
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
BRIEF RESEARCH REPORT article
Front. Conserv. Sci. , 07 October 2022
Sec. Global Biodiversity Threats
Volume 3 - 2022 | https://doi.org/10.3389/fcosc.2022.889501
This article is part of the Research Topic Disentangling the Complexity of Fence Effects on Wildlife and Ecosystems View all 8 articles
 Peter Tyrrell1,2,3*
Peter Tyrrell1,2,3* Robert Buitenwerf4,5
Robert Buitenwerf4,5 Peadar Brehony6
Peadar Brehony6 Mette Løvschal7,8Jake Wall9Samantha Russell1Jens-Christian Svenning4,5David W. Macdonald2
Mette Løvschal7,8Jake Wall9Samantha Russell1Jens-Christian Svenning4,5David W. Macdonald2 Johan T. du Toit10,11John Kamanga1
Johan T. du Toit10,11John Kamanga1Globally, rangelands are undergoing rapid social-ecological changes, yet the scale of these changes is rarely measured. Fencing, sedentarization, and land conversion limit access by wildlife and livestock to vital resources such as water and seasonal forage, leading to rangeland degradation. In addition, these changes limit connectivity between wildlife sub-populations, triggering a spiral of decreasing biodiversity and weakening ecosystem function. Moreover, the combination of land privatization, sedentarization and fencing endangers pastoral livelihoods by reducing resilience to drought and diminishing livestock holdings per person. We provide a unique, urgent, and vital snapshot across >30,000 km2 of southern Kenya’s rangeland, covering four ecosystems renowned for their rich megafauna and pastoral people. We document and explore the drivers of extensive fencing (~40,000 km), the proliferation of livestock enclosures (>27,000), and the conversion of rangelands for cultivation (~1,500 km2). Our analyses were based on an open-access database recently synthesized for the region. Fencing is generally more prolific in areas that have been converted from community tenure to private title, especially where land values are raised by agricultural potential and proximity to Kenya’s capital, Nairobi. These factors drive the transfer of land ownership from traditional pastoralists to speculators, eventually resulting in the transformation of rangeland into agricultural, industrial and urban land uses. Space for wildlife (and traditional pastoralism) is limited on private, subdivided land, where livestock enclosures are at their highest density, and where there is less unfenced land and less untransformed land, compared to conservation areas and pastoral commons. Conflicting planning incentives, policies, and economic forces are driving unsustainable and potentially irreversible social-ecological transitions over unprecedented spatial scales. The lesson from southern Kenya is that a range of financial, policy and governance-related interventions are required to allow people and nature to coexist sustainably in African savannas.
Rangelands cover >30% of the world’s terrestrial surface, supporting billions of people and nearly half of all livestock and sustaining biodiversity and vital ecosystem services (Briske, 2017). Yet many rangelands are undergoing radical social and ecological transitions, jeopardizing global plans to “prevent, halt, and reverse the degradation of ecosystems worldwide” during the UN Decade of Ecosystem Restoration1 (Reid et al., 2014). Biodiversity is decreasing mainly through land conversion and fragmentation (Hobbs et al., 2008; Reid et al., 2014). East Africa is one region that still hosts pastoralist social-ecological systems in which the rangeland is shared with a high abundance and diversity of large mammals (Western et al., 2020). However, these systems face profound degradation due to widespread agricultural expansion, sedentarization of pastoral people, and accelerating fencing, which is fragmenting ecosystems and threatening their sustainability (Reid et al., 2014; Jakes et al., 2018; Western et al., 2020; McInturff et al., 2020). The scale of changes in sedentarization and fencing are poorly understood, primarily because of a lack of large-scale spatial data (Hobbs et al., 2008).
Recently, we synthesized one of the largest known datasets for rangelands on the extent of fencing, agricultural land use, and the location of, and fencing around, livestock enclosures (landDX; Tyrrell et al., 2022). The study area covers 30,000 km2 and includes four large, adjacent ecosystems in southern Kenya with comparable social-ecological dynamics: Maasai Mara, Loita Hills, the South Rift Valley and Amboseli (Figure 1). We identified nearly 40,000 km of linear fencing—in a straight line these fences would encircle the earth—and 1,500 km2 of former rangeland converted to agricultural land, as well as 27,000 livestock enclosures. There is evidence of severe landscape fragmentation since the 1960s, when fences were virtually absent, with much of the change occurring very recently (Løvschal et al., 2017). Our aim in this research report is to describe some preliminary analyses based on landDX with the hope of attracting other researchers to use this open-access database. There is a dire need for such studies to address the multicausal and highly complex social and environmental challenges to the sustainability of rangeland ecosystems.
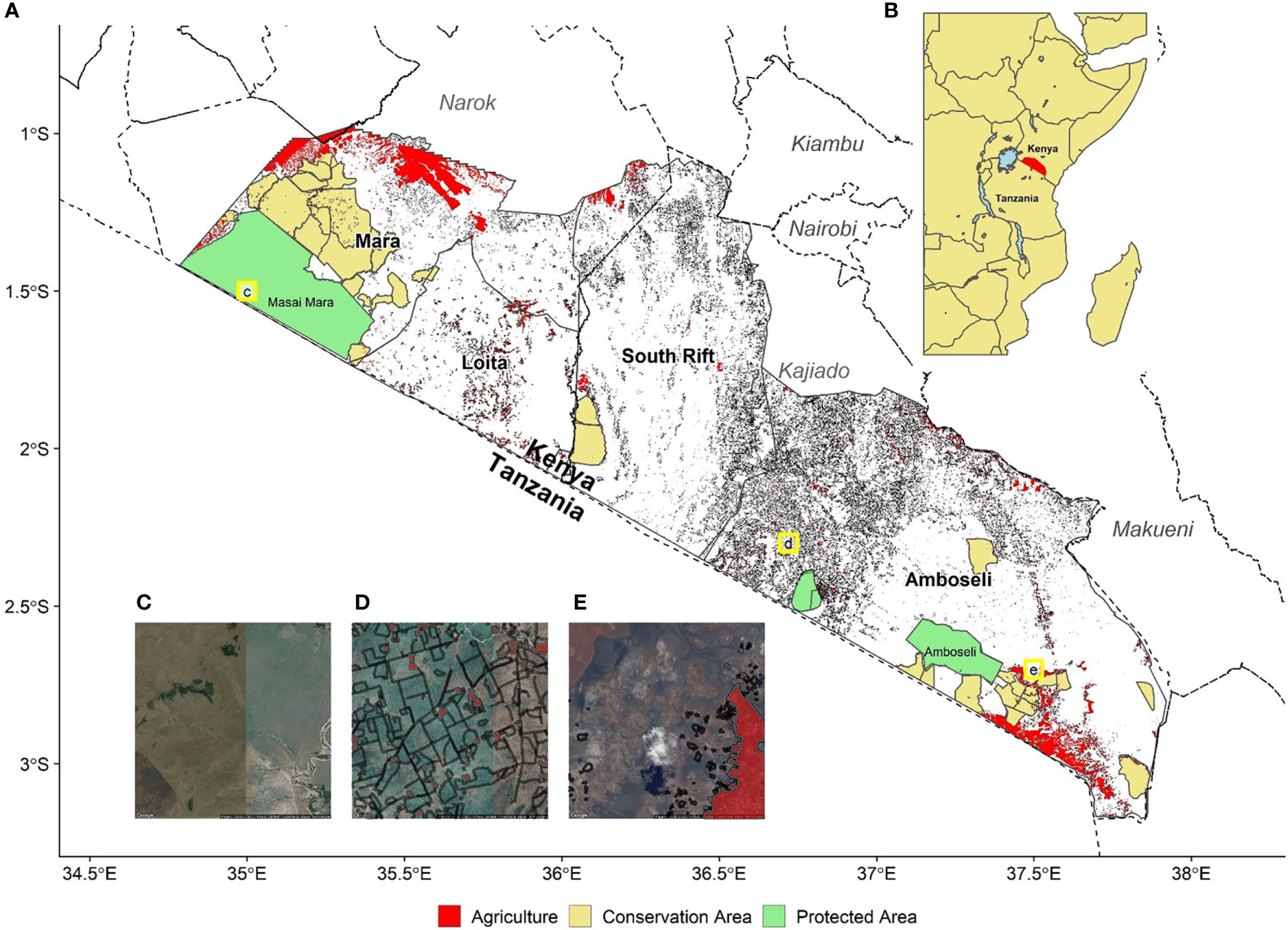
Figure 1 (A) ~40,000 km of fencing (black) and ~1,500 km2 of agriculture (red) across four southern Kenya ecosystems (left to right: Maasai Mara; Loita, South Rift Valley, Amboseli). Kenya county boundaries are shown for reference (dashed lines); (B) inset showing the study area (red) within East Africa; (C–E) zoomed in panels of three areas under different states of fencing and agricultural development, with Google Maps imagery for reference.
We quantified the extent of fencing, sedenterization, and land conversion across four iconic ecosystems with comparable but unique social-ecological dynamics: Amboseli; South Rift Valley; Loita; Maasai Mara. Data on land tenure in Southern Kenya were collected and split into four categories as defined by the Constitution (Government of Kenya, 2010) and Wildlife Conservation and Management Act, 2013 (Government of Kenya, 2013): (i) Protected area (State Forest Reserves, National Parks, and National Reserves), land where human settlement is limited and the land is under public ownership; (ii) Conservation Areas, private and community land which has been set aside for wildlife conservation; (iii) community land, land under de facto or de jure community ownership; and (iv) private land, land which has been subdivided with land title deeds issued to individuals. These data were collated from multiple sources including BigLife Foundation, Maasai Mara Conservancies Association, South Rift Association of Land Owners, Kenya Wildlife Conservancies Association, and the African Conservation Centre. Due to the many conflicting land claims, and potentially inaccurate boundaries, these data represent the best estimate of land tenure at a broad scale and are by no means definitive.
Ecosystem boundaries were digitized from original drawings of the Kenya Rangeland Ecological Monitoring Unit (KREMU), who split Kenya into 44 ecosystems (ecological units), based on topography, vegetation, and wildlife migration patterns and their seasonality (Stelfox and Peden, 1981). We adjusted the boundaries of the ecosystem to split the Nairobi and Amboseli ecosystems based on evidence from the literature and wildebeest collar data which track large ungulate migration patterns (Stabach, 2015; Said et al., 2016; Ojwang’ et al., 2017; Stabach et al., 2020). This resulted in four ecosystems covering the Southern Kenyan rangelands: Amboseli, South Rift, Loita, and Mara.
Generally, spatial data on fences and agricultural expansion are localized and dispersed among data owners and databases. Previously, Tyrrell et al. (2022) synthesized data from several research groups and conservation NGOs into a spatial-temporal database. The data include 27,000 livestock enclosures, nearly 40,000 km of fencing, and 1,500 km2 of agricultural land. As reported by Tyrrell et al. (2022), when data overlapped between research groups, the features with the more recent collection date were selected, and all data layers were clipped to the extent of data collection files provided with each data source.
In this paper, we used the data from the landDX open-access database (Tyrrell et al., 2022) to generate several metrics to determine the distribution and total impact of fencing and agricultural land use within each ecosystem. First, we calculated the extent of agriculture within each ecosystem. Second, we calculated the length of fencing in each ecosystem (Amboseli, South Rift, Loita, Mara). We then calculated the density of fences within non-agricultural land. We did this because the data we collected did not demarcate fences within agricultural land use. To demonstrate the spatial distribution of fencing within ecosystems, we created a 0.05° grid and calculated fence density, as kilometer of fencing per square kilometer, for each cell.
We performed a Spearman correlation test between fence density within the 0.05° grid to log10 transformed land price. Land prices (in USD) were calculated in Tyrrell et al. (2021) using scraped internet advertisements of land sales from January 2018, which were modelled using a Generalized Additive Model with socio-economic and ecological covariates (Tyrrell et al., 2021). We assigned the dominant land tenure category to each 0.05° grid cell and conducted non-parametric Kruskal-Wallis tests on fence data between the four land tenure groups, sequentially ordered by median fence density.
For each land tenure, we calculated the cumulative distribution function (CDF) of the distance from a fixed point in space to the nearest livestock enclosure, calculated using the empty space function F of a stationary point process (Baddeley et al., 2015). Protected areas are excluded because they have few or no settlements within their boundaries. We then fitted a Poisson point process model that assumes that the point process’ intensity (livestock enclosure) is a function of land tenure and compared it to the null model and a model with mean annual rainfall in addition to land tenure and evaluated the best fitting models using log-likelihood. These analyses were all conducted in R version 3.5.1 (R Core Team, 2018), with the packages sf (Pebesma, 2018), tidyverse (Wickham, 2019), raster (Hijmans, 2022), furrr (Vaughan and Dancho, 2020) , and exactextractr (Baston, 2022).
First, Amboseli, a 10,000 km2 area, provides habitat for several thousand elephants, hundreds of lions, and thousands of wildebeest and zebra. Private land in this area is now heavily fenced (21,150 km) with a median fence line density of 1.78 km.km-2 (Figures 1, 2). In addition to high densities of livestock enclosures, agriculture covers 428.09 km2or 3.4% of the area (Figure 1). This proliferation of fencing, sedenterization, and land transformation has fragmented the ecosystem, with severe implications for pastoral livelihoods (Kimiti et al., 2018) and the connectivity of wildlife populations (Groom and Western, 2013; Osipova et al., 2018). Yet, considerable areas of very low-density fencing remain around the Amboseli National Park (Figures 1, 2), especially in areas where communities have set aside parts of their land for conservation. Many of these communities are paid annual lease fees by conservation NGOs, supporting wildlife’s persistence around the National Park, although subdivision and fencing are also underway here.
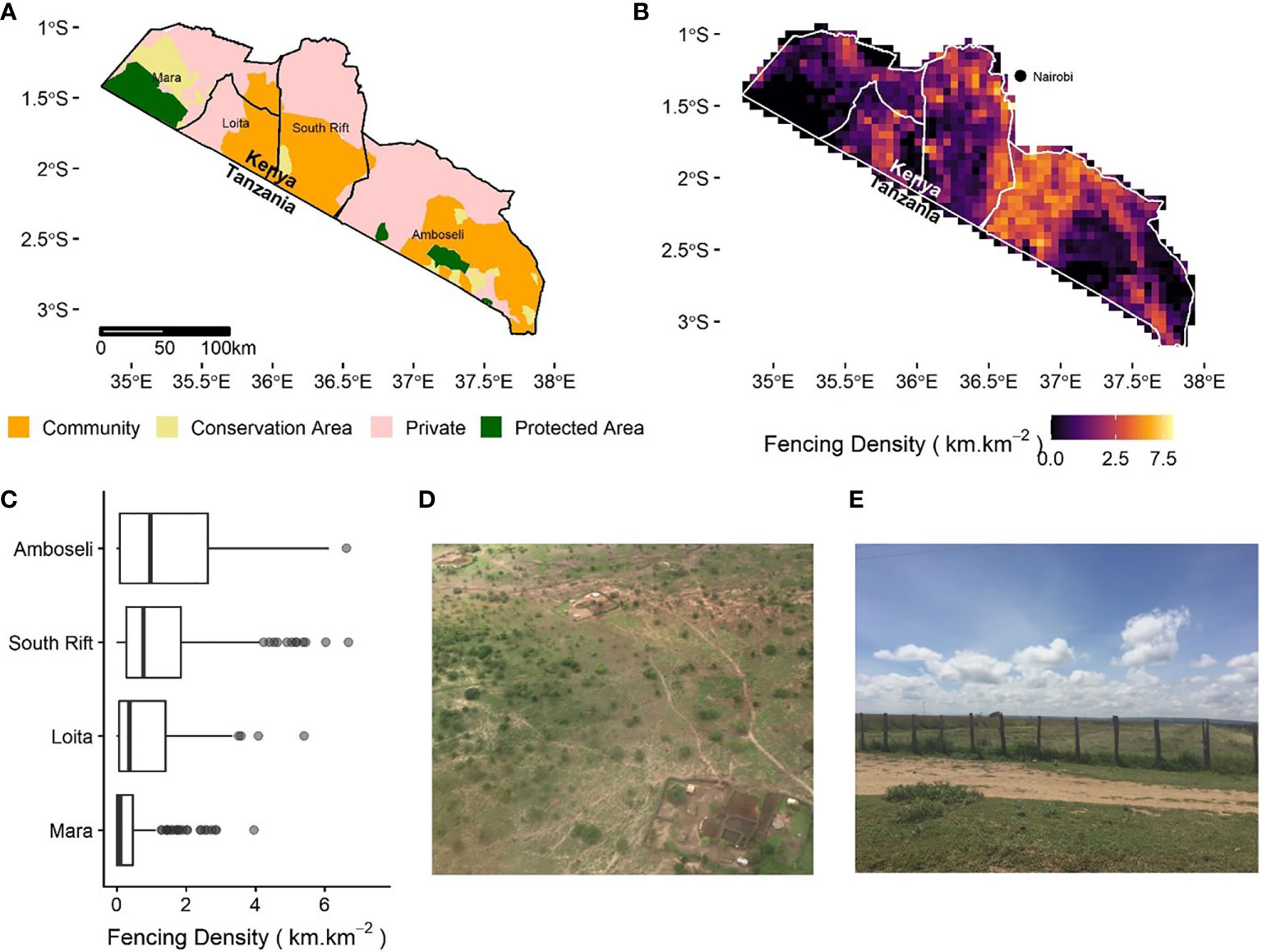
Figure 2 (A) The four ecosystems in southern Kenya: Amboseli, South Rift Valley, Loita Hills, and Maasai Mara. Government-protected areas (in green) are shown together with established community and private conservation areas (in yellow); the remaining land is subdivided and individually titled “Private” land without conservation efforts (in pink) and collectively owned or managed “Community” land (in orange); (B) fence density calculated as km.km-2 calculated on a 0.05-degree grid (~5 km resolution). Nairobi, Kenya’s capital city, is displayed for reference; (C) boxplot of fence density within each ecosystem, with median (vertical line), first and third quartiles (box) and 1.5 times the interquartile range (whiskers); (D) example of brush fences around settlements; (E) wire and post fencing to demarcate property in the Loita plains (north-east of the Mara ecosystem).
Second, the South Rift Valley, an 8,000 km2 area, is primarily under communal ownership with a diverse and abundant wildlife community. Here, private land in parts of the ecosystem has high densities of livestock enclosures and fences (1.3 km.km-2 median, 11,152 km in total: Figure 1) but areas in the south and west remain with very low-density settlement and fencing, including community conservation areas (Figures 1, 2). In these areas, wildlife and livestock still maintain considerable mobility, which is vital for their long-term survival (Tyrrell et al., 2017). The arid climate of the southern and western parts of the South Rift Valley makes them unsuitable for rainfed agriculture and the local community maintains traditional governance structures that manage the landscape for extensive pastoralism (Brehony, 2020). These factors have prevented land subdivision and conversion to cropland (presently only 0.9% of total area) or urban development (Western et al., 2020).
Third, the Loita ecosystem (~3,000 km2) has undergone considerable fencing (3,029 km total, 0.901 km km-2), sedentarization, and conversion to agriculture (80.06 km2, 2.3% of total area; Figures 1, 2), which now limits the mobility of large mammals between the Maasai Mara and South Rift Valley. The Loita Forest is a vital part of this ecosystem, where considerable areas remain with very low-density fencing and settlement (adjacent to the South Rift Valley). It is an important water source for both the Mara-Serengeti and South Rift Valley ecosystems, a central connection for large mammal movement across the greater Kenya-Tanzania borderlands region, a critical area for carbon sequestration, and a biodiversity hotspot (Broekhuis et al., 2018).
Finally, the Maasai Mara ecosystem (~7,000 km2), world-renowned for its large wildlife populations, still has a comparatively low intensity of fencing (3,039 km total, 0.477 km.km-2 median) and livestock enclosures due to the Maasai Mara National Reserve and the surrounding conservation areas, supported by eco-tourism lease fees. However, as reported by Løvschal et al. (2017), there has recently been a rapid expansion of fencing around the Maasai Mara National Reserve and within surrounding ‘conservation areas’ (Figures 1, 2), with associated wildlife declines including the near complete collapse of the Loita Plains migration of >300,000 wildebeest (Løvschal et al., 2018). Large-scale conversion to agriculture has occurred in key wet season grazing areas for wildlife, extending south from private lands in the north (8.3% of the ecosystem; 579.74 km2; Figures 1, 2).
Fencing has escalated most rapidly in areas with increasing land prices (Figure 3), driven by proximity to Kenya’s capital, Nairobi, and agricultural potential (Tyrrell et al., 2021). These factors encourage the transfer of land from traditional communities to urban speculators and its eventual transformation into agricultural, industrial, or urban land-uses (Figure 3) (Rutten, 1992; Tyrrell et al., 2021). The Nairobi National Park and Athi-Kapiti ecosystem, adjacent to Nairobi, are now almost completely fragmented with a collapse of the previously rich migratory system of wildlife and pastoralism (Said et al., 2016; Nkedianye et al., 2020). Economically, subdivision into private parcels may empower landowners, at least in the short-term, allowing them entry into the cash economy through land sales, loans on title deeds, and subsistence agriculture. However, land sales are often driven by wealthy and powerful investors external to the local community (Rutten, 1992). Without access to grazing land, pastoralists will eventually be forced to abandon their traditional lifestyle and culture, risking a poverty trap (Rutten, 1992). Poor planning and weak local leadership during the subdivision process itself, often led by local elites or government officials, can exacerbate inequalities within communities and worsen the ecological impacts of fencing and land conversion (Galaty, 1999; Mwangi, 2007).
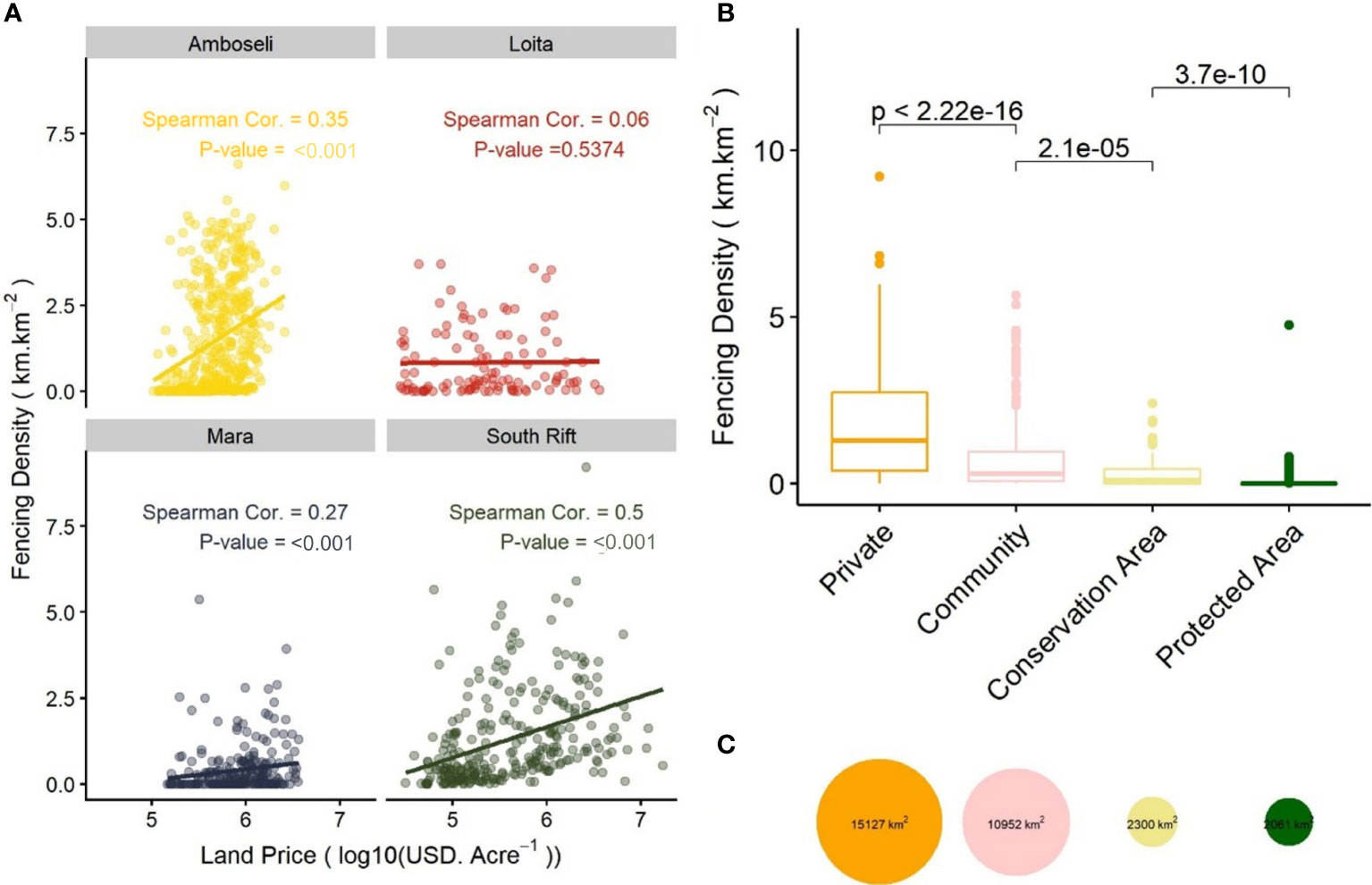
Figure 3 (A) The density of fences is positively correlated with the price of land in Amboseli, Mara and South Rift. The effect is strongest in Amboseli and the South Rift, where prices and land conversion are high and driven by the proximity to Nairobi, Kenya’s capital, and main roads. (B) Boxplots of fence density in each land-tenure type across the region. Private areas have the highest fence density, followed by community areas, conservation areas, and protected areas. The comparison text indicates p-values from a Kruskal-Wallis test between groups. (C) Bubble plot comparing the total area of each land-tenure type.
The density of livestock enclosures was lowest in protected areas followed by conservation areas, community land, and private subdivided land (Figure 4). Rainfall and land tenure both explained the distribution of livestock enclosure density (PPM, df= 4, p-value < 0.0001). Private land in Amboseli, the South Rift, and the Mara ecosystem showed some of the highest average densities (Figure 4) whereas the largest spaces between livestock enclosures were predictably in conservation areas (Figure 4). On the other end of the spectrum, private subdivided land had the least open space between livestock enclosures, which is related to the sedentarization of pastoral households during privatization and has worrying implications for rangeland sustainability (Groom and Western, 2013).
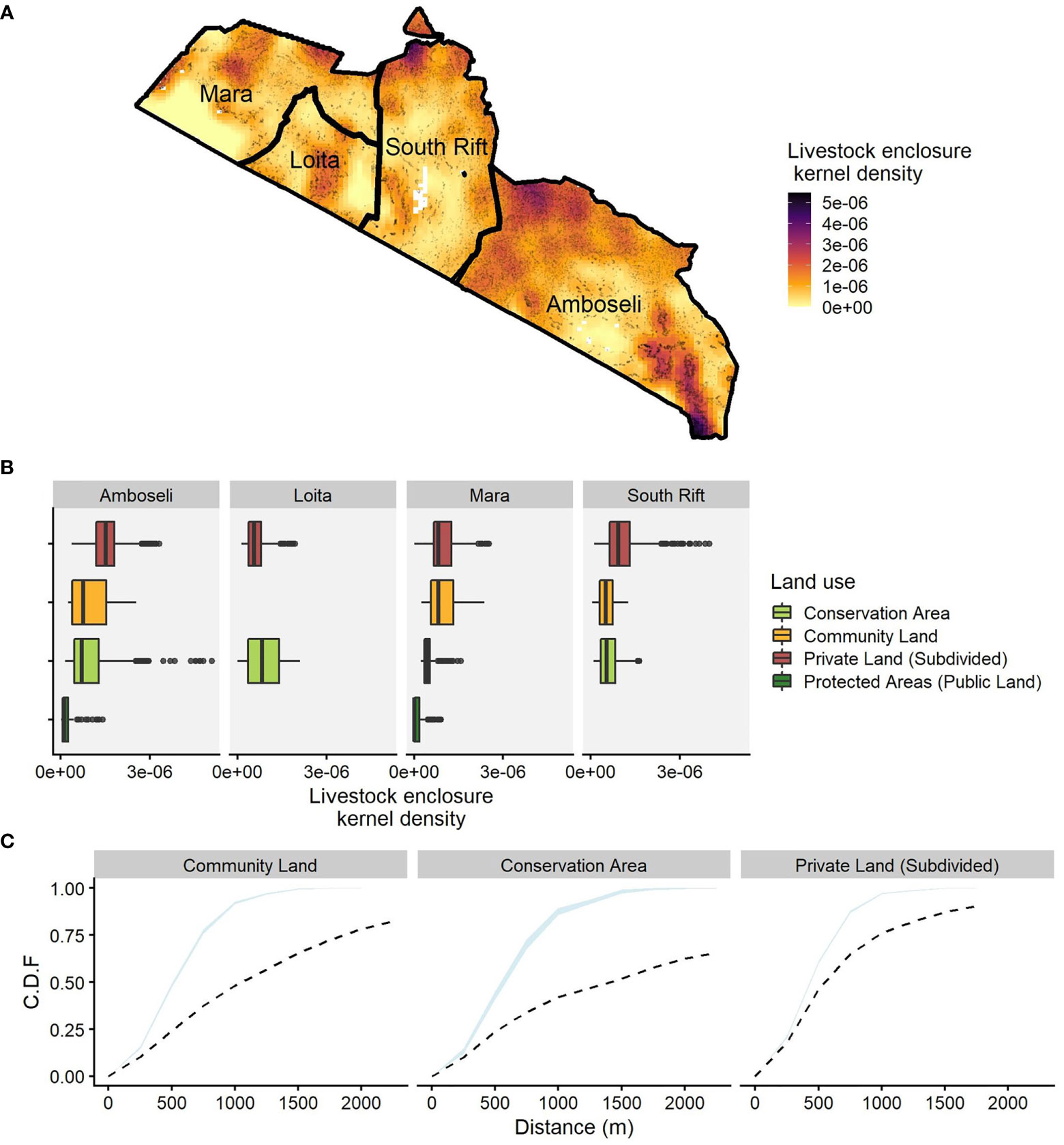
Figure 4 In southern Kenya, space for wildlife and livestock is limited in private, subdivided land. Here, livestock enclosures are at higher densities and less space exists between them than in community land or conservation areas. (A) Livestock enclosure kernel density calculated using 29,467 georeferenced locations (black dots); (B) boxplots showing different livestock enclosure kernel density values for the four ecosystems and four land tenure types; (C) the cumulative distribution function (CDF) of the distance from a fixed point in space to the nearest (and successively next) livestock enclosure, calculated using the empty space function F of a stationary point process for each land tenure type. Protected areas are excluded because they have few or no settlements within their boundaries. The blue ribbon represents the 96% point-wise intervals of the theoretical distribution of livestock enclosures under a stationary Poisson process of the same estimated intensity. Conservation areas have the shallowest slope, indicating the greatest distances between livestock enclosures, whereas the curve for private subdivided land increases steeply to an asymptote, indicating that little open space is available.
Across southern Kenya, pre-and post-colonial government policies have continually encouraged the privatization and subdivision of unfenced communally-owned land and the sedentarization of pastoralists (Mwangi and Ostrom, 2009). When coupled with a combination of structural forces, including Kenya’s fast-growing economy and population, land commodification has led to wide-scale fencing of privatized land to secure ownership (Rutten, 1992; Mwangi and Ostrom, 2009; Weldemichel and Lein, 2019) (Figure 3). The resulting shrinkage of space for pastoralism, in addition to the expansion of government-protected areas and conservancies, has perversely incentivized fencing on private land to protect individual property rights and resources (including forage for livestock) and to reduce human-wildlife conflict (Said et al., 2016; Løvschal et al., 2018; Weldemichel and Lein, 2019).
Fencing and land conversion alter the mobility of wildlife and livestock, including their access to vital resources such as water and seasonal forage, which are patchily distributed in rangeland ecosystems (Western and Gichohi, 1993; Homewood et al., 2001; Hobbs et al., 2008; Said et al., 2016; Jakes et al., 2018; McInturff et al., 2020). In addition, these changes limit connectivity between wildlife sub-populations thereby preventing wildlife from moving between seasonal ranges, restricting genetic exchange, exacerbating human-wildlife conflict, squeezing wildlife and livestock into smaller areas, altering ecosystem function, and hampering adaptation to climate change (Western and Maitumo, 2004; Newmark, 2008; Western et al., 2015a; Jakes et al., 2018; Osipova et al., 2018; Veldhuis et al., 2019; McInturff et al., 2020). Sedentarization results in a homogeneous year-round spread of livestock enclosures, reducing livestock mobility and increasing local livestock density and grazing pressure, which creates deeper annual and seasonal deficits in grass biomass than would occur from rainfall-induced droughts alone (Groom and Western, 2013; Western et al., 2015b; Western et al., 2021). Locally high stocking rates with low mobility alter the species composition, height, productivity, and nutritional content of forage resources, with ramifications for wildlife and livestock composition and productivity (Augustine et al., 2003; Boone et al., 2005; Western et al., 2009; Groom and Western, 2013; Young et al., 2013). Moreover, the combination of land privatization, sedentarization, and fencing endangers pastoral livelihoods by reducing resilience to drought and diminishing livestock holdings per person (Boone et al., 2005; Bedelian and Ogutu, 2017; Kimiti et al., 2018). These negative trajectories are likely to steepen with continued global climate change (Funk, 2020)
To maintain social-ecological systems with land-sharing between wildlife and local communities, land use planning for the remaining areas, particularly those currently undergoing subdivision, is vital and pressing. This must allow for people to equitably benefit from their land while maintaining the space and mobility required for livestock and wildlife (Western et al., 2020). In Amboseli, a comprehensive ecosystem management plan and a locally developed subdivision plan are indeed guiding the process of subdivision to ensure equitable allocations of land, and the maintenance of important wildlife corridors.2 In the community-owned and fertile Loita ecosystem, where conversion to private title is underway, bottom-up processes are also being incorporated into regional land-use planning. In the longer term, however, the response to these changes must be driven by regional (county) governments in collaboration with local communities, with significant support from the national government, local and international civil society stakeholders, and where relevant, tourism stakeholders (like the lucrative Mara). This support includes policy reforms to curtail land speculation and mitigate the impact of fencing and land subdivision, coupled with economic incentives for local landowners to maintain open rangelands, ensuring the social-ecological sustainability of these ecosystems (Norton-Griffiths and Said, 2009; Lindsey et al., 2020). Without such incentives, the current situation will continue with ad hoc propagation of fences encircling subdivisions of commodified land into small parcels.
The highest costs of wildlife conservation are ultimately borne by local communities (Norton-Griffiths and Said, 2009; Weldemichel and Lein, 2019). Solutions to offset these costs could include recognition by national and regional governments of the culturally important practice of pastoralism and local livelihoods, and the cultural and economic value of protecting it to avoid a poverty trap for a significant segment of the human population. Increased efforts are needed to develop payments for ecosystem services, to restructure and expand a potentially lucrative post-COVID eco-tourism industry, and to ensure an equitable distribution of benefits to local communities (Lindsey et al., 2020; Western et al., 2020). In some areas, there is evidence that removal of fencing and prevention of sedentarization can be incentivized (Western et al., 2020), but reducing the direct and indirect costs of co-existing with wildlife is also critical (Norton-Griffiths and Said, 2009).
Immediate investment could be required to support leases and purchases by conservation organizations of ecologically critical areas under imminent threat of fragmentation and conversion. There would also have to be co-created, long-term solutions for the people living in and off such areas. However, the level of investment needed is ominously high (Norton-Griffiths and Said, 2009). For example, in the private land around the Maasai Mara Reserve, over US$6,000,000 is already invested in conservation areas through land leases to offset the opportunity costs of conservation, although in some areas this still might not be enough to ensure equitable and sustainable livelihood options for the local community (Bedelian and Ogutu, 2017). Any support garnered would need to be underpinned by strong and accountable local leadership and governance, along with devolved benefits and decision-making rights to local people, in order to ensure its ultimate sustainability (Western et al., 2020). Kenya does have proactive policies for devolving the benefits and management rights of natural resources, including wildlife (Western et al., 2015b), but more is needed to ensure sustainability, fairness, and effectiveness at the landscape scale (Weldemichel and Lein, 2019). For the remaining areas not yet subdivided, there is a need to maintain and empower community tenure and governance systems, including adopting the recent Community Land Act, allowing for an alternative to uncontrolled and unequitable subdivision.
The open-access landDX database (Tyrrell et al., 2022) and our preliminary analysis of it across >30,000 km2 of southern Kenya provide a unique, urgent, and vital snapshot of the social-ecological changes occurring across global rangelands. Similar changes are occurring at different rates across eastern and southern Africa, North America, Australia, South America, and central Asia, yet few of these changes have been documented at the spatial scale we have covered (Reid et al., 2014; McInturff et al., 2020). As we enter the UN Decade on Ecosystem Restoration, global efforts must be strengthened to conserve and sustainably use rangelands. This requires swift and thoughtful action by governments, and appropriate support to local communities, to avoid irreversible losses of habitat, wildlife, ecosystem services, and the cultural heritages of pastoralist societies.
Publicly available datasets were analyzed in this study. This data can be found here: https://www.nature.com/articles/s41597-021-01100-9.
PT contributed to the conceptualization of the article, collected the data, analyzed the data, and wrote the first draft of the manuscripts. All authors contributed to the article writing and approved the submitted version.
PT was funded by the Ralph Mistler Scholarship at WildCRU. JCS considers this work a contribution to his VILLUM Investigator project “Biodiversity Dynamics in a Changing World”, funded by VILLUM FONDEN (grant 16549), and Center for Ecological Dynamics in a Novel Biosphere (ECONOVO), funded by Danish National Research Foundation. PT and PB received support from the Maasai Landscape Conservation Fund to complete this work.
We thank Guy Western, Steiner Sompeta, Femke Broekhuis, and David Western for discussions on this topic.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Augustine D. J., Mcnaughton S. J., Frank D. A. (2003). Feedback between soil nutrients and large herbivores in a managed savanna ecosystem. Ecol. Appl. 13, 1325–1337. doi: 10.1890/02-5283
Baddeley A., Rubak E., Turner R. (2015). Spatial point patterns: methodology and applications with r (CRC press).
Baston D. (2022). “Exactextractr: Fast extraction from raster datasets using polygons,” in R package version 0.8.2. Available at: https://CRAN.R-project.org/package=exactextractr.
Bedelian C., Ogutu J. O. (2017). Trade-offs for climate-resilient pastoral livelihoods in wildlife conservancies in the Mara ecosystem, Kenya. Pastoralism 7, 1–22. doi: 10.1186/s13570-017-0085-1
Boone R. B., BurnSilver S. B., Thornton P. K., Worden J. S., Galvin K. A. (2005). Quantifying declines in livestock due to land subdivision. Rangel. Ecol. Manage. 58, 523–532. doi: 10.2111/1551-5028(2005)58[523:QDILDT]2.0.CO;2
Brehony P. (2020). Conservation and eesilience to drought: A social-ecological perspective on conservation areas in kenya’s southern maasailand (Cambridge, UK: University of Cambridge).
Briske D. (2017). Rangeland systems: Processes, management and challenges (Springer Open). eBook. doi: 10.1007/978-3-319-46709-2
Broekhuis F., O’Meara R. H., O’Meara S., Barton M., Harrell C., Western G., et al. (2018). An assessment of mammals in naimina enkiyio forest, Kenya. Afr. J. Ecol. 56, 755–758. doi: 10.1111/aje.12532
Funk C. (2020). Ethiopia, Somalia And Kenya face devastating drought. Nature 586, 645. doi: 10.1038/d41586-020-02698-3
Galaty J. G. (1999). Grounding pastoralists: law, politics, and dispossession in East Africa. Nomadic. Peoples. 3, 56–73. doi: 10.3167/082279499782409334
Groom R. J., Western D. (2013). Impact of land subdivision and sedentarization on wildlife in kenya’s southern rangelands. Rangel. Ecol. Manage. 66, 1–9. doi: 10.2111/Rem-D-11-00021.1
Hijmans R. (2022). “Raster: Geographic data analysis and modeling,” in R package version 3.5-15. Available at: https://CRAN.R-project.org/package=raster.
Hobbs N. T., Galvin K. A., Stokes C. J., Lackett J. M., Ash A. J., Boone R. B., et al. (2008). Fragmentation of rangelands: Implications for humans, animals, and landscapes. Glob. Environ. Change 18, 776–785. doi: 10.1016/j.gloenvcha.2008.07.011
Homewood K., Lambin E. F., Coast E., Kariuki A., Kikula I., Kivelia J., et al. (2001). Long-term changes in Serengeti-Mara wildebeest and land cover: Pastoralism, population, or policies? Proc. Natl. Acad. Sci. 98, 12544–12549. doi: 10.1073/pnas.221053998
Jakes A. F., Jones P. F., Paige L. C., Seidler R. G., Huijser M. P. (2018). A fence runs through it: A call for greater attention to the influence of fences on wildlife and ecosystems. Biol. Conserv. 227, 310–318. doi: 10.1016/j.biocon.2018.09.026
Kimiti K. S., Western D., Mbau J. S., Wasonga O. V. (2018). Impacts of long-term land-use changes on herd size and mobility among pastoral households in amboseli ecosystem, Kenya. Ecol. Process. 7, 4. doi: 10.1186/s13717-018-0115-y
Løvschal M., Bøcher P. K., Pilgaard J., Amoke I., Odingo A., Thuo A., et al. (2017). Fencing bodes a rapid collapse of the unique greater Mara ecosystem. Sci. Rep. 7, 1–7. doi: 10.1038/srep41450
Løvschal M., Håkonsson D. D., Amoke I. (2018). Are goats the new elephants in the room? changing land-use strategies in greater Mara, Kenya. Land. Use Policy 1–7. doi: 10.1016/j.landusepol.2018.04.029
Lindsey P., Allan J., Brehony P., Dickman A., Robson A., Begg C., et al. (2020). Conserving africa’s wildlife and wildlands through the COVID-19 crisis and beyond. Nat. Ecol. Evol., 4, 1–11. doi: 10.1038/s41559-020-1275-6
McInturff A., Xu W., Wilkinson C. E., Dejid N., Brashares J. S. (2020). Fence ecology: frameworks for understanding the ecological effects of fences’. BioScience 70, 971–985. doi: 10.1093/biosci/biaa103
Mwangi E. (2007). Socioeconomic change and landuse in Africa: the transformation of property rights in kenya’s maasailand (New York: Palgrave MacMillan).
Mwangi E., Ostrom E. (2009). Top-down solutions: looking up from East africa’s rangelands. Environment 9157, 37–41. doi: 10.3200/ENVT.51.1.34-45
Newmark W. D. (2008). Isolation of African protected areas. Front. Ecol. Environ. 6, 321–328. doi: 10.1890/070003
Nkedianye D. K., Ogutu J. O., Said M. Y., Kifugo S. C., de Leeuw J., Van Gardingen P., et al. (2020). Comparative social demography, livelihood diversification and land tenure among the maasai of Kenya and Tanzania. Pastoralism 10. doi: 10.1186/s13570-020-00165-2
Norton-Griffiths M., Said M. Y. (2009). ““The future for wildlife on kenya’s rangelands: an economic perspective,”,” in Wild rangelands (Oxford UK: Wiley-Blackwell), 367–392. doi: 10.1002/9781444317091.ch14
Ojwang’ G. O., Wargute P. W., Said M. Y., Worden J. S., Davidson Z., Muruthi P., et al. (2017) Wildlife migratory corridors and dispersal areas: Kenya rangelands and coastal terrestrial ecosystems. Available at: http://www.kws.go.ke/content/launch-report-wildlife-corridors-and-dispersal-areas.
Osipova L., Okello M. M., Njumbi S. J., Ngene S., Western D., Hayward M. W., et al. (2018). Using step-selection functions to model landscape connectivity for African elephants: accounting for variability across individuals and seasons. Anim. Conserv. 22, 35–48. doi: 10.1111/acv.12432
Pebesma E. (2018). Simple features for r: Standardized support for spatial vector data. R. J. 10 (1), 439–446. doi: 10.32614/RJ-2018-009
R Core Team (2018) R: A language and environment for statistical computing. Available at: https://www.r-project.org/.
Reid R. S., Fernández-Giménez M. E., Galvin K. A. (2014). Dynamics and resilience of rangelands and pastoral peoples around the globe. Annu. Rev. Environ. Resour. 39, 217–242. doi: 10.1146/annurev-environ-020713-163329
Rutten M. M. E. M. (1992). Selling wealth to buy poverty: the process of the individualization of landownership among the maasai pastoralists of kajiado district, kenya, 1890--1990 (Saarbrücken: Breitenbach).
Said M. Y., Ogutu J. O., Kifugo S. C., Makui O., Reid R. S., de Leeuw J. (2016). Effects of extreme land fragmentation on wildlife and livestock population abundance and distribution. J. Nat. Conserv. 34, 151–164. doi: 10.1016/j.jnc.2016.10.005
Stabach J. A. (2015). Movement, resource selection, and the physiological stress response of white-bearded wildebeest (Fort Collins, Colorado: Colorado State University). doctoral disertation.
Stabach J. A., Hughey L. F., Reid R. S., Worden J. S., Leimgruber P., Boone R. B. (2020). Data from: Comparison of movement strategies of three populations of white-bearded wildebeest. Movebank. Data Repository. doi: 10.5441/001/1.h0t27719
Stelfox J. G., Peden D. G. (1981). The aerial survey programme of the Kenya rangeland ecological monitoring unit: 1976-79.
Tyrrell P., Amoke I., Betjes K., Broekhuis F., Buitenwerf R., Carroll S., et al. (2022). Landscape dynamics (landDX) an open-access spatial-temporal database for the Kenya-Tanzania borderlands. Sci. Data 9, 1–9. doi: 10.1038/s41597-021-01100-9
Tyrrell P., Naidoo R., Macdonald D. W., du Toit J. T. (2021). New forces influencing savanna conservation: increasing land prices driven by gentrification and speculation at the landscape scale. Front. Ecol. Environ. 19, 494–500. doi: 10.1002/fee.2391
Tyrrell P., Russell S., Western D. (2017). Seasonal movements of wildlife and livestock in a heterogenous pastoral landscape: Implications for coexistence and community based conservation. Glob. Ecol. Conserv. 12, 59–72. doi: 10.1016/j.gecco.2017.08.006
Vaughan D., Dancho M. (2020). “Furrr: Apply mapping functions in parallel using futures,” in R package version 0.2.1. Available at: https://CRAN.R-project.org/package=furrr.
Veldhuis M. P., Ritchie M. E., Ogutu J. O., Morrison T. A., Beale C. M., Estes A. B., et al. (2019). Cross-boundary human impacts compromise the Serengeti-Mara ecosystem. Science. 1428, 363. doi: 10.1126/science.aav0564
Weldemichel T. G., Lein H. (2019). “Fencing is our last stronghold before we lose it all.” a political ecology of fencing around the maasai Mara national reserve, Kenya. Land. Use Policy 87, 104075. doi: 10.1016/j.landusepol.2019.104075
Western D., Gichohi H. (1993). Segregation effects and the impoverishment of savanna parks: the case for ecosystem viability analysis. Afr. J. Ecol. 31, 269–281. doi: 10.1111/j.1365-2028.1993.tb00541.x
Western D., Groom R., Worden J. (2009). The impact of subdivision and sedentarization of pastoral lands on wildlife in an African savanna ecosystem. Biol. Conserv. 142, 2538–2546. doi: 10.1016/j.biocon.2009.05.025
Western D., Maitumo D. (2004). Woodland loss and restoration in a savanna park. Afr. J. Ecol. 42, 111–121. doi: 10.1111/j.1365-2028.2004.00506.x
Western D., Mose V. N., Maitumo D., Mburu C. (2021). Long-term changes in the plant ecology of an African savanna landscape and the implications for ecosystem theory and conservation management. Eco. Process. 10, 15. doi: 10.1186/s13717-021-00286-5
Western D., Mose V., Worden J., Maitumo D. (2015a). Predicting extreme droughts in savannah Africa: a comparison of proxy and direct measures in detecting biomass fluctuations, trends and their causes. PloS One 10, e0136516. doi: 10.1371/journal.pone.0136516
Western D., Tyrrell P., Brehony P., Russell S., Western G., Kamanga J. (2020). Conservation from the inside-out: Winning space and a place for wildlife in working landscapes. People Nat., 2, 1–13. doi: 10.1002/pan3.10077
Western D., Waithaka J., Kamanga J. (2015b). Finding space for wildlife beyond national parks and reducing conflict through community-based conservation : the Kenya experience. Parks. 21, 51–62. doi: 10.2305/IUCN.CH.2014.PARKS-21-1DW.en
Wickham H. (2019). Welcome to the tidyverse. J. Open Source Software. 4 (43), 1686. doi: 10.21105/joss.01686
Keywords: rangelands, fences (barriers), Kenya, subdivision, pastoralism, wildlife, community based conservation
Citation: Tyrrell P, Buitenwerf R, Brehony P, Løvschal M, Wall J, Russell S, Svenning J-C, Macdonald DW, du Toit JT and Kamanga J (2022) Wide-scale subdivision and fencing of southern Kenyan rangelands jeopardizes biodiversity conservation and pastoral livelihoods: Demonstration of utility of open-access landDX database. Front. Conserv. Sci. 3:889501. doi: 10.3389/fcosc.2022.889501
Received: 04 March 2022; Accepted: 22 September 2022;
Published: 07 October 2022.
Edited by:
Paul Jones, Alberta Conservation Association, CanadaReviewed by:
Helen Uchenna Agu, University of Nigeria, NigeriaCopyright © 2022 Tyrrell, Buitenwerf, Brehony, Løvschal, Wall, Russell, Svenning, Macdonald, du Toit and Kamanga. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Peter Tyrrell, cGV0ZXJkYXZpZHR5cnJlbGxAZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.