
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Conserv. Sci. , 31 January 2025
Sec. Conservation Social Sciences
Volume 5 - 2024 | https://doi.org/10.3389/fcosc.2024.1488378
This article is part of the Research Topic Preventing Zoonoses. Promoting Biophilia. View all 8 articles
 Ma. Fernanda Sánchez-Soto1
Ma. Fernanda Sánchez-Soto1 Osiris Gaona1
Osiris Gaona1 Ricardo Mercado-Juárez1,2
Ricardo Mercado-Juárez1,2 Alfredo Yanez-Montalvo1
Alfredo Yanez-Montalvo1 Arit de León-Lorenzana1
Arit de León-Lorenzana1 Gabriela Borja-Martínez2,3Daniela Zaldívar4Stephany Rodríguez-González2Erika N. Hernández-Villegas5
Gabriela Borja-Martínez2,3Daniela Zaldívar4Stephany Rodríguez-González2Erika N. Hernández-Villegas5 Andres Moreira-Soto6
Andres Moreira-Soto6 Jan Felix Drexler6
Jan Felix Drexler6 Gerardo Suzán7Ella Vázquez-Domínguez3
Gerardo Suzán7Ella Vázquez-Domínguez3 Luisa I. Falcón1*
Luisa I. Falcón1*The Yucatán Peninsula (YP) is home to approximately 60 bat species with differing feeding strategists that, collectively, are fundamental for the health of tropical forests. During the SARS-CoV-2 pandemic, biophobic (aversive) responses towards bats were recorded in urban and rural areas of the YP, making evident the need to monitor bat diversity, investigate species’ biology (e.g., microbiome) and, perhaps most importantly, conduct educational activities that foster an affinity for bats. We designed a multi-scale effort to characterize bat populations and their gut microbiome in urban (Mérida), agricultural (Tizimín), and conserved (Calakmul) landscapes of the YP, while conducting outreach activities to promote biophilic responses. In general, children showed positive responses toward bats, recognizing that they are important parts of their environment. A total of 308 bats from 18 species were sampled; frugivorous species were the most abundant guild (93%), represented mainly by Artibeus jamaicensis and Artibeus lituratus, followed by insectivorous species. Conserved and agricultural landscapes harbored 11 and 16 bat species respectively, both with higher diversity than the suburban areas (eight species). Findings suggest that land use differences in the YP are a factor affecting bat diversity, as well as bat microbiome diversity. Gut microbiome was mainly composed of Pseudomonadota, Bacillota and Actinobacterota (>90%), and abundant bacterial families included Enterobacteriaceae, Staphylococcaceae and Streptococcaceae. Our results have set the baseline for zoonotic disease screening and prevention in the YP, highlighting the importance of coexistence with bats given their key role in maintaining the health of ecosystems.
Bats are keystone species in tropical forests. They provide numerous ecosystem services beneficial to human well-being, such as seed dispersal, pollination, and insect control (Hougner et al., 2006; Kasso and Balakrishnan, 2013; Gannon et al., 2016; Ramirez-Francel et al., 2022). Nonetheless, effectively communicating these benefits to the human communities where bats are distributed, including policy makers and other stakeholders, remains challenging. Misconceptions and the lack of knowledge about bats have likely led to an underestimation of the ecological roles of bats and their significance to human lives and livelihoods (Kasso and Balakrishnan, 2013; Medellin et al., 2017; Ramirez-Francel et al., 2022).
In México, around 142 species of bats have been reported, approximately 60 of which inhabit the Yucatán Peninsula (YP) (Sosa-Escalante et al., 2013; Gaona et al., 2024). For the Maya people, who have dwelled in this region for the past 4,000 years, bats represent an important symbol as messengers of the dead. Camazotz, the vampire bat god, represents death and annihilates dying men on their way to the Underworld (Sieradzki and Mikkola, 2022). Cultural beliefs that associate bats with evil spirits, largely because of the animals’ nocturnal nature, foster fear, explicitly ‘chiroptophobia’ (Sieradzki and Mikkola, 2022). Further exacerbating this fear, some bat species are natural reservoirs of viruses that can cause infectious diseases in humans, including Marburg, Nipah, Hendra, Ebola, influenza, rabies, Middle East (MERS) and severe acute respiratory syndrome coronaviruses (SARS-CoV-2) (Donaldson et al., 2010; Letko et al., 2020). Fear of bats and the misunderstanding of their ecological importance increased during and after the COVID-19 pandemic (Lu et al., 2021). Chiroptophobia induces biophobic (biological aversive) responses that challenge human coexistence with bats, hindering efforts to conserve their diversity and habitat globally (Sieradzki and Mikkola, 2022), consequently threatening the services bats provide.
Pathogens (disease-causing microbes) have also been identified in a diversity of wild animals (Donaldson et al., 2010; Bai et al., 2015; Bolatti et al., 2022). The natural transmission of zoonotic pathogens (those passed from fauna to humans) is called spillover (Jones et al., 2009; Saba-Villarroel et al., 2023). The rising number and magnitude of the emergence of zoonotic diseases are closely related to the ongoing socioeconomic and ecological changes associated with landscape modification, which forces wildlife into closer contact with humans and domestic animals, thereby facilitating pathogen (Jones et al., 2009; Federici et al., 2022; Sánchez-Soto et al., 2024). Urbanization and agriculture intensification, driven by human growth and affluence, are undoubtedly one of the main factors increasing the risk of zoonotic diseases (Jones et al., 2013). Disease-causing bacteria (e.g. Leptospira, Rickettsia, Bartonella, Staphylococcus, Anaplasma) have been detected in some bat species (Stuckey et al., 2017; Lugo-Caballero et al., 2021; Torres-Castro et al., 2020 and Torres-Castro et al., 2021; Federici et al., 2022) as common components of their microbiome (Federici et al., 2022). A growing body of research has shown that the microbiome responds to land‐use and environmental changes, particularly where such changes lead to altered food resources (Amato et al., 2013; Barelli et al., 2015). Considering the zoonotic pathogens found in the microbiome of wild animals, recent studies have emphasized the importance of preserving their habitat from anthropogenic activities to minimize the spillover of potential infectious diseases (Federici et al., 2022).
The YP in México, like many other tropical regions, has a long history of landscape modification by human activities, which have failed to appropriately manage tropical forests and their biodiversity (García-Frapolli et al., 2007). Such land modification started during the development of the Mayan civilization (2,000 BCE–), followed by the prolific henequen production of the 19th century, until the current accelerated urbanization and expansion of economic activities in the region (Gómez-Pompa and Kaus, 1999; González-Iturbe et al., 2002). Specifically, regarding the three states that constitute the YP in México (Figure 1), urbanization and tourism mainly affect the coasts of Quintana Roo, while agriculture and livestock dominate the deforestation trends in Yucatán and Campeche, as a result of recent agricultural intensification policies (Ellis et al., 2017; Špirić et al., 2022). The YP loses over 280,000 ha of tropical forests every year (CCMSS, México 2024), thus under the current social and economic development model, biodiversity and ecosystem health will continue to be threatened (García-Frapolli et al., 2007).

Figure 1. Sampling sites in the Yucatán Peninsula, México. Urban, agricultural and conserved landscapes according to the predominant land use correspond to Mérida, Tizimín and Calakmul municipalities. Sampling site shapes were generated with a buffer of 30 km that correspond to the average dispersal of sampled bat species. Source: Map modified with permission from INEGI, México (2020) land use and vegetation map, 2021.
Information regarding biodiversity, especially bat ecology and the role of bats in providing essential ecosystem services, can enhance opportunities to promote biophilia (biological affinity), particularly within the communities where bats are distributed. This can in turn aid in the development of conservation strategies and, concurrently, in actions that favor insect control. By promoting a biophilic perspective that fosters human-wildlife coexistence, it is possible to build a future where people and nature thrive together (Hougner et al., 2006; Tidball, 2012). Working with local communities is fundamental for conservation of wildlife and natural habitats. We became particularly conscious during the CoVID-19 pandemic that negative perceptions were growing towards bats. To promote bat conservation, we thus paired field work with a pilot program to promote biophilic responses to bats through outreach activities, giving educational talks in primary schools in the localities where we did sampling. Public outreach and the dissemination of scientific knowledge are critical to the promotion of conservation and the implementation of strategies that build towards a sustainable future. To this end, the project objectives we report on in this paper were to 1) perform educational intervention intended to cultivate positive feelings toward bats in school-aged children (5-11 years old); 2) assess the perception of bats in the children who participated in this pilot program; and at the same time 3) analyze bat diversity and microbiome across three landscapes (urban, agricultural and conserved) to produce locally relevant data for integration into further education efforts. Our analyses included screening for bacterial taxa and Betacoronavirus (β-CoV) that may pose potential threats to public health.
The YP in southeastern México is characterized by a tropical warm and subhumid climate with an average annual temperature of 22°C-26°C and 800-1500 mm of rainfall (Vidal-Zepeda, 1990). The flat and low land relief is in the range of 0-250 m above sea level with a bedrock dominated by limestone (García-Rejon et al., 2008). Native vegetation in the most southern region is composed of tropical and subtropical evergreen high rainforests, characterized by a highly diverse floristic composition. Towards Campeche, Quintana Roo, and the south of the state of Yucatán, the vegetation is dominated by medium forest, semi-evergreen and sub deciduous forest. The northwestern region, including a large area of Yucatán, a small extension of Campeche, and dispersed along the Quintana Roo coast, is mostly covered by low deciduous and sub deciduous forests. The rocky and shallow soils of low forest result in a floristically and structurally simpler composition (Islebe et al., 2015).
The three study regions encompassed urban, agricultural and conserved landscapes (Figure 1). The urban region was located in the city of Mérida (population ∼1,008,000), including the suburban communities of Ucú and Caucel. The agricultural region (ecotone) was located in Tizimín (population ∼80,672) and the conserved region was located in the Calakmul Biosphere Reserve (population ∼31,714), which harbors the largest continuous tropical forest cover in the Americas, after the Amazon (INEGI, México 2020).
The SARS-CoV-2 pandemic evidenced the need to communicate the importance of bats globally because it was apparent that some people thought bats where to blame (Burki, 2020). As part of this effort, we worked with children (5-11 years old) in the localities of Mérida, Tizimín and Calakmul (Figure 1) during our bat sampling field work. This approach included informative talks in local elementary and preschools using teaching and game materials (i.e. coloring masks/mandalas, paper sheets, crayons, game boards, infographic material showing myths and truths of bats). We displayed a mist net to show how field work is performed. Talks focused on bat diversity, morphological attributes, behavior, feeding habits, common beliefs, and the environmental services bats provide. Importantly, information given during the talks, as well as the games and teaching materials, were prepared and presented in accordance to the school level of the children in each school (see Table 1 for each school region, number of children participating and school level, and activities). After the talks, a basic questionnaire was distributed to all the children to assess the children’s knowledge and perception towards bats (Supplementary Material S1). We highlighted the relevance of bats for humans’ lives and provided basic recommendations (e.g., not handling bats and avoiding disturbing the animals and their shelters by any means). Respect for wildlife was instilled and we used inclusive language with respect for cultural and gender diversities. The complete bat perception and environmental education strategy is described in Supplementary Material S2. Schools approved, permitted, and supervised the talks, the general perception survey, and the teaching and game materials used. Additionally, we conducted a review of news articles and reports on biophilic or biophobic actions towards bats in the YP, before, during, and after the SARS-CoV-2 pandemic. The publication source, date, location, bat species involved, action on bats, and the motives were recorded (see Supplementary Material S3).
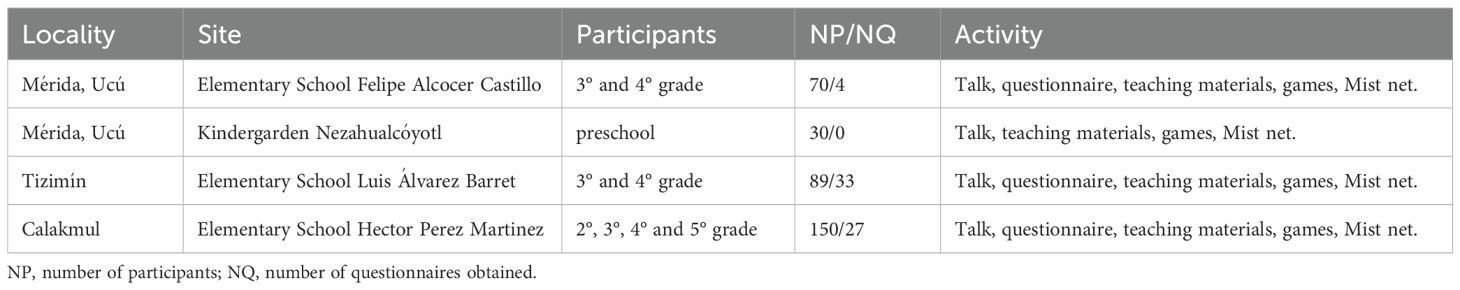
Table 1. Local rural communities where bat perception was surveyed in the Yucatán Peninsula.
Bats were sampled following the guidelines of the American Society of Mammalogists for capture, handling, and welfare for wild mammals (Gannon and Sikes, 2007; Gardner, 1979). We set three mist nets during seven consecutive nights (19:00-01:00h), in each landscape during the dry (November 2021 and March 2022) and rainy (May 2022) seasons. Bats were placed in sterile plastic bags with ventilation until they defecated. Fecal samples were collected using sterile gloves and placed in empty and sterilized Eppendorf tubes (1.5 ml). All samples were frozen in liquid nitrogen until processing in the laboratory. Anal and oral swabs were taken using sterile applicators, stored in cryogenic tubes with RNA later (500µls) and preserved in liquid nitrogen. Standard measurements such as body and forearm length, weight, gender, age (adult or young by checking wing bone ossification), and females as lactating or pregnant, were recorded for all captured bats (Anthony, 1988; Jones et al., 1996). All sampling was done under collector permit SGPA/DGVS/07572/2.
DNA extractions from fecal samples were performed with the QIAamp PowerFecal Kit (Qiagen, Valencia, CA) following the manufacturer’s instructions. To increase DNA yield we concentrated DNA with a 3M sodium acetate (10%) precipitation resuspended in 30 µL of molecular grade water and stored at -20°C until PCR amplification. Additionally, total nucleic acids were extracted from anal and oral swabs for viral analysis following the MagNA Pure DNA and Viral NA small volume protocol using a MagNAPure DNA instrument (Roche) at the Institute of Virology, Charité, Universitätsmedizin Berlin.
Fecal DNA samples were PCR amplified with universal bacterial/archaeal primers 515F/806R (16S rRNA hypervariable region V4) following the procedures reported by Apprill et al. (2015). PCR reactions (25 µl) contained 2-6 ng of total DNA, 2.5 µl Takara ExTaq PCR buffer 10X, 2 µl Takara dNTP mix (2.5 mM), 0.7 µl bovine serum albumin (BSA, 20 mg ml-1), 1 µl primers (10 μM), 0.125 µl Takara Ex Taq DNA Polymerase (5 U μl-1; TaKaRa, Shiga, Japan) and nuclease free-water. Samples were amplified in triplicate using a PCR protocol including an initial denaturalization step at 95°C (3 min), followed by 35 cycles of 95°C (30 s), 52°C (40 s) and 72°C (90 s), adding a 12 min final extension at 72°C. Triplicates were pooled and purified using the SPRI magnetic bead, AgencourtAMPure XP PCR purification protocol (Beckman Coulter, Brea, CA, USA). The purified 16S rRNA fragments (~20 ng per sample) were sequenced on an Illumina MiSeq platform (Yale Center for Genome Analysis, CT, USA), generating ~250 bp paired end reads. The sequence data are available in the NCBI BioProject PRJNA1153560.
Anal and oral nucleic acid samples were analyzed for β-CoV following a PCR approach. β-CoV RNA was amplified with a hemi nested PCR method (Annan et al., 2013) using primers Pan2cRdRP-FWD and Reverse (10 µM). First round reactions had 12.5µls final volume, BSA (1 mg/ml), MgSO4 (50mM) with an initial reverse transcription of 50°C;C for 20 min, followed by an initial denaturation of 95°C; 3 min and 20X of 94°C; 15 sec, 60°C; 15 sec and 72°C; 30 sec; 30X 95°C 15sec, 50°C; 15 sec, 72°C; 30 sec with a touchdown of -0.5°C;C per cycle. The second round PCR included 1µl of the first-round product and primers pan2cRdRp-FWD and Rnest (10µM) with the same amplification protocol in 25 µl volume, MgCl2 (50mM), dNTP (10mM each). All PCR reactions were done with the SuperScript™ III One-Step RT-PCR System with Platinum™ Taq High Fidelity DNA Polymerase (Invitrogen). Amplicons (398-bp) were verified with agarose gel-electrophoresis and Sanger sequencing, but no further analysis was performed since no positive sequences were detected.
We used fastp 0.23.2 (Chen et al., 2018) for quality control of the sequencing reads in fastq format and QIIME2 qiime2-amplicon-2023.9 (Bolyen et al., 2019) for quality processing of sequences. Sequence data were denoised with DADA2 and clustered by amplicon sequence variants (ASVs) (Callahan et al., 2016) with the q2‐dada2 plugin. All ASVs were aligned with MAFF (Katoh et al., 2002) with the complement q2‐alignment, which were used to construct a phylogenetic tree with fasttree2 (Price et al., 2010) and the q2‐phylogeny plugin. Taxonomy was assigned to ASVs using the SILVA 138 database (Quast et al., 2013). Finally, we removed all sequences classified as Eukaryote or unclassified at the phylum level in the R programming environment (R Core Team, 2024) and filC;tered the samples between the 25th and 75th quartiles based on the frequency and type of feeding.
We used the ‘tidyverse’ package (Wickham et al., 2019) in R for data manipulation and visualization, ‘qiime2R’ (Bisanz, 2018) for integration of results coming from QIIME 2, ‘phyloseq’ (McMurdie and Holmes, 2013) for analysis of amplicon sequencing data and ‘microbiome’ (Lahti and Shetty, 2019) to perform biodiversity-specific analyses from a subsampling of 200,000 sequences per sample. Beta-diversity was tested on Unifrac metrics through the PERMANOVA model implemented in the ‘adonis’ function of the vegan package in R. Graphics for data analysis were elaborated in ‘ggplot2’ (Wickham, 2009).
Sequence identification and analysis of pathogenic bacteria followed a meticulous process. We aimed to identify ASVs related at the family level to the most abundant bacterial families with known pathogens, which in this study were represented by Clostridiaceae, Mycoplasmataceae, Staphylococcaceae and Streptococcaceae, and at the order level to Enterobacterales, Rhizobiales, Rickettsiales and Xanthomonadales. We conducted restrictive blast searches (Altschul et al., 1990) against a specialized pathogen database (Yang et al., 2023) to identify sequences similar to organisms of epidemiological interest. The sequences with the best hits were extracted for further analysis. These selected sequences were then clustered with cd-hit (Li and Godzik, 2006), aligned from the V4 16S rRNA region with MAFFT, and phylogenies were constructed using FastTree. We used FigTree software (http://tree.bio.ed.ac.uk/software/figtree/) to visually represent the phylogenetic relationships between sequences. Finally, an analysis of each ASV that clustered to a known zoonotic bacterial agent was performed for each bat species per study area.
The number of children participating in the talks and in the drawing and gaming activities in each school are indicated in Table 1 (see Supplementary Material S1 for detailed information). We provided the questionnaire to all the children and obtained responses from 73; the total number of drawings was 94 (data not shown). The survey aimed to explore the children’s knowledge and positive or negative feelings toward bats after the talks we presented; we found that most (96%, 70/73) are familiar with bats and knowledgeable of their feeding strategies and habits (Supplementary Material S1). Interestingly, all children answering the questionnaire from Calakmul, the most conserved landscape, had a 100% positive perception of bats. Most children (78%) from agricultural Tizimín considered bats to be beneficial for ecosystems. Drawings showed a positive trend of the children’s perception and feelings towards bats after the educational intervention (Figure 2).
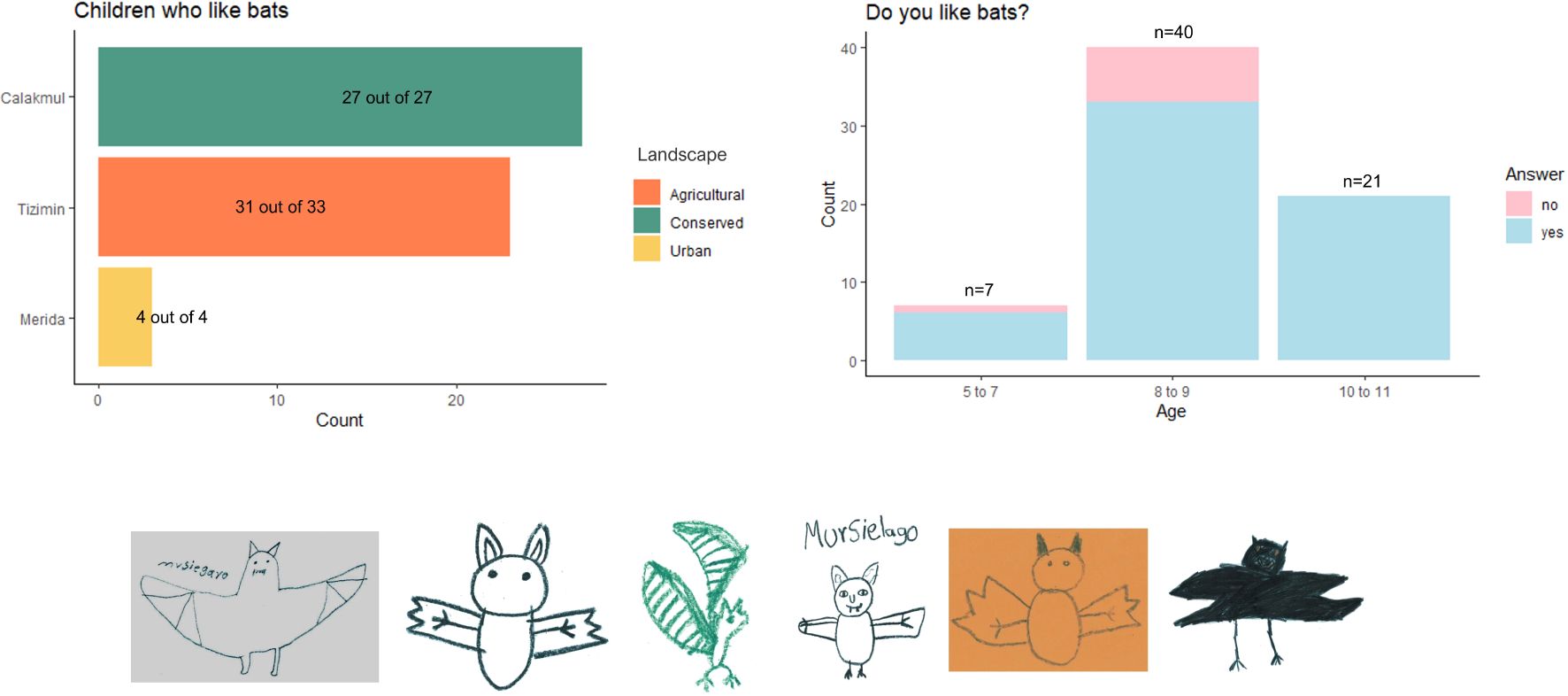
Figure 2. Surveys on children’s perception toward bats. Results based on a total of 73 questionnaires obtained. Number of children who like bats per landscape (top left); Bat perception provided by scholar children from different ages (top right); Some examples from 94 drawings performed by the children surveyed (bottom).
Our review of news articles and reports on biophilic actions toward bats in the YP revealed that few biophobic incidents have been recorded in the YP, with only 10 news reports since 2006 (Supplementary Material S3). During the SARS-CoV-2 pandemic, three incidents were reported of direct attacks from people to bat colonies, whereas previous to the pandemic, four reports occurred related to rabies (SENASICA, México, 2024), and the rest had no direct cause.
A total of 308 bats representing three families (Phyllostomidae, Vespertilionidae, and Mormoopidae), and 18 species were sampled, 42.2% of which (130/308) were found in the most conserved landscape (Calakmul), 31.5% (97/308) in the agricultural (Tizimín) and 26.3% (81/308) in the urbanized landscapes (Mérida suburbs) (Figure 3). The number of bat species and diversity (Shannon diversity index) were similar in all localities (Figure 3; Supplementary Material S4). A total of six bats species, four frugivorous Artibeus jamaicensis, A. lituratus, Glossophaga soricina and Sturnira parvidens and two insectivorous Pteronotus mesoamericanus and Rhogeessa aeneus, were observed in the three landscapes (conserved, agricultural and urban). Frugivorous strategists accounted for 93% of all observations (Figure 3), while the insectivorous strategists accounted for only 5%. Fruit eaters were mostly represented by A. jamaicensis, the dominant bat species (35%, 109/308) in all the sampling settings. Other frugivorous bats including A. lituratus (18%, 55/308), G. soricina (13%, 48/308), Carollia sowelli (10%, 32/308), Dermanura phaeotis (7.5%, 23/308) and S. parvidens (7.5%, 17/308) were relatively abundant in different landscapes (Figure 3). A. lituratus was more abundant in the agricultural and conserved landscapes, C. sowelli and D. phaeotis were better represented in conserved areas, while G. soricina showed higher abundances in the urbanized and S. parvidens in the agricultural landscapes. The insectivorous bats (15 individuals) were mostly observed in the agricultural landscape. Carnivorous and hematophagous bats included one species each, observed in agricultural and in the conserved landscapes respectively (Figure 3).
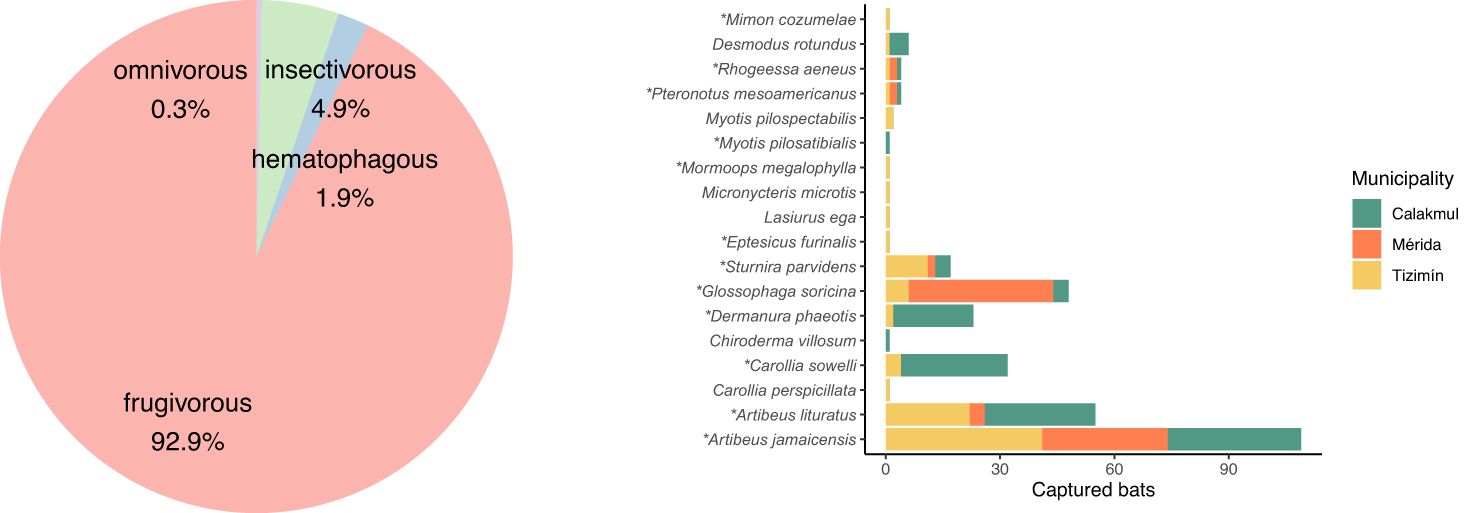
Figure 3. Identified bat species and associated feeding strategies. Proportion of observed bats per feeding strategy (left). Number of bats per species among conserved, agricultural and urban landscapes, which respectively correspond to Calakmul, Tizimín and Mérida (right). Bracket colors according to the feeding strategists indicated in the pie chart. Asterisks show bat species assessed in the gut microbiome analysis.
A total of 108,656,360 paired end raw sequences from fecal microbiome amplifications were obtained. After applying quality-control filters 94,519,756 (86.98%) of paired end sequences and 75,409,547 (69.40%) of non-chimeric denoising sequences remained. In total, we recovered 34,835 ASVs from 77 fecal samples and 12 bat species (Figure 3). The sampling effort was sufficient to assess the microbial diversity present in the fecal samples for most bat species (Supplementary Material S5) suggesting a similar composition according to the beta-diversity analysis (Figure 4; Supplementary Material S6). Alpha-diversity indexes of the fecal-associated microbiome was similar among different bat species at the landscape scale (Supplementary Materials S7, S8), and higher diversity was found in the agricultural landscape (Supplementary Material S8). No statistically different structure of the A. jamaicensis fecal microbiome composition was observed between agricultural and urban landscapes, which significantly differ from the conserved area (Supplementary Material S9).
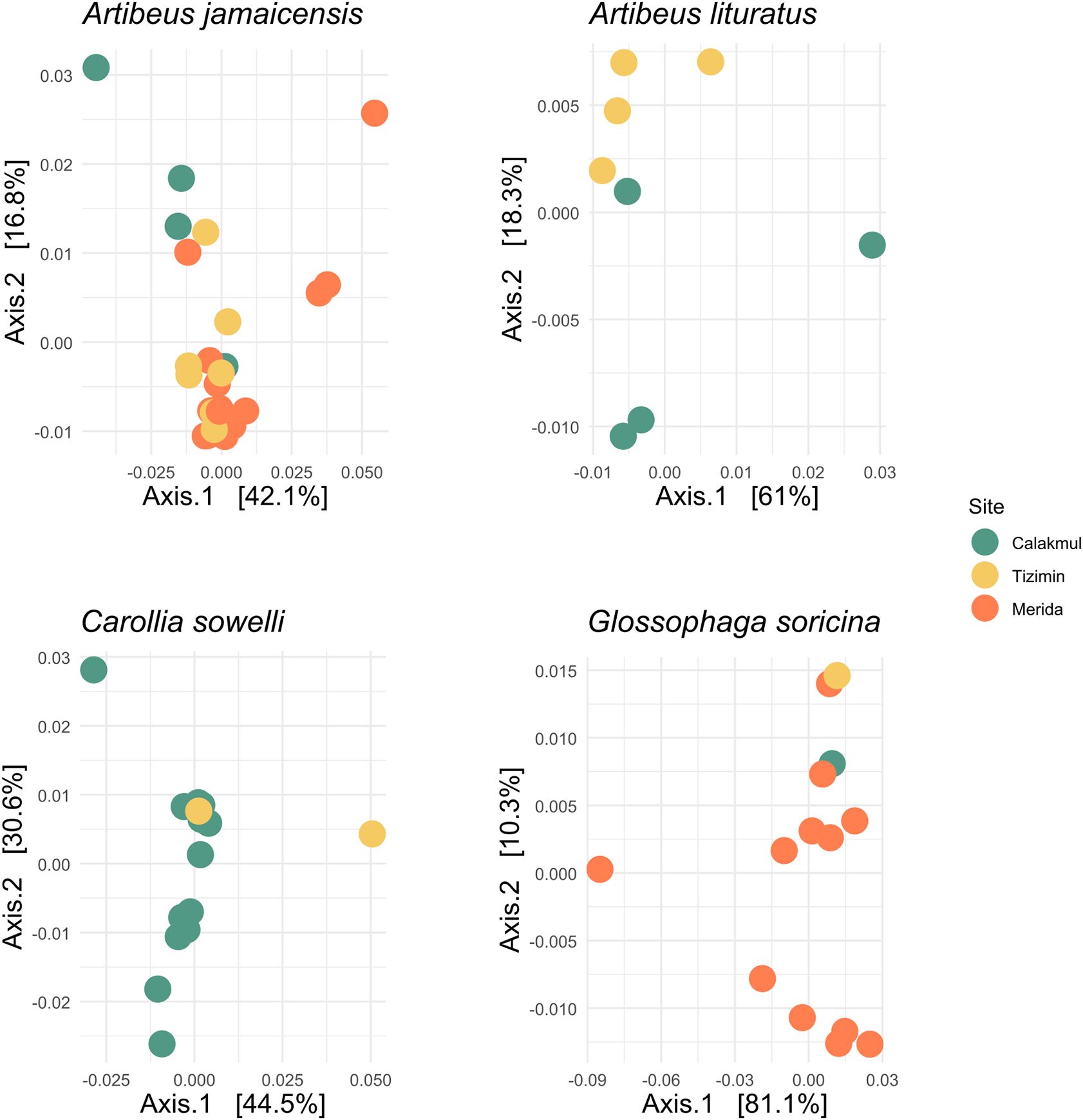
Figure 4. Gut microbiome community structure analysis for the most abundant bat species. PCoA ordination method performed with the weighted Unifrac distance matrix from bacterial 16S rRNA gene sequences. Sites correspond to conserved (Calakmul), agricultural (Tizimín) and urban (Mérida) landscapes.
Each bat species analyzed harbors a distinct fecal microbiome which is related to their feeding strategies and landscapes. The predominant bacterial phyla included Pseudomonadota, Bacillota, Actinomycetota, Bacteoridota, Desulfobacterota, Chloroflexota, Planctomycetota, Myxococcota and Verrucomicota (Figure 5). Transient chloroplasts were more abundant in frugivorous bats from the conserved Calakmul landscape (Figure 5).
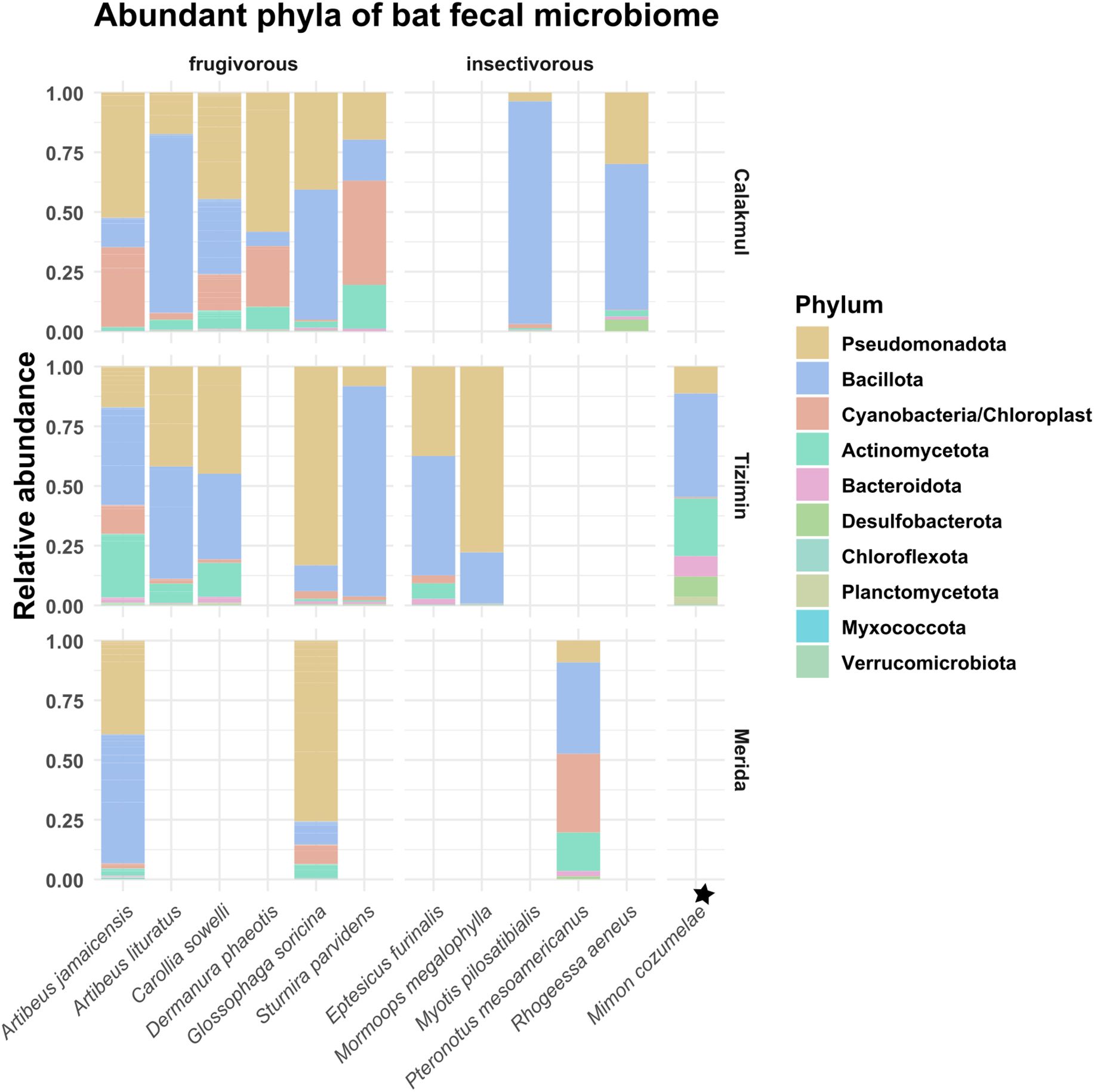
Figure 5. Gut microbiome composition. Relative abundances of the top 10 bacterial phyla for the frugivorous, insectivorous and carnivorous bats sampled in the conserved (Calakmul), agricultural (Tizimín) and urban (Mérida) landscapes. Bat species with only one individual is indicated with the symbol ★.
A few sequences were related to putative pathogenic bacterial families (Supplementary Material S10), which had different relative abundances in bat species (Figure 6). Overall, Enterobacteriaceae, Streptococcaceae, Staphylococcacea and Erwiniaceae formed a large proportion of the fecal microbiome (Figure 6). It is important to note that only the fecal microbiome of A. jamaicensis and G. soricina could be assessed across the three landscapes. In A. jamaicensis, Enterobacteriaceae and Staphylococcacea were abundant in the conserved landscape (Calakmul), while Streptococcaceae abundance increased in the agricultural landscape (Tizimín), and Erwiniaceae in the urban landscape (Mérida). In G. soricina, Enterobacteriaceae clearly dominated the fecal microbiome in all landscapes (Figure 6). Regarding viral pathogens, the focused surveillance of β-CoVs from anal and oral swabs of bats performed following a hemi nested PCR approach, showed no positive results.
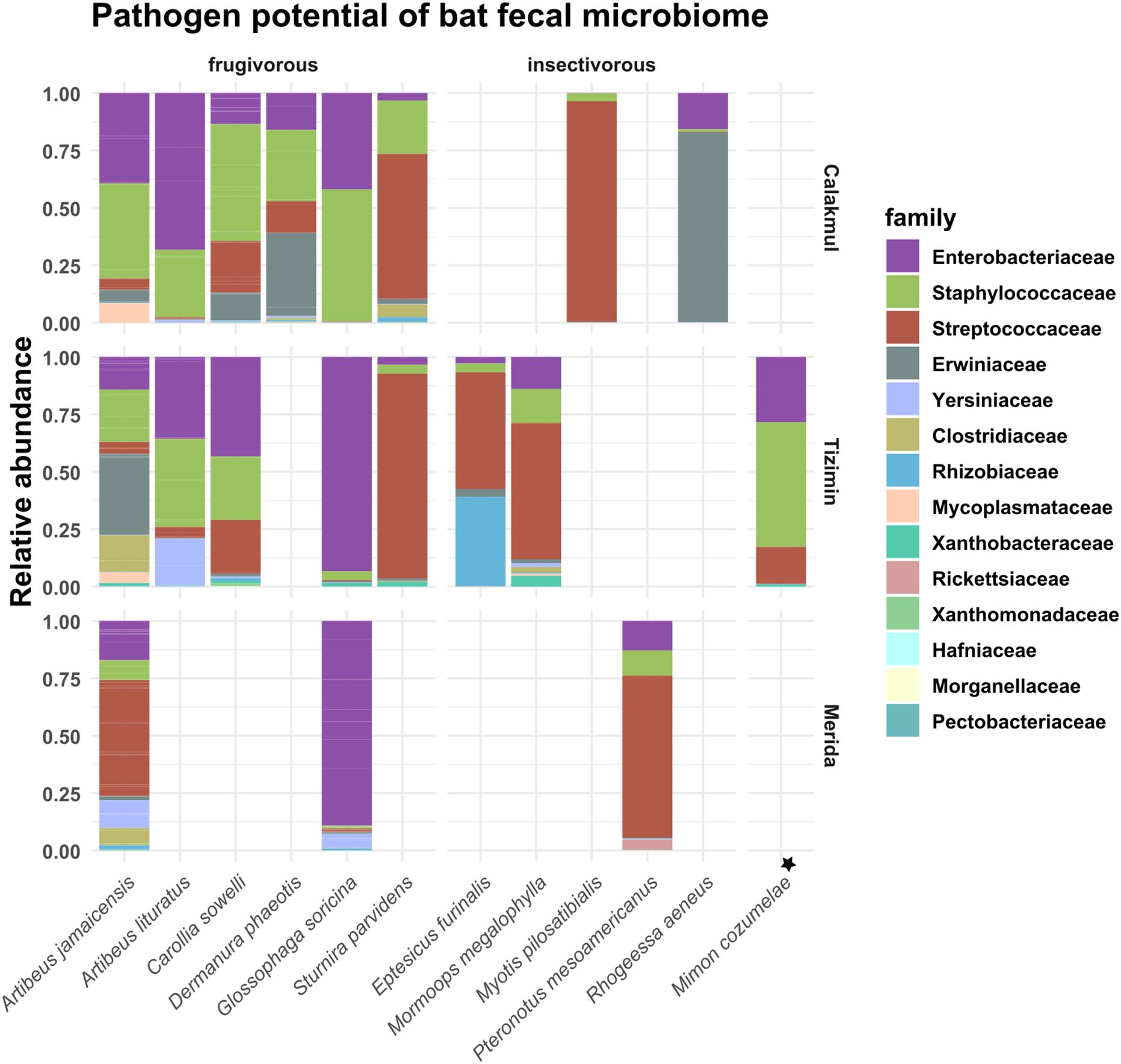
Figure 6. Putative pathogenic bacterial families. Relative abundances of bacterial families of epidemiological interest based on 16S rRNA gene sequences compared against specialized pathogen databases. Bat species with only one individual is indicated with the symbol ★.
This study of bats is framed within the basic principles of One Health which recognize the interconnection among humans, animals, and ecosystems (Mackenzie et al., 2014). We aimed to address the coexistence between humans and bats in the Yucatán Peninsula (YP), by combining an educational intervention with ecological and microbiome characterizations. This survey on the biophilic or biophobic actions towards bats, in light of the perception generated after the global pandemic of SARS-CoV-2, provides a basic understanding of local traditions and people’s relationship with the natural environment in YP regions (Figure 2). Importantly, because the localities we studied include Mayan populations, our approach and the educational pilot study performed were designed to include aspects of their customs and traditions. Interviewed children revealed that there is an appreciation for bats in local communities, which is not that surprising since they live in contact with wildlife on an everyday basis. Moreover, the cosmovision of the Mayan culture, constructed around a deep relationship with the natural world, respects all living and nonliving things, believing they possess a soul (Lucero, 2018). The bat-god ‘Camazotz’ is a Mayan deity which was ancestrally revered and respected (Sieradzki and Mikkola, 2022). The optimistic reception of the informative talks revealed a latent interest in wildlife among the communities in rural settings, confirming the potential to help bridge the gap between humans and nature.
The outreach activities implemented in primary schools promoted positive feelings towards bats. Collaborative participation with the local communities fosters a sense of coexistence with wildlife and of support for appropriate management of ecosystems. School-aged children effectively permeate knowledge to their younger siblings, parents and to other children through their games. This supports the long-term benefits of educating children, who will grow up to become informed adults. As ecologists we observe that educated children foster greater respect towards nature. Although this is a pilot program for Yucatán, similar educational activities have been replicated in other states of México (RELCOM Latinoamérica, 2024). To develop a long-standing project to promote biophilic responses, we propose that subsequent studies should perform questionnaires before and after educational interventions, broadening the target audience to include a similar number of participants from different ages, socioeconomic contexts and educational levels. Adapting the program for Maya speakers is key. It will also be important to incorporate our locally relevant research findings on bat diversity and microbiomes, making the connection to land use scenarios.
Previous studies performed in the peninsula have addressed various aspects concerning bat fauna, including their composition, morphology and seasonality behavior (Arita, 1997; Rydell et al., 2002; Hernández-Dávila et al., 2012). Others have explored bat responses to habitat fragmentation, the ecological role of frugivorous bats and the importance of habitat management in conserving bat assemblages (Montiel et al., 2006; Vilchis et al., 2007; Medellin et al., 2017; Cafaggi et al., 2024). The presence of zoonotic pathogens in certain bat species has begun to be studied (Machain-Williams et al., 2013; Torres-Castro et al., 2020), including our current research. None of these studies involved outreach to promote biophilic actions towards bats. While educational interventions in the region have focused on preventing specific zoonotic diseases (e.g. Chagas, dengue, rickettsiosis), primary through vector control (Valdez-Tah et al., 2015; Chávez-Arias et al., 2017; Dzul-Rosado et al., 2023), there is a lack of initiatives promoting a positive relationship between local communities and bats and other natural insect predators. Recognizing the interconnection between nature and society, conservation biology, and applied ecology emphasize that effective management of nature is both an outcome and a driver for social, economic and ecological changes (Cumming and Allen, 2017). Driven by this understanding, our research team is committed to generating scientific knowledge, while developing and implementing more effective strategies that foster stronger connections between nature and local communities.
Findings suggest that land use differences in the YP are a factor affecting bat diversity and their microbiome composition. Notably, bats are recognized as indicator or sentinel species (Jones et al., 2009; Wolf et al., 2022). During the present study, a higher diversity of bat species was registered in agricultural dominated (Tizimín) and conserved (Calakmul) (Figure 3) than in urban landscapes. The interconnection of natural habitats and the adjoining managed lands, where silvicultural activities are performed, are likely serving as biological corridors, supporting bat dispersal and providing habitat and food availability. This suggests that these corridors contribute to biodiversity conservation allowing connectivity between populations, communities and ecological processes in fragmented landscapes (Laurance, 2004 Bolívar-Cimé et al., 2013). The Calakmul Biosphere Reserve is part of the Mesoamerican Biological Corridor, a region of great biodiversity (Vester et al., 2007). It harbors the largest region of continuous rainforest in the Americas, second only to the Amazon. This reserve still sustains populations of large mammals, including Baird’s Tapir (Tapirus bairdii),jaguar (Panthera onca), and the largest diversity of bats reported (Vester et al., 2007) (Figure 3). Further, across the study sites (Figure 3), we found a higher abundance of opportunistic fruit eating bats, such as A. jamaicensis (Montiel et al., 2006; de Souza and Vizentin-Bugoni, 2020), and a scarcity of insectivores that are more environmentally sensitive to landscape changes (Threlfall et al., 2011). Particularly for the Calakmul reserve, these results might be explained by potential habitat loss due to the increasing establishment, expansion and intensification of agricultural activities (Vester et al., 2007; Špirić et al., 2022). Unfortunately, this process is doomed to continue because Calakmul is further threatened by fragmentation triggered by a controversial train that will run through the heart of this nature reserve (Ortega and Jaber, 2022).
The high abundance of frugivorous bats, especially of A. jamaicensis in urban areas, demonstrates their great adaptability to fairly hostile and resource-limited conditions (Moretto et al., 2023). This adaptability likely relies on diversifying their food sources, which enables them to displace the more selective or dietary specialized species that are not able to persist in such fragmented ecosystems (Meyer et al., 2008; Montiel et al., 2011). Bat species that tolerate urban and suburban habitats in the YP are vital for the sustainability of forests as seed dispersers, contributing to forest regeneration (Hougner et al., 2006; MacSwiney et al., 2017). Furthermore, bat species that are better adapted to human modified landscapes will continue to interact with urban human populations, which are expected to be poorly connected to, and knowledgeable of, natural landscapes (Moretto et al., 2023). Experiences that combine education and immersion in natural landscapes have a positive influence on human behavior (Dzul-Rosado et al., 2023). Thus, it is fundamental to continue with the design and implementation of educational strategies in the region that promote biophilic responses towards bats, while informing the different actors of the key environmental benefits they provide.
Microbiome assessments are essential tools to monitor microbes of zoonotic importance, and this information significantly impacts the perception towards bats and the way people indirectly interact with them in the YP. As part of this ongoing research, we aim to incorporate the microbiome knowledge here generated in future outreach activities to continue promoting bat conservation and public health. The low abundance of insectivorous bats observed in this survey suggests a potential loss of biological insect control. Molecular identification of prey remains in insectivorous bat feces has revealed the consumption of insects, including mosquitoes that can transmit diseases of medical importance (Burgar et al., 2021). Given the prevalence of mosquito-borne diseases in the YP, such as dengue, Zika and West Nile Virus (Sánchez-Soto et al., 2024), insectivorous bats can perform a role in controlling populations of these disease-vector insects.
Importantly, food resources influence the host microbiome. Hence, microbiome data offers valuable insights into the potential ecological consequences of land-use changes on bat populations. Our results show differences between the gut microbiome of the frugivorous A. jamaicencis from conserved areas and that from suburban and agricultural settings (Supplementary Material S9), likely as a result of differences in food resource quality, abundance and diversity, and/or by exposure to novel bacteria (Carrillo-Araujo et al., 2015; Ingala et al., 2021). This study can establish a reference of the diversity and composition of the gut microbiome in contrasting landscapes of the YP. It can serve as a basis for future studies to evaluate the effects of habitat modification on bat health and fitness, and the relation between habitat modification and potential pathogens spillover and transmission (Ingala et al., 2018). In this regard, an important research venue would be to survey bacteria of clinical interest in bat microbiomes (Cláudio et al., 2018), to perform zoonotic risk analysis to address effective control measures in priority areas with presence of specific pathogens.
Nonetheless, it is important to emphasize that the presence of opportunistic potential pathogens in the gut microbiome (Figure 6) does not indicate that bats in the YP are reservoir hosts for the transmission of bacterial zoonotic diseases (Wolkers-Rooijackers et al., 2018). Results show that the individual bats analyzed did not host β-CoV. In a recent study, Colunga-Salas and Hernández-Canchola (2020) analyzed sequences for the ORF1ab polyprotein of CoVs detected in 11 bat species distributed in México and confirmed that SARS-CoV-2 was not present. Although these bats did not present a zoonotic disease threat, there is evidence of certain viruses associated with A. jamaicensis, including influenza, dengue, Zika, West Nile and rabies, which could contribute to potential disease spread (Cabrera-Romo et al., 2016; Almeida-Campos et al., 2019; Torres-Castro et al., 2021). Yet, we underline the need for wildlife microbiome surveillance that aids in the assessment of public health risks in the region (Sánchez-Soto et al., 2024). Considering that bats are key species that indicate the health of ecosystems, their gut microbiome will also be representative at the landscape level and could be monitored with relative ease.
Far from generating a negative perception towards bats, addressing public concerns about these flying mammals is essential for bat conservation, since negative attitudes frequently stems from misinformation, fear, and a lack of awareness about the key environmental benefits they provide (Lu et al., 2021; Ramirez-Francel et al., 2022). Current dominant socioeconomic models have ignored the relation between human population growth, habitat modification, and ecosystem health. Our findings support that land use has an impact on bats’ diversity and their microbiome, whereby the diversity of bats decreases in the most urbanized sites and their gut microbiome tends to be more similar in modified landscapes. Bat diversity in the YP was dominated by frugivorous species, suggesting loss of other ecosystem services, such as insect control by bats that can consume large quantities of insects, including those that transmit diseases. The lower the insect predators, the higher the potential of some zoonotic diseases and spillover, such as dengue, Zika and others. Pollinating bats are also essential in the ecosystems; changes in land use and fragmentation affect this bat guild. Clearly, studies that promote biophilia responses in the YP are fundamental; while surveying bat populations and their microbiome dynamics offer an indirect means to understand ecosystem health. A biophilic approach promotes healthy bat populations in hand with sustainable development, while it is in healthy ecosystems that zoonotic diseases will be controlled.
The original contributions presented in the study are publicly available. This data can be found here: NCBI BioProject, accession PRJNA1153560.
Ethical review and approval was not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent to participate in this study was provided by the participants legal guardian/next of kin.
MS-S: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft, Writing – review & editing, Validation. OG: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Visualization, Writing – original draft, Writing – review & editing. RM-J: Data curation, Formal analysis, Methodology, Visualization, Writing – review & editing. AY-M: Methodology, Writing – review & editing. AL-L: Methodology, Writing – review & editing. GB-M: Data curation, Methodology, Writing – review & editing. DZ: Formal analysis, Methodology, Writing – original draft. SR-G: Writing – review & editing, Methodology. EH-V: Writing – review & editing, Methodology. AM-S: Methodology, Writing – review & editing, Funding acquisition, Project administration. JD: Funding acquisition, Resources, Project administration, Writing – review & editing. GS: Conceptualization, Funding acquisition, Project administration, Resources, Writing – review & editing. EV-D: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Resources, Visualization, Writing – review & editing, Project administration, Supervision. LF: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Funding for this work, including postdoctoral scholarships (AY-M and MS-S) was provided by UNAM, PAPIIT project No. IV200421 (EV-D, LF, GS). GLACIER (DAAD-UNAM) funded work in Charité (LIF, SR-G, EH-V), PASPA-UNAM funded work in Charité (LF). The publication of this paper was sponsored through an Interagency Agreement Between the US Fish & Wildlife Service and Smithsonian National Zoo & Conservation Biology Institute. It advances work on risk communication as a component of study directed by the American Rescue Plan Act. Additional in-kind partners in this sponsorship include the International Alliance Against Health Risks in the Wildlife Trade and the International Union for the Conservation of Nature (IUCN).
The authors deeply thank Juan Cruzado for field work and bat identification, Graciela García-Guzmán and Marco Tulio Solano De la Cruz (Instituto de Ecología, UNAM), Arne Kühne (Charité, Berlin), for their help with collector permits and laboratory work, and Alfredo Argaez, Soluciones Ambientales ITZENI, AC for technical assistance. Authors appreciate the reviewers and editor who greatly helped improve this article. Schools approved and permitted the general perception survey and the talks we gave to students about the relevance of bats.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author (LIF) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2024.1488378/full#supplementary-material
Almeida-Campos A. C. A., Góes L. G. B., Moreira-Soto A., de Carvalho C., Ambar G., Sander A. L., et al. (2019). Bat influenza A (HL18NL11) virus in fruit bats, Brazil. Emerg. Infect. Dis. 25, 333. doi: 10.3201/eid2502.181246
Altschul S. F., Gish W., Miller W., Myers E. W., Lipman D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/s0022-2836(05)80360-2
Amato K. R., Yeoman C. J., Kent A., Righini N., Carbonero F., Estrada A., et al. (2013). Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. ISME J. 7, 1344–1353. Available at: https://www.nature.com/articles/ismej201316 (Accessed July 8, 2024).
Annan A., Baldwin H. J., Corman V. M., Klose S. M., Owusu M., Nkrumah E. E., et al. (2013). Human betacoronavirus 2c EMC/2012–related viruses Anthony in bats, Ghana and Europe. Emerg. Infect. Dis. 19, 456. doi: 10.3201/eid1903.121503
Anthony E. L. P. (1988). “Age determination in bats,” in Ecological and behavioral methods for the study of bats. Ed. Kunz T. H. (Smithsonian Institution Press, Washington, DC), 47–58.
Apprill A., McNally S., Parsons R., Weber L. (2015). Minor revision to V4 region SSU rRNA 806R gene primer greatly increases detection of SAR11 bacterioplankton. Aquat. Microb. Ecolog. 75, 129–137. doi: 10.3354/ame01753
Arita H. T. (1997). Species composition and morphological structure of the bat fauna of Yucatan, Mexico. J. Anim. Ecol. 66, 83–97. doi: 10.2307/5967
Bai Y., Hayman D. T., McKee C. D., Kosoy M. Y. (2015). Classification of Bartonella strains associated with straw-colored fruit bats (Eidolon helvum) across Africa using a multi-locus sequence typing platform. PloS Negl. Trop. Dis. 9, e0003478. doi: 10.1371/journal.pntd.0003478
Barelli C., Albanese D., Donati C., Pindo M., Dallago C., Rovero F., et al. (2015). Habitat fragmentation is associated to gut microbiome diversity of an endangered primate: implications for conservation. Sci. Rep. 5, 14862. doi: 10.1038/ismej.2013.16
Bisanz J. E. (2018). qiime2R: Importing QIIME2 artifacts and associated data into R sessions (v0.99) [Computer software]. Available online at: https://github.com/jbisanz/qiime2r (Accessed February 29, 2024).
Bolatti E. M., Viarengo G., Zorec T. M., Cerri A., Montani M. E., Hosnjak L., et al. (2022). Viral metagenomic data analyses of five new world bat species from Argentina: identification of 35 novel DNA viruses. Microorganisms 10, 266. doi: 10.3390/microorganisms10020266
Bolívar-Cimé B., Laborde J., MacSwiney G. M. C., Muñoz-Robles C., Tun-Garrido J. (2013). Response of phytophagous bats to patch quality and landscape attributes in fragmented tropical semi-deciduous forest. Acta Chiropt. 15, 399–409. doi: 10.3161/150811013x679026
Bolyen E., Rideout J. R., Dillon M. R., Bokulich N. A., Abnet C. C., Al-Ghalith G. A., et al. (2019). Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 37, 852–857. doi: 10.1038/s41587-019-0209-9
Burgar J. M., Hitchen Y., Prince J. (2021). Effectiveness of bat boxes for bat conservation and insect suppression in a Western Australian urban riverine reserve. Austral Ecol. 46, 186–191. doi: 10.1111/aec.12980
Burki T. (2020). The origin of SARS-coV-2. Lancet Infect. Dis. 20, 1018–1019. doi: 10.1016/S1473-3099(20)30641-1
Cabrera-Romo S., Max Ramirez C., Recio-Tótoro B., Tolentino-Chi J., Lanz H., Del Angel R. M., et al. (2016). No evidence of dengue virus infections in several species of bats captured in central and southern Mexico. ZPHOAH 63, 579–583. doi: 10.1111/zph.12276
Cafaggi D., Marín G., Medellin R. A. (2024). Bats and Mayan temples: Bat diversity and the potential for conservation of archeological zones in Yucatan, Mexico. Biotropica 56, e13350. doi: 10.1111/btp.13350
Callahan B. J., McMurdie P. J., Rosen M. J., Han A. W., Johnson A. J. A., Holmes S. P. (2016). DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Carrillo-Araujo M., Taş N., Alcantara-Hernandez R. J., Gaona O., Schondube J. E., Medellin R. A., et al. (2015). Phyllostomid bat microbiome composition is associated to host phylogeny and feeding strategies. Front. Microbiol. 6. doi: 10.3389/fmicb.2015.00447
CCMSS, México (2024). Available online at: https://ccmss.org.mx/.
Chávez-Arias N. P., Chim J. E. V., Shanahan M. W., Buenfil D. J. B., Ceballos M. J. R., Valencia J. E. R., et al. (2017). Studying sociocultural factors associated with dengue fever in elementary school children in Yucatan, Mexico (London: SAGE Publications Ltd). doi: 10.4135/9781473998551
Chen S., Zhou Y., Chen Y., Gu J. (2018). Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinform. 34, i884–i890. doi: 10.1093/bioinformatics/bty560
Cláudio V. C., Gonzalez I., Barbosa G., Rocha V., Moratelli R., Rassy F. (2018). Bacteria richness and antibiotic-resistance in bats from a protected area in the Atlantic Forest of Southeastern Brazil. PloS One 13, e0203411. doi: 10.1371/journal.pone.0203411
Colunga-Salas P., Hernández-Canchola G. (2020). Bats and humans during the SARS-CoV-2 outbreak: The case of bat-coronaviruses in Mexico. Transbound Emerg. Dis. 68, 987–992. doi: 10.1111/tbed.13751
Cumming G. S., Allen C. R. (2017). Protected areas as social-ecological systems: perspectives from resilience and social-ecological systems theory. Ecol. Appl. 27, 1709–1717. doi: 10.1002/eap.1584
de Souza R., Vizentin-Bugoni J. (2020). Diversity of fruits in Artibeus lituratus diet in urban and natural habitats in Brazil: a review. J. Trop. Ecol. 36, 65–71. doi: 10.1017/S0266467419000373
Donaldson E. F., Haskew A. N., Gates J. E., Huynh J., Moore C. J., Frieman M. B. (2010). Metagenomic analysis of the viromes of three North American bat species: viral diversity among different bat species that share a common habitat. J. Virol. 84, 13004–13018. doi: 10.1128/jvi.01255-10
Dzul-Rosado K., Castillo-León T., Montalvo-Nah E., Arias-León J., Puerto-Manzano F. (2023). Perception of risk among children: Exploring the risk of TB-rickettsial disease based on the children’s drawing pictures in a Mayan community of Yucatan. Health Promot. Perspect. 13, 129. doi: 10.34172/hpp.2023.16
Ellis E. A., Gomez U. H., Romero-Montero J. A. (2017). Los procesos y causas del cambio en la cobertura forestal de la Península Yucatán, Mexico. Ecosist. 26, 101–111. doi: 10.7818/ecos.2017.26–1.16
Federici L., Masulli M., De Laurenzi V., Allocati N. (2022). An overview of bats microbiome and its implication in transmissible diseases. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.1012189
Gannon M. R., Bovard B. N., Butchkoski C. M., Reeder D. M., Turner G. G., Whidden H. P. (2016). “The value of bats: Keystone species in the Keystone State,” in Conservation and ecology of pennsylvania’s bats.ds. Butchkoski M. B. D. M., Reeder D. M., Turner G. G., Whidden H. P. (Pennsylvania Academy of Science, Pennsylvania, USA).
Gannon W. L., Sikes R. S. (2007). The animal care and use committee of the american society of mammalogists, guidelines of the american society of mammalogists for the use of wild mammals in research. J. Mammal. 88, 3, 809–823. doi: 10.1644/06-mamm-F-185R1.1
Gaona O., Colli A., Cruzado J., Falcón L., López S., Marruenda P., et al. (2024). Catálogo de murciélagos de la Península de Yucatán. México: Soluciones Ambientales Itzeni, A.C.
García-Frapolli E., Ayala-Orozco B., Bonilla-Moheno M., Espadas-Manrique C., Ramos-Fernández G. (2007). Biodiversity conservation, traditional agriculture and ecotourism: Land cover/land use change projections for a natural protected area in the northeastern Yucatan Peninsula, Mexico. Landsc. Urban Plan. 83, 137–153. doi: 10.1016/j.landurbplan.2007.03.007
García-Rejon J., Loroño-Pino M. A., Farfan-Ale J. A., Flores-Flores L., Rosado-Paredes E. D. P., Rivero-Cardenas N., et al. (2008). Dengue virus-infected Aedes aEgypti in the home environment. ASTMH 79, 940–950.
Gardner A. L. (1979). “Feeding habits,” in Biology of bats of the new world family phyllostomatidae. Part II. Eds. Baker R. J., Jones J., Knox J., Carter D. C. (Special Publications the Museum Texas Tech University, Lubbock, TX), 293–350.
Gómez-Pompa A., Kaus A. (1999). From pre-Hispanic to future conservation alternatives: lessons from Mexico. PNAS 96, 5982–5986. doi: 10.1073/pnas.96.11.598
González-Iturbe J. A., Olmsted I., Tun-Dzul F. (2002). Tropical dry forest recovery after long term Henequen (sisal, Agave fourcroydes Lem.) plantation in northern Yucatan, Mexico. For. Ecol. Manage. 167, 67–82. doi: 10.1016/s0378-1127(01)00689-2
Hernández-Dávila A., Vargas J. A., Martínez-Méndez N., Lim B. K., Engstrom M. D., Ortega J. (2012). DNA barcoding and genetic diversity of phyllostomid bats from the Yucatan Peninsula with comparisons to Central America. Mol. Ecol. Resour. 12, pp.590–pp.597. doi: 10.1111/j.1755-0998.2012.03125.x
Hougner C., Colding J., Söderqvist T. (2006). Economic valuation of a seed dispersal service in the Stockholm National Urban Park, Sweden. Ecol. Econ. 59, 364–374. doi: 10.1016/j.ecolecon.2005.11.007
INEGI, México (2020). Available online at: https://www.inegi.org.mx/.
Ingala M. R., Simmons N. B., Dunbar M., Wultsch C., Krampis K., Perkins S. L. (2021). You are more than what you eat: potentially adaptive enrichment of microbiome functions across bat dietary niches. Anim. Microbiome 3, 1–17. doi: 10.1186/s42523-021-00139-8
Ingala M. R., Simmons N. B., Perkins S. L. (2018). Bats are an untapped system for understanding microbiome evolution in mammals. Sphere 3, e00397–e00318. doi: 10.1128/mSphere.00397-18
Islebe G. A., Schmook B., Calmé S., León-Cortés J. L. (2015). “Introduction: biodiversity and conservation of the Yucatán Peninsula, Mexico,” in Biodiversity and conservation of the Yucatan Peninsula. Eds. Islebe G. A., Calmé S., León-Cortés J. L., Schmook B. (Springer International Publishing, Cham), 1–5. doi: 10.1007/978-3-319-06529-8
Jones B. A., Grace D., Kock R., Alonso S., Rushton J., Said M. Y., et al. (2013). Zoonosis emergence linked to agricultural intensification and environmental change. PNAS 110, 8399–8404. doi: 10.1073/pnas.1208059110
Jones G., Jacobs D. S., Kunz T. H., Willig M. R., Racey P. A. (2009). Carpe noctem: the importance of bats as bioindicators. Endanger. Species Res. 8, 93–115. doi: 10.3354/esr00182
Jones C., McShea W. J., Conroy M. J., Kunz T. H. (1996). “Capturing mammals,” in Measuring and monitoring biological diversity, standard methods for mammals. Eds. Wilson D. E., Cole F. R., Nichols J. D., Rudran R., Foster M. S.. Washington D.C.: Smithsonian Institute Press, 115–155.
Kasso M., Balakrishnan M. (2013). Ecological and economic importance of bats (Order Chiroptera). Int. Sch. Res. Notices 2013, 187415. doi: 10.1155/2013/187415
Katoh K., Misawa K., Kuma K., Miyata T. (2002). MAFFT: A novel method for rapid multiple sequence alignment based on fast Fourier transform. Nucleic Acids Res. 30, 3059–3066. doi: 10.1093/nar/gkf436
Lahti L., Shetty S. (2019). Microbiome R package [Computer software]. Available online at: https://microbiome.github.io (Accessed February 29, 2024).
Laurance S. G. (2004). “Landscape connectivity and biological corridors,” in Conclusion: agroforestry and biodiversity conservation in tropical landscapes. Eds. Schroth G., Da Fonseca G. A., Harvey C. A., et al (Washington USA: Island Press), 50–63.
Letko M., Seifert S. N., Olival K. J., Plowright R. K., Munster V. J. (2020). Bat-borne virus diversity, spillover and emergence. Nat. Rev. Microbiol. 18, pp.461–pp.471. doi: 10.1038/s41579-020-0394-z
Li W., Godzik A. (2006). Cd-hit: A fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinform 22, 1658–1659. doi: 10.1093/bioinformatics/btl158
Lu M., Wang X., Ye H., Wang H., Qiu S., Zhang H., et al. (2021). Does public fear that bats spread COVID-19 jeopardize bat conservation? Biol. Conserv. 254, 108952. doi: 10.1016/j.biocon.2021.108952
Lucero L. J. (2018). A cosmology of conservation in the ancient Maya world. J. Anthropol. Res. 74, 327–359. doi: 10.1086/698698
Lugo-Caballero C., Torres-Castro M., López-Ávila K., Hernández-Betancourt S., Noh-Pech H., Tello-Martín R., et al. (2021). Molecular identification of zoonotic Rickettsia species closely related to R. typhi, R. felis, & R. rickettsii in bats from Mexico. Indian J. Med. Res. 154, 536–538. doi: 10.4103/ijmr.IJMR_1083_19
Machain-Williams C., López-Uribe M., Talavera-Aguilar L., Carrillo-Navarrete J., Vera-Escalante L., Puerto-Manzano F., et al. (2013). Serologic evidence of flavivirus infection in bats in the Yucatan Peninsula of Mexico. J. Wildl. Dis. 49, 684–689. doi: 10.7589/2012-12-318
Mackenzie J. S., McKinnon M., Jeggo M. (2014). “One Health: from concept to practice,” in Confronting emerging zoonoses. Eds. Yamada A., Kahn L., Kaplan B., Monath T., Woodall J., Conti L. (Spinger, Tokyo, Japan), 163–189. doi: 10.1007/978-4-431-55120-1_8
MacSwiney M. C., Bolívar-Cimé B., Alfaro-Bates R., Ortíz-Díaz J. J., Clarke F. M., Racey P. A. (2017). Pollen movement by the bat Artibeus jamaicensis (Chiroptera) in an agricultural landscape in the Yucatan Peninsula, Mexico. Mam. Res. 62, 189–193. doi: 10.1007/s13364-016-0306-9
McMurdie P. J., Holmes S. (2013). Phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PloS One 8, e61217. doi: 10.1371/journal.pone.0061217
Medellin R. A., Wiederholt R., Lopez-Hoffman L. (2017). Conservation relevance of bat caves for biodiversity and ecosystem services. Biol. Conserv. 211, 45–50. doi: 10.1016/j.biocon.2017.01.012
Meyer C. F., Fründ J., Lizano W. P., Kalko E. K. (2008). Ecological correlates of vulnerability to fragmentation in Neotropical bats. J. Appl. Ecol. 45, 381–391. doi: 10.1111/j.1365-2664.2007.01389.x
Montiel S., Estrada A. L., León P. (2006). Bat assemblages in a naturally fragmented ecosystem in the Yucatan Peninsula, Mexico: species richness, diversity and spatio-temporal dynamics. J. Trop. Ecol. 22, 267–276. doi: 10.1017/S026646740500307X
Montiel S., Estrada A. L., León P. (2011). Reproductive seasonality of fruit-eating bats in northwestern Yucatan, Mexico. Acta Chiropt. 13, 139–145. doi: 10.3161/150811011X578688
Moretto L., Coleman J. L., Davy C. M., Fenton M. B., Korine C., Patriquin K. J. (2023). Urban bats: biology, ecology, and human dimensions (Switzerland: Springer Nature), 9783031131738.
Ortega R. P., Jaber I. G. (2022). A controversial train heads for the Maya forest. Sci. 375, 250–251. doi: 10.1126/science.ada0230
Price M. N., Dehal P. S., Arkin A. P. (2010). FastTree 2 – approximately maximum-likelihood trees for large alignments. PloS One 5, e9490. doi: 10.1371/journal.pone.0009490
Quast C., Pruesse E., Yilmaz P., Gerken J., Schweer T., Yarza P., et al. (2013). The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219
Ramirez-Francel L. A., García-Herrera L. V., Losada-Prado S., Reinoso-Flórez G., Sánchez-Hernández A., Estrada-Villegas, et al. (2022). Bats and their vital ecosystem services: a global review. Integr. Zool. 17, 2–23. doi: 10.1111/1749-4877.12552
R Core Team (2024). R: A language and environment for statistical computing (4.3.3) [R] (R Foundation for Statistical Computing). Available at: https://www.R-project.org/ (Accessed February 29, 2024).
RELCOM Latinoamérica. (2024). Available online at: https://www.relcomlatinoamerica.net/%C2%BFqu%C3%A9-hacemos/educaci%C3%B3n.html (Accessed November 20, 2024).
Rydell J., Arita H. T., Santos M., Granados J. (2002). Acoustic identification of insectivorous bats (order Chiroptera) of Yucatan, Mexico. J. Zool. 257, 27–36. doi: 10.1017/S0952836902000626
Saba-Villarroel P. M., Gumpangseth N., Songhong T., Yainoy S., Monteil A., Leaungwutiwong P., et al. (2023). Emerging and re-emerging zoonotic viral diseases in Southeast Asia: One Health challenge. Front. Public Health 11. doi: 10.3389/fpubh.2023.1141483
Sánchez-Soto M. F., Gaona O., Vigueras-Galván A. L., Suzán G., Falcón L. I., Vázquez-Domínguez E. (2024). Prevalence and transmission of the most relevant zoonotic and vector-borne pathogens in the Yucatan peninsula: A review. PloS Negl. Trop. Dis. 18, e0012286. doi: 10.1371/journal.pntd.0012286
SENASICA, Mexico (2024). Available online at: https://www.gob.mx/senasica/acciones-y-programas/campana-nacional-para-la-prevencion-y-control-de-la-rabia-en-bovinos-y-especies-ganaderas (Accessed July 8, 2024).
Sieradzki A., Mikkola H. (2022). “Bats in folklore and culture: a review of historical perceptions around the world,” in Bats–disease-prone but beneficial. Ed. Mikkola H. (British Library, London, United Kingdom). doi: 10.5772/interchopen.95729
Sosa-Escalante J. E., Pech-Canché J. M., MacSwiney M. C., Hernández-Betancourt S. (2013). Mamíferos terrestres de la península de Yucatán, Mexico: riqueza, endemismo y riesgo. Rev. Mex. Biodiv. 84, 949–969. doi: 10.7550/rmb.33285
Špirić J., Vallejo M., Ramirez M. I. (2022). Impact of productive activities on forest cover change in the Calakmul biosphere reserve region: Evidence and research gaps. Trop. Conserv. Sci. 15. doi: 10.1177/1940082922110
Stuckey M. J., Chomel B. B., Galvez-Romero G., Olave-Leyva J. I., Obregón-Morales C., Moreno-Sandoval H., et al. (2017). Bartonella infection in hematophagous, insectivorous, and phytophagous bat populations of Central Mexico and the Yucatan Peninsula. Am. J. Trop. Med. Hyg. 97, 413. doi: 10.4269/ajtmh.16-0680
Threlfall C., Law B., Penman T., Banks P. B. (2011). Ecological processes in urban landscapes: mechanisms influencing the distribution and activity of insectivorous bats. Ecography 34, 814–826. doi: 10.1111/j.1600-0587.2010.06939.x
Tidball K. G. (2012). Urgent biophilia: human-nature interactions and biological attractions in disaster resilience. Ecol. Soc 17. doi: 10.5751/ES-04596-170205
Torres-Castro M., Cuevas-Koh N., Hernández-Betancourt S., Noh-Pech H., Estrella E., Herrera-Flores B., et al. (2021). Natural infection with Trypanosoma cruzi in bats captured in Campeche and Yucatán, Mexico. Bioméd. 41, 131–140. doi: 10.7705/biomedica.5450
Torres-Castro M., Febles-Solis V., Hernandez-Betancourt S., Noh-Pech H., Estrella E., Peláez-Sánchez R., et al. (2020). Pathogenic leptospira in bats from campeche and yucatán, Mexico. Rev. MVZ Córdoba 25, pp.1–pp16. doi: 10.21897/rmvz.1815
Valdez-Tah A., Huicochea-Gómez L., Ortega-Canto J., Nazar-Beutelspacher A., Ramsey J.M. (2015). Social representations and practices towards triatomines and Chagas disease in Calakmul, México. Plos one 10 (7), e0132830.
Vester H. F., Lawrence D., Eastman J. R., Turner B. L., Calmé S., Dickson R., et al. (2007). Land change in the southern Yucatan and Calakmul Biosphere Reserve: effects on habitat and biodiversity. Ecol. Appl. 17, 989–1003.https://doi.org/10.1890/05-1106. doi: 10.1890/05-1106
Vidal-Zepeda R. (1990). Atlas Nacional de Mexico. (Mexico city: Instituto Nacional de Geografía, UNAM). Available at: https://geodigital.geografia.unam.mx/atlas_nacional/index.html/ (Accessed February 29, 2024).
Vilchis P., Clarke F. M., Racey P. A. (2007). The importance of cenotes in conserving bat assemblages in the Yucatan, Mexico. Biol. Conserv. 136, 499–509. doi: 10.1016/j.biocon.2006.12.021
Wickham H. (2009). Ggplot2: elegant graphics for data analysis (Springer Science and Business Media). Available at: http://www.springer.com/978-0-387-98140-6 (Accessed February 29, 2024).
Wickham H., Averick M., Bryan J., Chang W., McGowan L. D., François R., et al. (2019). Welcome to the tidyverse. JOSS 4, 1686. doi: 10.21105/joss.01686
Wolf J. M., Jeschke J. M., Voigt C. C., Itescu Y. (2022). Urban affinity and its associated traits: A global analysis of bats. Glob. Change Biol. 28, 5667–5682. doi: 10.1111/gcb.16320
Wolkers-Rooijackers J., Rebmann K., Bosch T., Hazeleger W. C. (2018). Fecal bacterial communities in insectivorous bats from the Netherlands and their role as a possible vector for foodborne diseases. Acta Chiropt. 20, 475–483. doi: 10.3161/15081109ACC2018.20.2.017
Keywords: Artibeus jamaicensis, Artibeus lituratus, bats, biophilia, habitat fragmentation, microbiome, One Health, Yucatán Peninsula
Citation: Sánchez-Soto MF, Gaona O, Mercado-Juárez R, Yanez-Montalvo A, de León-Lorenzana A, Borja-Martínez G, Zaldívar D, Rodríguez-González S, Hernández-Villegas EN, Moreira-Soto A, Drexler JF, Suzán G, Vázquez-Domínguez E and Falcón LI (2025) Ecological-based insights into bat populations in the Yucatán Peninsula under a One Health approach: coexistence or biophobia. Front. Conserv. Sci. 5:1488378. doi: 10.3389/fcosc.2024.1488378
Received: 29 August 2024; Accepted: 30 December 2024;
Published: 31 January 2025.
Edited by:
Jamie K. Reaser, Smithsonian Conservation Biology Institute (SI), United StatesReviewed by:
Jason Kirkey, Smithsonian Conservation Biology Institute (SI), United StatesCopyright © 2025 Sánchez-Soto, Gaona, Mercado-Juárez, Yanez-Montalvo, de León-Lorenzana, Borja-Martínez, Zaldívar, Rodríguez-González, Hernández-Villegas, Moreira-Soto, Drexler, Suzán, Vázquez-Domínguez and Falcón. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luisa I. Falcón, ZmFsY29uQGVjb2xvZ2lhLnVuYW0ubXg=; bHVpc2FpZmFsY29uQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.