 Tempe S. F. Adams
Tempe S. F. Adams Keith E.A. Leggett2
Keith E.A. Leggett2 Marlee A. Tucker
Marlee A. Tucker
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Conserv. Sci., 28 July 2022
Sec. Human-Wildlife Interactions
Volume 3 - 2022 | https://doi.org/10.3389/fcosc.2022.872472
The global impact of increased human activities has consequences on the conservation of wildlife. Understanding how wildlife adapts to increased human pressures with urban expansion and agricultural areas is fundamental to future conservation plans of any species. However, there is a belief that large wild free-ranging carnivores and ungulates, cannot coexist with people, limited studies have looked at wildlife movements through differing human-dominated landscapes at finer spatial scales, in Africa. This information is vital as the human population is only going to increase and the wildlife protected areas decrease. We used remote-sensor camera traps to identify the movement patterns of African elephant (Loxodonta africana) through six wildlife corridors in Botswana. The wildlife corridors were located in two different human-dominated landscapes (agricultural/urban), with varying degrees of human impact. While we found that elephants use corridors in both landscapes, they use the urban corridors both diurnally and nocturnally in contrast to agricultural corridors which were only nocturnal. Our results provide evidence for temporal partitioning of corridor use by elephants. We identified that seasonality and landscape were important factors in determining the presence of elephants in the corridors. Our findings demonstrate that elephant diel patterns of use of the wildlife corridor differs based on the surrounding human land-uses on an hourly basis and daily basis, revealing potential adaptation and risk avoidance behaviour.
Understanding the way different human pressures, whether that be resource extraction, habitat depletion, urbanization or agricultural expansion, impact the natural world is of fundamental importance if we are going to survive as a species (Jones et al., 2018; Gaynor et al., 2018a; Allen et al., 2020; Ghoddousi et al., 2021). Habitat fragmentation and depletion of protected wildlife areas is confining wildlife into smaller restricted areas, which must adapt to survive (Carter et al., 2012; Rodriguez-Soto et al., 2013; Bortolamiol et al., 2016; Shamoon et al., 2018). Human pressures are defined as actions taken by humans with the potential to harm nature (Martins et al., 2012; Venter et al., 2016). In order to create effective conservation management, it is vital for us to understand the impact that these pressures are having on species, and how they are potentially adapting to those pressures, if at all (Bortolamiol et al., 2016; Venter et al., 2016; Allen et al., 2020; Tiller et al., 2021; Ghoddousi et al., 2021). Despite this, there is very limited wildlife research (<2%) published on urban wildlife research globally, and the studies that are being published are primarily conducted in North America, Europe or Australia, the areas with the lowest rates of urbanization (Magle et al., 2012). It is apparent there is a need for urban wildlife research to be produced in the areas with the rapid rates of urban development throughout Africa, central- south Americas and Asia, which is also where the world’s highest rates of biodiversity exist (Balmford et al., 2001).
Changing land uses and the expansion of both agricultural and urban landscapes, are some of the most impactful human pressures on wildlife (Venter et al., 2016). This changing pressure is constantly impacting the functioning of the natural environment, often resulting in species vacating areas of high pressure by simply avoiding the area altogether, or they will adapt to the pressure by avoiding areas of high rates of human-wildlife interaction at times when humans are most active (Carter et al., 2012; Gaynor et al., 2018a; Gaynor et al., 2018b; Shamoon et al., 2018). There has been limited research that has looked at how differing human pressures and changing land-uses are impacting the utilization of wildlife corridors (Carter et al., 2012; Adams et al., 2017). Wildlife corridors are often used as key conservation tools as they allow movement between habitat patches. These connectivity paths can facilitate human-wildlife coexistence as they provide wildlife safe access to required resources (Caro et al., 2009), by reducing interactions between people and wildlife. With the increased uptake and use of wildlife corridors all over the world, begs the question how much surrounding human pressure can wildlife handle to continue using the corridors and how do we factor this into our future wildlife management plans.
The African elephant (Loxodonta africana) is a large, charismatic, umbrella species that exhibit cathemeral diel activity, but are also strongly influenced by human activities (Roever et al., 2013; Kansky and Knight, 2014; Songhurst et al., 2016; Adams et al., 2017; Gaynor et al., 2018a; Clauss et al., 2021). These qualities make them an ideal model species to monitor the impact of differing human-dominated landscapes on their movements. In addition, understanding the impacts of human pressure on elephants is of high importance, as approximately 60% of the elephant range lies outside formally protected areas (Wall et al., 2021).
Elephant movements need to be considered in future land-use development plans, if they are not, then humans and elephants will be forced into closer contact with one another, which will result in increased negative interactions. This will result in damage to property, competition over natural resources, space, and at its worst, loss of human or elephant lives (Osborn and Parker, 2003; Kansky and Knight, 2014; Hoare, 2015; Mumby and Plotnik, 2018). These negative interactions are largely becoming an increasingly controversial political issue (Hoare, 2003), as elephants represent an economic value for tourism enterprises, however living within the elephant range especially when you feel as though you are not personally benefitting from the elephant population can be a burden, and resentment can grow leading to increased public pressure on Government to develop sustainable, effective long term solutions to reduce the conflict (DeMotts and Hoon, 2012; Gupta, 2013).
Elephant-movement behaviour is influenced by environmental factors such as water and foraging availability (Boettiger et al., 2011), and rainfall (Thouless, 1996). Additionally, elephants are increasingly affected by humans and their development into previously protected wildlife areas (Buij et al., 2007; Blake et al., 2008; Roever et al., 2013). We have a limited understanding of how human disturbance and pressure influences elephants’ fine-scale movement behaviour (Graham et al., 2009; Roever et al., 2013; Songhurst et al., 2016; Adams et al., 2017), not only in agricultural landscapes, but also in increasingly urbanized areas.
According to Wittemyer et al. (2008) elephants entering communal areas have developed certain behavioural strategies to reduce human interaction, one being that elephants tend to enter agricultural and urban settlements at night to reduce their chances of coming into contact with people (Jackson et al., 2008; Graham et al., 2009; Roever et al., 2013; Songhurst et al., 2016; Adams et al., 2017). Elephants have been shown to actively avoid areas of human development, showing that their movements in human developed areas were directly impacted by human urban infrastructure (Buij et al., 2007). Similarly, other studies have found elephant movements were impacted by the construction of roads that acted as a barrier to elephants (Blom et al., 2004; Blake et al., 2008). Previous research gives evidence clearly showing that elephants avoid human development by altering their movement patterns, as a result these studies have stressed the need for relevant government bodies to conserve large uninterrupted protected areas to maintain wildlife populations (Blom et al., 2004). However, realistically with human pressure predicted to increase alternative wildlife management strategies such as the implementation and designation of wildlife corridors are required to conserve wide ranging species long term.
To date, we have limited knowledge in comparing how elephants use wildlife corridors and resources in areas with different levels of human pressure. There is no evidence if they have varying adaptive strategies based on the corridors surrounding human pressure, this is key information for implementing successful corridors and minimising conflict between humans and wildlife. Based on the previous literature, we would predict elephants would show greater avoidance where human pressure is higher in urban landscapes by reducing their use of the corridors compared to that of low human pressure in agricultural landscapes, where physical development is potentially less.
Botswana has the largest elephant population in Africa with approximately 130,000 individuals (Chase et al., 2019), together with one of the lowest human populations of all African countries (~2.3 million) (World Bank Database, 2019). Botswanan wildlife management authorities are concerned about the effect that the large elephant populations will have on expanding urban and agricultural development, and they predict increases in human-elephant conflict (HEC) over space and resources (Government of Botswana, 2021). This concern highlights the need for an improved understanding of elephants’ temporal movement behaviour and their potential level of adaptation to increased and differing human development. Elephants use of wildlife corridors and informed land-use management practices are required to facilitate human-elephant coexistence (Adams et al., 2017).
Our objective was to determine the effect of different levels of human footprint, a proxy of human activities (Venter et al., 2016) will impact how elephants use established wildlife corridors. We used a series of remote-sensor camera traps, to investigate elephant temporal corridor use moving through an agricultural landscape compared to an urban human-dominated landscape for over two years. We predicted that elephants would use the corridors differently between the two landscapes in order to minimize their interactions with people.
The study was conducted in two different human-dominated landscapes: the townships of Kasane and Kazungula, and the farming villages of the Chobe Enclave both located in the Chobe District (22,560 km2) (Figure 1). The Chobe District is in the north-eastern corner of Botswana and is bordered to the north by Namibia, and Zambia, and to the east by Zimbabwe (Garekae et al., 2019). A total of the 70% of the district is declared protected wildlife areas: Chobe National park 52% and forest reserves 18% (Garekae et al., 2017; Adams et al., 2020). There are four recognised seasons within northern Botswana: cold dry season (May-July), hot dry season (August-October), wet season (November-March) and post-wet season (April) (Adams et al., 2017). There are approximately 32,700 elephants in Chobe District, with an estimated 15,000 of them in Chobe National Park (Chase et al., 2019). Chobe district is an unfenced area, where wildlife can move freely throughout the different land-use designations. The elephants in the Chobe District also move across into Zimbabwe, Zambia and Namibia.
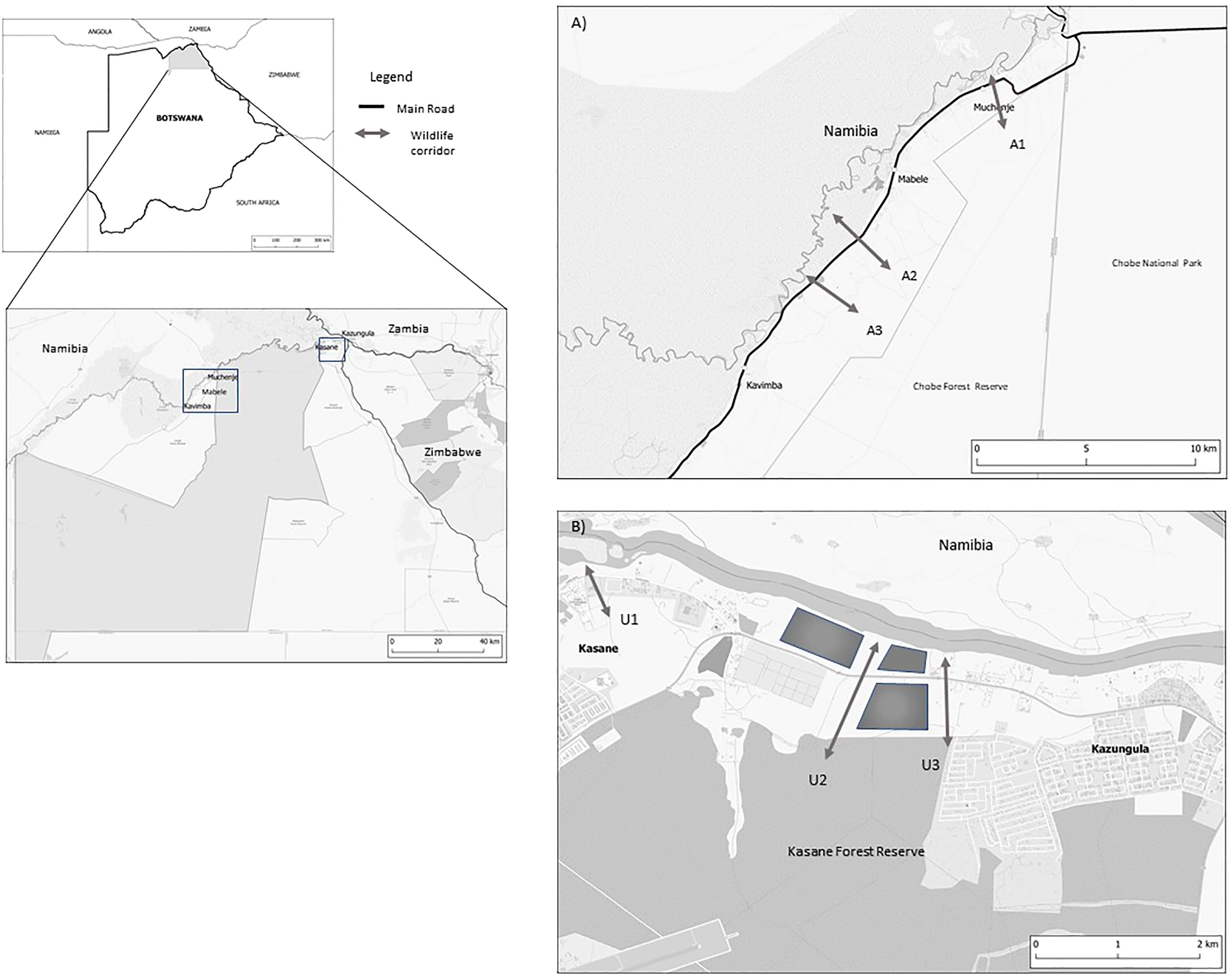
Figure 1 Maps showing the location of Chobe District within Botswana; (A) Map shows the locations of the Agricultural wildlife corridors in the Chobe Enclave; and (B) shows the location of the urban wildlife corridors in the urban centres of Kasane and Kazungula.
Kasane and Kazungula (KK) are the urban centre of the district and have a combined human population of 17,655. However the district is experiencing a rapid human population increase with Kazungula doubling in size from 2011-2022 (Census Office, 2022). The two towns are approximately 10 km apart from one another on the banks of the Chobe River. Kasane township is the district administrative centre, housing government offices, retail and industrial shopping centres and tourism lodges, hotels and guest houses (Garekae et al., 2017). KK are classified as urban centre (Adams et al., 2017) and estimated Human Footprint Index (HFI) of 12-18.9 which is equal to “very high pressure” (Venter et al., 2016).
The Chobe Enclave (Enclave) is a community trust managed concession which is comprised of five low density villages located along the Chobe River floodplain. It is a seasonal floodplain dominated by small-scale farming (livestock and crops), with cultivation occurring in the wet season, followed by harvest in the post-wet season (Jackson et al., 2008; Gupta, 2013). Given the level of cultivation, proximity to the floodplain, and its location alongside “protected areas”, the Enclave is an area of high human-elephant conflict (HEC) (Adams et al., 2020). The area has one of the highest dry season densities in the country with 2.94 elephants per km2. The Enclave is multi-ethnic however the BaSubiya constitute the majority of the inhabitants of the area. The main sources of income in the area is crop and livestock production, tourism lodges and wage subsidies (Garekae et al., 2017). As the Enclave is largely an agricultural area has a calculated HFI of 6.5-7.6, which equates to a “high pressure” human footprint index, however less than the KK index.
Unsurprisingly, the Chobe district is a human-elephant conflict government declared hotspot with elephants being responsible for 48% of problem animal control reports made to the department of Wildlife and National Park (Department Of Wildlife And National Parks Of Botswana, 2018). Elephants travel from the protected areas Chobe Forest Reserve and Chobe National Park through the Enclave and KK to access water on the floodplain and in the Chobe River and to browse on Vachellia ssp (Adams et al., 2017; Adams et al., 2020).
We monitored six designated (demarcated in land-use maps of the area) and undesignated (not demarcated in land-use maps of the area) wildlife corridors: three urban corridors (U) within the KK landscape and three agricultural corridors (A) within the Enclave (Ministry of Lands, Housing and Environment, 2000). All the corridors have similar geographical attributes, with elephants travelling from the Kasane and Chobe Forest Reserve through the corridors to access the Chobe Floodplain/river. They also all have a sealed road passing across each of them.
The urban corridors are located within the townships of Kasane and Kazungula. The U1 corridor (length c.750m, minimum width of c. 3 m) is designated as in an open space recreation area. It runs from outside Kasane to the Seboba Recreational park on the Chobe River, and is in proximity of fishing, religious ceremonies, commercial businesses and a bar. Both corridor U2 (length c. 1,700m, minimum width c. 250) and U3 (length c. 1,700m, minimum width 250 m) are located between Kasane and Kazungula. The corridors run alongside the two largest commercial farms in the area. The end point of the corridors is natural hot spring that flows out to the Chobe River, which is a popular fishing and recreational site (Adams et al., 2017). The urban corridors were monitored between 1 November 2012- 30 April 2014.
The agricultural corridors in the Enclave were located between the villages of Muchenje, Mabele and Kavimba. A1 (length c. 1,200m, minimum width c. 30m) is located on the edge of village of Muchenje and five photographic safari lodges, it runs from the top of a ridge from the Chobe Forest Reserve down through the community area ending out on the Chobe River Floodplain. Corridors A2 (length c. 1185, minimum width c. 30m) and A3 (length c. 1000 m, minimum width c. 40m), are located between the villages of Mabele and Kavimba, they also run from Chobe Forest Reserve and open out on the Chobe River Floodplain, are surrounded by crop fields and cattle Kraals. The agricultural corridors were monitored between during 11 May 2016- 10 August 2019.
Camera trap surveys are a popular technology which is increasingly used as a non-invasive tool to determine wildlife presence in high pressure human environments (Adams et al., 2017; Allen et al., 2020). We monitored the elephants’ presence in the corridors by installing one Bushnell Trophy Cam Brown HD (119437C) camera traps (Bushnell Corporation, Overland Park, USA) in each corridor (Carter et al., 2012; LaPoint et al., 2013; Adams et al., 2017). Cameras were attached to trees or wooden poles, c. 1.0-1.8 m above the ground, facing towards the most frequented used pathway that is used by elephants. Images were downloaded on a weekly basis. The cameras were triggered by motion and were set to continuously over for 24 hours with a 3 second interval between captures. For each photograph, we recorded the time, date and number of species in the image. Both elephant family units and bulls were recorded.
For all analyses, we classified a detection event as a photo capture. As our focus was on corridor use by elephants and how human activities influence this, we only included photos where an elephant or a human was present. Photos within 5 minutes of each other were considered one detection to avoid pseudo-replication. As activity patterns were similar for the three urban, and the three agricultural corridors (Figure S1A, B), we combined the detections across all corridors for each area to examine elephant and human activity patterns.
The timestamps of the independent photo captures were used to examine the 24-hour activity distributions. We used kernel density estimation to estimate the activity patterns and to quantify the overlap in elephant activity patterns between the urban and agricultural corridors. We used the ‘overlap’ package (Ridout and Linkie, 2009) in R v3.6.0 (R Core Team, 2019), which fit the data to a circular kernel density and estimates the activity level at each time period of the 24-hour cycle. We then compared the activity patterns between the urban and agricultural areas to get a coefficient of overlap, where 0 represents no overlap and 1 represents complete overlap. Bootstrapping 10 000 activity estimates, we estimated the 95% confidence intervals for the coefficient of overlap (Ridout and Linkie, 2009). The detections of elephants and humans in both the agricultural and urban corridors were larger than 75, so we used the Dhat4 overlap estimated according to Ridout and Linkie (2009). As we also had data on humans in all the corridors, we ran the daily activity and overlap analyses again, but this time comparing activity patterns between elephants and humans. We did this for the urban and the agricultural corridors separately.
Using the independent photo capture data, we recorded a single count of presence (1) or absence (0) for each day in each corridor. We used binomial linear mixed effects models to examine the presence or absence of elephants across the urban versus agricultural corridors. We included area (i.e. agriculture or urban) and season as predictor variables. We also included an interaction term between area and season to test for differences in elephant presence across the seasons between the agricultural and urban corridors. Corridor ID and year were included as random effects. Significance was determined for all analyses when p<0.05. Binomial models were run using the ‘lmerTest’ package (Kuznetsova et al., 2017) and model predictions were estimated using the ‘effects’ package (Fox & Wiesberg, 2018; 2019).
We recorded a total of 26,023 images of elephants and humans across the six corridors. A total of 5,956 images were captured in the urban corridors and a total of 20,067 images were captured in the agricultural corridors. For the final diel activity analyses (i.e., after selecting images > 5 minutes apart), we had 661 elephant detections and 550 human images in the urban corridors, and 3772 elephant detections and 528 human images in the agricultural corridors. For the daily corridor use analyses, we had a total of 2976 daily elephant presence values (agriculture n = 1987 and urban n = 989). There was a significant difference in elephant presence in the corridors between the different land-uses at different timescales (Table 1; Figure 4A).
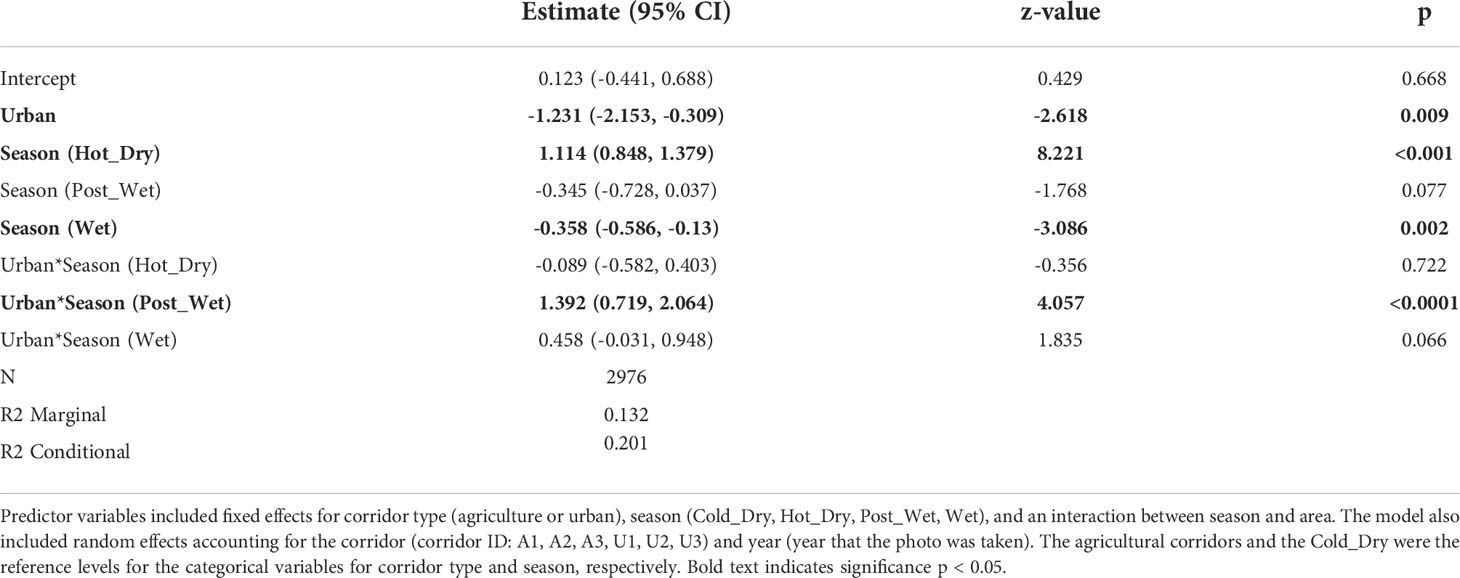
Table 1 Model coefficients, standard errors (SE), p values, sample sizes and variance explained (marginal and conditional R2) of linear mixed effects models predicting mean daily elephant detections (elephant detections divided by sampling days).
Diel activity of elephants differed between the urban and agricultural corridors, with a 67.8% (CI 64.0 – 70.2) overlap in the hourly temporal activity of elephants between the two land-use types (Figure 2). Elephants in the agricultural corridors were more nocturnal, with peaks in activity after 18:00 and before 06:00. In the urban corridors, elephants were active during the entire day. The differences in activity patterns between the two areas may be related to humans (Figure 3). Figure This was particularly strong for the agricultural corridors with only a 9.1% overlap in activity between humans and elephants (Figure 3A). In the urban corridors, the activity patterns overlapped 26.8%, indicating a reduction in the avoidance of humans (Figure 3B).
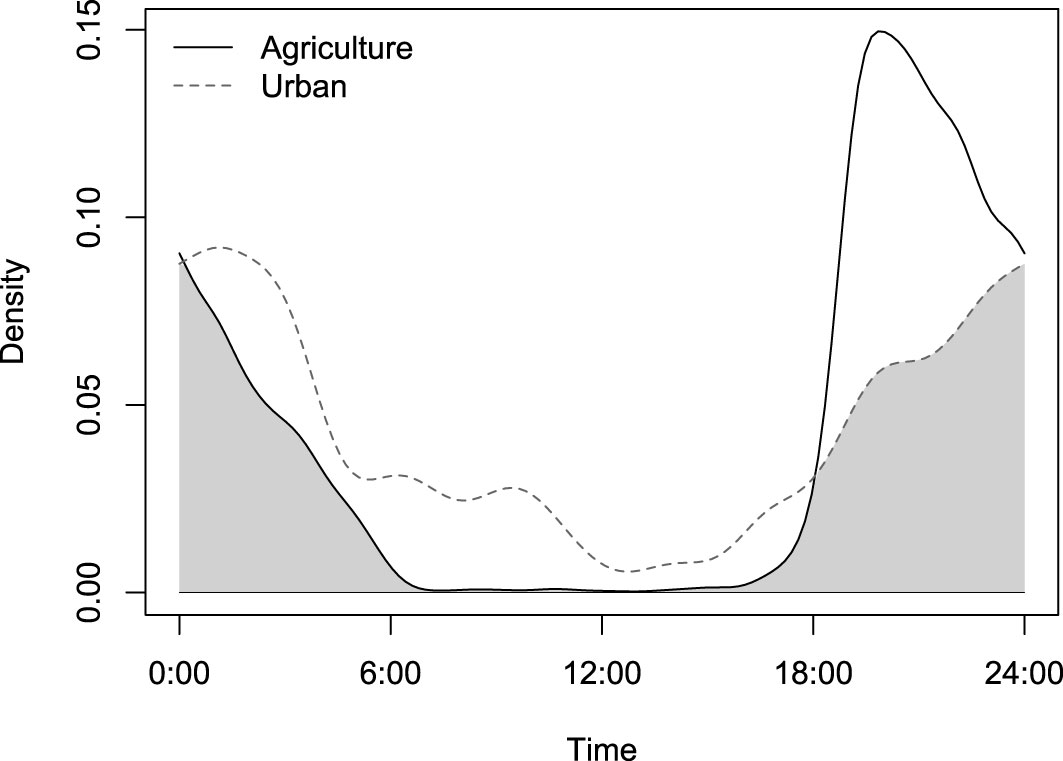
Figure 2 Temporal activity and overlap (kernel density) between elephants in agricultural (black solid line) and urban areas (grey broken line). The curves overlap 67.8% (CI 64.0 – 70.2; grey shaded area).
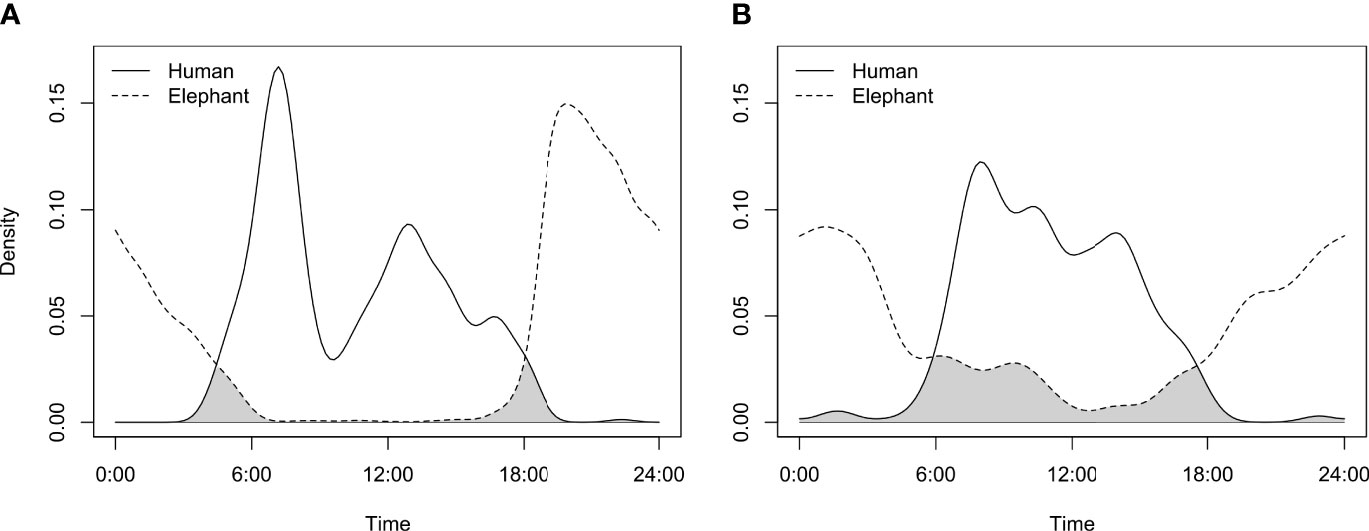
Figure 3 Temporal activity and overlap (kernel density) between elephants (grey broken line) and humans (black line) in (A) agricultural (9.1% overlap CI 5.4 – 8.5) and (B) urban areas (26.8% overlap CI 21.1 – 27.9). The grey shaded areas represent the overlap between the two distributions.
The daily presence of elephants in the corridors differed between the urban and agricultural landscapes, with a higher probability of presence in the agricultural corridors (56%) compared with urban corridors (32%) (Table 1; Figure 4A). Across all corridors, there were seasonal difference in elephant presence (Table 1), with higher probability of presence during the hot-dry season (69%) and the lowest probability during the wet season (38%). There were also seasonal changes in elephant presence within the agricultural and urban corridors (Table 1, Figure 3B). The highest probability of elephant presence in the urban corridors was during the post-wet season (48%), and the lowest during the cold-dry and wet seasons (both 44%). In the agricultural corridors, the highest probability of presence was during the hot-dry season (76%) and lowest during the cold-dry season (25%). Overall, there were general trends of higher probabilities in the agricultural corridors across the seasons compared to the urban corridors, except for the post-wet season.
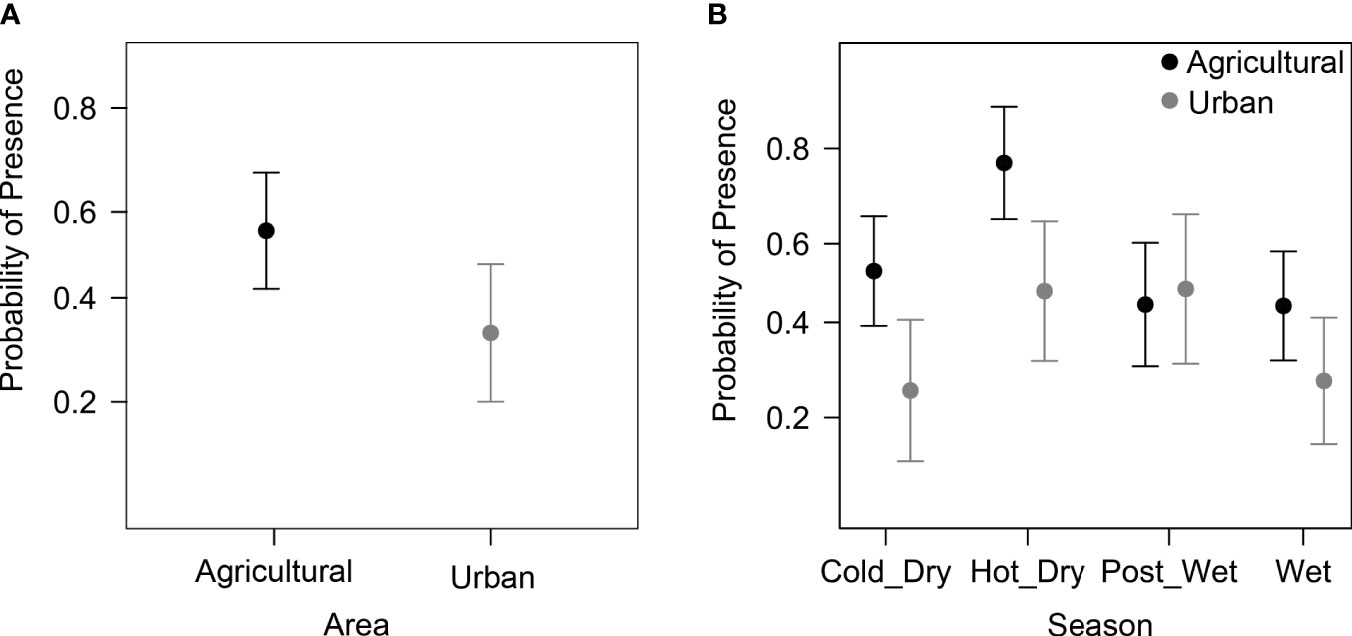
Figure 4 Predicted daily probability of elephant presence detections per day between (A) the agricultural and urban corridors and (B) across seasons and agricultural/urban corridors. These predictions are based on the model in Table 1. Points represent the mean predicted elephant presence and bars represent the 95% confidence intervals.
The results of this study explored how elephant movements are impacted by differing forms of human pressures and development in the Chobe District in northern Botswana. A major finding is that land-use seemingly affects diel use of the corridors by elephants. The results show that elephants in urban and agricultural landscapes have different diel activity patterns of use while moving through the wildlife corridors. Elephants in agricultural areas are utilizing the wildlife corridors predominantly nocturnally, between 1800 and 0600, when humans are less active. Comparatively the elephants tend to use the urban corridors throughout the day, even with overlapping human presence.
We did see overall that the agricultural corridors had a higher presence of elephants compared with the urban corridors. This could be due to the agricultural landscapes being less developed with less infrastructure such as roads and buildings and a lower human pressure measure than that of the urban landscape. This could also be due to the corridors in the agricultural land-use simply have increased open space, so there is physically more space for the elephants to pass through. As the area is made up of undeveloped land open cattle grazing land, fenced crop fields, kraals and low density villages (Figure 1) However, elephants need to balance this with the high risk of interacting with humans while moving through an agricultural landscape. It seems to overcome this, elephants are using the corridors at night to minimise the risk.
The elephant’s nocturnal patterns of use of the agricultural corridors are consistent with other elephants’ crop-raiding behavioural studies (Gunn et al., 2013; Shamoon et al., 2018; Tiller et al., 2021). Elephant crop-raiding behaviour is widely understood to be predominately a nocturnal activity. This is due to elephants avoiding the height of human activity and the risk of increased visibility (Gunn et al., 2013; Tiller et al., 2021), this could be the case for elephant use of the wildlife corridors through the area. However, the seasonal pattern of the elephant corridor use in the agricultural landscape was not consistent with crop maturation, unlike Snyer et al. (2019) study, which showed elephant movements through the agricultural areas in the western Serengeti increased at the time of crops maturing. Crops mature and begin to be harvested in the Chobe enclave between April-June, there appeared to be no increase in elephants use of the corridors at that time (Adams et al., 2020). This would indicate that the elephants do not increase the use of corridors at crop maturing time of year with the intent of raiding crops.
Buchholtz et al. (2021) found that elephants selected water access points in areas with less human development to navigate through agricultural areas, and it was suspected that this was an avoidance strategy to avoid interacting with humans. A study conducted by Gaynor et al. (2018a) revealed a strong effect of human daily patterns on wildlife temporal movement patterns. Overall, animals increased their nocturnality by an average factor of 1.36 in response to human disturbance, where the urban environment had the highest ratio compared to any other human disturbance (Gaynor et al., 2018a). Previous research supports the hypothesis that elephants alter their movement behaviour to reduce the risk of encountering humans by being most active in human-dominated areas when humans are least active (Graham et al., 2009; Gunn et al., 2013). This is consistent with our findings on the elephant movement behaviour in the agricultural corridors, however, this is not the case for the urban corridors. This indicates that not all human activities pose the same risk as one another.
The perceived level of threat of different human activities could be reflected in the elephant’s use of the corridors. Human-wildlife conflict is a hugely complex area of research where the human-human conflicts are often the underlying drivers of the conflict, whether it be between authorities and farmers or different cultural groups (Dickman, 2009; DeMotts and Hoon, 2012). Previous research has established that the response of people to conflict incidents with elephants can reflect humans’ attitudes towards the animals (Dickman, 2009). These attitudes often reflect the level of threat or loss an individual suffers through living in the same space as these animals, such as reduced crop yields or damage to property. To minimise interactions with humans, elephants can avoid areas of high risk, where the presence or absence of elephants in an area changes throughout the day (Jachowski et al., 2013; Gaynor et al., 2018b; Tiller et al., 2021). The concept of understanding risk, as well as reactions to it, is heavily influenced by humans’ social and cultural perceptions, values, and history (Dickman, 2009). Differences in the elephant movements between the urban and agricultural landscapes suggest that the urban environment of Kasane and Kazungula presents a less risky environment to the elephants despite the increased human pressure. Adams et al. (2017) found that the elephants slow down their movements in the urban corridors and spend high quantities of time within the urban environment, further indicating that these urban areas are a low fear-based landscape. Compared to that of the Enclave where there is no overlap of time between humans and elephants. As previously discussed the Enclave is an agricultural conflict hotspot in Botswana, where often farmers resort to shooting at elephants in an effort to deter them from entering their crops (Gupta, 2013), it is for this reason this area could potentially appear riskier to the elephants. In order to better understand the level of perceived risk for people in association with elephants a social survey could be conducted throughout the settlements of KK and the Enclave, determining peoples tolerance, opinions, and perceptions towards elephants and how they behave and react to an elephant when they interact with them. A study of this nature could give further explanation for why elephant movement behaviours differ in the different human land-uses, and whether a difference in people’s attitude could be directly related to the different ways elephants move through the same occupied area.
It appears elephant presence in both types of corridors is also influenced by seasonality and rainfall in the area. Overall, the elephants had a higher presence in the hot dry season rather than the wet, which due to the elephants need to access the Chobe river to drink (Adams et al., 2017). Although the urban corridors saw a peak in activity in the post-wet season, we are still unsure of why this is occurring, however it is thought to be associated with resources such as fruiting marula trees in the area.
African countries must balance urban and agricultural development goals with the maintenance of wildlife conservation areas (Buij et al., 2007; Carter et al., 2012), if they want to have any biodiversity and ecosystems. Designation and maintenance of wildlife corridors can be the fundamental wildlife management tool to allow both development and conservation to continue. Our study highlights the functionality of both urban and agricultural corridors in facilitating elephant movements through differing human pressured environments. Wildlife corridors are crucial for effective conservation in allowing connectivity through different human-dominated landscapes (Kiffner et al., 2016), however, corridors are being blocked with increased changes in surrounding land-uses. Increasing our understanding of how different land-uses and human pressures impact wildlife use of an area is fundamental to the future implementation of them as a form of wildlife management (Ghoddousi et al., 2021). Our study highlights the need to consider wildlife diel activity patterns in differing human-dominated landscapes for conservation and environmental management planning (Shamoon et al., 2018). This research gives further evidence highlighting the need for transparent documenting of human pressure within and around protected wildlife areas is now more critical than ever, to assist in the conservation of species.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethical review and approval was not required for the animal study because The ethics approval to conduct this study in Botswana was granted under the research permit permit EWT 8/34/4 XX (34), granted by the Department of Wildlife and National Parks to Elephants Without Borders. No further ethics approval was required.
TA, KL and MC conceived the ideas and designed the methodology; TA collected the data; TA and MT analysed the data; TA, MT led the writing of the manuscript, with feedback and support from KL and MC. All authors contributed critically to the drafts and gave final approval for the publication.
This work was supported by the Woodtiger Fund, The Stadler Family Charitable Foundation Inc, Paul G. Allen Family Foundation, Thomas C Bishop Charitable Foundation, James & Deborah Burrows Foundation, Foundation Segre and Christopher Parker
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We thank the Government of Botswana for granting the approval and support for this research under the permit EWT 8/34/4 XX (34), granted by the Department of Wildlife and National Parks to Elephants Without Borders. We would also like to thank the University of New South Wales for logistical support. In addition, we would like to thank M. Zambo, I. Mwezi, R. Sutcliffe and K. Landen for their assistance with editing and field work.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2022.872472/full#supplementary-material
Supplementary Figure 1 | (A, B): Temporal activity and overlap between elephants hourly presence in the different corridors in (A) agricultural and (B) urban areas.
Adams T. S. F., Chase M. J., Rogers T., Leggett K. (2017). Taking the elephant out of the room and into the corridor: can urban corridors work? Oryx 51, 347–353. doi: 10.1017/S0030605315001246
Adams T. S. F., Mwezi I., Jordon N. (2020). Panic at the disco: solar-powered strobe light barriers incursions by African elephants loxodonta africana in chobe district, Botswana. Oryx 55 (5), 739–746. doi: 10.101017/S0030605319001182
Allen M. L., Sibarani M. C., Utoyo L., Krofel M. (2020). Terrestrial mammal community richness and temporal overlap between tigers and other carnivores in bukit barisan selatan national park, Sumatra. Anim. Biodiv. Conserv. 43, 97–107. doi: 10.32800/abc.2020.43.0097
Balmford A., Moore J. L., Brooks T., Burgess N., Hansen L. A., Williams P., et al. (2001). Conservation conflicts across Africa. Sci., 291 (5513) 2616–2619. doi: 10.1126/science.291.5513.2616
Blake S., Deem S. L., Strindberg S., Maisels F., Momont L., Inogwabini-Bila I., et al. (2008). Roadless Wilderness area determines forest elephant movements in the Congo. PloS ONE 3 (10), e3546.
Blom A., Van Zalinge R., Mbea E., Heitkonig I. M. A., Prins H. H. T. (2004). Human impact on wildlife populations within a protected central African forest. Afri. J. Ecol. 42 (1), 23–31. doi: 10.1111/j.0141-6707.2004.00441.x
Boettiger A. N., Wittemyer G., Starfield R., Volrath F., Douglas-Hamilton I., Getz W. M. (2011). Interfering ecological behavioural driver of African elephant movement using a linear filtering approach. Ecology 92 (8), 1648–1657. doi: 10.1890/10-0106.1
Bortolamiol S., Cohen M., Jiguet F., Pennec F., Seguya A., And Krief S. (2016). Chimpanzee non-avoidance of hyper-proximity to humans. J. Wildl. Manag. 80 (5), 924–934. doi: 10.1002/jwmg.1072
Buchholtz E. K., Spragg S., Songhurst A., Stronza A., McCulloch G., Fitzgerald L. A. (2021). Anthropogenic impact on wildlife resources use: Spatial and temporal shifts in elephants’ access to water. Afri. J. Ecol. 59 (3), 614–623. doi: 10.1111/aje.12860
Buij R., Mc Shea W. J., Campbell P., Ee M. E., Dallmeier F., Guimondou S., et al. (2007). Patch-occupancy models indicate human activity as major determinant of forest elephant Loxodonta cyclotis seasonal distribution in an industrial corridor in Gabon. Biol. Conserv. 135, 189–201. doi: 10.1016/j.biocon.2006.10.028
Caro T., Jones T., Davenport T. R.B.. (2009). Realities of documenting wildlife corridor in tropical countries. Biol. Con. 142(11), 2807–2811. doi: 10.1016/j.biocon.2009.06.011
Carter N. H., Shrestha B. K., Karki J. B., Pradhan N. M. B., Liu J. (2012). Coexistence between wildlife and humans at fine spatial scales. PNAS 109 (38), 15360–15365. doi: 10.1073/pnas.1210490109
Census Office (2022). Government of Botswana: Central Statistics office. 2022 population & housing census preliminary results brief, Gaborone: Central statistics office.
Chase M., Schlossberg S., Sutcliffe R., Seonyatseng E. (2019). “Dry season aerial survey of elephants and wildlife in northern Botswana: July–October 2018,” in Elephants without borders & department of wildlife & national parks of Botswana(Kasane, Botswana).
Clauss M., Scriba M., Kioko J., Ganzhorn J. U., Kiffner C. (2021). Camera-trap data do not indicate scaling of diel activity and cathemerality with body mass in an East African mammal assemblage. Ecol. Evol. 11, 13846–13861. doi: 10.1002/ece3.8090
DeMotts R., Hoon P. (2012). Whose elephants? conserving, compensating, and competing in northern Botswana. Soc Natur. Resour. 25, 837–851. doi: 10.1080/08941920.2011.638362
Department Of Wildlife And National Parks Of Botswana (2018). “A summary of problem animals control reports in the chobe district from 2007–2017,” in Elephants without borders(Kasane, Botswana).
Dickman A. J. (2009). Complexities of conflict: the importance of considering social factors for effectively resolving human-wildlife conflict. Anim. Conserv. 13 (5), 458–466. doi: 10.1111/j.1469-1795.2010.00368.x
Fox J., Weisberg S. (2018). Visualizing fit and lack of fit in complex regression models with predictor effect plots and partial residuals. J. Stat. Software 87 (9), 1–27. doi: 10.18637/jss.v087.i09
Fox J., Weisberg S. SAGE Publications, California. (2019). An r companion to applied regression. 3rd Edition (Thousand Oaks). Available at: https://socialsciences.mcmaster.ca/jfox/Books/Companion/index.html.
Garekae H., Lepetu J., Thakadu O. T. (2019). Forest resource utilization and rural livelihoods: insights from chobe enclave, Botswana. S. Afr. Geogr. J. 102, 22–40. doi: 10.1080/03736245.2019.1606730
Garekae H., Thakadu O. T., Lepetu J. (2017). Socio-economic factors influencing household forest dependency in chobe enclave, Botswana. Ecol. Proc. 6, 1. doi: 10.1186/s13717-017-0107-3
Gaynor K. M., Branco P. S., Long R. A., Goncalves D. D., Granli P. K., Poole J. H. (2018b). Effects of human settlement and roads on diel activity patterns of elephants (Loxodonta africana). Afri. J. Afri. Ecol. 56, 872–881. doi: 10.1111/aje.12552
Gaynor K. M., Hojnowski C. E., Carter N. H., Brashares J. S. (2018a). The influence of human disturbance on wildlife nocturnality. Science 360 (6394), 1232–1235. doi: 10.1126/science.aar7121
Ghoddousi A., Buchholtz E. K., Dietsch A. M., Williamson M. A., Sharma S., Balkenhol N., et al. (2021). Anthropogenic resistance: account for human bheaviour in wildlife connectivity planning. One Earth 4, 39–48. doi: 10.1016/j.oneear.2020.12.003
Government of Botswana (2021). Botswana Elephant management plan and action plan 2021:2026 (Gaborone: Department of Wildlife and National Parks).
Graham M. D., Douglas-Hamilton I., Adams W. M., Lee P. C. (2009). The movement of African elephants in a human-dominated land-use mosaic. Anim. Conserv. 12, 445–455. doi: 10.1111/j.1469-1795.2009.00272.x
Gunn J., Hawkins D., Barnes R. F. W., Mofulu F., Grant R. A., Norton G. W. (2013). The influence of lunar cycles on crop-raiding elephants: evidence of risk avoidance. Afri. J. Ecol. 52, 129–137. doi: 10.1111/aje.12091
Gupta C. A. (2013). Elephant, safety nets and agrarian culture: understanding human-wildlife conflict and rural livelihood around chobe national park, Botswana. J. Polit. Ecol. 20, 238–254. doi: 10.2458/v20i1.21766
Hoare R. E. (2003). Fencing and other barriers against problem elephants (Nairobi, Kenya: African Elephant Specialists Group Technical Brief Series. IUCN/SSC African Elephant Specialists Group).
Hoare R. (2015). Lessons from 20 years of human–elephant conflict mitigation in Africa. Hum. Dimens. Wildl. 20, 289–295. doi: 10.1080/10871209.2015.1005855
Jachowski D. S., Slotow R., Millspaugh J. J. (2013). Corridors use and streaking behaviour by African elephant in relation to physiological state. Conserv. Biol. 167, 276–282. doi: 10.1016/j.biocon.2013.08.005
Jackson T. P., Mosojane S., Ferreira S. M., Van Aarde R. J. (2008). Solutions for elephant Loxondonta africana crop raiding in northern Botswana: moving away from symptomatic approaches. Oryx 4, 83–91.
Jones K. R., Venter O., Fuller R. A., Allan J. R., Maxwell S. L., Negret P. J., et al. (2018). One-third of global protected land is under intense human pressure. Science 360, 788–791. doi: 10.1126/science.aap9565
Kansky R., Knight A. T. (2014). Key factors driving attitudes towards large mammals in conflict with humans. Biol. Conserv. 179, 93–105. doi: 10.1016/j.biocon.2014.09.008
Kiffner C., Nagar S., Kollmar C., Kioko J. (2016). Wildlife species richness and densities in wildlife corridors of northern Tanzania. J. Nat. Conserv. 31, 29–27. doi: 10.1016/j.jnc.2016.02.006
Kuznetsova A., Brockhoff P. B., Christensen R. H. B. (2017). lmerTest Package: Tests in Linear Mixed Effects Models. J. Stat. Software 82 (13), 1–26. doi: 10.18637/jss.v082.i13
LaPoint S., Gallery P., Wikelski M., Kays R. (2013). Animal behavior, cost-based corridor mode corridors. Landscape Ecology 23, 1615–1630. doi: 10.1007/s10980-013-9910-0
Magle S. B., Hunt V. M., Veron M., Crooks K. R. (2012). Urban wildlife research: past, present and future. Biol. Conserve. 155, 23–332. doi: 10.1016/j.biocon.2012.06.018
Martins J. H., Camanho A. S., Gaspar M. B. (2012). A review of the application of driving forces–pressure–state–impact–response framework to fisheries management. Ocean. Coast. Manage. 69, 273–281. doi: 10.1016/j.ocecoaman.2012.07.029
Ministry of Lands, Housing and Environment (2000). Scott Wilson Field investigation 2000 (Gaborone, Botswana: Department of Town and Regional Planning).
Mumby H., Plotnik J. M. (2018). Taking the elephants’ perspective: Remembering elephant behavior, cognition and ecology in human– elephant conflict mitigation. Front. Ecol. Evol. 6, 122. doi: 10.3389/fevo.2018.00122
Osborn F. V., Parker G.E. (2003) Linking two elephant refuges with a corridor in the communal lands of Zimbabwe. African J. of Ecol. 41, 68–74.
R Core Team (2019). R: A language and environment for statistical computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org/.
Ridout M. S., Linkie M. (2009). Estimating overlap of daily activity patterns from camera trap data. J. Agric. Biol. Envi. Stat. 14 (3), 322–337. doi: 10.1198/jabes.2009.08038
Roever C. L., Van Aarde R. J., Chase M. J. (2013). Incorporating mortality into habitat selection to identify secure and risky habitats for savannah elephants. Biol. Cons. 164, 98–106.
Rodriguez-Soto C., Monroy-Vilchis O., Zarco-Gonzalez M. M. (2013). Corridors for jaguar (Panthera onca) in Mexico. Conserv. Strat. 21, 438–443. doi: 10.1016/j.jnc.2013.07.002
Shamoon H., Maor R., Saltz D., Dayan T. (2018). Increased mammal nocturnality in agricultural landscapes results in fragmentation due to cascading effects. Biol. Conserv. 226, 32–41. doi: 10.1016/j.biocon.2018.07.028
Snyer K. D., Mneney P., Benjamin B., Mkilindi P., Mbise N. (2019). Seasonal and spatial vulnerability to agricultural damage by elephants in the western Serengeti, Tanzania. Oryx 55 (1), 1390149. doi: 10.1017/S0030605318001382
Songhurst A., Mcculloch G., Coulson T. (2016). Finding pathways to human–elephant coexistence: a risky business. Oryx 50 (4), 713–740. doi: 10.1017/S0030605315000344
Thouless C. (1996). Home ranges and social organization of female elephants in northern Kenya. Afri. J. Ecol. 33, 284–297. doi: 10.1111/j.1365-2028.1996.tb00623.x
Tiller L. N., Humle T., Amin R., Deere N. J., Lago B. O., Leader-Williams N., et al. (2021). Changing seasonal, temporal and spatial crop-raiding trends over 15 years in a human-elephant conflict hotspot. Biol. Conserv. 254, 108941. doi: 10.1016/j.biocon.2020.108941
Venter O., Sanderson E. W., Magrach A., Allan J. R., Beher J., Jones K. R., et al. (2016). Sixteen years of change in the global terrestrial human footprint and implications for biodiversity conservation. Nat. Commun. 7, 12558. doi: 10.1038/ncomms12558
Wall J., Wittemyer G., Klinkenberg B., LeMay V., Blake S., Strindberg S., et al. (2021). Human footprint and protected areas shape elephant range across Africa. Curr 31 (11), 2437–2445. doi: 10.1016/j.cub.2021.03.042
World Bank (2019). Available at: https://data.worldbank.org/country/botswana (Accessed 23/7/2021).
Keywords: Botswana, camera traps, conservation, human pressure, human-elephant conflict, coexistence, land-use management, wildlife corridors
Citation: Adams TSF, Leggett KEA, Chase MJ and Tucker MA (2022) Who is adjusting to whom?: Differences in elephant diel activity in wildlife corridors across different human-modified landscapes. Front. Conserv. Sci. 3:872472. doi: 10.3389/fcosc.2022.872472
Received: 09 February 2022; Accepted: 29 June 2022;
Published: 28 July 2022.
Edited by:
Benoit Goossens, Cardiff University, United KingdomReviewed by:
Ee Phin Wong, University of Nottingham Malaysia Campus, MalaysiaCopyright © 2022 Adams, Leggett, Chase and Tucker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tempe S. F. Adams, dGVtcGVAdW5zdy5lZHUuYXU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.