 Laura Bosco
Laura Bosco Damaris Siegenthaler2
Damaris Siegenthaler2 Livio Ruzzante
Livio Ruzzante Alain Jacot
Alain Jacot Raphaël Arlettaz
Raphaël Arlettaz- 1Finnish Museum of Natural History, University of Helsinki, Helsinki, Finland
- 2Division of Conservation Biology, Institute of Ecology and Evolution, University of Bern, Bern, Switzerland
- 3Department of Ecology and Evolution, University of Lausanne, Lausanne, Switzerland
- 4Swiss Institute of Bioinformatics, Lausanne, Switzerland
- 5Swiss Ornithological Institute, Regional Office Valais, Sion, Switzerland
Alternative farming methods must be deployed to mitigate the detrimental impacts of intensive agriculture on climate, biodiversity, and ecosystem services. Organic and biodynamic farming are environmental-friendly practices that progressively replace conventional agriculture. While potential biodiversity benefits of organic vs. conventional farming have been studied repeatedly, the effects of biodynamic farming on biodiversity remain ill-understood. We investigated the effects of these three main management regimes, and their interaction with ground vegetation cover, on vineyard invertebrate communities in SW Switzerland. Invertebrates were sampled three times during the vegetation season in 2016, focusing on ground-dwelling (pitfall traps) and epiphytic (sweep-netting) invertebrates, and their abundance was modelled for single, additive, and interactive influences of management and ground vegetation cover. Overall, organic and, but to a lesser degree, biodynamic vineyards provided better conditions for invertebrate abundance than conventional vineyards. On the one hand, there was a significant interaction between management and ground vegetation cover for epiphytic invertebrates with a positive linear increase in abundance in organic, a positive curvilinear relationship in biodynamic but a negative curvilinear response to vegetation cover in conventional vineyards. The abundance of ground-dwelling invertebrates was primarily affected by the management regime alone, i.e. without any interaction with ground vegetation characteristics, leading to much higher abundances in organic compared to conventional vineyards, while biodynamic did not differ from the other two regimes. We interpret the patterns as follows: organic grape production offers more suitable habitat conditions for invertebrates due to a spatially more heterogenous but also less often disturbed (compared to biodynamic management) or destroyed (compared to conventional) ground vegetation cover, in line with the predictions of the intermediate disturbance hypothesis. Biodynamic and conventional viticultural management regimes often provide a habitat that is either too mineral (conventional: ground vegetation widely eliminated) or subject to soil disturbance happening frequently through ploughing (biodynamic). We conclude that alternative farming methods do promote biodiversity in vineyard agro-ecosystems, especially so organic management.
Introduction
Agriculture is one of the main drivers of climate disruption and biodiversity erosion (Rockström et al., 2009). Alternative management practices to conventional farming have thus to be developed and applied widely to mitigate its impacts (Tscharntke et al., 2021), which is reflected in several European farmland strategies such as e.g. the new Common Agricultural Policy (CAP), or the EU Green Deal including the farm-to-fork strategy aiming to ensure that 25% of European farmland will be organic by 2030. Most vineyard regions of the world have undergone a massive intensification of farming practices during the past decades, resulting in a dramatic simplification of the landscape, further inflating the ongoing biodiversity crisis (Tscharntke et al., 2012; Maxwell et al., 2016). Recently, however, trends towards more environmentally friendly vineyard management modes have arisen, leading to a general decline in pesticide applications, paralleled by a marked increase in organic grape production (Provost and Pedneault, 2016).
Today’s vineyards are typically farmed according to three management modes: conventional (C), organic (O) or biodynamic (BD). In conventional production, chemicals such as synthetic fertilizers and pesticides are allowed. To reduce competition for water and nutrients between the vines and the ground vegetation, conventional farmers use herbicides – often applied over the entire production surface – while organic and biodynamic farmers use mechanical methods or, less often, grazing by sheep, horses, or poultry to remove parts of the ground vegetation layer. Less herbicide application leads to more ground vegetation in vineyards, which is benefiting a more diverse flora and entomofauna (Winter et al., 2018; Bosco et al., 2019; Fried et al., 2019), with cascading effects up to vertebrates (Guyot et al., 2017; Bosco et al., 2020; Munoz-Saez et al., 2020; Petrescu Bakış et al., 2021) and for ecosystem services (Thomson and Hoffmann, 2009; Sanguankeo and León, 2011; Saenz-Romo et al., 2019). In addition to indirect benefits such as increased ground vegetation through fewer herbicide applications, a major cause of reduced biodiversity in conventional farming systems are direct lethal or sublethal effects of agrochemicals, which impact in particular soil biodiversity (Karimi et al., 2020).
In general, organic production only uses natural fungicides and fertilizers (Lampkin, 1999; Caprio et al., 2015) and is targeted towards achieving a well-balanced ecosystem (Provost and Pedneault, 2016). Biodynamic production can be considered as a form of organic production, but the entire farm is further viewed as an organism in itself, aiming for closed nutrient cycles (Vogt, 2007). In addition to organic management, biodynamic farming usually includes fermented manure, and plant preparations that are applied to the soil and crops for stimulation of soil nutrient cycling (Döring et al., 2015). Among all types of crops cultivated worldwide, it is primarily in grapevine production that biodynamic management is most widespread (Phillips and Rodgriguez, 2006). Whether biodynamic cultivation is more beneficial for biodiversity than organic farming remains controversial (Turinek et al., 2009; Döring et al., 2015; Morrison-Whittle et al., 2017; Döring et al., 2019). The effects of biodynamic farming on biodiversity have quite rarely been explored so far, as biodynamic and organic farming are often combined together when carrying out comparisons of trendy farming practices against conventional management (Döring et al., 2019). For instance, just a few studies have tested the effects of conventional vs. biodynamic management on fungal communities (Morrison-Whittle et al., 2017), or organic vs. biodynamic management on soil and grape quality (Carpenter-Boggs et al., 2000; Reeve et al., 2005), while effects on aboveground biodiversity have remained extremely scarce (Döring et al., 2019). On the other hand, organic viticulture has been shown to have beneficial effects on species richness and abundance across many taxa such as birds, insects, soil micro-organisms and plants (e.g. Nascimbene et al., 2012; Caprio et al., 2015; Caprio and Rolando, 2017; Masoni et al., 2017; Puig-Montserrat et al., 2017; Karimi et al., 2020), with only a handful of studies not evidencing any positive effects in vineyards (Bruggisser et al., 2010; Assandri et al., 2016). Such negative, mixed or neutral effects of organic management are often attributable to masking landscape effects (e.g., through stronger effects of landscape heterogeneity), which are beyond the mode of farmland management at parcel level (Hole et al., 2005).
In Switzerland, 30% of vineyards are situated in the southwest, at sun-exposed locations along the Rhône river, in the canton of Valais. About 90% of the vineyards in this region are managed according to the integrated production (IP) protocol (hereafter named conventional regime as this has become the basic farming standard nowadays), which has led to a marked reduction in synthetic pesticide utilization in the recent past. In Valais, a long-running trend in reducing herbicide application has resulted in more ground vegetation in vineyards overall. However, since the reliance on herbicides is not explicitly forbidden in the conventional regime, over 70% of Valais vineyards still have a very bare appearance, i.e., are mostly devoid of any ground vegetation (Bosco et al., 2018). The remaining 30% show a partial or full ground vegetation cover, resulting from a reduced use or full renouncement of herbicides, of which ca 25% follow an organic and less than 5% a biodynamic production protocol.
In organic and biodynamic vineyards, a variable fraction of the ground is generally vegetated: in effect, reduction or removal of the ground vegetation is only allowed mechanically or through grazing, and usually in every other inter-row under these two management regimes. There are subtle differences, however, in the way organic and biodynamic farming manage ground vegetation cover. In organic vineyards, ground vegetation is usually mown 2-3 times a year, more often so under the vine row than in the inter-row, and very rarely ploughed because there is no need of intervention in the soil. In contrast, the biodynamic management often includes ploughing, because of the requirement for special manure applications, with operations typically occurring 3-4 times a year. As a result, organic vineyards offer a heterogeneous but temporally more static arrangement of ground vegetation micro-habitats while biodynamic vineyards are subjected to a more radical disturbance of their ground vegetation layer, leading to a potentially spatially less heterogeneous and more static assemblage of micro-habitats. Examined from the predictions of the intermediate disturbance hypothesis, which posits that biodiversity peaks at an intermediate level of habitat disturbance (Norris and Kogan, 2000; Mackey and Currie, 2001; Sanguankeo and León, 2011), these three management regimes allow drawing predictions for biodiversity patterns in conventional, organic, and biodynamic vineyards. In effect, conventional management entails a major ecological disturbance in vineyards, due to a total and quasi permanent elimination of vegetation on the ground surface through the application of herbicides; biodynamic is characterized by a rather high level of disturbance as well (frequent ploughing operations) although it allows ground vegetation cover to develop; finally, organic management intervenes less radically and less often on the ground vegetation cover of vineyards, typically representing an intermediate ecological disturbance (Figure 1).
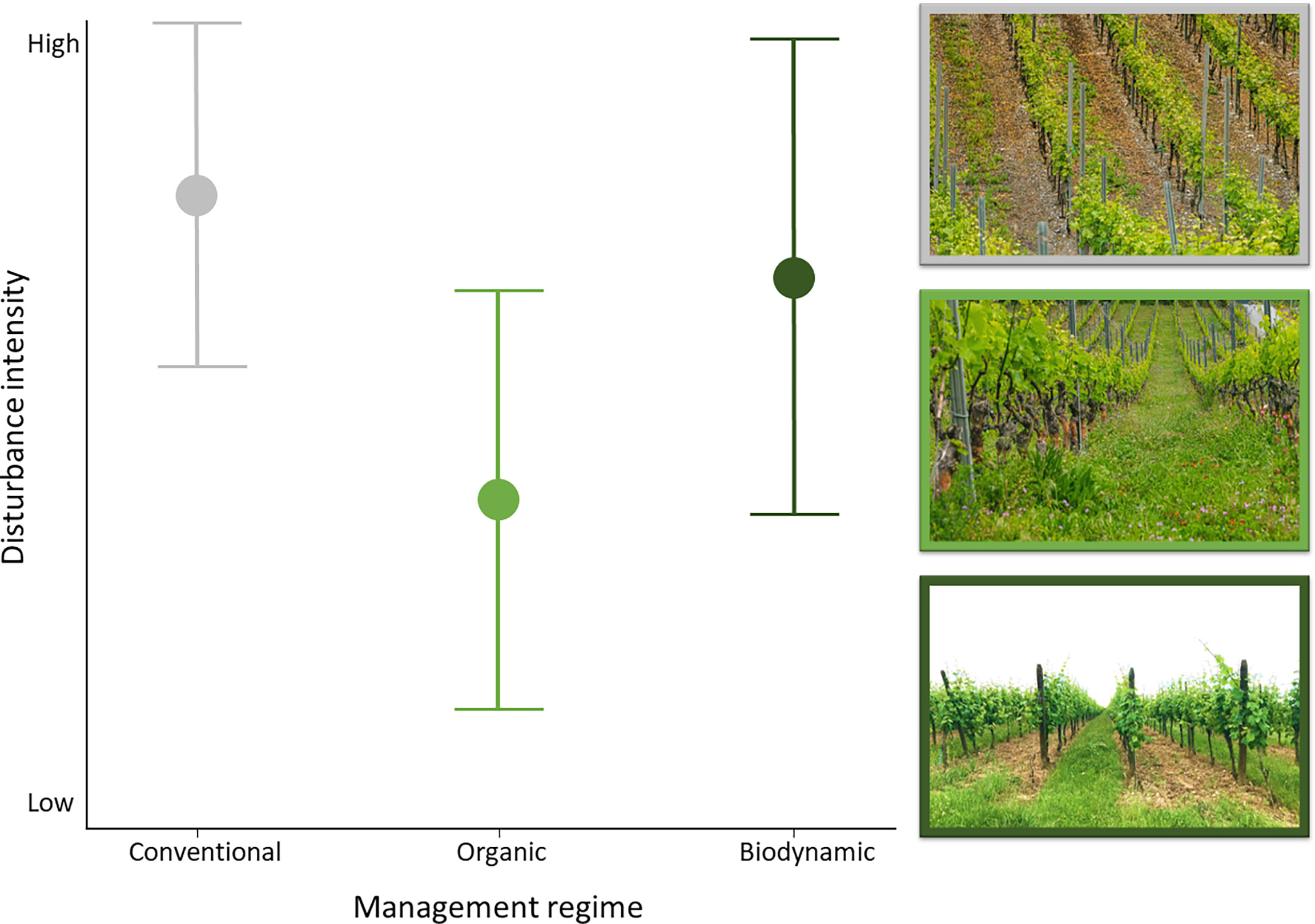
Figure 1 Schematic representation of hypothesized soil and ground vegetation disturbance levels dependent on the management regime (i.e., conventional in grey; organic in green; biodynamic in dark green) and example pictures for the three management modes. Top: a high amount of bare ground due to herbicide applications characterizes conventional vineyards. Middle: organic vineyards are characterized by a structurally complex, and plant species diverse ground vegetation layer, in this case covering 100% of the surface. Bottom: an extreme case of biodynamic management, where every other inter-row was recently ploughed. ("(C) C. Pfammatter").
In this study, we measured the invertebrate fauna inhabiting Swiss vineyards characterized by the three different management regimes, while accounting for ground vegetation conditions – this to decipher potential interactions with farming mode. We predicted that: i) overall invertebrate abundance is higher in organic and biodynamic vineyards compared to conventional vineyards (hypothesis H1); ii) invertebrate abundance is higher in organic than biodynamic vineyards, in line with the predictions of the intermediate disturbance hypothesis (H2); iii) the relationship between ground vegetation cover and invertebrate abundance depends on the underlying management regime (H3); and iv) different invertebrate taxa exhibit contrasted, group-specific responses to management regime and ground vegetation cover (H4). Our overarching objective was thus to test whether new, alternative agricultural practices such as biodynamic farming benefit vineyard biodiversity, while a more specific goal was to draw evidence-based guidance for more biodiversity-friendly vineyard management.
Material and Methods
Study Area and Sampling Design
This study was carried out in 2016 in the vineyards of the upper Rhône Valley, covering an area of ca 50 km2, between the communities of Salgesch (46°18′N, 7°34′E) and Fully (46°08′N, 7°07′E), Canton of Valais, SW Switzerland. Valais vineyards are located mostly on steep, often traditionally terraced, south-exposed slopes up to 900 m above sea level and on shallow slopes next to the plain (Arlettaz et al., 2012). These vineyards grow on different soil types and harbor a fair diversity of rare and specialized plant and animal species (Sierro and Arlettaz, 2003). We selected 35 vineyard fields (Supplementary Material Table S1) that could be attributed unequivocally to the three different management regimes: conventional (N=14), organic (N=12) and biodynamic production (N=9). The sample size for the latter category was constrained by the scarcity of that management mode in the study area. Within each management regime, we selected fields along a gradient from 0 to 100% ground vegetation cover (Supplementary Material Figure S5A), which was measured by visually estimating the average ground vegetation coverage (%) for the entire field (following Bosco et al., 2019). Mean ± SD ground vegetation cover was 32.7 ± 21.2%, 55.3 ± 25.8% and 58.9 ± 23.9% for conventional, organic, and biodynamic fields, respectively, leading to a significant difference between O-C (p<0.001) and BD-C (p<0.001) (one-way ANOVA). To avoid spatial clustering of vineyards with a given management regime, vineyard fields were selected in a stratified random manner to ensure a balanced spatial distribution across the entire study area. Given the limited availability of vineyards under biodynamic management, however, these fields were more clustered than the conventional and organic vineyards (see Supplementary Material Figure S1).
Invertebrate Sampling
We used two different sampling methods to record either epiphytic invertebrates, sampled with sweep-netting or ground-dwelling arthropod taxa, which were sampled with pitfall traps. We sampled during three sessions in 2016, with a duration of pitfall trap opening of one week per session (early spring: May, week 19/20; late spring: June, week 25/26 and summer: August, week 31/32).
Sweep-netting was performed along a transect running in an interrow next to one of the three pitfall trap locations. In case of vineyards vegetated only in every other interrow, we sampled in the vegetated interrow. The net was moved for 20 footsteps with one swing per footstep, while to avoid possible edge effects, we started the sweep-netting five steps off the vineyard edge (Bruggisser et al., 2010). Sweep-netting was carried out three times as well, once during every pitfall trapping session, and only on sunny days with no or low wind speed. For the pitfall trapping, we placed three traps in three different rows per vineyard field with at least 5 m distance between the traps and a distance of at least 10 m to the edge of the vineyard field. The traps were placed at the edge of the inter-rows to avoid destruction by mowing machines in case operations were carried out through the inter-row during the sampling periods. The pitfall traps were plastic cups of 9 cm diameter, 11.5 cm depth and 0.5 L volume. Every trap was filled with a 1:1 mixture of water and propylene glycol, which served as a preservation fluid while being harmless for the environment (Woodcock, 2005). A drop of detergent was added to reduce the surface tension of the solution.
The pitfall trap and sweep-netting samples were stored in 98% ethanol in plastic tubes until identification in the lab. All invertebrates except ants – due to their potentially highly clumped abundances related to nearby colonies – were counted and identified to order level. For the order Hemiptera, we identified each specimen at the suborder level, distinguishing between leafhoppers (Auchenorrhyncha), aphids (Sternorrhyncha) and heteropterans (Heteroptera) and hereafter summarizing orders and suborders as “groups”. The mean percentage of ground vegetation cover per vineyard field was estimated at every sampling session (i.e. three separate measures per field).
Statistical Analyses
Pitfall trap and sweep-netting data were analyzed separately while for both datasets there were three repeated measures for all 35 sampling fields, resulting in 105 observations per dataset. Out of a total of 315 pitfall traps, 18 had unfortunately been destroyed or removed. The three pitfall traps per field were pooled, to obtain averaged abundance values per vineyard field. Fields with missing traps (< 3 traps), and those where mice or lizards had been trapped were excluded due to the potential attraction of scavenger arthropods, resulting in a final total of 93 observations for pitfall trapping (see Supplementary Material Table S2 for full list of sample sizes per session). Invertebrate abundance was log-transformed to fit linear mixed models with a Gaussian distribution, using the R-package lme4 (Bates, 2015). We included the coordinates of sampling fields as a random effect (factor with 35 levels) to account for potential spatial autocorrelation of close-by traps and repeated samplings. Based on our hypotheses, we had three different modelling approaches. (1) The first model only included the factor management (three levels) as a fixed effect, to test for differences between management regimes (H1 and H2; see Introduction). (2) To test the third hypothesis, we fitted an additive model including management and ground vegetation cover as fixed effects as well as an interactive model including the term management*vegetation. For ground vegetation cover, the second polynomial order was always included in the models given a known curvilinear response evidenced in several studies of habitat selection by birds inhabiting vineyards (e.g. Arlettaz et al., 2012; Guyot et al., 2017; Maurer et al., 2020). Model assumptions such as normality of residuals and homogeneity of variance were visually checked using check_model from R package performance (Lüdecke et al., 2021). AIC weights and R2 conditional were calculated and posthoc Tukey comparisons for effects of the three management regimes were computed using the glht function from the R package multcomp (Hothorn et al., 2008). All analyses were performed using R version 3.6.2 (R Core Team, 2020).
Indicator Analysis
Finally, we performed indicator analyses to check whether certain invertebrate groups might be specifically linked to one of the management regimes (H4), using the function multipatt of the R-package indicspecies (Cáceres and Legendre, 2009). In addition, by categorizing ground vegetation cover into four density classes (bare: 0-20%, intermediate: 21-40%, high: 41-60%, very high: >60% cover), we investigated links between invertebrate groups and ground vegetation cover. An indicator value (IndVal) varying between 0 and 1 was calculated for each group with the maximum value 1 attributed to a group found in all sites of a class.
Results
Invertebrate Abundance
From sweep net samples (epiphytic invertebrates), a total of 8’286 invertebrates were counted, with a range of 4 to 430 and an average of 79.67 ± 73.7 (SD) specimens per vineyard field (for management specific averages see Supplementary Material Table S3). Among those, 11 groups could be identified, including the categories “other” (unidentified taxa occurring only marginally) and “larvae” (larvae of some taxa often look similar and were thus grouped together). Diptera (24%), Auchenorrhyncha (20%) and Heteroptera (14%) were the most abundant groups with average specimen numbers ( ± SD) of 19.23 ± 18.1, 15.87 ± 18.6, 11.16 ± 17.4 per vineyard, respectively (Supplementary Material Figure S2A).
From pitfall traps (ground-dwelling invertebrates), a total of 21’750 invertebrates were recorded, with a range of 74 to 555 and an average of 233.87 ± 100.5 specimens per vineyard. Sixteen groups, plus “other” and “larvae”, were recognized, with Coleoptera (23%), Diptera (17%) and Araneae (15%) being the most abundant (mean ± SD abundance=52.98 ± 33.8, 39.74 ± 33.8, 35.51 ± 25.9 respectively; Supplementary Material Figure S2B).
Differences Between Farming Regimes
For the sweep net samples, invertebrate abundance was significantly higher in organic and biodynamic compared to conventional vineyards, with no significant difference between organic and biodynamic fields, although the former showed higher mean figures than the latter (posthoc Tukey test, O-C: z=3.576, p=0.001; BD-C: z=2.811, p=0.014; BD-O: z=-0.468, p=0.886; Table 1A and Figure 2A).
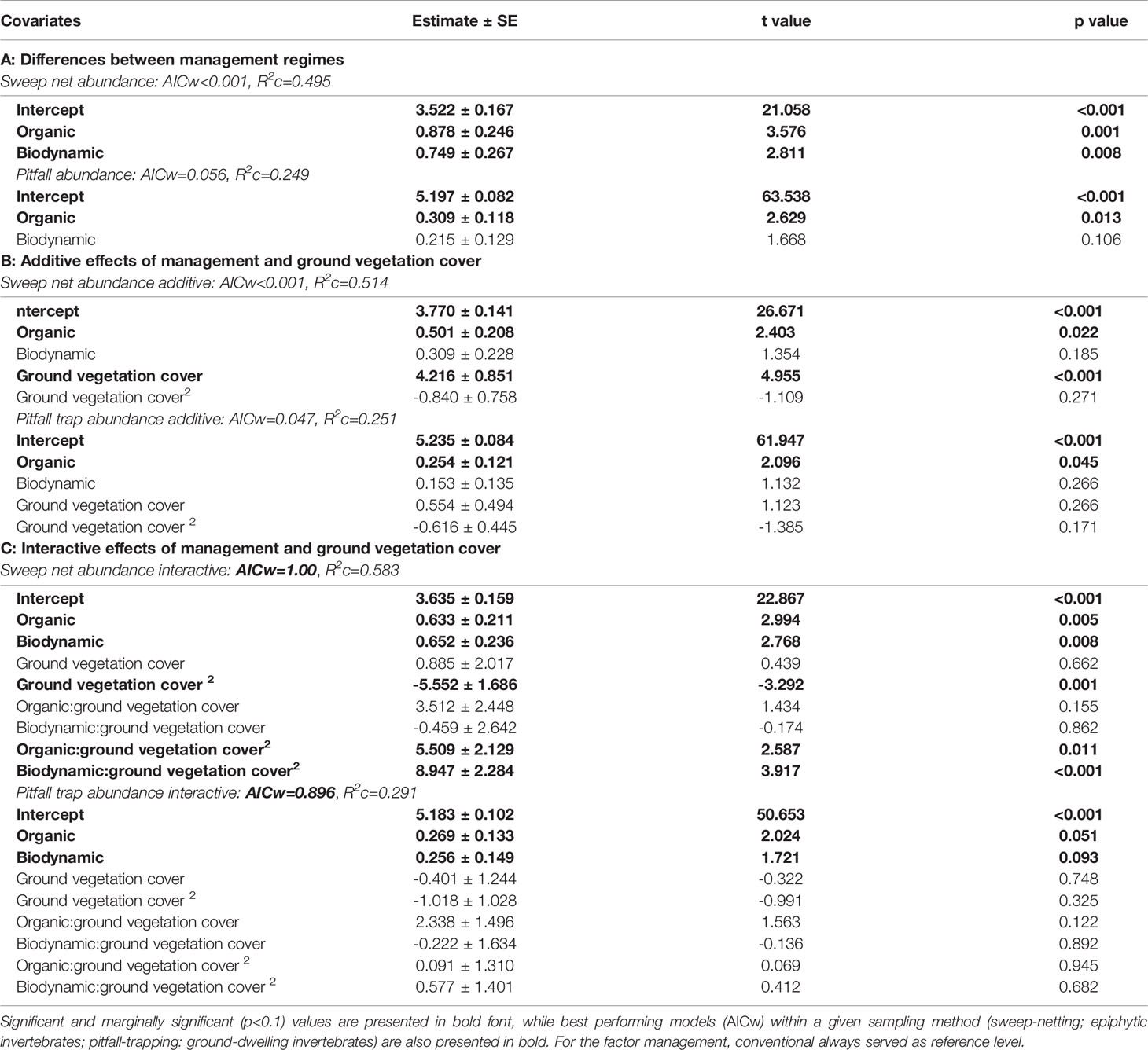
Table 1 Estimates of the three different models (A): management regime only; (B): management regime + ground vegetation cover; (C): management regime*ground vegetation cover) for invertebrate abundance with respect to sampling method.
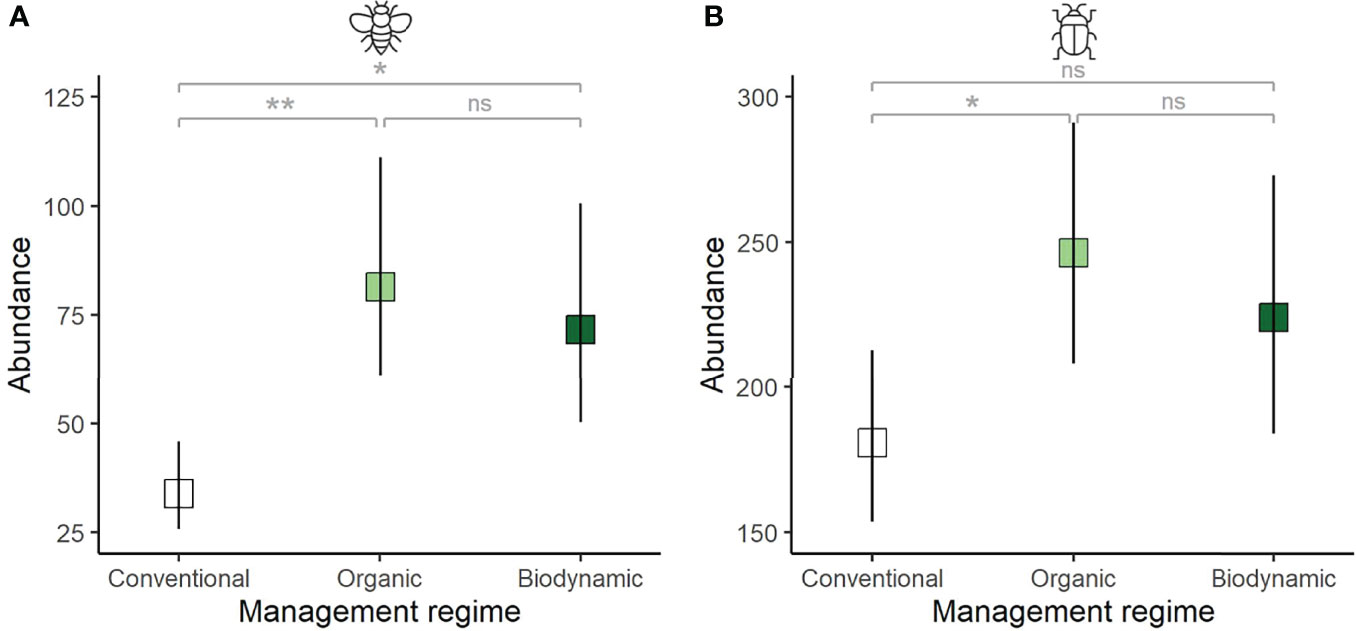
Figure 2 Differences in invertebrate abundance from (A) sweep net (represented by dipterans) and (B) pitfall trap samples (represented by beetles) with respect to management regime. Plotted are the model-based predictions, where boxes show mean estimates and vertical lines the 95% confidence intervals. Stars indicate significance of p values (**p<0.01; *p<0.05), ns, non-significant.
In pitfall trap samples, invertebrate abundance also showed significant differences between organic and conventional fields, while there were no statistically significant differences between biodynamic and conventional, or biodynamic and organic fields (posthoc Tukey test, O-C: z=2.629, p=0.0232; BD-C: z=1.668, p=0.217; BD-O: z=-0.722, p=0.750; Tables 1A and Figure 2B).
Combining Management Regime and Ground Vegetation Cover
When including ground vegetation cover into the modelling of sweep net invertebrate abundance, the significant difference between organic and conventional fields remained, but biodynamic did not differ any longer from the other two regimes (posthoc Tukey test, O-C: z=2.403, p=0.0428; BD-C: z=1.354, p=0.365; BD-O: z=-0.879, p=0.653; Table 1B). The linear term of ground vegetation cover showed a strong positive influence on the number of invertebrates per vineyard field (Table 1B and Figure 3). Introducing the interaction between management regime and ground vegetation cover into the model resulted in significantly different effects of ground vegetation cover in organic and biodynamic from effects in conventional vineyards: a positive linear increase in organic, a convex curvilinear relationship in biodynamic but a concave curvilinear response in conventional vineyards (Figure 4). The interactive model performed better than the simple and the additive models, as shown by its lowest AICc value for sweep net abundance (Table 1A–C; depicted in bold).
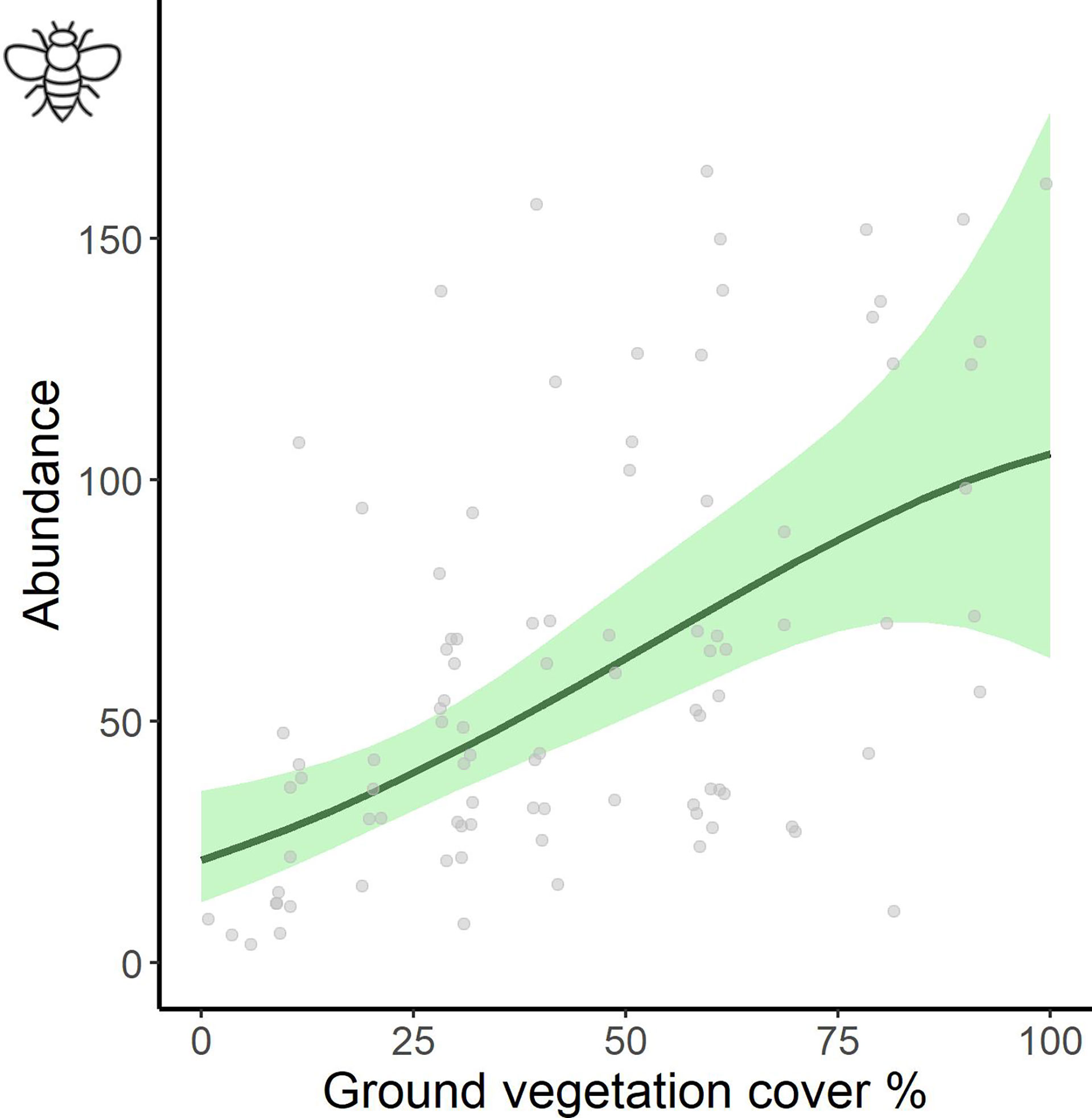
Figure 3 Effects of ground vegetation cover on invertebrate abundance as obtained from the additive model based on with sweep-netting samples (epiphytic invertebrates represented by dipterans). Plotted are model-predicted means (solid line), 95% confidence intervals (shaded area) and raw data points (grey dots).
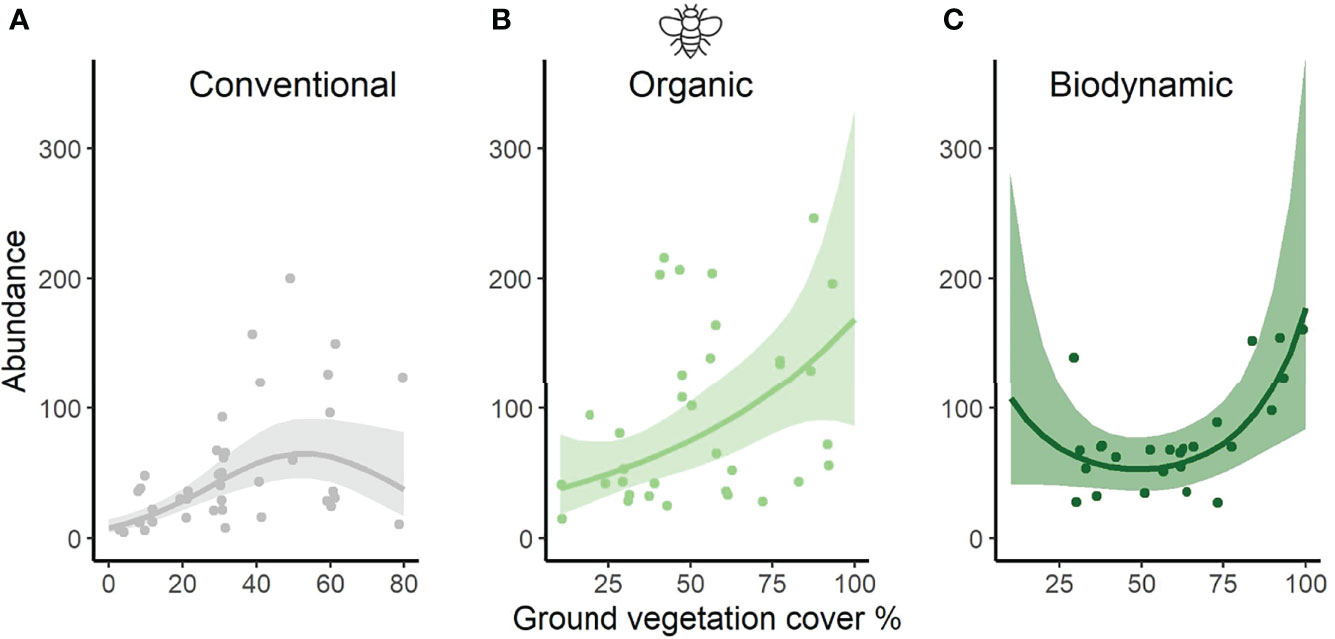
Figure 4 Effects of ground vegetation cover on invertebrate abundance as obtained from sweep-netting samples (epiphytic invertebrates represented by dipterans), modulated by management regime shown for (A) conventional, (B) organic, and (C) biodynamic management separately. Plotted are predicted estimates from the interaction term, with means (solid lines) and 95% confidence intervals (CI, shaded area). Points show the raw data, while the x-axes were restricted to the value ranges of ground vegetation coverage per management regime (C: 0-80%; O: 10-100%; BD: 10-100%). The effects of ground vegetation cover in organic and biodynamic vineyards are significantly different from the effects in conventional vineyards. The large overlap of the CI for organic and biodynamic translates into an absence of significant effects between them. Note the particularly small sample sizes for biodynamic in the lowest values (left hand) of the ground vegetation cover gradient and vice versa for conventional at the highest values (right hand) of the gradient.
In the pitfall trap samples, ground vegetation cover did not affect invertebrate abundance in the additive models, while organic vineyards again yielded significantly higher abundances compared to conventional vineyards (Table 1B). In the interactive model, we found no statistical difference in the effect of ground vegetation between the three management regimes (Table 1C).
Indicator Analyses
The analysis of the sweep net data established that there were three groups of invertebrates linked to organic (heteropterans, hymenopterans, dipterans) and one linked to biodynamic management (spiders; Table 2). As concerns the ground vegetation cover classes, seven groups (hymenopterans, cicadas, dipterans, heteropterans, butterflies, spiders, and larvae) were linked to a high ground cover (60-100%, Table 2).
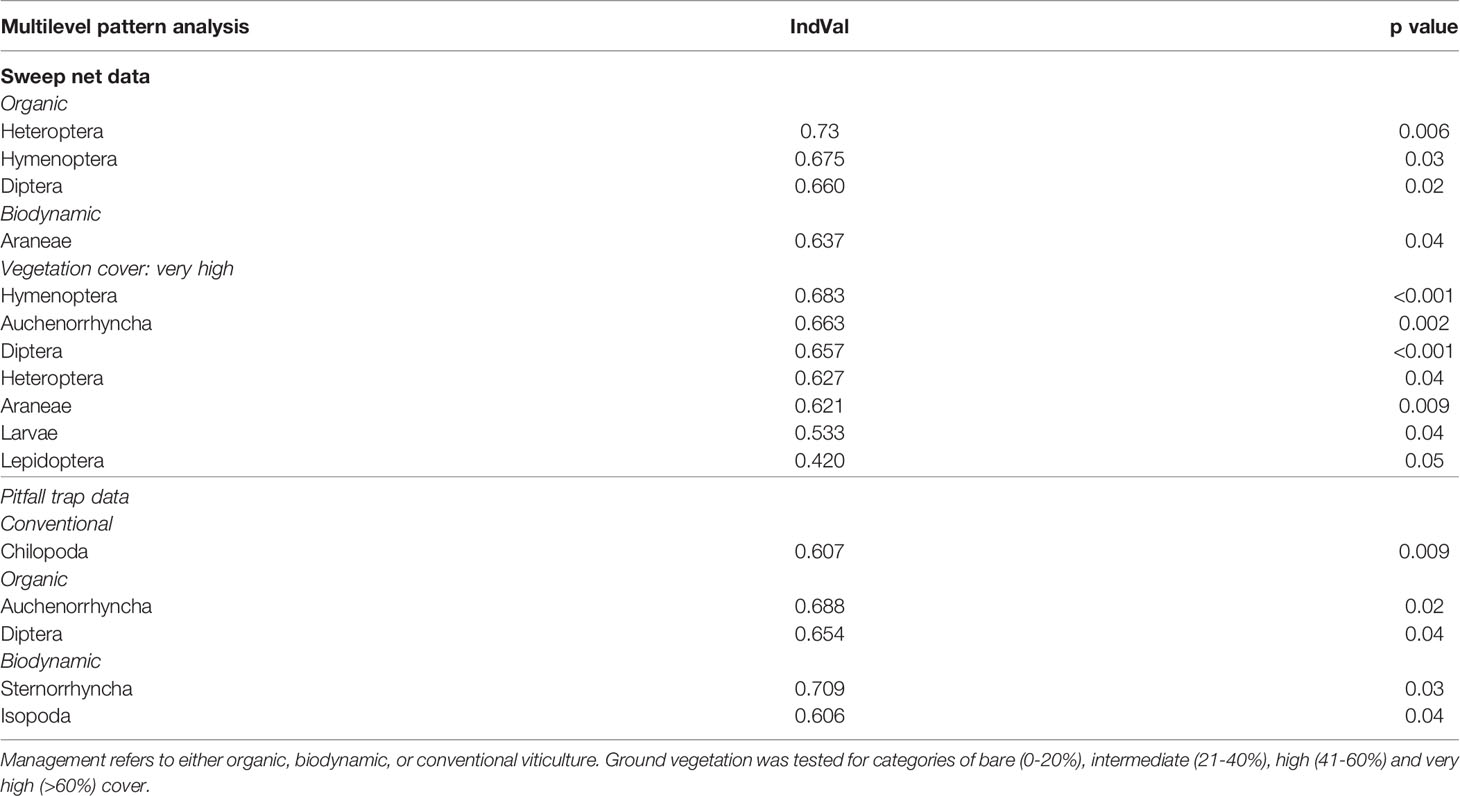
Table 2 Indicator analyses output, showing only significant links given by indicator values (IndVal) ranging from >0.5 to 1.
The analysis of the pitfall trap data indicated that centipedes were linked to conventional management, while Sternorrhyncha and isopods were associated with biodynamic, and finally cicadas and dipterans with organic farming (Table 2). No indicators of vegetation cover classes emerged from the pitfall trap data.
Discussion
This study demonstrates that there is a large potential to increase biodiversity in vineyards overall, providing that alternative farming modes, in replacement of conventional viticulture, are applied. In effect, organic and biodynamic viticulture offer a much wider palette of ecological niches, and thus many more resources for the development of diverse communities and abundant flora and fauna populations, in addition to the benefits stemming from a reduction of the adverse toxicological effects of no or fewer pesticide applications compared to conventional farming. Ultimately, however, much seems to depend on the way the ground vegetation layer is managed. These findings indicate that management-specific relationships exist between a given biodiversity measure (ground greening) and invertebrate abundance, highlighting the need to consider the mode of agricultural management when suggesting ecological revitalization measures for vineyards.
Organic Management Provides Better Conditions for Invertebrate Abundance
In line with our first hypothesis (H1), organic management promotes the abundance of ground-dwelling invertebrates in vineyards, much more so than conventional farming, while epiphytic invertebrates abound under both organic and biodynamic farming, being considerably less numerous in conventionally cultivated vineyards. A general positive influence of organic farming on biodiversity has been shown repeatedly (Puig-Montserrat et al., 2017; Barbaro et al., 2021; Tscharntke et al., 2021) and has mainly been linked to a reduced application of pesticides and their adverse effects on biodiversity, in particular soil organisms (Nascimbene et al., 2012; Trivellone et al., 2012; Masoni et al., 2017; Karimi et al., 2020). If organic and biodynamic management, contrary to conventional farming, are both beneficial to epiphytic invertebrates, it is likely because of an increased ground vegetation coverage, and an enhanced structural complexity and diversity of the flora (Roschewitz et al., 2005; Gabriel et al., 2006; Rundlöf et al., 2008). If habitat heterogeneity is generally the crux for promoting farmland biodiversity at the landscape scale (Benton et al., 2003; Vickery and Arlettaz, 2012; Barbaro et al., 2021; Tscharntke et al., 2021), we provide here evidence that it is beneficial also at the field scale.
We did not find any significant differences between organic and biodynamic viticulture for both epiphytic and ground-dwelling arthropods, contrary to our prediction (H2) drawn from the intermediate disturbance hypothesis. Neither did we find a difference in the abundance of ground-dwelling invertebrates between biodynamic and conventional vineyards, what would be in line with that hypothesis given that these two regimes can entail frequent disturbances, via ploughing (biodynamic) and intensive use of herbicides (conventional), both annihilating ground vegetation on a wide fraction of the vineyard surface (Mackey and Currie, 2001). Organic manages the ground vegetation mostly via mowing operations, and more rarely by ploughing a restricted fraction of the ground, leading to variegated habitat heterogeneity on a small spatial scale because here the disturbance doesn’t impact the entire soil surface. Optimal micro-habitat conditions for biodiversity may therefore be achieved by the shallower management of the soil that typically characterizes organic grape production, whereas the other two regimes may provide micro-habitats that are too homogeneous in space and time.
Ground Vegetation Management and Farming Regimes Are Intertwined
Even though we found that ground vegetation cover boosts the populations of epiphytic invertebrates (but not significantly concerning ground-dwelling invertebrates), the greater abundances of invertebrates in organic vs. conventional vineyards persisted after accounting for the effects of ground vegetation. This clearly indicates the existence of other underlying co-factors, beyond the mere soil and ground vegetation disturbance effects described above. Those may of course be the lethal or sublethal effects of agrochemicals (Trivellone et al., 2012; Masoni et al., 2017; Karimi et al., 2020), which chiefly depend on the management regime, but also more subtle qualitative differences in the composition of the plant communities that typically accompany the three management regimes (e.g., the ratio of herbs:grasses) and may entail various complexities of vegetation structure and/or floral resources (Puig-Montserrat et al., 2017; Winter et al., 2018). In general, however, the positive influence of increased ground vegetation cover confirms that a good field layer promotes not only overall invertebrate abundance (Bosco et al., 2018; Bosco et al., 2019; Saenz-Romo et al., 2019) and their insectivorous predators (Arlettaz et al., 2012; Guyot et al., 2017; Bosco et al., 2019; Bosco et al., 2020), but also key ecosystem service providers such as pest control agents (Thomson and Hoffmann, 2009; Saenz-Romo et al., 2019) and pollinating insects (Winter et al., 2018; Maurer et al., 2020).
The best fitting models for any response variables were the interactive ones, with a significant interaction between management regime and ground vegetation cover for epiphytic invertebrates, hence confirming our hypothesis H3. For epiphytic arthropods, the extent of ground vegetation cover induces a simple positive linear effect in organic, a ∪-shaped effect in biodynamic and a ∩-shaped effect in conventional vineyards. These differences in the trajectories indicate that an increase in ground greening does not per se lead to improved biodiversity on a field-scale but that the underlying management (including both ground vegetation and chemical inputs) needs to be considered. Ground vegetation and below-ground community composition is known to be sensitive not only to management-induced changes in carbon, nitrogen, and phosphorus cycles, partly due to mowing regime, but also to pesticide application (Reeve et al., 2005; Celette et al., 2009; Nascimbene et al., 2013; Fried et al., 2019; Karimi et al., 2020). Given that such factors differ considerably among the management regimes it is comprehensible that biodiversity patterns as seen in our study vary between conventional, organic, and biodynamic management. Contrary to epiphytic invertebrates, ground-dwelling invertebrates do not seem to respond to a similar extent to ground vegetation cover as there were hardly any differences in additive or interactive models. Their abundance seems thus mostly driven by the agricultural regime.
Indicator Groups
As predicted (H4), different invertebrate groups were specifically associated with the three management regimes and with the coverage of the ground vegetation layer. Among epiphytic arthropods, heteropterans, hymenopterans and dipterans were linked to organic farming, indicating that this regime favors in particular pollinating species (Rundlöf et al., 2008). Not surprisingly, Araneae emerged as an indicator group of biodynamic management; in effect, regular ploughing creates a very mineral substrate that is typically appreciated by spiders.
Among ground-dwelling invertebrates, on the other hand, centipedes (Chilopoda) were linked to conventional management, matching their ecology of generalist, free-ranging predators inhabiting mineral soils and the litter layer (Klarner et al., 2017). The suborder Sternorrhyncha (mainly aphids) and Isopoda were associated with biodynamic management, likely reflecting the dual habitat conditions prevailing in biodynamic fields: vegetated inter-rows with usually tall vegetation, likely promoting aphids, and ploughed and/or mulched inter-rows, favouring Isopods. Cicadas and dipterans were associated with organic farming, again likely due to a greater availability of floral resources (dipterans) and the presence of specific host plant species (cicadas).
Conclusions
This study not only provides new insights into the implications of the three dominant viticultural practices for above-ground animal biodiversity, but it is also the first, to the best of our knowledge, that compares the arthropod fauna of organic and biodynamic viticulture. The conclusion that organic, to an even greater extent than biodynamic management, supersedes conventional management from a biodiversity viewpoint provides novel arguments in the debate about which farming practice is most beneficial for the balance of nature. Even though ground vegetation appears to be a key factor for boosting biodiversity, additional underlying co-factors linked to the three management regimes further contribute to shaping vineyard invertebrate communities. However, our results must be interpreted context-specifically since we did not consider more detailed differences between the management regimes such as vegetation composition and diversity, precise soil management (e.g. the intensity of mowing, ploughing or grazing), quantity of pesticides applied and year effects. Nevertheless, as invertebrates constitute a staple commodity for many vertebrates situated higher up along the food chain, these findings have far-reaching implications for ecological functions, ecosystem services, and provide basic guidance for an environmentally more sustainable viticulture into the future. Whether these conclusions can be generalized to other types of agro-ecosystems, other organisms and other spatial scales deserves further investigations.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author Contributions
RA, DS, and LR designed and planned the study with support from AJ and LB. DS and LR did the field and lab work. LB, DS, and LR analyzed the data. DS and LB lead the writing of the manuscript. All authors contributed critically to the drafts and gave final approval for publication.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank all viticulturists for their collaboration, expert knowledge, and the free access to their ground for this project.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2022.837551/full#supplementary-material
References
Arlettaz R., Maurer M. L., Mosimann-Kampe P., Nusslé S., Abadi F., Braunisch V., et al. (2012). New Vineyard Cultivation Practices Create Patchy Ground Vegetation, Favouring Woodlarks. J. Ornith. 153 (1), 229–238. doi: 10.1007/s10336-011-0737-7
Assandri G., Bogliani G., Pedrini P., Brambilla M. (2016). Diversity in the Monotony? Habitat Traits and Management Practices Shape Avian Communities in Intensive Vineyards. Agric. Ecosys. Environ. 223, 250–260. doi: 10.1016/j.agee.2016.03.014
Barbaro L., Assandri G., Brambilla M., Castagneyrol B., Froidevaux J., Giffard B., et al. (2021). Organic Management and Landscape Heterogeneity Combine to Sustain Multifunctional Bird Communities in European Vineyards. J. Appl. Ecol. 58 (6), 1261–1271. doi: 10.1111/1365-2664.13885
Bates D, Mächler M, Bolker B, Walker S (2005). “Fitting Linear Mixed-Effects Models Using lme4.” in Journal of Statistical Software, 67(1), 1–48. doi: 10.18637/jss.v067.i01
Benton T. G., Vickery J. A., Wilson J. D. (2003). Farmland Biodiversity: Is Habitat Heterogeneity the Key? Trends Ecol. Evol. 18 (4), 182–188. doi: 10.1016/S0169-5347(03)00011-9
Bosco L., Arlettaz R., Jacot A. (2019). Ground Greening in Vineyards Promotes the Woodlark Lullula Arborea and Their Invertebrate Prey. J. Ornith. 160 (3), 799–811. doi: 10.1007/s10336-019-01666-7
Bosco L., Cushman S. A., Wan H. Y., Zeller K. A., Arlettaz R., Jacot A. (2020). Fragmentation Effects on Woodlark Habitat Selection Depend on Habitat Amount and Spatial Scale. Anim. Conserv 24(1), 84–94. doi: 10.1111/acv.12604
Bosco L., Wan H. Y., Cushman S. A., Arlettaz R., Jacot A. (2018). Separating the Effects of Habitat Amount and Fragmentation on Invertebrate Abundance Using a Multi-Scale Framework. Landsc. Ecol. 34 (1), 105–117. doi: 10.1007/s10980-018-0748-3
Bruggisser O. T., Schmidt-Entling M. H., Bacher S. (2010). Effects of Vineyard Management on Biodiversity at Three Trophic Levels. Biol. Conserv. 143 (6), 1521–1528. doi: 10.1016/j.biocon.2010.03.034
Cáceres M. D., Legendre P. (2009). Associations Between Species and Groups of Sites: Indices and Statistical Inference. Ecol. 90 (12), 3566–3574. doi: 10.1890/08-1823.1
Caprio E., Nervo B., Isaia M., Allegro G., Rolando A. (2015). Organic Versus Conventional Systems in Viticulture: Comparative Effects on Spiders and Carabids in Vineyards and Adjacent Forests. Agric. Syst. 136, 61–69. doi: 10.1016/j.agsy.2015.02.009
Caprio E., Rolando A. (2017). Management Systems may Affect the Feeding Ecology of Great Tits Parus Major Nesting in Vineyards. Agric. Ecosys. Environ. 243, 67–73. doi: 10.1016/j.agee.2017.03.013
Carpenter-Boggs L., Kennedy A. C., Reganold J. P. (2000). Organic and Biodynamic Management Effects on Soil Biology. Soil Sci. Soc Am. J. 64 (5), 1651–1659. doi: 10.2136/sssaj2000.6451651x
Celette F., Findeling A., Gary C. (2009). Competition for Nitrogen in an Unfertilized Intercropping System: The Case of an Association of Grapevine and Grass Cover in a Mediterranean Climate. Eu. J. Agron. 30 (1), 41–51. doi: 10.1016/j.eja.2008.07.003
Core Team R. (2020). "R: A Language and Environment for Statistical Computing" (Vienna, Austria: The R Foundation for Statistical Computing).
Döring J., Collins C., Frisch M., Kauer R. (2019). Organic and Biodynamic Viticulture Affect Biodiversity and Properties of Vine and Wine: A Systematic Quantitative Review. Am. J. Enol. Vitic. 70 (3), 221–242. doi: 10.5344/ajev.2019.18047
Döring J., Frisch M., Tittmann S., Stoll M., Kauer R. (2015). Growth, Yield and Fruit Quality of Grapevines Under Organic and Biodynamic Management. PloS One 10 (10), e0138445. doi: 10.1371/journal.pone.0138445
Fried G., Cordeau S., Metay A., Kazakou E. (2019). Relative Importance of Environmental Factors and Farming Practices in Shaping Weed Communities Structure and Composition in French Vineyards. Agric. Ecosys. Environ. 275, 1–13. doi: 10.1016/j.agee.2019.01.006
Gabriel D., Roschewitz I., Tscharntke T., Thies C. (2006). Beta Diversity at Different Spatial Scales: Plant Communities in Organic and Conventional Agriculture. Ecol. Appl. 16 (5), 2011–2021. doi: 10.1890/1051-0761(2006)016[2011:BDADSS]2.0.CO;2
Guyot C., Arlettaz R., Korner P., Jacot A. (2017). Temporal and Spatial Scales Matter: Circannual Habitat Selection by Bird Communities in Vineyards. PloS One 12 (2), e0170176. doi: 10.1371/journal.pone.0170176
Hole D. G., Perkins A. J., Wilson J. D., Alexander I. H., Grice P. V., Evans A. D. (2005). Does Organic Farming Benefit Biodiversity? Biol. Conserv. 122 (1), 113–130. doi: 10.1016/j.biocon.2004.07.018
Hothorn T., Bretz F., Westfall P. (2008). Simultaneous Inference in General Parametric Models. Biometr. J. 50 (3), 346–363. doi: 10.1002/bimj.200810425
Karimi B., Cahurel J. Y., Gontier L., Charlier L., Chovelon M., Mahé H., et al. (2020). A Meta-Analysis of the Ecotoxicological Impact of Viticultural Practices on Soil Biodiversity. Environ. Chem. Lett. 18, 1947–1966. doi: 10.1007/s10311-020-01050-5
Klarner B., Winkelmann H., Krashevska V., Maraun M., Widyastuti R., Scheu S. (2017). Trophic Niches, Diversity and Community Composition of Invertebrate Top Predators (Chilopoda) as Affected by Conversion of Tropical Lowland Rainforest in Sumatra (Indonesia). PloS One 12 (8), e0180915. doi: 10.1371/journal.pone.0180915
Lampkin N. (1999). “Organic Farming in the European Union–overview, Policies and Perspectives,” in Organic Frarming in the European Union-Perspectives for the 21st Century (Baden/Vienna: Proc. EU Commission/Austrian Governmnet Conference), 23–30.
Lüdecke D, Ben-Shachar M, Patil I, Waggoner P, Makowski D (2021). “performance: An R Package for Assessment, Comparison and Testing of Statistical Models.,” Journal of Open Source Software, 6(60), 3139. doi:10.21105/joss.03139.
Mackey R. L., Currie D. J. (2001). The Diversity–Disturbance Relationship: Is it Generally Strong and Peaked? Ecol. 82 (12), 3479–3492. doi: 10.1890/0012-9658(2001)082[3479:TDDRII]2.0.CO;2
Masoni A., Frizzi F., Brühl C., Zocchi N., Palchetti E., Chelazzi G., et al. (2017). Management Matters: A Comparison of Ant Assemblages in Organic and Conventional Vineyards. Agric. Ecosys. Environ. 246, 175–183. doi: 10.1016/j.agee.2017.05.036
Maurer C., Bosco L., Klaus E., Cushman S. A., Arlettaz R., Jacot A. (2020). Habitat Amount Mediates the Effect of Fragmentation on a Pollinator's Reproductive Performance, But Not on its Foraging Behaviour. Oecol. 193 (3), 523–534. doi: 10.1007/s00442-020-04658-0
Maxwell S. L., Fuller R. A., Brooks T. M., Watson J. E. (2016). Biodiversity: The Ravages of Guns, Nets and Bulldozers. Nat. News 536(7615), 143. doi: 10.1038/536143a
Morrison-Whittle P., Lee S. A., Goddard M. R. (2017). Fungal Communities are Differentially Affected by Conventional and Biodynamic Agricultural Management Approaches in Vineyard Ecosystems. Agric. Ecosys. Environ. 246, 306–313. doi: 10.1016/j.agee.2017.05.022
Munoz-Saez A., Kitzes J., Merenlender A. M. (2021). Bird-Friendly Wine Country Through Diversified Vineyards. Conserv. Biol 35 (1), 274–284. doi: 10.1111/cobi.13567
Nascimbene J., Marini L., Ivan D., Zottini M. (2013). Management Intensity and Topography Determined Plant Diversity in Vineyards. PloS One 8 (10), e76167. doi: 10.1371/journal.pone.0076167
Nascimbene J., Marini L., Paoletti M. G. (2012). Organic Farming Benefits Local Plant Diversity in Vineyard Farms Located in Intensive Agricultural Landscapes. Environ. Manage. 49 (5), 1054–1060. doi: 10.1007/s00267-012-9834-5
Norris R. F., Kogan M. (2000). Interactions Between Weeds, Arthropod Pests, and Their Natural Enemies in Managed Ecosystems. Weed Sci. 48 (1), 94–158. doi: 10.1614/0043-1745(2000)048[0094:IBWAPA]2.0.CO;2
Petrescu Bakış A.-L., Macovei I., Barros P., Gomes C., Carvalho D., Cabral J. A., et al. (2021). Is Biodiversity Linked With Farm Management Options in Vineyard Landscapes? A Case Study Combining Ecological Indicators Within a Hybrid Modelling Framework. Ecol. Indic 121, 1–12. doi: 10.1016/j.ecolind.2020.107012
Phillips J. C., Rodgriguez L. P. (2006). “Beyond Organic: An Overview of Biodynamic Agriculture With Case Examples,” in Agricultural and Applied Economics Association (AAEA) Conference, 2006 Annual meeting. Long Beach, CA, USA. doi: 10.22004/ag.econ.21036
Provost C., Pedneault K. (2016). The Organic Vineyard as a Balanced Ecosystem: Improved Organic Grape Management and Impacts on Wine Quality. Scient. Hortic. 208, 43–56. doi: 10.1016/j.scienta.2016.04.024
Puig-Montserrat X., Stefanescu C., Torre I., Palet J., Fàbregas E., Dantart J., et al. (2017). Effects of Organic and Conventional Crop Management on Vineyard Biodiversity. Agric. Ecosys. Environ. 243, 19–26. doi: 10.1016/j.agee.2017.04.005
Reeve J. R., Carpenter-Boggs L., Reganold J. P., York A. L., McGourty G., McCloskey L. P. (2005). Soil and Winegrape Quality in Biodynamically and Organically Managed Vineyards. Am. Enol. Vitic. 56 (4), 367–376.
Rockström J., Steffen W., Noone K., Persson Å, Chapin F. S. III, Lambin E., et al. (2009). Planetary Boundaries: Exploring the Safe Operating Space for Humanity. Ecol. Soc 14 (2), 1–33. http://www.ecologyandsociety.org/vol14/iss2/art32/
Roschewitz I., Gabriel D., Tscharntke T., Thies C. (2005). The Effects of Landscape Complexity on Arable Weed Species Diversity in Organic and Conventional Farming. J. Appl. Ecol. 42 (5), 873–882. doi: 10.1111/j.1365-2664.2005.01072.x
Rundlöf M., Nilsson H., Smith H. G. (2008). Interacting Effects of Farming Practice and Landscape Context on Bumble Bees. Biol. Conserv. 141 (2), 417–426. doi: 10.1016/j.biocon.2007.10.011
Saenz-Romo M. G., Veas-Bernal A., Martinez-Garcia H., Ibanez-Pascual S., Martinez-Villar E., Campos-Herrera R., et al. (2019). Effects of Ground Cover Management on Insect Predators and Pests in a Mediterranean Vineyard. Insects 10 (12), 1–16. doi: 10.3390/insects10120421
Sanguankeo P., León R. (2011). Weed Management Practices Determine Plant and Arthropod Diversity and Seed Predation in Vineyards. Weed Res. 51 (4), 404–412. doi: 10.1111/j.1365-3180.2011.00853.x
Sierro A., Arlettaz R. (2003). L’avifaune Du Vignoble En Valais Central: Évaluation De La Diversité À L’aide De Transects. Nos Oiseaux 50 (2), 89–100.
Thomson L. J., Hoffmann A. A. (2009). Vegetation Increases the Abundance of Natural Enemies in Vineyards. Biol. Contr. 49 (3), 259–269. doi: 10.1016/j.biocontrol.2009.01.009
Trivellone V., Paltrinieri L. P., Jermini M., Moretti M. (2012). Management Pressure Drives Leafhopper Communities in Vineyards in Southern Switzerland. Insect Conserv. Divers. 5 (1), 75–85. doi: 10.1111/j.1752-4598.2011.00151.x
Tscharntke T., Grass I., Wanger T. C., Westphal C., Batáry P. (2021). Beyond Organic Farming – Harnessing Biodiversity-Friendly Landscapes. Trends Ecol. Evol 36(10), 919–930. doi: 10.1016/j.tree.2021.06.010
Tscharntke T., Tylianakis J. M., Rand T. A., Didham R. K., Fahrig L., Batáry P., et al. (2012). Landscape Moderation of Biodiversity Patterns and Processes-Eight Hypotheses. Biol. Rev. 87 (3), 661–685. doi: 10.1111/j.1469-185X.2011.00216.x
Turinek M., Grobelnik-Mlakar S., Bavec M., Bavec F. (2009). Biodynamic Agriculture Research Progress and Priorities. Renew. Agric. Food Syst. 24 (2), 146–154. doi: 10.1017/s174217050900252x
Vickery J., Arlettaz R. (2012). “The Importance of Habitat Heterogeneity at Multiple Scales for Birds in European Agricultural Landscapes,” in Birds and Habitat: Relationships in Changing Landscapes. Ed. Fuller R. J. (Cambridge: Cambridge University Press), 177–204.
Vogt G. (2007). “The Origins of Organic Farming,” in Organic Farming: An International History, 9–29. London, UK
Winter S., Bauer T., Strauss P., Kratschmer S., Paredes D., Popescu D., et al. (2018). Effects of Vegetation Management Intensity on Biodiversity and Ecosystem Services in Vineyards: A Meta-Analysis. J. Appl. Ecol. 55 (5), 2484–2495. doi: 10.1111/1365-2664.13124
Keywords: agroecology, ground vegetation cover, invertebrate abundance, soil management, vineyards
Citation: Bosco L, Siegenthaler D, Ruzzante L, Jacot A and Arlettaz R (2022) Varying Responses of Invertebrates to Biodynamic, Organic and Conventional Viticulture. Front. Conserv. Sci. 3:837551. doi: 10.3389/fcosc.2022.837551
Received: 16 December 2021; Accepted: 23 June 2022;
Published: 15 July 2022.
Edited by:
James Bullock, UK Centre for Ecology and Hydrology (UKCEH), United KingdomReviewed by:
Mauro Fois, University of Cagliari, ItalyJosé M. Rey Benayas, University of Alcalá, Spain
Copyright © 2022 Bosco, Siegenthaler, Ruzzante, Jacot and Arlettaz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Laura Bosco, bGF1cmEuYm9zY29AaGVsc2lua2kuZmk=