on Animal Use (CEUA). The study was conducted in accordance with the local legislation and institutional requirements. Written informed consent was obtained from the individual(s) for the publication of any identifiable images or data included in this article.
DG: Conceptualization, Investigation, Visualization, Writing – original draft. PP: Conceptualization, Investigation, Visualization, Writing – review & editing. EZ: Conceptualization, Funding acquisition, Investigation, Methodology, Writing – review & editing. MO: Conceptualization, Funding acquisition, Investigation, Methodology, Visualization, Writing – review & editing. LR: Conceptualization, Investigation, Visualization, Writing – review & editing. JD: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Writing – original draft.
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001, FUNEP/TFCA/FUNBIO (043/2013), and CNPq (302495/2022-3).
The authors are grateful to Joaquim Mansano Garcia, PhD, and Marina Susuki Cursino, PhD, for their assistance with the design of protocols and FUNBIO’s project, as this study is part of it. To Maria E. Franco Oliveira, PhD, Gabriela Martins Siqueira, PhD., and Lincy Camacho Orjuela, MSc., for their assistance with the surgical procedures. To Marina Ragagnin de Lima, PhD, for her valuable support with the vitrification and thawing of embryos. To Pedro Nacib Jorge-Neto, PhD, and DVM Letícia Alecho Requena for their support with the embryo transfer technique. To Adriano Bonfim Carregaro PhD, Rozana Wenlder da Rocha, PhD, André Nicolai Elias, PhD, and Bárbara Ferrari, MSc. for their support with the anesthetic procedures. The authors are also grateful to each one of the institutions that allowed the use of animals belonging to their establishments, such as the Marsh Deer Conservation Center, the Sorocaba Zoo, Fábio Barreto Forest and Zoo, and the Piracicaba Municipal Zoo.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Argo C. M. G., Jabbour H. N., Goddard P. J., Webb R., Loudon A. S. I. (1994). Superovulation in red deer (Cervus elaphus) and Pere David’s deer (Elaphurus davidianus), and fertilization rates following artificial insemination with Pere David’s deer semen. J. Reprod. Fertility 100, 629–636. doi: 10.1530/jrf.0.1000629
Asher G. W., Jabbour H. N., Thompson J. G., Tervit H. R., Morrow C. J. (1995). Superovulation of farmed red deer (Cervus elaphus) and fallow deer (Dama dama): Incidence of ovulation and changes in plasma hormone concentrations during the pre-ovulatory period in relation to ova recovery and fertilisation. Anim. Reprod. Sci. 38, 137–154. doi: 10.1016/0378-4320(94)01351-L
Blanco M. R., Simonetti L., Rivera O. E. (2003). Embryo production and progesterone profiles in ewes superovulated with different hormonal treatments. Small Ruminant Res. 47, 183–191. doi: 10.1016/S0921-4488(02)00245-6
Bleisch A. D., Keller B. J., Bonnot T. W., Hansen L. P., Millspaugh J. J. (2017). Initial movements of re- introduced elk in the missouri ozarks. Am. Midland Nat. 178, 1–16. doi: 10.1674/0003-0031-178.1.1
Bo G. A., Adams G. P., Pierson R. A., Mapletoft R. J. (1995). Exogenous control of follicular wave emergence in cattle. Theriogenology 43, 31–40. doi: 10.1016/0093-691X(94)00010-R
Brann D. W., Mahesh V. B. (1991). Role of corticosteroids in female reproduction. FASEB journal: Off. Publ. Fed. Am. Societies Exp. Biol. 5, 2691–2698. doi: 10.1096/fasebj.5.12.1655548
Burke C. R., Day M. L., Bunt C. R., Macmillan K. L. (2000). Use of a small dose of estradiol benzoate during diestrus to synchronize development of the ovulatory follicle in cattle. J. Anim. Sci. 78, 145–151. doi: 10.2527/2000.781145x
Carregaro A. B., Ferrari B. G., Silva A. N.E., Xavier N. V., Duarte J. M. B. (2019). Azaperone and xylazine: A pharmacological combination to facilitate captive deer management for red brocket deer (Mazama americana). PLoS One 14, e0220288. doi: 10.1371/journal.pone.0220288
Castilho C., Garcia J. M. (2005). Divergência no crescimento folicular: efeito na competência oocitária para produção in vitro de embriões - revisão. Arch. Veterinary Sci. 10, 17–23. doi: 10.5380/avs.v10i3.5124
Comizzoli P. (2020). Integrating fertility preservation and cryo-banking into the conservation of rare and endangered deer species. Anim. Production Sci. 60, 1227. doi: 10.1071/AN18674
Cursino M. S., Salviano M. B., Abril V. V., Zanetti E. S., Duarte J. M. B. (2014). The role of chromosome variation in the speciation of the red brocket deer complex: the study of reproductive isolation in females. BMC evolutionary Biol. 14, 40. doi: 10.1186/1471-2148-14-40
D’Occhio M. J., Sudha G., Jillella D., Whyte T, Maclellan L. J., Walsh : , et al. (1997). Use of a GnRH agonist to prevent the endogenous LH surge and injection of exogenous LH to induce ovulation in heifers superstimulated with FSH: a new model for superovulation. Theriogenology 47, 601–613. doi: 10.1016/S0093-691X(97)00018-6
D’Occhio M. J., Jillella D., Lindsey B. R. (1999). Factors that influence follicle recruitment, growth and ovulation during ovarian superstimulation in heifers: opportunities to increase ovulation rate and embryo recovery by delaying the exposure of follicles to LH. Theriogenology 51, 9–35. doi: 10.1016/S0093-691X(98)00228-3
Diskin M. G., Austin E. J., Roche J. F. (2002). Exogenous hormonal manipulation of ovarian activity in cattle. Domest. Anim. Endocrinol. 23, 211–228. doi: 10.1016/S0739-7240(02)00158-3
Duarte J. M. B. (2008). A technique for the capture of free-ranging marsh deer (Blastocerus dichotomus). J. Zoo Wildlife Med. 39, 596–599. doi: 10.1638/2005-056.1
Duarte J. M. B., Braga F. G., Vogliotti A., Abril V. V., Piovezan U, Reis M. L., et al. (2012). Plano de Ação Nacional para a Conservação dos Cervídeos Ameaçados de Extinção, Espécies de Cervídeos Brasileiros Não Ameaçadas de Extinção. (Brasília: Instituto Chico Mendes de Conservação da Biodiversidade (ICMBio)). Available at: http://www.icmbio.gov.br/portal/faunabrasileira/plano-de-acao-nacional-lista/860-plano-de-acao-nacional-para-conservacao-dos-cervideos (Accessed May 22, 2024).
Duarte J. M. B., Varela D., Piovezan U., Beccaceci M. D., Garcia J. E. (2016). Blastocerus dichotomus. The IUCN Red List of Threatened Species. (Cambridge, UK: IUCN), e.T2828A22160916. doi: 10.2305/IUCN.UK.2016-1.RLTS.T2828A22160916.en
Duarte J. M. B., Garcia J. M. (1995). Assistant reproduction in Brazilian cervidae. Rev. Bras. Reprodução Anim. 19, 111–121.
Duarte J. M. B., Uhart M. M., Galvez C. E. S. (2010). “Capture and physical restraint,” in Neotropical Cervidology. Eds. Duarte J. M. B., González S. (FUNEP, São Paulo), 218–227.
Falconer D. S. (1989). Introduction to quantitative genetics. 3rd edn (Harlow: Longman: Scientific & Technical).
Favoretto S. M., Zanetti E. S., Duarte J. M. B. (2012). Cryopreservation of red brocket deer semen (Mazama americana): comparison between three extenders. J. Zoo wildlife medicine: Off. Publ. Am. Assoc. Zoo Veterinarians 43, 820–827. doi: 10.1638/2011-0195R1.1
Ferreira P. R. S. (2011). Genética da reintrodução: O caso do cervo-do-pantanal (Blastocerus dichotomus) na Estação Ecológica de Jataı́. (master's thesis). São Paulo State University (UNESP), São Paulo, Brazil. Available online at http://hdl.handle.net/11449/92542.
Figueira C. J. M., Pires J. S. R., Andriolo A., Costa M. J. R. P., Duarte J. M. B. (2005). Marsh deer (Blastocerus Dichotomus) reintroduction in the Jataí Ecological Station (Luís Antônio, SP): spatial preferences. Braz. J. Biol. 65, 263–270. doi: 10.1590/S1519-69842005000200009
Fischer J., Lindenmayer D. B. (2000). An assessment of the published results of animal relocations. Biol. Conserv. 96, 1–11. doi: 10.1016/S0006-3207(00)00048-3
González R., Berlinguer F., Espeso G., Ariu F., Del Olmo A., Garde J. J., et al. (2008). Use of a neuroleptic in assisted reproduction of the critically endangered Mohor gazelle (Gazella dama mhorr). Theriogenology 70, 909–922. doi: 10.1016/j.theriogenology.2008.05.052
Kaim M., Bloch A., Wolfenson D., Braw-Tal R., Rosenberg M., Voet H., et al. (2003). Effects of GnRH administered to cows at the onset of estrus on timing of ovulation, endocrine responses, and conception. J. Dairy Sci. 86, 2012–2021. doi: 10.3168/jds.S0022-0302(03)73790-4
Kumar P. S., Saravanan D., Rajasundaram R. C., Selvaraju M., Kathiresan D. (2003). Serum oestradiol and progesterone profiles and their relationship with superovulatory responses in Tellicherry goats treated with eCG and FSH. Small Ruminant Res. 49, 69–77. doi: 10.1016/S0921-4488(03)00071-3
Liu J., Sirois J. (1998). Follicle size-dependent induction of prostaglandin G/H synthase-2 during superovulation in cattle. Biol. Reprod. 58, 1527–1532. doi: 10.1095/biolreprod58.6.1527
McCorkell R. B., Woodbury M. R., Adams G. P. (2008). Induction of ovarian follicular wave emergence in wapiti (Cervus elaphus). Theriogenology 70, 1017–1023. doi: 10.1016/j.theriogenology.2008.04.030
Mertes K., Stabach J. A., Songer M., Wacher T., Newby J., Chuven J., et al. (2019). Management background and release conditions structure post-release movements in reintroduced ungulates. Front. Ecol. Evol. 7. doi: 10.3389/fevo.2019.00470
Monfort S. L., Wemmer C., Kepler T. H., Bush M., Brown J. L., Wildt D. E. (1990). Monitoring ovarian function and pregnancy in Eld’s deer (Cervus eldi thamin) by evaluating urinary steroid metabolite excretion. J. Reprod. fertility 88, 271–281. doi: 10.1530/jrf.0.0880271
Parker I. D., Watts D. E., Lopez R. R., Silvy N. J., Davis D. S., McCleery R. A., et al. (2008). Evaluation of the efficacy of florida key deer translocations. J. Wildlife Manage. 72, 1069–1075. doi: 10.2193/2007-025
Peixoto M. G. C. D., Bergmann J. A.G., Fonseca C. G., Penna V. M., Pereira C. S. (2006). Effects of environmental factors on multiple ovulation of zebu donors. Arquivo Brasileiro Medicina Veterinária e Zootecnia 58, 567–574. doi: 10.1590/S0102-09352006000400019
Pereira R. J. G. (2010). “Male reproduction,” in Neotropical Cervidology: Biology and Medicine of Latin American Deer. Eds. Duarte J. M. B., González S. (FUNEP/IUCN, Jaboticabal), 39–50.
Pereira R. J. G., Polegato B. F., Souza S., Negrão J. A., Duarte J. M. B. (2006). Monitoring ovarian cycles and pregnancy in brown brocket deer (Mazama gouazoubira) by measurement of fecal progesterone metabolites. Theriogenology 65, 387–399. doi: 10.1016/j.theriogenology.2005.02.019
Piovezan U., Tiepolo L. M., Tomas W. M., Duarte J. M. B., Varela D., Filho J. S. M. (2010). “Marsh deer Blastocerus dichotomus (Illiger 1815),” in Neotropical Cervidology: Biology and Medicine of Latin American Deer. Eds. Duarte J. M. B., González S. (FUNEP/IUCN, Jaboticabal, São Paulo), 66–76.
Polegato B. F., Zanetti E. S., Duarte J. M. B. (2018). Monitoring ovarian cycles, pregnancy and post-partum in captive marsh deer (Blastocerus dichotomus) by measuring fecal steroids. Conserv. Physiol. 6, cox073. doi: 10.1093/conphys/cox073
Ramos H. G. C. (2004). O ciclo do chifre do cervo-do-pantanal: Aspectos ecoloǵ icos e reprodutivos. (master’s thesis). São Paulo State University (UNESP), São Paulo, Brazil.
Rola L. D., Buzanskas M. E., Melo L. M., Chaves M. S., Freitas V. J. F., Duarte J. M. B. (2021). Assisted reproductive technology in neotropical deer: A model approach to preserving genetic diversity. Animals: an Open Access J. MDPI 11, 1961. doi: 10.3390/ani11071961
Rola L. D., Galindo D. J., Freitas V. J.F., Duarte J. M. B. (2023). “The South American deer,” in Assisted Reproduction in Wild Mammals of South America. (CRC Press, Boca Raton), 7–28. doi: 10.1201/9781003231691-3
Roldan E. R. S., Gomendio M., Garde J. J., Espeso G., Ledda S., Berlinguer F., et al. (2006). Inbreeding and reproduction in endangered ungulates: preservation of genetic variation through the Organization of Genetic Resource Banks. Reprod. Domest. Anim. = Zuchthygiene 41 Suppl 2, 82–92. doi: 10.1111/j.1439-0531.2006.00772.x
Ryckman M. J., Rosatte R. C., McIntosh T., Hamr J., Jenkins D. (2010). Postrelease dispersal of reintroduced elk (Cervus elaphus) in Ontario, Canada. Restor. Ecol. 18, 173–180. doi: 10.1111/j.1526-100X.2009.00523.x
Seddon P. J., Armstrong D. P., Maloney R. F. (2007). Developing the science of reintroduction biology. Conserv. Biol. 21 (2), 303–312. doi: 10.1111/j.1523-1739.2006.00627.x
Shorrock B. (1980). A Origem da Diversidade as Bases Geneticas da Evolução. TA Queiroz (Sao Paulo: Editora da Universidade de Sao Paulo).
Stringfellow D. A., Seidel S. M. (1999). Manual da Sociedade Internacional de Transferência de Embriões: um guia de procedimento e informação geral para uso da tecnologia de transferência de embriões, enfatizando precauções sanitárias. 3rd edn (Uberlândia: Sociedade Brasileira de Transferência de Embriões).
Szűcs M., Melbourne B. A., Tuff T., Weiss-Lehman C., Hufbauer R. A. (2017). Genetic and demographic founder effects have long-term fitness consequences for colonising populations. Ecol. Lett. 20 (4), 436–444. doi: 10.1111/ele.12743
Szabó M. P. J., Labruna M. B., Pereira M. C., Duarte J. M. B. (2003). Ticks (Acari: Ixodidae) on wild marsh-deer (Blastocerus dichotomus) from Southeast Brazil: infestations before and after habitat loss. J. Med. Entomology 40, 268–274. doi: 10.1093/jmedent/40.3.268
Szabó M. P. J., Castro M. B., Ramos H. G.C., Garcia M. V., Castagnolli K. C., Pinter A., et al. (2007). Species diversity and seasonality of free-living ticks (Acari: Ixodidae) in the natural habitat of wild Marsh deer (Blastocerus dichotomus) in Southeastern Brazil. Veterinary Parasitol. 143, 147–154. doi: 10.1016/j.vetpar.2006.08.009
Teixeira C. P., Azevedo C. S., Mendl M., Cipreste F. C., Young R. J. (2007). Revisiting translocation and reintroduction programmes: the importance of considering stress. Anim. Behav. 73 (1), 1–13. doi: 10.1016/j.anbehav.2006.06.002
Tuberville T. D., Norton T. M., Todd B. D., Spratt J. S. (2008). Long-term apparent survival of translocated gopher tortoises: A comparison of newly released and previously established animals. Biol. Conserv. 141, 2690–2697. doi: 10.1016/j.biocon.2008.08.004
Twagiramungu H., Guilbault L. A., Dufour J. J. (1995). Synchronization of ovarian follicular waves with a gonadotropin-releasing hormone agonist to increase the precision of estrus in cattle: a review. J. Anim. Sci. 73, 3141–3151. doi: 10.2527/1995.73103141x
Warne R. K., Chaber A. L. (2023). Assessing disease risks in wildlife translocation projects: A comprehensive review of disease incidents. Animals 13 (21), 3379. doi: 10.3390/ani13213379
Xu Z., Garverick H. A., Smith G. W., Smith M. F., Hamilton S. A., Youngquist R. S.. (1995). Expression of follicle-stimulating hormone and luteinizing hormone receptor messenger ribonucleic acids in bovine follicles during the first follicular wave. Biol. Reprod. 53, 951–957. doi: 10.1095/biolreprod53.4.951
Zanetti E. S., Duarte J. M. B. (2008). Livro de registro genealógico da população cativa de cervo-do-pantanal (Blastocerus dichotomus). (Jaboticabal: Fundação de Apoio a Pesquisa, Ensino e Extensão (FUNEP)).
Zanetti E. S., Duarte J. M. B. (2012). Comparison of three protocols for superovulation of brown brocket deer (Mazama gouazoubira). Zoo Biol. 31, 642–655. doi: 10.1002/zoo.20428
Zanetti E. S., Duarte J. M.B., Polegato B. F., Garcia J. M., Canola J. C. (2010a). “Assisted reproductive technology,” in Neotropical Cervidology: Biology and Medicine of Latin American Deer. Eds. Duarte J. M. B., Gonzales S. (FUNEP/IUCN, Jaboticabal, São Paulo), 255–270.
Zanetti E. S., Munerato M. S., Cursino M. S., Duarte J. M. B. (2014). Comparing two different superovulation protocols on ovarian activity and fecal glucocorticoid levels in the brown brocket deer (Mazama gouazoubira). Reprod. Biol. endocrinology: RB&E 12, 24. doi: 10.1186/1477-7827-12-24
Zanetti E. S., Polegato B. F., Duarte J. M. B. (2009). “Primeiro relato de inseminação artificial com tempo fixo em Veado-mateiro (Mazama americana),” in Palestras apresentadas no XVIII Congresso Brasileiro de Reprodução Animal.(Belo Horizonte, Minas Gerais: Colégio Brasileiro de Reprodução Animal), 14.
Keywords: Blastocerus dichotomus, conservation program, assisted reproductive techniques, genetics, ecology
Citation: Galindo DJ, Peres PHdF, Zanetti EdS, de Oliveira ML, Rola LD and Duarte JMB (2025) Embryo reintroduction to enhance genetic diversity in a Marsh deer population: first attempt, outcomes, challenges, and future perspectives. Front. Conserv. Sci. 6:1537563. doi: 10.3389/fcosc.2025.1537563
Received: 01 December 2024; Accepted: 24 February 2025;
Published: 14 March 2025.
Edited by:
José Luis Ros-Santaella, Czech University of Life Sciences Prague, CzechiaReviewed by:
Benjamin James Pitcher, Macquarie University, AustraliaCopyright © 2025 Galindo, Peres, Zanetti, de Oliveira, Rola and Duarte. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: José Maurício Barbanti Duarte, bWF1cmljaW8uYmFyYmFudGlAdW5lc3AuYnI=; David Javier Galindo, ZGdhbGluZG9oQHVubXNtLmVkdS5wZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.