 Kingsley J. Griffin
Kingsley J. Griffin Emma L. Johnston
Emma L. Johnston Alistair G. B. Poore
Alistair G. B. Poore Adriana Vergés
Adriana Vergés Graeme F. Clark
Graeme F. Clark- 1School of Biological, Earth, and Environmental Sciences, University of New South Wales, Sydney, NSW, Australia
- 2School of Life and Environmental Sciences, University of Sydney, Sydney, NSW, Australia
Physical disturbances typically cause ecological impacts within areas of direct contact (primary disturbances) but can also impact surrounding areas through other mechanisms (secondary disturbances). Secondary disturbances are often overlooked, especially in marine ecosystems where sufficiently detailed observation can be difficult to obtain. For example, boat moorings create circular clearings in seagrass meadows by physically scouring the seabed, but visible impacts extend beyond this into surrounding areas due to disturbances such as increased sediment transport, edge effects, and shading from boats. Previous studies on impacts of moorings have not distinguished between primary and secondary disturbances, and secondary disturbances are rarely accounted for in environmental management. We used spatial modelling to examine the primary and secondary disturbances associated with moorings in a meadow of the threatened seagrass Posidonia australis. We compared the disturbance from traditional ‘swing’ moorings with ‘environmentally friendly’ moorings (EFM) designed to reduce scour. Within the scour zone (0-5 m from moorings), we compared seagrass cover around swing moorings with cover around EFM. Further from moorings (5-25 m), we tested the degree of association between seagrass cover and secondary disturbances that may accumulate with mooring density and be influenced by mooring design. We found that mooring design affected the degree of direct disturbance, with cover of P. australis in the scour zone of standard moorings ~49% lower than that of EFMs (p < 0.01). Mooring density had cumulative negative effects on P. australis cover in the surrounding meadow, but the influence of mooring density was reduced when most moorings were EFMs. This suggests that secondary effects contingent on direct physical disturbance (e.g. sediment transport, fragmentation, edge effects) may be stronger than the general influence of moored boats (shading). We use the findings to simulate two broad policy scenarios for mooring designs in P. australis meadows: installing only EFM, or only traditional moorings. The simulations suggest that using only EFM would lead to a 14.2% increase (~164 m2), whereas installing only traditional moorings would lead to a 16% (~187 m2) loss of P. australis cover in this already highly disturbed meadow. Synthesis and applications: We demonstrate a nondestructive approach to assessing primary and secondary disturbances driving the distribution of a threatened seagrass and simulate a range of potential management scenarios that could assist in conserving the species.
Introduction
Anthropogenic disturbance and modification of natural habitats are major drivers of the systemic decline of biodiversity worldwide (Ojima et al., 1994; Waycott et al., 2009). Physical disturbance from human activities (e.g. clearing and building) can generate sharp environmental boundaries, but can also introduce secondary processes that disturb surrounding habitats. These so-called ‘secondary disturbances’ include changes in light, water, sound and nutrient regimes, disrupting animal movements and behaviour, or assisting the spread of invasive pests (Walker et al., 1987; Laurance and Yensen, 1991; Murcia, 1995). Secondary disturbances can be mechanistically linked or contingent on primary disturbances, and have lower relative intensity though they may be more persistent. The difficulty of detecting secondary disturbances means they are sometimes poorly understood and often overlooked by environmental impact assessments (Walker et al., 1987; Erftemeijer and Robin Lewis, 2006). Whilst several of the key mechanisms causing biodiversity loss are well investigated, the roles of and interactions between primary and secondary disturbances are still unknown for many species and stressors (Maxwell et al., 2017).
A prominent example of an environmental impact resulting from both primary and secondary disturbances in the marine environment is the impact of boat moorings on seagrasses. Moorings physically disturb the seabed within a ‘scour zone’ and potentially introduce a range of secondary disturbances in the surrounding area. For example, clearings from scour can disturb surrounding seagrasses by increasing sediment movement (Frost et al., 1999), burying plants, driving fragmentation, and impacting resource translocation (Tuya et al., 2013). Open space can increase exposure to storm damage and give fast colonising species (e.g. species of the genera Zostera and Halophila) a competitive advantage, which may outcompete slower-growing species (Kirkman, 1985). Shading by moored vessels is another potential secondary disturbance that extends beyond the scour zone (Evans et al., 2017). Each of these secondary disturbances that extend into the surrounding meadow may accumulate with increasing mooring density.
Elsewhere, efforts to conserve seagrasses have included mitigation of direct impacts (e.g. no anchoring areas), reducing pollution from neighbouring watersheds (e.g. Bryars and Rowling, 2009), or active restoration (e.g. Paling et al., 2007). The limited success of restoration and ongoing declines of seagrasses globally suggest that losses are challenging to reverse, emphasising the need to understand and mitigate the processes causing loss (Cullen-Unsworth and Unsworth, 2016). One option to reduce impacts from boating is through environmentally friendly mooring designs, which reduce physical disturbance and potentially some secondary disturbances. Trials of new moorings that avoid physical disturbance have shown some success (Demers et al., 2013), posing a potential strategy to mitigate impacts on seagrasses (Glasby and West, 2018).
Coastal monitoring programs are our chief method for detecting ecological changes, but most lack sufficient spatial and temporal resolution to attribute and manage change before losses occur (Field et al., 2007; Schultz et al., 2015). Without sampling at higher spatial and temporal resolution, we may fail to detect or mitigate local processes that contribute to declines in difficult-to-restore habitats, or miss the early-warning signs of ecosystem collapse (Duarte, 2002; Dakos et al., 2012; Kéfi et al., 2014; Schultz et al., 2015). This is especially true for secondary disturbances, which are typically more subtle and spatially distributed than primary disturbances. Most marine and coastal ecosystems lack sufficient spatial and temporal data to detect small-scale losses before they escalate (Pressey, 2004) or impacts of secondary disturbances.
Methods from landscape ecology use detailed observations over large areas to untangle the local and regional processes that drive ecological patterns, but practical limitations have typically precluded this style of study from marine subtidal environments (Pittman, 2017). However, recent technological developments have facilitated the use of these methods in marine systems, including sonar- and accelerometer-based positioning systems, remote autonomous vehicles (Hill et al., 2014), and high-resolution imaging converted to ecological data via computer-vision and machine-learning software (e.g. Beijbom et al., 2015; Griffin et al., 2017). Spatial analytical approaches allow for investigating relationships between impact exposure and species distributions, with taxonomic and spatial resolution limited only by the sampling methodology (Elith and Leathwick, 2009). When matching spatial data is available to describe the intensity of the stressor, distribution modelling can assess the relationship and predict species’ response across space (Elith and Leathwick, 2009). Spatially explicit ‘seascape’ analyses are still relatively rare in sub-tidal marine systems but might improve our understanding of sensitive habitats and processes that cause their decline (Brown and Hamilton, 2018). Similar approaches can be applied to risk analysis, impact assessment, and multiple-stressor studies, and can improve our ability to simulate and test the effectiveness of regulation and management in complex ecosystems (Dumbauld and McCoy, 2015; Martin et al., 2018).
In this study, we use a landscape approach to assess the impacts of coastal infrastructure on the threatened seagrass Posidonia australis, which forms a dominant and valuable habitat in southern Australia. Declines since the 1950s (Glasby and West, 2015) and ongoing risks from coastal development (Clark and Johnston, 2016) have led to P. australis being listed as a regionally endangered ecological community in some areas of New South Wales under the NSW Fisheries Management Act 1994 in 2010 and the Australian Environment Protection and Biodiversity Conservation Act 1999 in 2015. Despite the legislative protection, P. australis meadows in some estuaries have declined by up to 40% in recent years (Evans et al., 2018). Moorings have removed an estimated ~130,000 m2 of the species (Glasby and West, 2018). Individual scars may take centuries to recover (Meehan and West, 2000) due to slow growth and the fact that P. australis relies on intact rhizome material to stabilise sediments for new shoots (Meehan and West, 2000). Investigations into moorings effects on P. australis have concentrated on the rate and area of physical removal (Demers et al., 2013; Evans et al., 2018; Glasby and West, 2018). These losses, however, could be difficult to predict if secondary disturbances that accumulate with mooring density result in non-linear relationships between distance (exposure) to moorings and seagrass survival (Unsworth et al., 2017).
We aimed to investigate whether environmentally friendly moorings (EFM) reduced the primary and secondary disturbance from boat moorings for the threatened seagrass P. australis and two non-threatened and co-occurring species of seagrass (Zostera muelleri and Halophila ovalis). We surveyed a seagrass meadow at Manly Cove, Sydney Harbour, at a high spatial resolution (<1 m) to investigate the response of P. australis, Z. muelleri and H. ovalis to the proximity and density of the two mooring designs. We separately tested the effects of mooring design (i) close to the mooring in the area subject to physical disturbance by scour (0-5m from moorings), and (ii) further from the mooring (5–25m away) where there is no scour, but secondary disturbance may be operating. We also demonstrate how a spatially explicit approach can aid decision-making by simulating the implications of three potential management actions at the site. Specifically, we used simulations to predict how the cover of P. australis may change in response to (i) replacing any single traditional mooring with an EFM, (ii) replacing all EFMs with traditional moorings, and (iii) replacing all traditional moorings with EFMs.
Methods
Site history
Urban development, industrial pollution and land reclamation have left seagrasses in Sydney Harbour in an ongoing decline (Evans et al., 2018). As of 2019, only two small seagrass meadows dominated by P. australis remain, the largest of which is at Manly Cove (Evans et al., 2018). Boat moorings have been present at Manly Cove since the 1940s, gradually growing to the high number and density of moorings in place today (Figures 1A, B). The P. australis meadow at Manly Cove is shallow (depth 2-5 m), and the dense array of traditional ‘swing’ moorings have left a highly fragmented meadow, with a further boundary to the west made by frequent ferry traffic at Manly wharf (Figure 1). An “environmentally friendly” moorings (EFM) trial was installed at Manly Cove from 2009. The design of EFMs consisted of a fixed point driven into the seabed, acting as a base for a swivel arm angled upwards (Figure 1E). The 25 trial EFMs are interspersed with 74 traditional moorings (Figure 1D), separated to avoid overlap with other moorings rather than avoiding or targeting the seagrass within the study area. Given the continued regional decline of P. australis, its protected status, and its susceptibility to physical disturbance, management agencies are interested in the fine-scale impact of both mooring designs.

Figure 1. Manly Cove moorings in 1943 (A) and 2010 (B), and image survey track with seagrass cover in blue, Nov 2016 (C). Underwater view of traditional mooring (D) and environmentally friendly (E) mooring designs at Manly Cove. Source: (A–C) Images from Google Earth.
Survey of seagrass abundance and distribution
We collected dense seabed imagery georeferenced with an underwater positioning system to determine the distribution and abundance of seagrass across a meadow with boat moorings. We comprehensively photographed the seabed on SCUBA in the first week of September 2016 (Figure 1C). We used a 16 MP mirrorless digital camera in underwater housing and positioning equipment, including a Leica ‘Viva’ GNSS sensor and Sonardyne ‘Scout’ ultra-short-baseline (USBL). The above-water side of the positioning system collected high-accuracy (+/- <0.2 m) GPS positions and 3-axis vessel orientation data within an inertial motion unit in the Sonardyne Scout. The undersea side of the USBL positioning system sent and received acoustic ‘pings’ between an underwater transducer and roving unit attached to the underwater camera, giving a distance and heading between the boat and the camera. A surface-side computer processed and recorded the signals, which were later rectified in R (v 3.4.4) (R Core Team, 2018).
Approximately 5200 images were collected at a consistent distance of ~50 cm from the seabed while SCUBA diving, avoiding overlap by a minimum separation of 3 m between photos. Real-time navigation was not possible while underwater, but the track position was reviewed after each dive to avoid revisiting an area and to concentrate survey efforts on areas where seagrass was present. Where a mooring was sighted, an image was taken directly above the mooring equipment, from which we obtained a precise position and design type for each mooring. Targeted snorkel surveys were undertaken after the primary survey to ensure no moorings were missed. The images were geo-referenced from the positioning system, colour-balanced, and uploaded to the CoralNet semi-automatic image classification server (Beijbom et al., 2015).
Semi-automated classification of images with CoralNet
The CoralNet interface follows a point-count method (e.g. Kohler and Gill (2006), whereby n randomly positioned points are classified for biological data and summed to create a percentage-cover measure. CoralNet uses a software ‘robot’ that suggests the most probable classification based on pixel and texture patterns (Beijbom et al., 2015) in an implementation of ‘deep learning’ (Jia et al., 2014). As the operator confirms more training points, the predictions generally improve, and the operator can elect to automatically accept annotations based on their statistical confidence. CoralNet is cross-validated using k-fold (k = 5) Cohen’s Kappa (see Beijbom et al. (2015) for more details) and presents the user with a confusion matrix of the various classification tags.
We classified 50 randomly positioned points in each image with codes following the CATAMI benthic image classification scheme (Althaus et al., 2015) to species for the three seagrasses at the site. We initially set a 98% threshold of alleviation until enough training data was generated to produce accurate (>75%) classifiers, at which time alleviation was gradually increased. Images were discarded during scoring if they showed signs of excessive distortion, poor focus, or obstructed view. For a comprehensive discussion around optimising points-in-images, image dimensions, and image survey techniques, see Perkins et al. (2016).
Data analysis
Disturbance within and beyond the scour zone
To compare the degree of direct physical impacts of EFM and swing mooring gear on seagrass abundance, observations within the range of mooring scour (0 - 5 m from each mooring) were isolated from other survey data. Z-tests were performed to compare species mean cover within range of scour by the two mooring designs (Supplementary Materials). The 5 m radius of physical scour was determined from a visual estimate of the length of the heavy chain on each traditional swing mooring at Manly Cove. Although there was variation among moorings, this was a conservative visual estimate (see underwater images in Figure 1) supported by sediment grain size analysis (Griffin et al. in prep). The distance from the coastline (a proxy for depth and wave exposure gradients) was calculated for each survey location using the 1:125,000 coastline distributed by Geoscience Australia (Geoscience Australia, 2004), as detailed bathymetry was unavailable.
For each seagrass species, we specified models to test for relationships between seagrass cover within the scour zone and the predictor variables distance to coast, mooring type, and their interaction. We then applied backward model selection to remove non-credible terms, starting with the interaction. Each final model included significant predictors and the random spatial field.
For the analyses within and beyond the scour zone, we employed spatial models that used stochastic partial differential equations (SPDE, Lindgren et al. (2011) and integrated nested Laplace approximation (INLA, Rue et al. (2009) using the ‘R-INLA’ package (www.r-inla.org, Lindgren and Rue (2015). These models accounted for the spatial arrangement of seagrass (autocorrelation) as a random spatial field. Detailed examples and usage instructions for the INLA modelling framework are described in (Krainski et al., 2018).
To understand the cumulative impacts of EFM and swing moorings on the broader seagrass meadow beyond the scour zone, observations outside the reach of mooring gear but within a conceivable range of indirect influence (5 - 25m from a mooring) were isolated. This distance was estimated to capture the majority of secondary effects from each individual mooring (e.g. due to the drifting arc of the moored vessel) while avoiding overlap with neighbouring mooring zones. The potential for accumulation of secondary effects was represented by calculating the density of either design of moorings within a 0.5-hectare area (a circle of radius 39.89 m) surrounding each point (Figure 2). This area parameter was informed by both the moored vessel dimensions and mooring spacing at the site to include only neighbouring moorings that could perceivably contribute to secondary effects (and not those further away). We do not claim to delineate these zones with high precision. The areas chosen simply represent regions that are likely to be exposed to the anticipated effects.
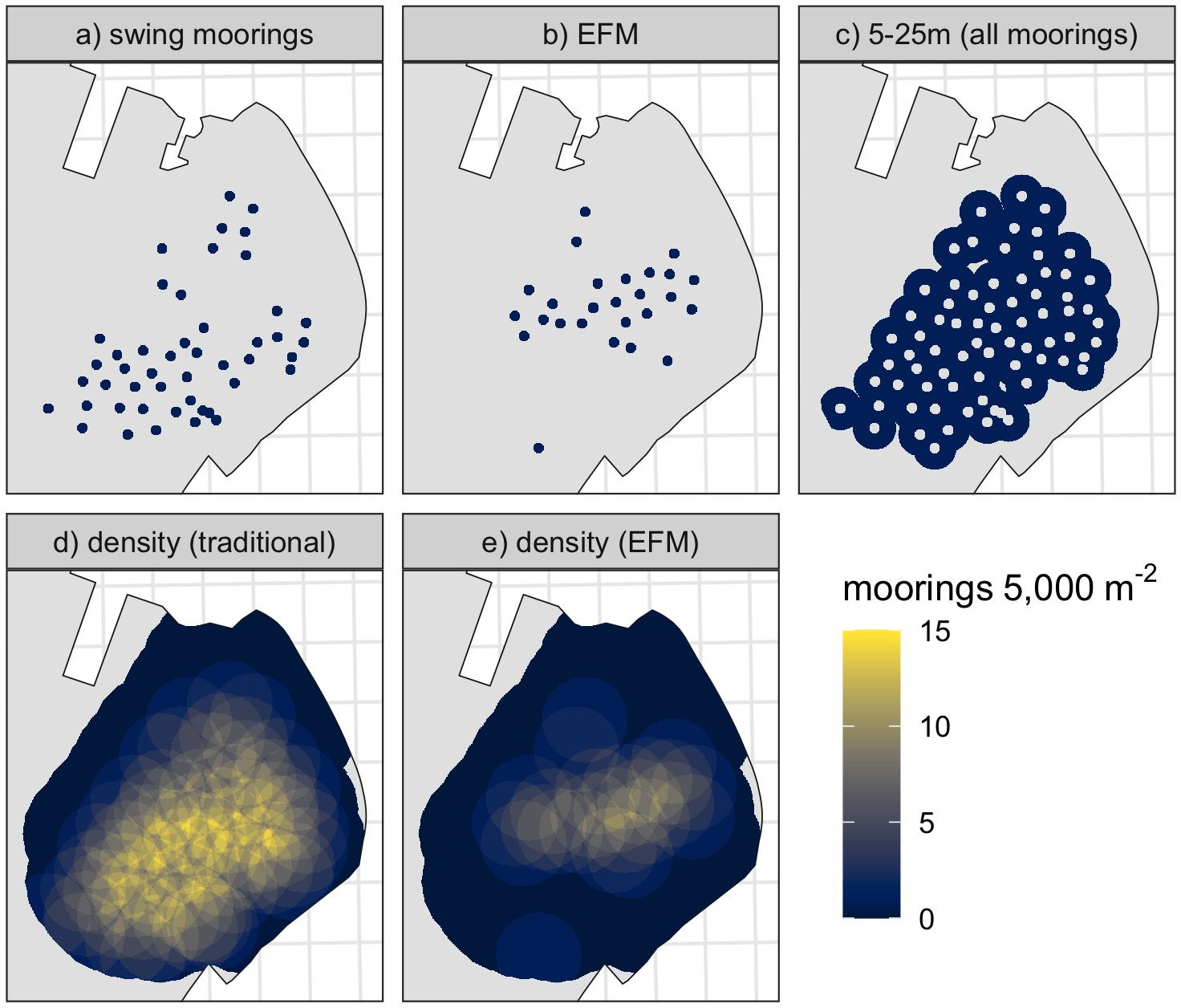
Figure 2. Spatial layers for covariates: (A) seabed 0-5m from traditional moorings, or (B) 0-5m from environmentally friendly screw moorings; (C) 5-25 m from all moorings; (D) density of traditional moorings, and (E) density of EFMs.
As in the previous analysis, we specified SPDE models to estimate the relationship of each seagrass species cover with the density of both moorings in the surrounding area (specified as an interaction), accounting for distance from the coastline and the spatial random field (autocorrelation). In this case, all model estimates for P. australis, including the interaction, had credible intervals that excluded zero and were retained. We maintained this model structure for the other two species to allow for direct comparison.
Meadow scale disturbance and simulated management effects on P. australis
To predict the effect of converting individual moorings on P. australis coverage, we simulated replacing 1 - 15 randomly selected traditional moorings at Manly Cove with EFMs to capture the variation introduced by cumulative effects. We repeated this ten times to capture the variance introduced by random mooring selection. The effects 0-5 m and 5-25 m away were then standardised for the number of moorings replaced, and seagrass cover change was calculated using the predicted relative cover of each cell.
To predict the impact of mooring policy change on P. australis, we simulated two scenarios: replacing existing EFMs with traditional moorings and replacing traditional moorings with EFMs. Then, we compared those two scenarios with the current arrangement. We generated new data layers for each scenario and obtained predictions using the model output and spatial random effect estimated from the above models.
Results
Disturbance within and beyond the scour zone
The cover of each seagrass species in the scour zone (0 to 5 m) was significantly lower around traditional moorings, relative to EFMs (p < 0.01). The effect of mooring design was greatest for P. australis, with cover in the scour zone around traditional moorings (5.09%) 49% less than around EFMs (10.38%; p < 0.01). Of the three seagrass species, cover of Z. muelleri differed least between mooring designs, but EFMs were still associated with significantly higher Z. muelleri cover in the scour area (2.6%; p < 0.01). The model predictions for the scour zone fit these observations and were acceptably robust (Figure 3; R2 = 0.648-0.809). Complete z-test tables of observed cover are available in the Supplementary Materials.
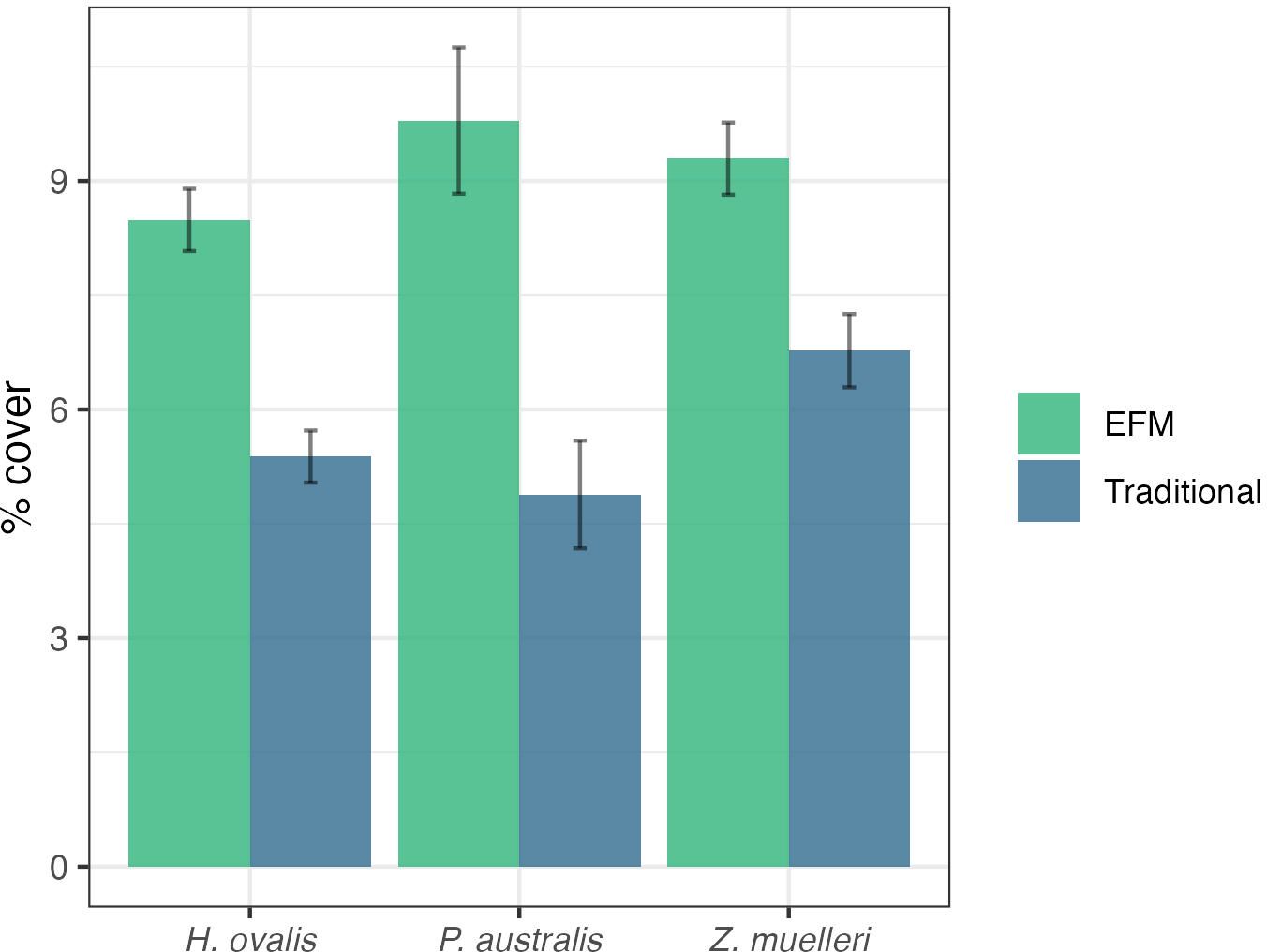
Figure 3. Predicted seagrass abundance within 5 m radius of environmentally friendly and traditional mooring installations (mean predicted abundance (%) and standard error around predictions; R2 = 0.809). Z-test scores for the observed values are provided in Supplementary Materials.
Beyond the range of physical disturbance (5 - 25 m from moorings), only P. australis abundance was significantly influenced by the density of neighbouring moorings (Figure 4). The abundances of Z. muelleri and H. ovalis were not significantly related to the density of either mooring design (Figure 4). Posidonia australis cover was associated with the interaction of traditional and EFM density measures, with a higher cover of P. australis where EFM density was higher, and where traditional mooring density was lower. The model fit for P. australis was acceptable (R2 = 0.57), performing well for low cover < 20% but generally under-predicting higher densities (Supplementary Material).
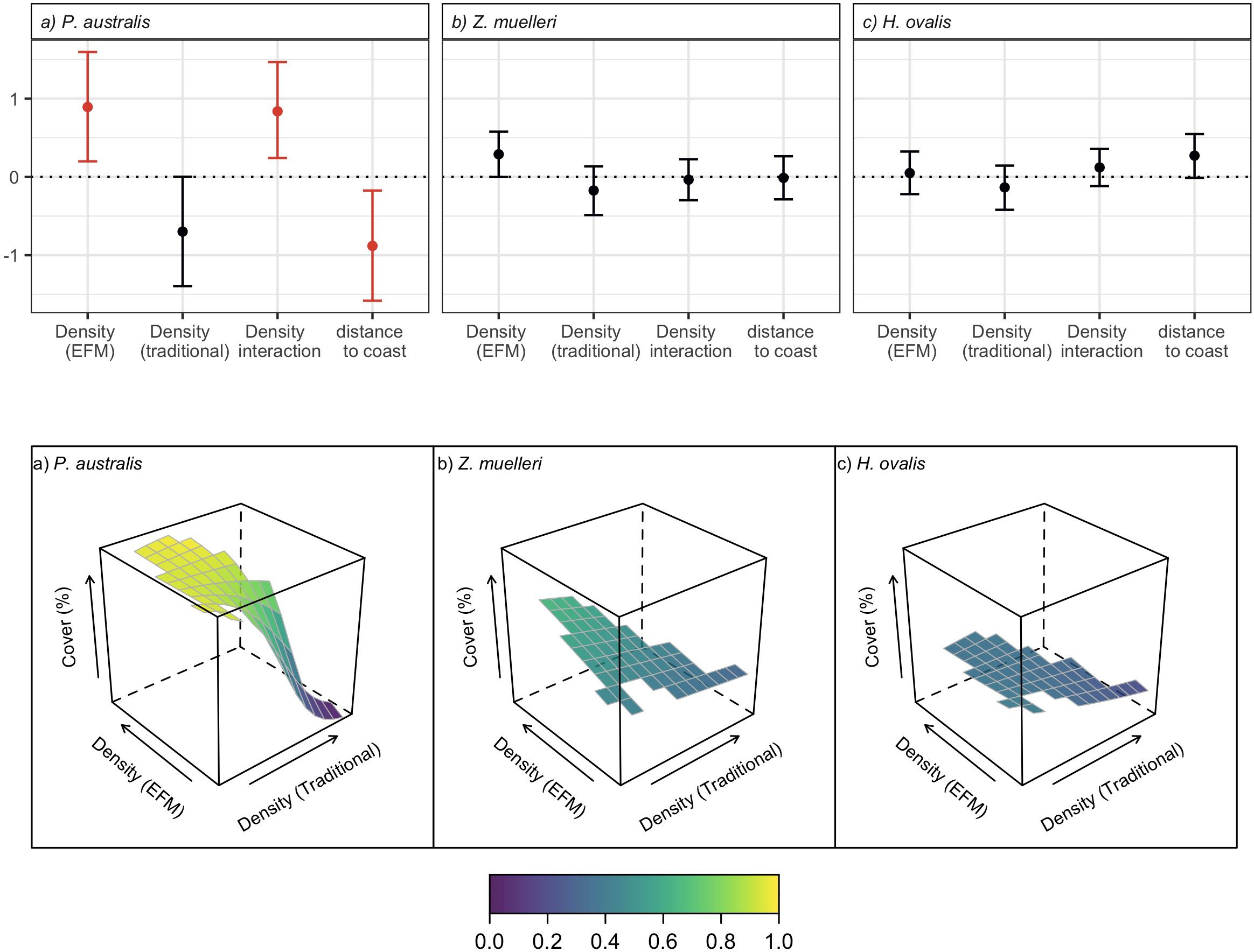
Figure 4. Upper pane: model estimates (scaled) and credible intervals (0.025 and 0.975 quantiles) for each of the terms in INLA SPDE models in the area 0-25 m from moorings for (A) P. australis; (B) Z. muelleri; and (C) H. ovalis. Estimates with credible intervals that exclude zero are presented in red. Lower pane: predicted seagrass cover 5 - 25 m from moorings under varying densities of both traditional and EF moorings, holding the spatial random field constant at the observed maximum for each species.
Meadow scale disturbance and simulated management effects on P. australis.
Replacing a traditional mooring with an EFM increased the predicted cover of Posidonia australis (Figure 5). The predicted increase in P. australis cover due to changing a traditional mooring to EFM was far greater in the non-scour area (mean increase of ~21.6 m2) than within the scour area (mean increase of ~1.6 m2). Replacing a single traditional mooring is predicted to deliver a mean increase of ~23 m2 of seabed covered by P. australis, which equates to a ~1.2% increase in P. australis cover in the area within a 25 m radius of the mooring, or the equivalent of a ~2.7 m radius circular patch with 100% seagrass cover.
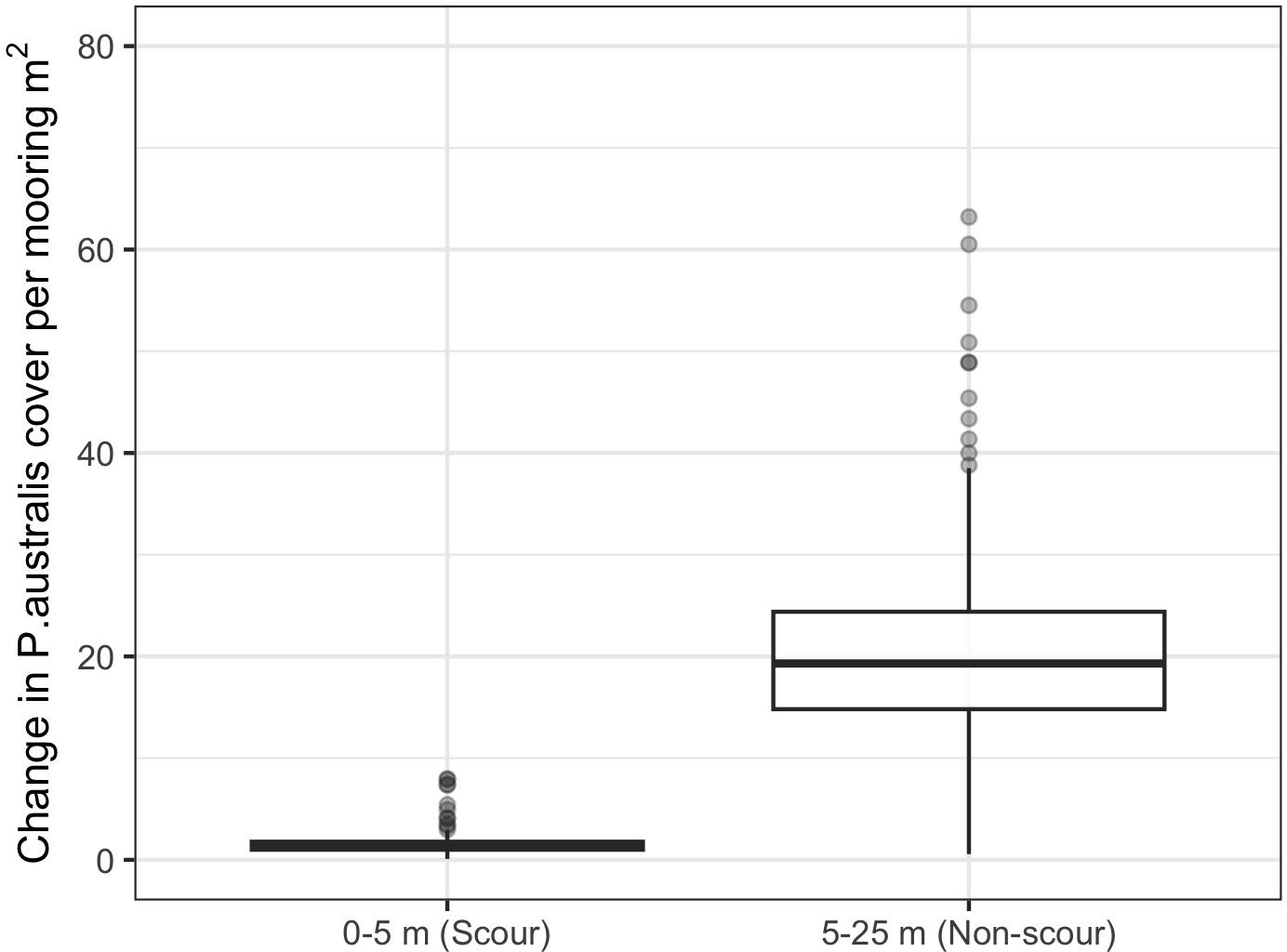
Figure 5. The predicted per-mooring change in cover of P. australis (m2 per mooring) in the scour zone (0-5 m from a mooring) and non-scour zone (5-25 m away) from the random replacement of 1-15 traditional moorings with EFMs.
Replacing all traditional moorings at Manly Cove with EFMs is predicted to increase P. australis cover by 164.2 m2 (+14.2% net change) while exchanging existing EFMs with traditional moorings is predicted to decrease the area covered with P. australis by 187.26 m2 (-16.2% net change) from the current area estimate of 1159.5 m2 (Figure 6). The frequency of high-cover areas of P. australis (>50%) is reduced under the traditional mooring scenario and increased under the EFM scenario. The simulations depend heavily on the present distribution of the seagrass, accounted for by the spatial random effect (autocorrelation), and this is representative of P. australis’ reliance on remnant shoots and rhizomes for natural recovery.
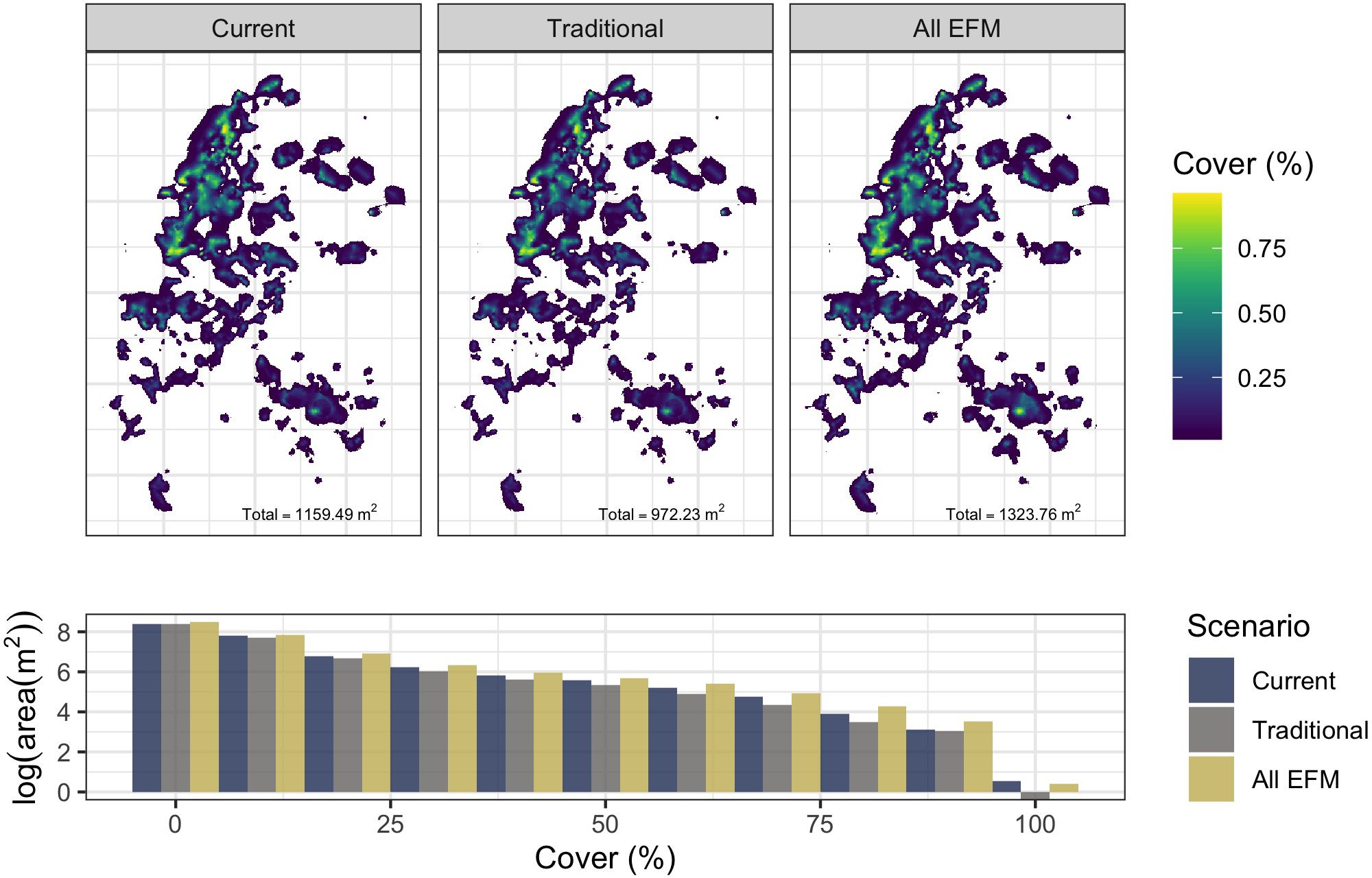
Figure 6. Upper pane: predicted cover of P. australis under current conditions (25/74 EFMs, left) and two scenarios: replacement of existing EFMs with traditional moorings (centre), and replacement of all traditional moorings with EFMs (right). The sum total area of predicted P. australis cover is shown in the lower right of each pane, with predictions lower than 0.01 excluded. Lower: frequency distribution of predicted P. australis cover scenarios.
Discussion
Our results suggest that in addition to boat moorings physically disturbing seagrasses, they also reduce seagrass cover in the surrounding meadow, which may reflect secondary disturbance processes. Furthermore, we found indications that reduced physical disturbance was associated with a concomitant improvement in the cover of the threatened seagrass Posidonia australis in the surrounding meadow. In areas subjected to direct disturbance, moorings designed to avoid contact with the seabed (EFMs) were associated with significantly higher seagrass cover than traditional ‘swing’ moorings. Potential impacts of secondary disturbance from moorings (i.e. impacts outside scour zones) were only detected for the threatened species Posidonia australis, a slow-growing seagrass with low recovery potential, while two faster-growing seagrasses, Zostera muelleri and Halophila ovalis showed no significant relationships with moorings outside of the scour zone. Mooring density was associated with lower P. australis cover, but this effect was reduced when most moorings were EFMs. We suggest that this amelioration of cumulative effects due to EFMs might reflect a reduction in the secondary disturbances that are contingent on physical scour (e.g. lower rates of sediment transport and edge effects) rather than by moored boats (e.g. shading, pollution).
Simulations found that the benefits of replacing traditional moorings with EFMs were greater in the non-scour than scour zones, increasing seagrass cover at the meadow scale. These results support the use of mooring designs that avoid physical disturbance to seagrasses, which in turn are likely to reduce secondary disturbances to the surrounding meadow. By modelling the density of seagrass, we were able to add details to estimates of P. australis at the site beyond what would be obtainable from satellite or aerial imagery. The sampling methodology was an example of extending traditional landscape ecology ideas (e.g. Walker and Walker, 1991) into underwater seascapes, and similar approaches may be more frequently or extensively conducted using hyperspectral remote sensing or automated vehicles in the future. This study demonstrates both the applicability of seascape ecology for impact identification and simulation; and emphasises the importance of identifying the key scales at which impacts might accumulate.
Bare circular clearings of vegetation are a characteristic signal of the impact of boat moorings in seagrass meadows, but this was the first effort to distinguish between direct physical disturbance from mooring gear and potential secondary disturbances due to habitat clearing (e.g. increased sediment transport, fragmentation and edge effects) or the moored vessels (e.g. shading). Studies of boat mooring impacts on seagrasses have previously quantified the extent of complete clearings by measuring the area of sandy vs vegetated seabed in aerial images (Glasby and West, 2018), diver-surveys (Demers et al., 2013), and/or camera-drops (Unsworth et al., 2017). These studies established relationships between the size of mooring scars and seagrass species, depth, mooring age, and vessel size. They determined that impacts from individual moorings scale up to cause substantial impacts at a regional scale. Glasby and West (2018) demonstrated that mooring scars in P. australis grew linearly after installation (over a 4.8-year period) despite no perceived change in the mooring equipment. We suggest that the expanding decline of seagrasses around moorings is partly due to secondary disturbance contingent upon the physical disturbance, and that these processes have significant implications for the persistence of seagrasses where traditional moorings are installed, especially in meadows of threatened P australis.
Species traits may explain the relative sensitivity of P. australis (c.f. Z. muelleri or H. ovalis) to the secondary disturbance from physical fragmentation by moorings and, in turn, the stronger response of P. australis when physical disturbance is reduced. Mooring scars in P. australis meadows are larger than those found in meadows of smaller, faster-growing species and expand faster over time (Glasby and West, 2018). Coloniser or early successional genera like Zostera, Heterozostera and Halophila are often interspersed within fragmented P. australis meadows and can be indicators of environmental stress (Irving et al., 2013). The study of human impacts in marine and terrestrial systems has benefitted from studies that identify how community dynamics can modify stressor responses (Johnston and Keough, 2003). Accordingly, more investigation is required to determine whether the patterns we observed are representative of monospecific meadows, especially for Z. muelleri and H. ovalis.
Apart from physical impacts, increasing mooring/boat density should result in less light reaching the seabed. Seagrasses are susceptible to major reductions in light - but the effects of chronic low-level shading are less well understood (Bulmer et al., 2016) and are likely to be both site and species-specific (Statton et al., 2018). Z. muelleri has been shown to have phenotypic plasticity in response to subtle changes in light conditions with negligible penalty (Manassa et al., 2017), whereas H. ovalis appears more sensitive to shading (Longstaff et al., 1999), and P. australis responses can vary based on genotypic diversity (Evans et al., 2017). These effects invite new considerations for both planning and conserving seagrasses and emphasise the need to factor in the cumulative effects of multiple primary and secondary stressors.
The implications of this study for the ongoing management of P. australis are clear - the physical disturbance of P. australis should be avoided, and non-contact designs such as EFMs should be used where seagrasses meadows exist to avoid creating clearings that are likely to expand. The density of moorings has a significant negative effect on P. australis and should not be overlooked as a source of long-term chronic decline. Our findings are likely to be conservative given that the study site was heavily impacted by a dense installation of traditional moorings and a legacy of disturbance associated with urbanisation. A hypothetical ‘pristine’ site is likely to benefit more from EFM designs than our study site by maintaining meadow contiguity, assuming moorings are necessary and traditional designs are the alternative option. The study site was chosen specifically as one of two remaining P. australis meadows in Sydney Harbour, from which we hoped to gain new insights into this species’ management (Johnston et al., 2015).
Variations in the effectiveness of non-contact mooring designs in reducing impacts on seagrasses found elsewhere indicate that further investigation and ongoing maintenance are needed (Hastings et al., 1995; Demers et al., 2013). Despite EFM density being positively associated with P. australis, the maximum density of EFMs we could assess was much less than that of traditional moorings and as such, it should be assumed that high densities of EFMs could also introduce smaller, but still detrimental effects for seagrasses. Further investigation is needed to allow us to generalise these patterns for other seagrasses, meadows, ‘non-contact’ mooring designs and impact sources, and to better understand the non-linear relationship between seagrass and mooring density. In scour areas, the lack of remnant seagrass is likely to make natural recovery slow, and active restoration, such as using beach-cast fragments, offers hope for recolonizing scour areas Ferretto et al., 2021).
There are many situations where environmental decision-making can be improved using data and simulation, in this case by assessing the effect of complex spatial arrangements of interacting stressors. Given the forecasted synergistic effects of climate change and urbanisation (Hughes et al., 2003; Mayer-Pinto et al., 2015), there is an urgent need to improve our understanding of factors driving coastal habitat loss and to implement effective mitigation. Most species continue to diminish after being listed as threatened, and whilst conservation efforts can reduce the loss of individual species, extinction has accelerated into the 21st century (Barnosky et al., 2011). An important aspect to combat this is the monitoring of fine-scale effects to avoid landscape-scale impacts (e.g. Romero et al., 2015), but many species/stressor combinations are yet to be investigated (Dafforn et al., 2016). Other sources of secondary impacts in marine systems (e.g. light or noise pollution) may have similar cumulative effects on species or communities (Bolton et al., 2017). In urbanised marine environments, multiple overlapping sources of pollutants or sediment input could have a similar spatially driven effect, along with other areas where human-made structures modify the marine environment (e.g. pylons, seawalls, and aquaculture installations) (Dafforn et al., 2015). When applied to the stabilisation and recovery of threatened species, simulations might also consider the use of rehabilitation and restoration techniques, especially where (as in the case of P. australis) the species has limited ability to recover naturally.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
KG: Formal analysis, Writing – review & editing, Project administration, Data curation, Writing – original draft, Software, Methodology, Investigation, Conceptualization. EJ: Conceptualization, Funding acquisition, Resources, Supervision, Writing – original draft, Writing – review & editing. AP: Conceptualization, Funding acquisition, Supervision, Writing – original draft, Writing – review & editing. AV: Conceptualization, Funding acquisition, Supervision, Writing – original draft, Writing – review & editing. GC: Conceptualization, Formal analysis, Investigation, Supervision, Validation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was funded by an ARC Linkage grant (LP140100855) awarded to EJ, AV and AP.
Acknowledgments
We thank Luke Hedge, Ruby Garthwin, Brendan Lanham, Aria Lee, and Samuel Burrell for assisting with field surveys.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer ES declared a past co-authorship with the author AV to the handling editor.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2024.1463637/full#supplementary-material
References
Althaus F., Hill N., Ferrari R., Edwards L., Przeslawski R., Schönberg C. H. L., et al. (2015). A standardised vocabulary for identifying benthic biota and substrata from underwater imagery: the CATAMI classification scheme. PloS One 10, e0141039. doi: 10.1371/journal.pone.0141039
Barnosky A. D., Matzke N., Tomiya S., Wogan G. O. U., Swartz B., Quental T. B., et al. (2011). Has the Earth’s sixth mass extinction already arrived? Nature 471, nature09678. doi: 10.1038/nature09678
Beijbom O., Edmunds P. J., Roelfsema C., Smith J., Kline D. I., Neal B. P., et al. (2015). Towards automated annotation of benthic survey images: variability of human experts and operational modes of automation. PloS One 10, e0130312. doi: 10.1371/journal.pone.0130312
Bolton D., Mayer-Pinto M., Clark G., Dafforn K., Brassil W., Becker A., et al. (2017). Coastal urban lighting has ecological consequences for multiple trophic levels under the sea. Sci. Total Environ. 576, 1–9. doi: 10.1016/j.scitotenv.2016.10.037
Brown C. J., Hamilton R. J. (2018). Estimating the footprint of pollution on coral reefs with models of species turnover. Conserv. Biol. 32, 949–958. doi: 10.1111/cobi.2018.32.issue-4
Bryars S., Rowling K. (2009). Benthic habitats of eastern gulf St Vincent: major changes in benthic cover and composition following European settlement of Adelaide. Trans. R. Soc. South Aust. 133, 318–338.
Bulmer R. H., Kelly S., Jeffs A. G. (2016). Light requirements of the seagrass, Zostera muelleri, determined by observations at the maximum depth limit in a temperate estuary, New Zealand. New Z. J. Mar. Freshw. Res. 50, 183–194. doi: 10.1080/00288330.2015.1120759
Clark G. F., Johnston E. L. (2016). “Coasts: Coasts,” in Australia State of the Environment (Australian Government Department of the Environment; Energy, Canberra).
Cullen-Unsworth L. C., Unsworth R. K. F. (2016). Strategies to enhance the resilience of the world’s seagrass meadows. J. Appl. Ecol. 53, 967–972. doi: 10.1111/1365-2664.12637
Dafforn K. A., Johnston E. L., Ferguson A., Humphrey C. L., Monk W., Nichols S. J., et al. (2016). Big data opportunities and challenges for assessing multiple stressors across scales in aquatic ecosystems. Mar. Freshw. Res. 67, 393–413. doi: 10.1071/MF15108
Dafforn K. A., Mayer-Pinto M., Morris R. L., Waltham N. J. (2015). Application of management tools to integrate ecological principles with the design of marine infrastructure. J. Environ. Manage. 158, 61–73. doi: 10.1016/j.jenvman.2015.05.001
Dakos V., Carpenter S. R., Brock W. A., Ellison A. M., Guttal V., Ives A. R., et al. (2012). Methods for detecting early warnings of critical transitions in time series illustrated using simulated ecological data. PloS One 7, e41010. doi: 10.1371/journal.pone.0041010
Demers M.-C. A., Davis A. R., Knott N. A. (2013). A comparison of the impact of ‘seagrass-friendly’ boat mooring systems on Posidonia australis. Mar. Environ. Res. 83, 54–62. doi: 10.1016/j.marenvres.2012.10.010
Duarte C. M. (2002). The future of seagrass meadows. Environ. Conserv. 29 (2), 192–206. doi: 10.1017/S0376892902000127
Dumbauld B. R., McCoy L. M. (2015). Effect of oyster aquaculture on seagrass Zostera marina at the estuarine landscape scale in Willapa Bay, Washington (USA). Aquacult. Environ. Interact. 7, 29–47. doi: 10.3354/aei00131
Elith J., Leathwick J. R. (2009). Species distribution models: ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. System. 40, 677–697. doi: 10.1146/annurev.ecolsys.110308.120159
Erftemeijer P. L. A., Robin Lewis R. R. III (2006). Environmental impacts of dredging on seagrasses: A review. Mar. pollut. Bull. 52, 1553–1572. doi: 10.1016/j.marpolbul.2006.09.006
Evans S. M., Griffin K. J., Blick R. A. J., Poore A. G. B., Vergés A. (2018). Seagrass on the brink: Decline of threatened seagrass Posidonia australis continues following protection. PloS One 13, e0190370. doi: 10.1371/journal.pone.0190370
Evans S. M., Vergés A., Poore A. G. B. (2017). Genotypic diversity and short-term response to shading stress in a threatened seagrass: does low diversity mean low resilience? Front. Plant Sci. 8. doi: 10.3389/fpls.2017.01417
Ferretto G., Callaghan C. T., Glasby T. M., Housefield G. P., Kendrick G. A., Langley M., et al. (2021). Naturally-detached fragments of the endangered seagrass Posidonia australis collected by citizen scientists can be used to successfully restore fragmented meadows. Biol. Conserv. 262, 109308. doi: 10.1016/j.biocon.2021.109308
Field S. A., O’connor P. J., Tyre A. J., Possingham H. P. (2007). Making monitoring meaningful. Austral Ecol. 5, 485–491. doi: 10.1111/j.1442-9993.2007.01715.x
Frost M. T., Rowden A. A., Attrill M. J. (1999). Effect of habitat fragmentation on the macroinvertebrate infaunal communities associated with the seagrass Zostera marina L. Aquat. Conserv.: Mar. Freshw. Ecosyst. 9, 255–263. doi: 10.1002/(SICI)1099-0755(199905/06)9:3<255::AID-AQC346>3.0.CO;2-F
Geoscience Australia. (2004). GEODATA COAST 100K 2004. Available at: https://ecat.ga.gov.au/geonetwork/srv/api/records/a05f7892-eae3-7506-e044-00144fdd4fa6.
Glasby T. M., West G. (2015). Estimating losses of Posidonia australis due to boat moorings in Lake Macquarie, Port Stephens and Wallis Lake, Vol. 147. doi: 10.1002/aqc.2872
Glasby T. M., West G. (2018). Dragging the chain: Quantifying continued losses of seagrasses from boat moorings. Aquat. Conserv.: Mar. Freshw. Ecosyst. 28, 383–394. doi: 10.1002/aqc.2872
Griffin K. J., Hedge L. H., González-Rivero M., Hoegh-Guldberg O. I., Johnston E. L. (2017). An evaluation of semi-automated methods for collecting ecosystem-level data in temperate marine systems. Ecol. Evol. 7, 4640–4650. doi: 10.1002/ece3.2017.7.issue-13
Hastings K., Hesp P., Kendrick G. A. (1995). Seagrass loss associated with boat moorings at Rottnest Island, Western Australia. Ocean Coast. Manage. 26, 225–246. doi: 10.1016/0964-5691(95)00012-Q
Hill N. A., Lucieer V., Barrett N. S., Anderson T. J., Williams S. B. (2014). Filling the gaps: Predicting the distribution of temperate reef biota using high resolution biological and acoustic data. Estuarine Coast. Shelf Sci. 147, 137–147. doi: 10.1016/j.ecss.2014.05.019
Hughes T. P., Baird A. H., Bellwood D. R., Card M., Connolly S. R., Folke C., et al. (2003). Climate change, human impacts, and the resilience of coral reefs. Science 301, 929–933. doi: 10.1126/science.1085046
Irving A. D., Tanner J. E., Gaylard S. G. (2013). An integrative method for the evaluation, monitoring, and comparison of seagrass habitat structure. Mar. pollut. Bull. 66, 176–184. doi: 10.1016/j.marpolbul.2012.10.017
Jia Y., Shelhamer E., Donahue J., Karayev S., Long J., Girshick R., et al. (2014). “Caffe: convolutional architecture for fast feature embedding,” in Proceedings of the 22Nd ACM International Conference on Multimedia MM ‘14, New York, NY, USA. 675–678 (ACM). Available at: https://dl.acm.org/doi/10.1145/2647868.2654889.
Johnston E. L., Keough M. J. (2003). Competition modifies the response of organisms to toxic disturbance. Mar. Ecol. Prog. Ser. 251, 15–26. doi: 10.3354/meps251015
Johnston E. L., Mayer-Pinto M., Hutchings P. A., Marzinelli E. M., Ahyong S. T., Birch G., et al. (2015). Sydney Harbour: What we do and do not know about a highly diverse estuary. Mar. Freshw. Res. 66, 1073–1087. doi: 10.1071/MF15159
Kéfi S., Guttal V., Brock W. A., Carpenter S. R., Ellison A. M., Livina V. N., et al. (2014). Early warning signals of ecological transitions: methods for spatial patterns. PloS One 9, e92097. doi: 10.1371/journal.pone.0092097
Kirkman H. (1985). Community structure in seagrasses in southern western Australia. Aquat. Bot. 21, 363–375. doi: 10.1016/0304-3770(85)90077-4
Kohler K. E., Gill S. M. (2006). Coral Point Count with Excel extensions (CPCe): A Visual Basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 32, 1259–1269. doi: 10.1016/j.cageo.2005.11.009
Krainski E. T., Gómez-Rubio V., Bakka H., Lenzi A., Castro-Camilo D., Simpson D., et al. (2018). Advanced Spatial Modeling with Stochastic Partial Differential Equations Using R and INLA (New York: Chapman; Hall/CRC).
Laurance W. F., Yensen E. (1991). Predicting the impacts of edge effects in fragmented habitats. Biol. Conserv. 55, 77–92. doi: 10.1016/0006-3207(91)90006-U
Lindgren F., Rue H. (2015). Bayesian spatial modelling with R-INLA. J. Stat. Softw. 63, 1–25. doi: 10.18637/jss.v063.i19
Lindgren F., Rue H., Lindström J. (2011). An explicit link between Gaussian fields and Gaussian Markov random fields: The stochastic partial differential equation approach. J. R. Stat. Soc.: Ser. B (Statistical Methodology) 73, 423–498. doi: 10.1111/j.1467-9868.2011.00777.x
Longstaff B. J., Loneragan N. R., O’Donohue M. J., Dennison W. C. (1999). Effects of light deprivation on the survival and recovery of the seagrass Halophila ovalis (R.Br.) Hook. J. Exp. Mar. Biol. Ecol. 234, 1–27. doi: 10.1016/S0022-0981(98)00137-3
Manassa R. P., Smith T. M., Beardall J., Keough M. J., Cook P. L. M. (2017). Capacity of a temperate intertidal seagrass species to tolerate changing environmental conditions: Significance of light and tidal exposure. Ecol. Indic. 81, 578–586. doi: 10.1016/j.ecolind.2017.04.056
Martin R. W., Waits E. R., Nietch C. T. (2018). Empirically-based modeling and mapping to consider the co-occurrence of ecological receptors and stressors. Sci. Total Environ. 613-614, 1228–1239. doi: 10.1016/j.scitotenv.2017.08.301
Maxwell P. S., Eklöf J. S., van Katwijk M. M., O’Brien K. R., de la Torre-Castro M., Boström C., et al. (2017). The fundamental role of ecological feedback mechanisms for the adaptive management of seagrass ecosystems - a review. Biol. Rev. Cambridge Philos. Soc. 92, 1521–1538. doi: 10.1111/brv.2017.92.issue-3
Mayer-Pinto M., Johnston E. L., Hutchings P. A., Marzinelli E. M., Ahyong S. T., Birch G., et al. (2015). Sydney Harbour: A review of anthropogenic impacts on the biodiversity and ecosystem function of one of the world’s largest natural harbours. Mar. Freshw. Res. 66, 1088–1105. doi: 10.1071/MF15157
Meehan A. J., West R. J. (2000). Recovery times for a damaged Posidonia australis bed in south eastern Australia. Aquat. Bot. 67, 161–167. doi: 10.1016/S0304-3770(99)00097-2
Murcia C. (1995). Edge effects in fragmented forests: Implications for conservation. Trends Ecol. Evol. 10, 58–62. doi: 10.1016/S0169-5347(00)88977-6
Ojima D. S., Galvin K. A., Turner B. L. (1994). The global impact of land-use change. BioScience 44, 300–304. doi: 10.2307/1312379
Paling E. I., Van Keulen M., Tunbridge D. J. (2007). Seagrass Transplanting in Cockburn Sound, Western Australia: A Comparison of Manual Transplantation Methodology Using Posidonia sinuosa Cambridge et Kuo. Restor. Ecol. 15, 240–249. doi: 10.1111/j.1526-100X.2007.00207.x
Perkins N. R., Foster S. D., Hill N. A., Barrett N. S. (2016). Image subsampling and point scoring approaches for large-scale marine benthic monitoring programs. Estuarine Coast. Shelf Sci. 176, 36–46. doi: 10.1016/j.ecss.2016.04.005
Pressey R. L. (2004). Conservation planning and biodiversity: assembling the best data for the job. Conserv. Biol. 18, 1677–1681. doi: 10.1111/j.1523-1739.2004.00434.x
Romero J., Alcoverro T., Roca G., Pérez M. (2015). “Bioindicators, Monitoring, and Management Using Mediterranean Seagrasses: What Have We Learned from the Implementation of the EU Water Framework Directive?,” in Experiences from Ground, Coastal and Transitional Water Quality Monitoring The Handbook of Environmental Chemistry (Springer, Cham), 161–182.
Rue H., Martino S., Chopin N. (2009). Approximate Bayesian inference for latent Gaussian models by using integrated nested Laplace approximations. J. R. Stat. society: Ser. b (statistical methodology) 71, 319–392. doi: 10.1111/j.1467-9868.2008.00700.x
Schultz S. T., Kruschel C., Bakran-Petricioli T., Petricioli D. (2015). Error, power, and blind sentinels: the statistics of seagrass monitoring. PloS One 10, 9. doi: 10.1371/journal.pone.0138378
Statton J., McMahon K., Lavery P., Kendrick G. A. (2018). Determining light stress responses for a tropical multi-species seagrass assemblage. Mar. pollut. Bull. 128, 508–518. doi: 10.1016/j.marpolbul.2018.01.060
Tuya F., Espino F., Terrados J. (2013). Preservation of seagrass clonal integration buffers against burial stress. J. Exp. Mar. Biol. Ecol. 439, 42–46. doi: 10.1016/j.jembe.2012.10.015
Unsworth R. K. F., Williams B., Jones B. L., Cullen-Unsworth L. C. (2017). Rocking the boat: damage to eelgrass by swinging boat moorings. Front. Plant Sci. 8. doi: 10.3389/fpls.2017.01309
Walker D. A., Walker M. D. (1991). History and pattern of disturbance in Alaskan arctic terrestrial ecosystems: A hierarchical approach to analysing landscape change. J. Appl. Ecol. 28, 244–276. doi: 10.2307/2404128
Walker D., Webber P., Binnian E. F., Everett K., Lederer N., Nordstrand E., et al. (1987). Cumulative impacts of oil fields on northern Alaskan landscapes. Science 238, 757–761. doi: 10.1126/science.238.4828.757
Keywords: conservation decision making, cumulative impact, seagrass mapping, secondary effects, simulation, spatial modelling, Posidonia australis, seascape ecology
Citation: Griffin KJ, Johnston EL, Poore AGB, Vergés A and Clark GF (2024) Reducing direct physical disturbance also mitigates hidden drivers of decline in a threatened seagrass meadow. Front. Conserv. Sci. 5:1463637. doi: 10.3389/fcosc.2024.1463637
Received: 12 July 2024; Accepted: 01 November 2024;
Published: 02 December 2024.
Edited by:
Sally Joan Watson, National Institute of Water and Atmospheric Research (NIWA), New ZealandReviewed by:
Michael Sievers, Griffith University, AustraliaElisabeth Marijke Anne Strain, University of Tasmania, Australia
Copyright © 2024 Griffin, Johnston, Poore, Vergés and Clark. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kingsley J. Griffin, a2luZ3NsZXlnODhAZ21haWwuY29t