Abstract
Habitat suitability assessment is the basis for wildlife conservation management and habitat restoration. It is a useful tool to understand the quality of wildlife habitat and its potential spatial distribution. In order to reveal the habitat suitability and connectivity of sambar (Rusa unicolor) to promote species and biodiversity conservation, this study collected records of sambar (Rusa unicolor) from over 2,000 camera traps in the forests of Southwest China in the past 5 years to assess the overall situation of their habitat. The results of the species distribution model revealed that the suitable habitat area for sambar in the five major mountain ranges (Minshan, Qionglai, Daxiangling, Xiaoxiangling, and Liangshan) in Southwest China is 18,231 km2, accounting for 17.02% of the total area. The most suitable habitat of sambar is primarily distributed in Qionglai, as well as the intersection areas of Daxiangling, Xiaoxiangling, and Minshan. The temperature annual range, temperature seasonality, elevation, and distance to road were important factors affecting the distribution of suitable habitat for sambar. Analysis of landscape pattern shows that there were 273 habitat patches, with a maximum patch area of 9,983 km2, accounting for 54.8% of the total suitable habitat area. However, the segmentation index and separation index of each habitat patch were 0.99 and 106.58, respectively, indicating a relatively high habitat fragmentation in the study area. The results of habitat connectivity analysis showed that the Qionglai mountains have the largest suitable habitat area and the highest connectivity among habitat patches. However, habitat connectivity between the five mountains is very low, suggesting that gene flow among these mountain ranges is probably limited. We therefore recommend strengthening protection of sambar and their habitat, with special attention to the establishment of corridors between the different mountain populations.
1 Introduction
Ungulates are one of the most important animal groups, playing an irreplaceable role in the ecosystem (Chanchani et al., 2010). Ungulates have a significant impact on the growth of forest vegetation, especially on the restoration of secondary forest vegetation. Furthermore, ungulates occupy a specific ecological niche, playing a key role in the food chain. Ungulates provide an important link between material circulation and energy flow (Hobbs, 1996). Due in part to their important ecological roles, wild ungulate research is the subject of substantial conservation research, integrating species, community (Dar et al., 2012; Ofstad et al., 2016), and molecular and macroecology levels (Kreulen, 1985; Webb et al., 2010). Macroecological studies based on 3s (Remote Sensing, Global Position System, and Geographic Information System) technology are valuable for evaluating the status of habitat, and studies of habitat selection and habitat suitability can play a crucial role in ungulate conservation. Ungulate species richness and abundance are good indicators of habitat quality, size, quality, and connectivity (Jun and Ming, 2003; Xiao et al., 2008).
Factors affecting the distribution of forest ungulates are especially complex due to the complexity of the topography and vegetation structure of the forest ecosystem (Augustine and Mcnaughton, 1998). Habitat evaluation is based on assessments of occupied habitat, key factors affecting survival and reproduction, and analysis of the ecological drivers influencing the distribution of suitable habitat for the species (Jun and Ming, 2003). Because the choice of factors can directly affect the results of habitat evaluation (Wang, 2006), it is necessary to consider information such as scale, landscape patterns, and habitat connectivity.
Sambar (Rusa unicolor) is a social forest-dwelling ungulate with migratory habits (Dar et al., 2012; Dahlan and Dawend, 2013) native to South Asia. It is classified as a national secondary protected animal in China and was listed as Endangered on the IUCN Red List in 2013. There are four sambar subspecies (Rusa unicolor dejeani, Rusa unicolor equina, Rusa unicolor hainana, Rusa unicolor swimhoi) in China (He et al., 2019). According to historical records, the sambar historically had a wide distribution in China, extending across the area south of the Qinling Mountains and the Huaihe River (He, 1983). However, due to anthropogenic activities (habitat destruction, hunting, etc.), climate change, and other factors, the population and habitat for sambar have gradually diminished (Wang and Sheng, 1995), with its distribution shifting from the northeast to southwest mountains (He, 1983). As a species sympatric with giant pandas in Southwest China (Wang et al., 2018), sambar have benefited from the network of protected areas established for giant pandas. Under this protective umbrella, the sambar population in its core distribution has increased significantly in the past few decades (Zhang et al., 2015). However, certain local populations remain endangered. For example, the sambar population in Liangshan Mountain has been reported to be locally extinct (Mao et al., 2021).
At present, most habitat evaluations on sambar in China have been based on single reserves and little information is available at larger scales across reserves, creating a knowledge gap that impedes conservation action. Range-wide studies of landscape features and species occupancy are needed to reveal important information about sambar habitat requirements, dispersal and migratory patterns, spatial and temporal population dynamics, and the degree of remaining habitat connectivity. To address these information needs, we developed a species distribution model constructed with sambar occurrence records from camera traps. Our results provide basic data useful for the conservation, management, and restoration of habitat for sambar and help illuminate their role in the ecosystem.
2 Materials and methods
2.1 Study area
The study area was located in Southwest China, including five mountains: Minshan, Qionglai, Daxiangling, Xiaoxiangling, and Liangshan (101°00′34″~105°37′33″E, 27°49′04″~34°18′59″N) located on the eastern edge of the Qinghai–Tibet Plateau. Most of this area was characterized by high mountains and deep valleys (elevation ranging from 70 to 6,250 m) (Via et al., 2010; Yang et al., 2017). Environmental factors such as elevation, temperature, rainfall, and soil type contribute to the diversity of ecosystems that range from arid to subtropical to alpine (Yang et al., 2017).
This region is considered one of the 34 global biodiversity hot spots (Myers et al., 2000), with rich animal and plant resources and containing vulnerable and endangered wild animals such as giant panda (Ailuropoda melanoleuca), golden monkey (Rhinopithecus roxellanae), and snow leopard (Panthera uncia). Moreover, due to its rich vegetation types and extensive forest ecosystems, there are a number of large forest-dwelling ungulates in this area including Sichuan takin (Budorcas tibetanus), forest musk deer (Moschus berezovskii), Chinese serow (Capricornis milneedwardsii), Chinese goral (Naemorhedus griseus), and sambar deer (Wang et al., 2021).
2.2 Data collection
2.2.1 Occurrence data
We collected sambar occurrence data obtained from 2,099 camera traps (Figure 1, 2) installed across five major mountain ranges (Minshan, Qionglai, Daxiangling, Xiaoxiangling, and Liangshan) in Southwest China from 2015 to 2021. Camera traps were placed on a 1-km2 grid in key protected areas. Camera trap placement was in some cases constrained by terrain, and locations in excessively steep and dangerous terrain were avoided. We extracted location information from all photographs containing sambar to construct presence-only species distribution models.
Figure 1
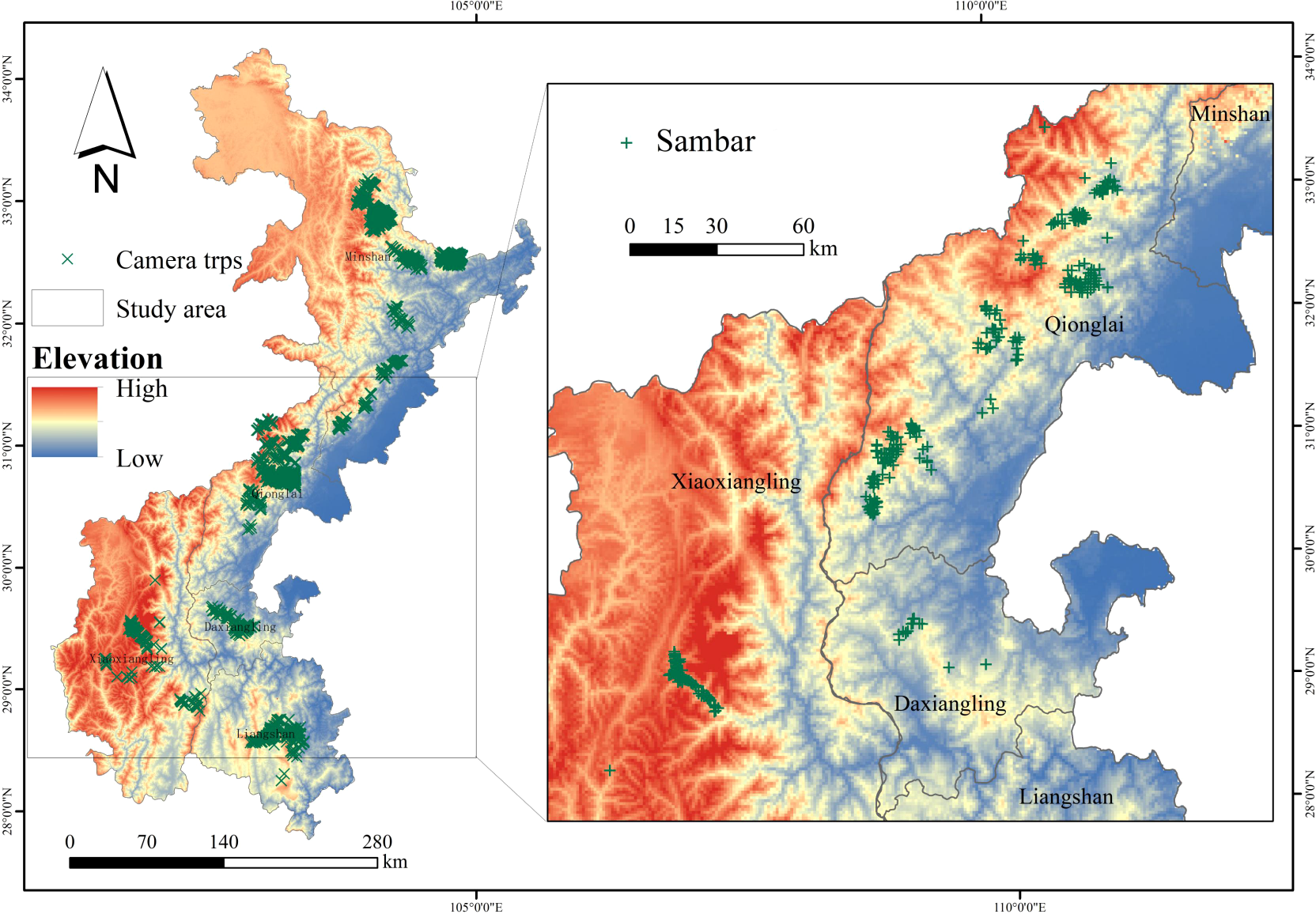
Camera traps and sambar in the study area.
Figure 2

Photos of sambar from camera traps.
2.2.2 Environment variables
Five environmental variables were selected to construct species distribution models, namely, climate, terrain, water source, vegetation, and disturbance. Climate variables included 19 bioclimatic factors (http://www.worldclim.org). Terrain variables included elevation, slope, and aspect and were extracted from the digital terrain elevation model (DEM) (http://www.gscloud.cn). The water source variable was the European distance layer generated based on the river and lake distribution layer of the five mountain systems to reflect the distance from each grid to the nearest water source. Vegetation variables included the normalized vegetation index (NDVI) (https://modis.gsfc.nasa.gov/) and surface vegetation classification (http://data.tpdc.ac.cn). The disturbance variables were the Euclidean distance layers of each grid from the nearest highway generated based on the road layer of the study area.
2.3 Data processing and model setting
2.3.1 Modeling potential distribution of habitat
We used the maximum entropy model (MaxEnt) to assess sambar habitat suitability. We used a 1-km2 occupancy grid, with each cell containing one or more camera traps with a sambar occurrence incorporated into a presence-only species distribution model. Surplus occurrence records from each grid cell were deleted based on the “ENMTools” package in R 4.0.3 to avoid spatial autocorrelation (Dan et al., 2010). We estimated the variance expansion factor (VIF) between the environmental variables and removed the environmental variables with VIF >10 to avoid multiple collinearities (Wang et al., 2021). The remaining variables were then used to build the preanalysis model, and environmental variables with contribution <1 were removed from the final MaxEnt model (Yang et al., 2020).
Construction and analysis of habitat suitability models were performed using software MaxEnt 3.4.1. Species occurrence points and environmental variables were introduced into the model, with 75% randomly selected as a model training set and the remaining 25% as a test set. The model regulation rate, convergence threshold, and maximum iterations were maintained as default. To ensure model stability, the model was performed with 10 bootstrap replicates and the mean value was used as model results. Model performance was assessed using the area under the subject operating characteristic curve (AUC) (Phillips et al., 2017). The contribution rate of environmental variables was estimated by a jackknife method to determine the importance of each environmental variable. Based on the habitat prediction results of the MaxEnt model, the habitat was divided into unsuitable habitat, suitable habitat, and highly suitable habitat. We define three categories by maximizing the sum of sensitivity and specificity, and the average of habitat suitability index for all suitable habitat (suitable habitat and highly suitable habitat).
2.3.2 Landscape pattern analysis
Based on the results of the species distribution model, the landscape pattern index of suitable habitat for sambar was calculated by using the landscape pattern analysis software Fragstats 4.2, including number of patches (NP), patch density (PD), mean patch size (MPS), largest patch size (LPS), and largest patch index (LPI). Habitat convergence and dispersion indicators included patch cohesion index (COHESION), segmentation index (DIVI-SION), and separation index (SPLIT). The definition, calculation method, and ecological significance of each landscape pattern index followed McGarigal and Marks (1995).
2.3.3 Habitat connectivity analysis
Based on the results of the species distribution model, we use the raster calculator in ArcGIS 10.4.2 to obtain the habitat connectivity resistance layer, by subtracting the habitat suitability index layer from 1. The habitat connectivity analysis software Circuitscape 4.0 was then used to calculate the habitat connectivity of sambar in the five major mountain ranges, and then the pairwise mode operation model was selected. The output of the circuit model was a probability graph representing species dispersal or movement in the landscape by current diffusion, describing the chance of the species moving between two or more predefined sites in the landscape (Liang et al., 2021).
3 Results
3.1 Suitable habitat evaluation for sambar
A total of 405 camera traps captured photos of sambar (Figure 1) in the five mountains, with presence detected in the Qionglai, Daxiangling, and Xiaoxiangling mountains. Sambar suitable habitat evaluation using the MaxEnt model showed that the model prediction accuracy (AUC = 0.958) is recognized as excellent model performance (Phillips et al., 2006; Phillips and Dudík, 2008).
The results of MaxEnt operation were reclassified by ArcGis10.4.2 to classify the suitable habitat of sambar. The model yielded estimates of 2,692 km2 (HSI >0.652) of highly suitable habitat, 15,539 km2 (0.331≤ HSI ≤0.652) of suitable habitat, and 88,872 km2 (HSI <0.331) of unsuitable habitat for sambar in the study area (Figure 3). The total suitable habitat area of sambar was 18,231 km2, accounting for 17.02% of the study area. The largest suitable habitat area was in the Qionglai mountains (8,176 km2), followed by Xiaoxiangling (4,474 km2), Minshan (3,174 km2), Liangshan (1,317 km2), and Daxiangling (1,090 km2). As shown in Figure 3, the most suitable habitat of sambar is mainly distributed in Qionglai mountains as well as the intersection area of three mountains of Xiaoxiangling, Minshan, and Qionglai.
Figure 3
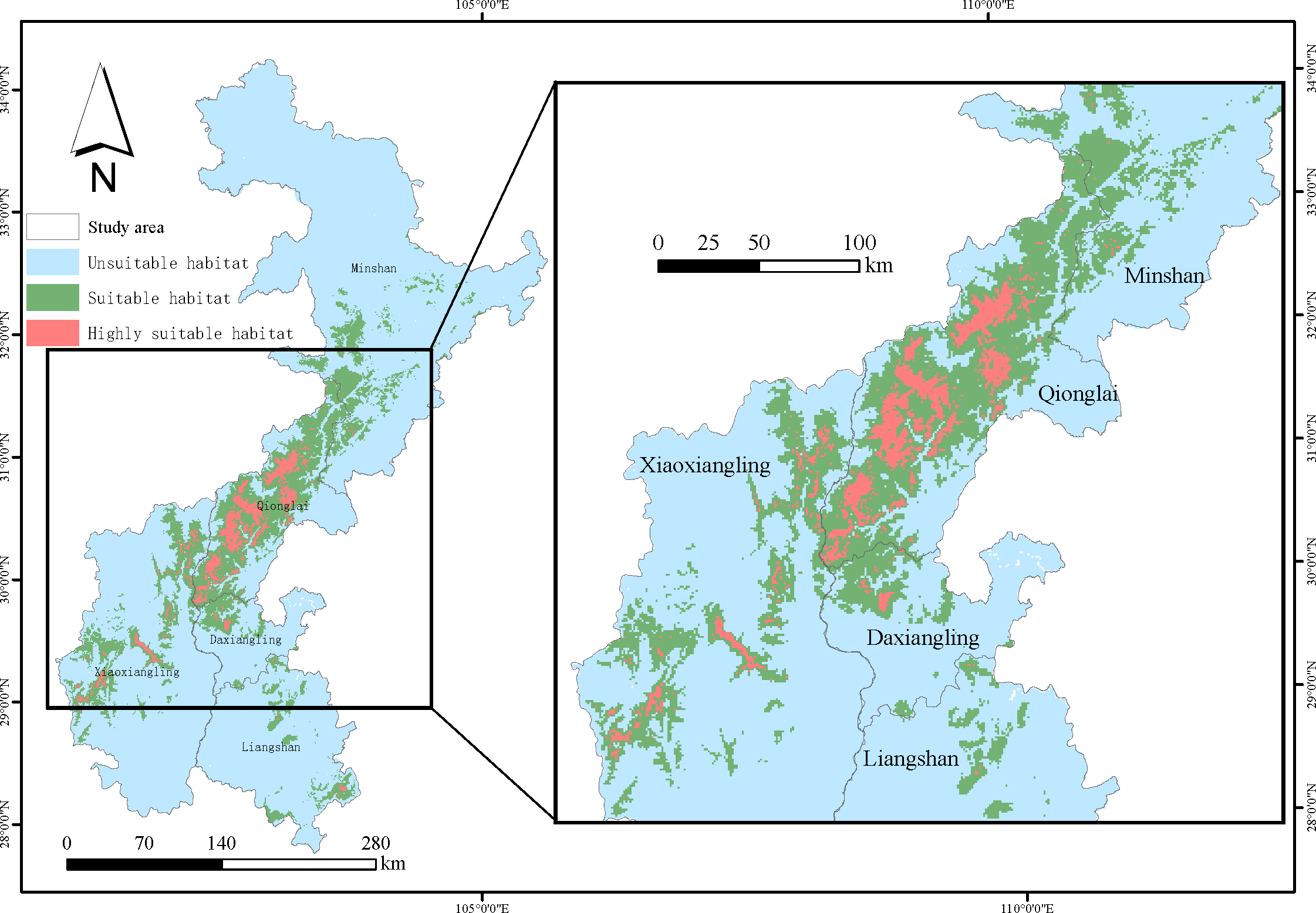
Suitable habitat of sambar, obtained from the MaxEnt species distribution model.
In the MaxEnt model, the importance of environmental variables was determined by jackknife resampling analysis to obtain the contribution rate of each environmental factor (Table 1). The four variables with the highest contribution rate were temperature annual range, temperature seasonality, altitude, and distance from the road, and their total contribution rate reached 82%. It showed that these four environmental factors were the key factors affecting the suitable habitat distribution of sambar.
Table 1
| Variables | Percent contribution |
|---|---|
| Temperature annual range (bio7) | 39.5 |
| Temperature seasonality (bio4) | 18.5 |
| Elevation | 12.1 |
| Distance to road | 11.9 |
| Normalized difference vegetation index (NDVI) | 7 |
| Precipitation seasonality (bio15) | 5.8 |
| Vegetation classification | 3.8 |
| Distance to the water | 1.8 |
| Slope | 1.5 |
Contribution rate of variables.
3.2 Landscape pattern
Eight indexes were selected to analyze the landscape pattern of sambar habitat (Table 2), including 273 habitat patches throughout the study area. The results show that the maximum area of habitat patches is 9,983 km2, accounting for 54.8% of the total suitable habitat area. It indicates the high quality of habitat patches suitable for sambar survival in the study area. However, overall, the segmentation index and separation index of each habitat patch were 0.99 and 106.58, respectively, indicating a relatively high habitat fragmentation of sambar in the study area.
Table 2
| Mountains | Landscape pattern index | |||||||
|---|---|---|---|---|---|---|---|---|
| NP | PD | MPS | LPS | LPI | COHESION | DIVISION | SPLIT | |
| Study area | 273 | 0.0025 | 66 | 9982 | 9.39 | 97.963 | 0.9906 | 106.58 |
| Qionglai | 19 | 0.0015 | 430 | 7498 | 57.51 | 99.513 | 0.6682 | 3.01 |
| Xiaoxiangling | 63 | 0.0023 | 71 | 1574 | 5.72 | 96.982 | 0.9931 | 143.92 |
| Minshan | 134 | 0.003 | 24 | 700 | 2.08 | 93.432 | 0.9992 | 1307.35 |
| Liangshan | 56 | 0.0038 | 24 | 332 | 2.46 | 91.202 | 0.9988 | 860.76 |
| Daxiangling | 15 | 0.0026 | 73 | 851 | 14.86 | 96.557 | 0.9774 | 44.33 |
Analysis of sambar habitat landscape pattern.
The number of patches in the Qionglai mountains was relatively small compared with the rest of the study area, and the mean patch size, largest patch size, largest patch index, and patch cohesion index were all significantly higher, indicating that the landscape pattern and habitat quality in the Qionglai mountains were better. Overall, the data from Daxiangling and Xiaoxiangling were similar and the quality of habitat patches suitable for sambar survival was lower than that of Qionglai. Minshan and Liangshan had higher patch numbers, the lowest patch density, and a significantly higher separation index than the other mountain ranges, which indicated that habitat patches in the two mountains were mainly dispersed as small patches with the highest habitat fragmentation and the lowest habitat quality.
3.3 Assessing sambar habitat connectivity
The results showed that most of the Qionglai mountains were characterized by high landscape connectivity, whereas the northern areas of the Daxiangling and Xiaoxiangling mountains adjoining Qionglai also contained high connectivity (Figure 4). Outside of these areas, the remaining area in Daxiangling and Xiaoxiangling had relatively low landscape connectivity. There was no evidence for high connectivity in the Minshan and Liangshan mountains. There was a disconnection between the three mountains. Connectivity between mountain ranges other than Qionglai was low, indicating that populations in these ranges are isolated from each other.
Figure 4
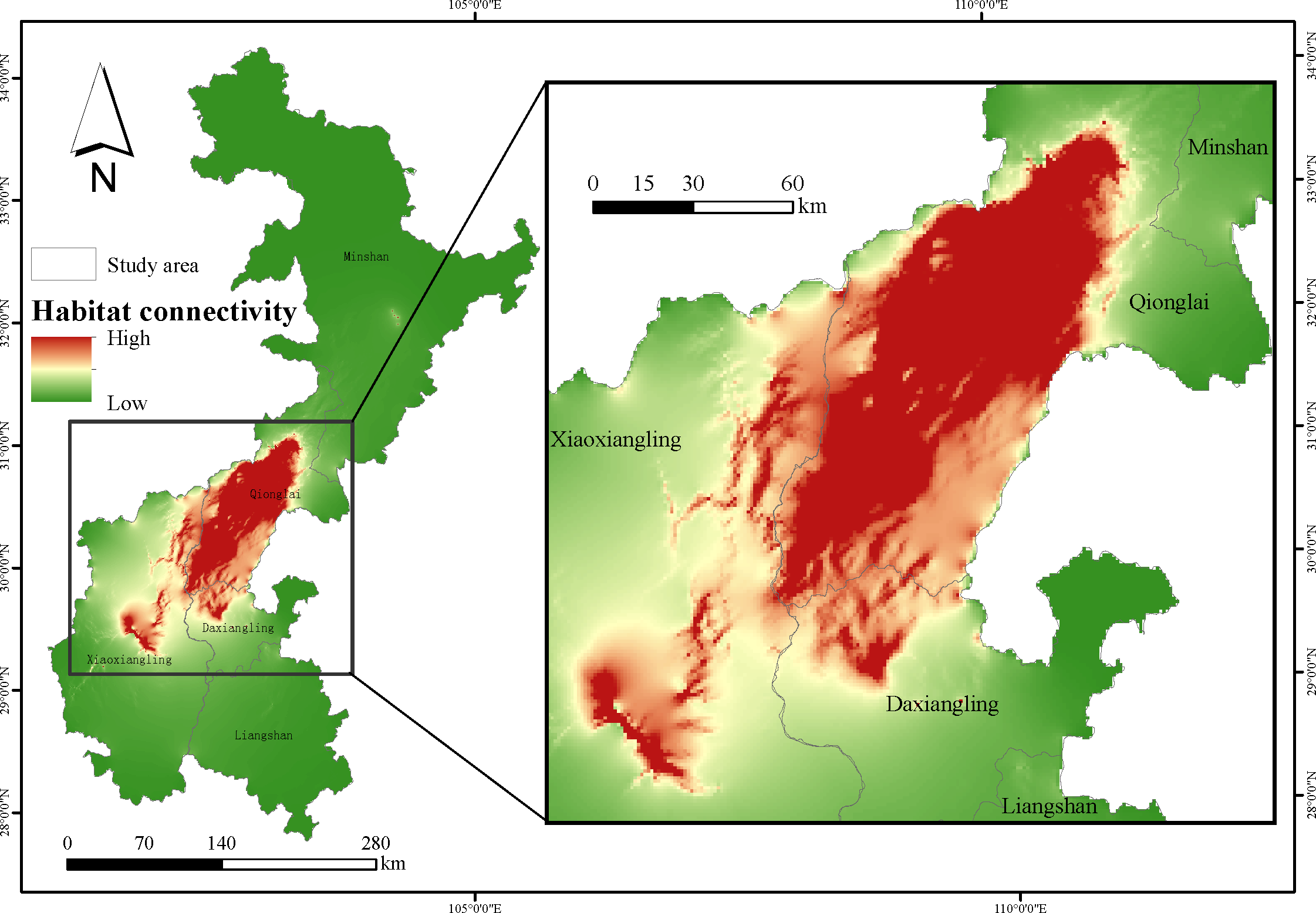
Habitat connectivity of sambar.
4 Discussion
The quantity and quality of habitat are closely related to animal survival and reproduction, and, consequently, habitat evaluation research is a foundational activity for biodiversity conservation (Yang et al., 2017). To reveal the drivers of animal movements for species that migrate or occupy large ranges, it is important to understand the distribution of suitable habitats and the ecological processes that underlay them. A thorough understanding of habitat quality requires knowledge of the potential extent of habitat area and the degree of connectivity among habitat patches to inform effective conservation policies.
Our findings on a scale of several mountain ranges indicate that sambar have suitable habitat distributed across all five mountain ranges, with suitable habitat for sambar comprising 17.02% of the total study area, a relatively small proportion. Qionglai, and the area at the intersection of three mountain ranges—Xiaoxiangling, Minshan, and Qionglai—contained the highest proportion of highly suitable habitat for sambar. These findings are consistent with previous studies indicating high population density of sambar in the Wolong, Saddle River, and Gongga mountains in Qionglai (Zhang et al., 2015; He et al., 2019), matching the distribution of highly suitable habitat in the present study.
It is well documented that the distribution of key environmental factors affects the distribution of animal populations (Elith and Leathwick, 2009). The most influential variables predicting sambar distribution in this study were annual temperature range, temperature seasonality, altitude, and distance from the road, which combined to account for 82% of model variance. These most highly ranked variables contributing to the model were both climatic factors, annual temperature range and temperature seasonality, indicating a key role for climate affecting the distribution of suitable habitat for sambar. Our findings obtained over a broad regional scale are consistent with previous results for studies conducted on a smaller scale, reinforcing and expanding on conclusions drawn from these studies. Important variables associated with sambar distribution include vegetation, elevation, slope, and climate (Kushwaha et al., 2004; Wang et al., 2018), and at least one study has found that climatic variables are the most influential in sambar habitat suitability models (Gong, 2020). In addition, the effect of elevation on the distribution of sambar in the present study is consistent with previous research indicating high habitat suitability at lower elevations (<3,000 m) (Pant et al., 1999; Hu et al., 2018).
Landscape pattern analysis showed that there were few large habitat patches suitable for sambar remaining in Southwest China. Habitat patches were distributed in an irregular and isolated pattern, indicating highly fragmented suitable habitat. The five mountain ranges included in this study varied substantively in the landscape’s suitability to support sambar survival. Sambar are large social, migratory forest ungulates (Dar et al., 2012) that require large home ranges to meet their foraging needs (Dahlan and Dawend, 2013). Species that are more sensitive to habitat area size are prone to greater negative impacts from habitat fragmentation, which may compromise range size, migration patterns, dispersal, and interspecies communication (Li et al., 2021). Qionglai, with its larger contiguous areas of suitable habitat, will likely support sambar populations much better than the fragmented landscape prevalent in Daxiangling, Xiaoxiangling, and Liangshan in Southwest China, where dispersal and long-term survival of sambar is compromised.
Evaluation of habitat connectivity was performed to assess the impact of habitat fragmentation on regional species persistence (Bowne et al., 1999; Tischendorf and Fahrig, 2000). This study found that habitat with high landscape connectivity was centrally distributed in the Qionglai mountains. Habitat in the region of Daxiangling and Xiaoxiangling adjacent to Qionglai also had high habitat connectivity, indicating that these regions are suitable for the survival and dispersal of sambar populations. It is helpful to fully understand the habitat situation of the species and develop a scientific conservation plan by combining evaluation of habitat suitability and connectivity (Ahmadi et al., 2017). We found that evidence of sambar presence was limited to Qionglai, Daxiangling, and Xiaoxiangling, which were characterized by high habitat suitability and habitat connectivity and low degree of habitat fragmentation, and the distribution of the species was associated with multiple factors. The poor connectivity between Qionglai, Daxiangling, and Xiaoxiangling will impede sambar migration, limiting gene flow between populations, and may eventually compromise population viability due to the loss of genetic diversity within small, isolated populations. Thus, our studies of landscape pattern and connectivity combined lead to the recommendation to establish an ecological corridor between the three mountains to improve habitat connectivity and promote sambar dispersal, migration, and gene flow.
Statements
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
PW, BF, and WB conceived the ideas and designed the experiments. BF, ZT, and WB collected the data. BF and WB contributed to the statistical analysis. PW, BF, XF, LZ, and WB conducted the analyses and coordinated in the writing of the manuscript. XD, JZ, and CZ revised it critically for important intellectual content. All authors contributed to the article and approved the submitted version.
Funding
This study was supported by China Postdoctoral Science Foundation (2021M690484), the Natural Science Foundation of Sichuan Province (2023NSFSC1152), the subsidy funds for national parks for forestry, grassland and ecological protection and restoration from the central government (51000022T000005003490), the Sichuan Science and Technology Program (2022YFS0468), and the fund of China West Normal University (22kA016).
Acknowledgments
We would like to thank all field teams and nature reserves for their assistance with sambar monitoring and field data collection.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
Ahmadi M. Balouchi B. N. Jowkar H. Hemami M. R. Fadakar D. Malakouti-Khah S. et al . (2017). Combining landscape suitability and habitat connectivity to conserve the last surviving population of cheetah in Asia. Diversity Dis.23, 592–603. doi: 10.1111/ddi.12560
2
Augustine D. J. Mcnaughton S. J. (1998). Ungulate effects on the functional species composition of plant communities: Herbivore selectivity and plant tolerance. J. Wildl. Manage.62, 1165–1183. doi: 10.2307/3801981
3
Bowne D. R. Peles J. D. Barrett G. W. (1999). Effects of landscape spatial structure on movement patterns of the hispid cotton rat (Sigmodon hispidus). Landscape Ecol.14, 53–65. doi: 10.1023/A:1008025827895
4
Chanchani P. Rawat G. S. Goyal S. P. (2010). Unveiling a wildlife haven: status and distribution of four trans-Himalayan ungulates in sikkim, India. Oryx44, 366–375. doi: 10.1017/S0030605310000293
5
Dahlan I. Dawend J. (2013). Growth and reproductive performance of sambar deer in sabal forest reserve of Sarawak, Malaysia. Trop. Anim. Health prod.45, 1469–1476. doi: 10.1007/s11250-013-0383-6
6
Dan L. W. Glor R. E. Turelli M. (2010). ENMTools: a toolbox for comparative studies of environmental niche models. Ecography33, 607–611. doi: 10.1111/j.1600-0587.2009.06142.x
7
Dar T. A. Habib B. Khan J. A. (2012). Group size, habitat use and overlap analysis of four sympatric ungulate species in shivalik ecosystem. Uttarakhand India Mammal.76, 31–41. doi: 10.1515/mammalia-2012-0502
8
Elith J. Leathwick J. R. (2009). Species distribution models: Ecological explanation and prediction across space and time. Annu. Rev. Ecol. Evol. Syst.40 (1), 677–697. doi: 10.1146/annurev.ecolsys.110308.120159
9
Gong X. (2020). Spatial distribution pattern and suitable habitat prediction of rusa unicolor and budorcas taxicolor in anzihe reserve, sichuan province. Univ. Chin. Acad. Sci. doi: 10.27525/d.cnki.gkchs.2020.000018
10
He Y. (1983). The history of sambar in eastern China. J. Guangxi Normal Univ. (Natural Sci. Edition)01 (1983), 79–86. doi: CNKI:SUN:GXSF.0.1983-01-012
11
He X. Fu Q. Wu Y. Wang B. Chen X. Ran J. (2019). Population structure and activity rhythm of rusa unicolor. Acta Theriol. Sinica.39, 134–141. doi: 10.16829/j.slxb.150195
12
Hobbs N. T. (1996). Modification of ecosystems by ungulates. J. Wildl. Manage.60 (4), 695–713. doi: 10.2307/3802368
13
Hu J. Yao G. Li D. Yang Z. Li Y. (2018). Summer habitat selection of sambar deer (Rusa unicolor) in wolong national nature reserve. Acta Theriol. Sinica.38, 277–285. doi: 10.16829/j.slxb.150142
14
Jun W. Z. Ming L. Y. (2003). Effects of habitat fragmentation on survival of animal populations. Acta Ecol. Sin.23 (11), 2424–2435. doi: 10.3321/j.issn:1000-0933.2003.11.027
15
Kreulen D. (1985). Lick use by large herbivores: a review of benefits and banes of soil consumption. Mammal. Rev.15, 107–123. doi: 10.1111/j.1365-2907.1985.tb00391.x
16
Kushwaha S. Khan A. Habib B. Quadri A. Singh A. (2004). Evaluation of sambar and muntjak habitats using geostatistical modelling. Curr. Sci.86 (10), 1390–1400. doi: stable/24109211
17
Liang J. Ding Z. Jiang Z. Yang X. Xiao R. Singh P. B. et al . (2021). Climate change, habitat connectivity, and conservation gaps: a case study of four ungulate species endemic to the Tibetan plateau. Landscape Ecol.36, 1071–1087. doi: 10.1007/s10980-021-01202-0
18
Li Q. Li S. Cao M. Xu H. (2021). Habitat suitability evaluation and corridor design of black jac (Muntiacus crinifrons) in qianjiangyuan national park. Journal of Ecology and Rural Environment. 37 (6), 778–785. doi: 10.19741/j.issn.1673-4831.2020.0822
19
Mao Z. Bai W. Fu L. Cai T. Huang Y. Hong Y. et al . (2022). Investigation on beasts of suspicious distribution in mabian dafengding nature reserve. J. China West Normal Univ. (Natural Sci. Edition)43 (1), 1–8. doi: 10.16246/j.issn.1673-5072.2022.01.001
20
McGarigal K. Marks B. J. (1995). Fragstats: Spatial pattern analysis program for quantifying landscape structure. USDA Forest Service - General Technical Report PNW. 351 (Portland).
21
Myers N. Mittermeier R. A. Mittermeier C. G. Fonseca G. Kent J. (2000). Biodiversity hotspots for conservation priorities. Nature403, 853–858. doi: 10.1038/35002501
22
Ofstad E. G. Herfindal I. Solberg E. J. Sther B. E. (2016). Home ranges, habitat and body mass: simple correlates of home range size in ungulates. proceedings of the royal society b. Biol. Sci.283 (1845), 20161234. doi: 10.1098/rspb.2016.1234
23
Pant A. Chavan S. G. Roy P. S. Das K. K. (1999). Habitat analysis for sambar in corbett national park using remote sensing and gis. J. Indian Soc. Remote Sens27, 133–139. doi: 10.1007/BF02991566
24
Phillips S. J. Anderson R. P. Dudík M. Schapire R. E. Blair M. E. (2017). Opening the black box: an open-source release of maxent. Ecography40 (7), 887–893. doi: 10.1111/ecog.03049
25
Phillips S. J. Anderson R. P. Schapire R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecol. Model.190, 231–259. doi: 10.1016/j.ecolmodel.2005.03.026
26
Phillips S. J. Dudík M. (2008). Modeling of species distributions with maxent: new extensions and a comprehensive evaluation. Ecography31, 161–175. doi: 10.1111/j.0906-7590.2008.5203.x
27
Tischendorf L. Fahrig L. (2000). On the usage and measurement of landscape connectivity. Oikos90, 7–19. doi: 10.1034/j.1600-0706.2000.900102.x
28
Via A. Tuanmu M. N. Xu W. Yu L. Liu J. (2010). Range-wide analysis of wildlife habitat: Implications for conservation. Biol. Conserv.143, 1960–1969. doi: 10.1016/j.biocon.2010.04.046
29
Wang Y. (2006). Study on landscape ecological pattern and restoration of forest musk deer’s habitat in fengxian county (Shanxi Province: East China Normal University).
30
Wang P. Bai W. Hang J. Zhang J. Liu D. Xia S. et al . (2018). Habitat use of differentiation between sympatric giant panda and sambar. Acta Ecol. Sinica.38 (15), 311–317. doi: 10.5846/stxb201801210162
31
Wang X. Sheng H. (1995). The status of sambar (Rusa unicolor) in China. J. Wildl.3, 7–8. doi: 10.19711/j.cnki.issn2310-1490.1995.03.002
32
Wang F. Winkler J. Via A. Mcshea W. J. Liu J. (2021). The hidden risk of using umbrella species as conservation surrogates: A spatio-temporal approach. Biol. Conserv.253, 108913. doi: 10.1016/j.biocon.2020.108913
33
Webb S. L. Gee K. L. Strickland B. K. Demarais S. DeYoung R. W. (2010). Measuring fine-scale white-tailed deer movements and environmental influences using GPS collars. Int. J. Ecol.2010, 1–12. doi: 10.1155/2010/459610
34
Xiao Y. Ouyang Z. Y. Zhao J. Z. Huang B. R. Zhu C. Q. (2008). An assessment of giant panda(Ailuropoda melanoleuca) habitat stressors in minshan region. Acta Ecol. Sin.28 (1), 267–273. doi: 10.3321/j.issn:1000-0933.2008.01.031
35
Yang N. Ma D. Zhong X. Yang K. Zhou Z. Zhou H. et al . (2020). Habitat suitability assessment of blue eared-pheasant based on MaxEnt modeling in wanglong national nature reserve, sichuan province. Acta Ecol. Sinica.40 (19), 7064–7072. doi: 10.5846/stxb201908081666
36
Yang H. Viña A. Tang Y. Zhang J. Wang F. Zhao Z. et al . (2017). Range-wide evaluation of wildlife habitat change: A demonstration using giant pandas. Biol. Conserv.213, 203–209. doi: 10.1016/j.biocon.2017.07.010
37
Zhang J. Li Y. Li R. (2015). Application of infrared camera technology in studies of mammal activity patterns. Sichuan J. Zool.34, 671–676. doi: 10.11984/j.issn.1000-7083.20150172
Summary
Keywords
habitat suitability, population distribution, habitat connectivity, wildlife conservation, sambar (Rusa unicolor)
Citation
Wang P, Feng B, Zhang L, Fan X, Tang Z, Dong X, Zhang J, Zhou C and Bai W (2023) Assessment of habitat suitability and connectivity across the potential distribution landscape of the sambar (Rusa unicolor) in Southwest China. Front. Conserv. Sci. 3:909072. doi: 10.3389/fcosc.2022.909072
Received
31 March 2022
Accepted
30 December 2022
Published
20 January 2023
Volume
3 - 2022
Edited by
Ronald R. Swaisgood, San Diego Zoo Wildlife Alliance, United States
Reviewed by
Sathyakumar Sambandam, Wildlife Institute of India, India; Daniel Ubiali, Universidade Federal Rural do Rio de Janeiro, Brazil
Updates
Copyright
© 2023 Wang, Feng, Zhang, Fan, Tang, Dong, Zhang, Zhou and Bai.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wenke Bai, baiwk2006@126.com
†These authors have contributed equally to this work
This article was submitted to Animal Conservation, a section of the journal Frontiers in Conservation Science
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.