 Kyle S. Tidwell
Kyle S. Tidwell Brett A. Carrothers
Brett A. Carrothers Daniel T. Blumstein
Daniel T. Blumstein Zachary A. Schakner
Zachary A. Schakner- 1United States Army Corps of Engineers, Bonneville Dam, Fisheries Field Unit, Cascade Locks, OR, United States
- 2Department of Ecology and Evolutionary Biology, University of California, Los Angeles, Los Angeles, CA, United States
- 3Office of Science and Technology, National Marine Fisheries Service, National Oceanic and Atmospheric Administration, Silver Spring, MD, United States
Protected Steller sea lions (Eumetopias jubatus) aggregate at Bonneville Dam on the Columbia River and prey upon multiple species of endangered salmon ascending the river. Hazing is a non-lethal activity designed to repel sea lions that includes aversive auditory and physical stimuli to deter animals from an area and has been employed with sea lion—fisheries interactions for more than 40 years but sea lion responses to hazing through time is not well-documented. We observed the behavior of Steller sea lions in periods with and without hazing during two spring Chinook salmon passage seasons to evaluate: (1) what effect hazing had on the number of animals present and their foraging behavior, and (2) whether they habituated to hazing. We found that hazing temporarily reduced the number of Steller sea lions, but only when actively hazed. During hazing, Steller sea lions were more likely to move away from hazers on the dam, decreased their foraging, and increased their time investigating the environment. However, these effects were temporary; their behavior returned to initial observation levels once hazing ceased. Furthermore, their responsiveness to hazing declined throughout the season, indicating habituation and raising concern for the application and long-term efficacy of hazing in managing predation on endangered salmon.
Introduction
The Marine Mammal Protection Act in 1972 is a conservation success story, accelerating the rebound of many Pacific Northwest pinniped populations [National Oceanic and Atmospheric Administration (NOAA), 2014; NOAA, 2015; Laake et al., 2018]. However, at the same time, largely because a mix of human development (Sobocinski et al., 2018; Cline et al., 2019), habitat destruction (Feist et al., 2003), and over-fishing (PFMC, 2016), 13 species of Columbia River salmon and steelhead have crashed and are now threatened or endangered (NMFS, 2019). An unintended consequence of pinniped recovery is that pinniped populations increasingly prey on endangered salmon (Schakner et al., 2016; Chasco et al., 2017). For more than 40 years, wildlife managers in the Pacific Northwest have sought to alter the foraging behavior of sea lions near impoundments to protect salmon runs. In the late 1980s, California Sea Lions (CSL, Zalophus californianus) predation significantly reduced endangered winter steelhead (Oncorhynchus mykiss) at Ballard Locks near Lake Washington in Washington State (Jeffries and Scordino, 1997; Fraker and Mate, 1999). Managers responded with non-lethal and later lethal interventions, but the winter steelhead run was ultimately extirpated. A similar situation has unfolded in the Columbia River near Bonneville Lock and Dam (Figure 1). Sea lion predation has directly increased the risk of extinction for several endangered salmon species (NMFS, 1997).
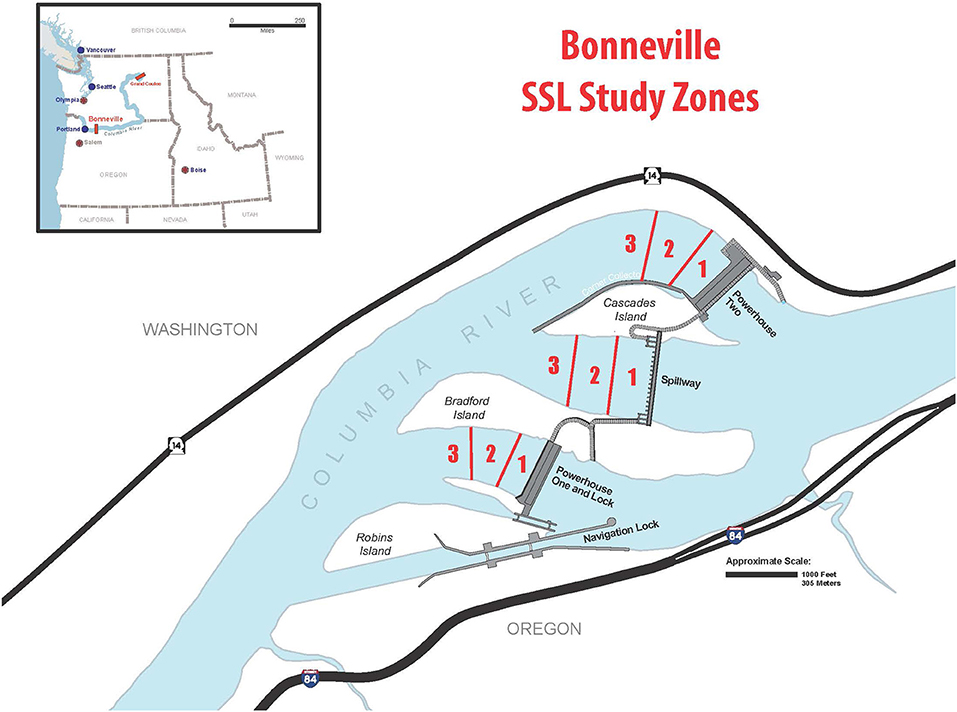
Figure 1. Bonneville Dam study area. Zones used to assign the location of Steller Sea Lions during study are defined. Observers and hazers stood on the dam and implemented study protocols on sea lions in the water of each tailrace.
Bonneville Lock and Dam is the first dam encountered by fish migrating upstream, located at river mile 146 (river kilometer 235). It spans the Columbia River between Oregon and Washington and consists of three concrete structures separated by islands. The structures form tailraces that guide fish to ladder entrances that facilitate passage (Figure 1). Thus, the dam bottlenecks fish passage (Kareiva et al., 2000; Quinones et al., 2015), concentrating the fish, making them vulnerable to predation (Naughton et al., 2011; Falcy, 2017). Since 2002, both CSLs and Steller sea lions (SSL, Eumetopias jubatus) have foraged on endangered salmon at Bonneville Dam (Stansell, 2004; Tidwell et al., 2020).
To comply with statutory guidance (NMFS biop), the U.S. Army Corps of Engineers initiated a program at Bonneville Dam to monitor the abundance of sea lions and the amount of predation and implement/evaluate hazing on pinnipeds at the dam. Hazing, a technique where deterrents are applied to immediately modify undesirable behavior (Schirokauer and Boyd, 1998; Hopkins et al., 2010), was initiated in 2002 with the ultimate aim of decreasing the predation on threatened salmonids as they migrate upstream. Deterrents applied at the dam include acoustic deterrent devices, paintballs, rubber shotgun projectiles, cracker shells, and seal bombs (i.e., underwater pyrotechnics) and physical barriers called Sea Lion Exclusion Devices that are placed on fish ladder entrances that prevent sea lion access to the fish ladders of the dam (NMFS, 2000, 2008, 2019). The combination of hazing, lethal removal, and non-lethal relocation of CSLs (over 283 lethally removed), have reduced CSL predation but created an opening for SSL at the dam. As a result, SSL are now the more abundant and persistent sea lion species near the dam (Figure 1, Tidwell et al., 2020).
Despite the long-term application of hazing at Bonneville, SSL behavioral response to hazing is uncertain (Scholl and Hanan, 1986; Schakner and Blumstein, 2013). Early reports from Bonneville Dam indicated that SSLs reportedly fled downstream following the initial bout of hazing. However, after repeated hazing, they fled for shorter periods and eventually remained near the dam despite hazing (Stansell, 2004; Norberg et al., 2005) (Figure 2). Hazing has been applied at Bonneville Dam with boats and on land from the dam's face. Evaluation of boat-based hazing of pinnipeds has been conducted at Bonneville Dam and elsewhere and deemed potentially effective at disrupting foraging but effectiveness is situationally dependent and has limited effectiveness over the long term (Lottis, 2009; Hatch et al., 2019). The declining response of SSLs to hazing stimuli indicates habituation, which is a recognized limitation of hazing with non-lethal deterrents (Schakner and Blumstein, 2013). Habituation, a non-associative learning process, exemplified by reducing responsiveness to stimulus exposure over time (Rankin et al., 2009), is frequently postulated in applied pinniped management (Geiger and Jeffries, 1987; Gearin et al., 1988) but rarely demonstrated empirically (as in Schakner and Blumstein, 2020). To test this, we exposed recurring SSL to controlled hazing periods (i.e., with and without hazing) throughout two spring Chinook passage seasons. We assessed land-based hazing exclusively because it has never been evaluated at an impoundment and is most feasible for management to apply in pinniped-fisheries interactions. We evaluated: (1) the effect of hazing on the foraging behavior of SSL near the dam and (2) whether hazing repelled individuals from the dam, and (3) whether SSL habituated to hazing stimuli. We predicted that hazing would have no impact on the number of individuals at the dam and that behavioral responses observed at the beginning of the season would diminish with subsequent hazing applications.
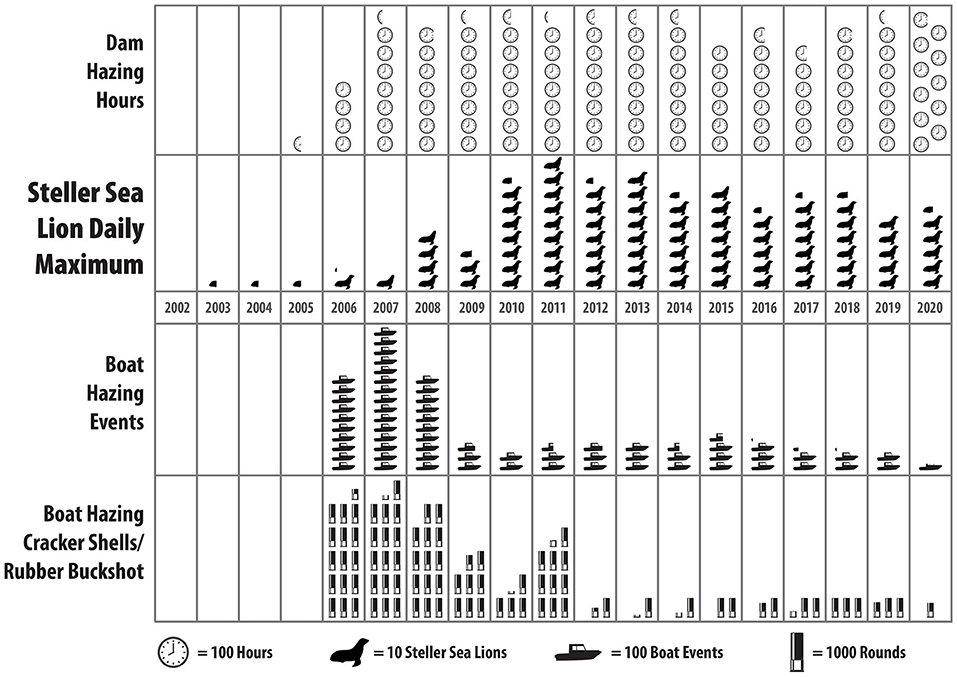
Figure 2. Non-lethal deterrent effort (Dam-and Boat Based hazing and the number of munitions spent to dissuade animals) and the number of Steller Sea Lions at Bonneville Dam between 2002 and 2020.
Methods
Study Subjects and Duration of Study
Spring Chinook salmon pass Bonneville Dam between early March and the end of June. Managers define the run as occurring between March 15 and June 15. During these months, Chinook comprise more than 95% of the fish passage estimates. All SSLs near Bonneville Dam are adult males. The females of the species remain at the coast and near the rookeries year-round while males migrate for foraging purposes. The SSLs at Bonneville Dam forage near the dam during the fall, winter, and spring and only leave the dam in June and July to breed and return to the dam in August. Alphanumeric cattle brands identify many, and some have been observed at the dam for up to 13 years. Not all SSL in the sample were individually identified, but monitoring program data indicate that 90% of individually identifiable SSL stay at the dam for 8–10 months of the year. As the spring Chinook run increases in April and May, more SSL move upstream to forage at Bonneville Dam.
Data were collected in conjunction with the sea lion predation monitoring program, which randomly assigns 1-h blocks of observation across the dam's three tailraces during all daylight hours using a systematic random design that provides equal distribution of at least 20 percent of daylight hours across all three tailraces (Tidwell et al., 2018). Behavioral sampling for this study was conducted for 1 week each month between March 1, 2018, and May 25, 2018, and again between April 14, 2020, and May 20, 2020 (Figure 3; Supplementary Table 1). During the sample period tailraces are subject to daily manipulation of water flow for hydropower production and since SSL presence across was randomly distributed across all three tailraces it was not individually monitored.
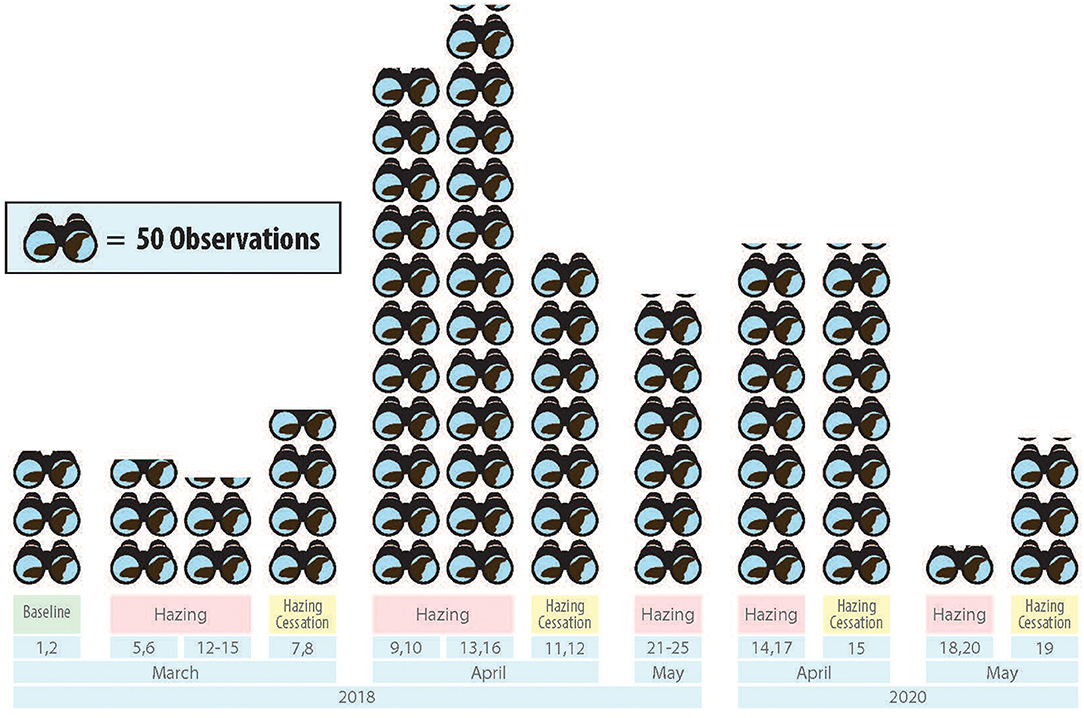
Figure 3. Treatment and sampling chronology for the study of Steller Sea Lion response to hazing at Bonneville Dam. Observations are the unique number of times surface active animals were recorded for study purposes during defined observation periods.
Treatment Groups
Pinniped hazing at Bonneville Dam is conducted under the authority of Sections 101(a)(4)(A)(iv) and 109(h)(1)(C) of the MMPA, which allows federal entities to dissuade and deter pinnipeds from federal property when needed (MMPA, 1994). Continuous daylight hazing is required as part of the lethal removal authority for CSL and had to be implemented. However, NOAA approved cessation and modification of hazing on a number of specific days to evaluate hazing effects. Dam-based hazing is applied from hazing personnel located on the dam structures overlooking the tailraces. No boat-based hazing occurred during the weeks of study in 2018 or 2020.
We evaluated SSL response to hazing by contrasting their behavior in a baseline condition without hazing (termed Baseline Treatment) to periods when hazing was applied (termed Hazing Treatment) and their behavior on days immediately following hazing (termed Hazing Cessation Treatment). Baseline observations occurred on the 2 days preceding the initiation of hazing in 2018. Of the three sampled weeks in March-May 2018 we collected data on two baseline days, 15 hazing days, and four post hazing days. In April-May 2020, we observed four hazing days and two post-hazing days (Figure 3).
Due to COVID-19 constraints, Bonneville Dam access was restricted in March 2020, and observers were not allowed to be present, but hazers were required to start hazing. As such, there was no sampling for the month and no baseline data were recorded in 2020.
Sampling
Five trained observers used 8 × 42 binoculars to monitor and sample SSL behavior in all three tailrace areas for a minimum of 1 h per sample using interval scan sampling and an ethogram. Because the mean dive time (length of time the animal is below the water's surface) for SSLs at Bonneville Dam is 6 min (Tidwell unpublished data), the observers sampled for 60 s every 3 min each hour to increase the likelihood of observing each SSL when it was at the surface. Observers recorded the data for each SSL at the beginning of the 60-s observation period rather than at the end of the observation period. The number of sea lions in each sampled tailrace was recorded before every 1-h observation period. If more than one SSL was present during an observation period, observers recorded all the data for the first SSL before recording data for additional SSL. All observers had extensive experience with SSL behavior from previous years. Where possible, they recorded individual SSL IDs during behavioral observations. Observers used scan sampling to provide a general snapshot of the population's behavior rather than focal follows of identified individuals, and, in most cases, behavioral observations were not linked with individual animals.
During the observation period at each tailrace, observers categorically scored the behavior, vigilance, and location of each SSL and whether the dam-based hazing was occurring (see the ethogram in Table 1). Each of these variables had three possible values intended to reveal the extent to which SSLs responded to hazing.
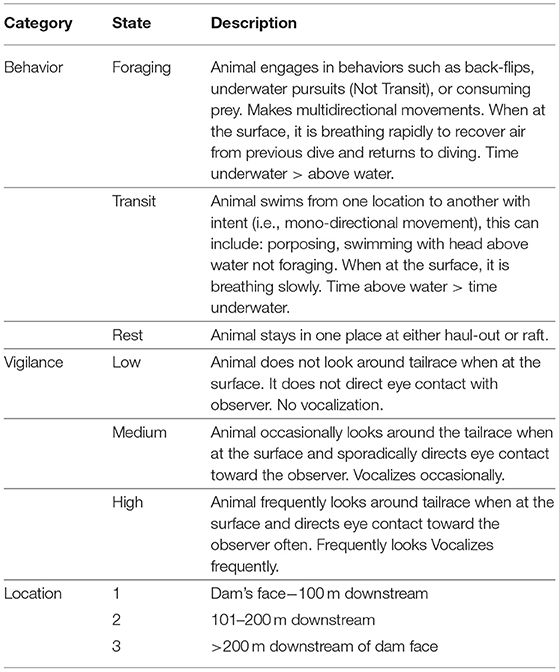
Table 1. Ethogram of Steller sea lion behavior used to categorize behavioral response to non-lethal hazing at Bonneville Dam.
Observers recorded the behavioral state of each individual by classifying them into three categories: foraging, traveling or resting. Animals transitioned between these states throughout the observation period; for example, if an animal was startled by a stimulus, including hazing, it might shift from foraging to traveling. These behaviors were scored based on the animal's body posture, movement pattern, and breathing intensity while at the surface. For example, an SSL was judged to be foraging if it briefly surfaced to breathe before quickly diving again, often without exposing its dorsal surface (e.g., in a cartwheel fashion), or surfaced to breathe and used its flippers to maintain a stationary position in the water column with its dorsal surface exposed after an extended dive. A traveling sea lion was at the surface longer, moved directionally with more speed, and breathed at a slower rate. A resting sea lion was defined as being either hauled out on land or resting in a raft with other sea lions. In the Bonneville Dam tailraces, resting in a raft is rarely observed but was included to be thorough.
Vigilance was scored as low, medium, or high based on the SSL's behavior and on almost two decades of SSL observation at Bonneville Dam that suggested it was sufficient to capture the variation in response to hazing. Low vigilance (score = 1) was characterized by an animal that exhibited no response to its surrounding environment and would never vocalize. High vigilance (score = 3) was characterized by an animal that was actively assessing the environment, looking at observers, other SSLs, or tailrace objects. The animal would actively look toward observers and vocalize often. Medium vigilance (score = 2) was a moderate expression of vigilance, and was characterized by an animal that occasionally vocalized or looked toward observers, but did not spend most of the surface active time investigating or vocalizing. Vigilance was scored independently of behavioral state, that is, animals could be foraging with high vigilance or traveling with high vigilance.
Location data were collected to assess how hazing might change SSL proximity to the dam. Location was characterized by defining three zones based on distance from the dam (Figure 1). Zone 1 was nearest the dam and is where the majority of SSL foraging occurs (Tidwell et al., 2018). Therefore, because hazing occurs from the dam and is nearest to Zone 1, animals that are more readily dispersed are expected to spend more time in Zone 2 or 3 than would otherwise be the case.
Continuous variables recorded included: date, number of pinnipeds present, and type of hazing. To assess how hazing might alter behavior over time we noted the number of hazing treatments (we refer to this as hazing number). For example, in March there were two Hazing Treatments, followed by a Hazing Cessation Treatment, then a third Hazing Treatment. We scored the third Hazing Treatment as Hazing Number = 3.
Statistical Analysis
For all statistical analyses, we used R 2.14 (R Development Core Team, 2014). The factors influencing the number of SSL present were analyzed using a generalized linear model (GLM) with a Poisson distribution and a log-link function. The following predictor variables were included in the “full” model structure; Hazing (presence/absence), date, year, as well as the interaction between hazing by zone interaction (Table 2). We compared models with different degrees of complexity and interpreted the estimates in the one with the lowest Akaike information criterion AIC value (Table 2—Burnham and Anderson, 2002).
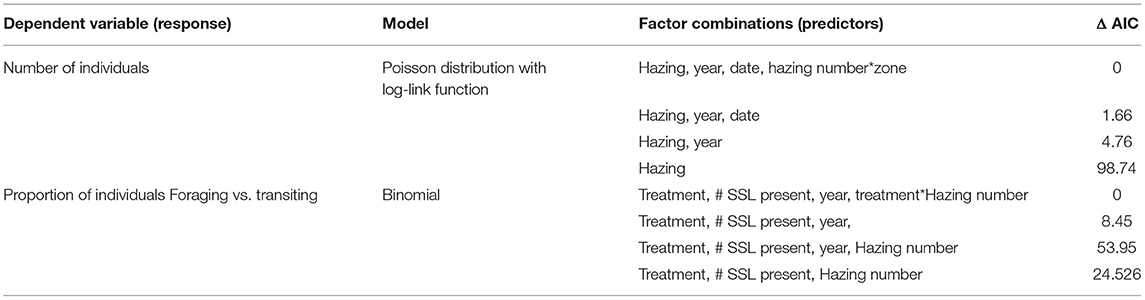
Table 2. Models that were evaluated during model selection.
To study factors that predicted SSL behavior (Foraging vs. Traveling), we fitted GLMs with a binomial distribution. The following predictor variables were included in the “full” model structure (Table 1): treatment type (Baseline, Hazing, and Hazing Cessation), year, hazing number, and number of SSL present, as well as the interaction between hazing number and treatment. Again, we compared models with different degrees of complexity and used AIC to identify the most efficient model.
Significance was assessed using confidence intervals (C.I.s, 95%) calculated with the “confint” function (method “Wald”) in lme4. Model assumptions were evaluated based on the distribution of residuals and quantile-quantile plots and by plots of residuals vs. fitted values (“DHARMa,” Florianhartig, 2019). All model parameter coefficients and C.I.s are shown on the scale of the response variable.
The factors affecting SSL vigilance were analyzed using an ordinal logistic regression model [Polr model function of MASS package (Venables and Ripley, 2007)]. This approach was chosen because of the discrete ordered nature of observed vigilance categories (Table 1). For this analysis, we used discrete vigilance categories low, medium and high as the response variable and treatment, number of SSL, year, Hazing number and Hazing number * treatment interaction as predictors. For the ordered logistic regression, the odds ratio (OR) and 95% confidence intervals are shown on the scale of the response variable. Model fit and diagnostics were validated using surrogate residuals calculated in the R package “sure” (Greenwell et al., 2017).
Results
In 2018 and 2020, we conducted 28 days of sampling and recorded 3,289 observations of SSL behavior during the various treatments (Figure 3). The number of SSL recorded ranged from 1 to 10 SSL with an average of 5.3 in 2018, and from 2 to 10 SSL with an average of 4.9 in 2020. In 2018 we observed a single animal resting and elected to exclude this from further analysis because its rarity at the Bonneville Dam.
We found that hazing reduced the overall number of SSL and altered their distribution (Table 3). The model predicted a 31 percent decrease in the number of SSL during the (Table 3). The interaction term of Hazing and Zone predicted a 47 percent increase in the number of SSL in Zone 3 (i.e., further away from the dam) during hazing. However, this effect appears temporary because date was marginally significant, indicating a trend toward increasing in the total number of animals in the observation area throughout the sampling period. In general more animals were predicted in Zone 2. Year was also significant, there were on average 18 percent fewer individuals at the dam during the 2020 field season, compared to 2018.
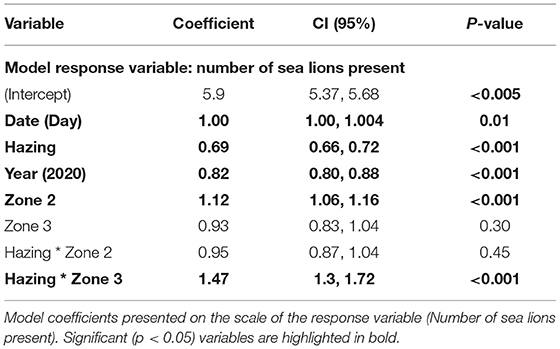
Table 3. General linear model predicting the factors influencing the number of SSL present at Bonneville during observation.
We found that the proportion of animals that are foraging relative to transiting changed with treatment type, the number of SSL, year, and the interactive effect of hazing and Hazing number. The application of the Hazing Treatment reduced SSL foraging and predicted an eight-fold increase in observations of SSL traveling (Table 4) which was not seen during the Baseline and Hazing Cessation treatments. Moreover, we found a significant, positive interaction between Hazing Treatment and Hazing number with behavior. Thus, with each subsequent application of the Hazing Treatment we found SSL were less likely to respond by traveling, and more likely to forage (Table 4). We also found a positive correlation between foraging behavior and the number of animals present. Year was also significant, with increased foraging behavior observed in the 2020 sampling season compared to 2018.
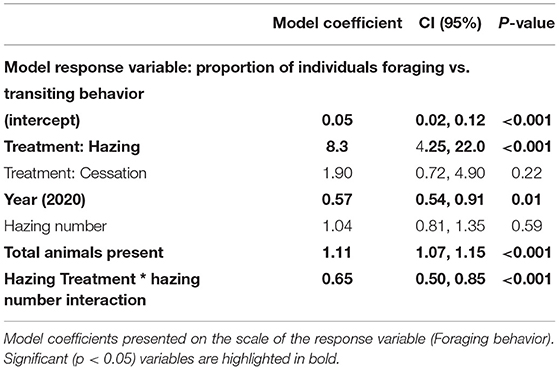
Table 4. Generalized linear model for sea lion foraging behavior (proportion of individuals foraging vs. traveling) during observation.
We found vigilance varied with Hazing Treatment, Year, hazing number, and the interactive effect of Hazing Treatment and hazing number (Table 5). The application of the Hazing Treatment predicted a three-fold increase in vigilance. However, this was not seen for the Hazing Cessation Treatment, thus vigilance appears to return to Baseline during the Hazing Cessation Treatment (Table 5). Moreover, we found a significant interaction between Hazing Treatment and hazing number, thus with each subsequent exposure to hazing, vigilance was reduced by nearly 25%. Additionally, year was significant, implying overall vigilance among the population was lower in the 2020 sampling season, compared to 2018.
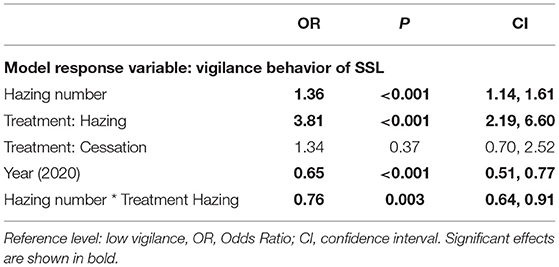
Table 5. Model estimates of the ordinal logistic regression model for factors predicting Vigilance behavior.
Discussion
For more than 40 years, wildlife managers in the Pacific Northwest have sought to alter the foraging behavior of sea lions near impoundments to reduce predation on protected salmon runs (Geiger and Jeffries, 1987; Jeffries and Scordino, 1997). By experimentally manipulating hazing throughout two spring Chinook salmon passage seasons, we were able to evaluate the effect of hazing on SSL behavior. We found hazing altered SSL behavior at the onset of the study, but the duration of effect was limited and returned to baseline measures equivalent to no hazing during the 3 months of study. Specifically, during the beginning of the spring Chinook run (i.e., in March), SSL were more likely to: move away from the face of the dam, leave the area being hazed, transit more than forage, and engage in increased vigilance. However, these effects were ephemeral; behavioral responses of both foraging and vigilance returned to initial observation levels immediately after hazing ceased. Additionally, by the end of the spring Chinook run (i.e., by May), despite 3 months of hazing, we found no difference in responsiveness to hazing from the initial observations. This pattern occurred in both years of study. From this, we conclude that hazing failed to fundamentally change the longer-term number and distribution of SSL or disrupt foraging behavior as the season progresses.
While habituation has long been hypothesized to underlie the lack of responsiveness to deterrents at Bonneville, we explicitly tested for and demonstrated habituation at the population level, as evidenced by diminished behavioral responses after repeated exposure to hazing stimuli. Habituation is rapid when there is short amount of time between exposures to stimuli (Staddon and Higa, 1996). At Bonneville, the foraging benefits from bottlenecked prey are high, resulting in motivation to forage there. Thus, if hazing initially repels animals away from the dam or reduces their foraging success, a motivation to forage may drive them back. Since the motivation to forage increases the rate at which an individual is exposed to deterrent stimuli, it accelerates habituation. Our study confirms the common observation that habituation is a common response to non-lethal hazing.
Hazing is frequently employed to mitigate a variety of pinniped/management issues but is rarely observed to change pinniped abundance, distribution, or foraging behavior (Long et al., 2015; Schakner and Blumstein, 2020). We know of only one successful application of hazing to reduce sea lion predation after hazing has stopped. In the Rogue River system on the southern coast of Oregon, CSL, SSL, and harbor seals (Phoca vitulina) were hazed 3 months to minimize predation fisheries. In addition to hazing, the port community restricted all positive attractants to pinnipeds. They actively deterred pinnipeds from hauling out and banned the disposal of fishing discards into the water at docks, which is known to attract sea lions and seals. This strategy successfully shifted pinniped distribution and prevented their presence around fishing over the long term (Lottis et al., 2007; Lottis, 2009). The success at Rogue river suggests that the structure of the focal area and prey concentrations largely dictate the success of the deterrent. Repellents generally work best when there are options for animals to move to a refuge, and there are few or a controlled number of positive attractants (Kemper et al., 2003; Breck et al., 2017; Gunther et al., 2018). Unfortunately, the large size of the Columbia River makes the application of many of the above deterrents difficult. Moreover, because the salmon at Bonneville Dam are especially concentrated at the dam tailrace and fish ladder entrances, it is nearly impossible to prevent animals from being attracted to such super-abundant resources, and hence, it may be impossible to successfully deter sea lions once they have arrived at the dam.
Our evaluation of hazing SSL at Bonneville Dam is consistent with previous observations and confirms that habituation underlies the waning behavioral response to hazing. At present, hazing fails to repel individuals from the dam and does not reduce SSL foraging behavior. Since hazing does not modify sea lion behavior over time, the continued application of conventional pinniped hazing intensities and techniques at Bonneville Dam raises practical issues. It draws into question whether it is useful to continue to do something that is both costly (Scordino, 2010) and ineffective. However, since hazing is currently a required management action, further investigation might assess how altering the current schedule and intensity of hazing might reduce habituation. Moreover, if lethal management actions are taken against SSL at Bonneville Dam, future research should investigate how newly recruited, and thus naive SSL, respond to hazing to determine how quickly habituation occurs and if it can be avoided. Hazing might generally be more effective for animals before they learn the value of resources and/or become a “problem.”
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics Statement
Ethical review and approval was not required for the animal study because pinniped hazing at Bonneville Dam is conducted under the authority of Sections 101(a)(4)(A)(iv) and 109(h)(1)(C) of the Marine Mammal Protection Act which allows federal entities to dissuade and deter pinnipeds from federal property when needed (MMPA, 1994). Our data were collected when federal operators were hazing sea lions under normal conditions and as such, were covered by the above provisions of the MMPA.
Author Contributions
KT: conceptualization, data curation, investigation, methodology, project administration, resources, supervision, validation, analysis, writing—original draft, and writing—review and editing. BC: data curation, investigation, methodology, project administration, validation, visualization, and writing—review and editing. DB: analysis, interpretation of results, and writing—review and editing. ZS: conceptualization, formal analysis, writing—original draft, and writing-review and editing. All authors contributed to the article and approved the submitted version.
Funding
Funding for data collection was provided by the U.S. Army Corps of Engineers, Portland District and funding for open access publication was provided by NOAA Office of Science and Technology.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We acknowledge and thank all our agency partners who continue to coordinate and assist with this work. Special thanks to the USDA personnel that conducted dam-based hazing, the Columbia River Inter-Tribal Fish Commission that conducted boat-based hazing, and the pinniped observation crews lead by Robert Stansell and Robert Wertheimer over the years that have supported data collection efforts. Data collection for this study was conducted by Kristen Bayley, Rebecca Cates, BC, Lindsay Magill, Bjorn van Der Leeuw, and KT. Figures were developed by John Leonard of the Portland District. We also thank our two reviewers for constructive suggestions that have helped clarify and improve our presentation.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2021.760866/full#supplementary-material
References
Breck, S. W., Poessel, S. A., and Bonnell, M. A. (2017). Evaluating lethal and nonlethal management options for urban coyotes. Hum. Wildl. Interact. 11, 133–145. doi: 10.26077/q5s9-vk08
Burnham, K. P., and Anderson, D. R. (2002). Model Selection and Multimodel Inference: A Practical Information-Theoretic Approach, 2nd Edn. New York, NY: Springer-Verlag.
Chasco, B. E., Kaplan, I. C., Thomas, A. C., Acevedo-Gutiérrez, A., Noren, D. P., Ford, M. J., et al. (2017). Competing tradeoffs between increasing marine mammal predation and fisheries harvest of Chinook salmon. Sci. Rep. 7:15439. doi: 10.1038/s41598-017-14984-8
Cline, T. J. J, and Ohlberger Schindler, D. E. (2019). Effects of warming climate and competition in the ocean for life-histories of Pacific salmon. Nat. Ecol. Evol. 3, 935–942. doi: 10.1038/s41559-019-0901-7
Falcy, M. (2017). Population Viability of Willamette River Winter Steelhead: An Assessment of the Effect of Sea Lions at Willamette Falls. ODFW Report.
Feist, B. E., Steel, E. A, Press, G. R, and Bilby, R. R. (2003). The Influence of scale on salmon habitat restoration priorities. Anim Conserv. 6, 271–282. doi: 10.1017/S1367943003003330
Florianhartig (2019). DHARMa: Residual Diagnostics for Hierarchical. (Multi-Level / Mixed) Regression Models. R package version 0.4.4. Available online at: http://florianhartig.github.io/DHARMa/
Fraker, M. A., and Mate, B. R. (1999). “Seals, sea lions, and salmon in the Pacific Northwest,” in Conservation and Management of Marine Mammals, eds J. R. Twiss Jr. and R. R. Reeves (Washington, DC: Smithsonian Institution Press), 156–178.
Gearin, P. J., Pfeifer, R., Jeffries, S. J., DeLong, R. L., and Johnson, M. A. (1988). Results of the 1986-87 California Sea Lion-Steelhead Trout Predation Control Program at the Hiram M. Chittenden Locks. NWAFC Processed Report 88-30, Alaska Fisheries Science Center, Newport, OR, United Sattes.
Geiger, A. C., and Jeffries, S. J (1987). Acoustical deterrents in marine mammal conflicts with fisheries. B. R. Mate and J. T. Harvey (eds.). Oregon State University Sea Grant College Program No. ORESU-W-86-001.
Greenwell, B., McCarthy, A., Boehmke, B., and Liu, D. (2017). Sure: Surrogate Residuals for Ordinal and General Regression Models. R package version 0.2.0. Available online at: https://cran.r-project.org/web/packages/sure/index.html (accessed February 21, 2021).
Gunther, K. A., Wilmot, K. R., Cain, S. L., Wyman, T. C., Reinertson, E. G., and Bramblett, A. M. (2018). Managing human-habituated bears to enhance survival, habitat effectiveness, and public viewing. Hum. Wildl. Interact. 12:7. doi: 10.26077/83cn-mh23
Hatch, D. R., Lessard, R., and Whiteaker, J. M. (2019). Sea Lion Monitoring and Non-Lethal Hazing, 1/1/2018 - 12/31/2018 Annual Report, 2009-004-00. p. 46.
Hopkins, J. B. III., Herrero, S., Shideler, R. T., Gunther, K. A., Schwartz, C. C., and Kalinowski, S.T. (2010). A proposed lexicon of terms and concepts for human-bear management in North America. Ursus 21, 154–168.
Jeffries, S. J., and Scordino, J. (1997). “Efforts to protect a winter Steelhead run from California sea lions at the Ballard Locks,” in Pinniped Populations, Eastern North Pacific: Status, Trends, and Issues, eds G. Stone, J. Goebel, and S. Webster (Boston, MA, Monterey, CA: New England Aquarium, and Monterey Bay Aquarium), 107–115.
Kareiva, P., Marvier, M., and McClure, M. (2000). Recovery and management options for spring/summer Chinook salmon in the Columbia River Basin. Science 290, 977–979. doi: 10.1126/science.290.5493.977
Kemper, C. Ms, Pemberton, D., Cawthorn, M., Heinrich, S., Mann, J., et al. (2003). “Aquaculture and marine mammals: co-existence or conflict,” in Marine Mammals: Fisheries, Tourism and Management Issues, eds N. Gales, M. Hindell, and M. R. Kirkwood (Melbourne, VIC: CSIRO), 209–225.
Laake, J. L., Lowry, M. S., DeLong, R. L., Melin, S. R., and Carretta, J. V. (2018). Population growth and status of California sea lions. J. Wildl. Manage. 82, 83–595. doi: 10.1002/jwmg.21405
Long, K. J., DeAngelis, M. L., Engleby, L. K., Fauquier, D. A., Johnson, A. J., Kraus, S. D., et al. (2015). Marine Mammal Non-Lethal Deterrents: Summary of the Technical Expert Workshop on Marine Mammal Non-Lethal Deterrents, 10-12 February 2015, Seattle, Washington. https://www.google.com/search?sxsrf=AOaemvL9pUlseuzRkoXPPBVP3zJrgRMCTA:1636115015162&q=Washington,+DC&stick=H4sIAAAAAAAAAOPgE-LQz9U3sEyrLFQCs4oyzLK1tLKTrfTzi9IT8zKrEksy8_NQOFYZqYkphaWJRSWpRcWLWPnCE4szMvPSS_LzdBRcnHewMgIAFDMcNlYAAAA&sa=X&ved=2ahUKEwj356iym4H0AhU4lEsFHZIvDZEQmxMoAXoECEIQAw Washington, DC: U.S. Dep. Commer.
Lottis, M. (2009). Field Report–Control of Nuisance Sea Lions in the Rogue River Estuary, 2008. Port of Gold Beach Report, Gold Beach, OR, United States.
Lottis, M., Tarwater, V., Nelson, S., Brown, R., Wright, B., Reimer, S., et al. (2007). Field Report—Control of Nuisance Sea Lions in the Rogue River Estuary. Gold Beach, OR, United States, 27p.
National Oceanic Atmospheric Administration (NOAA) (2014). Marine Mammal Stock Assessment. Available online at: https://www.fisheries.noaa.gov/national/marine-mammal-protection/marine-mammal-stock-assessment-reports-species-stock#pinnipeds—phocids-(earless-seals-or-true-seals) (accessed February 21, 2021).
Naughton, G. P., Keefer, M. L., Clabough, T. S., Jepson, M. A., Lee, S. R., Peery, C. A., et al. (2011). Influence of pinniped-caused injuries on the survival of adult Chinook salmon (Oncorhynchus tshawytscha) and Steelhead trout (Oncorhynchus mykiss) in the Columbia River basin. Can. J. Fish. Aquat. Sci. 68, 1615–1624. doi: 10.1139/f2011-064
NMFS (1997). Investigation of Scientific Information on the Impacts of California Sea Lions and Pacific Harbor Seals on Salmonids and on the Coastal Ecosystems of Washington, Oregon, and California. Seattle, WA: U.S. Department of Commerce, NOAA Technical Memorandum.
NOAA (2015). Potential Deterrence Methods for Pacific Harbor Seals, California Sea Lions and Eastern U.S. Stock Steller Sea Lions, Updated November 2015. Available online at: https://www.fisheries.noaa.gov/west-coast/marine-mammal-protection/deterring-nuisance-pinnipeds (accessed February 21, 2021).
Norberg, B., Stansell, R. J., Griffin, G., Brown, R. F., Jeffries, S. J., and Gearin, P. J. (2005). Field report – Preliminary Observations of Non-lethal Deterrence Measures for California Sea Lion Predation at Bonneville Dam. Seattle, WA: National Marine Fisheries Service, Protected Resourced Division. p. 23.
PFMC (2016). Review of 2015 Ocean Salmon Fisheries: Stock Assessment and Fishery Evaluation Document for the Pacific Coast Salmon Fishery Management Plan. Portland, OR: Pacific Fishery Management Council.
Quinones, R. M., Grantham, T. E., Harvey, B. N., Kiernan, J. D., Klasson, M., Wintzer, A. P., et al. (2015). Dam removal and anadromous salmonid (Oncorhynchus spp.) conservation in California. Rev. Fish Biol. Fish 25, 195–215. doi: 10.1007/s11160-014-9359-5
R Development Core Team. (2014). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing. Available online at: http://www.R-project.org/ (accessed February 21, 2021).
Rankin, C. H., Abrams, T., Barry, R. J., Bhatnagar, S., Clayton, D. F., Colombo, J., et al. (2009). Habituation revisited: an updated and revised description of the behavioral characteristics of habituation. Neurobiol. Learn. Mem. 92, 135–138. doi: 10.1016/j.nlm.2008.09.012
Schakner, Z. A., and Blumstein, D. T. (2013). Behavioral biology of marine mammal deterrents: a review and prospectus. Biol. Conserv. 167, 380–389. doi: 10.1016/j.biocon.2013.08.024
Schakner, Z. A., and Blumstein, D. T. (2020). “The California sea lion: Thriving in a human-dominated world,” in Ethology and Behavioral Ecology of Otariids and the Odobenids, eds R. Harcourt and C. Campangna (Cham: Springer Publishing), 347–365.
Schakner, Z. A., Buhnerkempe, M. G., Tennis, M. J., Stansell, R. J., van der Leeuw, B. K., Lloyd-Smith, J. O., et al. (2016). Epidemiological models to control the spread of information in marine mammals. Proc. R. Soc. B Biol. Sci. 283:20162037. doi: 10.1098/rspb.2016.2037
Schirokauer, D. W., and Boyd, H. M. (1998). Bear-human conflict management in Denali National Park and Preserve, 1982–94. Ursus 395–403.
Scholl, J., and Hanan, D. (1986). Acoustic Harassment Devices Tested in Combination With Crackershells on Pinnipeds Interacting With the Southern California Partyboat Fishery. California Department of Fish and Game, coastal marine mammal study, Annual Report from study period July 1 1982 – June 30 1983. Administrative report LJ-85-10C. Available online at: https://nsgl.gso.uri.edu/oresu/oresuw86001/oresuw86001_part3.pdf
Scordino, J. (2010). West Coast Pinniped Program Investigations on California Sea Lion and Pacific Harbor Seal Impacts on Salmonids and Other Fishery Resources. Report to the Pacific States Marine Fisheries Commission, Portland, OR, United States
Sobocinski, K., Greene, C., and Schmidt, M. (2018). Using a qualitative model to explore the impacts of ecosystem and anthropogenic drivers upon declining marine survival in Pacific salmon. Environ. Conserv. 45, 278–290. doi: 10.1017/S0376892917000509
Staddon, J. E. R., and Higa, J. J. (1996). Multiple time scales in simple habituation. Psychol. Rev. 103, 720–733. doi: 10.1037/0033-295X.103.4.720
Stansell, R. J. (2004). Evaluation of Pinniped Predation on Adult Salmonids and Other Fish in the Bonneville Dam Tailrace, 2002-2004. Cascade Locks, OR: U.S. Army Corps of Engineers, Bonneville Lock and Dam. Available online at: http://pweb.crohms.org/tmt/documents/FPOM/2010/Task%20Groups/Task%20Group%20Pinnipeds/ (accessed February 21, 2021).
Tidwell, K. S., McCanna, D. A., Cates, R. I., Ford, C. B., and van der Leeuw, B. K. (2020). Evaluation of Pinniped Predation on Adult Salmonids and Other Fish in the Bonneville Dam Tailrace, 2019. Cascade Locks, OR: U.S. Army Corps of Engineers, 60.
Tidwell, K. S., van der Leeuw, B. K., Magill, L. N., Carrothers, B. A., and Wertheimer, R. H. (2018). Evaluation of pinniped predation on adult salmonids and other fish in the Bonneville Dam tailrace, 2017. Cascade Locks, OR: U.S. Army Corps of Engineers, 54.
Keywords: Columbia River, Eumetopias jubatus, habituation, hazing, human-wildlife conflict, non-lethal deterrence, Steller sea lion
Citation: Tidwell KS, Carrothers BA, Blumstein DT and Schakner ZA (2021) Steller Sea Lion (Eumetopias jubatus) Response to Non-lethal Hazing at Bonneville Dam. Front. Conserv. Sci. 2:760866. doi: 10.3389/fcosc.2021.760866
Received: 18 August 2021; Accepted: 31 October 2021;
Published: 29 November 2021.
Edited by:
Jindong Zhang, China West Normal University, ChinaReviewed by:
Courtney R. Shuert, University of Windsor, CanadaColleen Cassady St. Clair, University of Alberta, Canada
Copyright © 2021 Tidwell, Carrothers, Blumstein and Schakner. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kyle S. Tidwell, a3lsZS5zLnRpZHdlbGxAdXNhY2UuYXJteS5taWw=