
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Conserv. Sci., 12 December 2022
Sec. Human-Wildlife Interactions
Volume 3 - 2022 | https://doi.org/10.3389/fcosc.2022.989368
This article is part of the Research TopicCoexistence Between Conservation and Food Security in Social-Ecological SystemsView all 5 articles
 Christian Kiffner1,2*
Christian Kiffner1,2* Sandra Uthes2
Sandra Uthes2 Emu-Felicitas Ostermann-Miyashita1,3
Emu-Felicitas Ostermann-Miyashita1,3 Verena Harms4
Verena Harms4 Hannes J. König1,3
Hannes J. König1,3Predation on livestock presents a daunting challenge for human–carnivore coexistence in agricultural landscapes. In Germany, the recolonization of wolves is ongoing and its consequences are insufficiently understood. Knowledge about which livestock species are susceptible to wolf predation, which farm types are predisposed to attacks by wolves, and when predation on livestock occurs is valuable for mitigating stakeholder conflicts. To this end, we analyzed 14 years of monitoring data and assessed the livestock prey spectrum, identified correlates between predation on livestock, farm type and livestock category, and described temporal patterns of livestock loss caused by a recolonizing wolf population in the state of Brandenburg (Germany). Among a total of 1387 recorded cases, 42% were unequivocally attributed to wolves (SCALP criteria C1 and C2) and 12% of cases were not caused by wolves. The number of head of livestock killed during a single wolf attack was mediated by farm type and livestock species; losses per event were greater in full-time farms vs. other farm types and greater in sheep, farmed deer and other livestock species, compared to cattle. While sheep were the most commonly killed livestock species, the increase in wolf territories over the investigation period was associated with a widening of the domestic prey species spectrum. Count regression models provided evidence for the increasing frequency of predation events over the 14-year period, along with an exponential increase in wolf territories. Predation on livestock occurred throughout the year, yet seasonality of events was evident and differed across livestock categories. Predation on sheep peaked in the fall, coinciding with the post-weaning period of wolf offspring. Predation on cattle peaked in the spring, coinciding with the cattle calving period. These results call for renewed investment in the implementation of prevention methods for all susceptible domestic species, particularly during times of elevated predation risk.
The recovery of large carnivore populations in cultural landscapes of Europe is a remarkable wildlife conservation success story (Chapron et al., 2014; Cimatti et al., 2021), and yet presents daunting challenges for human–carnivore coexistence in agricultural landscapes (Nyhus, 2016; Pimenta et al., 2018; König et al., 2020; Linnell et al., 2020; Pettersson et al., 2021). Predation on livestock by large carnivores such as grey wolves (Canis lupus; hereafter: wolf/wolves) represents a persistent problem both in areas recently recolonized by wolves and in those with well-established wolf populations (Ciucci and Boitani, 1998; Kaczensky, 1999; Gula, 2008; Gervasi et al., 2021a; Gervasi et al., 2021b; Fedyń et al., 2022; Khorozyan and Heurich, 2022). Predation on livestock can perpetuate or even aggravate the already predominantly negative attitudes towards wolves among parts of society (Dressel et al., 2015; Jürgens and Hacket, 2017). From the perspective of livestock herders, predation on livestock causes direct costs, i.e. loss of livestock, and indirect costs, i.e. opportunity and transaction costs associated with prevention and management of livestock loss (Barua et al., 2013; Widman and Elofsson, 2018; Widman et al., 2019). Recurring livestock loss due to wolf attacks are an often stated justification for lethal removal of (typically strictly protected) wolves (Naughton-Treves and Treves, 2005; Treves et al., 2016; Kiffner et al., 2019).
However, before such drastic – and, in the long term, often ineffective (Harper et al., 2008; Wielgus and Peebles, 2014; Treves et al., 2016; Eklund et al., 2017; Khorozyan and Waltert, 2019b) – methods are implemented, it is worthwhile to disentangle correlates of predation on livestock. Although cause and effect are ideally tested in randomized and more robust study designs (Treves et al., 2019; Khorozyan, 2022), identifying correlates of livestock predation could greatly contribute to guiding evidence-based prevention methods (van Eeden et al., 2018) and hence reduce predation on livestock and resulting stakeholder conflicts (Mayer et al., 2022). Here, we focus on this task by using monitoring data from Brandenburg, a German state with a large and rapidly expanding wolf population (Reinhardt et al., 2019). A first step towards this end is to separate reported livestock losses caused by wolves from reported losses due to other causes. Research suggests that the causes of livestock mortality are occasionally misidentified and incorrectly attributed to wolves. Indeed, livestock losses can also be caused by diseases, domestic dogs (Ciucci and Boitani, 1998) or other wild carnivore species, such as red foxes (Vulpes vulpes) (Patalano and Lovari, 1993). Using clear criteria for verification (Molinari-Jobin et al., 2021) and subsequently focusing on analyzing confirmed events allows identifying robust correlates of predation events by wolves.
One of the most fundamental aspects of the livestock-carnivore interface is to identify which livestock species are susceptible to predation by free ranging carnivores. In the European context, research on livestock predation by large carnivores often focuses on sheep (Ovis aries) (Gervasi et al., 2021a; Khorozyan and Heurich, 2022). However, a rich body of research indicates that wolves also prey on cattle (Bos taurus), farmed deer (e.g. fallow deer Dama dama and red deer Cervus elaphus), and other domestic mammal species (Treves et al., 2002; Musiani et al., 2003; Iliopoulos et al., 2009; Hanley et al., 2018; Pimenta et al., 2018; Fedyń et al., 2022), reflecting the wide food niche of wolves (Jedrzejewski et al., 2012). Based on the positive correlation between prey species richness and food niche breadth observed in wolves (Okarma, 1995), and assuming that an expanding wolf population encounters a greater diversity of potential prey species over time, we expected that the domestic prey spectrum would increase over time.
Another basic, yet important aspect is to assess patterns and correlates for the number of killed livestock per predation event. Wolves occasionally kill more than a single head of livestock per predation event, a phenomenon which is termed “surplus” or “mass” killing (Kruuk, 1972; Delgiudice, 1998; Fedyń et al., 2022). Identifying correlates for this wolf predation behavior is relevant to the issue of management because the financial damage is clearly linked to the number of lost livestock. Moreover, this wolf behavior may be interpreted as intentional and as a menace, thus reinforcing the negative image of wolves among parts of society (Jürgens, 2022). However, multiple biologically plausible hypotheses have been proposed to explain this wolf predation behavior. In this context, we hypothesized that the number of livestock individuals killed per event is a phenomenon primarily found in smaller-bodied livestock species (Gazzola et al., 2008). Moreover, the farm type could influence the number of killed livestock individuals via factors such as flock size of livestock (Eklund et al., 2017) and the implementation of preventive methods (such as appropriately installed electrified fences and livestock guardian dogs) to reduce the impact of wolf predation events (Reinhardt et al., 2012).
Finally, predicting when livestock are most at risk from predation is highly relevant to the issue of management. To date, predicting predation risk imposed by expanding wolf populations has mostly been addressed by using spatial models (Mech et al., 2000; Treves et al., 2004; Dondina et al., 2015). While such models can be accurate in spatially predicting predation risk (Treves et al., 2011; Treves and Rabenhorst, 2017), mechanistic and spatially-explicit predation risk models typically require detailed information on the distribution of livestock, alternative wild prey species and carnivores (Khorozyan et al., 2015; Janeiro-Otero et al., 2020; Bautista et al., 2021; Kuiper et al., 2022). These data are often not available at large spatial scales. However, the temporal dimension of livestock loses associated with an expanding wolf population can provide important insights for reducing the frequency of livestock predation on livestock (Fedyń et al., 2022; Khorozyan and Heurich, 2022). While it is plausible that predation events scale with range expansion of wolves (Harper et al., 2005; Khorozyan and Heurich, 2022), the frequency of predation events is not always related to the relative abundance of wolves (Gervasi et al., 2021a). Moreover, knowledge about seasonal dynamics of predation on livestock is valuable for guiding preventive interventions aimed at reducing its frequency. Multiple hypotheses have been put forward to explain seasonality in predation on livestock, including variation in livestock availability, variable susceptibility of livestock species (e.g. large-bodied species may be especially susceptible to wolf predation during the calving period), and variation in nutritional demands of wolf packs – such as elevated energy demand during weaning (Musiani et al., 2005; Iliopoulos et al., 2009). To date, these temporal patterns have not been assessed for wolf predation on livestock in Germany.
Drawing upon a long-term database (spanning 14 years) from Brandenburg, the state with the largest wolf population in Germany, and one that is still expanding (Reinhardt et al., 2019), we first describe patterns of livestock predation reports and then test the previously outlined hypotheses in relation to (1) domestic prey species spectrum, (2) number of killed livestock per event, and (3) temporal trends of predation on livestock. Finally, we discuss these results in relation to human-carnivore coexistence in agricultural landscapes.
We conducted this case study in the state of Brandenburg (hereafter: Brandenburg), Germany. Located in the northeastern part of Germany, Brandenburg covers a surface area of 29,654 km2. Approximately half of the area (48.6%) is used for agriculture; forests cover slightly more than one third (34.8%) of the area. The remaining surface area (combined 16.6%) is covered by settlements, roads, water bodies and other vegetation types (AFS, 2021). The terrain is mostly flat or slightly undulating and the sandy or sandy loamy soils are relatively poor. In combination with comparatively low amounts of annual precipitation (~550 mm year-1), cash crop production is challenging compared to in other parts of Germany (MIL, 2010). Livestock husbandry is an important element in rural areas, ranging from livestock kept for leisure (hereafter: hobby), livestock farms managed to augment income (hereafter: part-time), specialized farms for livestock production (hereafter: full-time), to the keeping of animals for educational or research purposes (hereafter: other). Livestock keeping on fenced pastures is mostly dominated by cattle and sheep. In Germany, the general approach to livestock keeping is broadly similar across the different farm categories: livestock are kept in fenced paddocks during day and night and farmers typically attend to the livestock on a daily basis for a limited amount of time. Generally, paddocks have open shelters which are not predator-proof; fencing usually includes mobile electric fences and (barbed) wire fences. Full-time farmers who specialize in sheep typically move their herd to grazing areas (such as landscape conservation areas) where they receive payments for prescribed grazing during the vegetation period. In these areas, sheep are either kept in areas fenced with mobile electric fences or herded with the help of herding or livestock guardian dogs. While there is likely some variability in terms of herding practices even within the farm types, the recorded farming type is likely associated with herd sizes. Although these data are not available, we assume that full-time farms have larger herd sizes, part-time and other farm types have intermediate herd sizes and hobby farms mostly have small herd sizes.
Often, income through small ruminant (i.e. sheep and goat) production (e.g. meat, milk, and wool) is financially not viable; owners of small ruminant herds generate a substantial part of their income for grazing their livestock as part of landscape management efforts (Peters, 1997). In some areas, farmed deer and European mouflon (Ovis gmelini musimon) are kept for either venison production or for educational purposes.
Under the European Habitat Directive, the member states of the European Union are required to monitor the status of wolf populations and associated predation events (Marucco and Boitani, 2012); in Germany, the individual states are responsible for implementation of these measures.
As background information on wolf predation on livestock, we obtained wolf monitoring data from the German Wolf Monitoring database (https://www.dbb-wolf.de/Wolfsvorkommen/territorien/zusammenfassung). These publicly available data include annual numbers of (1) wolf packs, (2) wolf pairs, and (3) territorial individuals.
To describe the wolf population over time, we aggregated these data to the number of wolf territories [sum of (1) to (3)] per year. The wolf monitoring database considers May 1 through April 30 of the consecutive year as one monitoring year. During the time period of this study (2007-2020), the number of wolf territories increased exponentially (Figure 1); a log-linear model described the number of wolf territories well (F1,11 = 225.2; p-value>0.001; Adj. R²=0.95):
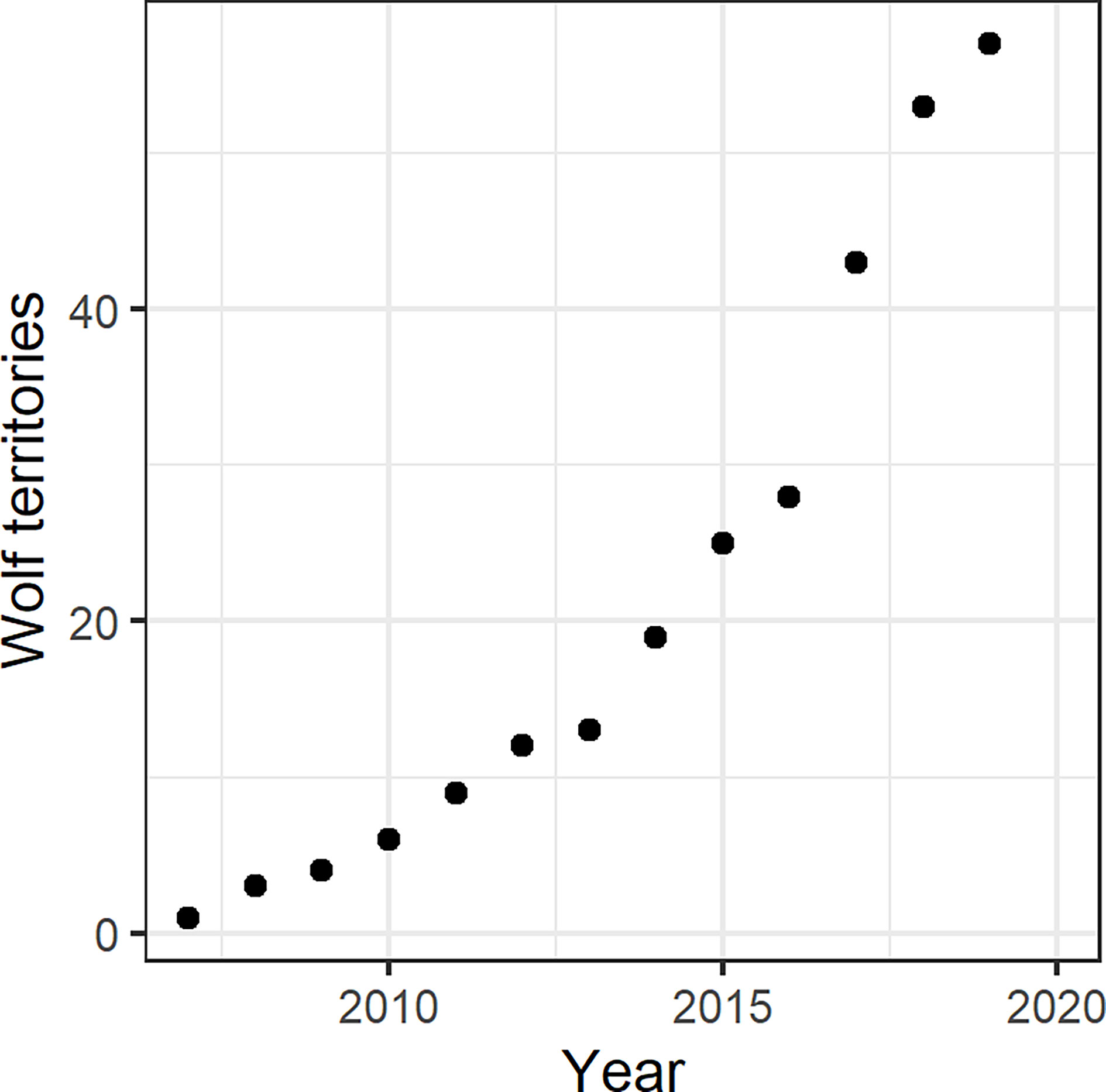
Figure 1 Number of wolf territories (sum of wolf packs, wolf pairs and territorial wolves) in Brandenburg State (Germany) from 2007/2008 to 2019/2020. The years are based on the wolf monitoring calendar (May 1 through April 30 of the following year).
For this study we used data from 01/01/2007 to 31/12/2020. Registered livestock keepers are eligible for compensation of costs associated with confirmed livestock loss caused by wolves. The compensation scheme has been in place since 2007, yet has undergone some changes. Initially, compensation was facilitated by the “de minimis aid” (i.e. small amount of state aid that does not have to be reported to the European Commission) and was limited to € 15,000 per farm over a three-year period. With the introduction of a compensation policy and subsequent notification obligation to the European Commission in 2014, the financial compensation limit per farm was removed. Since then, minimum standards for livestock protection have been required in order to receive compensation payments. Specifics of these minimum standards differ by livestock species, yet generally include sufficiently tall and electrified fences. Unfortunately, no attempts have been made by the corresponding authority to assess potential impacts of changes in the compensation scheme on number of predation events.
The main criteria for successfully claiming compensation hinges on an independent assessment to confirm that wolves caused the livestock loss. As there is some uncertainty involved in this step (see below), losses where wolves cannot be excluded as mortality cause, are also compensated. If compensation is granted, farmers receive payments according to the market value of the killed animals. Therefore, farmers who suspect that livestock losses are due to wolf predation have a financial incentive to report such events. However, we cannot exclude the possibility that some livestock keepers did not report predation events. If livestock owners suspect attacks by wolves, they contact independent assessors. These assessors typically visit the site on the same or the following day and assess whether the livestock loss was caused by wolves or by other causes. Site assessments include a search for signs of wolf presence at and around the carcass (e.g. bite marks on the carcass, wolf spoor in the surroundings) as well as swab sample collection at bite sites of the carcass to collect saliva traces of the species that killed the livestock. These swab samples are typically sent to the Conservation Genetics lab at the Senckenberg Research Institute for DNA extraction and PCR-based confirmation of wolf DNA from the swab samples (Harms et al., 2015). In few cases (e.g. adult cattle, horses and other particularly valuable animals), carcasses are transported off site and are subject to an autopsy at the “Landeslabor Berlin-Brandenburg”. The resulting data from the site visit and subsequent investigations are consolidated in a central database. Based on the information provided in this database, we assigned the causes of livestock losses using the SCALP criteria. Initially developed by the project “Status and Conservation of the Alpine Lynx Population (SCALP)” to categorise sightings of lynx (Lynx lynx) (Molinari-Jobin et al., 2021), these criteria are also used for evidence of wolves (https://www.dbb-wolf.de/wolf-management//scalp-criteria) and fall under five different categories:
● C1: records with confirmed hard facts with material evidence such as genetically identifiable material (e.g. hair, excrement) of wolves1
● C2: records such as prey killed by the species or tracks, which have been verified by an expert
● C3: unconfirmed records, where field signs indicative of wolves were too old or poorly visible
● NP: events which were not possible to be verified (due to absence of tracks or where DNA tests failed), were classified as not possible
● False: if the cause of mortality was clearly attributable to a different cause (e.g. disease)
To describe patterns of reported livestock losses, we investigated how the number of records per category was distributed across livestock species and years. Due to the relatively large number of livestock species in the database, we consolidated them into four categories for most of our analyses: sheep, cattle, farmed deer, and other species. In few cases, wolves killed two (n=31) or four (n=1) livestock species; to account for the livestock species identity, we duplicated these events and split them by species before analysis, resulting in a total of 1387 records.
To describe the domestic prey spectrum and identify correlates among the reported livestock predation events, we restricted the data to cases where clear evidence (C1) or confirmed hints (C2) were available, resulting in a sample size of 578 confirmed wolf predation events.
To test our main hypotheses, we used three separate modelling approaches: (1) a rarefaction model to assess the species richness of reported livestock predation events, and (2) a count regression model to test the impact of farm type and livestock species identity on the number of killed livestock per reported event. Since incorporating the temporal dimension into the model which tested the effect of farm type and livestock category was not feasible due to strong collinearity of the independent variables, we used (3) count regression models to describe temporal trends of species-specific reported events. For all analyses we used the software R 4.1 (R Core Team, 2021).
(1) To assess the species composition among livestock killed by wolves (C1 and C2), we calculated the proportion of reported predation events by wolves for each livestock species (or category). To further analyze the domestic prey spectrum of wolves and assess if the livestock species richness has reached an asymptote, we estimated species accumulation curves in EstimateS (Colwell, 2009), by randomizing the number of killed livestock species over the years. For this analysis we used the identity of the livestock species and not the aggregated species category.
(2) To assess how farm type (5-level factor) and livestock species category (4-level factor) affected reported predation on livestock (C1 and C2), we modelled the number of livestock individuals killed per reported predation event using a count regression model with log-link and negative binomial error distribution to account for the observed overdispersion in the number of killed livestock individuals per reported predation event. We predicted effect sizes of this model using the effects package (Fox and Weisberg, 2019).
(3) To investigate temporal trends of reported predation events caused by wolves, we used count regression models and fitted a separate model for each livestock category. We defined the number of reported and confirmed predation events per month (C1 and C2) as target variable and specified calendar year (linear predictor) and month (12-level factor) as explanatory variables. Because the target variable showed signs of overdispersion for sheep and cattle, we used negative binomial regressions for these livestock categories; models for farmed deer and other species were specified as Poisson regression.
To visually assess the fit of count regression models [(2) and (3)], we evaluated ‘rootograms’ of the models using the countreg package (Kleiber and Zeileis, 2016).
Among the four livestock categories and averaged over the entire observation period (2007-2020), the proportion of verified wolf attacks (C1 and C2 combined) was highest in farmed deer (60%), followed by sheep (53%), other livestock (32%) and cattle (25%) (Figure 2A).
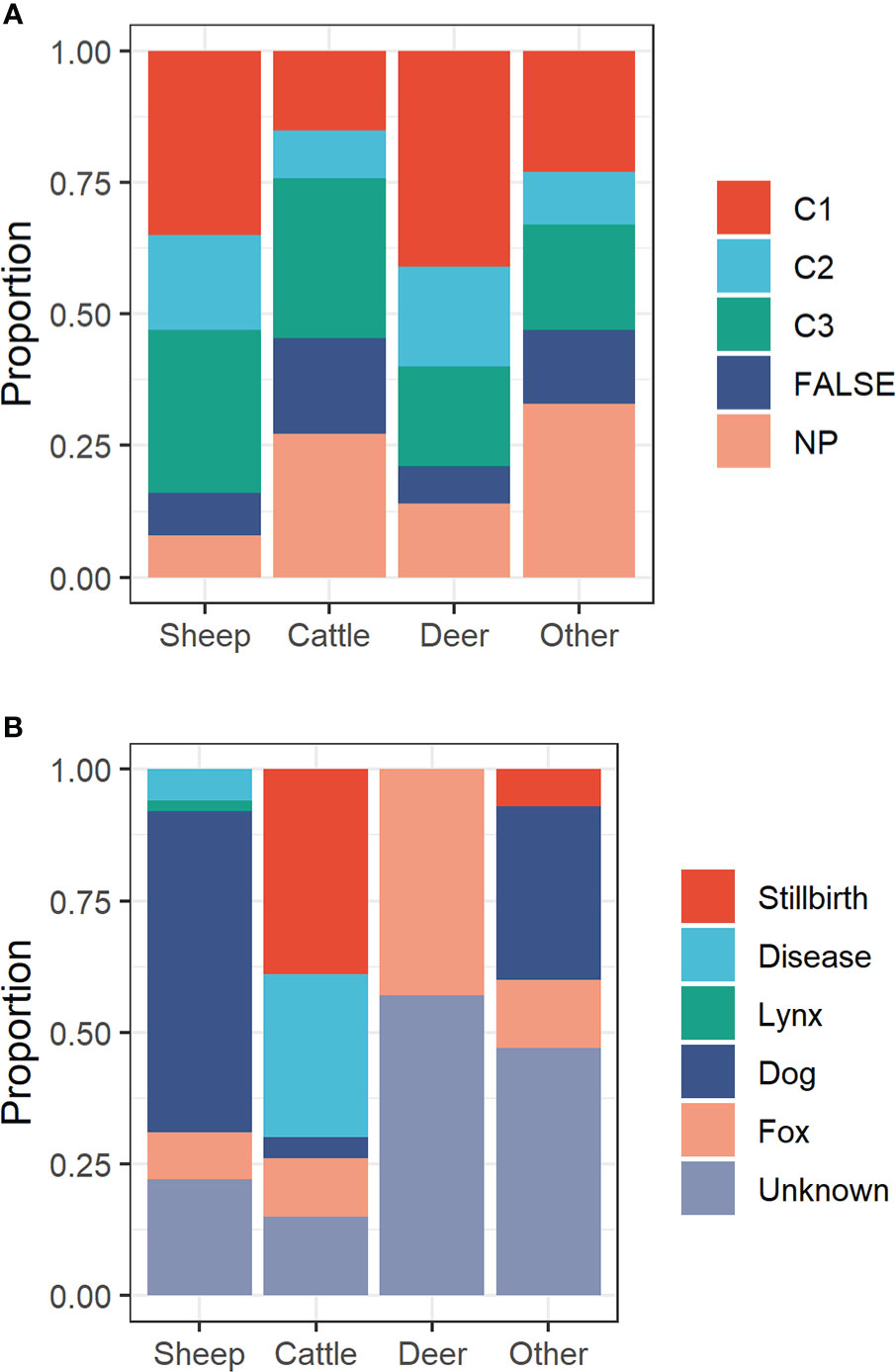
Figure 2 (A) Proportion of reported predation events on livestock by wolves in Brandenburg state (Germany), stratified by monitoring category (C1: clear evidence; C2: confirmed hint; C3: unconfirmed hint; FALSE: livestock loss not caused by wolves; NP: evaluation not possible) and livestock category. (B) Proportion of livestock causes for livestock losses from the FALSE-subset (where wolves were excluded as possible mortality cause).
In several cases, wolves were excluded as cause of mortality (number of “FALSE” reports: sheep=54; cattle=93; farmed deer=7; other livestock=15). For a large portion of these cases (sheep=22%; cattle=15%; farmed deer=57%; other livestock=47% of “FALSE” reports), it was not possible to establish the exact cause of mortality (Figure 2B). In sheep, non-wolf related mortality was frequently caused by dogs (61% of “FALSE” reports). Non-wolf-related cattle mortality was mostly attributed to stillbirth (39%) or disease (31%). In farmed deer, foxes were occasionally (43%) identified as the mortality cause. In one out of three cases (33%), dogs killed other livestock species (Figure 2B).
From 2007 to 2020, the number of reports (C1, C2, C3, NP and FALSE combined) increased substantially in all four livestock categories (Figures 3A–D). In particular, the number of reports increased relatively abruptly from 2015 onwards, especially for cattle (Figure 3B), farmed deer (Figure 3C), and other livestock species (Figure 3D).
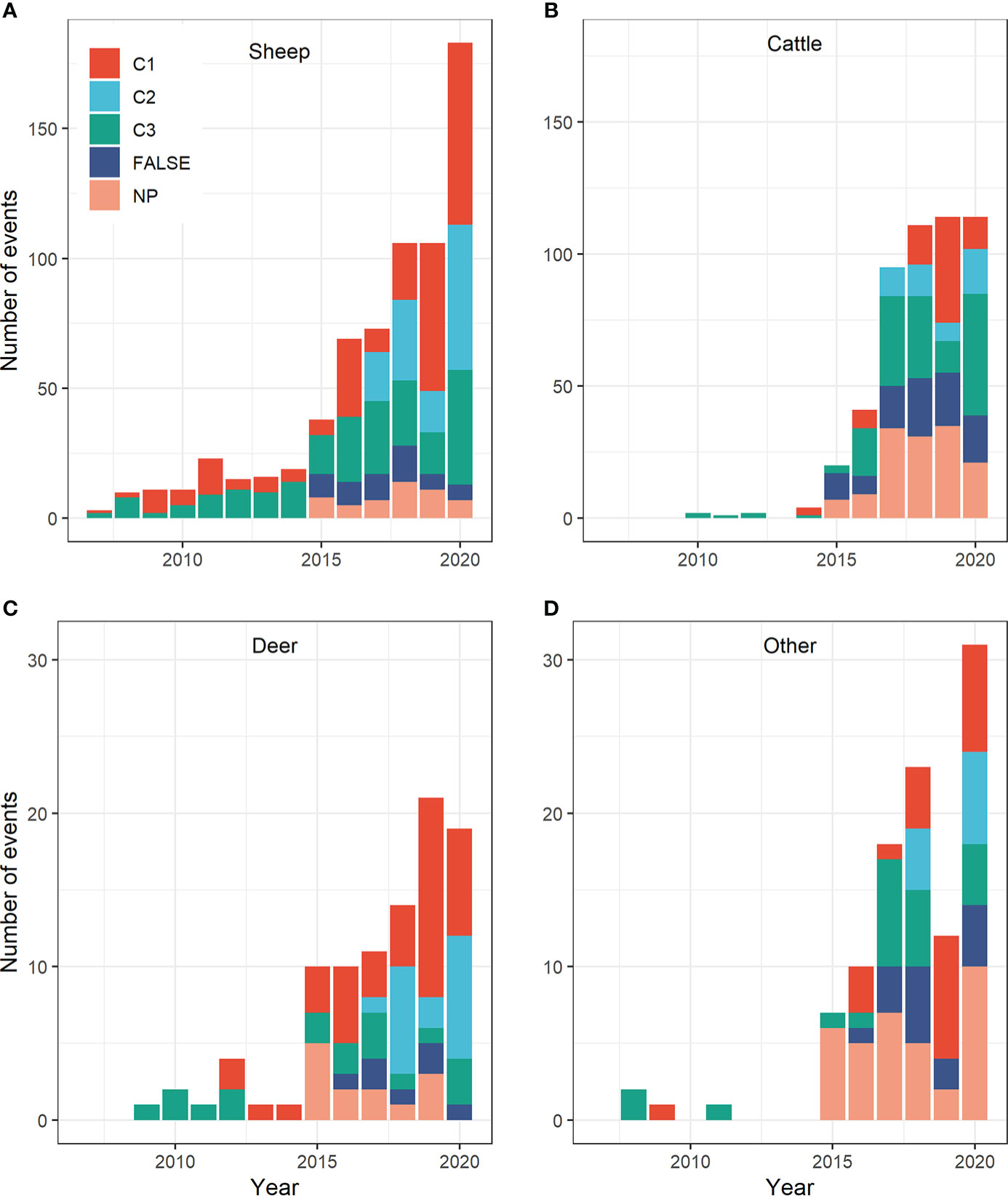
Figure 3 Number of reported wolf predation events in Brandenburg state (Germany), stratified by monitoring category (C1: clear evidence; C2: confirmed hint; C3: unconfirmed hint; FALSE: livestock loss not caused by wolves; NP: evaluation not possible) and calendar year for (A) sheep, (B) cattle, (C) farmed deer, and (D) other livestock species. Note the different y-scales in the upper and lower panels.
In terms of livestock species composition, reported and verified predation events (C1 and C2) occurred primarily in domestic sheep (n=364 events; 63.0% of all events), followed by cattle (n=124; 21.5%), and farmed deer (n=57; 9.9%) (Figure 3A). Other species accounted only for a small share (n=33; 5.7%) and included goat (Capra aegagrus hircus), mouflon, alpaca (Vicugna pacos), horse (Equus caballus), and rhea (Rhea americana). Based on the available reports, wolves preyed on nine different farmed species (farmed deer include fallow deer and red deer). Treating each calendar year as sample replicate, livestock species accumulation curves did not reach a plateau (Figure 4).
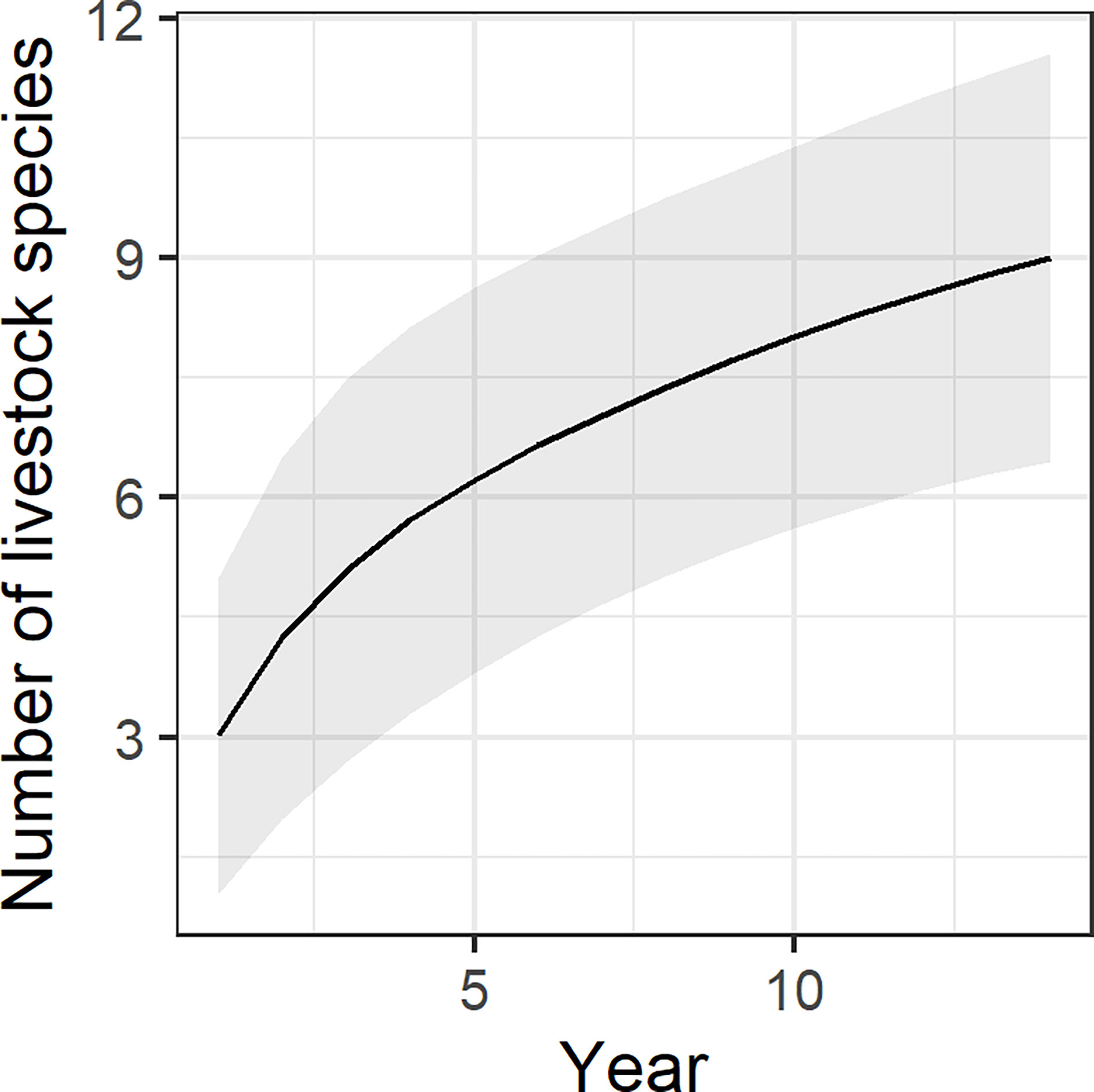
Figure 4 Estimated number of livestock species killed by wolves in Brandenburg State (Germany) based on reported wolf predation events. The species accumulation curve was modelled as a function of 14 complete sampling years (2007 to 2020) and shows the increase in dietary breadth of wolves.
The count regression model suggested that both farm type and livestock category affected the mean number of killed livestock individuals per reported (C1 and C2) predation event (Table 1; Figure S1). Compared to full-time farms (with a mean of four killed livestock per reported event; Figure S1), the mean number of killed livestock per reported predation event was lower in part-time farms, farms of unknown types, hobby farms (in each category a mean of approximately three individuals killed per reported event; Figure S1), and was lowest in other farm types (a mean of one individual killed per reported event; Figure S1). In terms of livestock category, the model revealed stark differences. Wolves were predicted to kill a mean of four sheep per reported predation event (Table 1; Figure S1). However, the range of killed sheep individuals varied considerably (Figure 5A). In line with the observed frequency of cattle killed per reported event (Figure 5B), the model predicted a mean of one killed individual per reported event (Figure S1). For farmed deer, the model predicted a mean of four individuals killed per reported event (Figure S1). However, the number of killed individuals varied considerably (Figure 5C). For other livestock species, the model predicted a mean of two individuals killed per reported event (Figure S1), but the number of killed individuals also varied in this category (Figure 5D). Model fit was decent, except for cases with single livestock individuals killed per event (Figure S2). Reported livestock predation by wolves showed substantial temporal dynamics (Figure 6). In all four livestock groups, the number of reported predation events increased significantly over the years (Figures 3, 6); the species-specific models indicated particularly strong annual increases in cattle and other livestock species, followed by farmed deer and sheep (Table 2). The monthly distribution of reported predation events also differed by species (Figure 6; Table 2). Reported predation on sheep occurred least often in June and peaked in September and October (Table 2). In more recent years (particularly in the year 2020), reported predation on sheep frequently occurred in March (Figure 6A), yet this heaping was not statistically different from the frequency of reported predation events in January (Table 2). Temporal patterns of reported predation on cattle differed from those in sheep (Figures 6A, B) with peaks occurring in the spring months (March, April, and May, and with marginal significance also in June) and from August through October (Table 2). Reported predation on farmed deer (Figure 6C) and on other species (Figure 6D), was not significantly associated with specific months (Table 2). Model fit was good, with little evidence for over- or underfitting (Figure S3).
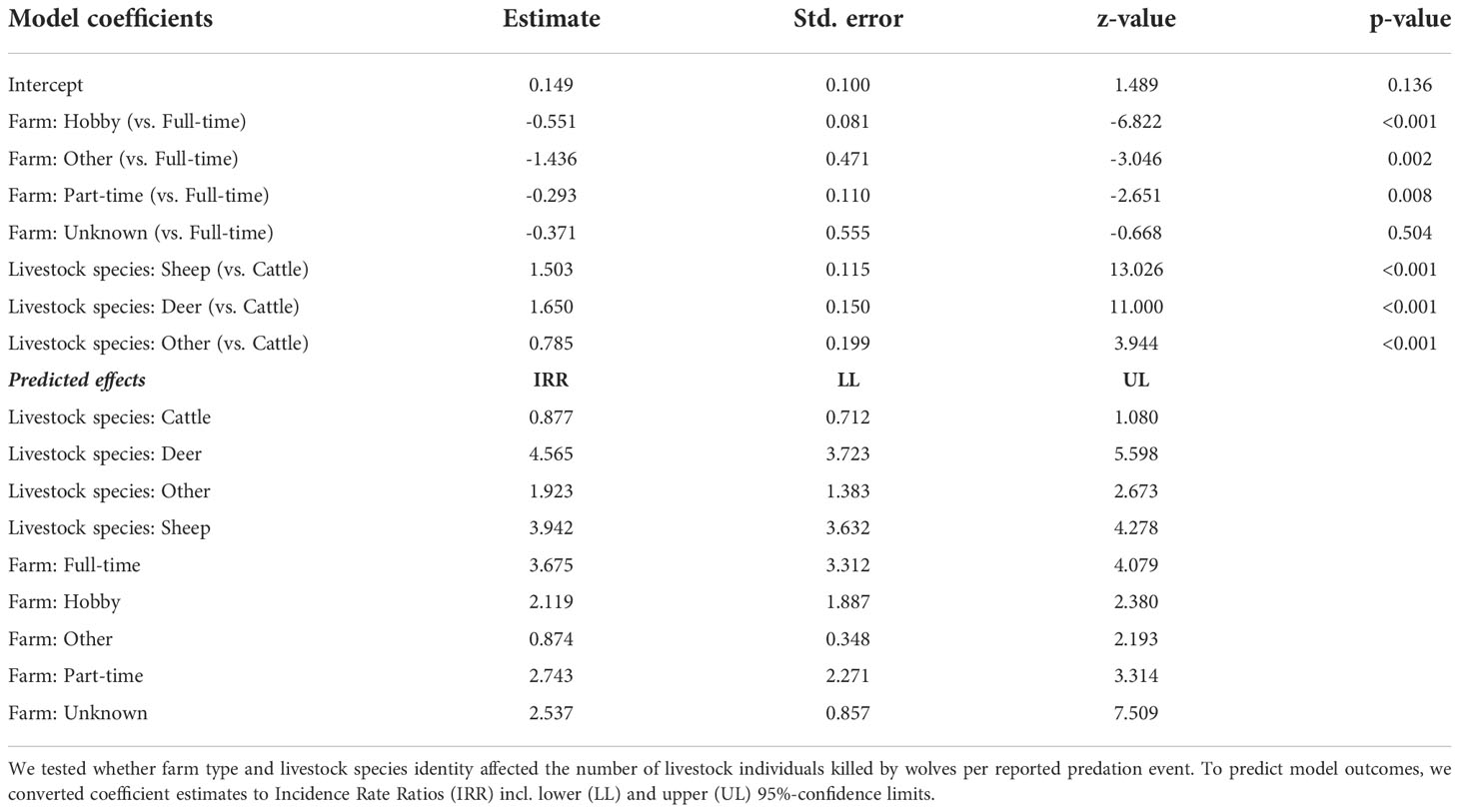
Table 1 Summary statistics of the negative binomial regression model to assess correlates of predation on livestock by wolves in Brandenburg State (Germany).
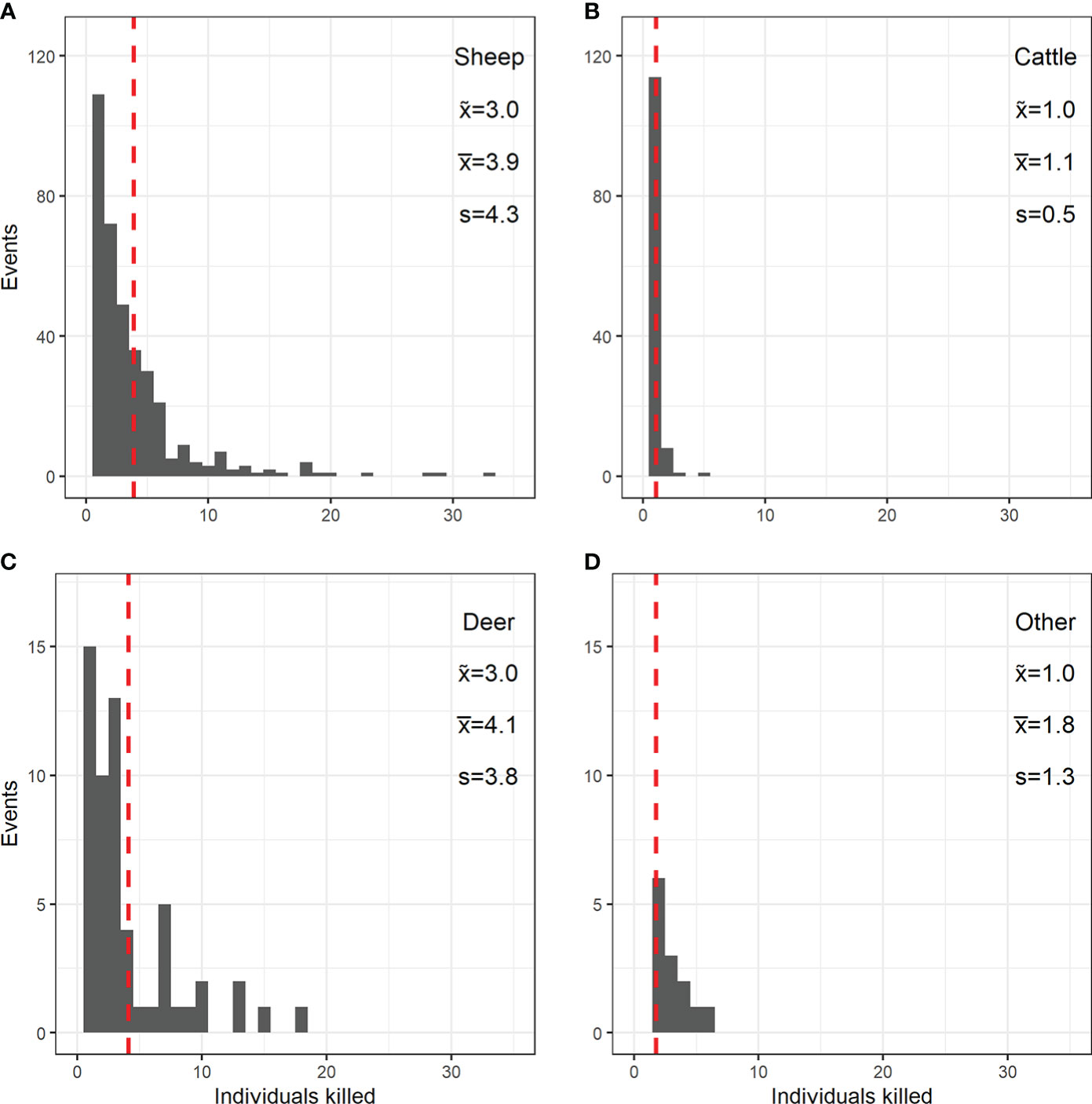
Figure 5 Distribution of number of livestock individuals [(A) sheep; (B) cattle, (C) farmed deer, and (D) other livestock species] killed by wolves in Brandenburg state (Germany) from 2007 to 2020 according to reported wolf predation events. To describe the distribution for each species category, we report the median (), the mean (; this value is plotted as dashed red line), and the standard deviation (s). Note the different y-scales in the upper and lower panels.
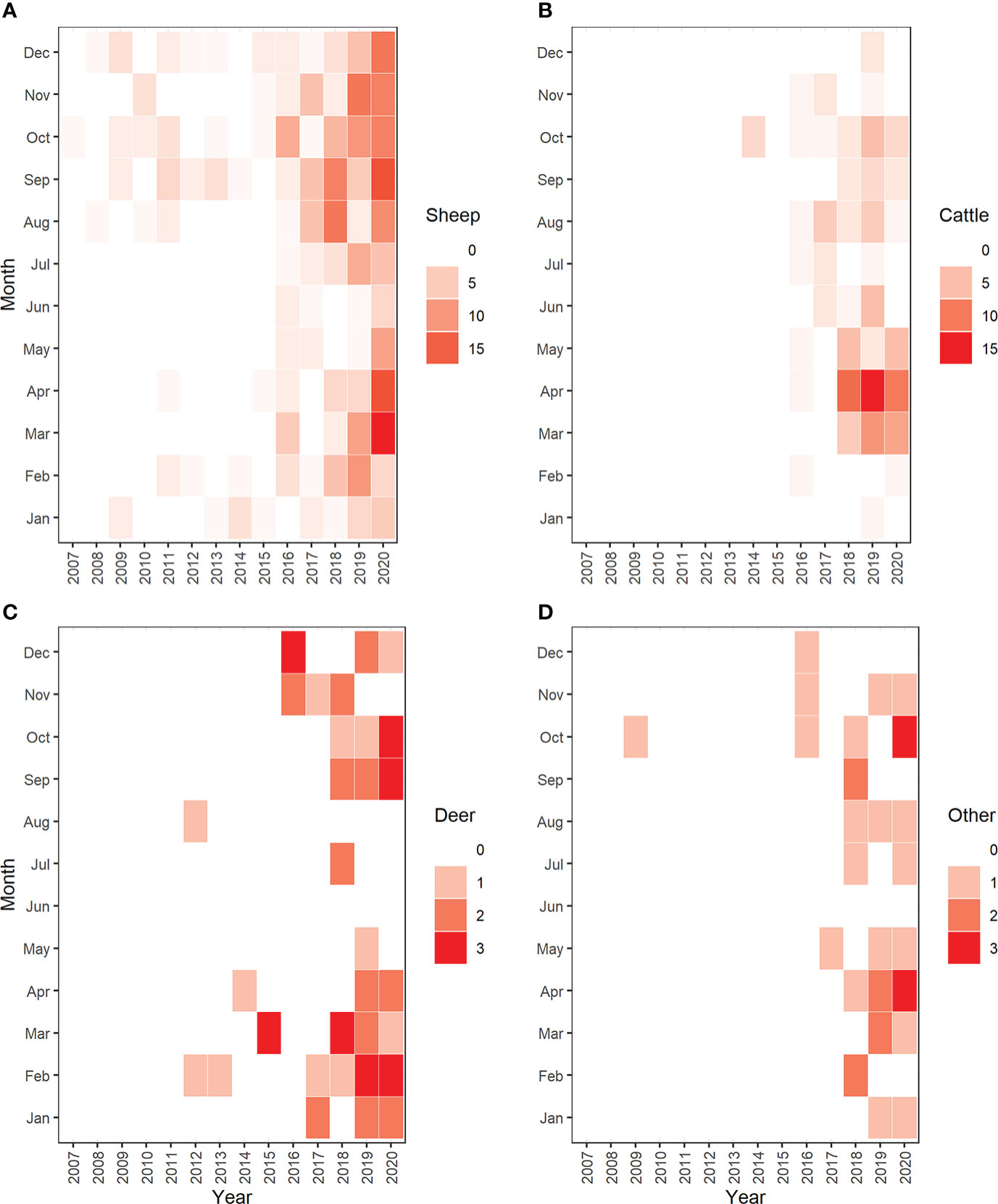
Figure 6 Yearly and monthly distribution of reported wolf predation events on (A) sheep, (B) cattle, (C) farmed deer, and (D) other livestock species in Brandenburg State (Germany).
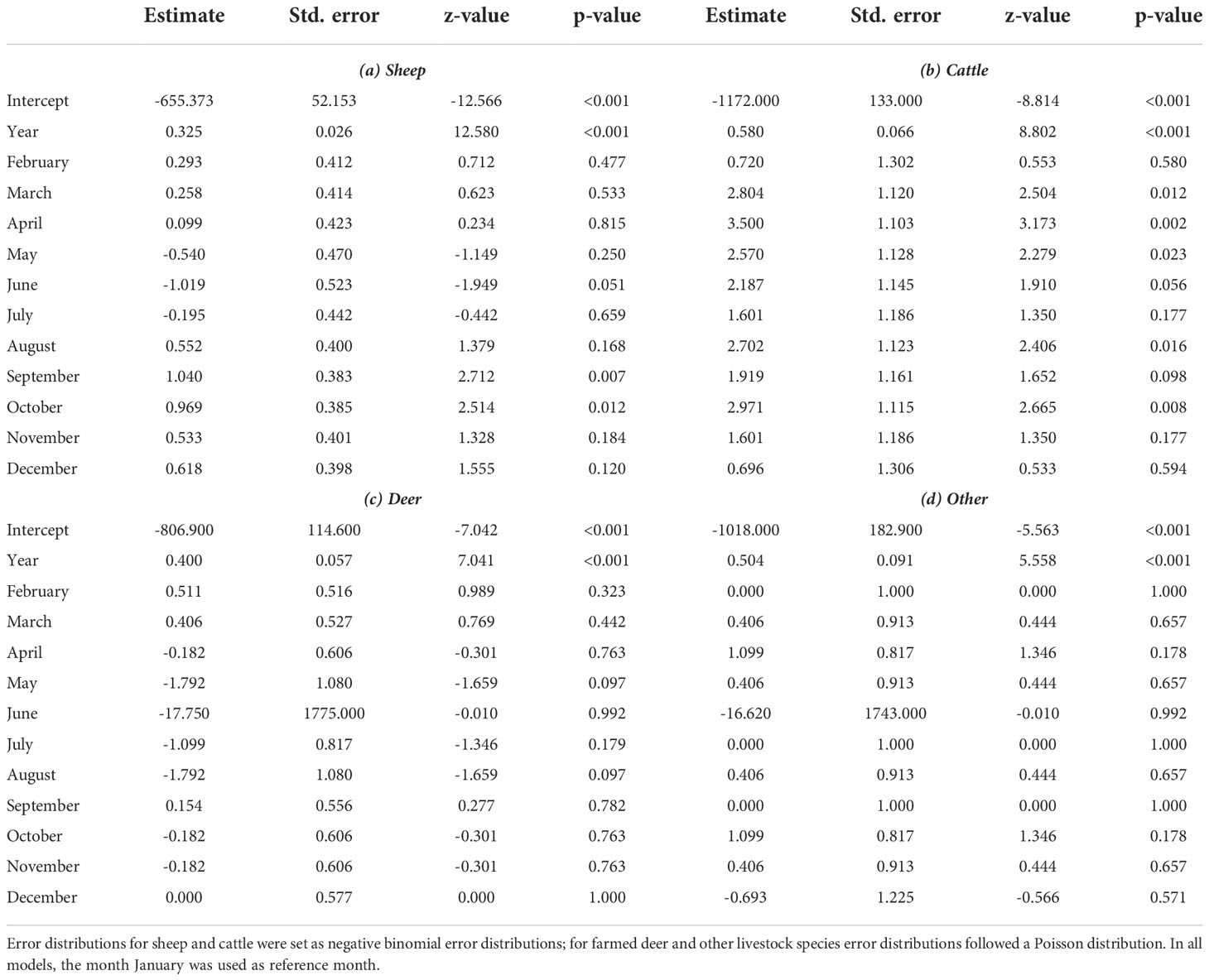
Table 2 Summary statistics of count regression models, testing the effect of year and month on the number of reported predation events on (a) sheep, (b) cattle, (c) farmed deer, and (d) other species by wolves in Brandenburg State (Germany).
Livestock loss due to wolves is arguably the most frequent and contentious issue associated with the recovery of wolf populations in agricultural landscapes (Ripple et al., 2014; van Eeden et al., 2018) and strongly affects negative attitudes among stakeholders and the general public (Dressel et al., 2015), which in turn presents challenges to determining options for sustainable coexistence of wolves and humans in agricultural landscapes (König et al., 2020). In order to provide the evidence basis for mitigating stakeholder conflicts, we assessed patterns and correlates of livestock predation by wolves. The analysis of reported predation on livestock by a recolonizing wolf population in Brandenburg over a period of 14 years revealed the following major results: (1) about 42% of reported livestock losses were clearly confirmed to be caused by wolves, but this varied by species category; (2) livestock species and farm type influenced the number of killed livestock during a reported wolf attack; (3) the spectrum of reported livestock species killed by wolves and the frequency of reported predation events increased alongside an expanding wolf population; and (4) reported predation on livestock was characterized by species-specific seasonal patterns.
Compensation schemes for damages related to predation by carnivores, although heavily debated (Nyhus et al., 2003; Bulte and Rondeau, 2005), are a central instrument for facilitating the persistence of large carnivores in landscapes shared among people, livestock and wildlife (Dickman et al., 2011). In central Europe, compensation payments typically depend on an independent assessment by trained assessors to confirm wolf predation as mortality cause (Iliopoulos et al., 2009; Gervasi et al., 2021b). Despite substantial forensic efforts, the exact mortality cause could not always be established (NP=17%), especially among cattle and other livestock species. Overall, our case study suggests relatively high confirmation rates (C1 + C2 = 42%; C3 = 29%), and a relatively low frequency of events in which wolves were mistakenly assumed as mortality cause (False=12%). Cattle loss was occasionally (18%) claimed as wolf depredation but was actually caused by other processes such as stillbirths, diseases or other causes (Figure 2A). Further studies could address whether these false claims are due to actual difficulties in establishing mortality causes or whether they are, at least partially, driven by intentions to offset costs associated with loss of valuable livestock such as cattle. While the overall degree of over-reporting livestock loss is relatively small (i.e. 12%), particularly when compared massive over-reporting (several orders of magnitude) in other livestock-carnivore systems (e.g. Kissui et al., 2022), these results underscore the importance of independent confirmation procedures to avoid fraudulent claims of livestock loss (Nyhus et al., 2003).
On the other hand, cases in which wolves were identified as mortality cause could theoretically have been cases of wolves scavenging on livestock that died due to other reasons (Ciucci et al., 2020). However, we assume that such instances were rare because farmers typically monitor their livestock on a daily basis and immediately remove livestock carcasses, thus minimizing the time period for potential scavenging.
Considering that establishing the exact cause of livestock mortality is occasionally difficult and to increase farmers’ tolerance for wolves, compensation payments are only withheld if the assessment clearly excludes wolves as possible mortality cause. While the overall patterns of predation on livestock by wolves are presumably well reflected in this database, the reported numbers likely represent minimum numbers. We suspect that a certain amount of livestock predation is not reported: in several studies farmers have been found to refrain from reporting due to perceived high administrative burden (Ravenelle and Nyhus, 2017). Farmers may also have not reported predation events if their livestock were not registered by the authorities, or if they were aware that they did not have minimum livestock protection standards in place and were thus not eligible for compensation.
In line with patterns of predation on livestock by wolves in Europe (e.g. Ciucci and Boitani, 1998; Gula, 2008; Iliopoulos et al., 2009; Gervasi et al., 2021a; Gervasi et al., 2021b), sheep were the most frequently reported livestock category preyed upon by wolves in Brandenburg (Figure 3A). We assume that this is due to their relatively high availability in the agricultural landscape (Khorozyan and Heurich, 2022). Nevertheless, reports indicate that wolves preyed on a wide range of livestock species (Figure 4). In particular from 2015 onwards, reported predation on cattle occurred relatively frequently (Figures 3B, 6B). Although the onset of increases in cattle reports coincided with the change in the compensation scheme (Figure 3B), we assume that this did not markedly affect reporting among cattle farmers. Even prior to 2015, cattle loss due to wolves was eligible for compensation. Moreover, the key prerequisite for successfully claiming compensation –i.e. having minimum standards for livestock protection in place – was only applicable after the change of the compensation scheme. Thus, systematic under-reporting is unlikely to explain the observed temporal pattern. Overall, the wide and increasing range of reported livestock species killed by wolves (incl. horses, cattle and exotic species such as alpaca or rhea) resonates well with the wide food niche of wolves (Okarma, 1995). Clearly, predation on such a wide variety of species including pets or companion animals may not only represent substantial direct and indirect economic damages (which may not always be entirely covered by compensation schemes) but may also inflict grief and even trauma to owners of livestock or pets (Rujoiu and Rujoiu, 2014; Spain et al., 2019).
To date, the factors driving people tolerance of wildlife are poorly understood (Treves and Bruskotter, 2014), yet people who had experienced damages by wolves generally have more negative attitudes towards wolves (Dressel et al., 2015). Therefore, reducing predation by protecting livestock is considered to be one (but not the only) central aspect to fostering tolerance for wolves. Hence, the wide range of the domestic prey spectrum warrants the wider usage of preventive husbandry practices, especially the use of adequately installed and sufficiently powered mobile electrified fences (Reinhardt et al., 2012), not only for paddocks containing small stock such as sheep and goats but also for paddocks containing other domestic species that are potentially subject to wolf depredation. To offset not only the economic cost associated with the loss of livestock, the implementation of preventive methods is financially supported by the state. Payments for livestock damage prevention measures, including monetary compensation for improved fences and purchasing and maintenance of livestock guardian dogs have been provided throughout the project period. These preventive measures are considered a key tool to promote coexistence between people, their livestock, and wolves in Germany. Financial volume for supporting livestock protection measures by far exceeds compensatory payments (https://www.dbb-wolf.de/wolf-management/herd-protection/prevention-and-compensation-payments).
In terms of surplus or mass killing by wolves, our results confirm previous research (e.g. Ciucci and Boitani, 1998; Gazzola et al., 2008) highlighting that this phenomenon occurs mainly in sheep. However, we show that mass killing also occurs in farmed deer. Rather than a general or temporary loss of natural anti-predatory instincts (which could be expected in domestic sheep but not necessarily in farmed deer, due to their much more recent and incomplete domestication) (Kruuk, 1972), we suggest that it is the “unusual condition” of livestock pastures (i.e. relative high number of accessible, yet confined, individual animals) that facilitates surplus killing of sheep and farmed deer by wolves (Linnell et al., 2008). In contrast, the relatively large prey handling time, and the threat imposed by potentially aggressive adult cattle in fenced pastures (Laporte et al., 2010), likely limit surplus killing in cattle herds.
The recorded farm type also affected reported predation on livestock with full-time farms typically experiencing a greater number of losses during a single wolf attack (Table 1). Unfortunately, the monitoring protocol included no key variables that affect predation on livestock, such as presence, type and specific conditions of implemented livestock protection methods (Eklund et al., 2017), husbandry practices (Ciucci and Boitani, 1998) and the flock size of livestock (Vos, 2000; Iliopoulos et al., 2009). Therefore, we can only assume that these variables differ by the coarse farm type category (e.g. greater flock sizes in full-time farms). To provide a clearer evidence basis for possible effects of livestock husbandry practices, we strongly suggest that these variables are systematically and explicitly recorded in future monitoring protocols.
Similar to other case studies (Harper et al., 2005; Fedyń et al., 2022; Khorozyan and Heurich, 2022), our study suggests that recolonizing and expanding wolf populations are associated with a greater frequency of reported predation events (Figures 1, 3 and 6). While such relationships between wolf recolonization and predation on livestock could be expected, we caution against generalizing this correlation. For example, Italian regions recently recolonized by wolves had lower rates of predation on livestock compared to areas with long-established wolf populations (Gervasi et al., 2021b). More detailed studies aimed at identifying the social status of wolves (i.e. resident vs. dispersing) that prey upon livestock may provide more insights in this context. For example, genetic evidence from wolf attacks on livestock in northern Germany and Denmark suggests that predation on livestock is disproportionally caused by dispersing wolves (Mayer et al., 2022) – a wolf category that is unfortunately not directly covered by the current wolf monitoring in Brandenburg.
Our results provide circumstantial evidence that livestock in our study area are not effectively protected against predation by wolves. Whereas husbandry practices are generally imperfectly effective in reducing livestock predation by large carnivores (Eklund et al., 2017; van Eeden et al., 2018; Khorozyan and Waltert, 2019a), we believe that there is substantial potential for farmers to adapt to the presence of wolves (Carter and Linnell, 2016), thereby increasing the efficacy of livestock protection methods. For example, seasonal adjustment of husbandry practices could reduce livestock losses. While generally occurring in any month, predation on sheep peaked in the fall (Figure 6A). This period coincides with the post-weaning period of wolf offspring, a time when nutritional demands by wolf packs are high (Van Ballenberghe and Mech, 1975). In line with similar seasonal patterns of wolf predation on sheep and goat reported from Greece (Iliopoulos et al., 2009), increased nutritional demands by wolf packs present a plausible explanation for the observed seasonal patterns. In contrast, predation on cattle mostly peaked in the spring, especially from March to May (Figure 6B). As this period coincides with the calving period and killed cattle are predominantly calves (among 251 cattle killed by wolves in Germany in 2021, 75% were younger than two months; Dokumentations- und Beratungsstelle des Bundes zum Thema Wolf, 2022), it is very likely that the presence of calves makes cattle more susceptible to wolf predation, a hypothesis that has been previously suggested to explain seasonality of cattle predation in North America (Musiani et al., 2005). While more detailed studies on the interrelationships between wolf behavior, livestock availability and husbandry practices may be required to elucidate the underlying reasons for the observed temporal patterns in predation events on livestock [which are broadly similar but not identical to those observed in North America (Musiani et al., 2005), Greece (Iliopoulos et al., 2009) and Italy (Meriggi et al., 1991)], the interaction between seasonality and livestock category observed in this study presents an important clue towards reducing predation on livestock. Overall, our analyses suggest that both wolf-related as well as livestock related variables affect the seasonality of livestock predation.
Coexistence of wolves and humans in cultural landscapes is a dynamic process, requiring adaption by both wolves and humans (Carter and Linnell, 2016). While wolves have adapted to anthropogenic landscapes, where abundant populations of wild ungulates, such as roe (Capreolus capreolus) and red deer, and wild boar (Sus scrofa) (Wagner et al., 2012) allow for their sustained population growth, the occasional predation on a wide spectrum of livestock species can further diminish the tolerance for wolves. Indeed, cryptic and illegal killing of wolves (often in retaliation for predation on livestock) is a suspected factor influencing wolf populations in Germany (Sunde et al., 2021). Furthermore, predation on pet species and killing of large numbers of livestock (e.g. up to 33 sheep in a single reported event; Figure 5A) can contribute to negative emotions towards wolves not only among livestock owners but also the wider public (Arbieu et al., 2019; Randler et al., 2020; Rode et al., 2021), particularly if sensationalized by the media (Chandelier et al., 2018; Delclaux and Fleury, 2020). As these negative attitudes may limit societal support for wolf conservation, minimizing the frequency and extent of livestock predation is crucial.
Although the speed of wolf population growth is expected to slow down in the near future, more and more areas of Germany are predicted to be occupied by wolves (Fechter and Storch, 2014; Kramer-Schadt et al., 2020). Concomitantly, the risk of predation on livestock will expand to previously unaffected areas if protective methods have not been put in place. Although specific comparisons with other case studies (e.g. in terms of temporal patterns and susceptibility of livestock species) reflect the uniqueness of specific human-wildlife conflict case studies (Zimmermann et al., 2021), our results hint towards ways for improving livestock prevention methods in Brandenburg and possibly other parts of central Europe as well. Our study suggests that employing effective livestock protection methods should be considered for all susceptible livestock and pet species. During the recolonization process, identification, training, and sensitization of key stakeholders, including livestock keepers, farmers, hunters, foresters, state administrative staff, responsible policy makers, and also the general public, are key factors for minimizing possible conflicts (König et al., 2021). In addition, quantifying “tolerable levels” of livestock loss could provide important insights for informing coexistence strategies (Carter and Linnell, 2016).
In Germany, the current strategy for facilitating wolf-livestock coexistence is to minimize livestock loss. To achieve this goal, each German state enacts minimum standards for livestock damage protection methods. These standards vary across states and livestock species and meeting those standards is prerequisite for successfully claiming compensation payments. Minimum standards generally include sufficiently tall and electrified fences; the addition of livestock guardian dogs has proven effective and is particularly recommended (but not required) for full-time farmers who keep large herds of sheep (Reinhardt et al., 2012). Moreover, in line with previous recommendations for reducing wolf predation on livestock in other parts of Europe (Pimenta et al., 2017), we suggest that improving prevention methods during the spring period for cattle (i.e. the main calving season) and during the fall period for sheep (i.e. post-weaning in wolf packs) could substantially reduce the frequency of predation events by wolves. In combination with spatial risk maps (Treves and Rabenhorst, 2017), such seasonally adapted methods could greatly reduce predation on livestock. For example, moving cattle calving grounds away from activity centers of large carnivores [in the cited example: cheetah (Acinonyx jubatus)] drastically reduced livestock losses on Namibian farmlands (Melzheimer et al., 2020). Improving livestock protection could also include keeping susceptible livestock (such as cattle during the calving period) in enforced enclosures at nighttime. While diel patterns of attacks by wolves are difficult to establish, circumstantial evidence from our study site (i.e. farmers typically detect dead livestock in the morning) and evidence from other studies suggests that livestock predation by wolves occurs mostly in the evening, at night, or early morning (Ciucci and Boitani, 1998; Iliopoulos et al., 2009). Enforced nighttime enclosures are cost-effective in other parts of the world (e.g. Kissui et al., 2019), and despite causing additional effort and expenses, they may be a suitable tool for reducing predation on livestock in central Europe, particularly during seasons with higher risk of predation.
Our analyses summarize the magnitude of reported livestock loss due to predation by wolves and call for renewed investment in research and implementation of preventative methods. Additionally, we recommend that potential benefits of wolves, such as reduction of animal-vehicle collisions (Raynor et al., 2021), dampening of pathogen transmission (Tanner et al., 2019), and fostering vegetation growth through a release from excessive herbivory (Callan et al., 2013), are assessed by dedicated monitoring efforts and equally highlighted to provide a more nuanced and balanced perspective in the contentious debate about wolves in human-dominated landscapes.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical review and approval was not required for the animal study because monitoring was not invasive. Written informed consent for participation was not obtained from the owners because data resulted from state-wide, mandatory monitoring.
CK, HK, and VH conceived and planned the experiments. VH coordinated the data collection. CK carried out the analyses. CK, SU, E-FO-M, VH, and HK contributed to the interpretation of the results. CK took the lead in writing the manuscript. All authors provided critical feedback and helped shape the research, analysis and manuscript.
This article was made possible through funding by the Marianne and Dr. Fritz Walter Fischer-Stiftung for the junior research group LandSTRAT. We sincerely thank S. Kramer-Schadt and two reviewers for helpful comments on earlier drafts.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2022.989368/full#supplementary-material
Supplementary Figure 1 | Predicted effects of livestock species category and farm type on the number of killed livestock individuals per reported wolf predation event. Predictions are based on the negative binomial regression model summarized in Table 1.
Supplementary Figure 2 | Rootogram of the fitted negative binomial regression model to assess the influence of farm type and livestock species category on the number of killed livestock individuals per reported event. Bars indicate observed and red dots indicate expected (given by the model) values. If the bar of the observed values is above the zero line the model overpredicts this particular count bin; if the bar exceeds the zero line it underpredicts the number of predation events.
Supplementary Figure 3 | Rootograms of the fitted count regression models to predict temporal trends of reported livestock predation events caused by wolves in Brandenburg State (Germany). Bars indicate observed and red dots indicate expected (given by the model) values. If the bar of the observed values is above the zero line the model overpredicts this particular count bin; if the bar exceeds the zero line it underpredicts the number of predation events. Models were fitted with negative binomial error distribution for sheep and cattle, and with Poisson error distribution for farmed deer and other species.
AFS (2021) Flächennutzung. Available at: https://www.statistik-berlin-brandenburg.de/flaechennutzung.
Arbieu U., Mehring M., Bunnefeld N., Kaczensky P., Reinhardt I., Ansorge H., et al. (2019). Attitudes towards returning wolves (Canis lupus) in Germany: Exposure, information sources and trust matter. Biol. Conserv. 234, 202–210. doi: 10.1016/j.biocon.2019.03.027
Barua M., Bhagwat S. A., Jadhav S. (2013). The hidden dimensions of human-wildlife conflict: Health impacts, opportunity and transaction costs. Biol. Conserv. 157, 309–316. doi: 10.1016/j.biocon.2012.07.014
Bautista C., Revilla E., Berezowska-Cnota T., Fernández N., Naves J., Selva N. (2021). Spatial ecology of conflicts: Unravelling patterns of wildlife damage at multiple scales. Proc. R. Soc B Biol. Sci. 288. doi: 10.1098/rspb.2021.1394
Bulte E. H., Rondeau D. (2005). Why compensating wildlife damages may be bad for conservation. J. Wildl. Manage. 69, 14–19. doi: 10.2193/0022-541X(2005)069<0014:WCWDMB>2.0.CO;2
Callan R., Nibbelink N. P., Rooney T. P., Wiedenhoeft J. E., Wydeven A. P. (2013). Recolonizing wolves trigger a trophic cascade in Wisconsin (USA). J. Ecol. 101, 837–845. doi: 10.1111/1365-2745.12095
Carter N. H., Linnell J. D. C. (2016). Co-Adaptation is key to coexisting with large carnivores. Trends Ecol. Evol. 31, 575–578. doi: 10.1016/j.tree.2016.05.006
Chandelier M., Steuckardt A., Mathevet R., Diwersy S., Gimenez O.. (2018). Content analysis of newspaper coverage of wolf recolonization in France using structural topic modeling. Biol. Conserv. 220, 254–261. doi: 10.1016/j.biocon.2018.01.029
Chapron G., Kaczensky P., Linnell J. D. C., Von Arx M., Huber D., Andrén H., et al. (2014). Recovery of large carnivores in Europe’s modern human-dominated landscapes. Science (80-.). 346, 1517–1519. doi: 10.1126/science.1257553
Cimatti M., Ranc N., Benítez-López A., Maiorano L., Boitani L., Cagnacci F., et al. (2021). Large Carnivore expansion in Europe is associated with human population density and land cover changes. Divers. Distrib. 27, 602–617. doi: 10.1111/ddi.13219
Ciucci P., Boitani L. (1998). Wolf and dog depredation on livestock in central Italy. Wildl. Soc Bull. 26, 504–514. http://www.jstor.org/stable/3783763.
Ciucci P., Mancinelli S., Boitani L., Gallo O., Grottoli L. (2020). Anthropogenic food subsidies hinder the ecological role of wolves: Insights for conservation of apex predators in human-modified landscapes. Glob. Ecol. Conserv. 21, e00841. doi: 10.1016/j.gecco.2019.e00841
Colwell R. K. (2009). EstimateS 8.2. 0. Statistical estimation of species richness and shared species from samples. User’s Guide and Application. Department of Ecology and Evolutionary Biology, University of Connecticut, Storrs.
Delclaux J., Fleury P. (2020). Medium‐term evolution in French national newspaper coverage of the interrelations between biodiversity and agriculture. Conserv. Sci. Pract. 3, e140. doi: 10.1111/csp2.140
Delgiudice G. D. (1998). Surplus killing of white-tailed deer by wolves in northcentral Minnesota. J. Mammal. 79, 227–235. doi: 10.2307/1382858
Dickman A. J., Macdonald E. A., Macdonald D. W. (2011). A review of financial instruments to pay for predator conservation and encourage human-carnivore coexistence. Proc. Natl. Acad. Sci. 108, 13937–13944. doi: 10.1073/pnas.1012972108
Dokumentations- und Beratungsstelle des Bundes zum Thema Wolf (2022). Wolfsverursachte Schäden, Präventions- und Ausgleichszahlungen in Deutschland. DBBW - Dokumentations- und Beratungsstelle des Bundes zum Thema Wolf. Available at: https://www.dbb-wolf.de/mehr/literatur-download/berichte-zu-praevention-und-nutztierschaeden. (last access: November 22, 2022).
Dondina O., Meriggi A., Dagradi V., Perversi M., Milanesi P. (2015). Wolf predation on livestock in an area of northern Italy and prediction of damage risk. Ethol. Ecol. Evol. 27, 200–219. doi: 10.1080/03949370.2014.916352
Dressel S., Sandström C., Ericsson G. (2015). A meta-analysis of studies on attitudes toward bears and wolves across Europe 1976-2012. Conserv. Biol. 29, 565–574. doi: 10.1111/cobi.12420
Eklund A., López-Bao J. V., Tourani M., Chapron G., Frank J. (2017). Limited evidence on the effectiveness of interventions to reduce livestock predation by large carnivores. Sci. Rep. 7, 2097. doi: 10.1038/s41598-017-02323-w
Fechter D., Storch I. (2014). How many wolves (Canis lupus) fit into Germany? The role of assumptions in predictive rule-based habitat models for habitat generalists. PloS One 9 (7), e101798. doi: 10.1371/journal.pone.0101798
Fedyń I., Bojarska K., Gerber N., Okarma H. (2022). Blood trail of expansion? Long-term patterns of livestock depredation by wolves in Poland. Ecol. Res. 37, 370–380. doi: 10.1111/1440-1703.12301
Fox J., Weisberg S. (2019). An R companion to applied regression. 3rd edition (Thousand Oaks: Sage).
Gazzola A., Capitani C., Mattioli L., Apollonio M. (2008). Livestock damage and wolf presence. J. Zool. 274, 261–269. doi: 10.1111/j.1469-7998.2007.00381.x
Gervasi V., Linnell J. D. C., Berce T., Boitani L., Cerne R., Ciucci P., et al. (2021a). Ecological correlates of large carnivore depredation on sheep in Europe. Glob. Ecol. Conserv. 30, e01798. doi: 10.1016/j.gecco.2021.e01798
Gervasi V., Salvatori V., Catullo G., Ciucci P. (2021b). Assessing trends in wolf impact on livestock through verified claims in historical vs. recent areas of occurrence in Italy. Eur. J. Wildl. Res. 67, 82. doi: 10.1007/s10344-021-01522-1
Gula R. (2008). Wolf depredation on domestic animals in the polish Carpathian mountains. J. Wildl. Manage. 72, 283–289. doi: 10.2193/2006-368
Hanley Z. L., Cooley H. S., Maletzke B. T., Wielgus R. B. (2018). Forecasting cattle depredation risk by recolonizing gray wolves. Wildlife Biol. 2012, 018. doi: 10.2981/wlb.00419
Harms V., Nowak C., Carl S., Muñoz-Fuentes V. (2015). Experimental evaluation of genetic predator identification from saliva traces on wildlife kills. J. Mammal. 96, 138–143. doi: 10.1093/jmammal/gyu014
Harper E. K., Paul W. J., Mech L. D. (2005). Causes of wolf depredation increase in Minnesota from 1979-1998. Wildife Soc Bull. 33, 888–896. doi: 10.2193/0091-7648(2005)33[888:COWDIM]2.0.CO;2
Harper E., Paul W. J., Mech D. L., Weisberg S. (2008). Effectiveness of lethal, directed wolf-depredation control in Minnesota. J. Wildl. Manage. 72, 778–784. doi: 10.2193/2007-273
Iliopoulos Y., Sgardelis S., Koutis V., Savaris D. (2009). Wolf depredation on livestock in central Greece. Acta Theriol. (Warsz). 54, 11–22. doi: 10.1007/bf03193133
Jedrzejewski W., Niedziałkowska M., Hayward M. W., Goszczyński J., Jedrzejewska B., Borowik T., et al. (2012). Prey choice and diet of wolves related to ungulate communities and wolf subpopulations in Poland. J. Mammal. 93, 1480–1492. doi: 10.1644/10-MAMM-A-132.1
Janeiro-Otero A., Newsome T. M., Van Eeden L. M., Ripple W. J., Dormann C. F. (2020). Grey wolf (Canis lupus) predation on livestock in relation to prey availability. Biol. Conserv. 243, 108433. doi: 10.1016/j.biocon.2020.108433
Jürgens U. M. (2022). “I am wolf, I rule!” - attributing intentions to animals in human-wildlife interactions. Front. Conserv. Sci. 3, 803074. doi: 10.3389/fcosc.2022.803074
Jürgens U. M., Hacket P. M. W. (2017). The big bad wolf: the formation of a stereotype. Ecopsychology 9, 33–43. doi: 10.1089/eco.2016.0037
Kaczensky P. (1999). Large carnivore depredation on livestock in Europe. Ursus 11, 59–71. Available at: http://www.jstor.org/stable/3872986.
Khorozyan I. (2022). Defining practical and robust study designs for interventions targetet at terrestrial mammalian predators. Conserv. Biol. 36, e13805. doi: 10.1111/cobi.13805
Khorozyan I., Ghoddousi A., Soofi M., Waltert M. (2015). Big cats kill more livestock when wild prey reaches a minimum threshold. Biol. Conserv. 192, 268–275. doi: 10.1016/j.biocon.2015.09.031
Khorozyan I., Heurich M. (2022). Large-scale sheep losses to wolves (Canis lupus) in Germany are related to the expansion of the wolf population but not to increasing wolf numbers. Front. Ecol. Evol. 10, 778917. doi: 10.3389/fevo.2022.778917
Khorozyan I., Waltert M. (2019a). A framework of most effective practices in protecting human assets from predators. Hum. Dimens. Wildl. 24, 380–394. doi: 10.1080/10871209.2019.1619883
Khorozyan I., Waltert M. (2019b). How long do anti-predator interventions remain effective? Patterns, thresholds and uncertainty. R. Soc Open Sci. 6. doi: 10.1098/rsos.190826
Kiffner C., Chapron G., König H. J. (2019). Germany’s wolves in the crosshairs. Science 365 (6458), 1089. doi: 10.1126/science.aay8053
Kissui B. M., Kiffner C., König H. J., Montgomery R. A. (2019). Patterns of livestock depredation and cost-effectiveness of fortified livestock enclosures in northern Tanzania. Ecol. Evol. 9, 11420–11433. doi: 10.1002/ece3.5644
Kissui B. M., Kisimir E. L., Lichtenfeld L. L., Naro E. M., Montgomery R. A., Kiffner C. (2022). “Human-carnivore coexistence in the Tarangire ecosystem,” in Tarangire: Human-wildlife coexistence in a fragmented ecosystem (Cham: Springer), 295–317. doi: 10.1007/978-3-030-93604-4_14
Kleiber C., Zeileis A. (2016). Visualizing count data regressions using rootograms. Am. Stat. 70, 296–303. doi: 10.1080/00031305.2016.1173590
König H. J., Ceauşu S., Reed M., Kendall H., Hemminger K., Reinke H., et al. (2021). Integrated framework for stakeholder participation: Methods and tools for identifying and addressing human–wildlife conflicts. Conserv. Sci. Pract. 3, e399. doi: 10.1111/csp2.399
König H. J., Kiffner C., Kramer-Schadt S., Fürst C., Keuling O., Ford A. T. (2020). Human-wildlife coexistence in a changing world. Conserv. Biol. 34, 786–794. doi: 10.1111/cobi.13513
Kramer-Schadt S., Wenzler M., Gras P., Knauer F. (2020). Habitatmodellierung und Abschätzung der potenziellen Anzahl von Wolfsterritorien (Deutschland: Bundesamt für Naturschutz). Available at: https://www.bfn.de/fileadmin/BfN/service/Dokumente/skripten/Skript556.pdf.
Kruuk H. (1972). Surplus killing by carnivores. J. Zool. 166, 233–244. doi: 10.1111/j.1469-7998.1972.tb04087.x
Kuiper T., Loveridge A. J., Macdonald D. W. (2022). Robust mapping of human–wildlife conflict: controlling for livestock distribution in carnivore depredation models. Anim. Conserv 25 (2), 195–207. doi: 10.1111/acv.12730
Laporte I., Muhly T. B., Pitt J. A., Alexander M., Musiani M. (2010). Effects of wolves on elk and cattle behaviors: Implications for livestock production and wolf conservation. PloS One 5, e11954. doi: 10.1371/journal.pone.0011954
Linnell J. D. C., Cretois B., Nilsen E. B., Rolandsen C. M., Solberg E. J., Veiberg V., et al. (2020). The challenges and opportunities of coexisting with wild ungulates in the human-dominated landscapes of Europe’s anthropocene. Biol. Conserv. 244, 108500. doi: 10.1016/j.biocon.2020.108500
Linnell J. D. C., Odden J., Smith M. E., Aanes R., Swenson J. E., Ronn A., et al. (2008). Large Carnivores that kill livestock: do "problem individuals really exist? Wildl. Soc Bull. 27, 698–705.
Marucco F., Boitani L. (2012). Wolf population monitoring and livestock depredation preventive methods in Europe. Hystrix 23 (1), 1–4. doi: 10.4404/hystrix-23.1-6364
Mayer M., Olsen K., Schulz B., Matzen J., Nowak C., Thomsen P. F., et al. (2022). Occurrence and livestock depredation patterns by wolves in highly cultivated landscapes. Front. Ecol. Evol. 10, 783027. doi: 10.3389/fevo.2022.783027
Mech L. D., Harper E. K., Meier T. J., Paul W. J., Mech L. D., Harper E. K., et al. (2000). Assessing factors that may predispose Minnesota farms to wolf depredations on cattle. Wildl. Soc Bull. 28, 630–635.
Melzheimer J., Heinrich S. K., Wasiolka B., Mueller R., Thalwitzer S. (2020). Communication hubs of an asocial cat are the source of a human – carnivore conflict and key to its solution. Proc. Natl. Acad. Sci. 117 (52), 33325–33333. doi: 10.1073/pnas.2002487117
Meriggi A., Rosa P., Brangi A., Matteucci C. (1991). Habitat use and diet of the wolf in northern Italy. Acta Theriol. (Warsz) 36, 141–151. doi: 10.4098/at.arch.91-11
MIL (2010) Entwicklungsplan für den ländlichen Raum Brandenburgs und Berlins 2007–2013. Available at: https://eler.brandenburg.de/sixcms/media.php/9/EPLR_2007-2013.pdf.
Molinari-Jobin A., Breitenmoser U., Breitenmoser-Würsten C., Cerne R., Drouhet-Hoguet N., Fuxjäger C., et al. (2021). SCALP: Monitoring the Eurasion lynx in the Alps and beyond. CATnews 14, 50–52.
Musiani M., Mamo C., Boitani L., Callaghan C., Gates C. C., Mattei L., et al. (2003). Wolf depredation trends and the use of fladry barriers to protect livestock in western north America. Conserv. Biol. 17, 1538–1547. doi: 10.1111/j.1523-1739.2003.00063.x
Musiani M., Muhly T., Gates C. C., Callaghan C., Smith M. E., Tosoni E. (2005). Seasonality and reoccurrence of depredation and wolf control in western north America. Wildl. Soc Bull. 33, 876–887. doi: 10.2193/0091-7648(2005)33[876:saroda]2.0.co;2
Nyhus P. J. (2016). Human–wildlife conflict and coexistence. Annu. Rev. Environ. Resour. 41, 143–171. doi: 10.1146/annurev-environ-110615-085634
Nyhus P., Fischer H., Madden F., Osofsky S. (2003). Taking the bite out of wildlife damage: The challenges of wildlife compensation schemes. Conserv. Pract. 4, 37–43. doi: 10.1111/j.1526-4629.2003.tb00061.x
Okarma H. (1995). The trophic ecology of wolves and their predatory role in ungulate communities of forest ecosystems in Europe. Acta Theriol. (Warsz). 40, 335–386. doi: 10.4098/AT.arch.95-35
Patalano M., Lovari S. (1993). Food habits and trophic niche overlap of the wolf Canis lupus, L.1758 and the red fox Vulpes vulpes (L.1758) in a Mediterranean mountain area. Rev. d’Ecologie Terre la Vie 48, 279–294. Available at: https://hal.archives-ouvertes.fr/hal-03529120.
Peters K. J. (1997). Alternative utilization of small ruminants - the case of Germany. In sheep and goat husbandry in the central and Eastern European countries - a struggle to survive. Ed. Kukovics S. (Rome: FAO). Available at: http://www.fao.org/regional/europe/pub/rts50/272.htm.
Pettersson H. L., Quinn C. H., Holmes G., Sait S. M., López-Bao J. V. (2021). Welcoming wolves? Governing the return of large carnivores in traditional pastoral landscapes. Front. Conserv. Sci. 2, 710218. doi: 10.3389/fcosc.2021.710218
Pimenta V., Barroso I., Boitani L., Beja P. (2017). Wolf predation on cattle in Portugal: Assessing the effects of husbandry systems. Biol. Conserv. 207, 17–26. doi: 10.1016/j.biocon.2017.01.008
Pimenta V., Barroso I., Boitani L., Beja P. (2018). Risks a la carte: Modelling the occurrence and intensity of wolf predation on multiple livestock species. Biol. Conserv. 228, 331–342. doi: 10.1016/j.biocon.2018.11.008
Randler C., Wagner A., Rögele A., Hummel E., Tomažič I. (2020). Attitudes toward and knowledge about wolves in SW German secondary school pupils from within and outside an area occupied by wolves (Canis lupus). Animals 10 (4), 607. doi: 10.3390/ani10040607
Ravenelle J., Nyhus P. J. (2017). Global patterns and trends in human–wildlife conflict compensation. Conserv. Biol. 31 (6), 1247–1256. doi: 10.1111/cobi.12948
Raynor J. L., Grainger C. A., Parker D. P. (2021). Wolves make roadways safer, generating large economic returns to predator conservation. Proc. Natl. Acad. Sci. 118, e2023251118. doi: 10.1073/pnas.2023251118
R Core Team (2021) R: A language and environment for statistical computing. Available at: http://www.r-project.org/.
Reinhardt I., Kluth G., Nowak C., Szentiks C. A., Krone O., Ansorge H., et al. (2019). Military training areas facilitate the re-colonization of wolves in Germany. Conserv. Lett. 12, e12635. doi: 10.1111/conl.12635
Reinhardt I., Rauer G., Kluth G., Kaczensky P., Knauer F., Wotschikowsky U. (2012). Livestock protection methods applicable for Germany - a country newly recolonized by wolves. Hystrix 23, 62–72. doi: 10.4404/hystrix-23.1-4555
Ripple W. J., Estes J. A., Beschta R. L., Wilmers C. C., Ritchie E. G., Hebblewhite M., et al. (2014). Status and ecological effects of the world’s largest carnivores. Science (80-.). 343, 1241484. doi: 10.1126/science.1241484
Rode J., Flinzberger L., Karutz R., Berghöfer A., Schröter-Schlaack C.. (2021). Why so negative? Exploring the socio-economic impacts of large carnivores from a European perspective. Biol. Conserv. 255, 108918. doi: 10.1016/j.biocon.2020.108918
Rujoiu O., Rujoiu V. (2014). Pet loss and human emotion: Romanian students’ reflections on pet loss. J. Loss Trauma 19, 474–483. doi: 10.1080/15325024.2013.806150
Spain B., O’Dwyer L., Moston S. (2019). Pet loss: Understanding disenfranchised grief, memorial use, and posttraumatic growth. Anthrozoos 32, 555–568. doi: 10.1080/08927936.2019.1621545
Sunde P., Collet S., Nowak C., Thomsen P. F., Hansen M. M., Schulz B., et al. (2021). Where have all the young wolves gone? Traffic and cryptic mortality create a wolf population sink in Denmark and northernmost Germany. Conserv. Lett. 14 (5), e12812. doi: 10.1111/conl.12812
Tanner E., White A., Acevedo P., Balseiro A., Marcos J., Gortázar C. (2019). Wolves contribute to disease control in a multi-host system. Sci. Rep. 9, 7940. doi: 10.1038/s41598-019-44148-9
Treves A., Bruskotter J. (2014). Tolerance for predatory wildlife. Science (80-.). 476, 12–14. doi: 10.1126/science.1252690
Treves A., Naughton-Treves L. (2005). Evaluating lethal control in the management of human–wildlife conflict. In Woodroffe R., Thirgood S., Rabinowitz A. Eds. People Wildlife: Conflict Coexistence? (Cambridge: Cambridge University Press), 86–106. doi: 10.1017/CBO9780511614774.007
Treves A., Jurewicz R. R., Naughton-treves L., Rose R. A., Willging R. C., Wydeven A. P., et al. (2002). Wolf depredation on domestic animals in Wisconsin, 1976-2000. Wildl. Soc Bull. 30, 231–241.
Treves A., Krofel M., McManus J. (2016). Predator control should not be a shot in the dark. Front. Ecol. Environ. 14, 380–388. doi: 10.1002/fee.1312
Treves A., Krofel M., Ohrens O., van Eeden L. M. (2019). Predator control needs a standard of unbiased randomized experiments with cross-over design. Front. Ecol. Evol. 7, 462. doi: 10.3389/fevo.2019.00462
Treves A., Martin K. A., Wydeven A. P., Wiedenhoeft J. E. (2011). Forecasting environmental hazards and the application of risk maps to predator attacks on livestock. Bioscience 61, 451–458. doi: 10.1525/bio.2011.61.6.7
Treves A., Naughton-Treves L., Harper E. K., Mladenoff D. J., Rose R. A., Sickley T. A., et al. (2004). Predicting human-carnivore conflict: a spatial model derived from 25 years of data on wolf predation on livestock. Conserv. Biol. 18, 114–125. doi: 10.1111/j.1523-1739.2004.00189.x
Treves A., Rabenhorst M. F. (2017). Risk map for wolf threats to livestock still predictive 5 years after construction. PloS One 12, e0180043. doi: 10.1371/journal.pone.0180043
Van Ballenberghe V., Mech L. D. (1975). Weights, growth, and survival of timber wolf pups in Minnesota. J. Mammal. 56, 44–63. doi: 10.2307/1379605
van Eeden L. M., Crowther M. S., Dickman C. R., Macdonald D. W., Ripple W. J., Ritchie E. G., et al. (2018). Managing conflict between large carnivores and livestock. Conserv. Biol. 32, 26–34. doi: 10.1111/cobi.12959
Vos J. (2000). Food habits and livestock depredation of two Iberian wolf packs (Canis lupus signatus) in the north of Portugal. J. Zool. 251, 457–462. doi: 10.1017/S0952836900008050
Wagner C., Holzapfel M., Kluth G., Reinhardt I., Ansorge H. (2012). Wolf (Canis lupus) feeding habits during the first eight years of its occurrence in Germany. Mamm. Biol. 77, 196–203. doi: 10.1016/j.mambio.2011.12.004
Widman M., Elofsson K. (2018). Costs of livestock depredation by large carnivores in Sweden 2001 to 2013. Ecol. Econ. 143, 188–198. doi: 10.1016/j.ecolecon.2017.07.008
Widman M., Steen M., Elofsson K. (2019). Indirect costs of sheep depredation by large carnivores in Sweden. Wildl. Soc Bull. 43, 53–61. doi: 10.1002/wsb.951
Wielgus R. B., Peebles K. A. (2014). Effects of wolf mortality on livestock depredations. PloS One 9, e113505. doi: 10.1371/journal.pone.0113505
Keywords: Canis lupus, human-wildlife coexistence, human-wildlife conflict, human-wildlife interactions, pastoralism
Citation: Kiffner C, Uthes S, Ostermann-Miyashita E-F, Harms V and König HJ (2022) Patterns of livestock loss associated with a recolonizing wolf population in Germany. Front. Conserv. Sci. 3:989368. doi: 10.3389/fcosc.2022.989368
Received: 08 July 2022; Accepted: 15 November 2022;
Published: 12 December 2022.
Edited by:
Gladman Thondhlana, Rhodes University, South AfricaReviewed by:
Lorraine Boast-Millar, Cheetah Conservation Botswana, BotswanaCopyright © 2022 Kiffner, Uthes, Ostermann-Miyashita, Harms and König. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian Kiffner, Y2hyaXN0aWFuLmtpZmZuZXJAemFsZi5kZQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.