 David García de León
David García de León José M. Rey Benayas
José M. Rey Benayas Enrique Andivia
Enrique Andivia
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Conserv. Sci. , 22 December 2021
Sec. Plant Conservation
Volume 2 - 2021 | https://doi.org/10.3389/fcosc.2021.789612
This article is part of the Research Topic Balancing Biodiversity Conservation and Food Production in European Agroecosystems View all 5 articles
Hedgerows are linear landscape features of woody vegetation usually located around agricultural fields. An increasing number of studies have addressed the effects of hedgerows on biodiversity and ecosystem services. This study is aimed to synthesize these effects and compare the levels of biodiversity and ecosystem services in farmland with hedgerows and (1) farmland without hedgerows and (2) nearby natural habitat at the global scale. We hypothesized that farmland with hedgerows (1) enhances biodiversity and ecosystem services as compared to farmland without hedgerows but (2) supports lower levels of biodiversity and ecosystem services than natural habitat. Our systematic literature review retained 835 observations from 170 primary studies, which were analyzed following the standard methodology in meta-analyses. Our results partially support both hypotheses. Farmland with hedgerows exhibited higher levels of biodiversity and provisioning services than farmland without hedgerows (H1). Farmland with hedgerows provided similar levels of biodiversity (edge effects) but lower levels of ecosystem services than natural habitat (H2). The effects of hedgerows on biodiversity and ecosystem services depended on control ecosystem type (grassland/meadow or forest/woodland) but were largely independent of climate type (temperate or tropical) and the focus of spatial scale (field or landscape). In conclusion, conservation and restoration of hedgerows contribute to people in several ways by enhancing biodiversity and multifunctionality in agricultural landscapes.
Agricultural production must be more sustainable if the society wants to redress the major source of direct and indirect environmental impacts on the planet, particularly biodiversity loss and climate change (IPBES, 2017, 2019; IPBES and IPCC, 2021). To achieve this goal, the EU Biodiversity Strategy for 2030 aims to convert 10% of farmland into high-diversity landscape features (European Commission, 2020), as species-rich semi-natural habitat and natural habitat remnants play a key role in maintaining biodiversity and a range of ecosystem services in agricultural landscapes (Jahnová et al., 2016; Dainese et al., 2017). High-diversity features in agricultural landscapes include a wide range of non-cropped habitats such as ponds, herbaceous and woody buffer strips, forest patches, and hedgerows (Rey Benayas and Bullock, 2012).
Hedgerows are linear landscape features of woody vegetation usually located around agricultural fields (Lajos et al., 2020). They can be either spontaneous (naturally grown) or planted. Spontaneous hedgerows reflect, to some extent, the woody community composition of remnant forests in a particular region (Rey Benayas et al., 2020). Hedgerows harbor a high diversity of plants (Freemark et al., 2002; Aavik et al., 2008) in terms of both plant abundance (Aavik et al., 2008) and richness (Evans et al., 2011). They can function as plant colonization sources (Endels et al., 2004; Rey Benayas and Bullock, 2015) and modulate plant community composition, which benefit native plants against exotic plants (Freemark et al., 2002).
Other species benefit from hedgerows, including small mammals such as mice (Fialho et al., 2019) and bats (Vandevelde et al., 2014; Heim et al., 2018). Large mammals such as the roe deer (Capreolus capreolus) and the Iberian wolf (Canis lupus) find hedgerows a substitutable habitat for woodland (Rodríguez-Freire and Crecente-Maseda, 2008; Morellet et al., 2011). They provide nest sites (Hotaling et al., 2018), food (Evans et al., 2003; Barbaro et al., 2008), or shelter (Ceresa et al., 2012) for many bird species (Burgess et al., 2015). Invertebrates such as less-common species of insects (Morandin and Kremen, 2013; Ponisio et al., 2016) and spiders find hedgerows a valuable habitat as well (Wu et al., 2009; Mestre et al., 2018), as they provide relatively stable microenvironmental conditions that allow them to overcome adverse changes in humidity, temperature, or wind, and recolonize other habitats when favorable conditions return. Butterflies and moths are subjected to complex effects of hedgerows (Berwaerts et al., 1998; Feber et al., 2007) as crops with hedgerows might or might not exhibit richer and more abundant communities of these taxa than crops without hedgerows; however, forests clearly benefit these taxa (Ouin and Burel, 2002). Also, Dainese et al. (2015) concluded that hedgerows result in higher species richness of both butterflies and parasitoid flies, with butterflies showing a significantly higher species richness in complex hedgerows.
Hedgerows provide multiple ecosystem services, some of which may directly enhance food and forage production through shelter (Hauser, 2008), pest regulation (Koellner and Scholz, 2008; Pfister et al., 2017), pollination (Sardiñas and Kremen, 2015; Byrne and Del Barco-Trillo, 2019), soil water content (Luptáčik et al., 2012; Wolka et al., 2018; Holden et al., 2019), and soil quality (Monokrousos et al., 2006). For instance, Van Vooren et al. (2017) found that hedgerows in temperate areas intercepted nitrogen from the surface and subsurface flow and phosphorus and soil sediment from the surface flow that increased the crop yield.
It has been proposed that hedgerows are an intermediate habitat between crops and forests in terms of biodiversity and provision of ecosystem services (Slade et al., 2013) and that a well-developed hedgerow network may be seen as a form of forest in agricultural landscapes (Rey Benayas and Bullock, 2015). Despite their numerous advantages, hedgerows also portrait some drawbacks (Capotorti et al., 2019). They occupy some land, make machinery mobility more difficult, and may spread some pests (Barr et al., 1995). Hedgerows by roadside margins may increase the mortality of some species such as mammals and birds (OrlŁowski, 2010). Recent review studies that have addressed the overall effect of hedgerows on biodiversity and ecosystem services have found contrasting results for individual taxa or ecosystem services (Van Vooren et al., 2017; Montgomery et al., 2020; Albrecht et al., 2021). For instance, the increase of ecosystem services such as pest regulation provided by hedgerows seems to be, overall, nonsignificant (Montgomery et al., 2020); however, Van Vooren et al. (2017) found an increase in natural enemy species richness. On the other hand, there is a lack of primary studies on individual ecosystem services other than pest regulation, pollination, and water regulation (Albrecht et al., 2021).
It is likely that pastures, herbaceous crops, woody crops, grasslands, and meadows without hedgerows would behave differently if hedgerows were planted around them (Jahnová et al., 2016; Timberlake et al., 2019). For instance, crop type seems to be a significant predictor of native species richness (Boutin et al., 2008). In addition, different climate types (Pinkus-Rendón et al., 2006) and focus of the spatial scale (e.g., field vs. landscape scale) might potentially have contrasting benefits by the presence of hedgerows as well (Freemark et al., 2002; Aavik et al., 2008).
This study is aimed to synthesize the published scientific evidence on the effects of hedgerows on local biodiversity and provision of ecosystem services and compare the levels of such biodiversity and ecosystem services provided by hedgerows and nearby natural habitats (mostly forest and grassland) at the global scale. We hypothesized that hedgerows (1) enhance contributions of nature to people in agricultural landscapes but (2) support lower levels of biodiversity and ecosystem services than natural habitat. We expect that this study provides further support for the need for widespread hedgerow restoration and more wildlife-friendly farming and multifunctional landscapes.
On July 25, 2019, we performed a scientific literature search on Web of Science for years 1969 to 2019 with the following combination of keywords in the title or the abstract: [(hedgerow* OR “living fence*” OR “woody field margin” OR “woody edge” OR “woody buffer strip”) AND (benefit* OR *diversity OR function* OR service*)] and these words in Spanish, namely, [(“cerca* viva*” OR seto* OR “borde leñoso” OR “margen leñoso” OR “franja leñosa”) AND (beneficio* OR *diversidad OR funcion* OR servicio*)]. This search identified 985 studies that might report quantitative evidence of levels of biodiversity and/or ecosystem services rendered by hedgerows.
One co-author read the abstracts of all 985 studies. After reading these abstracts, 294, 314, and 377 articles were deemed as useful, possibly useful, or non-useful, respectively, in terms of including quantitative information related to biodiversity or ecosystem services in hedgerows and paired control ecosystems. The 608 useful and possibly useful studies were carefully revised to identify and extract relevant data; this task was performed by the same author in the interest of consistency for data gathering. We finally extracted data from 294 studies (detailed PRISMA flow diagram in Supplementary Figure 1).
A database was built to systematize the extracted information of the following variables for each article: article's identifier, reference of the study, response variables (indicators), units of response variables, effect sign (direct or inverse), taxa (see below), type of ecosystem service (see below), sample size, response value at farmland with hedgerows and control ecosystems (namely, farmland without hedgerows or nearby natural habitat), variation and type of variation (variance, standard deviation (SD), standard error, interquartile range, confidence interval) of the response value, farmland type (crop or pasture), crop type (herbaceous or woody), natural habitat type (forest/woodland or grassland/meadow), country, climate type (tropical or temperate), focus of the spatial scale (field or landscape), overall effect of hedgerows on biodiversity and ecosystem services (positive, neutral, or negative), and type of inference for this overall effect.
Once completed, another coauthor independently revised the database to double-check the procedure and detect possible inconsistencies, errors, and noticeable weird information. This revision corrected the signs of many relationships and excluded 124 studies: 81 of them because they did not report the variability of response variables (i.e., a robust effect size measure could not be used with these articles) and 43 because they referred to non-woody margins. Appendix 1 lists the 170 primary studies that provided data that were finally analyzed. The response variables were linked to the taxonomic groups (biodiversity) plants, invertebrates, and vertebrates, and one of the four categories of ecosystem services according to the Millennium Ecosystem Assessment (MEA, 2005), namely, supporting, regulating, provisioning, and cultural ecosystem services. Farmland type, crop type, natural habitat type, climate type, and focus of the spatial scale were used as context factors (moderators) of hedgerow effects.
We analyzed the database following standard methodologies in meta-analyses (Koricheva et al., 2013). We split the database into four datasets for further analyses. The first and second datasets allowed to compare the biodiversity levels between farmland with hedgerows and farmland without hedgerows (Data File 1; R script BioHEFA) and between farmland with hedgerows and natural habitat (Data File 2; R script BioHENH). The third and fourth datasets allowed to compare the ecosystem services between farmland with hedgerows and farmland without hedgerows (Data File 3; R script ESHEFA) and between farmland with hedgerows and natural habitat (Data File 4; R script ESHENH). As using the broad ecosystem-service categories might result in lumping together cases that clearly have opposite signs, which will level out the overall response and reduce the likelihood of significant findings, we performed supplementary analyses on the ecosystem services with, at least, three primary studies and ten observations (Table 1; Data Files 5, 6; R scripts ESHEFA_specific, ESHENH_specific).
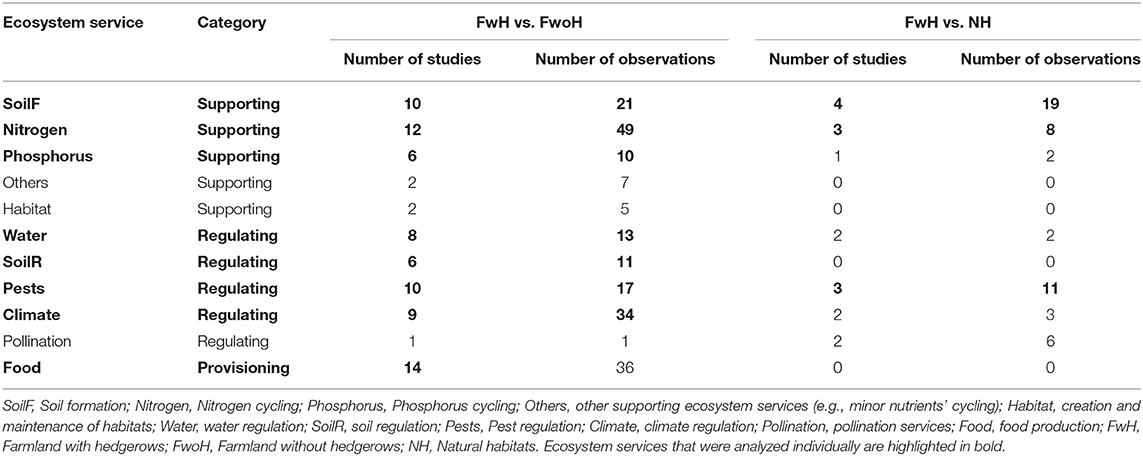
Table 1. Number of primary studies and observations providing data for specific ecosystem services.
Standardized mean difference (SMD) was computed as a robust estimate of effect size of farmland with hedgerows vs. control ecosystems using the escalc function in metafor R package (Viechtbauer, 2010). The sign of SMD was changed if an effect in each observation was based on an inverse relationship (e.g., invasive species and di-services). The SMD is commonly used to estimate the magnitude of the effect by comparing an experimental group and a control group (Hedges and Olkin, 1986; Gurevitch et al., 2001).
A null mixed model adjusted for meta-analysis was fit (Borenstein et al., 2009) to assess whether farmland with hedgerows showed different biodiversity or ecosystem services levels from either farmland without hedgerows or natural habitat. By using a mixed model, we assumed that each case study was a random sample of a larger overall population and considered the primary study (article) as a nesting factor to avoid violating the assumption that effect sizes are independent from each other. We evaluated the extent of heterogeneity by using the statistic I2 (Higgins and Thompson, 2002), which estimates the percentage of variability due to heterogeneity rather than due to the sampling error.
We also evaluated if moderators explained a significant part of the heterogeneity in the true effect by using simple meta-regressions. Moderators were farmland type (crop or pasture), crop type (herbaceous or woody), natural habitat type (forest/woodland or grassland/meadow), climate type (tropical or temperate), and focus of the spatial scale (field or landscape). Finally, we evaluated if the difference between the effect sizes of factor levels was significantly different from zero by conducting an omnibus test based on a chi-square distribution with the number of coefficients tested as degrees of freedom and following default settings. All statistical analyses were performed in metafor R package (R Core Team, 2021).
Data filtering retained 835 case studies from 170 primary studies conducted in 39 countries of all continents but Antarctica (Figure 1; Data File 7; script Figure 1). Most records (70% of observations and 74% of primary studies) were related to biodiversity (abundance = 38 and 41%; diversity = 31 and 32%; plants = 35 and 25%; invertebrates = 21 and 23%; vertebrates = 14 and 16%), followed by supporting services (16%, 20%), regulating services (13%, 26%), provisioning services (5%, 15%), and cultural services (<1%, excluded for further analyses due to low sample size). The number of observations and primary studies was as follows: crop = 64 and 56%, pasture = 4 and 7%, herbaceous crop = 54 and 42%, woody crop = 17 and 16%, forest/woodland habitat = 29 and 24%, grassland/meadow habitat = 3 and 5%, tropical climate = 22 and 21%, temperate climate = 78 and 72%, field scale = 67 and 49%, and landscape scale = 33 and 29% (see Table 2 for absolute numbers).
Figure 1. World map showing the countries where the selected primary studies were run.
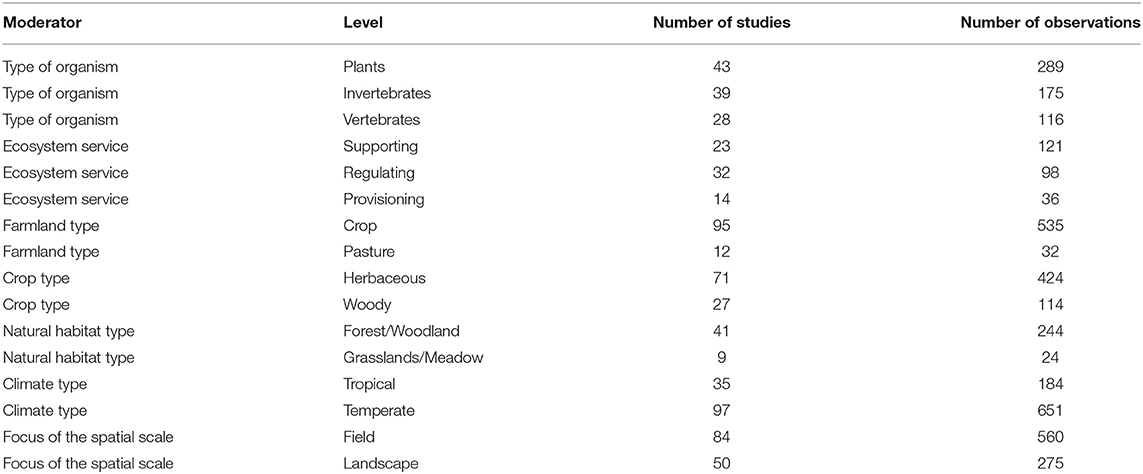
Table 2. Number of studies and observations per level of each moderator and context factor.
The effect of farmland with hedgerows as compared to farmland without hedgerows on biodiversity was positive, negative, and neutral in 243, 107, and 13 observations, respectively (N = 363 observations from 72 primary studies). Meta-analysis showed significantly higher biodiversity levels in farmland with hedgerows than in farmland without hedgerows (Figure 2; estimated effect size = 0.64 ± 0.18, p < 0.01). Heterogeneity was very high (I2 95% CI: 99.6–99.8%), suggesting that most heterogeneity not explained by sampling error and thus justifying the inclusion of moderators in the model.
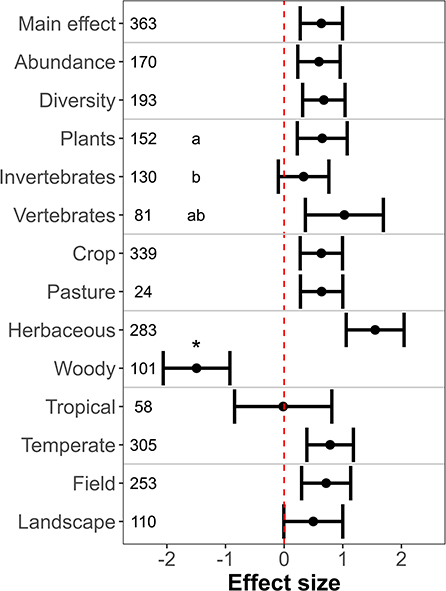
Figure 2. Biodiversity levels in farmland with hedgerows as compared to farmland without hedgerows. X-axis indicates the mean effect size (standardized mean difference) ± 95% confidence interval. The dashed vertical red line shows the null effect. A confidence interval that overlaps this line means that the effects are not significant. Significant differences between levels of moderators are indicated by asterisks (two levels) and different letters (three levels), respectively.
Farmland with hedgerows was equally beneficial for both abundance and diversity metrics (Figure 2; QM = 3.24, p = 0.07). Type of organism explained a significant part of the heterogeneity of effect sizes (Figure 2); while plants were more positively affected by hedgerows than invertebrates (QM = 7.69, p = 0.01), enhancement of vertebrate biodiversity was similar to that of plants (QM = 0.96, p = 0.33) and invertebrates (QM = 3.19, p = 0.07).
Hedgerows enhanced biodiversity levels in both cropped farmland and pastures, with no significant differences between these two farmland types (Figure 2; QM = 0.13, p = 0.72). However, hedgerows did not show a positive effect on biodiversity for crop types; hedgerows enhanced biodiversity on herbaceous crops, but the biodiversity of woody crops was higher without hedgerows (Figure 2; QM = 186.88, p < 0.01). Farmland with hedgerows supported higher biodiversity than farmland without hedgerows in temperate regions only (Figure 2; estimated effect size: temperate = 0.78 ± 0.20, p < 0.01; tropical = −0.02 ± 0.42, p = 0.97), and the difference between tropical and temperate regions was not significant (Figure 2; QM = 2.91, p = 0.09). Likewise, both field and landscape studies were equally positively benefitted by hedgerows (Figure 2; QM = 0.61, p = 0.43), but hedgerows effects at the landscape scale were barely significant.
The effect of farmland with hedgerows as compared to natural habitat on biodiversity was negative, positive, and neutral in 116, 92, and 9 observations, respectively (N = 217 observations from 43 primary studies). Consequently, meta-analysis revealed no significant differences between farmland with hedgerows and natural habitat in terms of biodiversity (Figure 3; estimated effect size = 0.11 ± 0.14, p = 0.43; I2 95% CI: 87.70–95.16%).
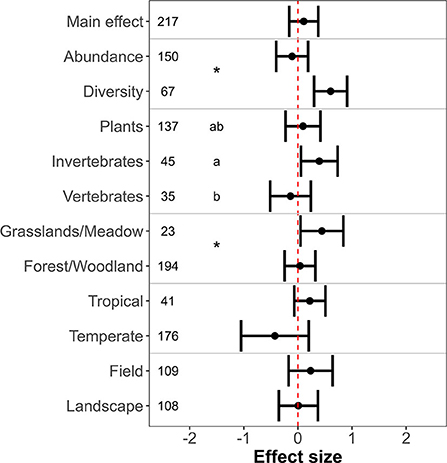
Figure 3. Biodiversity levels in farmland with hedgerows as compared to natural habitat. X-axis indicates the mean effect size (standardized mean difference) ± 95% confidence interval. The dashed vertical red line shows the null effect. A confidence interval that overlaps this line means that the effects are not significant. Significant differences between levels of moderators are indicated by asterisks (two levels) and different letters (three levels), respectively.
There were significant differences in the effect of hedgerows between biodiversity metrics, organism types, and natural habitat type (Figure 3). Abundance of organisms did not differ between farmland with hedgerows and natural habitat, but diversity was higher in farmland with hedgerows (Figure 3; QM = 128.78, p < 0.01). Only invertebrates exhibited higher biodiversity levels in farmland with hedgerows than in natural habitat (Figure 3; estimated effect size = 0.39 ± 0.17, p = 0.02). Biodiversity differed between natural habitat types; farmland with hedgerows had a higher level than grasslands/meadows but similar to forest/woodlands (Figure 3; QM = 5.39, p = 0.02). Neither climate type (QM = 3.38, p = 0.07) nor focus of spatial scale (QM = 0.66, p = 0.41) influenced the magnitude of effect size (Figure 3).
The effect on ecosystem services of farmland with hedgerows as compared to farmland without hedgerows was positive, negative, and neutral in 127, 65, and 12 observations, respectively (N = 204 observations from 43 primary studies). Hedgerows did not enhance overall ecosystem service levels (Figure 4; estimated effect size = 0.30 ± 0.33, p = 0.36; I2 95% CI: 95.75–98.63%). However, meta-analysis revealed significant differences when comparing ecosystem services and farmland types (Figure 4). Farmland with hedgerows showed positive effects on provisioning services (estimated effect size: 1.44 ± 0.38, p < 0.01), but not on regulating (estimated effect size: −0.22 ± 0.35, p = 0.54) or supporting services (estimated effect size: −0.08 ± 0.35, p = 0.82). In addition, farmland with hedgerows provided more supporting than regulating services (QM = 5.61, p = 0.02). Crops showed higher ecosystem service levels than pastures (Figure 4; QM = 22.06, p < 0.01). The magnitude of effect size did not vary between crop types (Figure 4; QM = 0.09, p = 0.76), climate type (Figure 4; QM = 0.40, p = 0.53), and the spatial scale (Figure 4; QM = 0.13, p = 0.72). The complementary analyses on specific ecosystem services confirmed the pattern observed for the broad ecosystem-service categories (Supplementary Figure 2; Data File 5; R script ESHEFA_specific).
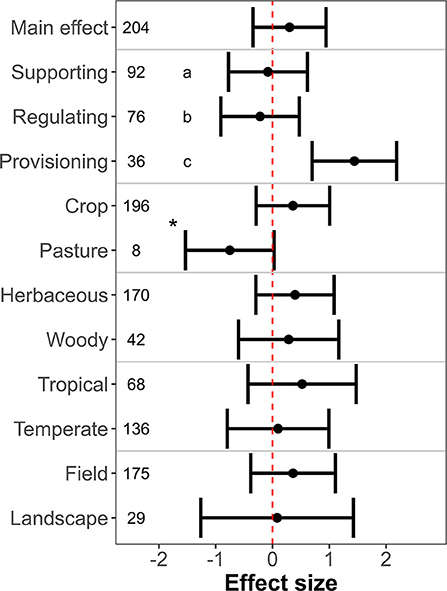
Figure 4. Ecosystem services levels in farmland with hedgerows as compared to farmland without hedgerows. X-axis indicates the mean effect size (standardized mean difference) ± 95% confidence interval. The dashed vertical red line shows the null effect. A confidence interval that overlaps this line means that the effects are not significant. Significant differences between levels of moderators are indicated by asterisks (two levels) and different letters (three levels), respectively.
The effect of farmland with hedgerows on ecosystem services as compared to natural habitat was negative, neutral, and positive in 32, 11, and 8 observations, respectively (N = 51 observations from 11 primary studies). Meta-analysis showed that natural habitat provided higher levels of ecosystem services than farmland with hedgerows (Figure 5; estimated effect size: −0.49 ± 0.20, p = 0.02; I2 95% CI: 58.20–94.84%).
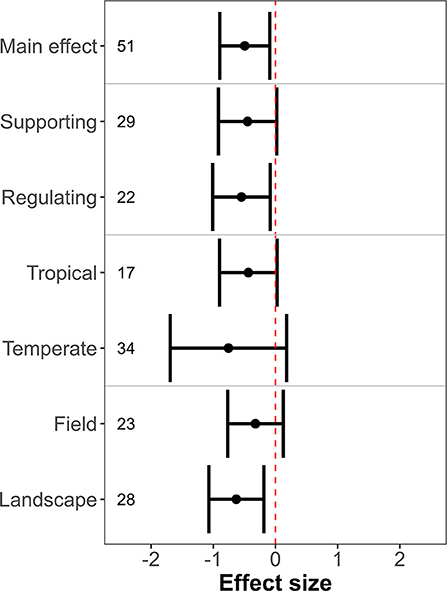
Figure 5. Ecosystem services levels in farmland with hedgerows as compared to natural habitat. X-axis indicates the mean effect size (standardized mean difference) ± 95% confidence interval. The dashed vertical red line shows the null effect. A confidence interval that overlaps this line means that the effects are not significant.
The magnitude of the effect size was similar between regulating and supporting services (Figure 5; QM = 0.21, p = 0.64), tropical and temperate climates (Figure 5; QM = 0.36, p = 0.55) and field and landscape scales (Figure 5; QM = 1.39, p = 0.24). There was insufficient statistical power to test the differences in effects size related to natural habitat types and provisioning services.
The complementary analyses for specific ecosystem services (soil formation, nitrogen cycling, and pest regulation) showed that natural habitats exhibited greater pest regulation services than farmland with hedgerows (Supplementary Figure 3; Data File 6; R script ESHENH_specific). These ecosystem services depended on climate type (hedgerows were more beneficial in temperate than in tropical regions; estimated effect size: temperate = −0.29 ± 0.06, p < 0.01; tropical = −1.00 ± 1.13, p < 0.01; QM = 23.37, p < 0.01).
Our results support the starting hypothesis (H1) of higher levels of biodiversity and provisioning ecosystem services in farmland with hedgerows as compared to farmland without hedgerows. However, despite farmland with hedgerows harbored as much biodiversity levels as natural habitat, natural habitat provided higher levels of ecosystem services than farmland with hedgerows (H2). Overall, the current meta-analysis agrees with a previous one (Van Vooren et al., 2017) conducted on arable crops in temperate regions. By broadening the scope to tropical regions and pastureland, we provide new insights on the contribution of hedgerows to people. In addition, the comparison of farmland with hedgerows and natural habitat supports the idea that farmland with hedgerows is an “intermediate,” more wildlife and ecosystem service friendly habitat than farmland without hedgerows (Rey Benayas and Bullock, 2012). The recent review by Albrecht et al. (2021) found that plant-species identity is crucial to optimize the contributions of hedgerows to ecosystem service delivery and ecological intensification of agriculture in the future. Table 3 synthesizes the results and hypotheses of this study that are discussed in the following paragraphs.
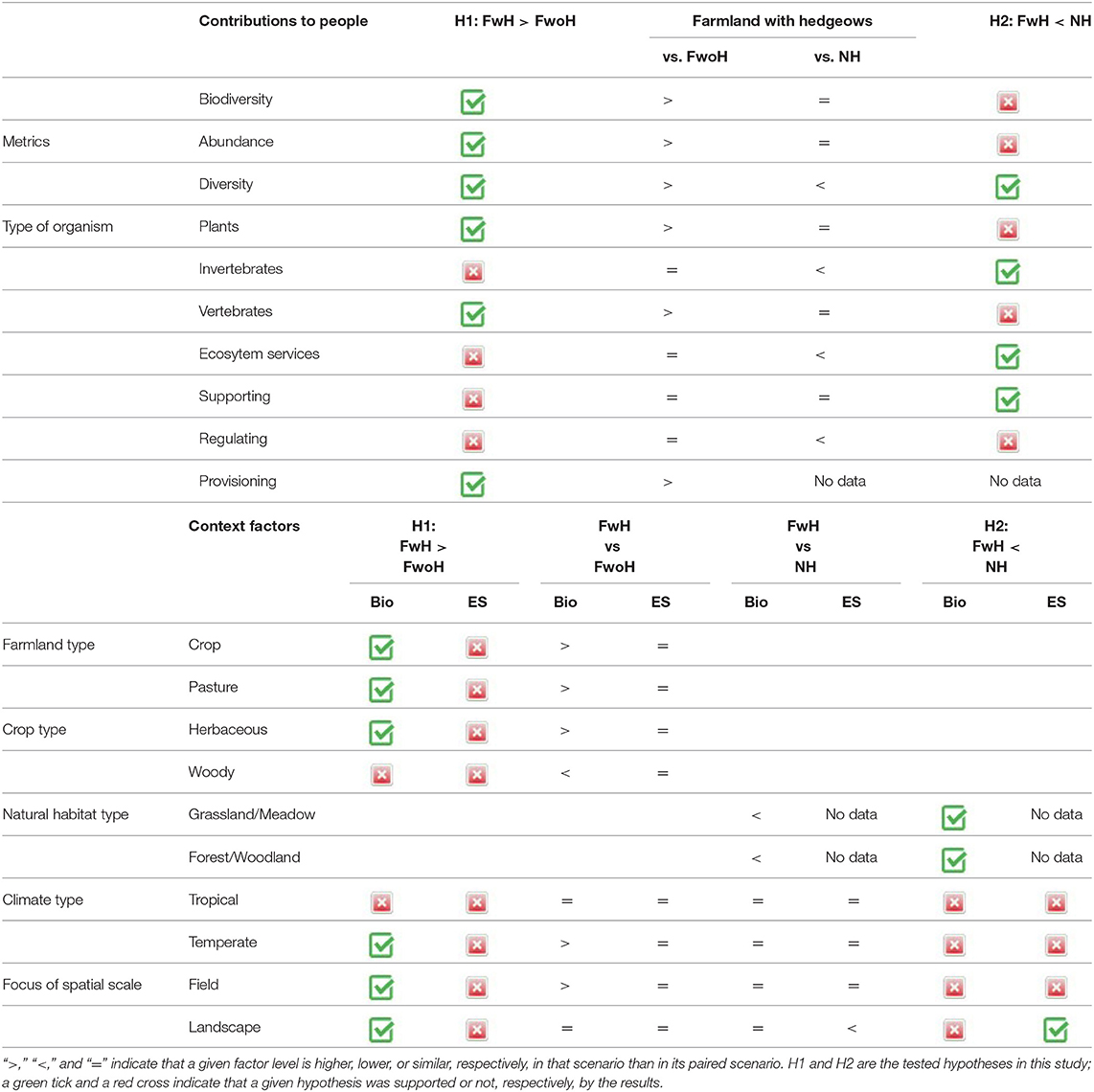
Table 3. Summary of comparisons of biodiversity (Bio) and ecosystem service (ES) levels between farmland with hedgerows (FwH) and farmland without hedgerows (FwoH), and between farmland with hedgerows (FwH) and natural habitat (NH).
Hedgerows enhanced the biodiversity levels in farmland, and both metrics (abundance and diversity) and farmland types (cropland and pastureland) studied here were similarly benefitted (Figure 2). Plants were the most positively affected taxa by hedgerows in agreement with previous studies. For instance, Aavik et al. (2008) found that semi-natural elements in the landscape enhanced plant species richness both at the field and at the landscape scales, and Evans et al. (2011) showed that uncultivated semi-natural habitats such as mature hedgerows are more species rich and have higher biomass than cropped habitat.
Hedgerows around herbaceous crops provide woody habitat otherwise absent or very scarce in agricultural landscapes. However, woody crops contain part of the hedgerows' functional diversity (i.e., the cultivated shrubs or trees) that may explain why herbaceous crops are more positively affected in terms of biodiversity than woody crops. For instance, the cultivated shrubs and trees facilitate the establishment of non-cropped perennial plants and other species (Oliveira et al., 2015). Castro-Caro et al. (2015) also reported that hedges, and ground covers to a lesser extent, efficiently increased the abundance and richness of passerine communities in olive groves, particularly of insectivorous birds. Other vertebrate species benefit from the presence of hedgerows. For instance, Verboom and Huitema (1997) suggested that landscapes with a high density of linear elements have a surplus value for serotine bats because bats use hedgerows as orientation clues, foraging habitat, and shelter from wind and predators.
The plant species that are found only in non-cropped habitat are primarily native and perennials, whereas a high proportion of the species found only in cultivated fields are annual and introduced species (Jobin et al., 1996). More exotic plants in farmland without hedgerows than in farmland with hedgerows have also been reported by Boutin et al. (2008). Plant species richness and composition varied substantially among the five landscapes studied by Freemark et al. (2002), ranging from a row-crop monoculture landscape to a diverse mosaic of cropped and non-cropped habitat landscape. The more complex the hedgerows, the species-richer farmland is (Klimek et al., 2014). Established shrubs may attract seed-dispersing wildlife and create proper environments for plant recruitment, generating reforestation feedback (Suárez-Esteban et al., 2013). In addition, trees planted into living fences generally have greater survival, relative growth, and height compared to those planted in open pasture (Love et al., 2009).
The difference in biodiversity levels between farmland with hedgerows and farmland without hedgerows was maximum in temperate regions (Figure 2). Such an extra benefit of hedgerows in temperate regions with respect to the tropics may be due to more ameliorated microclimate conditions for plant recruitment and wildlife animals (Ceresa et al., 2012; Burgess et al., 2015). For instance, the spider Nephila clavipes persists in highly fragmented tropical farmland landscapes thanks to the prevalence of relatively stable conditions (Pinkus-Rendón et al., 2006), whereas butterflies have been found to be favored by intermediate levels of landscape complexity (Dainese et al., 2015). The influence of drivers other than climate (e.g., hedgerow complexity) may explain why our analysis exhibited non-significant effects of hedgerows on invertebrate biodiversity (Figure 2).
Hedgerows were more beneficial at the field than at the landscape scale (Figure 2) since there is often some natural habitat other than hedgerows to support biodiversity at the landscape scale. The landscape context is important to the delivery of ecosystem services (Dainese et al., 2017). For instance, Fritz and Merriam (1996) concluded that habitat qualities that result from the architectural differences between hedgerows and forest may constrain plant movement. In landscapes of fragmented forest, hedgerows may provide corridors for woodland plants (Fritz and Merriam, 1996). However, non-fragmented (i.e., well-connected) forests do not need hedgerows for connectivity.
Ecosystem service levels in farmland with hedgerows were higher, albeit statistically insignificant, than in farmland without hedgerows. Our synthesis failed to highlight that hedgerows, in general, increased supporting or regulating services; however, hedgerows resulted in a higher amount of provisioning services – the ones that are sought in farmland habitat (Figure 4).
The complementary analyses confirmed the lack of significant differences for individual supporting and regulation services between farmland with hedgerows and farmland without hedgerows. This finding agrees with Albrecht et al. (2021) who concluded inconsistent and highly variable effects of hedgerows on crop pollination and yield and non-significant effects on pest control. Many agricultural systems use hedgerows as a supplementary source for cropping (for instance, intercropping of annual crops between Inga edulis contour hedgerows; Alegre and Rao, 1996). Hauser (2008) found that hedgerows dominated by cassava Manihot esculenta increased groundnut and maize production. In their meta-analysis, Van Vooren et al. (2017) reported that, up to 20 times the hedgerow height, yield is increased by an average of 6% as compared with farmland without hedgerows.
Conserving and restoring hedgerows may enhance some supporting services such as habitat creation (linked to biodiversity levels) and a range of regulating services. Shrubs and trees of hedgerows usually have well-developed root systems compared to herbaceous crops, which increases soil organic matter and sequesters carbon. Boutin's (2006) analyses of soil properties showed that organic matter and nutrient levels are often higher in hedgerows than in open fields. A hedgerow network protects landscapes from soil organic carbon erosion by runoff (Lacoste et al., 2016). Legume shrubs, which naturally fertilize soils, are common in hedgerow composition (Lemage et al., 2021). The architecture of some species such as the broom Retama sphaerocarpa provides shelter for many species (Matías et al., 2011). Franco (1998) and Van Vooren et al. (2017) reported that hedgerows retain 85 and 90% of suspended solids, 73 and 67% of total nitrogen, and 70 and 73% of total phosphorus in runoff, respectively.
Pastures were less benefitted by hedgerows than crop fields in terms of ecosystem services (Figure 4). Crop fields usually support a more intensive management scheme (tillage, fertilization, pesticide application) than pastures, where livestock fertilize the soil on crop fields. Heavy machinery compact soils and reduce supporting and regulating services (Power, 2010; The Research Council of Norway, 2011). Thus, hedgerows (i.e., unploughed, unfertilized features) around crop fields may become a reservoir for ecosystem services on top of biodiversity (Speir, 2015). In short, hedgerows play a relevant role in biodiversity conservation and climate change mitigation (Montgomery et al., 2020).
In farmland with hedgerows, abundance and diversity levels were similar and higher, respectively, than in natural habitat (Figure 3). At the population level, seedlings and juveniles are, usually, proportionally less abundant in hedgerows than in forests (Wehling and Diekmann, 2010), and the establishment of new populations of forest species in hedgerows is lower than in forests (Schmucki and De Blois, 2009). However, hedgerows represent a strong edge effect (forest and open fields) that may favor diversity (Litza and Diekmann, 2019).
We found a higher diversity of invertebrates in farmland with hedgerows than in natural habitat, particularly in grasslands and meadows. While these herbaceous habitats have seasonal food availability and fluctuant microclimate conditions, farmland with hedgerows can feed invertebrates along the year with complementary food resources and offer overwintering refuges. Thus, Jahnová et al. (2016) recorded more diverse ground-dwelling beetle communities in meadow margins than inside the meadows. Hedgerows help in ensuring a continuous supply of nectar on farmland; for instance, Timberlake et al. (2019) suggested that semi-natural habitat maintains complementary flowering phenology. The higher diversity of invertebrates in farmland with hedgerows than in natural habitat might be taxon specific. The finding of this study contrasts with Berwaerts et al. (1998) who found that the genetic diversity of speckled wood butterflies (Pararge aegeria L.) of large continuous woodland was significantly higher than in a highly fragmented area with very small woodland fragments and hedgerows scattered in an intensive agricultural landscape. Similarly, Buddle and Higgins (2020) suggested that lower spider diversity in hedgerows than in riparian forests may be due in part to the invasive honeysuckle shrub Lonicera maackii, which reduces hedgerow complexity on the ground layer.
Farmland with hedgerows provided higher amounts of ecosystem services than farmland without hedgerows, but lower amounts than natural habitat (Figure 5), thus supporting our H2 as other studies did. Hannon and Sisk (2009) found that hedgerow herbaceous flora is intermediate to that of crop fields and woodlands; consequently, regulating services such as pollination provided by natural habitat were higher than those provided by farmland with hedgerows. Byrne and Del Barco-Trillo (2019) found greater densities of bumblebees in unmanaged grasslands than in farmland with hedgerows. In this study, regulating services were particularly greater in natural habitat than in farmland with hedgerows at the landscape scale, whereas this effect is diluted at the field scale. However, we were unable to disentangle what moderators drove such a scale effect due to shortage of data.
The presence of hedgerows enhances the contributions of nature to people in agricultural landscapes (first hypothesis supported), including provisioning services. Farmland with hedgerows provides similar levels of biodiversity (an unexpected result mostly due to invertebrates) but lower levels of ecosystem services than natural habitat (second hypothesis partially supported). It is justified to recommend practitioners and policy makers conserving hedgerow networks and planting new hedgerows where they are absent or scarce to reach the goals of the EU Biodiversity Strategy for 2030 and maximize biodiversity and ecosystem services globally.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
JR conceived and designed the study. DG collected data and wrote first draft of the manuscript. DG and JR built the database. DG and EA analyzed the data. All authors contributed to revisions.
This study was supported by the Madrid Regional Government (REMEDINAL, ref. TE-CM S2018/EMT-4 338) and University of Alcala (CM/JIN/2019-023). DG was grateful to the Madrid Regional Government and the University of Alcalá for grant number 2018-T2/BIO-10995.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Sergio García Taboada, Navila Monteagudo, Baiba Ozolina, Jekaterina Bikova, and MarioVasquez Loy contributed to data gathering. Sara Villén-Pérez provided the R script to build Figure 1.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2021.789612/full#supplementary-material
Supplementary Figure 1. PRISMA flow diagram showing the process of study selection and exclusion criteria (adapted from Page et al., 2021).
Supplementary Figure 2. Levels of specific ecosystem services in farmland with hedgerows as compared to farmland without hedgerows. X-axis indicates the mean effect size (standardized mean difference) ± 95% confidence interval. The dashed vertical red line shows the null effect. A confidence interval that overlaps this line means that the effects are not significant. The significance of differences between specific ecosystem services are indicated in R script ESHEFA_specific.
Supplementary Figure 3. Levels of specific ecosystem services levels in farmland with hedgerows as compared to natural habitats. X-axis indicates the mean effect size (standardized mean difference) ± 95% confidence interval. The dashed vertical red line shows the null effect. A confidence interval that overlaps this line means that the effects are not significant. The significance of differences between specific ecosystem services are indicated in R script ESHENH_specific.
Data File 1. Contains the data used for comparing biodiversity of farmland with hedgerows and farmland without hedgerows; Identifier: it codes the primary study (Appendix 1); New.sign: it informs whether a given relationship is direct (+) or inverse (–); New.Response: it codes all response variables; New.Taxa: it categorizes all possible taxa into Plants, Invertebrates and Vertebrates; nHedgerow: The number of observations in farmland with hedgerows; nControl: The number of observations in farmland without hedgerows; Hedgerow.Value: The average value of the response variable in farmland with hedgerows; Hedgerow.SD: The standard deviation of the response variable in farmland with hedgerows; Farmland.Value: The average value of the response variable in farmland without hedgerows; Farmland.SD: The standard deviation of the response variable in farmland without hedgerows; Farmland.type: it indicates either crop or pasture; Crop.type: it indicates either herbaceous or woody; Climate.type: it indicates temperate or tropical; Spatial.Scale.Focus: it indicates field or landscape.
Data File 2. Contains the data used for comparing the biodiversity of farmland with hedgerows and natural habitats: nControl: The number of observations in natural habitats; Natural.habitat.Value: it provides the average of response variable at natural habitats; Natural.habitat.SD: The standard deviation of the response variable in the natural habitat; Natural.habitat.type; it indicates whether the control is a forest/woodland or a grassland/meadow; for other variables, see description in Data File 2.
Data File 3. Contains the data used for comparing the ecosystem services levels of farmland with hedgerows and farmland without hedgerows; Ecosystem.Service: it categorizes New.Response into Provisioning, Regulating, and Supporting; for other variables, see description in Data File 2.
Data File 4. Contains the data used for comparing the ecosystem services levels of farmland with hedgerows and natural habitats. Response: it indicates the ecosystem service involved in a given observation; Natural.habitat.type was excluded from these analyses due to a lack of power from grasslands/meadow level of factor; for other variables, please see description at Data File 3.
Data File 5. Contains the data used for comparing the levels of specific ecosystem services of farmland with hedgerows and farmland without hedgerows. The levels of New.Response are: SoilF: soil formation, Nitrogen: nitrogen cycling, Phosphorus: phosphorus cycling, Water: water regulation, SoilR: soil regulation, Climate: climate regulation, Food: food production; for other variables, see description of variables in Data File 3.
Data File 6. Contains the data used for comparing the levels of specific ecosystem services of farmland with hedgerows and natural habitats. See description of variables in Data Files 4, 5.
Data File 7. Contains the data needed to build Figure 1.R scripts | Figure_1a.docx contains the code applied on Data File 1; for building Figure 1; BioHEFA.docx contains the statistical analyses performed on Data File 2; BioHENH.docx contains the statistical analyses performed on Data File 3; ESHEFA.docx contains the statistical analyses performed on Data File 4; ESHENH.docx contains the statistical analyses performed on Data File 5; ESHEFA_specific.docx contains the statistical analyses performed on Data File 5; ESHENH_specific.docx contains the statistical analyses performed on Data File 6.
Appendix 1. List of studies used for each comparison, including the full reference and the identifier. This list eases the reproduction of meta-analyses with the provided data (Data Files 1–6). Specifically, section (a) includes the studies used for comparing biodiversity between farmland with hedgerows and farmland without hedgerows; section (b) the studies used for comparing biodiversity between farmland with hedgerows and natural habitats; section (c) the studies used for comparing ecosystem services provided by farmland with hedgerows and farmland without hedgerows; and section (d) the studies used for comparing the ecosystem services provided by farmland with hedgerows and natural habitat.
Aavik, T., Augenstein, I., Bailey, D., Herzog, F., Zobel, M., and Liira, J. (2008). What is the role of local landscape structure in the vegetation composition of field boundaries? Appl. Veg. Sci. 11, 375–386. doi: 10.3170/2008-7-18486
Albrecht, M., Kleijn, D., Williams, N. M., Tschumi, M., Blaauw, B. R., Bommarco, R., et al. (2021). The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: a quantitative synthesis. Ecol. Lett. 23, 1488–1498. doi: 10.1111/ele.13576
Alegre, J. C., and Rao, M. R. (1996). Soil and water conservation by contour hedging in the humid tropics of Peru. Agric. Ecosyst. Environ. 57, 17–25. doi: 10.1016/0167-8809(95)01012-2
Barbaro, L., Couzi, L., Bretagnolle, V., Nezan, J., and Vetillard, F. (2008). Multi-scale habitat selection and foraging ecology of the Eurasian hoopoe (Upupa epops) in pine plantations. Biodivers. Conserv. 17, 1073–1087. doi: 10.1007/s10531-007-9241-z
Barr, J., Britt, C. P., Sparks, T. H., and Churchwood, J. M. (1995). Hedgerow Management and Wildlife. A Review of Research on the Effects of Hedgerow Management and Adjacent Land on Biodiversity. (London: The Department for Environment, food and rural affairs (DEFRA) of the United Kingdom's government). p. 113. Available online at : https://hedgelink.org.uk/cms/cms_content/files/65_hedgerow_management_and_wildlife.pdf.
Berwaerts, K., Van Dyck, H., Van Dongen, S., and Matthysen, E. (1998). Morphological and genetic variation in the speckled wood butterfly (Pararge aegeria L.) among differently fragmented landscapes. Netherlands J. Zool. 48, 241–253. doi: 10.1163/156854298X00093
Borenstein, M., Hedges, L. V., Higgins, J. P. T., and Rothstein, H. R. (2009). Introduction to Meta-Analysis. Chichester: John Wiley & Sons, Ltd. p. 421.
Boutin, C. (2006). Comparison of the vegetation and seedbanks of soybean fields, adjacent boundaries, and hedgerows in Ontario. Can. J. Plant Sci. 86, 557–567. doi: 10.4141/P05-129
Boutin, C., Baril, A., and Martin, P. A. (2008). Plant diversity in crop fields and woody hedgerows of organic and conventional farms in contrasting landscapes. Agric. Ecosyst. Environ. 123, 185–193. doi: 10.1016/j.agee.2007.05.010
Buddle, C. M., and Higgins, S. (2020). The University of Notre Dame ground-dwelling spider assemblages inhabiting riparian forests and hedgerows in an agricultural landscape. Am. Midland Natural. 151, 15–26. doi: 10.1674/0003-0031(2004)151[0015:GSAIRF]2.0.CO;2
Burgess, M. D., Bright, J. A., Morris, A. J., Field, R. H., Grice, P. V., Cooke, A. I., et al. (2015). Umweltschutzprogramme in der landwirtschaft und ihr einfluss auf die besiedlung durch goldammern (Emberiza citronella) and grauammern (Emberiza calandra). J. Ornithol. 156, 153–163. doi: 10.1007/s10336-014-1113-1
Byrne, F., and Del Barco-Trillo, J. (2019). The effect of management practices on bumblebee densities in hedgerow and grassland habitats. Basic Appl. Ecol. 35, 28–33. doi: 10.1016/j.baae.2018.11.004
Capotorti, G., De Lazzari, V., and Ortí, M. A. (2019). Local scale prioritisation of green infrastructure for enhancing biodiversity in peri-urban agroecosystems: a multi-step process applied in the Metropolitan City of Rome (Italy). Sustain 11:3322. doi: 10.3390/su11123322
Castro-Caro, J. C., Barrio, I. C., and Tortosa, F. S. (2015). Effects of hedges and herbaceous cover on passerine communities in Mediterranean olive groves. Acta Ornithol. 50, 180–192. doi: 10.3161/00016454AO2015.50.2.006
Ceresa, F., Bogliani, G., Pedrini, P., and Brambilla, M. (2012). The importance of key marginal habitat features for birds in farmland: An assessment of habitat preferences of red-backed shrikes Lanius collurio in the Italian Alps. Bird Study 59, 327–334. doi: 10.1080/00063657.2012.676623
Dainese, M., Inclán Luna, D., Sitzia, T., and Marini, L. (2015). Testing scale-dependent effects of semi-natural habitats on farmland biodiversity. Ecol. Appl. 25, 1681–1690, doi: 10.1890/14-1321.1
Dainese, M., Montecchiari, S., Sitzia, T., Sigura, M., and Marini, L. (2017). High cover of hedgerows in the landscape supports multiple ecosystem services in Mediterranean cereal fields. J. Appl. Ecol. 54, 380–388. doi: 10.1111/1365-2664.12747
Endels, P., Adriaens, D., Verheyen, K., and Hermy, M. (2004). Population structure and adult plant performance of forest herbs in three contrasting habitats. Ecography 27, 225–241. doi: 10.1111/j.0906-7590.2004.03731.x
European Commission (2020). EU Biodiversity Strategy for 2030. Available online at: https://ec.europa.eu/environment/strategy/biodiversity-strategy-2030_en (accessed December 09, 2021).
Evans, D. M., Pocock, M. J. O., Brooks, J., and Memmott, J. (2011). Seeds in farmland food-webs: resource importance, distribution and the impacts of farm management. Biol. Conserv. 144, 2941–2950. doi: 10.1016/j.biocon.2011.08.013
Evans, K. L., Bradbury, R. B., and Wilson, J. D. (2003). Selection of hedgerows by swallows Hirundo rustica foraging on farmland: the influence of local habitat and weather. Bird Study 50, 8–14. doi: 10.1080/00063650309461284
Feber, R. E., Johnson, P. J., Firbank, L. G., Hopkins, A., and Macdonald, D. W. (2007). A comparison of butterfly populations on organically and conventionally managed farmland. J. Zool. 273, 30–39. doi: 10.1111/j.1469-7998.2007.00296.x
Fialho, M. Y. G., Cerboncini, R. A. S., and Passamani, M. (2019). Linear forest patches and the conservation of small mammals in human-altered landscapes. Mamm. Biol. 96, 87–92. doi: 10.1016/j.mambio.2018.11.002
Franco, D. (1998). “Hedgerows and non point source pollution: field test and landscape planning,” in Key Concepts in Landscape Ecologys, eds Dover J. W, Bunce R. G. H. (Garstang: IALE UK Colin Cross), 393–398.
Freemark, K. E., Boutin, C., and Keddy, C. J. (2002). Importance of farmland habitats for conservation of plant species. Conserv. Biol. 16, 399–412. doi: 10.1046/j.1523-1739.2002.00387.x
Fritz, R., and Merriam, G. (1996). Fencerow and forest edge architecture in Eastern Ontario farmland. Agric. Ecosyst. Environ. 59, 159–170. doi: 10.1016/0167-8809(96)01059-6
Gurevitch, J., Curtis, P. S., and Jones, M. H. (2001). Meta-analysis in ecology. Adv. Ecol. Res. 32, 199–247. doi: 10.1016/S0065-2504(01)32013-5
Hannon, L. E., and Sisk, T. D. (2009). Hedgerows in an agri-natural landscape: potential habitat value for native bees. Biol. Conserv. 142, 2140–2154. doi: 10.1016/j.biocon.2009.04.014
Hauser, S. (2008). Groundnut/cassava/maize intercrop yields over three cycles of planted tree fallow/crop rotations on ultisol in southern Cameroon. Biol. Agric. Hortic. 25, 379–399. doi: 10.1080/01448765.2008.9755063
Hedges, L., and Olkin, I. (1986). Statistical Methods for Meta-Analysis. 1st Edn. Elsevier. Available online at: https://www.elsevier.com/books/statistical-methods-for-meta-analysis/hedges/978-0-08-057065-5 (accessed December 09, 2021).
Heim, O., Lenski, J., Schulze, J., Jung, K., Kramer-Schadt, S., Eccard, J. A., et al. (2018). The relevance of vegetation structures and small water bodies for bats foraging above farmland. Basic Appl. Ecol. 27, 9–19. doi: 10.1016/j.baae.2017.12.001
Higgins, J. P. T., and Thompson, S. G. (2002). Quantifying heterogeneity in a meta-analysis. Stat. Med. 21, 1539–1558. doi: 10.1002/sim.1186
Holden, J., Grayson, R. P., Berdeni, D., Bird, S., Chapman, P. J., Edmondson, J. L., et al. (2019). The role of hedgerows in soil functioning within agricultural landscapes. Agric. Ecosyst. Environ. 273, 1–12. doi: 10.1016/j.agee.2018.11.027
Hotaling, N., Kuenzel, W., and Douglas, L. (2018). Breeding season bird use of restored wetlands in Eastern Maryland. Southeast. Nat. 1, 233–252. doi: 10.1656/1528-7092(2002)001[0233:BSBUOR]2.0.CO;2
IPBES (2017). IPBES Classification, 1–8. Available online at: https://www.ipbes.net/system/tdf/downloads/pdf/ipbes-5-inf-24.pdf?file=1&type=node&id=534 (accessed December 09, 2021).
IPBES (2019). Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. Available online at: https://ipbes.net/global-assessment%0Ahttps://ipbes.net/global-assessment-report-biodiversity-ecosystem-services (accessed December 09, 2021).
IPBES IPCC (2021). Scientific Outcome. 234. Available online at: https://www.ipbes.net/sites/default/files/2021-06/2021_IPCC-IPBES_scientific_outcome_20210612.pdf (accessed December 09, 2021).
Jahnová, Z., Knapp, M., Boháč, J., and Tulachová, M. (2016). The role of various meadow margin types in shaping carabid and staphylinid beetle assemblages (Coleoptera: Carabidae, Staphylinidae) in meadow dominated landscapes. J. Insect Conserv. 20, 59–69. doi: 10.1007/s10841-015-9839-5
Jobin, B., Boutin, C., and DesGranges, J. L. (1996). Habitats fauniques du milieu rural québécois: une analyse floristique. Can. J. Bot. 74, 323–336. doi: 10.1139/b96-041
Klimek, S., Lohss, G., and Gabriel, D. (2014). Modelling the spatial distribution of species-rich farmland to identify priority areas for conservation actions. Biol. Conserv. 174, 65–74. doi: 10.1016/j.biocon.2014.03.019
Koellner, T., and Scholz, R. W. (2008). Assessment of land use impacts on the natural environment: Part 2: generic characterization factors for local species diversity in Central Europe. Int. J. Life Cycle Assess. 13, 32–48. doi: 10.1007/s11367-006-0292-2
Koricheva, J., Gurevitch, J., and Mengersen, K. (2013). Handbook of Meta-Analysis in Ecology and Evolution. Princeton: Princeton University Press United States. 592 pp.
Lacoste, M., Viaud, V., Michot, D., and Walter, C. (2016). Model-based evaluation of impact of soil redistribution on soil organic carbon stocks in a temperate hedgerow landscape. Earth Surf. Process. Landforms 41, 1536–1549. doi: 10.1002/esp.3925
Lajos, K., Császár, O., Sárospataki, M., Samu, F., and Tóth, F. (2020). Linear woody landscape elements may help to mitigate leaf surface loss caused by the cereal leaf beetle. Landsc. Ecol. 35, 2225–2238. doi: 10.1007/s10980-020-01097-3
Lemage, B., Tsegaye, M., and Anmaw, Y. (2021). Evaluation and demonstration of leguminous shrubs hedgerows intercropping with maize crop. Int. J. Agric. Res. Innov. Technol. 11, 60–68. doi: 10.3329/ijarit.v11i1.54467
Litza, K., and Diekmann, M. (2019). Hedgerow age affects the species richness of herbaceous forest plants. J. Veg. Sci. 30, 553–563. doi: 10.1111/jvs.12744
Love, B. E., Bork, E. W., and Spaner, D. (2009). Tree seedling establishment in living fences: a low-cost agroforestry management practice for the tropics. Agrofor. Syst. 77, 1–8. doi: 10.1007/s10457-009-9244-8
Luptáčik, P., Miklisová, D., and Kováč, L. (2012). Diversity and community structure of soil Oribatida (Acari) in an arable field with alluvial soils. Eur. J. Soil Biol. 50, 97–105. doi: 10.1016/j.ejsobi.2011.12.008
Matías, L., Gómez-Aparicio, L., Zamora, R., and Castro, J. (2011). Effects of resource availability on plant recruitment at the community level in a Mediterranean mountain ecosystem. Perspect. Plant Ecol. Evol. Syst. 13, 277–285. doi: 10.1016/j.ppees.2011.04.005
MEA (2005). Ecosystem and Human Well-Being. Available online at: https://www.millenniumassessment.org/documents/document.356.aspx.pdf (accessed December 09, 2021).
Mestre, L., Schirmel, J., Hetz, J., Kolb, S., Pfister, S. C., Amato, M., et al. (2018). Both woody and herbaceous semi-natural habitats are essential for spider overwintering in European farmland. Agric. Ecosyst. Environ. 267, 141–146. doi: 10.1016/j.agee.2018.08.018
Monokrousos, N., Papatheodorou, E. M., Diamantopoulos, J. D., and Stamou, G. P. (2006). Soil quality variables in organically and conventionally cultivated field sites. Soil Biol. Biochem. 38, 1282–1289. doi: 10.1016/j.soilbio.2005.09.023
Montgomery, I., Caruso, T., and Reid, N. (2020). Hedgerows as ecosystems: service delivery, management, and restoration Annu. Rev. Ecol. Evol. Syst. 51, 81–102, doi: 10.1146/annurev-ecolsys-012120-100346
Morandin, L. A., and Kremen, C. (2013). Hedgerow restoration promotes pollinator populations and exports native bees to adjacent fields. Ecol. Appl. 23, 829–839. doi: 10.1890/12-1051.1
Morellet, N., van Moorter, B., Cargnelutti, B., Angibault, J. M., Lourtet, B., Merlet, J., et al. (2011). Landscape composition influences roe deer habitat selection at both home range and landscape scales. Landsc. Ecol. 26, 999–1010. doi: 10.1007/s10980-011-9624-0
Oliveira, C. C. C., Pereira, L. C. S. M., Lima, A., Shimabukuro, Y. E., and Torezan, J. M. D. (2015). Plant diversity in hedgerows amidst Atlantic Forest fragments. Acta Bot. Brasilica 29, 239–243. doi: 10.1590/0102-33062015abb0028
OrlŁowski, G. (2010). Effect of boundary vegetation and landscape features on diversity and abundance of breeding bird communities of abandoned crop fields in southwest Poland. Bird Study 57, 175–182. doi: 10.1080/00063650903449946
Ouin, A., and Burel, F. (2002). Influence of herbaceous elements on butterfly diversity in hedgerow agricultural landscapes. Agric. Ecosyst. Environ. 93, 45–53. doi: 10.1016/S0167-8809(02)00004-X
Page, M. J., McKenzie, J. E., Bossuyt, P. M., Boutron, I., Hoffmann, T. C., Mulrow, C. D., et al. (2021). The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. BMJ 372:n71. doi: 10.1136/bmj.n71
Pfister, S. C., Sutter, L., Albrecht, M., Marini, S., Schirmel, J., and Entling, M. H. (2017). Positive effects of local and landscape features on predatory flies in European agricultural landscapes. Agric. Ecosyst. Environ. 239, 283–292. doi: 10.1016/j.agee.2017.01.032
Pinkus-Rendón, M. A., León-Cortés, J. L., and Ibarra-Núñez, G. (2006). Spider diversity in a tropical habitat gradient in Chiapas, Mexico. Divers. Distrib. 12, 61–69. doi: 10.1111/j.1366-9516.2006.00217.x
Ponisio, L. C., M'Gonigle, L. K., and Kremen, C. (2016). On-farm habitat restoration counters biotic homogenization in intensively managed agriculture. Glob. Chang. Biol. 22, 704–715. doi: 10.1111/gcb.13117
Power, A. G. (2010). Ecosystem services and agriculture: Tradeoffs and synergies. Philos. Trans. R. Soc. B Biol. Sci. 365, 2959–2971. doi: 10.1098/rstb.2010.0143
R Core Team (2021). R: A Language and Environment for Statistical Computing. Available online: at: https://cran.r-project.org/ (accessed December 09, 2021).
Rey Benayas, J. M., Altamirano, A., Miranda, A., Catalán, G., Prado, M., Lisón, F., et al. (2020). Landscape restoration in a mixed agricultural-forest catchment: planning a buffer strip and hedgerow network in a Chilean biodiversity hotspot. Ambio 49, 310–323. doi: 10.1007/s13280-019-01149-2
Rey Benayas, J. M., and Bullock, J. M. (2012). Restoration of biodiversity and ecosystem services on agricultural land. Ecosystems 15, 883–899. doi: 10.1007/s10021-012-9552-0
Rey Benayas, J. M., and Bullock, J. M. (2015). Vegetation restoration and other actions to enhance wildlife in European agricultural landscapes. In: Pereira HM, Navarro LM, editors. Rewilding European Landscapes (Switzerland: Springer International Publishing), 127–142.
Rodríguez-Freire, M., and Crecente-Maseda, R. (2008). Directional connectivity of wolf (Canis lupus) populations in northwest Spain and anthropogenic effects on dispersal patterns. Environ. Model. Assess. 13, 35–51. doi: 10.1007/s10666-006-9078-y
Sardiñas, H. S., and Kremen, C. (2015). Pollination services from field-scale agricultural diversification may be context-dependent. Agric. Ecosyst. Environ. 207, 17–25. doi: 10.1016/j.agee.2015.03.020
Schmucki, R., and De Blois, S. (2009). Population structures and individual performances of Trillium grandiflorum in hedgerow and forest habitats. Plant Ecol. 202, 67–78. doi: 10.1007/s11258-008-9460-8
Slade, E. M., Merckx, T., Riutta, T., Bebber, D. P., Redhead, D., Riordan, P., et al. (2013). Life-history traits and landscape characteristics predict macro-moth responses to forest fragmentation. Ecology 94, 1519–1530. doi: 10.1890/12-1366.1
Speir, A. (2015). Soil compaction and pasture performance UGA Forage Team Newsletter III (IV), 2–5. Available online: at: https://extension.uga.edu/content/dam/extension-county-offices/madison-county/anr/newsletters/Forage%20Team%20Newsletter%20Volume%203-Issue%204%20December%202015.pdf (accessed December 09, 2021).
Suárez-Esteban, A., Delibes, M., and Fedriani, J. M. (2013). Unpaved road verges as hotspots of fleshy-fruited shrub recruitment and establishment. Biol. Conserv. 167, 50–56. doi: 10.1016/j.biocon.2013.07.022
The Research Council of Norway (2011). Heavy agricultural machinery can damage the soil, nordic researchers find. ScienceDaily May, 110505083737. Available online: at: www.sciencedaily.com/releases/2011/05/110505083737.htm (accessed December 09, 2021).
Timberlake, T. P., Vaughan, I. P., and Memmott, J. (2019). Phenology of farmland floral resources reveals seasonal gaps in nectar availability for bumblebees. J. Appl. Ecol. 56, 1585–1596. doi: 10.1111/1365-2664.13403
Van Vooren, L., Bert, R., Steven, B., Pieter, D. F., Victoria, N., Paul, P., et al. (2017). Ecosystem service delivery of agri-environment measures: a synthesis for hedgerows and grass strips on arable land. Agric. Ecosyst. Environ. 244, 32–51. doi: 10.1016/j.agee.2017.04.015
Vandevelde, J. C., Bouhours, A., Julien, J. F., Couvet, D., and Kerbiriou, C. (2014). Activity of European common bats along railway verges. Ecol. Eng. 64, 49–56. doi: 10.1016/j.ecoleng.2013.12.025
Verboom, B., and Huitema, H. (1997). The importance of linear landscape elements for the pipistrelle Pipistrellus pipistrellus and the serotine bat Eptesicus serotinus. Landsc. Ecol. 12, 117–125. doi: 10.1007/BF02698211
Viechtbauer, W. (2010). Conducting meta-analyses in R with the metafor package. J. Stat. Softw. 36, 1–48. doi: 10.18637/jss.v036.i03
Wehling, S., and Diekmann, M. (2010). Prediction of changes in the occurrence of forest herbs in hedgerow networks along a climate gradient in north-western Europe. Biodivers. Conserv. 19, 2537–2552. doi: 10.1007/s10531-010-9858-1
Wolka, K., Mulder, J., and Biazin, B. (2018). Effects of soil and water conservation techniques on crop yield, runoff and soil loss in sub-Saharan Africa: a review. Agric. Water Manag. 207, 67–79. doi: 10.1016/j.agwat.2018.05.016
Keywords: biodiversity, ecosystem services, farmland, multi-functionality, natural habitat
Citation: García de León D, Rey Benayas JM and Andivia E (2021) Contributions of Hedgerows to People: A Global Meta-Analysis. Front. Conserv. Sci. 2:789612. doi: 10.3389/fcosc.2021.789612
Received: 05 October 2021; Accepted: 17 November 2021;
Published: 22 December 2021.
Edited by:
Christopher Kaiser-Bunbury, University of Exeter, United KingdomReviewed by:
Matteo Dainese, Eurac Research, ItalyCopyright © 2021 García de León, Rey Benayas and Andivia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David García de León, ZGF2aWQuZ2FyY2lhbGhAdWFoLmVz
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.