 Austin J. Housley
Austin J. Housley Gary I. Baird2
Gary I. Baird2 David R. Burchfield
David R. Burchfield Steven L. Petersen
Steven L. Petersen Jericho C. Whiting
Jericho C. Whiting
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Conserv. Sci., 11 April 2025
Sec. Plant Conservation
Volume 6 - 2025 | https://doi.org/10.3389/fcosc.2025.1522727
Introduction: Rare and endemic plants that inhabit sand dunes are of conservation concern across the world. The St. Anthony Evening Primrose (Oenothera psammophila, (A. Nelson & J.F. Macbr.) W.L. Wagner, Stockhouse & Klein) is a rare endemic plant that occurs on the St. Anthony Sand Dunes (SASD) in Idaho, USA. This evening primrose is classified by the Bureau of Land Management (BLM) as a range-wide imperiled species with high endangerment risk and was listed as a candidate species under the Endangered Species Act in 1975, but was removed from candidacy in 1996 after extensive population estimates. Little research has been conducted on this species since these population estimates conducted by the BLM in 1994.
Methods: We conducted surveys in 2024 to compare the current geographic distribution and number of individuals of this species with similar data collected in 1994. We hypothesized that the distribution and number of these plants have decreased in the last 30 years.
Results: In 1994, it was documented that the St. Anthony Evening Primrose occupied 1,403 ha with 38,426 plants. In 2024, we documented that the St. Anthony Evening Primrose covered 829 ha (41% decrease; X2 = 106.85, p < 0.0001) with 19,737 plants (49% decrease; X2 = 3035.9, p < 0.0001). The distribution and number of plants decreased in two of the three regions in our study area but did not decrease in one of those regions. From 1994 to 2018–2021, there has been a 16,909% increase in the number of Off-Highway Vehicles (OHV) on the SASD during June, which is likely correlated to the decrease in distribution and number of St. Anthony Evening Primrose.
Discussion: Although we documented decreases in population distribution and number of plants in two regions of our study area, our data precluded us from quantifying the extent of those decreases that were caused by natural dune movement, variation in precipitation and soil moisture, or OHV use. Our results, however, do provide biologists with information about this species that can be used for land-use management plans and comparisons with future studies of the St. Anthony Evening Primrose. Lastly, our results support reevaluating the S-rank for this species.
Rare plant species that are endemic to sand dune systems are generally of conservation concern. These environments are constantly in flux due to dune migration patterns and increasing human recreational activities. Subsequently, plant species growing on dunes are typically sensitive to alterations in their habitats (Casazza et al., 2007; Thompson and Schlacher, 2008; Knisley et al., 2018; Hogan et al., 2020). For example, plants can become affected by shifting dunes that overtake and bury individual plants (Chadwick and Dalke, 1965; Lee, 1983; Maun, 1998, 2004; Hoover et al., 2018). In recent years, the diversity of recreational activities, especially off-highway vehicle (OHV) use, have increased on sand dunes and other public lands, and those activities now account for the greatest threat to rare endemic species that occur in these areas (Tilman and Lehman, 2001; Wagoner, 2006; Groom et al., 2007; Thompson and Schlacher, 2008; Hernández-Yáñez et al., 2016; Knisley et al., 2018; Hogan et al., 2020).
In western North America, two species of evening primrose (section Pachylophus, genus Oenothera) are endemic to sand dune systems: the Colorado Dunes Evening Primrose (O. harringtonii W.L.Wagner, Stockh. & W.M.Klein) and the St. Anthony Dunes Evening Primrose (O. psammophila (A. Nelson & J.F. Macbr.) W.L. Wagner, Stockhouse & Klein, hereafter the St. Anthony Evening Primrose or the evening primrose). The St. Anthony Evening Primrose has the smallest distribution of these two evening primrose species (Patsis et al., 2021). The St. Anthony Evening Primrose is an herbaceous perennial plant endemic to the St. Anthony Sand Dunes (SASD) in Idaho, USA (Lee, 1983; Patsis et al., 2021). This species was once classified as part of the O. caespitosa (Nutt.) group but was subsequently separated because of morphological and genetic variations (Patsis et al., 2021). The Bureau of Land Management (BLM) currently classifies the St. Anthony Evening Primrose as a range-wide imperiled species with high endangerment risk, and this plant was listed as a candidate species under the Endangered Species Act (ESA) between 1975 and 1996 (USFWS, 1996; Mancuso et al., 2019). The St. Anthony Evening Primrose was subsequently removed as a candidate species from the ESA after a series of population inventories and threat analysis studies conducted in the mid-1990s (Michaelson and Johnson, 1994; Rice, 1995; McFarling and Mahnami, 1996). These studies concluded that at the time, threats from OHV use, trampling, and a potential spread of noxious species of weeds were not significant for survival of this evening primrose species (USFWS, 1996; McFarling and Mahnami, 1996). The St. Anthony Evening Primrose is currently ranked as G3S3 and BLM Type 2- Rangewide/Globally Imperiled Species with high endangerment risk (Mancuso et al., 2019; NatureServe, 2025).
The distribution and number of St. Anthony Evening Primrose has potentially been affected by increased OHV use on the SASD in recent years. In 2003, over 100,000 recreationists visited the SASD, and this number has increased 7–10% annually since that time (Wagoner, 2006). By 2016, this number had increased to over 250,000 recreationists (Hughes and Paveglio, 2019). Recreationists visit the SASD to camp, ride horses, hike, and ride OHVs (Wagoner, 2006; Hughes and Paveglio, 2019). Of these activities 87% come to ride OHVs across the dune environment (Wagoner, 2006). While once thought to be inconsequential for this evening primrose species (Lee, 1983; McFarling and Mahnami, 1996), OHV use has since been documented to have negative effects on other plant species in similar dune systems (Groom et al., 2007; Hogan et al., 2020). These negative effects range from soil substrate compaction to direct plant mortality from OHV tires, which results in a reduction of species richness, diversity, and vegetation cover throughout the area (Kutiel et al., 2000a). The most recent research that documented the distribution and number of this evening primrose occurred in the mid-1990s (Michaelson and Johnson, 1994; Rice, 1995; McFarling and Mahnami, 1996); therefore, current information about this rare and endemic species is needed.
The St. Anthony Evening Primrose historically experienced higher rates of recruitment than mortality (Lee, 1983). The most common cause of mortality is from plants on the peripheries of interdune areas that cannot establish new recruits before becoming covered by increasing sand as the dunes move (Lee, 1983; McFarling and Mahnami, 1996). It is possible for a subpopulation of this evening primrose to be eliminated after extended periods of low recruitment years and low precipitation (Lee, 1983). However, a single spring of greater than average precipitation can replace most lost individuals (Lee, 1983). Historically, recruitment rates were similar between high-use OHV sites and control sites and that dune migration played the greatest role in decreasing recruitment (Lee, 1983; McFarling and Mahnami, 1996). The dispersal mechanisms of this species are not well documented but are thought to be a combination of wind and small mammal (Dipodomys spp. Gray) dispersal (Lee, 1983; McFarling and Mahnami, 1996). Additionally, dune movement rates have remained consistent at a rate of 0.29–2.4 m/year since the mid-1990s (Hoover et al., 2018). Although the St. Anthony Evening Primrose historically demonstrated resilience to natural variation in recruitment rates, increased OHV use in recent years could potentially affect the distribution of this species.
Endemic species are often of conservation concern due to their restricted geographic range and typically unique habitat adaptations (Hernández-Yáñez et al., 2016). Endemism provides additional challenges in the context of increased anthropogenic recreation, such as OHVs, because these species are not readily capable of adapting to changes in their environment (Hernández-Yáñez et al., 2016; Coelho et al., 2020). Areas with unique geologies, such as dune systems, tend to increase the presence of endemic species because these species are well adapted to grow and disperse in these locations (Locklear, 2017). Because endemic species, like the St. Anthony Evening Primrose, are specialized to existing in only these unique areas it is essential to have a more complete understanding of their conditions.
Understanding the historical and current distributions of plants, especially sand dune specialists, is essential for their conservation (Chadwick and Dalke, 1965; Lee, 1983; Mills and Schwartz, 2005; Hernandez-Stefanoni and Ponce-Hernandez, 2006; Casazza et al., 2007). Changes in historic species distribution and population numbers can be used as an indicator to track the effects of anthropogenic impacts on rare and endemic sand dune species like the St. Anthony Evening Primrose (Lee, 1983; Groom et al., 2007; Hogan et al., 2020). Herein, we documented the current number of St. Anthony Evening Primrose plants on the main dune complex of the SASD and showed how the distribution of the subpopulations and number of plants have potentially changed with increased OHV use since the previous surveys. We hypothesized that the distribution and number of the St. Anthony Evening Primrose have decreased in the last 30 years. Because of increased OHV use on the SASD, we predicted that the geographical distribution of the St. Anthony Evening Primrose has decreased in the areas most frequented by OHVs. We also predicted that the number of the St. Anthony Evening Primrose plants decreased in those areas. The results of our research will help with the conservation of this species on the SASD, as well as the conservation of rare and endemic plants on other dune systems.
The SASD occur on the Snake River Plain of Idaho, USA, and cover approximately 4,300 ha of that area with white quartz sand (Wagner, 2005; Hoover et al., 2018). The SASD formed approximately 10,000 years ago when the shores of Mud and Market lakes, located ~32 km to the west, shrank at the end of the last ice age (Rich et al., 2015). The climate of this area is a dry desert with hot summers and cold winters with most of the precipitation occurring in early spring with elevations ranging from 1,341 to 3,858 m (Johnson et al., 2024). The prevailing winds moved sand from the historical lake shores northeastward where it accumulated on basalt outcroppings forming the SASD (NASA Earth Observatory, 2010). Currently, the dunes can be anywhere from 15 to 121 m high and are constantly shifting and moving with high wind speeds typical of that region (Rich et al., 2015). The shape of individual dunes is typically steeper on the leeward side with gentler slopes on the windward sides (Chadwick and Dalke, 1965). Over the past 70 years, the SASD have been moving in a northeast direction approximately 0.29–2.4 m/year (Hoover et al., 2018). Currently, our study area is managed by the BLM and is designated as part of the Sand Mountain Wilderness Study Area with closures from January 1–April 1 yearly for ungulate migration (Bureau of Land Management, 2024). We focused our study on the largest contiguous dune in the SASD and divided the study area into three sections based on different OHV use/geologic features for sampling purposes (Lee, 1983; B. Cisna, personal communication 2024). These sections were selected from the 1994 primrose inventory, and we designated them as the West, Middle, and East regions. In 1994, those regions contained approximately 647, 418, and 338 hectares of evening primrose habitat, respectively (Michaelson and Johnson, 1994; Figure 1).
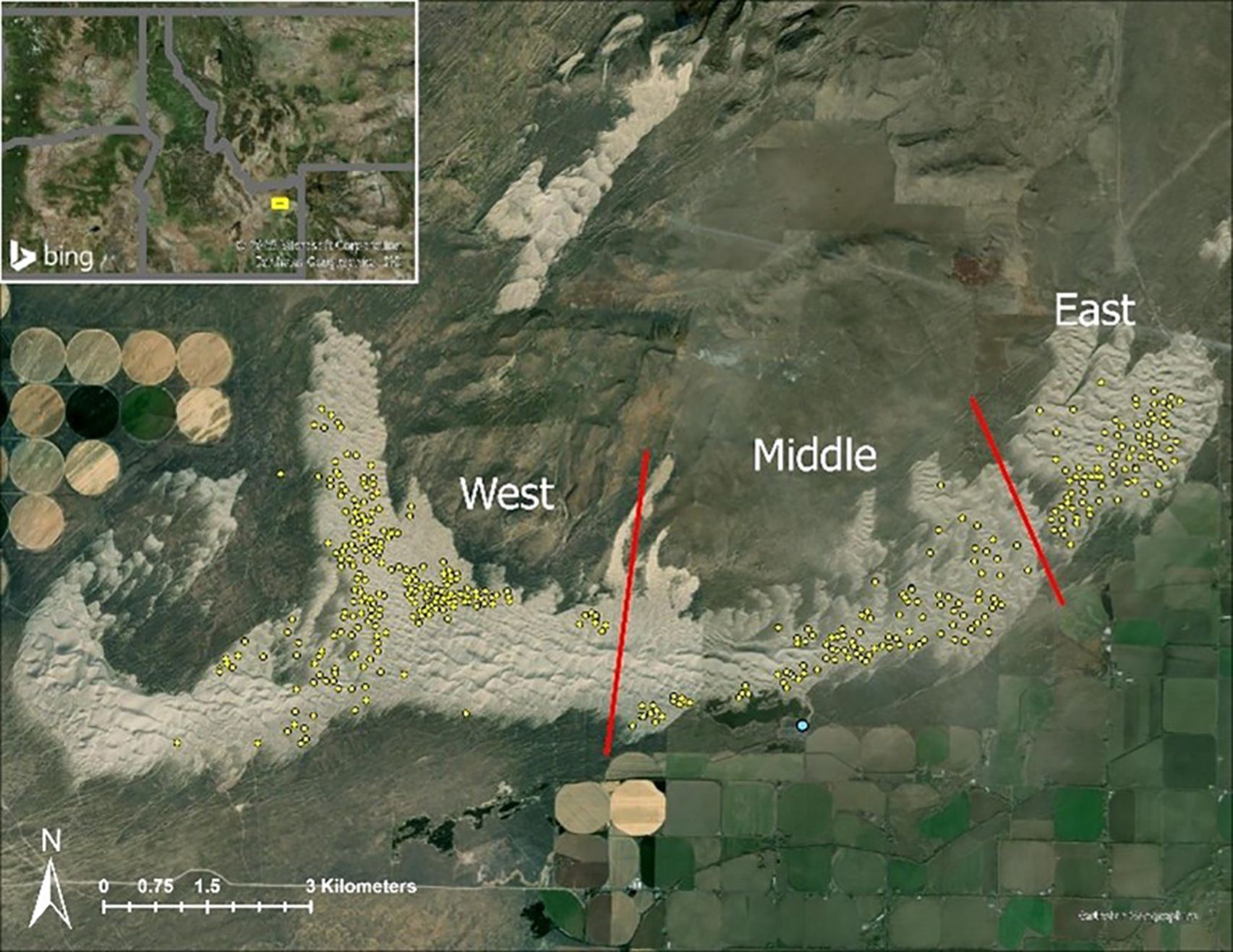
Figure 1. Our study area where we documented the distribution and number of St. Anthony Evening Primrose. Yellow dots represent the 1994 subpopulations of this evening primrose. Regions were divided into the West, Middle, and East based on topological and amount of OHV traffic. The light blue dot represents the location of the TRAFx OHV Counter.
The area surrounding the dunes were primarily composed of a mix of Rocky Mountain juniper (Juniperus scopulorum Sarg.) and sagebrush (Artemisia tridentata spp. Nutt. and A. tripartita Rydb.) steppe shrubland to the north and east, while the southern and western areas are a mix of juniper/shrubland community and farmland (Chadwick and Dalke, 1965). Other plants that occurred in that region were rabbitbrush (Ericameria spp. Nutt., Chrysothamnus visicidiflorus Hook.), horsebrush (Tetradymia spp. DC), native grasses (Eriocoma hymenoides Roem. & Schult., Hesperostipa comata Trin. & Rupr., Elymus L., and Leymus spp. Hochst.), and other nonnative grass species (Agropyron cristatum (L.) Gaetrn., and Bromus spp. L.) both on and around the SASD (Chadwick and Dalke, 1965; Lee, 1983). Other herbaceous forbs such as the pale evening primrose (O. pallida Lindl.) and dune scurf pea (Ladeania lanceolata (Pursh) Reveal & A. N. Egan) commonly occurred with the St. Anthony Evening Primrose (Chadwick and Dalke, 1965; Lee, 1983). Potential impacts from herbivory on the St. Anthony Evening Primrose occurred from migratory ungulates including Rocky Mountain elk (Cervus canadensis Erxleben), mule deer (Odocoileus hemionus Rafinesque), and moose (Alces alces Linneaus) (Lee, 1983). Rabbits (Leporidae Fischer) and kangaroo rats (Dipodomys spp.) may also have foraged on the leaves and seed pods of the evening primrose (Lee, 1983). Weather data were recorded from the nearest station (Pocatello region—about 145 km southwest of the SASD, Table 1, NOAA, 2025).
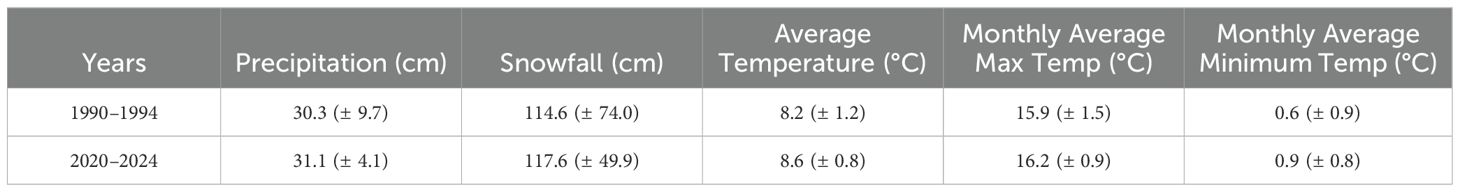
Table 1. Annual (± SD) precipitation and snowfall, with mean (± SD) temperature and maximum/minimum monthly temperature for the 4 years preceding and 1 year of the 1994 BLM population inventory, and for 4 years previous to our study and 1 year of our study.
In June 1983, it was documented that 38 OHVs entered the SASD from the East region (Lee, 1983). For a comparison of OHVs entering from the Middle region (from a more recently developed BLM campground) in June from 2018 to 2021 to the number of people entering the East region in 1983, we used data from a TRAFx OHV Counter (TRAFx Research Ltd., Alberta, Canada). That counter was installed in 2018 by the BLM and ran consecutively for 4 years at the SASD main entrance, which was located in the southwest portion of the Middle Region (Figure 1). That counter recorded the number of OHV riders entering and exiting the SASD (Manley et al., 2004). The counter used magnetometers to identify vehicle traffic by sensing metals within a radius of 1–6 m as they passed through the main entrance (Manley et al., 2004). These devices have been used to effectively monitor the effects of OHV traffic on plant and wildlife species (Trip, 2014; Jones et al., 2017).
The only large-scale population survey of the St. Anthony Evening Primrose occurred in 1994 (Michaelson and Johnson, 1994). The subpopulations were delineated by walking the perimeter with a Trimble Navigation Pathfinder Professionals (CMT) Global Positioning System (GPS) unit and counting the number of St. Anthony Evening Primrose plants within an interdune space from July–September 1994 (McFarling and Mahnami, 1996). During the surveys, the GPS units used were restricted by selective availability—i.e., accuracy of points could vary 20–80 m (McFarling and Mahnami, 1996; Hulbert and French, 2001). For each subpopulation created they documented elevation, latitude, longitude, number of plants (living and dead), and calculated an area of each subpopulation (McFarling and Mahnami, 1996). Those researchers identified 87,324 individual St. Anthony Evening Primrose plants, 38,426 (44%) of which were located on the main dune complex (Michaelson and Johnson, 1994; Figure 1).
For our surveys, we chose to revisit the subpopulation points created during the previous surveys in 1994. Using the locations (i.e., latitude and longitude) from the 1994 survey, we uploaded and projected points into a map using ArcGIS Pro 3.2.2 (ESRI, Redlands, CA) (Figure 1). Between 1 and 4 individuals navigated to each of these points and systematically searched the interdune spaces where the St. Anthony Evening Primrose was expected to grow within 100–300 m (depending on the size of the interdune space) of the point in all directions. When we observed evening primrose, we recorded each individual plant location using Field Maps (ESRI, Redlands, CA). We determined that individuals were different plants when they were separated by ≥ 30.5 cm. We then recorded plants as living or dead. Plants were determined as dead when there were no indications of green colored vegetation indicating photosynthetic activity was not taking place on any part of the mat/mound forming a single St. Anthony Evening Primrose. Our surveys started on April 17, 2024, and continued through June 18, 2024, and occurred for approximately 4–7 hours/day from 8:00 a.m. to 3:00 p.m. MST.
Our 2024 points were uploaded into ArcGIS Online and then downloaded into ArcGIS Pro to compare how the geographic distribution and population number had changed since 1994. To calculate geographic distribution, we used the Density-Based Clustering Scan (DBSCAN), Mean Center, and Kernal Density tools in ArcGIS Pro. We first used DBSCAN to identify individual plants and to generate subpopulations of at least 2 plants within 50 m (Malambo et al., 2019; Priya et al., 2020). After identifying locations for potential individuals and subpopulations, we then used the Mean Center tool to calculate the weighted centroids for each subpopulation in 2024. Using the Total Number of Plants field in both 1994 and 2024 layers, we calculated the Kernal Density and produced maps showing the change in distribution of subpopulations from 1994 to 2024.
We designated the Kernal Density tool settings as follows—Population Field: total number of living and dead plants, Area Units: km2, Output Cell Size: default, Output Cell Values: densities, Method: geodesic. We then adjusted the classes of the 2024 heatmap to match the classes of the 1994 heatmap to standardize heatmap categories. To test the differences between the distribution and number of plants in the different regions between 1994 and 2024, we used a chi-square test with a post-hoc pairwise comparison and a Bonferroni correction in the RVAideMemoire package (Herve, 2023). All statistical analysis was completed in R version 4.3.21 (R Core Team, 2023).
In 1994, the St. Anthony Evening Primrose plants occupied 1,403 ha of the SASD (Figure 2). In 2024, the area occupied by those plants was reduced to 829 ha (Figure 2). From 1994 to 2024, the area occupied by those plants that were identified with the Kernal Density estimator decreased 41% (Figures 2, 3; X2 = 106.85, p-value < 0.0001) with only 673 ha overlapping between both years. The total hectares occupied by the evening primrose decreased 45% in the West region (p-value < 0.0001) and 71% in the Middle region (Figure 3; p-value < 0.0001). There was no evidence of a decrease in the hectares occupied by the evening primrose in the East region (Figure 3; p-value = 1). In 1994, the evening primrose occurred in a larger area in the West region, and in 2024 the larger area with evening primrose occurred in the East region (Figure 2).
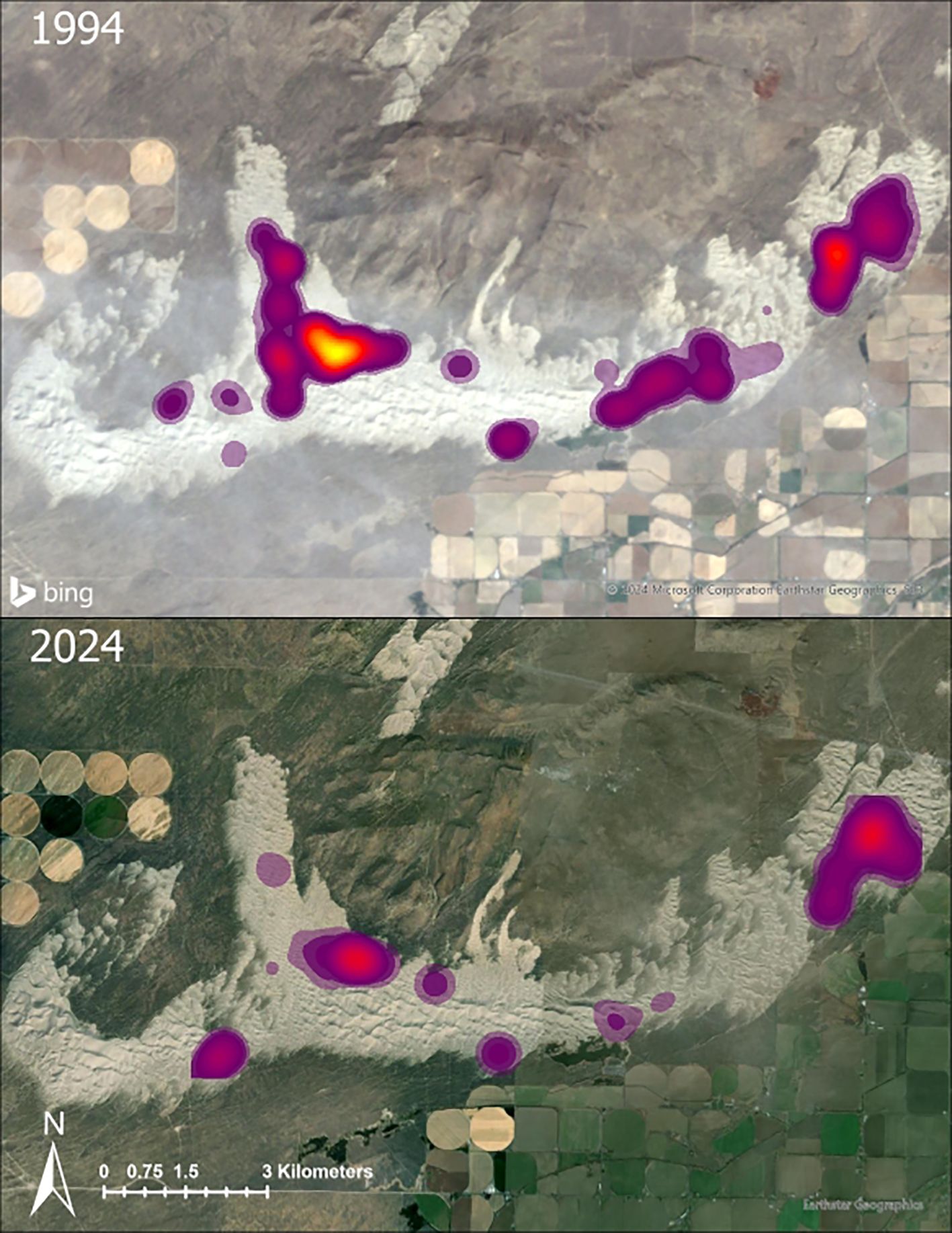
Figure 2. Comparison of the St. Anthony Evening Primrose distributions using Kernal Density estimates between 1994 (top) and 2024 (bottom) in Idaho, USA. The basemap of the 1994 Inventory (top) is Landsat 5 imagery and the 2024 Inventory (bottom) was done with current Landsat imagery.
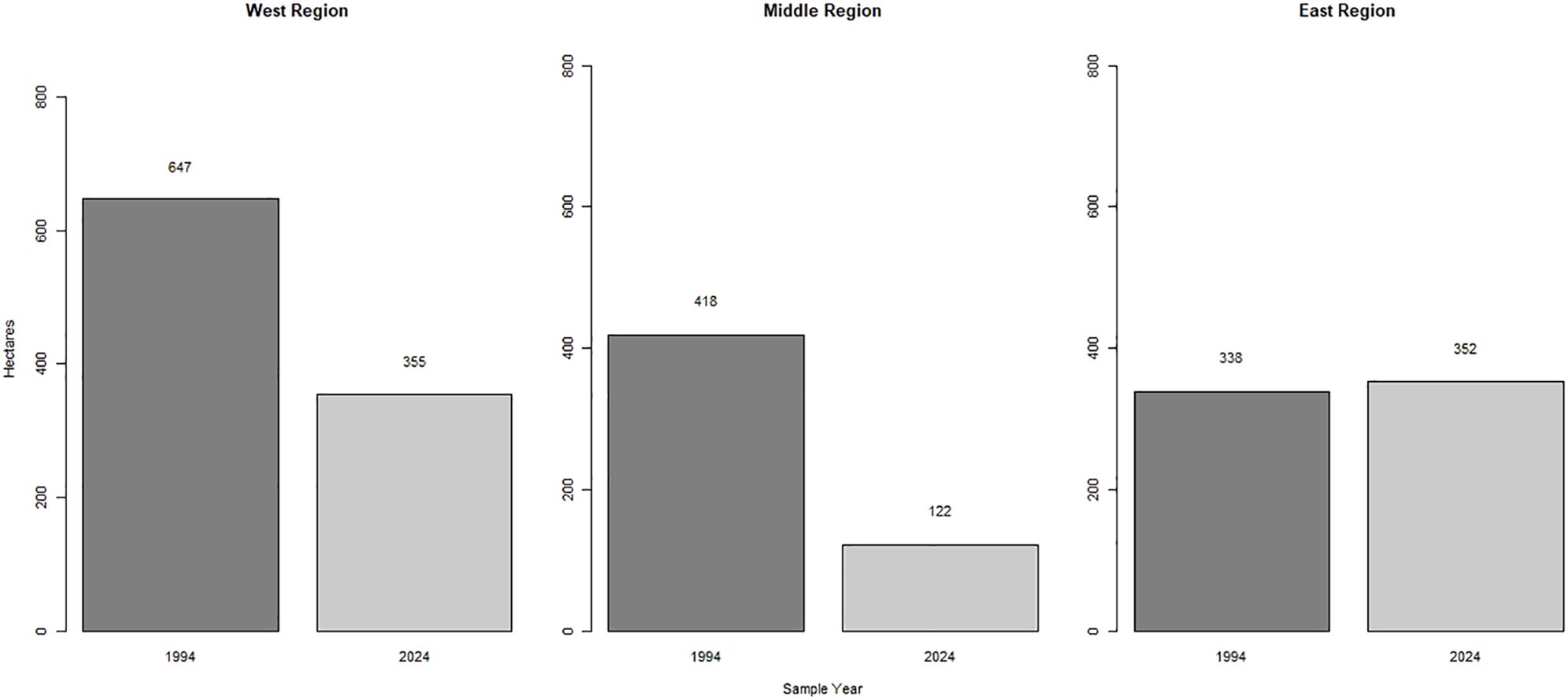
Figure 3. Total ha occupied by the St. Anthony Evening Primrose by region between 1994 and 2024 on the St. Anthony Sand Dunes in southeastern Idaho.
In 1994, there were 38,426 St. Anthony Evening Primrose plants (35,640 living, 2,786 dead) and 428 subpopulations. In 2024, we documented 19,737 St. Anthony Evening Primrose plants (18,443 living, 1,294 dead) and 331 subpopulations (Figure 4). In the last 30 years the total number of plants decreased 49% on the SASD (X2 = 3035.9, p-value < 0.0001). The total number of plants decreased 59% in the West region (p-value < 0.0001) and 73% in the Middle region (p-value < 0.0001). There was no evidence that there was a decrease in the number of plants in the East region (Figure 4; p-value = 0.29). In 1994, the highest concentration of plants occurred in the West region, and in 2024 the highest concentration of plants occurred in the East region (Figure 2). We documented a 16,909% increase in OHV riders in June from 1983 to 2019–2021 (Figure 5). Precipitation, snowfall, and temperatures were comparable between the 1994 study period and our 2024 study (Table 1).
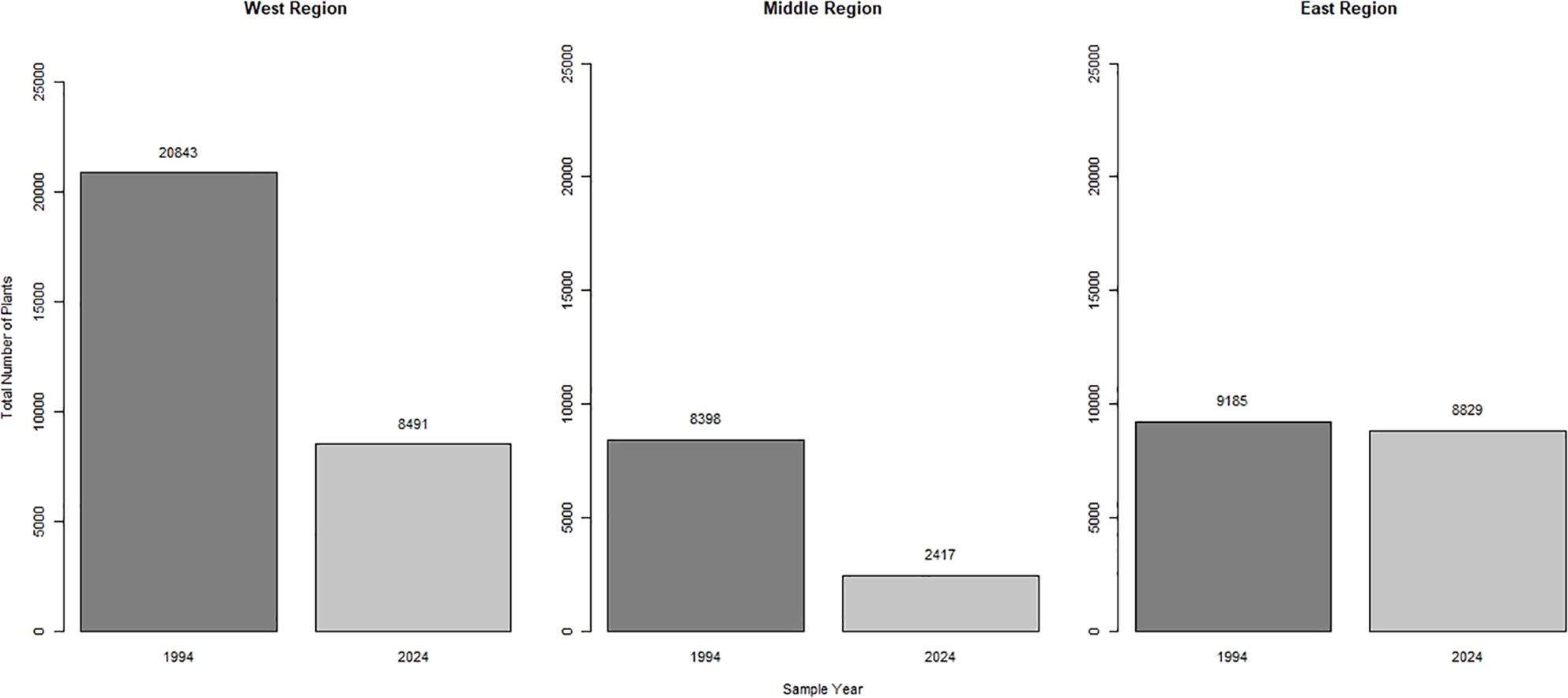
Figure 4. Total number of St. Anthony Evening Primrose plants by region between 1994 and 2024 on the St. Anthony Sand Dunes in southeastern Idaho.
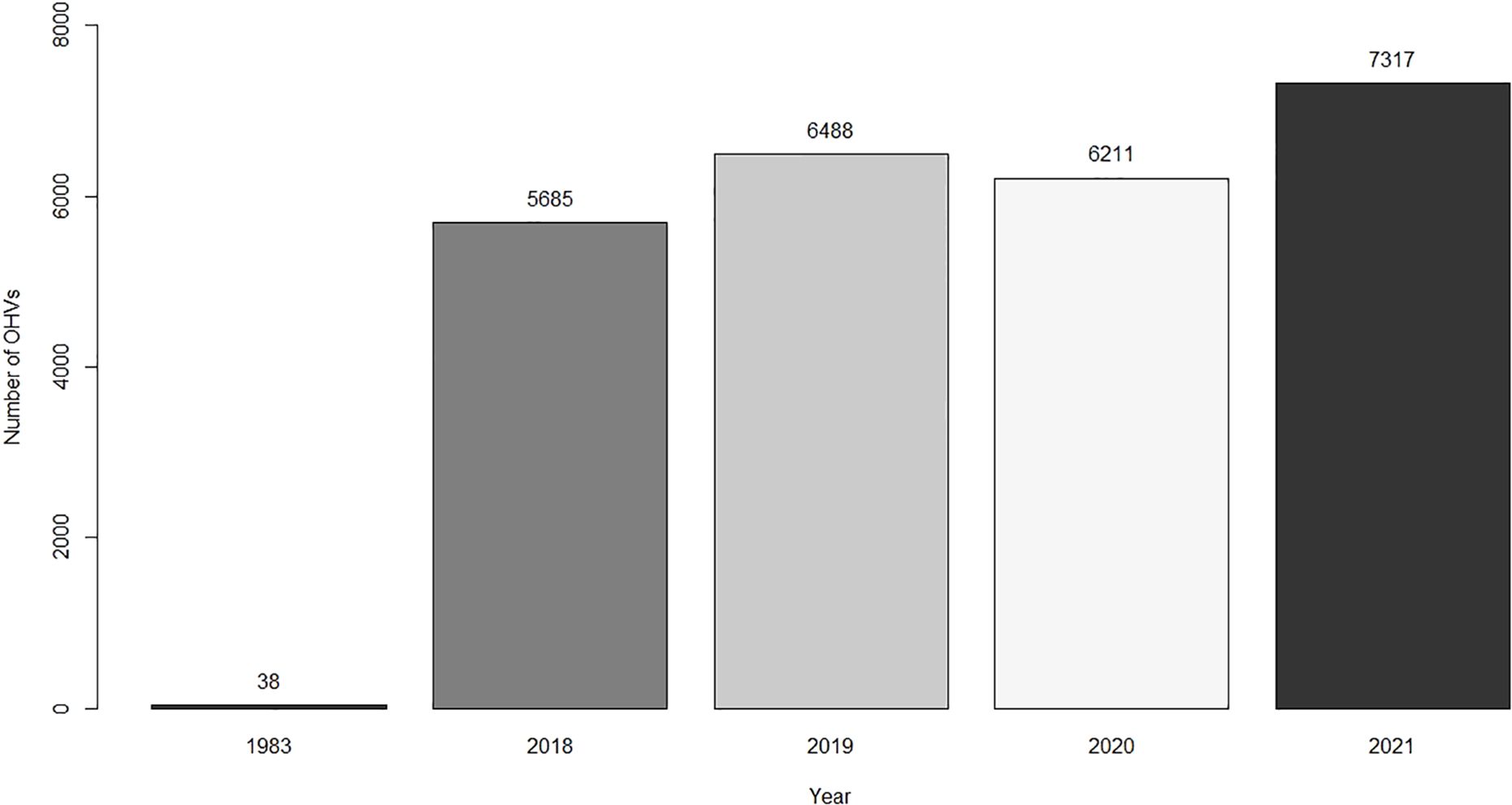
Figure 5. Total number of OHVs on the St. Anthony Sand Dunes during June. The 1983 results were compared with the total number of OHVs entering the Egin Lakes campground in the Middle region of our study area in southeastern Idaho.
We predicted that the distribution of the St. Anthony Evening Primrose would decrease in areas most frequented by OHVs. Indeed, we observed a 63% decrease in the distribution of St. Anthony Evening Primrose in the West and Middle regions (Figure 2). Currently, these regions are the highest use areas for OHV riders due to the main dune entrance in the Middle region and large dune hills for OHV recreation (Figures 1, 5; Cisna, Personal Communication, 2024). Previous research about evening primrose species on dune systems concluded that OHV riders had little impact on the overall distribution of the plants because the plants were able to recruit new individuals at a greater rate than they were degraded (Lee, 1983; Pavlik and Barbour, 1988; McFarling and Mahnami, 1996). Current research, however, indicates that the number of OHV riders has substantially increased since those studies (Wagoner, 2006; Hughes and Paveglio, 2019), and other studies have documented that increasing OHV use negatively affects the distribution of native vegetation (Hogan et al., 2020). For example, the distribution of vegetation on one coastal dune system in New Jersey, USA, was reduced by 86% by OHV use (Kelly, 2014). Our data provides important insights on how OHV use can potentially negatively affect the distribution of an endemic dune plant species.
We also predicted that the number of St. Anthony Evening Primrose would decrease in areas that were most frequently used by OHVs. We observed a combined 63% decrease in those plants in the Middle and West regions (Figure 4). This represents a decline from 29,241 to 10,908 plants and 345 to 232 subpopulations in the Middle and West regions. (Figure 4). The most likely cause of this decline is the 16,909% increase in OHV recreational activity on the SASD (Figure 5). Previous research concluded in the mid-1990s that OHV use on the SASD was inconsequential for the survival of the St. Anthony Evening Primrose plants (Rice, 1995; McFarling and Mahnami, 1996). Research in other similar systems, however, has determined that that recent increases in OHV activity presents one of the most significant threats to the survival of rare and endemic species (Mills and Schwartz, 2005; Hernández-Yáñez et al., 2016; Custer et al., 2017; Knisley et al., 2018). For example, Peirson’s Milkvetch (Astragalus magdalenae var. peirsonii, Greene) is a threatened species under the ESA that is endemic to dune systems in California, USA, and Sonora, Mexico (Groom et al., 2007). In California, areas that had light OHV use for 3 months experienced a 33% reduction in the survival of this milkvetch and a 4–5 times reduction in its density (Groom et al., 2007). Similarly, our research indicates a decrease in the total number of plants in the Middle and West region of the SASD across 30 years considering an increase in OHV use.
Importantly, our data did not allow us to directly quantify the decrease in the distribution and number of St. Anthony Evening Primrose due to OHV use compared with natural dune movement or variation in precipitation and soil moisture. The SASD are comprised of active and stable dune hills that move in a northeast direction approximately 0.29–2.4 m/year (Rich et al., 2015; Hoover et al., 2018). We predicted that the fringes of the distribution and the number of St. Anthony Evening Primrose plants would decrease naturally from dune movement where the plants would become covered by the shifting dunes (Chadwick and Dalke, 1965). Moreover, in other sand dunes ecosystems, precipitation has been shown to affect the productivity and diversity of plant growth (Amir et al., 2014; Homberger et al., 2024). Our precipitation data indicated that the mean precipitation was higher in 2020 to 2024, but the variation was close to the standard deviations for the 1994 study period and does not solely account for the decline of the evening primrose population (Table 1). These natural factors could potentially have had a negative effect on the distribution and number of the St. Anthony Evening Primrose; however, previous studies documented that the primrose is well-adapted and suited to the changes of its environment (Lee, 1983; McFarling and Mahnami, 1996). Future research should test the effects of dune movement, annual precipitation, and OHV use on the reduction of the distribution and number of the St. Anthony Evening Primrose across several years.
Climate change is often implicated for the variation in the populations of rare endemic species. Precipitation differences play a significant role in the number of individual plants on a year-to-year basis (Casazza et al., 2007; Amir et al., 2014; Homberger et al., 2024). Additionally, whether the precipitation comes in the form of rainfall or snowfall can influence the recruitment of new individuals. Past research of the St. Anthony Evening Primrose has indicated that a single year of above average precipitation can result in the recruitment of enough new individuals to replace those lost during years of less precipitation (Lee, 1983). Precipitation amounts (both rainfall and snowfall) were comparable between 1990–1994 and 2020–2024 (Table 1). Because of this comparable precipitation, future studies should refine their focus to variables such as dune movement and increased OHV use.
Our surveys indicated that the East region of the SASD experienced no significant change in distribution or number of the St. Anthony Evening Primrose compared with similar data from surveys conducted in 1994 (Michaelson and Johnson, 1994). We hypothesis that the reason for that lack of change in the East region is primarily because of the difference in habitat composition between the East region and the West and Middle regions of the SASD. The East region is characterized by a higher density of smaller dune hills (~15 – 30 m in height) and larger basalt ridges and domes in the interdune spaces, which precludes OHV riders from easily entering areas occupied by the evening primrose (Chadwick and Dalke, 1965; Lee, 1983; Rich et al., 2015; Cisna, Personal communication, 2024). The St. Anthony Evening Primrose populations are well adapted to growing on elevated basalt rock outcroppings (Chadwick and Dalke, 1965; Lee, 1983). Other studies have also documented that rock features serve as a natural deterrent of most OHV activity on vegetation (Custer et al., 2017).
Vegetation on dunes systems is essential for several ecosystem services (wildlife habitat, insect populations, soil quality, and climate regulation) for both coastal and inland dune systems (Van der Biest et al., 2017; Knisley et al., 2018; McGuirk et al., 2022). Dune systems also provide habitat for many rare and endemic plant species (Moreno-Casasola, 1988; Kutiel et al., 2000b; Groom et al., 2007; Lamb and Guedo, 2012; Celsi and Giussani, 2019). Additionally, many sand dunes in the USA contain endemic species of tiger beetles (Knisley, 2011; Knisley et al., 2014, 2018). The Idaho dunes tiger beetle (Cicindela arenicola) is endemic to several dune systems in Idaho including the SASD and occupies similar habitats to the St. Anthony Evening Primrose (Knisley, 2011; Knisley et al., 2014). The ecological importance of sand dunes underscores the need for proper management of these areas. For instance, coastal dune systems in Europe have decreased 70% due to increased urbanization in the last century, putting the remaining flora and fauna at great risk (Feola et al., 2011). Because of those reductions, there is a need to document the trends of the unique species inhabiting these dune systems.
Historical comparisons of species spatial distribution and population numbers are useful for determining long-term population trends (Mills and Schwartz, 2005; Hernandez-Stefanoni and Ponce-Hernandez, 2006). Other studies have either focused on smaller areas or on several species across a dune system to analyze the direct effects of human and OHV impacts (Tilman and Lehman, 2001; Casazza et al., 2007; Groom et al., 2007; Hogan et al., 2020). We documented a 41% decrease in spatial distribution and a 49% decrease in the number of St. Anthony Evening Primrose plants on the largest contiguous dune field of the SASD. This dune field contained 44% (38,426 out of 87,324 plants) of St. Anthony Evening Primrose individuals identified by the historical BLM population estimates, and this area alone represents a 23% decline (18,699 fewer plants) from the historic population size (Michaelson and Johnson, 1994). These reductions are most likely the result of a 16,909% increase in OHV use on the SASD. We also documented no reduction in the distribution and number of plants in one region of our study area. Because of increased OHV use on the SASD and the significant decreases in the distribution and number of St. Anthony Evening Primrose there is a need to reevaluate the current S3 status of this species (NatureServe, 2025). Future research should focus on assessing OHV use thresholds (e.g., capacity and patterns) on the SASD to assist and promote the conservation of the St. Anthony Evening Primrose and its habitat (Kutiel et al., 2000a; Thompson and Schlacher, 2008; Hernández-Yáñez et al., 2016; Knisley et al., 2018). Identifying the effects of increased OHV use on the SASD will contribute to the conservation of this and other dune specialists. Lastly, our results provide biologists with current information about this rare endemic plant that can be used for land-use management plans. Our results can also provide an understanding of how tracking plant population changes throughout long-time periods can aid in the conservation of other rare endemic species.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
AH: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Writing – original draft, Writing – review & editing. SP: Project administration, Resources, Supervision, Writing – review & editing. JW: Conceptualization, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Writing – original draft, Writing – review & editing. GB: Conceptualization, Methodology, Supervision, Writing – review & editing. DB: Conceptualization, Software, Supervision, Visualization, Writing – review & editing.
The author(s) declare that financial support was received for the research and/or publication of this article. We thank Brigham Young University and Brigham Young University - Idaho for providing the OHVs necessary to traverse the study area for data collection and field sampling. We also thank Zachary Linton for his assistance as a full-time field intern and his invaluable support to Housley during the field season. Additional thanks to the various students from the Brigham Young University – Idaho Fisheries, Range, and Wildlife Society who participated in the field sampling efforts. We also thank the Idaho Falls Zoo Staff for providing us with the opportunity to present our preliminary findings at the Zoo Exploration Stations and for providing funding for Housley and Linton to complete the field sampling efforts.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Amir R., Kinast S., Tsoar H., Yizhaq H., Zaady E., Ashkenazy Y. (2014). The effect of wind and precipitation on vegetation and biogenic crust covers in the Sde-Hallamish sand dunes. J. Geophysical Research: Earth Surface 119, 437–450. doi: 10.1002/2013JF002944
Bureau of Land Management (2024). St. Anthony sand dunes. Available online at: https://www.blm.gov/visit/st-anthony-sand-dunes (Accessed 2024, Jul 12).
Casazza G., Zappa E., Mariotti M. G., Médail F., Minuto L. (2007). Ecological and historical factors affecting distribution pattern and richness of endemic plant species: the case of the Maritime and Ligurian Alps hotspot. Diversity distributions 14, 47–58. doi: 10.1111/j.1472-4642.2007.00412.x
Celsi C. E., Giussani L. M. (2019). Geographical distribution and habitat characterization of Poa schizantha (Poaceae), a narrow endemic of the coastal sand dunes of the southern Pampas, Argentina. Botanical J. Linn. Soc. 192, 296–313. doi: 10.1093/botlinnean/boz069
Chadwick H. W., Dalke P. D. (1965). Plant succession on dune sands in fremont county, idaho. Ecol. (Durham) 46, 766–780. doi: 10.2307/1934008
Coelho N., Gonçalves S., Romano A. (2020). Endemic plant species conservation: biotechnological approaches. Plants 9, 345. doi: 10.3390/plants9030345
Custer N. A., Defalco L. A., Nussear K. E., Esque T. C. (2017). Drawing a line in the sand: Effectiveness of off-highway vehicle management in California’s Sonoran desert. J. Environ. Manage. 193, 448–457. doi: 10.1016/j.jenvman.2017.02.033
Feola S., Carranza M. L., Schaminée J. H. J., Janssen J. A. M., Acosta A. T. R. (2011). EU habitats of interest: an insight into Atlantic and Mediterranean beach and foredunes. Biodiversity Conserv. 20, 1457–1468. doi: 10.1007/s10531-011-0037-9
Groom J. D., McKinney L. B., Ball L. C., Winchell C. S. (2007). Quantifying off-highway vehicle impacts on density and survival of a threatened dune-endemic plant. Biol. Conserv. 135, 119–134. doi: 10.1016/j.biocon.2006.10.005
Hernandez-Stefanoni J. L., Ponce-Hernandez R. (2006). Mapping the spatial variability of plant diversity in a tropical forest: comparison of spatial interpolation methods. Environ. Monit. Assess. 117, 307–334. doi: 10.1007/s10661-006-0885-z
Hernández-Yáñez H., Kos J. T., Bast M. D., Griggs J. L., Hage P. A., Killian A., et al. (2016). A systematic assessment of threats affecting the rare plants of the United States. Biol. Conserv. 203, 260–267. doi: 10.1016/j.biocon.2016.10.009
Herve M. (2023). RVAideMemoire: Testing and plotting procedures for biostatistics (Version 0.9-83-7) [R package]. CRAN. Available online at: https://cran.r-project.org/package=RVAideMemoire.
Hogan J. L., Brown C. D., Wagner V. (2020). Spatial extent and severity of all-terrain vehicles use on coastal sand dune vegetation. Appl. Vegetation Sci. 24, 1–13. doi: 10.1111/avsc.12549
Homberger J., Lynch A., Riksen M., Limpens J. (2024). Growth response of dune-building grasses to precipitation. Ecohydrology, e2634. doi: 10.1002/eco.2634
Hoover R. H., Gaylord D. R., Cooper C. M. (2018). Dune mobility in the St. Anthony Dune Field, Idaho, USA: Effects of meteorological variables and lag time. Geomorphology 309, 29–37. doi: 10.1016/j.geomorph.2018.02.018
Hughes C. A., Paveglio T. B. (2019). Managing the St Anthony Sand Dunes: Rural resident support for off-road vehicle recreation development. J. Outdoor Recreation Tourism 25, 57–65. doi: 10.1016/j.jort.2018.12.001
Hulbert I. A. R., French J. (2001). The accuracy of GPS for wildlife telemetry and habitat mapping. J. Appl. Ecol. 38, 869–878. doi: 10.1046/j.1365-2664.2001.00624.x
Johnson A. J., Galanti J. M., Englestead D., Whiting J. C. (2024). Migratory patterns and fidelity to wintering areas of Greater Sage-Grouse (Centrocercus urophasianus). Wilson J. Ornithology 136, 7–16. doi: 10.1676/22-00070
Jones A. S., Anderson J. J., Dickson B. G., Boe S., Rubin E. S. (2017). Off-highway vehicle road networks and kit fox space use. J. Wildlife Manage. 81, 230–237. doi: 10.1002/jwmg.21204
Kelly J. F. (2014). Effects of human activities (raking, scraping, off-road vehicles) and natural resource protections on the spatial distribution of beach vegetation and related shoreline features in New Jersey. J. Coast. Conserv. 18, 383–398. doi: 10.1007/s11852-014-0324-1
Knisley C. B. (2011). Anthropogenic disturbances and rare tiger beetle habitats: benefits, risks, and implications for conservation. Terrestrial Arthropod Rev. 4, 41–61. doi: 10.1163/187498311X555706
Knisley C. B., Gowan C., Fenster M. S. (2018). Effects of off-highway vehicles on sandy habitat critical to survival of a rare beetle. Insect Conserv. Diversity 11, 185–193. doi: 10.1111/icad.2018.11.issue-2
Knisley C. B., Kippenhan M., Brzoska D. (2014). Conservation status of United States tiger beetles. Terrestrial Arthropod Rev. 7, 93–145. doi: 10.1163/18749836-07021077
Kutiel P., Eden E., Zhevelev Y. (2000a). Effect of experimental trampling and off-road motorcycle traffic on soil and vegetation of stabilized coastal dunes, Israel. Environ. Conserv. 27, 14–23. doi: 10.1017/S0376892900000035
Kutiel P., Peled Y., Geffen E. (2000b). The effect of removing shrub cover on annual plants and small mammals in a coastal sand dune ecosystem. Biol. Conserv. 94, 235–242. doi: 10.1016/S0006-3207(99)00172-X
Lamb E. G., Guedo D. D. (2012). The distribution, abundance, and environmental affinities of the endemic vascular plant taxa of the Athabasca Sand Dunes of northern Saskatchewan. Écoscience 19, 161–169. doi: 10.2980/19-2-3525
Lee V. R. (1983). Oenothera psammophila (Nels. and Macbr.) Wagner, Stockhouse & Klein ined (the Fremont County: Idaho Sand Dune System).
Locklear J. H. (2017). Endemic plants of the central grassland of north america: distribution, ecology, and conservation status. J. Botanical Res. Institute Texas 11, 193–234. doi: 10.17348/jbrit.v11.i1.1172
Malambo L., Popescu S. C., Horne D. W., Pugh N. A., Rooney W. L. (2019). Automated detection and measurement of individual sorghum panicles using density-based clustering of terrestrial lidar data. ISPRS J. Photogrammetry Remote Sens. 149, 1–13. doi: 10.1016/j.isprsjprs.2018.12.015
Mancuso M., Halford A., Coulson K. (2019). BLM idaho rare plants (Bureau of Land Management). Available online at: https://www.blm.gov/sites/blm.gov/files/uploads/BLM_ID_Rare%20Plants_2019.pdf (Accessed 19 February 2025).
Manley P., Stumpf J., Davis W., Craig D., Mick K. (2004). ‘Pilot test of programmatic approach to monitoring winter conditions and trends of wildlife populations and habitats in off-highway vehicle use areas: First winter pilot test, winter 2002-2003 final report’. U.S. Forest Service. Available online at: https://www.researchgate.net/profile/Patricia-Manley-2/publication/242266355_Pilot_Test_of_Programmatic_Approach_to_Monitoring_Winter_Conditions_and_Trends_of_Wildlife_Populations_and_Habitats_in_Off-Highway_Vehicle_Use_Areas_First_Winter_Pilot_Test_Winter_2002-2003_Final_Repo/links/54b7fd1f0cf269d8cbf629a2/Pilot-Test-of-Programmatic-Approach-to-Monitoring-Winter-Conditions-and-Trends-of-Wildlife-Populations-and-Habitats-in-Off-Highway-Vehicle-Use-Areas-First-Winter-Pilot-Test-Winter-2002-2003-Final-Repo.pdf (Accessed March 26, 2025).
Maun M. A. (1998). Adaptations of plants to burial in coastal sand dunes. Can. J. Bot. 76, 713–738. doi: 10.1139/b98-058
Maun M. A. (2004). “Burial of plants as a selective force in sand dunes,” in Coastal dunes: ecology and conservation. Eds. Martínez M. L., Psuty N. P. (Springer Berlin Heidelberg, Berlin, Heidelberg), 119–135.
McFarling R., Mahnami D. (1996). Monitoring of st. Anthony evening primrose (Oenothera psammophila) - A candidate species using global positioning system (GPS) (Internal records of the Bureau of Land Management: Bureau of Land Management Data Archives).
McGuirk M. T., Kennedy D. M., Konlechner T. (2022). The role of vegetation in incipient dune and foredune development and morphology; A review. J. Coast. Res. 38, 414–428. doi: 10.2112/JCOASTRES-D-21-00021.1
Michaelson V., Johnson L. (1994). Summary of observations from the 1994 survey of St. Anthony Evening Primrose (Idaho Falls: Memorandum to Russ McFarling, District Biologist).
Mills M. H., Schwartz M. W. (2005). Rare plants at the extremes of distribution: broadly and narrowly distributed rare species. Biodiversity Conserv. 14, 1401–1420. doi: 10.1007/s10531-004-9666-6
Moreno-Casasola P. (1988). Patterns of plant species distribution on coastal dunes along the gulf of Mexico. J. Biogeography 15, 787–806. doi: 10.2307/2845340
NASA Earth Observatory (2010). St. Anthony sand dunes, idaho. Available online at: https://earthobservatory.nasa.gov/images/46100/st-anthony-sand-dunes-idaho (Accessed 2025, February 19).
NatureServe (2025). Oenothera psammophila. Available online at: https://explorer.natureserve.org/Taxon/ELEMENT_GLOBAL.2.138325/Oenothera_psammophila (Accessed 19 February 2025).
NOAA (2025). Climate data for pocatello, idaho (Accessed 2025, February 19). National Weather Service.
Patsis A., Overson R. P., Skogen K. A., Wickett N. J., Johnson M. G., Wagner W. L., et al. (2021). Elucidating the evolutionary history of oenothera sect. Pachylophus (Onagraceae): A phylogenomic approach. Systematic Bot. 46, 799–811. doi: 10.1600/036364421X16312067913471
Pavlik B. M., Barbour M. G. (1988). Demographic monitoring of endemic sand dune plants, eureka valley, california. Biol. Conserv. 46, 217–242. doi: 10.1016/0006-3207(88)90069-9
Priya R., Ramesh D., Khosla E. (2020). Biodegradation of pesticides using density-based clustering on cotton crop affected by Xanthomonas malvacearum. Environment Dev. Sustainability 22, 1353–1369. doi: 10.1007/s10668-018-0251-7
R Core Team (2023). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing).
Rice K. (1995). 1995 monitoring of the st. Anthony sand dunes evening primrose (Oenothera psammophila). (Internal records of the Bureau of Land Management: Bureau of Land Management).
Rich J., Rittenour T. M., Nelson M. S., Owen J. (2015). OSL chronology of middle to late Holocene aeolian activity in the St. Anthony dune field, southeastern Idaho, USA. (USA: Quaternary International) 362, 77–86. doi: 10.1016/j.quaint.2014.09.071
Thompson L. M. C., Schlacher T. A. (2008). Physical damage to coastal dunes and ecological impacts caused by vehicle tracks associated with beach camping on sandy shores: a case study from Fraser Island, Australia. J. Coast. Conserv. 12, 67–82. doi: 10.1007/s11852-008-0032-9
Tilman D., Lehman C. (2001). Human-caused environmental change: impacts on plant diversity and evolution. Proc. Natl. Acad. Sci. United States America 98, 5433–5440. doi: 10.1073/pnas.091093198
Trip N. (2014). A comparison of all-terrain vehicle (ATV) trail impacts in boreal forest, heath and bog habitats within the avalon wilderness reserve and surrounding area. Master's thesis (St. John’s, Newfoundland and Labrador: Memorial University of Newfoundland). Available online at: https://research.library.mun.ca/6320/1/van_Vierssen_Trip_Nyssa_MacAllister_0514_MSc.pdf (Accessed March 26, 2025).
USFWS (1996). Endangered and threatened wildlife and plants; review of plant and animal taxa that are candidates for listing as endangered or threatened species. Federal Register 61 (40), 6472–6482. Available online at: https://www.fws.gov/sites/default/files/federal_register_document/96-4412.pdf.
Van der Biest K., De Nocker L., Provoost S., Boerema A., Staes J., Meire P. (2017). Dune dynamics safeguard ecosystem services. Ocean Coast. Manage. 149, 148–158. doi: 10.1016/j.ocecoaman.2017.10.005
Wagner W. L. (2005). Systematics of oenothera sections contortae, eremia, and ravenia (Onagraceae). Systematic Bot. 30, 332–356. doi: 10.1600/0363644054223558
Wagoner N. M. (2006). Saint Anthony Sand Dunes special recreation management area: An exploration of conflict, motorized recreation groups, and community-management relations (Logan, Utah: Utah State University). Available online at: https://ci.nii.ac.jp/ncid/BA8456106X (Accessed March 26, 2025).
Keywords: endemism, sand dune ecology, population clustering, historic comparison, OHV recreation
Citation: Housley AJ, Baird GI, Burchfield DR, Petersen SL and Whiting JC (2025) Changes in the distribution and number of St. Anthony Evening Primrose between 1994 and 2024. Front. Conserv. Sci. 6:1522727. doi: 10.3389/fcosc.2025.1522727
Received: 04 November 2024; Accepted: 21 March 2025;
Published: 11 April 2025.
Edited by:
Daniel Sánchez-Mata, Complutense University of Madrid, SpainReviewed by:
Thomas Edward Marler, University of Guam, GuamCopyright © 2025 Housley, Baird, Burchfield, Petersen and Whiting. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Austin J. Housley, aG91c2xleWFAYnl1LmVkdQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.