 Pavla Hejcmanová1
Pavla Hejcmanová1 Kateřina Gašparová1
Kateřina Gašparová1 Tamara Fedorova1
Tamara Fedorova1 Mara Vukelić1
Mara Vukelić1 Julian Fennessy2,3Abdoul Razack Moussa Zabeirou1,2,4Thomas Rabeil5
Julian Fennessy2,3Abdoul Razack Moussa Zabeirou1,2,4Thomas Rabeil5 Karolína Brandlová1*
Karolína Brandlová1*- 1Department of Animal Science and Food Processing, Czech University of Life Sciences Prague, Prague, Czechia
- 2Giraffe Conservation Foundation, Windhoek, Namibia
- 3School of Biology and Environmental Science, University College Dublin, Dublin, Ireland
- 4Sahara Conservation, Saint-Maur-des-Fossés, France
- 5Wild Africa Conservation, Niamey, Niger
Resting is an integral component of animal behavior, contributing to one’s fitness through careful optimization strategies. In large herbivores, resting periods are driven by availability of food, presence of predators, and thermoregulation. A combination of these drivers leads to high variability in resting behaviors and their time allocation throughout the day. However, these drivers are rarely evaluated in the wild. Megaherbivores, including giraffe (Giraffa spp.), adopt social resting strategies, which enable them to optimize the cost–benefit ratio, with rest and vigilance varying with group size and composition. We investigated resting behavior of the West African giraffe (G. camelopardalis peralta) living in a human-populated landscape dominated by agropastoralism activities in Niger. Through direct observation, we evaluated the influence of group size and composition, and presence of livestock and humans on giraffe resting behavior. We concluded that giraffe increased their resting time with shorter distance to other giraffe and livestock. Livestock did not negatively impact giraffe behavior; rather, they provided a kind of “safe environment”. Human presence resulted in only minor changes in vigilance of giraffe and did not significantly affect their resting time. Our findings highlight a positive instance of human–giraffe coexistence in a human-dominated landscape, attributed to the long-term benefits of conservation efforts.
Introduction
Resting is an integral and vital part of an animal behavioral state and rhythm (Siegel, 2008). Resting behaviors vary, from inactivity to sleeping, each having multiple essential functions reflecting an animal’s actual ecological and physiological demands (Dallaire, 1986; Craig et al., 2016; Rattenborg et al., 2017). Ultimately, the primary purpose of resting is to manage and strategically allocate energy to enhance reproductive success (Schmidt, 2014). The impact of resting or not resting on an individual’s fitness is varied, making the allocation of resting time a critical decision that involves both benefits and risks. Such decisions require careful optimization strategies, accounting for timing, location with actual environmental, i.e., ecological and human-related context, and duration (Shukla et al., 2021).
The allocation of time between active and resting behaviors is driven by trade-offs between three major needs, i.e., satisfying nutritional requirements through securing food, evading predators, and coping with heat load (Owen-Smith and Goodall, 2014; Mole et al., 2016; Vermeulen et al., 2024). This balance is shaped by the individual’s site- and time-specific context. For example, the search for food and the time spent foraging fluctuate with seasonal variation in forage abundance, quality, and environmental heterogeneity. However, the behavioral activity cycles of large herbivores, particularly ruminants, are constrained by the digestive passage rates, which force animals into periods of physical inactivity to process the food (Hirakawa, 1997; Jeschke and Tollrian, 2005). Animals therefore exhibit more significant plasticity in time allocation for antipredator behavior, enabling them to switch between foraging, active vigilance, and inactivity, depending on the level of perceived risk, as proposed by risk allocation hypothesis (Lima and Bednekoff, 1999; Luttbeg, 2017). While resting behavior and sleep of large mammals vary considerably (e.g., Duggan et al., 2016; Burger et al., 2020b), studies in the wild are limited (e.g., Siegel, 2008; Burger et al., 2020a). The presence, abundance, and behavior of predators strongly influence these patterns. For instance, in regions with nocturnal predators, prey species remain active, avoiding predator-frequented habitats, and rest during the daylight hours (e.g. Beekman and Prins, 1989; Fischhoff et al., 2007; Owen-Smith and Goodall, 2014; Owen-Smith and Traill, 2017). To reduce vulnerability, megaherbivores often adopt a social resting strategy, gathering in groups with conspecifics or other prey species, which provides protection and conserves energy (Shukla et al., 2021).
Giraffe (Giraffa spp.) are megaherbivores with a social resting strategy (Shukla et al., 2021), living in multilevel fission–fusion social systems with fluctuating group size modulated by social preferences (Gloneková et al., 2021; Carter et al., 2013; Burger et al., 2020b). Many herbivore species allocate less time to vigilance and more to resting with increasing group size and higher risk of predation (Creel et al., 2014; Beauchamp et al., 2021). A giraffe’s group size may not primarily be driven by not only predation avoidance but also other factors such as habitat and presence of calves (Muller et al., 2018). For instance, giraffe allocation of time to vigilance is reported to be greater in woodland environments (Marealle et al., 2020). Additionally, Marealle et al. (2020) observed that both presence of calves and predation risk increased the proportion of vigilant individuals and that the proportion of vigilant individuals decreased with an increase in group size. Moreover, the presence of a big bull giraffe influenced vigilance significantly with male vigilance decreasing as they invested more in mating, whereas females remained vigilant. The vigilance in giraffe therefore seems to also be influenced by presence of specific herd members (Cameron and du Toit, 2005).
Fear from predators is one of the strongest determinants of resting and vigilant behaviors with direct impacts on social structure, and ultimately fitness. With regard to wildlife, humans are “super predators” across the globe; hence, their presence can incite more fear than other predators and may have stronger effects on behavior and vigilance (Ciuti et al., 2012; Zbyryt et al., 2018; Zanette et al., 2023). Intensive human presence in a landscape can be of conservation concern as it can lead to disturbances in wildlife activity patterns, and increased levels of stress and energy expenditures (e.g., Jayakody et al., 2008; Gaynor et al., 2018; Ripari et al., 2022; Tucker et al., 2023), leading to reduced animal fitness and altered population dynamics. Wildlife often responds to anthropogenic disturbance by changing space use patterns (Chen and Koprowski, 2015), allocating more time to vigilance (Scheijen et al., 2021), or shifting specific activities to the night. Specifically, intensifying land use through livestock grazing in wildlife habitats forces them to change their ecological niche and shift their spatiotemporal use of the ecosystem by, e.g., contraction of home ranges, decreasing animal movement, or becoming more nocturnal (Loft et al., 1991; Scholte et al., 2022; Stabach et al., 2022; Brown et al., 2023). However, wildlife–livestock interactions seem to be predominantly indirect, with direct interactions requiring increasing investigation.
The West African giraffe (G. camelopardalis peralta), historically native throughout the Sudano-Sahelian belt in West Africa, now almost exclusively persist in an open, human-inhabited landscape in Niger, called the “Giraffe Zone,” and in a small, recently established population in the Gadabedji Biosphere Reserve, eastern Niger (Gašparová et al., 2024). Although poaching, habitat loss, degradation, and fragmentation were historically the main causes of their decline, the situation has changed significantly in recent decades. Today, despite the close human–giraffe coexistence, human–giraffe conflict is minimal thanks to the long-term conservation efforts of the Nigerian government, local communities, and NGOs (Gašparová et al., 2023). The effect of such coexistence in the “Giraffe Zone” has only peripherally been evaluated. In addition, the potential risks of pathogen transmissions (Gašparová et al., 2020), livestock may present an indirect disturbance for the giraffe. Accompanied by pastoralists and inhabiting areas near human settlements, giraffe may experience reduce forage due to wood cutting and limited crop production by local communities (Bond et al., 2021). However, the presence of livestock may also benefit the giraffe as there is no natural predators present, aside from humans (Bond et al., 2019). Mixed species groups of herbivores are common across taxa and habitats, allowing prey species to benefit from the presence of other animals by decreasing individual vigilance and increasing foraging time in areas with predators (Stensland et al., 2003). The West African giraffe therefore represents an ideal model to investigate the influence of livestock, human, and other disturbances on resting behavior of megaherbivores in a densely inhabited rural area.
Our study aimed to disentangle the patterns of West African giraffe resting within diurnal (daylight) activities in a human-dominated landscape and to explore whether the presence of livestock and people modulate the time they allocate to active or resting behaviors. Specifically, we hypothesized that presence of another giraffe or livestock does not affect, or can even increase, the resting behavior of an individual giraffe by creating a “group effect” (as suggested by Muller et al., 2018), which acts as a “detection and dilution effect” against potential predators (Stensland et al., 2003; Makenbach et al., 2013). In contrast, human presence may have disturbing and fear-inducing effects on an individual. We first identified the group size and composition of each giraffe herd and observed their activity time budgets, i.e., the total allocation and behavior durations according to time of day, and social category, i.e., sex and age. For activity patterns, we particularly focused on resting behaviors, i.e., laying, calm standing, and ruminating. Furthermore, we tested whether the active and resting behavior of an individual giraffe was impacted with the presence of conspecifics, livestock, and people. Finally, we tested the effect of group size and distance of conspecifics, livestock, and people on giraffe activity and resting time.
Materials and methods
Study site
The “Giraffe Zone” is an unofficial protected area situated across the central plateaus of Kouré and North Dallol Bosso, ca. 60 km southeast of the capital Niamey (Figure 1). The area is densely populated with people (49/km2), and habitat overlaps with pastoralists and their primarily subsistent agriculture activities and livestock (goat, sheep, and cattle) (Brown et al., 2023).

Figure 1. Map of the study site: (A) illustrative location of the ‘Giraffe Zone’ in Niger, and (B) positions of West African giraffe observations within the ‘Giraffe Zone’.
Data collection
To assess West African giraffe activity and their response to people and livestock presence, we observed them over a 2-month period in the dry season (January to March 2020). In this period, the average diurnal temperatures were 31.2°C (January), 34.7°C (February), and 38.9°C (March, Worlddata.info, 2024). Direct observations of giraffe were conducted using binoculars during daylight hours from 08:00 to 18:00 using CyberTracker®. Furthermore, we divided each day into three periods: morning (8:00 to 11:00), midday (11:01 to 15:00), and evening (15:01 to 18:00).
Upon spotting a giraffe herd, data collection was initiated. The observer (KG) recorded her own GPS position, number of giraffe in the herd, and their individual sex and age class. Distance and absolute angle of observer to each individual giraffe in the group, livestock, people, and house, if any, was then measured using a Rangefinder laser device (Nikon Monarch 3000 Stabilized). One giraffe within the herd was then assigned as a focal animal, and the distances were measured from that focal individual to herd members, livestock, people, and houses. The distance between focal giraffe to other giraffe, livestock, people, and house was then calculated with the help of basic trigonometric function. After that, the focal individual was observed for 20 min and all activities and their durations were recorded. When the focal sampling was finished, we repeated measuring the distance and angle between the focal individual and all other giraffe, livestock, people, and houses. Following this, another giraffe in the herd was selected for focal sampling. The recorded activities included feeding, walking, vigilance, and social interactions as active behaviors, whereas resting behavior included standing still and laying on ground, both with and without ruminating. According to the definition of Suscke et al. (2021), “vigilance” was recorded when individuals were performing an activity but constantly turning their heads around to monitor conspecifics, observers, or their surroundings.
In total, more than 300 giraffe in 54 groups (1–21 individuals) were recorded. For the data analyses, giraffe were classified into three categories: adult-size male, adult-size female, and calf/juvenile of any sex. We undertook 143 focal samplings and measured 521 distances between giraffe and other giraffe, livestock, people, and houses, with a total time of almost 48 h during 27 days (1–9 per day). The extent of the study was considerably smaller than originally planned due to safety and security restrictions imposed after a terrorist attack in the “Giraffe Zone” in May 2020 and subsequently by the COVID-19 pandemic.
Data analysis
For analyses, detailed activity categories were divided into active and resting behavior. First, we calculated the proportion of behaviors from data pooled across the data set and visualized them according to daytime and sex-age categories. Then, to assess the length of behavior bouts, we tested differences of activity duration among sex-age categories and across the daytime (separately) using Kruskal–Wallis test followed by pairwise comparisons of Wilcoxon rank sum test with Bonferroni correction.
To test the effects of group size and composition, and distance from another giraffe, livestock (cattle, sheep/goat), people, and houses on the activity of the focal giraffe (active versus resting coded as 1 and 0, respectively), we applied the logistic regression approach with logit link function, assessing models by AIC criterion, Tjur’s R squared coefficient of discrimination, and level of significance 0.05. All the analyses were performed in R 4.3.3 version using R base, tidyverse, lme4, MASS, tidy models, sjplot, and cowplot packages (R Core Team, 2024).
Results
Giraffe group size and composition
We observed a total of 54 independent events. Giraffe were mostly in herds (n = 45; 83.3%) ranging from 2 to 21 animals, with two (3.7%) and seven (13%) observations of a single adult male and female, respectively. The overall median herd size was four individuals. Males were present in 22 (48.8%) giraffe herds. Herds with males were larger (median size = 8) compared with herds without males (median size = 4). Calves were present in 28 (51.9%) giraffe herds with a median herd size of six, compared with herds without calf with a median size of three. Mixed herds consisting of females, males, and calves had a median size of 10.
Giraffe diurnal activity
Giraffe were mostly active during the day, spending 75% (n = 333) of their time in active behaviors compared with 25% (n = 97) of time resting (Figure 2). The proportion of active and resting behaviors was similar across sex and age categories, with slightly prolonged active behaviors in the evening (Figure 3).
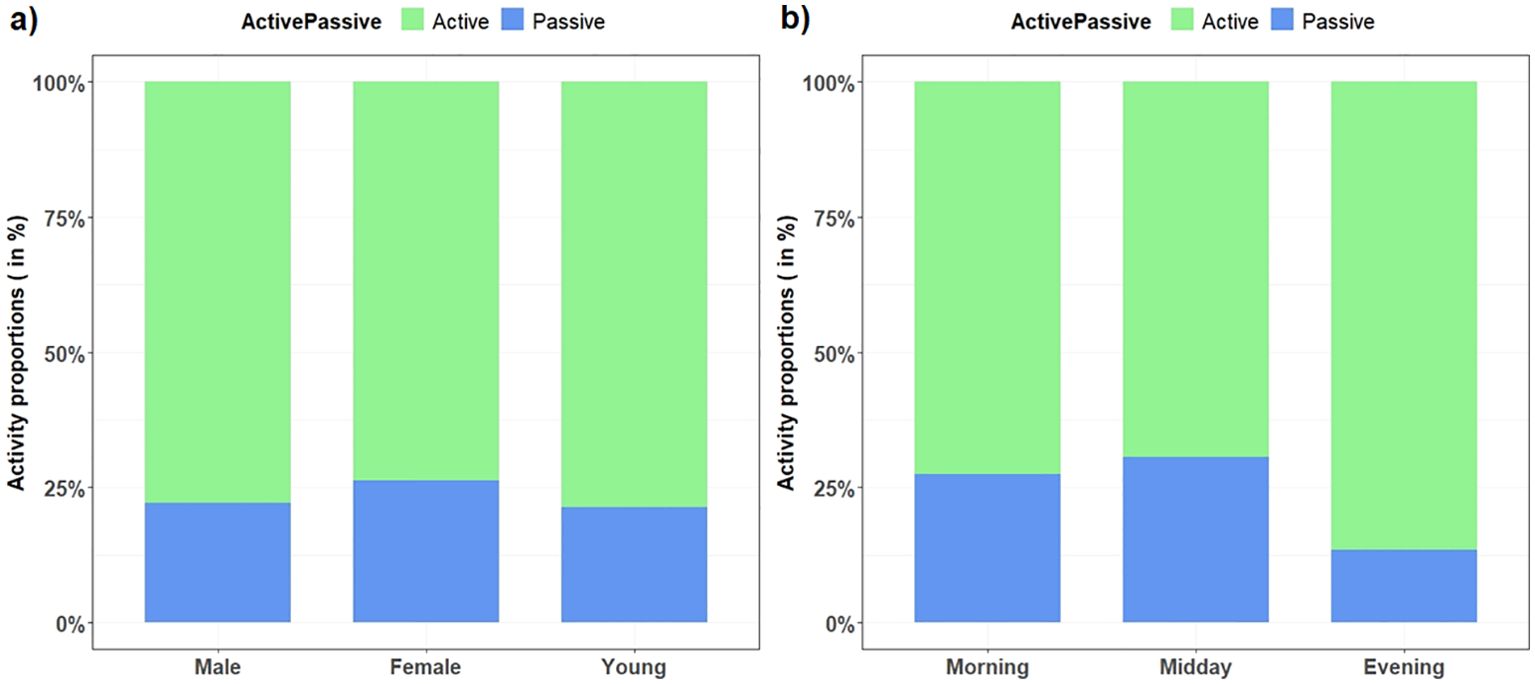
Figure 2. Proportions of active and resting behaviors of West African giraffe in the ‘Giraffe Zone’, Niger divided by (A) sex and age categories, and (B) time of day.
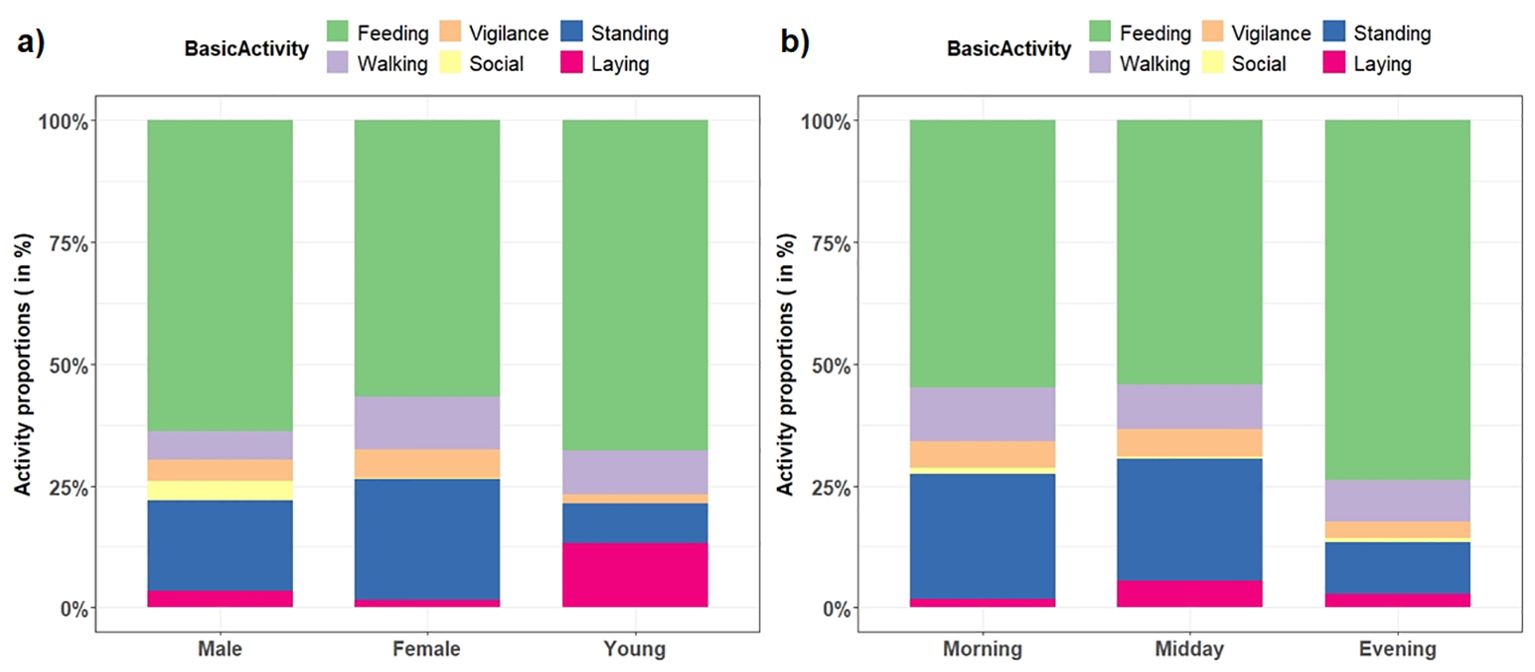
Figure 3. Proportions of basic activities of West African giraffe in the ‘Giraffe Zone’, Niger divided by (A) sex and age categories, and (B) time of day.
Giraffe allocated most of the daylight time to feeding (59.7%), followed by standing (21.3%), walking (9.6%), vigilance (5%), laying (3.5%), and social interactions (0.9%) (see Figure 2 for proportions of total time allocated to activities across sex and age categories and time of day). On average, laying was infrequent but observed for 13.3 min, and feeding for 7.7 min. Vigilance and walking during the focal bouts were significantly shorter than other activities (Kruskal–Wallis test, chi-sq = 79, df=5, p < 0.001) (see Table 1 for mean duration of activities). The mean duration of activities was similar for sex and age categories (all p > 0.05, Kruskal–Wallis tests) and for time of day, except for feeding which was greater in the evening compared with the morning (Kruskal–Wallis test, chi-sq= 10, df=2, p = 0.006) (Figure 4).
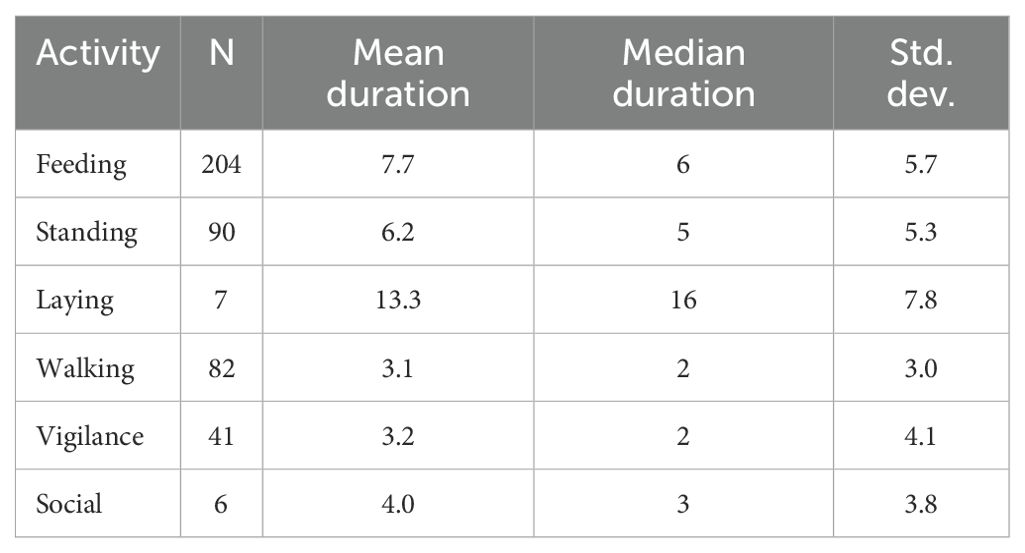
Table 1. Number (N) and duration (mean duration and median duration) of basic diurnal activities (in minutes) of West African giraffe in the ‘Giraffe Zone’, Niger.
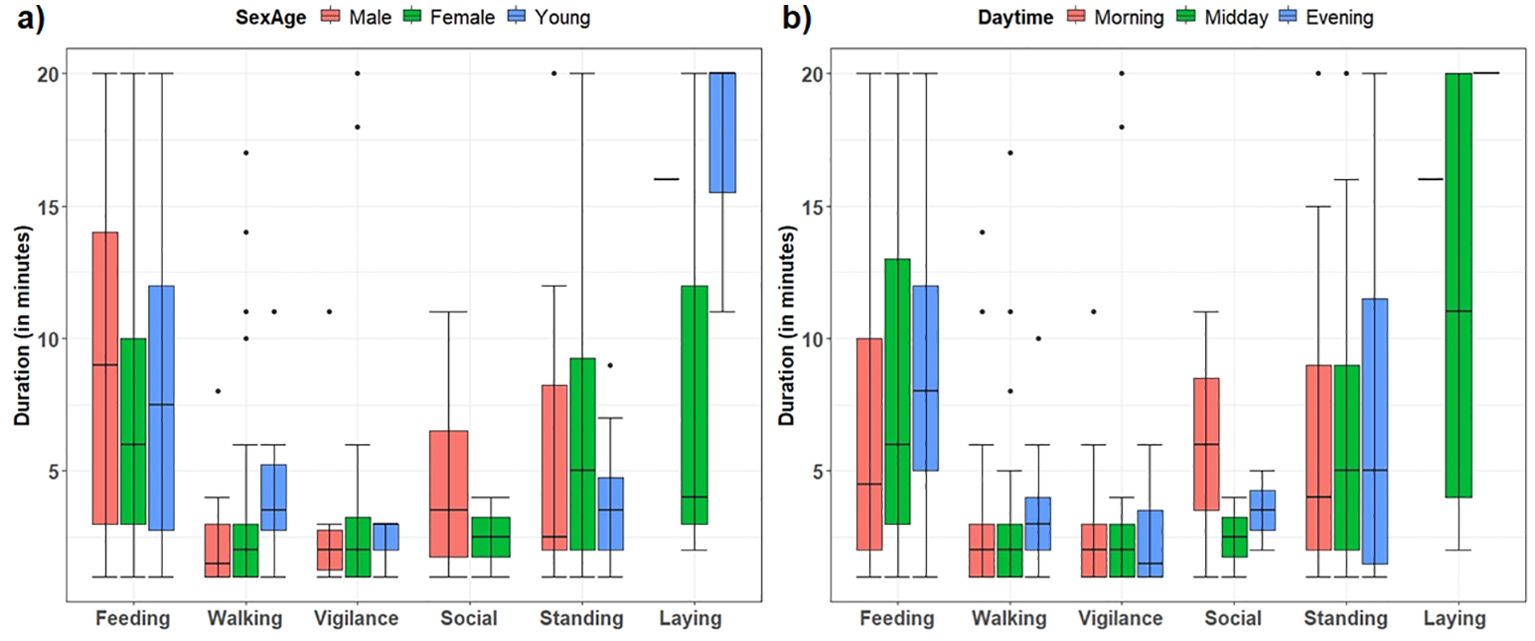
Figure 4. Duration of behavioral activity bouts (in minutes) behaviors of West African giraffe in the ‘Giraffe Zone’, Niger, divided by (A) sex and age categories, and (B) time of day. Line in the box indicates median, and error bars are minimum (Q1-1.5*IQR) and maximum (Q3 + 1.5*IQR).
Giraffe response to animals and people
Giraffe were seen in the presence of other giraffe only (i.e., no livestock, nor people) in 42% of observations at an average distance to the other giraffe of 57.7 m (SE ± 2.9 m, range 1–259 m). In 23% of observations, giraffe were observed in the presence of sheep/goat at an average distance of 82.3 m (SE ± 4.8 m, range 20–251 m), and in 15% of cases in the presence of cattle at an average distance of 109.8 m (SE ± 11.2 m, range 11–360 m). Giraffe were observed also in the presence of people (14%) and their houses (9%) at an average distance of 95.2 m (SE ± 7.4 m) and 220 m (SE ± 12.4 m), respectively.
Giraffe only spent a small portion of daytime resting. We observed a tendency for them to rest more when in the presence of other herbivores, whether these were other giraffe or livestock. In contrast, their activity increased as the distance from these animals grew. Specifically, the probability of a giraffe being active significantly increased with the distance from another giraffe (p = 0.03), and similar tendency (p < 0.1) was recorded for the distance to cattle and sheep/goat (Table 2, Figures 5A, B). The giraffe herd size had no effect on the time spent being active or resting.
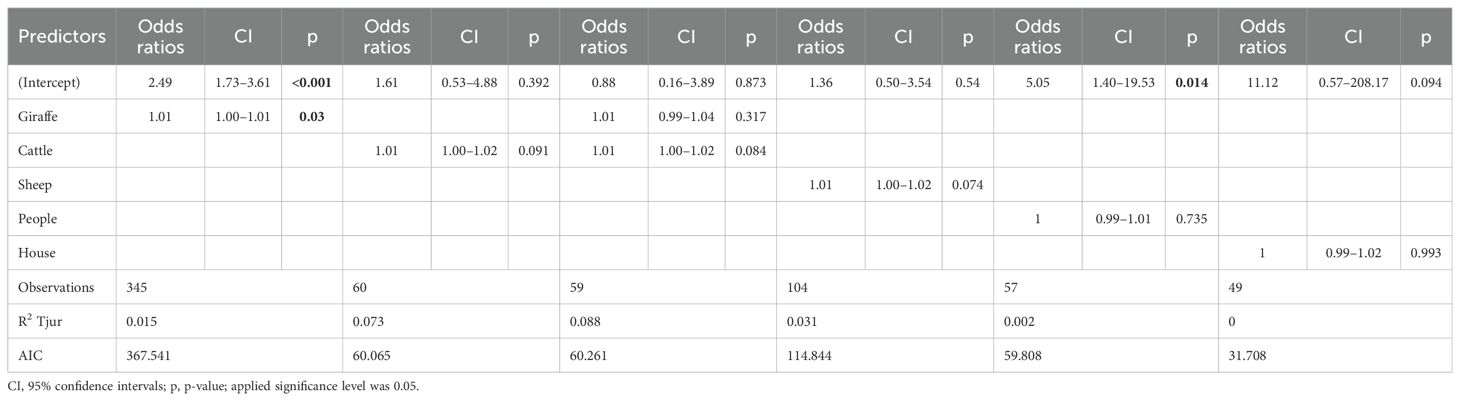
Table 2. Regression models showing the effects of distance of another giraffe, domesticated animals (cattle, sheep/goat), people (pastoralists, women and kids walking outside of village), and human structures on West African giraffe activity in the ‘Giraffe Zone’, Niger.
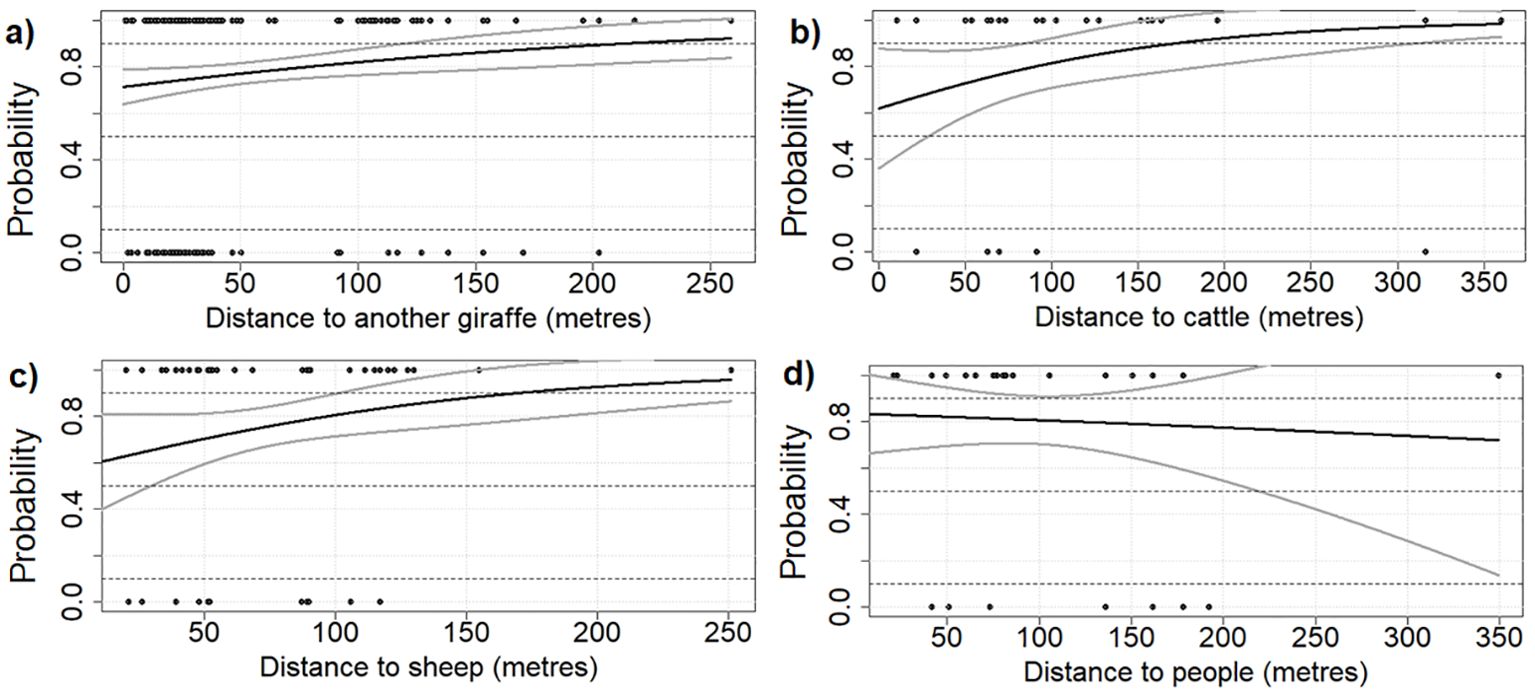
Figure 5. Probability of active (1) and resting (0) behavior of West African giraffe in the ‘Giraffe Zone’, Niger in response to distance to another animals: (A) giraffe, (B) cattle, (C) sheep/goat), and (D) people.
There was no effect of giraffe distance to people and their houses (p > 0.05, Table 2, Figures 5C, D). There were, however, interesting anecdotal observations. On the rare occasion, we observed that giraffe became more vigilant in the presence of people, especially if they carried a wooden stick, which is common among the herders. On one occasion, we observed that giraffe were vigilant toward women with postharvest remnants of millet which looked like a wooden stick. Regarding interactions between giraffe and livestock, we observed one instance of a giraffe approaching a tree with a donkey tied to it. The donkey appeared afraid and tried to escape. When the giraffe saw that the donkey was not moving away, it changed direction and moved off to another tree. Lastly, we recorded giraffe on one occasion feeding together with sheep on branches cut by people and pods of Faidherbia albida on the ground.
Discussion
We investigated the diurnal activity and resting behaviors of West African giraffe in an agropastoral human-dominated landscape. Our findings suggest that giraffe were not disturbed by the presence of livestock and, in fact, preferred the presence of other animals—whether conspecifics or domestic animals—when resting. They remained highly active during the day, and they exhibited increased resting behaviors in proximity to livestock or other giraffe.
The social behavior and population structure of giraffe have increasingly been studied since the early 2000s (Muller and Harris, 2021). While we did not primarily focus on their social structure, herd size was considered an important aspect in assessing their resting behavior. We predominantly observed smaller herds, similar to that of Kordofan giraffe (G. c. antiquorum) in the DRC (D’haen et al., 2019) and in contrast to larger Nubian giraffe (G. c. camelopardalis) herds in Kenya (n = 5–8) (Muller et al., 2018), largely attributed to habitat type. Herd sizes of the West African giraffe vary seasonally, notably smaller during the dry season (mean = 6, median = 4 individuals) compared with the rainy season (mean = 9; Le Pendu et al., 2000). The low number of lone males’ contrasts to almost all other studies, e.g., in Kenya, lone Nubian giraffe represented 17% of observations of which males were 85% (Muller et al., 2018). Such low observations of males might also be attributed to the female-dominated sex ratio, similarly to D’haen et al. (2019).
The West African giraffe inhabiting the “Giraffe Zone” were active most of the day, engaging at times in all activities, from feeding to walking, and vigilant scanning to social interactions. As large-bodied ruminant browser, giraffe inherently invest the majority of time in active searching for food and browsing to meet their metabolic demand (Demment and Van Soest, 1985; du Toit and Yetman, 2005). Yet, searching for food implies frequent, short-distance walking which forms typical fine spatiotemporal scale movements of browsing animals between discrete patches of resources (Gordon and Prins, 2008), i.e., scattered clumps of trees (e.g., Combretum spp., F. albida). In the “Giraffe Zone,” those patches are interspersed by pastoral areas and crop fields (Ciofolo, 1995; Leroy et al., 2009; Gašparová et al., 2023), creating a mosaic agropastoral landscape. Consequently, West African giraffe move greater distances daily and maintain larger home ranges than giraffe elsewhere to meet their demands (Brown et al., 2023).
The allocated day time browsing time of the West African giraffe during our study was high (59%) in comparison with previous findings in the same landscape, i.e., dry (46%) and rainy (22.8%) seasons (Ciofolo and Le Pendu, 2002). As the study was conducted during the dry season, a higher forage time allocation was anticipated (Ciofolo and Le Pendu, 2002). Allocation of time for browsing depends on site-specific resource availability and environmental heterogeneity, varying across giraffe species and populations. For instance, Masai giraffe (G. tippelskirchi) in the Masai Mara National Reserve, Kenya, spent 36% (females) and 39% (males) of time browsing (Adolfsson, 2009) compared with Masai giraffe in Serengeti National Park (NP), Tanzania, 53.2% (females) and 43.2% (males) (Pellew, 1984). High variability in allocation of time spent feeding occurs in browsers across species regardless of body size: 31.7% in black rhino (Diceros bicornis) (Kiwia, 1986), 36% in giant eland (Tragelaphus derbianus derbianus; Hejcmanová et al., 2013), 34.3% in greater kudu (T. strepsiceros; du Toit and Yetman, 2005), and 45% in African savannah elephant (Loxodonta africana; Shannon, 2005). Time allocation also depends on a variety of other non-metabolic factors such as higher stress from anthropogenic disturbances or increased population density which, in fact, is the case of the West African giraffe (Gašparová et al., 2024).
The West African giraffe dedicated 25% of their day to resting, a behavior consistent across age and sex classes and time of day, although marginally increased during midday. Giraffe were mostly standing when resting, including relaxed standing and ruminating. There were, however, several occasions when giraffe laid down, always when together with calves, and sometimes in the presence of sheep/goat, but never with cattle. Resting time was lower compared with Nubian giraffe, which also displayed variance in resting times across the day and seasons, with higher resting time observed in calves compared with adults (Gitau et al., 2024). However, midday resting was similar to Angolan giraffe (G. giraffa angolensis) (Hart et al., 2020) and other African ruminants, e.g., buffalo (Syncerus caffer) (Owen-Smith and Goodall, 2014; Megaze et al., 2018), blue wildebeest (Connochaetes taurinus) (Owen-Smith and Traill, 2017), and sable antelope (Hippotragus niger), especially during the dry season. The difference in resting patterns is likely environmental as the study of Nubian giraffe was conducted in a protected area with more diverse habitats and wildlife, including predators and without human and livestock presence (Gitau et al., 2024). From our findings, West African giraffe in the human-dominated landscape decreased resting time during the day, possibly shifting their resting to night when people are less active and there are no other predators.
The proportion of vigilance in the West African giraffe was on average low (5%) with calves being the least vigilant in comparison with adult males and females. Due to the absence of predators in the “Giraffe Zone,” our findings differed from those elsewhere, e.g., Paulse et al. (2023) recorded vigilance from 7% to 11% of daylight activity in adult giraffe and up to 47% in juveniles.
Inhabiting an agropastoral landscape with human settlements and co-occurrence with domestic animals naturally modulate the West African giraffe activity patterns. Importantly, they did not compete with the livestock for food resources because of their height and preferences. As with other browsers (Fritz et al., 1996), giraffe are little disturbed by livestock and even associate with them as we observed. Kinga et al. (2018) described that large browsers are attracted to livestock at short distances (up to 500 m) in pastoral grazing lands with free-ranging livestock herds accompanied by herders. For approximately 40% of their time, West African giraffe associated with livestock at short distances and within visibility that the Sahelian tiger bush savanna allowed, similar to time they spent with other giraffe (42%). As such, they preferred to “be in group” regardless of “a specific group” or species (Muller et al., 2018), but notably often not the same giraffe in a herd. This finding corresponds to their social resting strategy, further supported by the tendency of giraffe to relax and rest in proximity of another animals, even if our data confirmed significant effect only for distance to another giraffe. They were often observed browsing close to sheep/goat yet appeared less relaxed in the presence of cattle. This disturbance may in fact be a result of cattle herds increasing dust and giraffe seeking to avoid it (Tawey, pers. comm., AVEN), or because cattle are often accompanied by herders.
Interestingly, the West African giraffe did not alter their activity or resting patterns in response to human presence. Our observations of giraffe in alert when people appeared within sight or earshot (e.g., kids shouting) were anecdotal. Giraffe were also alert when herders with visible sticks accompanied larger cattle herds. These sticks are commonly used by locals to protect their crops and mango trees from giraffe approaching villages in search for food, by making threatening gestures (Suraud, 2011; Gašparová et al., 2023). Positively, people living in the “Giraffe Zone” generally hold favorable attitudes toward giraffe, are aware of their threatened status, and rarely harm them (Leroy et al., 2009; Suraud, 2011; Gašparová et al., 2023). In recent decades, the West African giraffe in Niger have always lived in a human-dominated landscape. Therefore, they most likely evolved a certain level of habituation and tolerance toward people (e.g., Blumstein, 2016; Scheijen et al., 2021), with limited disruption to their behavioral patterns while keeping vigilance under specific circumstances. Living in proximity to human settlements is not unique to the West African giraffe, although not always in similar ways. For example, Masai giraffe living in proximity to people in Tanzania showed social disruptions and/or looser social associations within larger giraffe herds (Bond et al., 2021).
Our findings on the activity and resting behavior patterns of West African giraffe and their response to livestock and people is positive for their conservation with the population adapted to the anthropogenic landscape which they inhabit. We demonstrated that long-term awareness campaigns and conservation measures by NGOs and government at a local level are beneficial for their coexistence.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because giraffes were observed using binoculars.
Author contributions
PH: Conceptualization, Data curation, Formal analysis, Methodology, Writing – original draft, Writing – review & editing. KG: Data curation, Funding acquisition, Investigation, Writing – original draft, Writing – review & editing. TF: Writing – original draft, Writing – review & editing. MV: Data curation, Writing – review & editing. JF: Funding acquisition, Writing – review & editing. AM: Resources, Writing – review & editing. TR: Resources, Writing – review & editing. KB: Funding acquisition, Methodology, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was financially supported by Czech University of Life Sciences, Faculty of Tropical AgriSciences IGA FTZ 20243107, Giraffe Conservation Foundation, Hlávkova nadace ‘Nadání’, and a Rufford Small Grant (grant number: 27641-1).
Acknowledgments
We thank the Nigerien Wildlife Authorities and their staff for the permission to undertake the work and the Giraffe Conservation Foundation who provided valuable technical and financial support. In addition, we thank Czech University of Life Sciences Prague, Hlávkova nadace ‘Nadání’, Ivan Carter Wildlife Conservation Alliance, IUCN SOS and the European Commission, Rufford Small Grant, Sahara Conservation Fund, for their funding and/or logistical support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adolfsson U. G. (2009). Diurnal behaviour and utilization of shade in Masai giraffes (Giraffa camelopardalis tippelskirchi) (Uppsala, Sweden: Swedish University of Agricultural Sciences).
Beauchamp G., Li Z., Yu C., Bednekoff P. A., Blumstein D. T. (2021). A meta-analysis of the group-size effect on vigilance in mammals. Behav. Ecol. 32, 919–925. doi: 10.1093/beheco/arab048
Beekman J. H., Prins H. H. T. (1989). Feeding strategies of sedentary large herbivores in East Africa, with emphasis on the African buffalo, Syncerus coffer. Afr. J. Ecol. 27, 129–147. doi: 10.1111/j.1365-2028.1989.tb00937.x
Blumstein D. T. (2016). Habituation and sensitization: new thoughts about old ideas. Anim. Behav. 120, 255–262. doi: 10.1016/j.anbehav.2016.05.012
Bond M. L., König B., Lee D. E., Ozgul A., Farine D. R. (2021). Proximity to humans affects local social structure in a giraffe metapopulation. J. J. Anim. Ecol. 90, 212 221. doi: 10.1111/1365-2656.13247
Bond M. L., Lee D. E., Ozgul A., König B. (2019). Fission-fusion dynamics of a megaherbivore are driven by ecological, anthropogenic, temporal, and social factors. Oecologia 19, 335–347. doi: 10.1007/s00442-019-04485-y
Brown M. B., Fennessy J. T., Crego R. D., Fleming C. H., Alves J., Brandlová K., et al. (2023). Ranging behaviours across ecological and anthropogenic disturbance gradients: a pan-African perspective of giraffe (Giraffa spp.) space use. Proc. Biol. Sci. 290, 20230912. doi: 10.1098/rspb.2023.0912
Burger A. L., Fennessy J., Fennessy S., Dierkes P. W. (2020b). Nightly selection of resting sites and group behavior reveal antipredator strategies in giraffe. Nat. Ecol. Evol. 10, 2917–2927. doi: 10.1002/ece3.6106
Burger A. L., Hartig J., Dierkes P. W. (2020a). Shedding light into the dark: Age and light shape nocturnal activity and sleep behaviour of giraffe. Appl. Anim. Behav. Sci. 229, 105012. doi: 10.1016/j.applanim.2020.105012
Cameron E. Z., du Toit J. T. (2005). Social influences on vigilance behaviour in giraffes, Giraffa camelopardalis. Anim. Behav. 69, 1337–1344. doi: 10.1016/j.anbehav.2004.08.015
Carter K. D., Brand R., Carter J. K., Shorrocks B., Goldizen A. W. (2013). Social networks, long-term associations and age-related sociability of wild giraffes. Anim. Behav. 86, 901–910. doi: 10.1016/j.anbehav.2013.08.002
Chen H. L., Koprowski J. L. (2015). Animal occurrence and space use change in the landscape of anthropogenic noise. Biol. Conserv. 192, 315–322. doi: 10.1016/j.biocon.2015.10.003
Ciofolo I. (1995). West Africa’s last giraffes: the conflict between development and conservation. J. Trop. Ecol. 11, 577–588. doi: 10.1017/S0266467400009159
Ciofolo I., Le Pendu Y. (2002). The feeding behaviour of giraffe in Niger. Mammalia. 66, 183–194. doi: 10.1515/mamm.2002.66.2.183
Ciuti S., Northrup J. M., Muhly T. B., Simi S., Musiani M., Pitt J. A., et al. (2012). Effects of humans on behaviour of wildlife exceed those of natural predators in a landscape of fear. PloS One d7, e50611. doi: 10.1371/journal.pone.0050611
Craig M., Dewar M., Harris M. A., Della Sala S., Wolbers T. (2016). Wakeful rest promotes the integration of spatial memories into accurate cognitive maps. Hippocampus 26, 185–193. doi: 10.1002/hipo.22502
Creel S., Schuette P., Christianson D. (2014). Effects of predation risk on group size, vigilance, and foraging behavior in an African ungulate community. Behav. Ecol. 25, 773–784. doi: 10.1093/beheco/aru050
D’haen M., Fennessy J., Stabach J. A., Brandlová K. (2019). Population structure and spatial ecology of Kordofan giraffe in Garamba National Park, Democratic Republic of Congo. Nat. Ecol. Evol. 9, 11395–11405. doi: 10.1002/ece3.5640
Demment M. W., Van Soest P. J. (1985). A nutritional explanation for body-size patterns of ruminant and nonruminant herbivores. Am. Nat. 125, 641–672. doi: 10.1086/284369
Duggan G., Burn C. C., Clauss M. (2016). Nocturnal behavior in captive giraffe (Giraffa camelopardalis)—A pilot study. Zoo. Biol. 35, 14–18. doi: 10.1002/zoo.21248
du Toit J. T., Yetman C. A. (2005). Effects of body size on the diurnal activity budgets of African browsing ruminants. Oecologia. 143, 317–325. doi: 10.1007/s00442-004-1789-7
Fischhoff I. R., Sundaresan S. R., Cordingley J., Rubenstein D. I. (2007). Habitat use and movements of plains zebra (Equus burchelli) in response to predation danger from lions. Behav. Ecol. 18, 725–729. doi: 10.1093/beheco/arm036
Fritz H., De Garine-Wichatitsky M., Letessier G. (1996). Habitat use by sympatric wild and domestic herbivores in an African savanna woodland: the influence of cattle spatial behaviour. J. Appl. Ecol. 33, 589–598. doi: 10.2307/2404987
Gašparová K., Blanco J., Glikman J. A., Fennessy J., Zabeirou A. R. M., Mahamadou A. R. A., et al. (2023). Social development and biodiversity conservation synergies for the West African giraffe in a human–wildlife landscape. Environ. Conserv. 50, 259–266. doi: 10.1017/S0376892923000243
Gašparová K., Fennessy J., Moussa Zabeirou A. R., Abagana A. L., Rabeil T., Brandlová K. (2024). Saving the last west African giraffe population: A review of its conservation status and management. Animals 14, 702. doi: 10.3390/ani14050702
Gašparová K., Fennessy J., Rabeil T., Brandlová K. (2020). Threat analysis: West African giraffe (Giraffa camelopardalis peralta) in Republic of Niger (Niamey, Niger: Czech University of LIfe Sciences Prague & Giraffe Conservation Foundation), 18pp. Available at: https://giraffeconservation.org/wp-content/uploads/2020/05/Threat-analysis-West-African-giraffe-in-Niger_April-2020.pdf.
Gaynor K. M., Hojnowski C. E., Carter N. H., Brashares J. S. (2018). The influence of human disturbance on wildlife nocturnality. Science. 360, 1232–1235. doi: 10.1126/science.aar7121
Gitau C. G., Mbau J. S., Ngugi R. K., Ngumbi E. (2024). Activity budget and foraging patterns of Nubian giraffe (Giraffa camelopardalis camelopardalis) in Lake Nakuru National Park, Kenya. Ecol. Evol. 14, e11463. doi: 10.1002/ece3.11463
Gloneková M., Brandlová K., Pluháček J. (2021). Further behavioural parameters support reciprocity and milk theft as explanations for giraffe allonursing. Sci. Rep. 11, 7024. doi: 10.1038/s41598-021-86499-2
Hart E. E., Fennessy J., Hauenstein S., Ciuti S. (2020). Intensity of giraffe locomotor activity is shaped by solar and lunar eitgebers. Behav. processes 178, 104178. doi: 10.1016/j.beproc.2020.104178
Hejcmanová P., Vymyslická P., Žáčková M., Hejcman M. (2013). Does supplemental feeding affect behaviour and foraging of critically endangered western giant eland in an ex situ conservation site? Afr. Zool. 48, 250–258. doi: 10.1080/15627020.2013.11407590
Hirakawa H. (1997). Digestion-constrained optimal foraging in generalist mammalian herbivores. Oikos 78, 37–47. doi: 10.2307/3545798
Jayakody S., Sibbald A. M., Gordon I. J., Lambin X. (2008). Red deer Cervus elephus vigilance behaviour differs with habitat and type of human disturbance. Wildl. Biol. 14, 81–91. doi: 10.2981/0909-6396(2008)14[81:RDCEVB]2.0.CO;2
Jeschke J. M., Tollrian R. (2005). Predicting herbivore feeding times. Ethol 111, 187–206. doi: 10.1111/j.1439-0310.2004.01052.x
Kinga G. W., Mironga J., Odadi W. O. (2018). Analysis of the spatial relationship between cattle and wild ungulates across different land-use systems in a tropical Savanna landscape. Int.J eccol. 2018, 2072617. doi: 10.1155/2018/2072617
Kiwia H. Y. D. (1986). Diurnal activity pattern of the black rhinoceros. Diceros bicornis (L.) in Ngorongoro Crater, Tanzania. Afr. J. @ Ecol. 24, 89–96. doi: 10.1111/j.1365-2028.1986.tb00347.x
Le Pendu Y., Ciofolo I., Gosser A. (2000). The social organization of giraffes in Niger. Afr. J. Ecol. 38, 78–85. doi: 10.1046/j.1365-2028.2000.00214.x
Leroy R., De Visscher M. N., Halidou O., Boureima A. (2009). The last African white giraffes live in farmers’ fields. Biodivers. Conserv. 18, 2663–2677. doi: 10.1007/s10531-009-9628-0
Lima S. L., Bednekoff P. A. (1999). Temporal variation in danger drives antipredator behavior: the predation risk allocation hypothesis. Am. Nat. 153, 649–659. doi: 10.1086/303202
Loft E. R., Menke J. W., Kie J. G. (1991). Habitat shifts by mule deer: the influence of cattle grazing. J. Wildl. Manage. 55, 16–26. doi: 10.2307/3809236
Luttbeg B. (2017). Re-examining the causes and meaning of the risk allocation hypothesis. Am. Nat. 189, 644–656. doi: 10.1086/691470
Makenbach S. A., Waterman J. M., Roth J. D. (2013). Predator detection and dilution as benefits of associations between yellow mongooses and Cape ground squirrels. Behav. Ecol. Sociobiol. 67, 1187–1194. doi: 10.1007/s00265-013-1544-3
Marealle W. N., Holmern T., Røskaft E. (2020). Factors Affecting Group Size and Vigilance Behaviour of Maasai Giraffe (Giraffa camelopardalis tippelskirchi) on the Serengeti-Ngorongoro Ecosystem, Tanzania. EANSO 2, 14–23. doi: 10.37284/eajenr.2.1.133
Megaze A., Balakrishnan M., Belay G. (2018). Diurnal activity budget of African buffalo (Syncerus caffer Sparrman 1779) in chebera churchura national park, Ethiopia. Afr. J. Ecol. 56, 436–444. doi: 10.1111/aje.12359
Mole M. A., Rodrigues DÁraujo S., Van Aarde R. J., Mitchell D., Fuller A. (2016). Coping with heat: behavioural and physiological responses of savanna elephants in their natural habitat. Conserv. Physiol. 4, cow044. doi: 10.1093/conphys/cow044
Muller Z., Cuthill I. C., Harris S. (2018). Group sizes of giraffes in Kenya: the influence of habitat, predation and the age and sex of individuals. J. Zoo. 306, 77–87. doi: 10.1111/jzo.12571
Muller Z., Harris S. (2021). A review of the social behaviour of the giraffe Giraffa camelopardalis: a misunderstood but socially complex species. Mammal Revi. 52, 1–15. doi: 10.1111/mam.12268
Owen-Smith N., Goodall V. (2014). Coping with savanna seasonality: comparative daily activity patterns of African ungulates as revealed by GPS telemetry. J. Zool. 293, 181–191. doi: 10.1111/jzo.12132
Owen-Smith N., Traill L. W. (2017). Space use patterns of a large mammalian herbivore distinguished by activity state: fear versus food? J. Zool. 303, 281–290. doi: 10.1111/jzo.12490
Paulse J., Couldridge V., Cupido C., Deacon F. (2023). The diurnal activity budgets of extralimital giraffe (Giraffa camelopardalis giraffa) in the Western Cape Province, South Africa. Afr. J. Ecol. 61, 746–751. doi: 10.1111/aje.13135
Pellew R. A. (1984). The feeding ecology of selective browser, the giraffe (Giraffa camelopardalis tippelskirchi). J. Zool. 202, 57–81. doi: 10.1111/j.1469-7998.1984.tb04288.x
Rattenborg N. C., de la Iglesia H. O., Kempenaers B., Lesku J. A., Meerlo P., Scriba M. F. (2017). Sleep research goes wild: new methods and approaches to investigate the ecology, evolution and functions of sleep. Philos. Trans. R. Soc Lond. B Biol. Sci. 372, 20160251. doi: 10.1098/rstb.2016.0251
R Core Team (2024). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org.
Ripari L., Premier J., Belotti E., Bluhm H., Breitenmoser-Würsten C., Bufka L., et al. (2022). Human disturbance is the most limiting factor driving habitat selection of a large carnivore throughout Continental Europe. Biol. Conserv. 266, 109446. doi: 10.1093/jmammal/gyz044
Scheijen C. P., van der Merwe S., Ganswindt A., Deacon F. (2021). Anthropogenic influences on distance traveled and vigilance behavior and stress-related endocrine correlates in free-roaming giraffes. Animals. 11, 1239. doi: 10.3390/ani11051239
Schmidt M. H. (2014). The energy allocation function of sleep: a unifying theory of sleep, torpor, and continuous wakefulness. Neurosci. Biobehav.Rev. 47, 122–153. doi: 10.1016/j.neubiorev.2014.08.001
Scholte P., Pays O., Adam S., Chardonnet B., Fritz H., Mamang J. B., et al. (2022). Conservation overstretch and long-term decline of wildlife and tourism in the Central African savannas. Conserv. Biol. 36, e13860. doi: 10.1111/cobi.13860
Shannon G. (2005). The effects of sexual dimorphism on the movements and foraging ecology of the African elephant. (Durban, South Africa: University of KwaZulu-Natal).
Shukla I., Kilpatrick A. M., Beltran R. S. (2021). Variation in resting strategies across trophic levels and habitats in mammals. Ecol. Evol. 11, 14405–14415. doi: 10.1002/ece3.8073
Stabach J. A., Hughey L. F., Crego R. D., Fleming C. H., Hopcraft J. G. C., Leimgruber P. (2022). Increasing anthropogenic disturbance restricts wildebeest movement across East African grazing systems. Front. Ecol. Evol. 10. doi: 10.3389/fevo.2022.846171
Stensland E. V. A., Angerbjörn A., Berggren P. E. R. (2003). Mixed species groups in mammals. Mamm. Rev. 33, 205–223. doi: 10.1046/j.1365-2907.2003.00022.x
Suraud J. P. (2011). Identifying conservation constraints for the last West African giraffe: population dynamics, determining factors and spatial distribution pattern. University of Lyon, France.
Suscke P., Presotto A., Izar P. (2021). The role of hunting on Sapajus xanthosternos’ landscape of fear in the Atlantic Forest, Brazil. Am. J. Primatol. 83, e23243. doi: 10.1002/ajp.23243
Tucker M. A., Schipper A. M., Adams T. S., Attias N., Avgar T., Babic N. L., et al. (2023). Behavioral responses of terrestrial mammals to COVID-19 lockdowns. Science 380, 1059–1064. doi: 10.1126/science.abo6499
Vermeulen M. M., Fritz H., Strauss W. M., Hetem R. S., Venter J. A. (2024). Seasonal activity patterns of a Kalahari mammal community: Trade-offs between environmental heat load and predation pressure. Ecol. Evol. 14, e11304. doi: 10.1002/ece3.11304
Worlddata.info (2024). The climate in Niger. Available online at: https://www.worlddata.info/africa/niger/climate.php (Accessed September 14, 2024).
Zanette L. Y., Frizzelle N. R., Clinchy M., Peel M. J., Keller C. B., Huebner S. E., et al. (2023). Fear of the human “super predator” pervades the South African savanna. Curr. Biol. 33, 4689–4696. doi: 10.1016/j.cub.2023.08.089
Keywords: Giraffa camelopardalis peralta, wildlife - livestock interaction, anthropogenic disturbance, resting behaviour, human - wildlife coexistence, human-dominated landscape, Niger
Citation: Hejcmanová P, Gašparová K, Fedorova T, Vukelić M, Fennessy J, Moussa Zabeirou AR, Rabeil T and Brandlová K (2024) Diurnal activity and resting time allocation of the West African giraffe in an agropastoral human-dominated landscape. Front. Conserv. Sci. 5:1459960. doi: 10.3389/fcosc.2024.1459960
Received: 05 July 2024; Accepted: 30 September 2024;
Published: 21 October 2024.
Edited by:
Anton Baotic, Austrian Academy of Sciences (OeAW), AustriaReviewed by:
Andrea Presotto, East Carolina University, United StatesJamie Paulse-Ross, University of the Free State, South Africa
Copyright © 2024 Hejcmanová, Gašparová, Fedorova, Vukelić, Fennessy, Moussa Zabeirou, Rabeil and Brandlová. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Karolína Brandlová, YnJhbmRsb3ZhQGZ0ei5jenUuY3o=