 Katia Maria Paschoaletto Micchi de Barros Ferraz1*
Katia Maria Paschoaletto Micchi de Barros Ferraz1* Isabella de Freitas Bento1Anna Beatriz Queiroz Di Souza1Caio da Silveira Nunes1
Isabella de Freitas Bento1Anna Beatriz Queiroz Di Souza1Caio da Silveira Nunes1 Maria Augusta de Mendonça Guimarães1Monicque Silva Pereira1,2Livia Lima da Silva1Letícia Keiko Nunes de Campos1Anita Seneme Gobbi1José Caio Quadrado Alves1Loisa Fabrícia Prates Alvarez1
Maria Augusta de Mendonça Guimarães1Monicque Silva Pereira1,2Livia Lima da Silva1Letícia Keiko Nunes de Campos1Anita Seneme Gobbi1José Caio Quadrado Alves1Loisa Fabrícia Prates Alvarez1 Silvio Marchini1Ana Carla Medeiros Morato de Aquino1Vinicius Kenji de Moraes Sato1Roberta Montanheiro Paolino1,3
Silvio Marchini1Ana Carla Medeiros Morato de Aquino1Vinicius Kenji de Moraes Sato1Roberta Montanheiro Paolino1,3- 1Laboratório de Ecologia, Manejo e Conservação de Fauna Silvestre, Departamento de Ciências Florestais, Escola Superior de Agricultura “Luiz de Queiroz”, Piracicaba, SP, Brazil
- 2Secretaria de Meio Ambiente, Infraestrutura e Logística do Estado de São Paulo, São Paulo, SP, Brazil
- 3Laboratório de Ecologia e Conservação, Faculdade de Filosofia, Ciências e Letras de Ribeirão Preto, Universidade de São Paulo, Ribeirão Preto, SP, Brazil
Land sharing strategies for conciliating biodiversity conservation and human development usually do not consider the need to deal with human-wildlife conflicts, a type of human-wildlife interaction (HWI). To measure, monitor and solve conflicts is fundamental to achieve the coexistence necessary to promote environmental and social justice. Here, we present a new approach to assess and evaluate HWI aiming to inform decision-making regarding conflicts. We developed a method to classify and map events between humans and wildlife according to their nature, context and effect for both sides, distinguishing what an encounter (unidirectional) is and what an interaction (bidirectional effect) is. We typified and categorized HWI regarding their effects (positive or negative) for both sides. We compiled opportunistic observations from events between humans and wildlife in a shared landscape (campus Luiz de Queiroz, Piracicaba, São Paulo, Brazil) from February 2022 until April 2024. We created a standardized table, performed descriptive statistics, used Minimum Bounding Geometry and Kernel Density, a simple method idealized to assist scientists and managers in different contexts. We had a total of 570 events, of which 297 were characterized as encounters and 273 as HWI. We recorded 42 animal taxa related to the events, of which 36 interacted with humans, being Nasua nasua, Cairina moschata, and Didelphis albiventris the most frequent ones. We identified 16 types of HWI that can occur in shared landscapes, 10 were categorized as Human-Wildlife Conflict, six as Unsustainable Use, four as Wildlife Damage and three as Convivencia, with some classified in more than one category. Among them, 10 occurred in our studied landscape with Unintentional Feeding being the most frequent one. We classified for the first time Unexpected Encounter, Accident Avoidance and Chase Away as HWI. Spaces of interaction were close to main buildings, central lawn and cats’ feeding sites. Our approach was useful to prioritize species and stakeholders, and to identify the large amount of food supply due to inadequate waste disposal and domestic cat feeding as the primary reason for most of HWI in our studied landscape. We recommend a comprehensive characterization of HWI to find interconnections and guide strategies for coexistence.
1 Introduction
Humans and wildlife have been increasingly co-occurring in shared landscapes leading to a vast number of interactions (Soulsbury and White, 2015; Schell et al., 2020). Human-wildlife interactions (HWI) are events involving direct or indirect contact between humans and animals (Whitehouse-Tedd et al., 2021), with effects for both sides (bidirectional effects) that can vary substantially in nature, frequency and severity (Soulsbury and White, 2019). Typical examples of HWI that result from direct contact between humans and animals are wildlife-vehicle collisions, intentional feeding, poaching or retaliation, whereas examples of interactions that result from indirect contact are crop damage, livestock predation and unintentional feeding.
HWI may vary according to land sharing/land sparing context (Crespin and Simonetti, 2019). In urban shared landscapes, we might expect more frequent conflicts such as nuisance wildlife, wildlife-vehicle collisions, intentional and non-intentional feeding, unexpected encounter, property damage and zoonotic disease. In rural shared/spared landscape, we might expect more frequently crop damage, livestock predation, poaching, harvesting, and retaliation. This emphasizes the importance of local assessment of interactions between humans and wildlife considering different contexts to better inform decision-making.
Assessing and evaluating HWI are essential for guiding decision-making towards coexistence. Coexistence is a sustainable though dynamic state, where humans and wildlife co-adapt for sharing landscapes and human interactions with wildlife are effectively governed to ensure wildlife populations persist in socially legitimate ways that ensure tolerable risk levels (Carter and Linell, 2016; Pooley et al., 2020). So, coexistence is ecologically and socially complex, context-specific, dynamic and, at some point, it generally requires agreement – or at the very least, cooperation – between different groups of people about the wildlife in question (IUCN, 2023).
Coexistence has emerged into the mainstream of conservation science to better understand and manage interactions between humans and wildlife with negative outcomes (IUCN, 2020; 2023). Interactions with negative outcomes and underpinning by social tensions between groups of people are known as human-wildlife conflicts (Dickman, 2010). The human-wildlife conflict is defined as “struggles that emerge when the presence or behavior of wildlife poses an actual or perceived, direct and recurring threat to human interests or needs, leading to disagreements between groups of people and negative impacts on people and/or wildlife” (IUCN, 2020). Other definitions, such as biodiversity conflicts, focus more on the competition between people about wildlife than on human-wildlife interaction and its effects on the other (Young et al., 2010; White et al., 2009). Also, the anthropological perspective of conflicts focuses not only on the material dimension of conflicts, but more on the social and cultural ones, which reveals the tensions and divisions in human society that affect conflicts (Knight, 2000).
Human-wildlife conflicts have tremendously impacted the whole global community due to the adverse effects on wildlife and ecosystems, human health, safety, equity, social dynamics, sustainable development, commodity production and businesses, ultimately impeding coexistence (Nyhus, 2016; Gross et al., 2021). Despite the importance of recognizing and managing human-wildlife conflicts for species conservation, a paradigm’ shift, from human-wildlife conflicts to human-wildlife coexistence, has been proposed and it has opened a forefront for more positive and inclusive relations with wildlife and nature (Frank and Glikman, 2019).
To advance, the science of coexistence requires a better understanding and evaluation of nature, context, dynamics, and effects of HWI, especially in shared landscapes where the conflicts of living together are even more costly and inevitable. Thus, in this study, we aim to present an approach for assessing and evaluating HWI at multispecies level, accounting the complexity and diversity of interactions, as a basis for guiding decisions towards coexistence in shared landscapes. The results presented here are part of the ongoing transdisciplinary research project Wildlife Neighbors: Towards Human-Wildlife Coexistence, which seeks to transform coproduced knowledge into accessible, useful, and actionable knowledge, supporting the decision-making process for transformative changes towards human-wildlife coexistence.
To illustrate this approach, we used HWI recorded in the last two years in a shared landscape in southeastern Brazil, characterized by an interesting scenario, common from urban spaces, which brings together people, wildlife, wildlife-unfriendly infrastructure, feeding sites for abandoned cats, abundant and available organic waste, and restored green spaces. All these elements together make up a landscape conducive to human-wildlife conflicts that are difficult for public managers to resolve. We analyzed the hotspots of the most typical HWI according to the landscape attributes to (i) better understand the reasons for the high frequency of the HWI and (ii) properly plan for the impacts mitigation in the spaces delimited.
In the last two decades, the campus has undergone a profound process of land cover change, due to a Conduct Adjustment Agreement (CAA) established by the Public Ministry to ensure the restoration of the riparian forests and Legal Reserve in compliance with the Brazilian Forest Code (Brazil, 2012). The total increase in forested habitats since the CAA was established is 55,7 ha1. Therefore, nowadays the landscape of the Luiz de Queiroz campus is totally favorable for hosting a high diversity of wildlife (Alexandrino et al., 2013; Bovo et al., 2018; Alexandrino et al., 2021), which seems to be increasing as new species of birds and mammals have been more frequently recorded recently.
Planned to host a public park in the urban perimeter, the campus has artificial lakes that contribute to maintaining a significant diversity of wildlife, particularly waterfowl (such as ducks, geese and teals) in areas with intense human use. For cultural reasons, humans intentionally feed the waterfowls, especially the muscovy ducks (Cairina moschata), what result in a high frequency of encounters between these species and humans. The food provided for the waterfowls also attracts other vertebrate species.
Additionally, the campus has many abandoned cats, which have been cared for by volunteers (employees, students, external users) for over 20 years, and who are responsible for installing and maintaining more than 60 cat feeding sites throughout the area. Many wildlife species have been frequently sighted feeding at cat feeding sites.
In a landscape with so many people and high diversity and abundance of wildlife, encounters and interactions between humans and animals are inevitable and have increased considerably in recent years, with the majority of the interactions resulting in negative outcomes for both.
2 Methods
2.1 Study area
We have been carrying out a HWI assessment and evaluation in a shared landscape (campus USP “Luiz de Queiroz”, 22 42’ 30” S and 47 38’ 30” W, 546 meters of elevation, and 914,5 ha), in Piracicaba, São Paulo state, Brazil (Figure 1). The Luiz de Queiroz campus is part of the University of São Paulo since 1934 and is composed by buildings for students and professionals (graduate and undergraduate students, professors, researchers and employees), open green areas for leisure activities, agricultural fields (sugar cane, corn, rice) and livestock (cattle, sheep, goats, horses) for experimental researches and classes, Pinus and eucalyptus plantation for forestry management, artificial water bodies and streams, semi-deciduous seasonal Atlantic forest remnants, riparian forests, and paved and unpaved streets. The Luiz de Queiroz campus is bordered by the Piracicaba River on one side and by avenues and residential houses (Piracicaba city) on the other. Around 5,000 people, 4,000 vehicles and 400 cyclists circulate on the campus daily for teaching, researching, outreach and leisure activities.
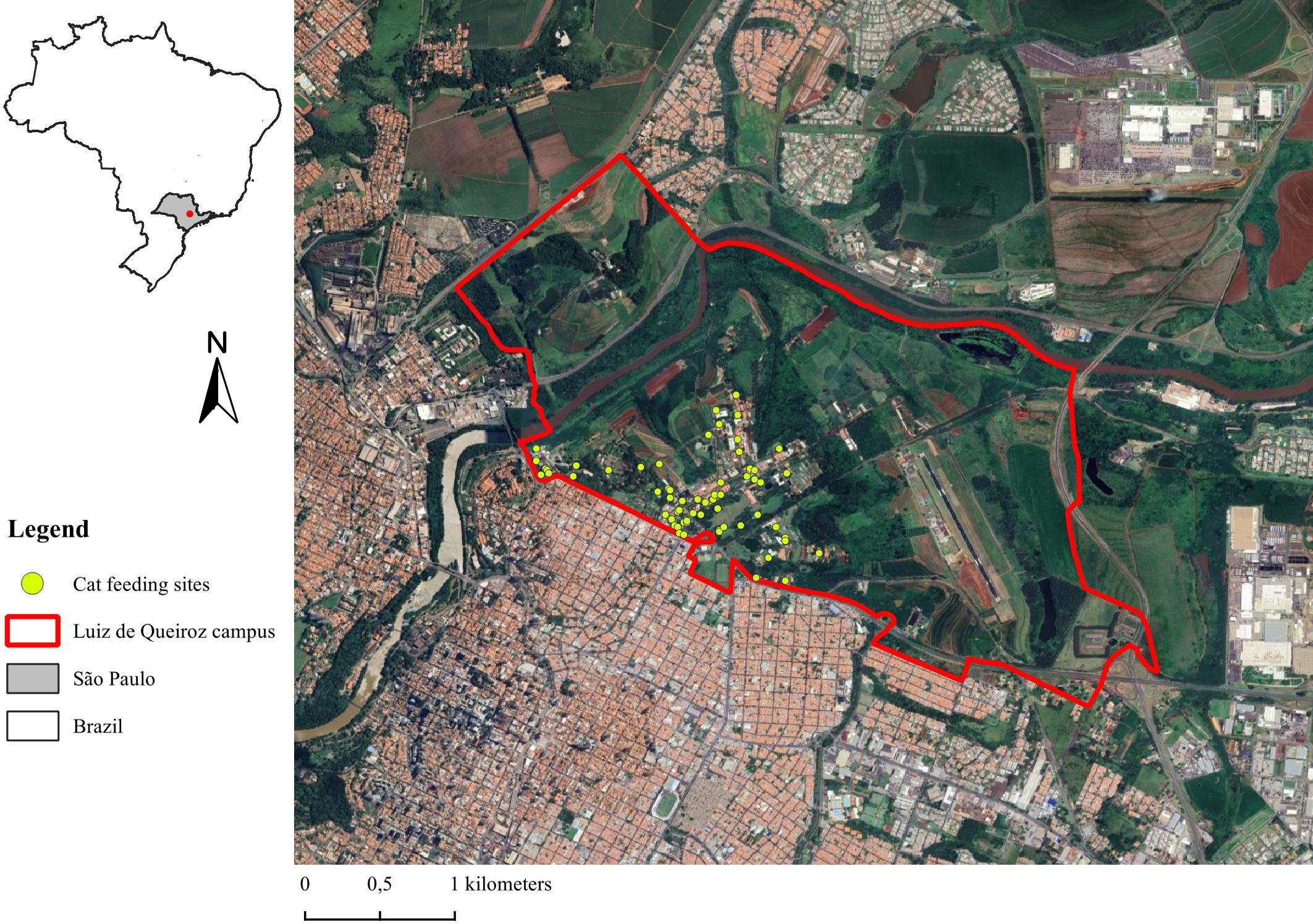
Figure 1. Luiz de Queiroz campus, Piracicaba, São Paulo, Brazil.
2.2 Data collection and classification
We compiled opportunistic observations from events between humans and wildlife in the campus from February 2022 until April 2024. Events were gathered by two different sources: (i) from the research group (primary data), during wildlife surveys or opportunistic sightings; and (ii) from external collaborators (secondary data), made by someone interested in sharing the observations with our research team, usually sent via WhatsApp, but also by email or phone call. As the Wildlife Neighbors Project became more popular on the campus, others became more interested in collaborating by sharing events with wildlife. The data collected by the research team was also complemented by other records from projects conducted on campus such as the monitoring of feral cats (2022), the assessment of the impact of cat feeding sites on wildlife (2022), the monitoring and population management of coatis (2023-2024), and emergency wildlife rescue (2023-2024).
To organize the events recorded, we created a standardized table (Supplementary Table S1) containing as much information as possible. This table included information about the (i) event itself, such as date, time, GPS coordinates, place, nature, context, effect for humans, effect for animal, type (if interaction), picture/video; and (ii) details about the involved animal, such as species and number of individuals.
The effects of each event could be positive, negative or neutral (i.e., no effect). Events with neutral effects for one of the parts (humans or wildlife) involved in the interactions were classified as unidirectional, while events with effects (negative or positive) for both, regardless of who, were classified as bidirectional. Then, based on uni or bidirectional effects, events were classified as encounters or interactions, respectively.
To avoid subjectiveness, we did not evaluate reactions from animals or people to interactions. For example, some people enjoy feeding wildlife, others do not like it, but, regardless of the feeling/reaction, the consequences of feeding wildlife are almost always negative for the animals as they become habituated to human food and places, resulting in behavioral and metabolic changes.
For the events classified as interactions, we created a typology, attributing names for each interaction based on its characteristics. Some of the interactions are typically already known as HWI such as Wildlife Damage, Livestock Predation and Retaliation, while others, such as Unintentional Feeding, Unexpected Encounter, Accident Avoidance, and Chase Away were classified as HWI for the first time in this paper. We also included in the list of interactions others not recorded in our landscape, but frequently reported in others, in order to provide a more complete picture of the variety of the HWI that can occur in shared landscapes. For some specific situations, the event was classified into more than one type of interaction, according to its nature and interrelation (e.g., Unintentionally Feeding and Nuisance Wildlife).
Regarding species classification, the recorded bird species were categorized based on their taxonomic order, underlining that similarities in morphological and behavioral traits within a taxon imply similar management strategies for addressing conflicts between closely related species. The snakes were classified into venomous and non-venomous categories, as this distinction dictates the urgency in implementing management measures to address potential conflicts. It’s important to note that alternative groupings may also be considered, depending on specific characteristics relevant to the management context. Toads, frogs and tree frogs were grouped in the Anura Order. If a species could not be identified to a specific level, it was categorized as ‘non-identified,’ such as ‘non-identified snakes’ or ‘non-identified birds’.
2.3 Human-wildlife interactions analysis
Since our purpose was to identify interactions that should be managed to change the negative outcomes for promoting coexistence, we focused our analysis only on HWI. We analyzed all the attributes of the interactions (type, species, place, nature, context, and frequency) and the effects (positive or negative) by descriptive statistics in excel. We also framed the interactions, under conservation and social perspectives, following the conceptual framework proposed by Marchini et al. (2021), into four categories: (i) Unsustainable Use: the effect is negative for wildlife, but positive for humans, (ii) Human-Wildlife Conflicts: the effect is negative for both, (iii) Wildlife Damage: the effect is positive for wildlife, but negative for humans, and (iv) Convivencia: the effect is positive for both wildlife and humans.
We plotted all recorded HWI in the landscape. We delimited the human-wildlife spaces for the most typical interactions (e.g., human-coati) by using the Minimum Bounding Geometry (convex_hull) in the ArcGis 10.3 (ESRI, 2014). Then, we used the Kernel Density tool (default search radius = bandwidth) to calculate the density of features (HWI) in a neighborhood around those features in the ArcGis 10.3 (ESRI, 2014).
3 Results
We recorded 570 events involving humans and wildlife in the studied shared landscape, of which 297 were characterized as encounters (with unidirectional effects either for humans or for wildlife) and 273 were characterized as HWI (effects for both humans and wildlife). From these events, we recorded 42 taxa, of which 36 taxa interacted with humans, of which coatis (Nasua nasua), muscovy ducks (Cairina moschata) and white-eared opossums (Didelphis albiventris) were the most frequent ones (Table 1).
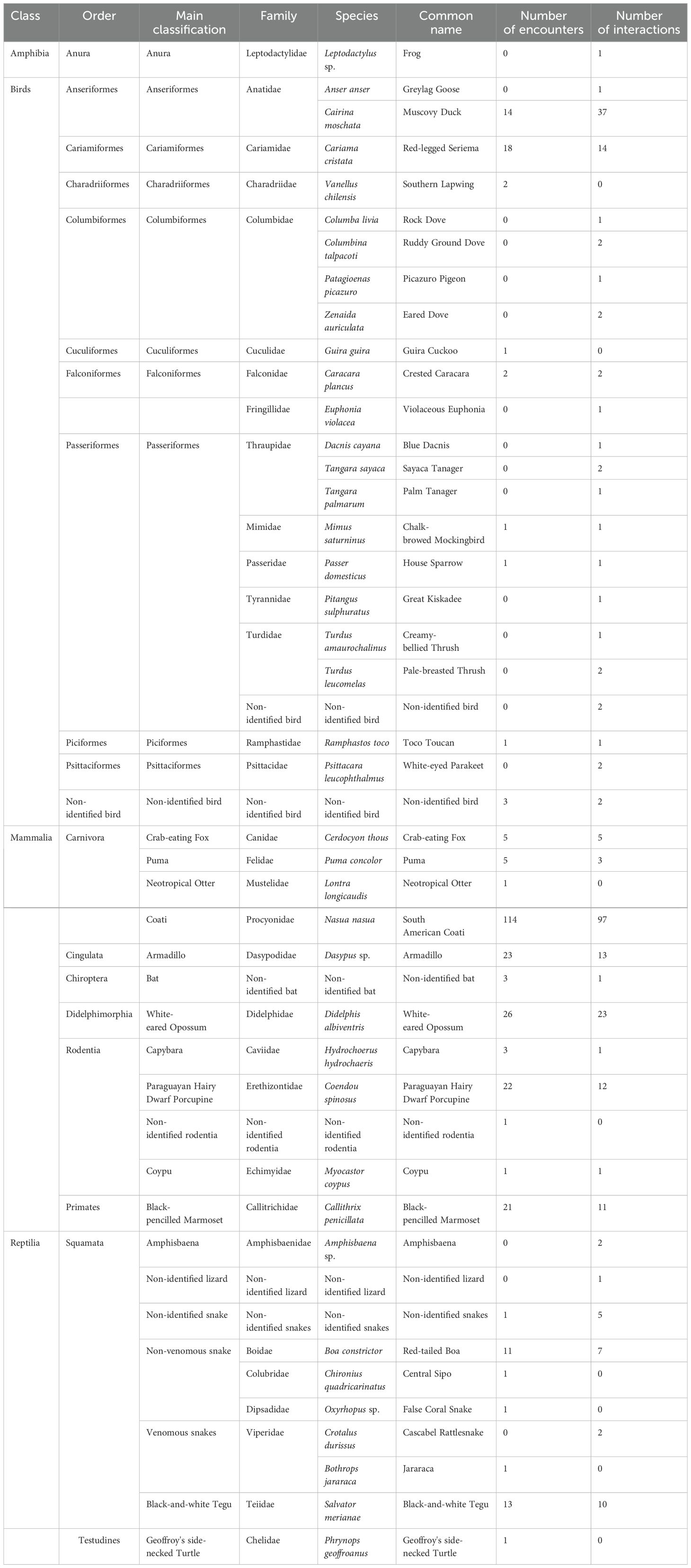
Table 1. List of taxons, classifications, common names, scientific names, number of encounters and number of interactions recorded in the shared landscape.
3.1 Typology of human-wildlife interactions
We identified 16 types of HWI that can occur in shared landscapes, of which eight resulted from direct contact between humans and wildlife, four resulted from indirect contact and five could result from direct or indirect contact (Table 2). Regarding HWI categorization according to their effects on humans and wildlife, 10 were classified as Human-Wildlife Conflict, six as Unsustainable Use, four as Wildlife Damage and three as Convivencia. Between them, six HWI were classified in more than one category, because their effect on humans and animals may vary depending on the situation.
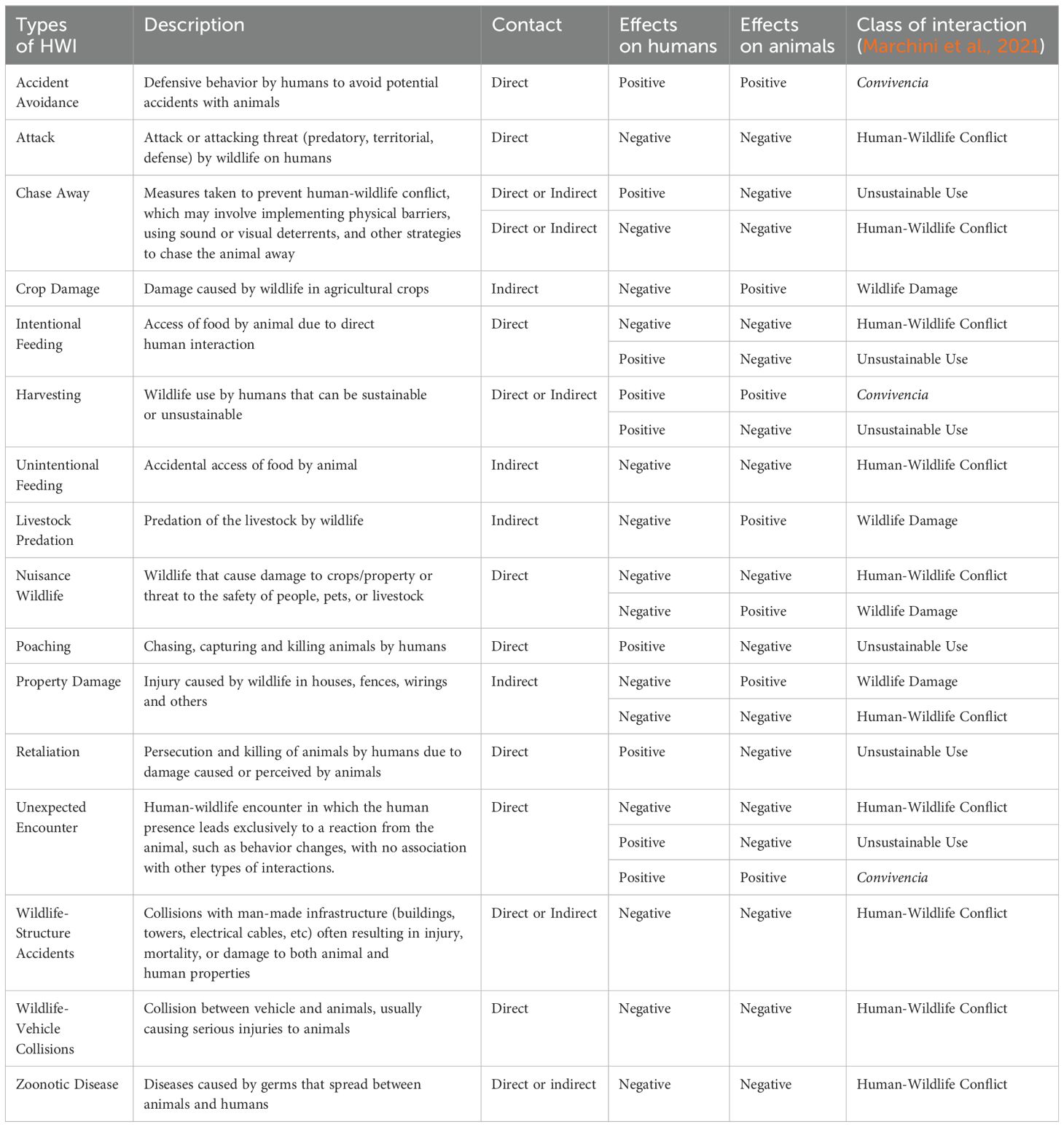
Table 2. Types of human-wildlife interactions (HWI), description, direct/indirect contact, and effects for humans and animals.
3.2 Human-wildlife interactions assessment and evaluation
From the 16 types of HWI that can occur in shared landscapes, we identified ten types of HWI in our study area. The most frequent interaction recorded in our landscape was Unintentional Feeding (34.08%), followed by Unexpected Encounter (15.02%), Accident Avoidance (14.65%), Wildlife-Vehicle Collision (10.62%) and Nuisance Wildlife (10.62%) (Figure 2).
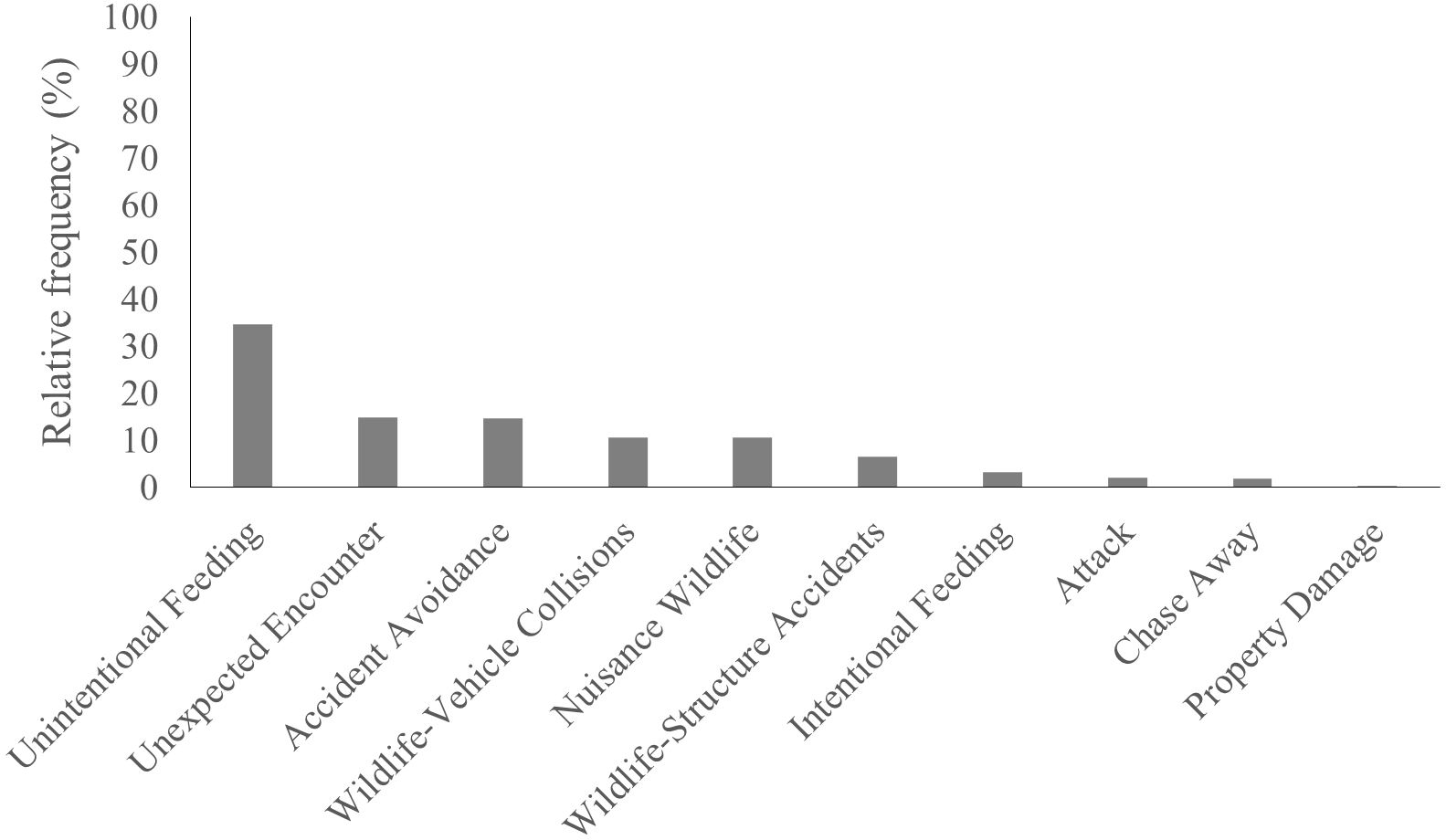
Figure 2. Number of events of each human-wildlife interaction in the shared landscape.
Unintentional Feeding occurred more with coatis (53.68%) and muscovy ducks (21.05%), while Unexpected Encounter (36.59%), Accident Avoidance (30%) and Wildlife-Vehicle Collision (24.14%) were mostly related to coatis. Nuisance Wildlife was equally related to coatis and white-eared opossum (27.59% each) (Figure 3). A result that stood out is that 55% of Nuisance Wildlife events led to emergency rescues, especially with white-eared opossums (N = 6), red-tailed boa (Boa constrictor; N = 4) and coatis (N = 2).
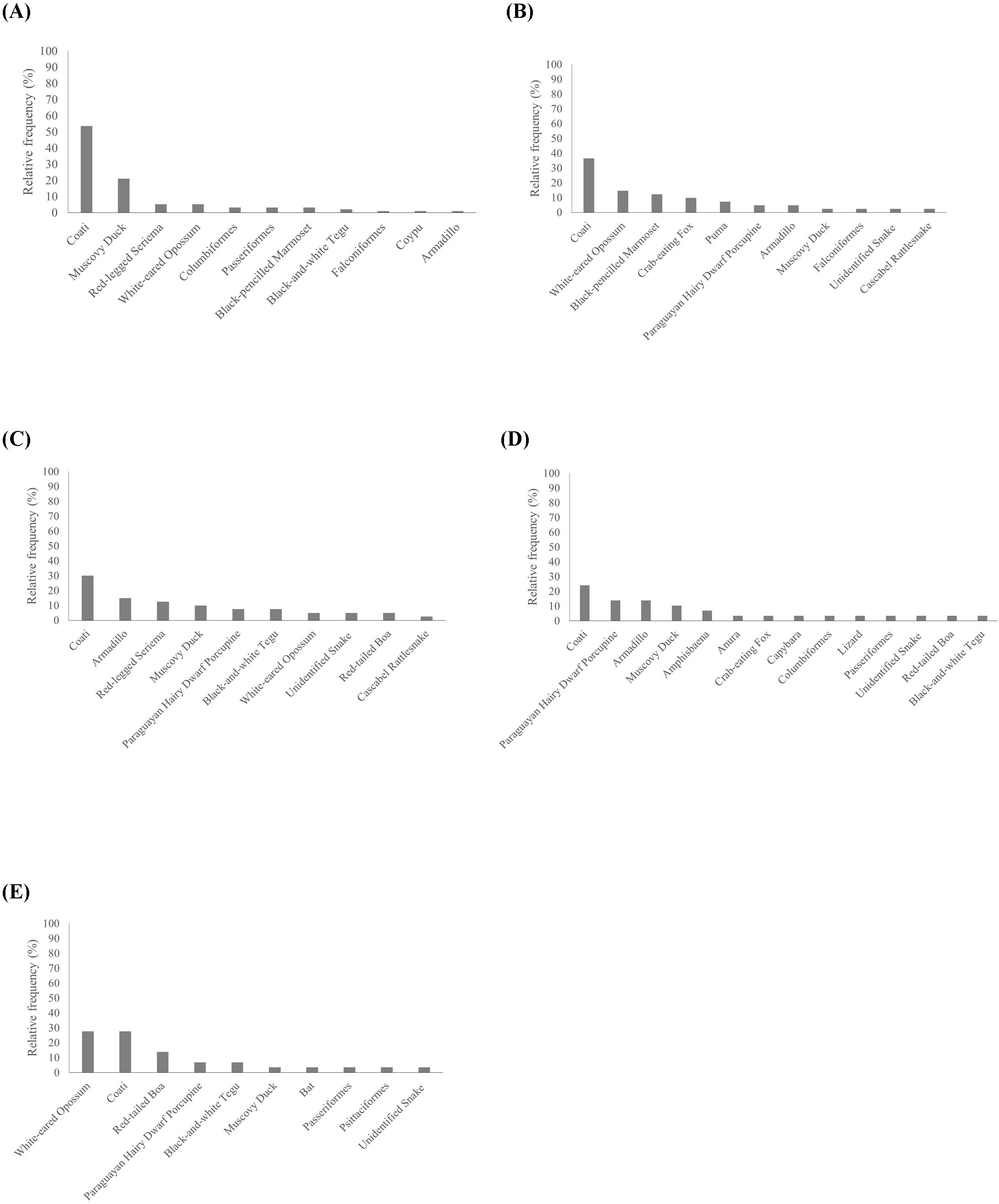
Figure 3. Relative frequency of each taxon for Unintentional Feeding (A), Unexpected Encounter (B), Accident Avoidance (C), Wildlife-Vehicle Collision (D) and Nuisance Wildlife (E) in the shared landscape.
Of the 29 Wildlife-Vehicle Collisions registered, 58.62% involved mammals, 20.69% (N = 6) reptiles, 17.24% (N = 5) birds, and 3.45% (N = 1) amphibians. Of these, 86.20% (N = 25) of the animals were found deceased, while the others either escaped and couldn’t be rescued or were taken to veterinary clinics for treatment. Coatis were the most frequently involved species in this interaction, accounting for 24.14% (N = 7) of the cases, followed by the armadillos (Dasypus sp.) and Paraguayan hairy dwarf porcupines (Coendou spinosus), each comprising 13.79% (N = 4) of the cases. Other taxa involved were Anseriformes (N = 3), Anura (N = 1), Columbiformes (N = 1), Passeriformes (N = 1), non-identified snake (N = 1), red-tailed boa (N = 1), black-and-white tegu (Salvator merianae; N = 1), non-identified lizard (N = 1), capybara (Hydrochoerus hydrochaeris; N = 1), crab-eating fox (Cerdocyon thous; N =1) and amphisbaena (N = 2).
Out of the 40 Accident Avoidance incidents, coatis were involved in the most frequent occurrences, comprising 30% (N =12) of the events. The next more frequent taxa were the armadillos, which accounted for 15% (N = 6), and the Cariamiformes order, represented here by the red-legged seriema (Cariama cristata), which constituted 12.50% (N = 5) of the incidents.
We also identified 18 interactions classified as Wildlife-Structure Accidents, in which 83.33% (N = 15) of the taxons involved were birds, including Anseriformes (N = 1), Cariamiformes (N = 1), Columbiformes (N = 2), Passeriformes (N = 8), Piciformes (N = 1) and non-identified birds (N = 2). The other three Wildlife-Structure Accidents registered happened with mammals: two with coatis and one with the Paraguayan hairy dwarf porcupine. Regarding the birds, 80% (N = 12) of accidents were due to bird-window collisions with glass panes. The other interactions occurred with other man-made infrastructure, such as electrical cables or fences. Considering all the Wildlife-Structure Accidents, in 83.33% (N = 15) of these cases, the animals were already found dead due to these accidents. In the remaining three cases, the animals were transported to a veterinary clinic for necessary care. Unfortunately, the Paraguayan hairy dwarf porcupine did not survive, the toco toucan (Ramphastos toco) couldn’t regain flight due to internal injuries, and the seriema is currently still recovering from treatment.
We classified nine of the interactions identified in the studied shared landscape as Human-Wildlife Conflicts, due to the negative effects for humans and animals, totaling 230 events, of which 41.30% were Unintentional Feeding, 17.39% Unexpected Encounter, 12.61% Wildlife-Vehicle Collision and 12.61% Nuisance Wildlife. We did not record any interactions related to Wildlife Damage (Figure 4).
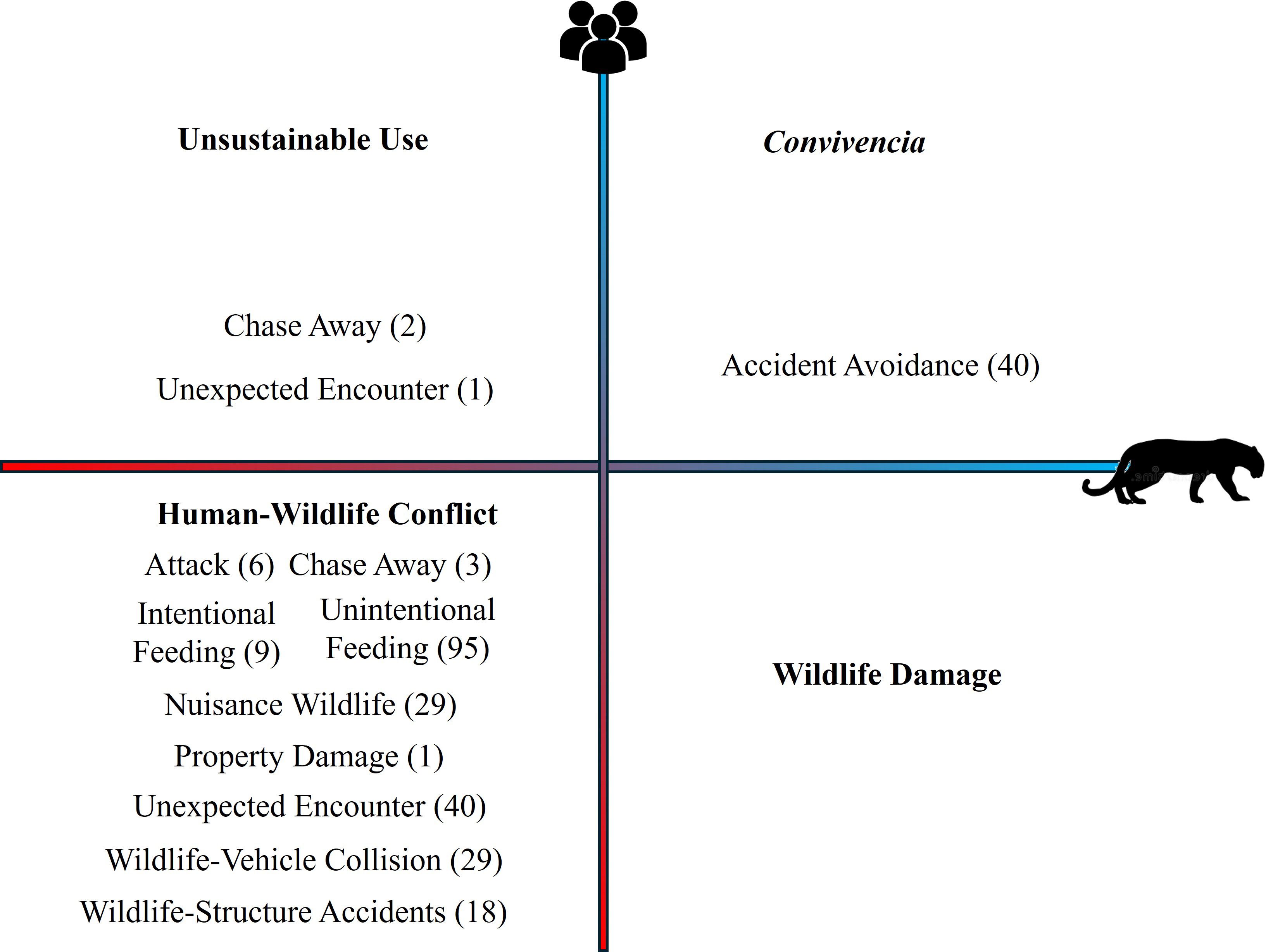
Figure 4. Human-wildlife interactions framed in four categories (Unsustainable Use, Human-Wildlife Conflict, Wildlife Damage, Convivencia) according to the effects under conservation and social perspectives (adapted from Marchini et al., 2021). Red color indicates negative effects. Blue color indicates a positive effect. The number in the parenthesis indicates the number of human-wildlife interactions recorded in the shared landscape.
The most frequent species involved in interactions with humans was the coati (97 human-coati interactions) (Figure 5), being 52.58% due to Unintentional Feeding, 15.46% due to Unexpected Encounter and 12.37% due to Accident Avoidance. The second most frequent species was the muscovy duck (37 human-duck interactions), due to Unintentional Feeding (54.05%), Intentional Feeding (16.22%), Accident Avoidance (10.81%) and Wildlife-Vehicle Collisions (8.11%). The third most frequent species was the white-eared opossum (23 human-opossum interactions), due to Nuisance Wildlife (34.78%), Unexpected Encounter (26.09%) and Unintentional Feeding (21.74%).
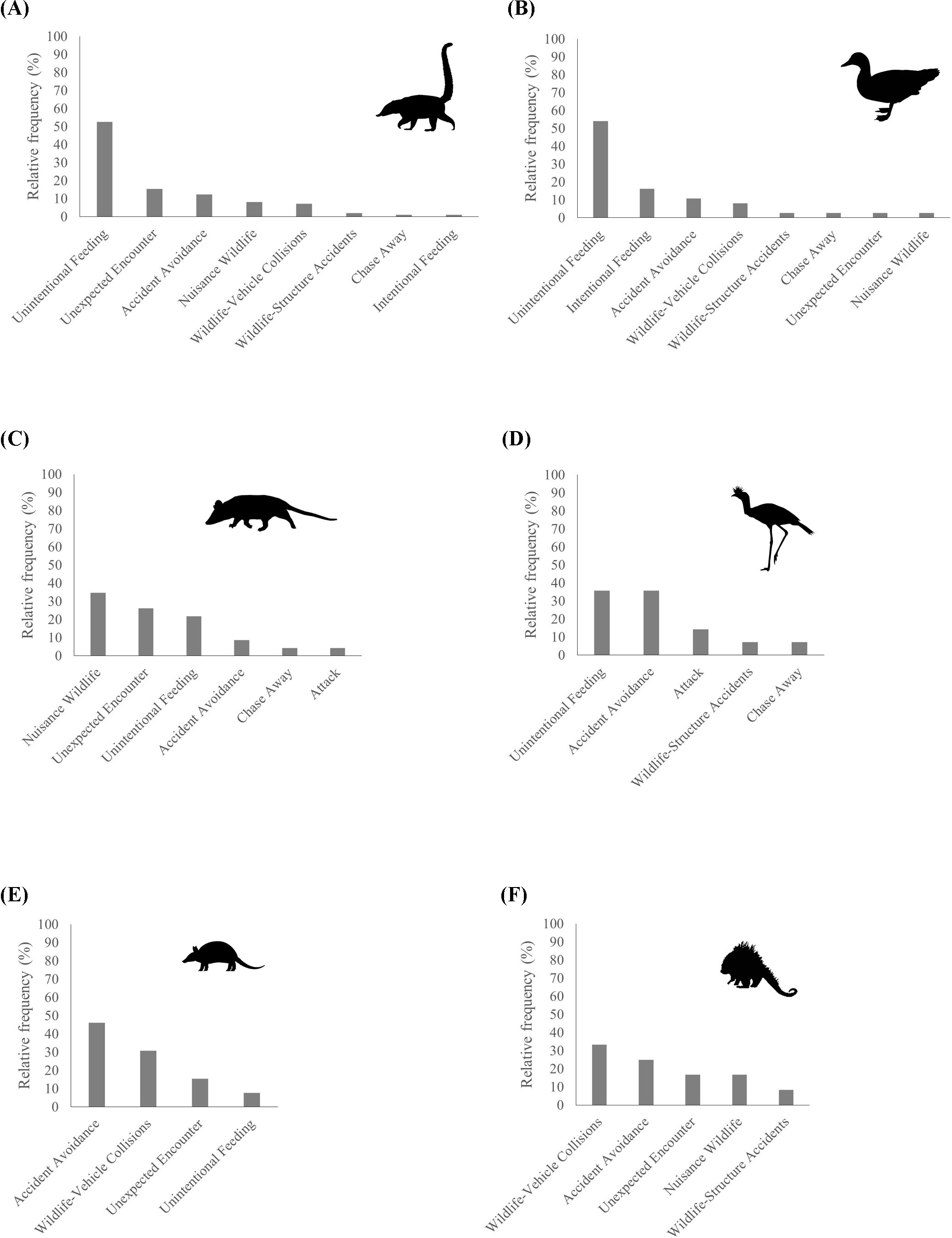
Figure 5. Relative frequency of human-wildlife interactions for coati (Nasua nasua) (A), muscovy duck (Cairina moschata) (B), white-eared opossum (Didelphis albiventris) (C), red-legged seriema (Cariama cristata) (D), armadillo (Dasypus sp.) (E) and Paraguayan hairy dwarf porcupine (Coendou spinosus) (F) in the shared landscape.
The human-coati space of interactions was widely distributed on the campus (Figure 6A), while the human-duck (Figure 6B) and human-opossum (Figure 6C) spaces of interactions were restricted and concentrated on the central lawn of the campus (open green space used for leisure activities). Although, most of the interactions with coatis, ducks and opossums were close to the main buildings, central lawn and cats’ feeding sites. Within these human-coati unintentional feeding interactions, 38 (74.51%) were observed at cats’ feeding sites. For human-opossum unintentional feeding interactions, 3 (60%) of them occurred at cat feeding sites, and for human-duck unintentional feeding interactions, all of them (N = 20; 100%) were related to cat feeding sites. Coatis, ducks and opossums were frequently observed consuming cat food on these sites.
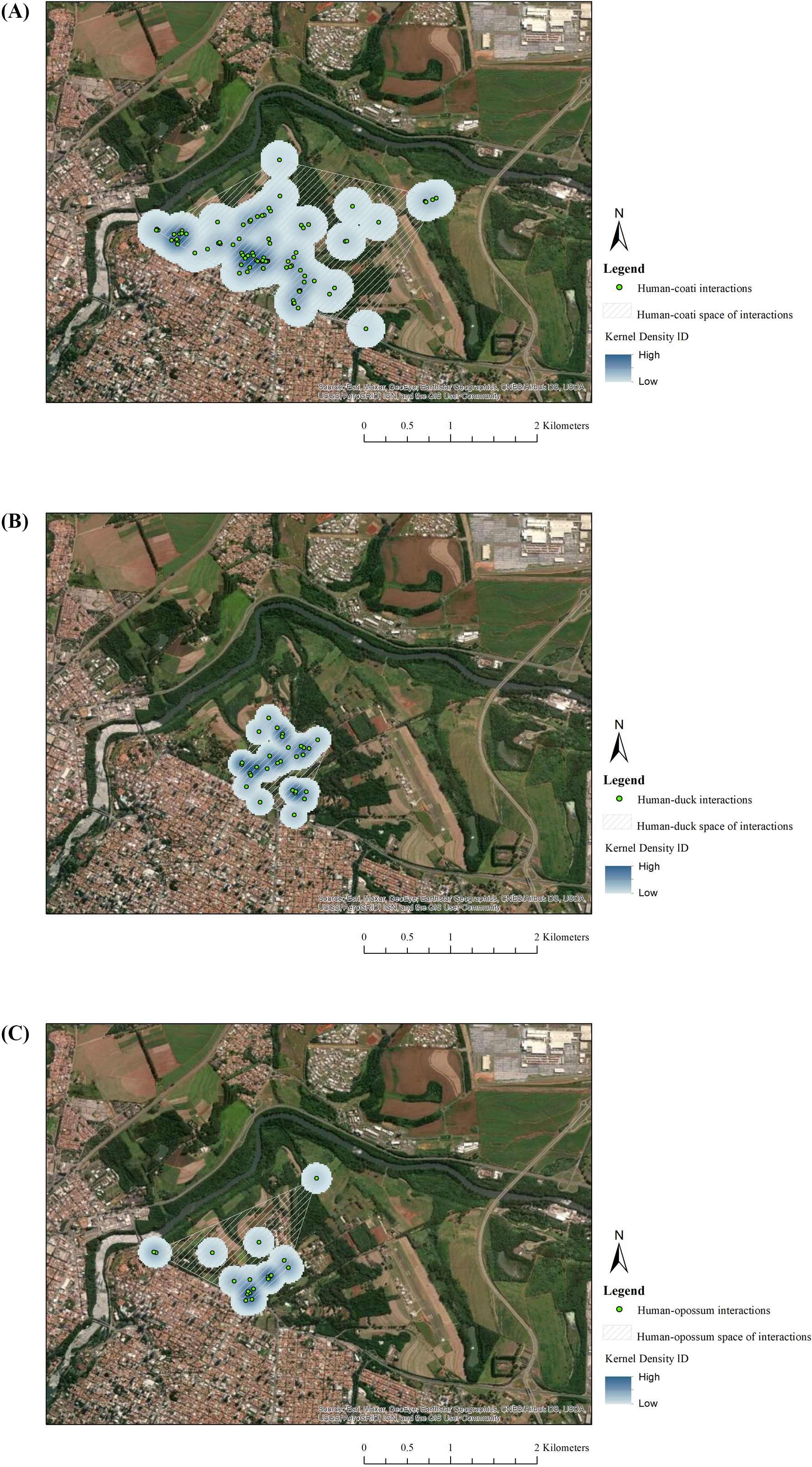
Figure 6. Human-coati interaction density and the space of interaction (A), human-muscovy duck interaction density and the space of interaction (B) and human-white-eared opossum interaction and the space of interaction (C) in the shared landscape.
4 Discussion
More than 80% of interactions recorded in the shared landscape were framed as Human-Wildlife Conflicts because of their negative outcomes, which highlights the enormous challenge for decision-makers in seeking solutions that avoid conflicts or minimize their effects and increase people’s tolerance toward the presence of wildlife. The context of the shared landscape studied explains most of the current conflicts. The Luiz de Queiroz campus is used for multiple purposes such as for teaching, research, sports, leisure activities, agricultural experiments, livestock raising, and wildlife conservation. Garbage cans are distributed throughout the urbanized area, especially close to buildings, open green spaces, restaurants and streets. Most garbage cans are open, accumulating a significant amount of leftover food, especially on weekends, as the campus is one of the most frequented urban green spaces for leisure activities in the region. In addition to the high availability of organic waste, the abundance of cat food at numerous feeding sites is an important attractiveness factor for wildlife, leading to the high frequency of Unintentional Feeding by generalist species. Since Intentional Feeding is neither common nor encouraged practice on the campus, this interaction´s frequency was low.
All the most frequent HWI (Unintentional Feeding, Unexpected Encounter, Accident Avoidance, Wildlife-Vehicle Collision and Nuisance Wildlife) recorded in the area form an intricate web, with elements that are context-dependent and interconnected (Figure 7). The steady and great availability of food is probably the main factor that is attracting wildlife close to humans and to areas with higher human traffic, buildings and vehicle flow, consequently increasing interactions of nuisance and unexpected encounters. It also increases the risk of Wildlife-Vehicle Collisions, Accident Avoidance and Wildlife-Structure Accidents (Basilio et al., 2020; Da Silva et al., 2022). In urban spaces that were not initially planned to considerer wildlife presence and use, accidents involving human structures can become frequent, such as animals getting electrocuted by power lines, trapped in building gaps, or injured on barbed wire fences (Martín et al., 2022). In situations like these, it is necessary to adapt the spaces to the specific needs of each shared landscape, aiming to reduce damage risks for both wildlife and people.
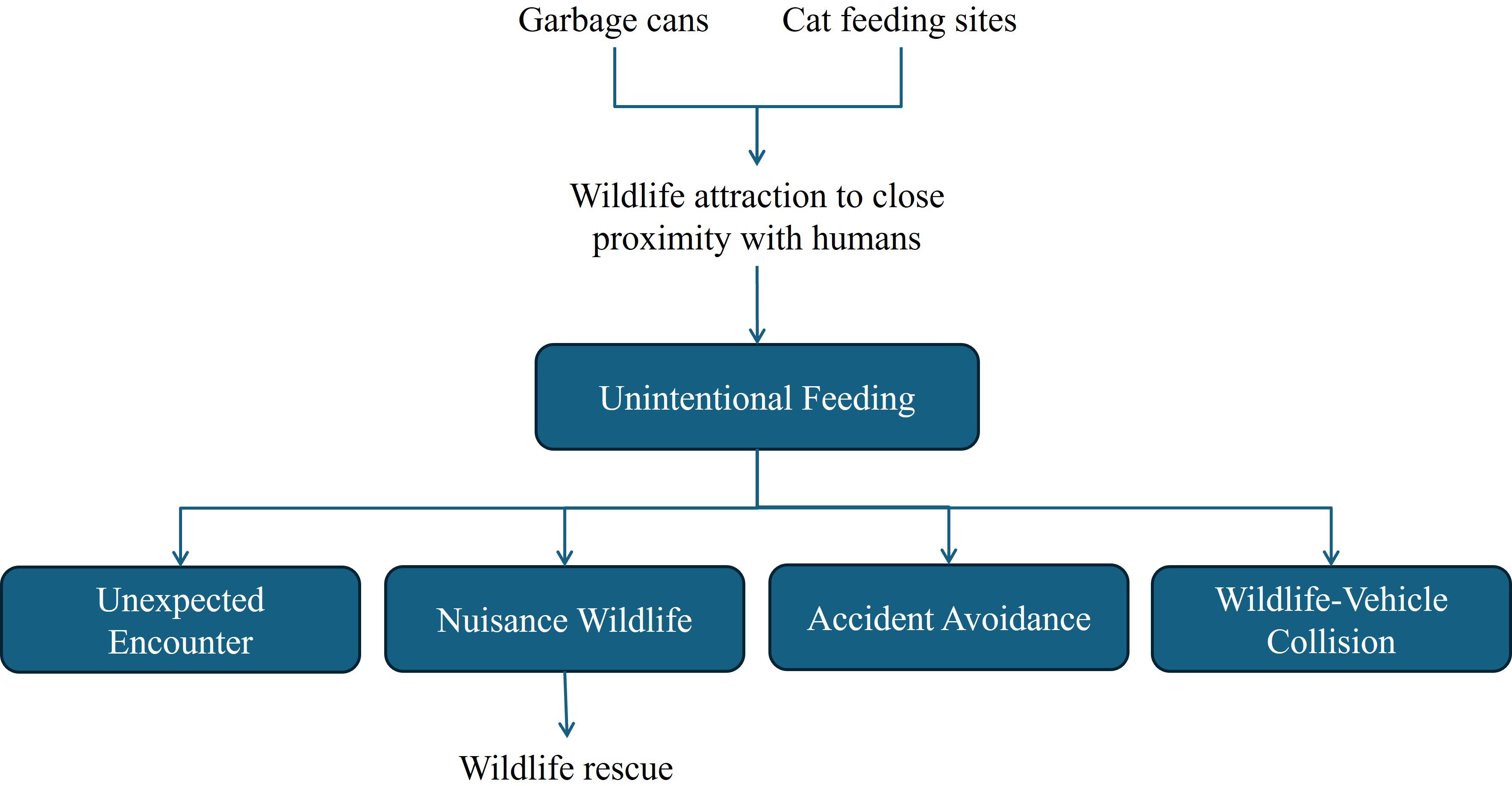
Figure 7. Interrelation between the most frequent human-wildlife interactions in the shared landscape.
Feeding wildlife (intentionally or unintentionally) can profoundly alter natural animal behavior, physiology, reproduction and population levels (Orams, 2002; Griffin and Ciuti, 2023). Evidence suggests that wildlife residing in urban areas may not exhibit the same life history traits as their rural counterparts because of adaptation to human-induced stresses (Ditchkoff et al., 2006). A steady human-supplied food source has a high potential to attract wildlife closer to humans, and habituate them to human presence and urban spaces, which will certainly result in an increase in human-wildlife conflicts like disease transmission, physical attacks, roadkills, property damage, nuisance and others (Soulsbury and White, 2015) thereby demanding conflict resolution.
The great availability of food can stimulate population growth in these animals (Abbas et al., 2011), leading to more interactions and consequently, more conflicts. While adaptation to the anthropogenic environments might initially seem advantageous for animal survival, it can lead to habituation to human-provided food (Blumstein, 2016), with young animals learning to forage in trash bins and cat feeding sites instead of their natural environment, which brings them closer to domestication and the loss of natural foraging behaviors. Furthermore, as they are in fragmented environments, the top-down regulation may not be effective when top predators are at very low densities (Terborgh et al., 2001).
Unexpected Encounter, the second most frequent interaction in the shared landscape, was more related to interactions between humans and coatis, mostly when coatis were moving towards or coming from cat feeding sites. This interaction was the only one with three different possibilities, according to the bidirectional effects, of being classified under social and ecological perspectives (according to Marchini et al., 2021). Especially for this interaction the description of nature must be sufficiently detailed to facilitate its framing as Unsustainable Use, Human-Wildlife Conflict and Convivencia.
Accident Avoidance was the only interaction with positive effects for humans and wildlife as the collision was avoided in all cases. However, it is undeniable that the risk of an accident exists, since the encounters between cars and animals almost resulted in accidents. Therefore, even though this interaction is positive (framed as Convivencia in the diagram), a more effective preventive measure to promote coexistence would be eliminating the risk of accidents. Most accident avoidances and vehicle collisions occurred on streets with intense vehicle circulation within the campus, and close to the main buildings and vegetation. Also, the lower number of Wildlife-Vehicle Collisions involving amphibians and reptiles does not necessarily indicate fewer collisions with these taxa, but rather may reflect the challenge in detecting smaller-bodied specimens.
Our results demonstrated that the coati, followed by the muscovy duck and the white-eared opossum, were the species most frequently involved in interactions with humans. These species have in common a generalist diet (Alves-Costa et al., 2004; Sá et al., 2014; Islam et al., 2020), which facilitates the adaptation of these individuals to anthropogenic landscapes given the steady availability of food sources (Andren, 1994; Gascon et al., 1999; Silva et al., 2005; Kupfer et al., 2006; Devictor et al., 2008). The coati has a broad dietary niche, consuming from fruits to insects and gastropods (Alves-Costa et al., 2004). The muscovy duck is native to South America and has been domesticated in parts of it. In natural environments, the muscovy duck feeds on plants and insects (Islam et al., 2020) through filtration, while in anthropogenic environments its foraging is influenced by the availability of food (Harun et al., 1998; Chapman and Jones, 2009, 2011). In our shared landscape, resident populations of muscovy ducks are mainly found around three artificial lakes, which are high-traffic areas for people, including visitors who feed these animals daily as part of a local cultural tradition. The white-eared opossum is widely distributed in Brazil (Emmons and Feer, 1997; Reis et al., 2014), has solitary and nocturnal habits (Loretto and Vieira, 2005), and feeds from plant material to vertebrates such as bats and snakes (Reis et al., 2014). In this study, this species was frequently observed in garbage cans and on streets near cat’ feeding sites.
Shared landscapes bring even more challenges for coexistence planning and management as conflicts and land-sharing are uncoupled (Crespin and Simonetti, 2019). Human-wildlife conflicts are rampant in a land-sharing context where wildlife co-occurs with humans, crops or livestock, ultimately hindering coexistence if not properly managed (Soulsbury and White, 2015; Crespin and Simonetti, 2019). Unfortunately, the viability of land-sharing/sparing approaches has not considered the importance of coexistence between humans and wildlife (Crespin, 2018). Therefore, efforts to change human behavior may be crucial in promoting coexistence in shared landscapes, like the ability to effectively prevent human-wildlife conflict, to recover from these conflicts and to manage emergency issues precisely (Chen et al., 2023). Also, a key part of shared landscape management is not only to reduce conflicts but also to explore the positive aspects of human-wildlife interactions (Soulsbury and White, 2019).
Indeed, solid waste management significantly impacts the chain of HWI. In this sense, a more appropriate plan for promoting coexistence should consider not only the implementation of wildlife-proof waste disposal structures but also, crucially, environmental education and conflict resolution strategies capable of addressing the collective and individual benefits of behaviors conducive to harmonious interactions with wildlife. At the Luiz de Queiroz campus, dialogue strategies between campus authorities and groups responsible for providing food for abandoned cats have shown promising results in behavior change, as the parties involved have begun to forge new agreements to facilitate coexistence. Following guidance from experts in wildlife conservation, the groups have agreed to gradually relocate the feeding sites away from areas with a high presence of humans, thereby contributing to the reduction of Nuisance Wildlife such as coatis. Measures like these are expected to mitigate the effects of indirect feeding and could influence the chain of HWI that leads to human-wildlife conflicts in this landscape. Further studies about this case are still ongoing, but they underscore the importance of viewing HWI as an interdisciplinary issue that should also be addressed through the lens of social sciences (Soulsbury and White, 2019; Hull et al., 2023). Therefore, by eliminating the causal factors, it is likely that all subsequent interactions will be reduced. Ultimately, the need for emergency wildlife rescues would also decrease. Emergency wildlife rescue is an effective and immediate solution to human-wildlife conflict. Although, it is not sustainable in the long term, and it should not be necessary once coexistence is achieved.
We presented here an approach to assess and evaluate HWI aiming to inform decision-making regarding conflict resolutions to promote coexistence in shared landscapes. This approach should focus on understanding the complexity and diversity of the interactions at multi-species level, providing useful results to guide the interventions that should be made to prioritize and manage HWI with negative outcomes. The HWI assessment and evaluation approach must (i) survey the interactions, (ii) classify the interactions based on their attributes, (iii) map the interactions in the landscape, and (iv) analyze the interactions’ attributes and effects. Based on the results, the decision-maker can prioritize the interaction, species and/or even the location for intervention planning tailored to a specific situation to minimize conflicts, especially in shared landscapes where conflict resolution is more urgent and necessary.
5 Final considerations
The best way to encompass and understand the complexity and diversity of interactions in a shared landscape is to properly assess and evaluate the HWI, as proposed in this paper. Interactions between humans and wildlife occur within a singular and complex context that is crucial for guiding decision-makers with solid, evidence-based solutions. This context encompasses a range of elements that shape and influence such interactions, going beyond isolated situations within the landscape context in which they are embedded (Hull et al., 2023). To provide a more accurate and reliable basis for analysis, we strongly advocate for HWI assessment and evaluation, considering specific factors inherent to each context, such as restored areas, visitor presence, and wildlife attraction factors. Recognizing and understanding this backdrop is essential for developing more effective and sustainable solutions that consider not only the available data, but also the environmental and social context in which these interactions take place.
The nature of interaction is paramount for a more refined analysis. Therefore, the interaction must be described with as many details as possible, so that its effects on people and wildlife can be assessed. This will determine which category the interaction fits into and, consequently, the most appropriate management action. For example, the interaction Chase Away can be situated in two quadrants of the diagram (Marchini et al., 2021), depending on the effect on the person. In general, it is a situation of human-wildlife conflict, as it causes stress both for the person who chases away the animal and for the animal itself. However, we have encountered situations in our landscape where the person who chases away the animal was clearly enjoying the interaction, usually with birds. This type of interaction requires specific management actions, not only focusing on cognitive aspects, such as awareness campaigns, but also on emotional aspects, as the interaction provokes feelings of joy and euphoria in the person. Several studies on moral emotions have made significant contributions to the discussion of a “willingness to act” that leads to actions directed toward the common good (La Taille, 2002, 2010). Incorporating this layer of analysis into studies on HWI also presents great potential.
Other interactions, also related to emotional aspects, require specific actions, such as Intentional Feeding. In many cases, the information that food is harmful to wildlife is not enough, due to the feelings of joy caused by the proximity to the animal. In these cases, it may be more effective to recognize that there is a desire for interaction than to simply prohibit it, suggesting ways of approaching that are not harmful to the animals, such as taking pictures and birdwatching. The emphasis on coexistence, instead of on conflicts, can transform conflicts/risks into opportunities so that humans and wildlife can live in proximity sustainably sharing resources (Jacobs and Vaske, 2019).
All human actions that lead to an increase in food availability for wildlife will certainly result in an increase in human-wildlife conflicts. To improve decision-making in solving conflicts, it is crucial to prioritize actions that address the root cause of the problem in a chain of impacts, identifying and correcting the primary cause of conflict to prevent subsequent effects. Our results highlight the urgency to implement measures to reduce the conflicts related to the presence of wildlife in the proximity of humans in the shared landscape. Therefore, in this context, it is paramount for decision-makers to focus their efforts on tackling a combination of interventions, with multiple and alternative methods (Treves et al., 2009), that accomplish the three realms - landscape, individual, institution and politics (Jacobs, 2006) - of the HWI. Affective aspects will be crucial in this final realm, linking morality to politics to produce agreements and coordinated actions. At landscape level, we recommend (i) replace open by closed garbage cans; (ii) implement mitigation measures in the hotspots of accident avoidance, such as speed reducers, speed enforcement cameras and warning signs, and wildlife crossings; (iii) adopt wildlife-inclusive urban planning and design (Kay et al., 2021). At individual level, we recommend (i) inform and advise users towards a more conscious use of the area, taking away their leftover or disposing them properly in closed garbage cans; (ii) advise the cat feeders about the negative effects of supplying feeding sites with exaggerate amount of cat food in areas with high wildlife diversity; (iii) disseminate information about the negative impacts of feeding wildlife (attacks, disease transmission, nuisance and others); and, (iv) conduct training courses for campus employees and visitors on best practices in coexistence with wildlife. And, at the institutional and political level, we suggest (i) improve regulations and policies for proper solid waste management in wildlife areas; (ii) create guidelines and regulations to promote ‘leave no trace’ practices; (iii) implement protocols for nuisance wildlife and emergency rescues; (iv) create accessible and inexpensive common spaces for conflict resolution among stakeholders. This has proven to be particularly important in the case of cat feeders. While such integrated approaches are unlikely to fully resolve the complex and unique nature of most human-wildlife interactions, they will contribute toward making better decisions while promoting human-wildlife coexistence (König et al., 2021). By adopting appropriate tools and management, public policies, and societal support, people and wildlife can coexist in human-dominated landscapes (Nyhus, 2016).
Mutual adaptation (co-adaptation) between humans and wildlife species is essential to facilitate coexistence in space and time. This means that, to a certain extent, both can change their behavior, learn from experience and pursue their interests concerning each other (Carter and Linell, 2016). This is particularly necessary in urban environments such as the Luiz de Queiroz campus. Promoting a perspective of the common good supported by notions of coexistence makes it possible to produce a pathway to co-adaptation, leading to a more harmonious scenario with wildlife. In this sense, by transforming the attitude of users and visitors in the area into more positive and conscious ones, the related conflicts can be reduced. Therefore, understanding the patterns of interaction through the HWI approach, as part of a process of co-adaptation, is crucial for decision-makers aiming to achieve a scenario of coexistence.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because the data colleted and used in this article was only observations about human-wildlife interactions.
Author contributions
KF: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Writing – original draft, Writing – review & editing. IB: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. AS: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. CN: Data curation, Formal analysis, Methodology, Writing – review & editing. MG: Conceptualization, Formal analysis, Writing – original draft, Writing – review & editing. MP: Conceptualization, Writing – review & editing. LS: Formal analysis, Investigation, Methodology, Writing – review & editing. LC: Formal analysis, Methodology, Writing – review & editing. AG: Data curation, Methodology, Writing – review & editing. JA: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. LA: Data curation, Methodology, Writing – review & editing. SM: Conceptualization, Writing – review & editing. AA: Methodology, Writing – review & editing. VS: Data curation, Methodology, Writing – review & editing. RP: Conceptualization, Investigation, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by Superintendence of Environmental Management/University of São Paulo (SGA-USP), Luiz de Queiroz College of Agriculture, Luiz de Queiroz campus City Hall, and São Paulo Research Foundation (FAPESP) (#2023/11823-0). KF thanks Conselho Nacional de Pesquisa e Desenvolvimento Científico e Tecnológico (CNPq) for the grant (#303940/2021-2). RP thanks SGA-USP for the post-doctoral scholarship received from the USPSusten Program. LA and JA thank the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001.
Acknowledgments
We thank the Forest Science Department, Luiz de Queiroz College of Agriculture, Luiz de Queiroz campus City Hall, and the Superintendence of Environmental Management of the University of São Paulo. We especially thank Ana Maria Meira de Lello, Jefferson Lordello Polizel, Luciano Mendes, Roberta Helena Fiorotto Rodrigues Bacha, Roberto Arruda de Souza Lima for all the logistic support for the Vizinhos Silvestres Project. We thank all the non-academic stakeholders of the Vizinhos Silvestres Project, Isabela Caroline da Silva and Larissa Domingues Orlandin for their contribution in data collection and analysis. We also thank all the collaborators that shared their own HWI observations with our team.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2024.1456072/full#supplementary-material
Footnotes
- ^ Unpublished data. Relatório do Termo de Ajustamento de Conduta apresentado ao Ministério Público, dados do Grupo de Adequação Ambiental do Campus, Escola Superior de Agricultura "Luiz de Queiroz", Universidade de São Paulo, novembro de 2021.
References
Abbas F., Morellet N., Hewison A. J. M., Merlet J., Cargnelutti B., Lourtet B., et al. (2011). Landscape fragmentation generates spatial variation of diet composition and quality in a generalist herbivore. Oecologia 167, 401–411. doi: 10.1007/s00442-011-19tra94-0
Alexandrino E. R., Bovo A. A. A., Luz D. T. A., Costa J. C., Betini G. S., Ferraz K.M.P.M.B., et al. (2013). Aves do campus “Luiz de Queiroz” (Piracicaba, SP) da Universidade de São Paulo: Mais de 10 anos de observações neste ambiente antrópico. Atualidades Ornitológicas. 173, 40–52. doi: 10.11606/9786587391052
Alexandrino E. R., Bovo A. A. A., Moraes F., Betini G. S., Costa J. C., Luz D. T. A., et al. (2021). “Aves do campus “Luiz de Queiroz,” (ESALQ/USP, Piracicaba, SP).
Alves-Costa C. P., Da Fonseca G. A., Christófaro C. (2004). Variation in the diet of the brown-nosed coati (Nasua nasua) in southeastern Brazil. J. mammal. 85, 478–482. doi: 10.1644/1545-1542(2004)085<0478:VITDOT>2.0.CO;2
Andren H. (1994). Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat - a review. Oikos 71, 355–366. doi: 10.2307/3545823
Basilio L. G., Moreno D. J., Piratelli A. J. (2020). Main causes of bird window collisions: a review. An. Acad. Bras. Ciênc. 92, 1–11. doi: 10.1590/0001-3765202020180745
Blumstein D. T. (2016). Habituation and sensitization: new thoughts about old ideas. Anim. behavior. 120, 255–262. doi: 10.1016/j.anbehav.2016.05.012
Bovo A. A. A., Magioli M., Percequillo A. R., Kruszynski C., Alberici V., Mello M. A. R., et al. (2018). Human-modified landscape acts as refuge for mammals in Atlantic Forest. Biota Neotropica 18, e20170395. doi: 10.1590/1676-0611-BN-2017-0395
Brazil (2012). Lei n° 12.651, de 25 de maio de 2012 - Dispõe sobre a proteção da vegetação nativa; altera as Leis n°s 6.938, de 31 de agosto de 1981, 9.393., de 22 de dezembro de 2006; revoga as Leis n°s 4.771, de 15 de setembro de 1965, e 7.754, de 14 de abril de 1989, e a Medida Provisória n° 2.166-67, de 24 de agosto de 2001; e dá outras providências (Brasília: Presidência da República).
Carter N. H., Linell J. D. C. (2016). Co-adaptation is key to coexisting with large carnivores. Trends Ecol. Evol. 31, 575–578. doi: 10.1016/j.tree.2016.05.006
Chapman R., Jones D. N. (2009). Just feeding the ducks: quantifying a common-wildlife interaction. Sunbird. 3, 19–28. Available at: https://www.semanticscholar.org/paper/Just-Feeding-the-Ducks%3A-Quantifying-a-Common-Chapman-Jones/004f3e915144900dab0ec17979530d2609783922. Accessed December 9, 2024.
Chapman R., Jones D. N. (2011). Foraging by native and domestic ducks in urban lakes: Behavioral implications of all that bread. Corella 35, 101–106. Available at: https://www.semanticscholar.org/paper/Foraging-by-native-and-domestic-ducks-in-urban-of-Chapman-Jones/2d7bf3834bdbb48c2fedabbae2b1f02f74592ceb. (Accessed December 9, 2024).
Chen J., Wang Y., Sun J., Wang Y., Li W., Zhang J., et al. (2023). Mitigating the effects of human activities to promote human-wildlife coexistence in urban ecosystems. Sci. Bullet. 68, 2295–2297. doi: 10.1016/j.scib.2023.08.007
Crespin S. (2018). Completing the land-sharing strategy: reaching human-wildlife coexistence through alternative resource management. Universidad de Chile, Chile, Santiago.
Crespin S. J., Simonetti J. A. (2019). Reconciling farming and wild nature: Integrating human–wildlife coexistence into the land-sharing and land-sparing framework. Ambio 48, 131–138. doi: 10.1007/s13280-018-1059-2
Da Silva A. C. F. B., De Menezes J. F. S., Santos L. G. R. O. (2022). Roadkill risk for capybaras in an urban environment. Landscape Urban Planning 222 (104398), 1–7. doi: 10.1016/j.landurbplan.2022.104398
Devictor V., Julliard R., Jiguet F. (2008). Distribution of specialist and generalist species along spatial gradients of habitat disturbance and fragmentation. Oikos 117, 507–514. doi: 10.1111/j.2008.0030-1299.16215.x
Dickman A. J. (2010). Complexities of conflict: the importance of considering social factors for effectively resolving human-wildlife conflict. Anim. Conserv. 13, 458–466. doi: 10.1111/j.1469-1795.2010.00368.x
Ditchkoff S. S., Saalfeld S. T., Gibson C. J. (2006). Animal behavior in urban ecosystems: Modifications due to human-induced stress. Urban Ecosys. 9, 5–12. doi: 10.1007/s11252-006-3262-3
Emmons L. H., Feer F. (1997). Mamíferos da Floresta Neotropical: uma guia de campo. 2. ed (Chicago, IL: University of Chicago Press).
Frank B., Glikman J. A. (2019). “Human-Wildlife conflicts and the need to include coexistence,” in Human-Wildlife interactions: Turning conflict into coexistence. Eds. Frank B., Glikman J. A., Marchini S., 1–19. Cambridge University Press Cambridge: Cambridge University Press (Conservation Biology). doi: 10.1017/9781108235730.004
Gascon C., Thomas E., Richard O., Bierregaard J. R., Malcom J. R., Stouffer P. C., et al. (1999). Matrix habitat and species richness in tropical forest remnants. Biol. Conserv. 91, 223–229. doi: 10.1016/S0006-3207(99)00080-4
Griffin L. L., Ciuti S. (2023). Should we feed wildlife? A call for further research into this recreational activity. Conserv. Sci. Pract. 5, e12958. doi: 10.1111/csp2.12958
Gross E., Jayasinghe N., Brooks A., Polet G., Wadhwa R., Hilderink-Koopmans F. (2021). A Future for All: The Need for Human-Wildlife Coexistence (Gland, Switzerland: WWF).
Harun M. A. S., Veeneklaas R. J., Van Kampen M., Mabasso M. (1998). Breeding Biology of Muscovy duck Cairina moschata in Natural Incubation: The Effect of Nesting Behaviour on Hatchability. Poultry Sci. 77, 1280–1286. doi: 10.1093/ps/77.9.1280
Hull V., Bian X., Episcopio-Sturgeon D. J., Rivera C. J., Rojas-Bonzi V., Morzillo A. T. (2023). Living with wildlife: a review of advances in social-ecological analysis across landscapes. Landscape Ecol. 38, 4385–4402. doi: 10.1007/s10980-023-01778-9
Islam S., Barua K., Ghosh S., Akter N., Sultan N., Islam S., et al. (2020). The Potentials of Raising Muscovy duck (Cairina moschata) in Bangladesh - A Review. Bangladesh J. Vet. Anim. Sci. 8 (2), 146–154. doi: 10.60015/bjvas/V08I2A30
IUCN (2023). IUCN SSC guidelines on human-wildlife conflict and coexistence. 1st ed. (Gland, Switzerland: IUCN).
IUCN SSC Species Survival Commission Human-Wildlife Conflict Task Force (2020). Position Statement on the Management of Human-Wildlife Conflict (Gland, Switzerland: IUCN).
Jacobs M. (2006). The Production of Mindscapes: A Comprehensive Theory of Landscape Experience (Wageningen University and Research). Awarding Institution: Wageningen University Place of Publication [S.l.]. doi: 10.18174/40182
Jacobs M., Vaske J. J. (2019). “Understanding Emotions as Opportunities for and Barriers to Coexistence with Wildlife,” in Human-Wildlife interactions: Turning conflict into coexistence. Eds. Frank B., Glikman J. A., Marchini S. (Cambridge University Press, 1–19). doi: 10.1017/9781108235730.004
Kay C. A. M., Rohnke A. T., Sander H. A., Stankowich T., Fidino M., Murray M. H., et al. (2021). Barriers to building wildlife-inclusive cities: insights from the deliberations of urban ecologists, urban planners and landscape designs. People Nature. 2, 62–70. doi: 10.1002/pan3.10283
Knight J. (2000). Natural enemies: people-wildlife conflicts in anthropological perspective (London and New York: Routledge).
König H. J., Carter N., Ceauşu S., Lamb C., Ford A. T., Kiffner C. (2021). Human-wildlife coexistence in science and practice. Conserv. Sci. Pract. 3, 1–5. doi: 10.1111/csp2.401
Kupfer J. A., Malanson G. P., Franklin S. B. (2006). Not seeing the ocean for the islands: the mediating influence of matrix-based processes on forest fragmentation effects. Global Ecol. Biogeogr. 15, 8–20. doi: 10.1111/j.1466-822X.2006.00204.x
La Taille Y. (2002). Shame and its relationships with morality. Psicol.: Reflexão e Crítica. 15, 13–25. doi: 10.1590/S0102-79722002000100003
La Taille Y. (2010). Moral and Ethic: a psychological reading. Psicol.: Teoria e Pesquisa (special issue). 26, 105–114. doi: 10.1590/S0102-37722010000500009
Loretto D., Vieira M. V. (2005). The effects of reproductive and climatic seasons on movements in the Black-Eared Opossum (Didelphis aurita Wied-Neuwied 1826). J. Mammal. 86, 287–293. doi: 10.1644/BEH-117.1
Marchini S., Ferraz K.M.P.M B., Foster V., Reginato T., Kotz A., Barros Y., et al. (2021). Planning for human-wildlife coexistence: conceptual framework, workshop process, and a model for transdisciplinary collaboration. Front. Conserv. Sci. 2. doi: 10.3389/fcosc.2021.752953
Martín J. M., López J. R. G., Sousa H. C., Barrios V. (2022). Wildlife and power lines: Guidelines for preventing and mitigating wildlife mortality associated with electricity distribution networks (Gland, Switzerland: IUCN).
Nyhus P. H. (2016). Human-wildlife conflict and coexistence. Annu. Rev. Environ. Res. 41, 143–147. doi: 10.1146/annurev-environ-110615-085634
Orams M. B. (2002). Feeding wildlife as a tourism attraction: a review of issues and impacts. Tourist Manage. 23, 281–293. doi: 10.1016/S0261-5177(01)00080-2
Pooley S., Bhatia S., Vasava A. (2020). Rethinking the study of human-wildlife coexistence. Conserv. Biol. 35, 784–793. doi: 10.1111/cobi.13653
Reis N. R., Fregonezi M. N., Perachi A. L., Shibatta O. A., Sartore E. R., Rossaneis B. K., et al. (2014). Mamíferos Terrestres de Médio e Grande Porte da Mata Atlântica: guia de campo. 1 ed (Rio de Janeiro: Technical Books).
Sá E. F. G. G., Mendes D. A. S., Cheung K. C. (2014). Levantamento e caracterização de Didelphis albiventris (Lund 1840) em fragmentos de Cerrado na Fazenda Cervinho, Bandeirantes, MS, Brasil. Multitemas 45, 83–102. Available at: https://www.multitemas.ucdb.br/multitemas/article/view/237. (Accessed December 9, 2024).
Schell C. J., Stanton L. A., Young J. K., Angeloni L. M., Lambert J. E., Breck S. W., et al. (2020). The evolutionary consequences of human–wildlife conflict in cities. Evolution. Applications. 14, 178–197. doi: 10.1111/eva.13131
Silva M., Hartling L., Opps S. B. (2005). Small mammals in agricultural landscapes of Prince Edward Island (Canada): effects of habitat characteristics at three different spatial scales. Biol. Conserv. 126, 556–568. doi: 10.1016/j.biocon.2005.07.007
Soulsbury C. D., White P. C. (2015). Human–wildlife interactions in urban areas: a review of conflicts, benefits and opportunities. Wildl. Res. 42, 541–553. doi: 10.1071/WR14229
Soulsbury C. D., White P. C. (2019). “A framework for assessing and quantifying human-wildlife interactions in urban areas,” in Human-Wildlife interactions: Turning conflict into coexistence, vol. 1-19 . Eds. Frank B., Glikman J. A., Marchini S., 107–128. Cambridge University Press Cambridge: Cambridge University Press (Conservation Biology).
Terborgh J., Lopez L., Nuñez P., Rao M., Shahabuddin G., Orihuela G., et al. (2001). Ecological meltdown in predator-free forest fragments. Science 294, 1923–1926. doi: 10.1126/science.1064397
Treves A., Wallace R. B., White S. (2009). Participatory planning of interventions to mitigate human-wildlife conflicts. Conserv. Biol. 23, 1577–1587. doi: 10.1111/j.1523-1739.2009.01242.x
White R. M., Fischer A., Marshall K., Travis J. M., Webb T. J., Di Falco S., et al. (2009). Developing an integrated conceptual framework to understand biodiversity conflicts. Land Use policy. 26, 242–253. doi: 10.1016/j.landusepol.2008.03.005
Whitehouse-Tedd K., Abell J., Dunn A. K. (2021). House Evaluation of the use of psychometric scales in human-wildlife interaction research to determine attitudes and tolerance toward wildlife. Conserv. Biol. 35, 533–547. doi: 10.1111/cobi.13599
Keywords: feeding wildlife, human-wildlife conflicts, nuisance wildlife, urban space, wildlife
Citation: Ferraz KMPMdB, Bento IdF, Di Souza ABQ, Nunes CdS, Guimarães MAdM, Pereira MS, da Silva LL, de Campos LKN, Gobbi AS, Alves JCQ, Alvarez LFP, Marchini S, de Aquino ACMM, Sato VKdM and Paolino RM (2025) Assessing and evaluating human-wildlife interactions for coexistence in shared landscapes. Front. Conserv. Sci. 5:1456072. doi: 10.3389/fcosc.2024.1456072
Received: 27 June 2024; Accepted: 29 November 2024;
Published: 06 January 2025.
Edited by:
Silvio J. Crespin, University of Concepcion, ChileReviewed by:
Halina Teresa Kobryn, Murdoch University, AustraliaAndrea Presotto, East Carolina University, United States
Copyright © 2025 Ferraz, Bento, Di Souza, Nunes, Guimarães, Pereira, da Silva, de Campos, Gobbi, Alves, Alvarez, Marchini, de Aquino, Sato and Paolino. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Katia Maria Paschoaletto Micchi de Barros Ferraz, a2F0aWEuZmVycmF6QHVzcC5icg==