 Helen Pheasey1,2*
Helen Pheasey1,2* George Glen3
George Glen3 Nicole L. Allison1Luis G. Fonseca4Didiher Chacón4Jaime Restrepo5Roldán A. Valverde5,6
Nicole L. Allison1Luis G. Fonseca4Didiher Chacón4Jaime Restrepo5Roldán A. Valverde5,6- 1Caño Palma Biological Station, Canadian Organization for Tropical Education and Rainforest Conservation, Limón, Costa Rica
- 2Durrell Institute of Conservation and Ecology, School of Anthropology and Conservation, University of Kent, Canterbury, United Kingdom
- 3Archie Carr Center for Sea Turtle Research and Department of Biology, University of Florida, Gainesville, FL, United States
- 4Latin American Sea Turtles, Tibás, Costa Rica
- 5Sea Turtle Conservancy, Gainesville, FL, United States
- 6Department of Biological Sciences, Southeastern Louisiana University, Hammond, LA, United States
Estimates of illegal wildlife trade vary significantly and are often based on incomplete datasets, inferences from CITES permits or customs seizures. As a result, annual global estimates of illegal wildlife trade can vary by several billions of US dollars. Translating these figures into species extraction rates is equally challenging, and estimating illegal take accurately is not achievable for many species. Due to their nesting strategies that allow for census data collection, sea turtles offer an exception. On the Caribbean coast of Costa Rica, three sea turtle species (leatherback, Dermochelys coriacea; green, Chelonia mydas; and hawksbill, Eretmochelys imbricata) are exploited by poachers. Despite the consumption of turtle eggs and meat being illegal, they are consumed as a cultural food source and seasonal treat. Conservation programmes monitor nesting beaches, collect abundance data and record poaching events. Despite the availability of robust long-term datasets, quantifying the rate of poaching has yet to be undertaken. Using data from the globally important nesting beach, Tortuguero, as well as beaches Playa Norte and Pacuare on the Caribbean coast of Costa Rica, we modelled the spatial and temporal distribution of poaching of the three sea turtle species. Here, we present data from 2006 to 2019 on a stretch of coastline covering c.37 km. We identified poaching hotspots that correlated with populated areas. While the poaching hotspots persisted over time, we found poaching is declining at each of our sites. However, we urge caution when interpreting this result as the impact of poaching varies between species. Given their low abundance on these beaches, the poaching pressure on leatherback and hawksbill turtles is far greater than the impact on the abundant green turtles. We attribute the decline in poaching to supply-side conservation interventions in place at these beaches. Finally, we highlight the value of data sharing and collaborations between conservation NGOs.
Introduction
The illegal international trade in wildlife is estimated to be between US$8 and US$21 billion a year (Scheffers et al., 2019). This is severe enough to threaten biodiversity, damage source countries' economies, and is amongst the world's most lucrative criminal enterprises (Rosen and Smith, 2010; Nellemann et al., 2016). Our globalised economy means international trade chains are accelerating habitat degradation and species losses at locations far from the consumer (Lenzen et al., 2012). In response, 183 parties are signatory to the Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES), which regulates and monitors trade in c.35,000 species (CITES, 2021). All parties are required to submit annual reports of their trade data in CITES-listed species, and export quotas are imposed to limit the number of specimens that can enter the trade (Robinson and Sinovas, 2018). Management decisions and recommendations are often based on CITES trade data. However, CITES only concerns permitted species transiting international borders, and does not specifically account for illegal trade. Whilst CITES has no influence on domestic measures, other conventions have a more direct influence on in-country activities. The Inter-American Convention on Sea Turtles, for example, requires its 15 parties to commit to domestic measures to protect the six species of sea turtle that inhabit the Americas. Amongst these include, prohibiting the deliberate take and domestic trade of sea turtles or their eggs, and compliance with CITES (NOAA, 2021). However, decisions can only be made on available data. Social desirability bias, coupled with the fear of being caught, often causes violators to behave covertly, with reluctance to speak openly about wrongdoing (Nederhof, 1985; Gavin et al., 2009). This presents challenges when studying illegal behaviour and limits the availability of data, resulting in the need to extrapolate overexploitation estimates from small datasets.
Illegal wildlife trade is undertaken by three offender types: minor offenders, organised traders, and major crime syndicates (Wilson-Wilde, 2010). This results in a diverse array of tactics to extract and traffic wildlife, and requires an equally multifaceted approach to quantify and curtail wildlife crime. Conservationists and law enforcement officials use carcass counts, population estimates, black market trade prices, and meat sales in their attempts to quantify poaching (Milner-Gulland et al., 2001; Wittemyer et al., 2014). Bushmeat sales and market surveys offer an opportunity to assess wildlife trade in a cheaper and more practical way than attempting to estimate species abundances in areas of high hunting pressure (Fa, 2007; Allebone-Webb et al., 2011). However, due to the clandestine nature of illegal wildlife trade, it can be difficult to ascertain the availability of certain products, and surveys are often limited to items that are openly for sale or visually identifiable (Barber-Meyer, 2009; Moyle and Conrad, 2014; Pheasey et al., 2021). Targeting trafficking routes, either at transit hubs or through roadblocks, contributes seizure data. However, by definition, this is biassed towards only those items that are intercepted. These methods are resource-heavy, require trained personnel, and are often beyond the capacity of many source countries (Lee et al., 2005). In all cases, the accuracy of value estimates and quality of data directly correlates with the detectability of illicit goods. In other areas of criminology, more complete crime statistics are achievable due to the availability of both law enforcement observations and victim reports. Wildlife crime, by its nature, lacks victim reports (Lemieux, 2014).
Sea turtle nesting beaches may offer an exception. On beaches, the detectability of poached sea turtles or clutches is relatively high, and can be treated in much the same way as victim reports. This offers an opportunity to study terrestrial poaching with greater accuracy, and in some cases can offer near complete data on poaching rates. Direct take of turtles or eggs is considered a significant threat to many sea turtle populations (Mortimer and Donnelly, 2008; Seminoff et al., 2015; IUCN, 2021). Sea turtles are hunted for their meat, eggs and shell which has caused significant population declines (Heppell et al., 2003; Tomillo et al., 2008). On the Caribbean coast of Costa Rica, sea turtles are a traditional food source, and despite the practise being illegal, meat and eggs are still consumed today. Three species nest annually on Costa Rica's Caribbean coast, green (Chelonia mydas), hawksbill (Eretmochelys imbricata), and leatherback (Dermochelys coriacea). The most abundant species to nest in the Caribbean is the green turtle, which is categorised as endangered by the IUCN Red List (IUCN, 2021). The critically endangered hawksbill turtle nests in low numbers on this coastline, between May and September (WIDECAST, 2008). Whilst the Northwest Atlantic leatherback turtle population is classified as least concern by the IUCN, the population appears to have been in decline since 2004 (Troëng et al., 2004; Wallace et al., 2013; Wallace and Eckert, 2018). Due to its large size and unpalatable meat, the leatherback turtle is only exploited for its eggs in Costa Rica. Green turtles are prized for their meat and eggs, and the hawksbill turtle suffers additional exploitation for its shell, which is crafted into jewellery and trinkets (Mortimer and Donnelly, 2008).
In Costa Rica, sea turtles are protected under two conservation laws: Costa Rican law #8325 and a more general wildlife law #7317. Despite this, law enforcement is under-resourced and much of the trade occurs in remote locations, close to the nesting beaches (Pheasey et al., 2020, 2021). As seasonal nesters with stereotypic nesting behaviour, sea turtles are easy targets for human hunters. However, this predictability, coupled with the distinctive tracks they leave on the beach, enables census data collection during the nesting season. In the same manner, poaching activity also leaves distinctive traces in the sand, e.g., footprints or disturbance to the nest site. Drawing parallels with theories in criminology, these traces are the equivalent of victim reports and enable a greater degree of crime detection, often unavailable in other cases of illegal trade in terrestrial fauna. This makes accurate estimates of poaching much easier than for many other organisms. Despite this, few data have been published on poaching rates of sea turtles and their eggs (see Koch et al., 2006; Tomillo et al., 2008; Mancini and Koch, 2009; Senko et al., 2014, for exceptions).
Here, we contribute to filling this knowledge gap by collaborating and sharing our poaching data from 14 years of monitoring three nesting beaches on the Caribbean coast of Costa Rica. Our aims were to: 1. Report temporal trends in poaching rates at the three nesting beaches we monitor 2. Identify spatial patterns of poaching and correlations with beach exit points or properties. 3. Report spatiotemporal patterns in poaching.
Materials and Methods
Study Sites
We collected temporal and spatial poaching data from three sea turtle nesting beaches from 2006 to 2019. Our research transects are situated on the Caribbean coast of Costa Rica, on beaches Playa Norte, Tortuguero and Pacuare (Figure 1). Beach transects were measured in miles and reported in increments of 0.125 mi (c.200 m). Atlantic leatherback turtles nest annually on this stretch of coastline between February and June. The green turtle main nesting period is June–October with nesting density highest between July and September, whereas hawksbill turtles nest sporadically in low numbers throughout both seasons. Data were collected under permits from the Costa Rican Ministry of Environment and Natural Resources (MINAE).
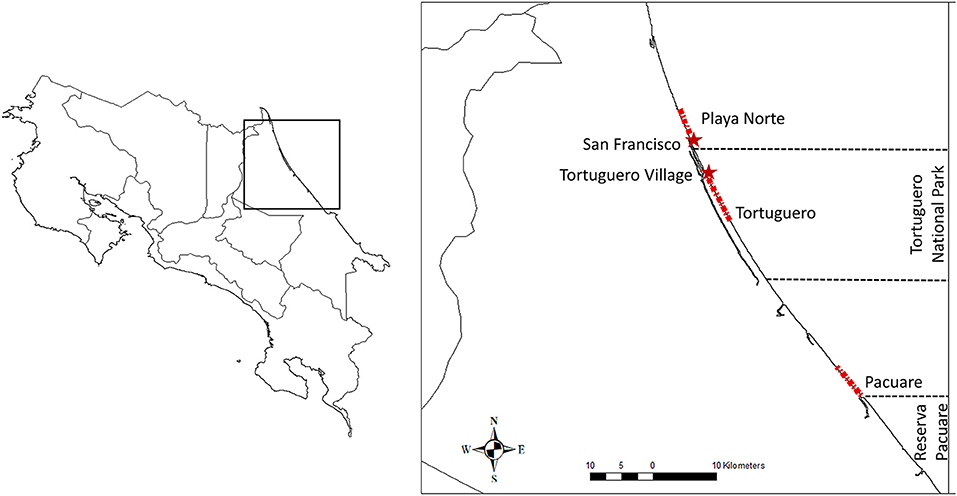
Figure 1. Playa Norte, Tortuguero, and Pacuare research transects monitored by Caño Palma Biological Station, Sea Turtle Conservancy, and Latin American Sea Turtles, respectively.
Playa Norte
Playa Norte is located within the “Barra del Colorado” Wildlife Refuge. This protected area lies to the north of the Tortuguero National Park (TNP). Playa Norte is a 5 km stretch of coastline which has <300 turtles nest in a typical year. The beach is monitored by Caño Palma Biological Station (CPBS) who conduct night patrols and morning census surveys throughout the season. Some domestic dwellings and small holdings are situated along the beach transect and the village of San Francisco is located to the south. The village houses c.600 residents. Employment and education levels are low in the area and the nearest police station is in Tortuguero. It is illegal to enter the beach at night without permits. Clutches remain in situ. Data were recorded daily and are available from 2006 to 2019.
Tortuguero
Tortuguero and Playa Norte are separated by the Laguna Tortuguero river mouth (c.700 m). Tortuguero hosts the largest nesting beach for green turtles (c.27,000 nesting females a year) in the western hemisphere. Currently, the economy thrives on tourism revenue from the c.80,000 visitors that arrive annually to observe nesting turtles (Harrison et al., 2005; Troëng and Rankin, 2005; Campbell, 2007). Tourists pay to observe turtles until midnight as part of guided tours, after which it is illegal to be on the beach without research permits. Research takes place over the c.8 km of the 29 km beach and is patrolled throughout the season by the Sea Turtle Conservancy (STC). Morning track census were run every 3 days during leatherback turtle season (February–June) and covered the full 29 km of beach. Nightly patrols take place in green turtle season (June–October) along the 8 km transect. Nests are monitored in situ. Here we use data from 2006 to 2019.
Pacuare
Latin American Sea Turtles (LAST) monitor Pacuare, the most southerly beach in this study. This is a c.5 km sandbank, with <40 permanent residents, situated c.40 km north of Puerto Limon. All beaches in this study see an influx of migrants during sea turtle nesting season, but this is particularly apparent in Pacuare (Pheasey, 2020). Migrants are attracted to the beach for its natural resources, particularly sea turtles. This is a public beach, situated outside the protected Reserva Pacuare, meaning there are no restrictions to entry. LAST patrol teams compete with poachers in an effort to relocate eggs to a hatchery with 24 h security. As clutches are rarely left in situ, all data are collected during night patrols. LAST began work in Pacuare in 2012 and data are available until 2019.
Data Collection
We conducted night patrols and morning census between 1st March and 31st October. However, these dates were flexible and coincided with nesting events. Our patrol teams comprised international volunteers, and aimed to collect data on nesting turtles. Patrols at Playa Norte and Pacuare also aimed to deter poaching. However, these teams were not acting in a law enforcement capacity. Patrols were a minimum of 4 h per team between 20.00 and 04.00. Patrol hours were fixed at Tortuguero but flexible at the other beaches, dependent on the number of nesting events and patrol teams available. In Pacuare and Playa Norte, teams remained with green and hawksbill turtles for the duration of the nesting event to ensure the turtle was protected from poachers. Due to the volume of turtles at Tortuguero this was not possible, and teams aimed to encounter and collect data on as many turtles as possible, within the 4-h patrol. When teams encountered a turtle before or during oviposition on Playa Norte or Tortuguero, they triangulated the nest to ensure it could be monitored throughout the incubation period, during morning census. Poaching pressure at Pacuare was too intense to leave nests in situ and therefore all encountered clutches were relocated to a hatchery (see Chacón et al., 2007 for nest relocation protocols).
Morning track surveys at Tortuguero and Playa Norte were undertaken to check the status of existing nests and record any new nests. Nest data include date, species, location and nest status (natural, poached, or lost for other reasons, i.e., mammalian predation). Indicators of poaching include visibly empty egg chambers, stick holes, dog and bare foot-prints and a few empty eggshells scattered near the nest. Any two or more of these indicate the clutch had been removed. In Pacuare, it was common for night patrol teams to witness eggs being poached and this was recorded. Evidence that a turtle has been poached include, characteristic drag marks–a turtle had been “flipped” onto its back and dragged into the vegetation–often accompanied by barefoot prints, particularly signs of heels heavily set in the sand, or signs that attempts have been made to disguise the tracks. These events were sometimes confirmed by the presence of a butchered carcass or a live turtle in the vegetation. When a live turtle was found, it was either rescued by patrol teams or at Pacuare, used as bait by the Coast Guards, who intercepted poachers on their return to the turtle. Under no circumstances would conservation patrol teams engage with poachers.
At Playa Norte and Tortuguero, triangulated nests were monitored throughout the incubation period and exhumed once the hatchlings had exited the nest. These exhumations aimed to record the success of the nest and confirm if it was poached (see Chacón et al., 2007 for nest exhumation protocols).
Data Analysis
It is possible to directly compare the number of clutches poached to the number of nests laid. However, not all females that exit the sea successfully nest, meaning we do not know the number of females poached to the number that enter the beach. Therefore, we only analysed poached clutches in the temporal data. High variation in the number of nesting events among species at different sites, meant there were insufficient data to analyse species separately (Supplementary Figure 1). However, our interest was in how human behaviour (poaching) changed, rather than interspecies differences. Therefore, we analysed the poaching events for all species in combination. Due to the differences in poaching pressure and public access at each beach, it was more appropriate to analyse data from each beach separately. At Tortuguero, data were collected at different scales during leatherback and green turtle seasons. Leatherback turtle nesting season covered the full 29 km of beach and data were collected every 3 days. During green turtle season data were collected over the 8 km transect every night. We analysed these seasons separately and only included green turtle season in the spatial analysis. At Pacuare, due to LAST relocating clutches to a hatchery, we analysed poaching rates and the number of clutches relocated to the hatchery (herein: rescued clutches) as separate datasets.
To test how the rate of poaching and relative percentage of poached clutches changed over spatial and temporal scales, we fitted generalised additive models (GAM) and generalised linear models (GLM) respectively. GAMs are a modelling technique that relate the predictors to the response variables, using smoothing functions, i.e., splines. This method is useful where the relationship between the response over the predictors is non-linear (Wood, 2017). We used the transect mile markers (MILE) and YEAR as our predictor variables and the response was the proportion of poached clutches, or rescued clutches in Pacuare. Where appropriate, we fitted thin plate regression splines for all smoothing functions other than those for YEAR (Wood, 2017). For YEAR, we used cubic regression splines. For both spatial and temporal models, we determined which distribution fit our data best using the Schwarz (or Bayesian) Information Criterion (BIC) (Schwarz, 1978; Brewer et al., 2016). ΔBIC is defined as the difference between the model with the lowest BIC and a competing model. We considered ΔBIC <2 indistinguishable in their ability of predicting the response (Burnham and Anderson, 2002).
Spatial
To examine the spatial data, we compared the poaching rates over the transect mile markers (MILE) and examined how the distribution of poaching along the beach has changed over time (YEAR). If the response was highly curvilinear over the explanatory variable, we proceeded to fit a GAM, otherwise a GLM was fitted. When the choice of modelling technique was unclear (GLM or GAM), we used the residual sum-of-squares and mean absolute error to check goodness-of-fit. This was done over an information-based approach, which are inappropriate in this situation because GLM and GAM use different methods to compute the likelihood, e.g., GAMs used penalised likelihood maximisation instead of minimising the log-likelihood (Marx and Eilers, 1998). For the proportion of poached clutches, we used a binomial error distribution with a logit link function. The proportion of poached clutches was computed as the number of poached clutches relative to the total nest abundance on that mile of the beach. When overdispersion was present (the dispersion parameter >1), we refitted the model with a quasibinomial distribution. For the count models that looked at how poaching is distributed over mile of the beach, and if poaching hotspots changed over the years, we fitted a GAM with a Poisson error distribution and a log link function. If overdistribution was present, we refitted the model with a negative binomial. The negative binomial distribution allows us to account for spatial heterogeneity in poaching events (Fisher et al., 1943). When there was substantial autocorrelation in the data, which we assessed using the acf() function in the package stats, we fitted an AR(1) model.
Temporal
To analyse how the rate of poaching has changed over time we used GLMs with a binomial error distribution and a logit link function, with YEAR as the temporal variable. The logit function related the mean of the response (proportion of poaching events) to the dependent variable (YEAR) via
where
and TPoached and TNests are total poached and total nest laid, α is a constant, ε is the binomial, or quasibinomial error term, and βi is the ith coefficient for the ith covariate in the model. For most models, the only covariate included was YEAR and β represented the effect of YEAR on the poaching rate.
Similar to the spatial analysis, the proportion of poached clutches was computed as the number of poached clutches relative to the total nest abundance on that mile of the beach. When overdispersion was present (the dispersion parameter >1), we refitted the model with a quasibinomial distribution. If there was substantial autocorrelation in the data, we fitted an AR(1) model. Due to the daily data collection at Tortuguero during green season, and at Playa Norte for both seasons, we were able to test for an effect of day of week (DOW) on poaching.
All data were analysed in R version 3.6.2 (R Core Team, 2019) using packages lmtest, zoo, dplyr, RColorBrewer, colorspace, MASS, bbmle, reshape2, ggplot2, ggthemes, mgcv, glmmTMB, MuMIn, itsadug, splines.
Results
The species assemblage of nesting turtles varied between beaches, but annual nesting events were recorded for all three species. Tortuguero and Playa Norte received the highest number of green nests (n = 299, 389, and 6318, respectively−2006–2019) while Pacuare was dominated by leatherback turtles (n = 1137−2012–2019). Poaching of clutches, as opposed to females, accounted for the majority of poaching events (Table 1; Supplementary Figure 2). Leatherback turtles were not poached at any of our sites.
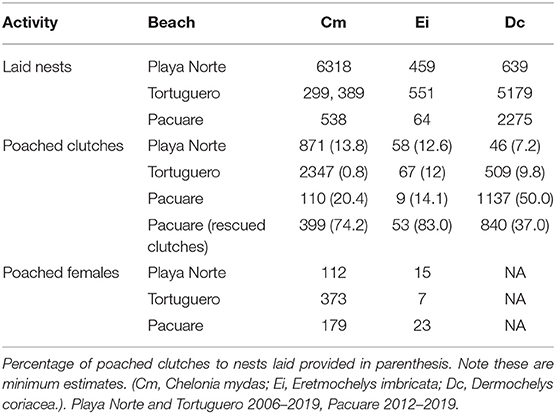
Table 1. Number of nests laid, clutches poached, clutches rescued (Pacuare), and female turtles poached at each site.
Spatial
At all beaches, mile maker had a significant effect on the distribution of poaching (Table 2A; Supplementary Table 1). At Playa Norte, we saw a general trend of relative poaching decreasing away from mile 0, with the exception of a peak increase in poaching around mile marker 0.750. This corresponds with proximity to San Francisco village and a property at 0.750. In Tortuguero, we identified peaks in activity between mile markers 3–3.375, where Tortuguero village borders the National Park. The spike in poaching around mile −0.375 appears to be driven by unusually high poaching activity at that marker in 2006 (Figure 2A). At Pacuare, poaching was more uniformly distributed across the transect, with most rescued clutches occurring between miles 2 and 3 (Figure 3). The spatial distribution of poached clutches has persisted over time at all beaches (Table 2B; Figures 2A–C).
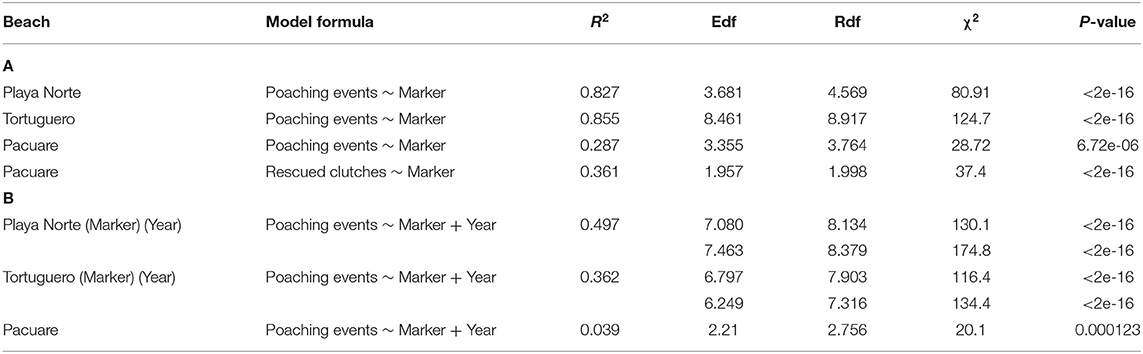
Table 2. The most parsimonious models for the spatial data with binomial distribution for poaching of clutches at each beach against (a) mile marker and (b) year (Edf, Estimated degrees of freedom; Rdf, Relative degrees of freedom).
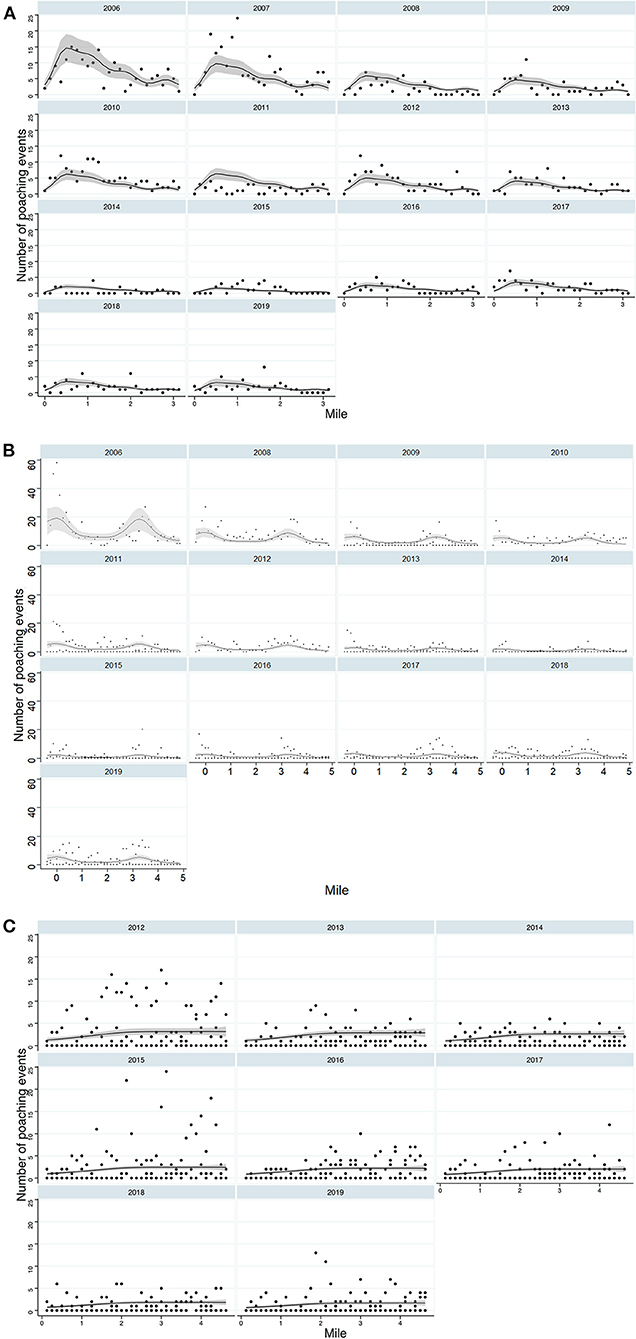
Figure 2. Spatiotemporal distribution of poaching on each beach, (A) Playa Norte, (B) Tortuguero (C) Pacuare. This includes females and nests. Solid line indicates the model fit and the grey shaded areas are 95% CI.
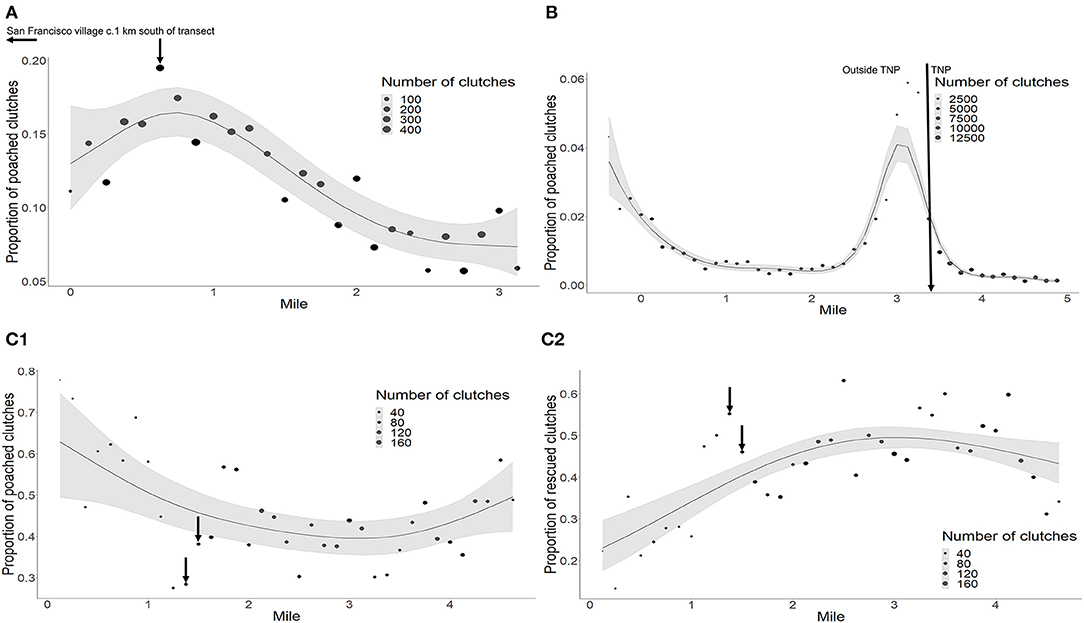
Figure 3. Spatial poaching trends across the beach transects. (A) Playa Norte, arrow depicts location with highest poaching frequency and a property with easy access to the beach. Data are recorded in miles (transect), geographical locations outside transects are in km. (B) Tortuguero, arrow depicts National Park border, (C1,C2). Pacuare, arrows depict LAST hatchery and volunteer accommodations. Point size indicates the number of clutches laid on that mile of beach. Solid line indicates the model fit, grey shaded areas are 95% CI.
Temporal
The percentage of poached clutches has been decreasing annually at each of our study sites (Playa Norte: log-odds = −0.17, 95% CI: −0.25−0.09; Tortuguero leatherback turtle season: log-odds = −0.19, 95% CI: −0.28−0.11; Tortuguero green turtle season: log-odds = −0.08, 95% CI: −0.11−0.04; Pacuare: log-odds = −0.10, 95% CI: −0.20–0.00). At Pacuare, the log-odds of rescued clutches has increased by 0.27, 95% CI: 0.24–0.30 (Figure 4). Including DOW and the interaction between DOW and Year did not improve the fit of the model for Playa Norte (ΔBICWeek = 34.9, ΔBICWeek*Year = 69.0). This suggests that at Playa Norte, DOW does not significantly affect poaching rates, implying poaching is consistent across days of the week and this has not changed across the duration of the study. However, in Tortuguero during green season, including the interaction between DOW and Year greatly improved the model fit (Tortuguero green season: ΔBICYear = 70.8, ΔBICYear+DOW = 25.7). This suggests that the rate and direction of poaching is not the same across years, but is temporally stratified by day of the week. Poaching for each day except for Thursday showed a declining trend (Supplementary Figure 3).
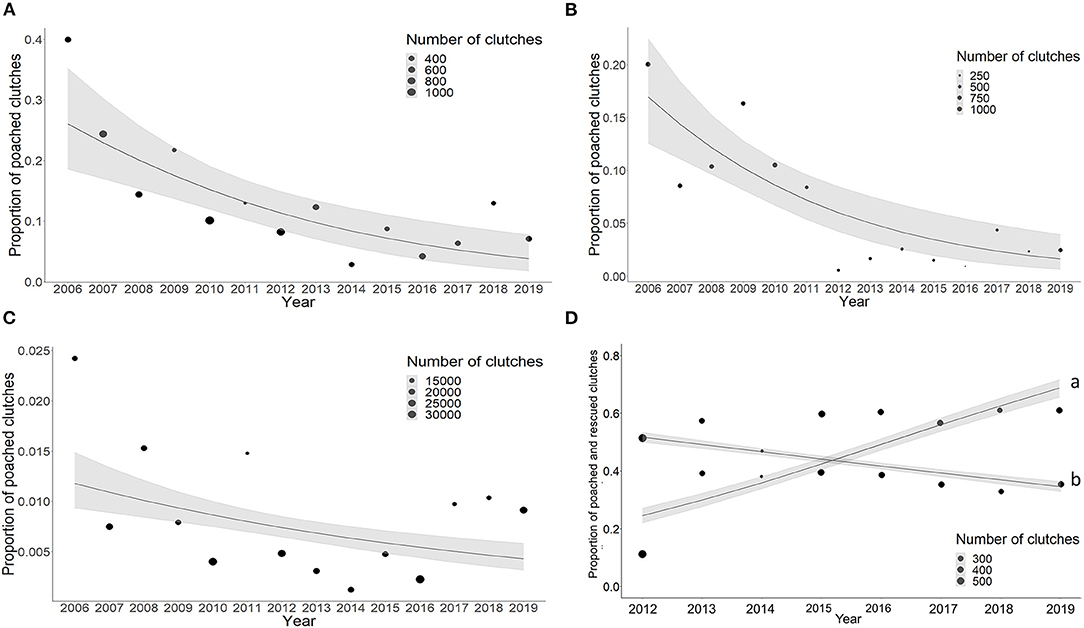
Figure 4. Annual decrease in percent of poached clutches at each study site. The shaded areas indicate the standard error. (A) Playa Norte, (B) Tortuguero leatherback season, (C) Tortuguero green season, (D) Pacuare, (a) proportion of rescued clutches, (b) poached clutches. Point size indicates the number of clutches laid on that mile of beach. Solid line indicates the model fit, grey shaded areas are 95% CI.
Discussion
Our findings show that at Playa Norte and Tortuguero, there were hotspots of poaching activity which correlated with populated areas. At Pacuare, we did not identify clear poaching hotspots, possibly as there is no nearby village, so poaching events were more uniform across the transect. While the spatial distribution of poaching remained consistent over the study period, poaching rates were declining at each site. Tortuguero green turtle season had the smallest percentage decrease in poaching (8%). This may be due to the long running conservation efforts and ecotourism in place at this site since the 1950's Ake, 2013). The initial impacts of these actions may not be visible in recent data. Conversely, in adjacent Playa Norte, where there were almost no tourism related incentives, the proportion of poaching was higher. However, overall poaching was decreasing. In Pacuare, decreasing poaching was coupled with increasing frequency of rescued clutches. This suggests that despite being an invasive method, relocating clutches to the hatchery is an effective anti-poaching strategy.
The ability to quantify poaching with a reasonable degree of accuracy offered an opportunity to identify whether the population was being overexploited. Across the beaches we recorded a loss of only 1.1% of green turtle clutches to poachers. With such a proportionately small and declining rate, it is unlikely that poaching of eggs is impacting this population. If human consumption is having a detrimental effect on the Caribbean green turtles, it is more likely to be taking place at their feeding grounds in the Miskito region of Nicaragua. Fishers there may legally harvest green turtles, and it is estimated that 7,000 are taken for human consumption annually (Eckert et al., 2020). In a study spanning 17 years, Lagueux et al. (2017) found that this population was exhibiting an overall decrease in mean body size of mature turtles, and a possible decline in juvenile recruitment. Changes in population characteristics such as these, may be indicative of overexploitation. Of more immediate concern however, is the impact of poaching on leatherback and hawksbill turtles at our study sites. Leatherback turtles accounted for 2.6% of nesting events in our study, yet lost almost a quarter of clutches to poachers (21%), the majority from Pacuare. The recent status review of the Northwest Atlantic leatherback population identified fisheries bycatch and beach erosion as the key threats to this species (Wallace and Eckert, 2018). With such great losses identified in our study, we argue that poaching also poses a significant threat. Our most vulnerable species, the hawksbill turtle exhibited only 0.34% of nesting events whilst suffering 12.5% clutch losses. Over the course of our study period, we recorded the poaching of 45 nesting females. The current IUCN assessment of hawksbill turtles highlighted the trade in shell and eggs as the most significant threat to this species (Mortimer and Donnelly, 2008). With such low nesting density in the region, the unit value and therefore loss, of each individual, is far greater than for the other species.
Wildlife crime has traditionally been a concern of conservationists, which has focused on species population declines and protected areas (Kurland et al., 2017). However, more recently it has caught the attention of criminologists, leading to the emergence of the conceptual framework Conservation Criminology. This field aims to bridge the gap between the two disciplines, and allows for the principles of traditional criminology to be applied to poaching (Gluszek et al., 2021). In this context, poaching is understood to be an interaction between offenders, guardians and victims, recognising wildlife as the victim (Lemieux, 2014). Situational Crime Prevention (SCP) focuses on the temporal and spatial dynamics of traditional crimes. It offers five crime mitigation mechanisms: 1. make it harder, 2. less rewarding, 3. increase the risk of offending, 4. remove excuses and 5. reduce provocations (Kurland et al., 2017). A number of studies have discussed the theoretical application of SCP to poaching (see Lemieux and Clarke, 2009; Pires and Moreto, 2011; Lemieux, 2014; Petrossian et al., 2016). These mechanisms are in operation to a greater or lesser degree at our study sites, and may serve to explain how a multipronged approach to conservation might explain the decline in poaching.
Anyone on the beach at night, be it patrol team or tourist, automatically increases the surveillance on the beach, potentially deterring anyone not wishing to be seen poaching. This makes it harder and less rewarding by increasing the poacher's search effort and reducing their likelihood of success. In Pacuare, this is amplified by removing clutches from the beach. Relocating clutches to a hatchery, however, is not a panacea for all beaches. It would appear be particularly beneficial at Playa Norte which received the highest abundance of the most valuable species, the hawksbill. However, there are several reasons why this intervention is unfeasible. Costal erosion on this beach is becoming increasingly sever and unpredictable, meaning there is no one location that could accommodate a hatchery for the duration of the nesting season. Further, hatcheries require 24 h surveillance, meaning at least one patrol per night would have to guard the hatchery. CPBS does not have the personnel to spare a patrol team for this. A possible intervention could be to use polystyrene cool boxes as incubators, stored inside the grounds of the biological station. However, this is a high risk strategy with a danger of egg mortality during transit across a canal, and the potential loss or damage to the boxes as the base severely floods twice a year. A detailed risk assessment would be needed before this intervention should be considered. However, the chief concern for this species is the loss of females and this can only be prevented through direct or indirect surveillance.
In Tortuguero, a natural resource surveillance committee (COVIRENA) has recently begun operating. This programme recruits volunteers who support public officials in helping to protect the area's natural resources (SINAC (Sistema Nacional de Áreas de Conservación), 2019). This additional manpower serves the added benefit of increasing the risk, or perceived risk, of arrest for poaching. When deciding whether to engage in illegal behaviour, potential wrongdoers undertake an implicit cost-benefit analysis: if benefit outweighs risk, it pays to commit a crime (Mancini et al., 2011). Our spatial analysis found the majority of poaching events occurred closer to populated areas. This suggests that despite the increased risk of being witnessed, or intercepted by law enforcement, the benefits outweigh the risks of poaching. This was particularly apparent in Tortuguero, where the abundance of turtles offers a low search effort and high reward, for a seemingly low risk activity. To that end, villagers from San Francisco reportedly poach on Tortuguero beach (Mejías-Balsalobre et al., 2021). If true, this implies that despite the increased risk of arrest, the benefit of poaching on Tortuguero is greater than the search costs associated with finding turtles closer to home. However, in Costa Rica, the likelihood of arrest and prosecution for poaching is low. In 2013, Costa Rican prisons were 137% over-capacity (Woods, 2015), leading to reforms resulting in lighter or no sentences for minor crimes. Moreover, there is a growing body of evidence suggesting tougher penalties do not equate to lower crime rates in wildlife offences (Challender and MacMillian, 2013; Wilson and Boratto, 2020).
The legality surrounding sea turtle poaching in Costa Rica is well-understood by law abiding citizens and poachers alike, making the 4th mechanism, removing excuses, less relevant here. However, removing provocations is applicable when we look to market forces. Recent studies in the region indicate that turtle eggs offer an opportunity to generate small but fast revenue. Easy sales occur door-to-door in short supply chains, directly from poacher to consumer (Hart et al., 2013; Pheasey, 2020; Mejías-Balsalobre et al., 2021). Understanding supply and demand dynamics enables targeted conservation interventions (McNamara et al., 2016). If trade appears to be demand driven, focusing on supplying alternatives, changing the source of the commodity from wild harvested, or attempting to change consumer preferences, may be effective. Conversely, supply-side dynamics may focus on alternative livelihoods for poachers, increased enforcement, or poverty alleviation interventions that move away from a reliance on the species in question (McNamara et al., 2016). Turtle eggs in the Caribbean are only available for a few months of the year and few households in Costa Rica depend on wild foods to fulfil protein needs or other livelihood benefits (Arauz-Almengor et al., 2001; Pheasey, 2020). This suggests the end consumer does not depend on turtle eggs or meat. On Playa Norte and Pacuare, searching for nests is time consuming and physically demanding. It is unlikely therefore, that people with employment, living close to a nesting beach would spend time searching for a nest, and are more likely to only take one they encounter opportunistically or close to home. Mejías-Balsalobre et al. (2021) found this to be the case in Tortuguero, where despite the high volume of turtles and therefore reduced effort of poaching, most consumers questioned in the study, stated they purchased turtle eggs from poachers, rather than actively poaching themselves. Therefore, while the demand is high, the illegal trade is predominately supply driven. Our conservation interventions reflect this by focusing predominantly on the poacher rather than the consumer, by attempting to deter and reduce poaching.
While poachers utilise the predictability of sea turtle nest site fidelity and seasonal nesting to target their activities, the specific timing of nesting events is harder to predict. Nevertheless, poaching behaviour mirrors nesting events. Our analysis uses a method which offers greater accuracy in identifying where poachers are likely to be active, regardless of the turtles' movements. This presents an opportunity in SCP enabling targeted law enforcement, and identifies specific locations where to focus resources. Moreover, we present a methodological advance in understanding poaching which is applicable to other species. Many freshwater turtle species nest along rivers, leaving the same distinctive tracks on the riverbanks. In Amazonia, river turtles of the genus Podocnemis are utilised for their meat and eggs, which is illegal or restricted according to the country in question (Conway-Gómez, 2008). Our method is directly applicable to monitoring the extraction of these species. Moreover, we suggest that many bird species offer similar life history traits that enable the application of our method, in particular many Psittacidae species. Parrots are amongst the most heavily trafficked of birds and often exhibit nest site fidelity and seasonal nesting, making them vulnerable to poaching of nests in the same way as turtles. Nest monitoring and data collection is also similar to that of turtle research, with data collection including nest success and poaching rates (Wright et al., 2001). These parallels offer the opportunity to adapt our analysis to the study of parrot nest raiding rates. We suggest our method is directly applicable to the quantification of extraction of these species and their eggs, which can help quantify illegal behaviour and inform policy decision making in much the same way as sea turtles.
Returning to our suggestion of treating empty nests as the equivalent of victim reports, we offer further discussion on the implications of regarding turtles as victims. The inclusion of non-human animals in victimology is complex. It incorporates debates on animal rights, utilitarianism and hierarchical speciesism. This is largely beyond the scope of this paper (for a detailed discourse see Flynn and Hall, 2017). In victimology, victims are subjects of harm, and this is incorporated into legislative decision making and sanction calculation. In Costa Rica, while turtles are not formally recognised as victims, prosecutions nevertheless follow the same model; incorporating the degree of harm inflicted (i.e., quantity of eggs or meat poached) in the calculation of sanctions. Tougher sanctions are afforded to more endangered species (Saborío Rodríguez, 2017; Castro Morales et al., 2019). However, formally recognising turtles as victims has the potential to be counterproductive in conservation. As conservationists we are often pragmatic about the non-consumptive use of a species. We recognise the overall benefits from ecotourism on both local economy and conservation of turtles, outweighs the disturbance to some individual nesters. If recognising turtles as victims were to lead to them being allocated rights, would this have an impact on ecotourism? Would the rights of the turtle to nest undisturbed be violated by tourists, and would the knock on effect mean fewer tourists and an increase in poaching? Therefore, whilst we encourage the inclusion of evidence of poaching as a form of victim reporting, and degree of harm as a tool for calculating sanctions, we urge caution in taking this philosophy further.
Due to the long lifespans and widespread distribution of sea turtles, there is great value in research that encompass long term data across multiple sites. Other studies have attempted to assess the impact of overexploitation on sea turtles; however, these have been restricted in both time and space (Koch et al., 2006; Mancini and Koch, 2009; Senko et al., 2014). Our study is a rare example of data sharing between NGOs and straddles natural and social sciences, as well as drawing upon theories of criminology. We encourage more multidisciplinary studies into the behaviour of humans in the exploitation of endangered species. Specifically, we recommend further research and data sharing between NGOs working on leatherback and hawksbill turtle nesting beaches, so further work can be done to enhance the conservation of these declining populations.
Data Availability Statement
The original contributions presented in the study are publicly available. This data can be found here: https://doi.org/10.22024/unikent/01.01.181.
Ethics Statement
Ethical review and approval were not required for this study as it involved analysing existing datasets collected under in-country permits.
Author Contributions
HP conceived and designed the study, organised the database, and wrote the majority of the manuscript. GG performed the statistical analysis and wrote the accompanying sections of the manuscript. NLA supported the statistical analysis and assisted with the first draft of the manuscript. All authors contributed to the revision, read, and approved the submitted version.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
The authors would like to thank the staff and volunteers who supported the data collection and running of our respective field stations, and MINAE for issuing permits to collect data. Additionally, we wish to thank the editor and reviewers for their support in improving this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2021.705556/full#supplementary-material
References
Allebone-Webb, S. M., Kümpel, N. F., Rist, J., Cowlishaw, G., Rowcliffe, J. M., and Milner-Gulland, E. J. (2011). Use of data to assess bushmeat hunting sustainability in Equatorial Guinea. Conserv. Biol. 25, 597–606. doi: 10.1111/j.1523-1739.2011.01681.x
Arauz-Almengor, M., Mo, C. L., and Vargas, M. E. (2001). Preliminary evaluation of olive ridley egg commerce from ostional wildlife refuge, costa rica. Mar. Turtle Newslett. 63, 10–13. Available on line at: http://www.seaturtle.org/mtn/archives/mtn63/mtn63p10.shtml
Barber-Meyer, S. M. (2009). Dealing with the clandestine nature of wildlife-trade market surveys. Conserv. Biol. 24, 918–923. doi: 10.1111/j.1523-1739.2010.01500.x
Brewer, M. J., Butler, A., and Cooksley, S. L. (2016). The relative performance of AIC, AICC, and BIC in the presence of unobserved heterogeneity. Methods Ecol. Evol. 7, 679–692. doi: 10.1111/2041-210X.12541
Burnham, K. P., and Anderson, D. R. (2002). Model Selection, and Multimodel Inference: A Practical Information-Theoretic Approach. Berlin: Springer Science, and Business Media.
Campbell, L. (2007). Local conservation practice and global discourse: a political ecology of sea turtle conservation. Ann. Assoc. Am. Geogr. 97, 313–334. doi: 10.1111/j.1467-8306.2007.00538.x
Castro Morales, C., Troëng, S., Monterrosa, L., Campbell, D., and Chamorro, E. (2019). Valoración del daño ecológico causado al medio ambiente referente a la caza de tortuga verde (Chelonia mydas). Available online at: http://www.latinamericanseaturtles.com/archivos/documentos/ValoracioneconomicaCm.pdf (accessed July 21, 2021).
Chacón, D., Sánchez, J., Calvo, J., and Ash, J. (2007). Manual para el manejo y la conservación de las tortugas marinas en Costa Rica; con énfasis en la operación de proyectos en playa y viveros. San José, CA: Sistema Nacional de Áreas de Conservación (SINAC), Ministerio de Ambiente y Energía (MINAE). Gobierno de Costa Rica.
Challender, D. W. S., and MacMillian, D. C. (2013). Poaching is more than an Enforcement Problem. Conserv. Lett. 7, 484–494. doi: 10.1111/conl.12082
CITES (2021). Convention on International Trade in Endangered Species. Available online at: https://www.cites.org/eng/disc/how.php (accessed July 21, 2021).
Conway-Gómez, K. (2008). Market integration, perceived wealth and household consumption of river turtles (Podocnemis spp.) in eastern lowland Bolivia. J. Lat. Amer. Geogr. 7, 85–108. doi: 10.1353/lag.2008.0013
Eckert, K., Azanza Ricardo, J., Barrientos-Muñoz, K. G., Barrios-Garrido, H., Berkel, J., and Bevan, E (2020). Sea Turtles of the Caribbean, SWOT Report, Vol 15. Available online at: https://www.seaturtlestatus.org/articles/2020/2/25/sea-turtles-of-the-caribbean (accessed July 21, 2021).
Fa, J. E. (2007). “Bushmeat markets–white elephants or red herrings?” in Bushmeat and Livelihoods: Wildlife Management and Poverty Reduction, eds G. Davies and D. Brown (Malden, MA: Blackwell), 47–60.
Fisher, R. A., Steven Corbet, A., and Williams, C. B. (1943). The relation between the number of species and the number of individuals in a random sample of an animal population. J. Anim. Ecol. 12, 42–58. doi: 10.2307/1411
Flynn, M., and Hall, M. (2017). The case for a victimology of nonhuman animal harms. Cont. Just. Rev. 20, 299–318. doi: 10.1080/10282580.2017.1348898
Gavin, M. C., Solomon, J. N., and Blank, S. G. (2009). Measure ad monitoring illegal use of natural resources. Conserv. Biol. 24, 89–100. doi: 10.1111/j.1523-1739.2009.01387.x
Gluszek, S., Viollaz, J., Mwinyihali, R., Wielnd, M., and Gore, M. L. (2021). Using conservation criminology to understand the role of restaurants in the urban wild meat trade. Conserv. Sci. Pract. 3:e368. doi: 10.1111/csp2.368
Harrison, E., Troëng, S., Guzman, A., and Rees, M. (2005). Report on the 2004 Green Turtle Program at Tortuguero, Costa Rica. San Pedro, CA: Caribbean Conservation Corporation.
Hart, K. A., Gray, T., and Steed, S. M. (2013). Consumptive versus non-consumptive use of sea turtles? stakeholder perceptions about sustainable use in three communities near Cahuita National Park, Costa Rica. Mar. Policy 42, 236–244. doi: 10.1016/j.marpol.2013.03.008
Heppell, S. S., Snover, M. L., and Crowder, L. B. (2003). “Sea turtle population ecology,” in The Biology of Sea Turtles, vol II, eds P. L. Lutz, J. A. Musick, and J. Wyneken (Boca Raton, FL: CRC Press), 275–306.
IUCN (2021). IUCN Red List of Threatened Species. Gland: International Union for the Conservation of Nature. Available online at: http://www.iucnredlist.org (accessed July 21, 2021).
Koch, V., Nichols, W. J., Peckham, H., and de la Toba, V. (2006). Estimates of sea turtle mortality from poaching and bycatch in Bahía Magdalena, Baja California Sur, Mexico. Biol. Conserv. 128, 327–334. doi: 10.1016/j.biocon.2005.09.038
Kurland, J., Pires, S. F., McFann, S. C., and Moreto, W. D. (2017). Wildlife crime: a conceptual integration, literature review, and methodological critique. Crime Sci. 6:4. doi: 10.1186/s40163-017-0066-0
Lagueux, C. J., Campbell, C. L., and Strindberg, S. (2017). Artisanal green turtle (Chelonia mydas) fishery of Caribbean Nicaragua: II. characterization and trends in size, sex, and maturity status of turtles killed, 1994–2011. Mar. Biol. 164:60. doi: 10.1007/s00227-017-3092-2
Lee, R. J., Gorog, A. J., Dwiyaherni, A., Siwu, S., Riley, J., Alexander, H., et al. (2005). Wildlife trade and implications for law enforcement in Indonesia: a case study from North Sulawesi. Biol. Conserv. 123, 477–488. doi: 10.1016/j.biocon.2005.01.009
Lemieux, A. M., and Clarke, R. V. (2009). The international ban on ivory sales and its effects on elephant poaching in Africa. Br. J. Criminol. 49, 451–471. doi: 10.1093/bjc/azp030
Lenzen, M., Moran, K., Kanemoto, K., Foran, B., Lobefaro, L., and Geschke, A. (2012). International trade drives biodiversity threats in developing nations. Nature 486, 109–112. doi: 10.1038/nature11145
Mancini, A., and Koch, V. (2009). Sea turtle consumption and black market trade in Baja California Sur, Mexico. Endanger. Species. Res. 7, 1–10.
Mancini, A., Senko, J., Borquez-Reyes, R., Guzman Póo, J., Seminoff, J. A., and Koch, V. (2011). To poach or not to poach an endangered species: elucidating the economic and social drivers behind illegal sea turtle hunting in Baja California Sur, Mexico. Hum. Ecol. 39, 743–756. doi: 10.1007/s10745-011-9425-8
Marx, B. D., and Eilers, P. H. C. (1998). Direct generalized additive modeling with penalized likelihood. Comput. Stat. Data Anal. 28, 193–209. doi: 10.1016/S0167-9473(98)00033-4
McNamara, J., Rowcliffe, M., Cowlishaw, G., Alexander, J. S., Ntiamoa-Baidu, Y., Brenya, A., et al. (2016). Characterising wildlife trade market supply- demand dynamics. PLoS ONE 11:e0162972. doi: 10.1371/journal.pone.0162972
Mejías-Balsalobre, C., Restrepo, J., Borges, G., García, R., Rojas-Cañizales, D., Barrios-Garrido, H., et al. (2021). Local community perceptions of sea turtle egg use in Tortuguero, Costa Rica. Ocean Coast. Manag. 201:105423. doi: 10.1016/j.ocecoaman.2020.105423
Milner-Gulland, E. J., Kholodova, M. V., Bekenov, A., Bukreeva, O. M., Grachev, I. A., Amgalan, L., et al. (2001). Dramatic declines in saiga antelope populations. Oryx 35, 340–345. doi: 10.1046/j.1365-3008.2001.00202.x
Mortimer, J. A., Donnelly, M., and (IUCN SSC Marine Turtle Specialist Group). (2008). Eretmochelys imbricata. The IUCN Red List of Threatened Species 2008: e.T8005A12881238. Available online at: http://dx.doi.org/10.2305/IUCN.UK.2008.RLTS.T8005A12881238.en (accessed July 21, 2021).
Moyle, B., and Conrad, K. (2014). The Chinese illegal ivory market: a pilot study. SSRN Electron J. doi: 10.2139/ssrn.2459207
Nederhof, A. J. (1985). Methods of coping with social desirability bias: a review. Eur. J. Soc. Psychol. 15, 263–280. doi: 10.1002/ejsp.2420150303
Nellemann, C., Henriksen, R., Kreilhuber, A., Stewart, D., Kotovou, M., Raxter, P., et al. (2016). The rise of environmental crime–a growing threat to natural resources peace, development and security. A UNEP-INTERPOL Rapid Response Assessment. United Nations Environment Programme and RHIPTO Rapid Response–Norwegian Center for Global Analyses. Available online at: www.rhipto.org (accessed July 21, 2021).
NOAA (2021). Inter-American Convention for the Protection and Conservation of Sea Turtles. Available online at: https://www.fisheries.noaa.gov/national/endangered-species-conservation/inter-american-convention-protection-and-conservation-sea#:~:text=The%20Inter%2DAmerican%20Convention%20(IAC,the%20benefit%20of%20sea%20turtles) (accessed July 21, 2021).
Petrossian, G., Pires, S. F., and van Uhm, D. P. (2016). An overview of seized illegal wildlife entering the United States. Glob. Crime 17, 181–201. doi: 10.1080/17440572.2016.1152548
Pheasey, H. (2020). Methods of and motives for laundering a wildlife commodity beyond captive farming-based systems: The harvest of olive ridley sea turtle eggs (Ph.D. thesis). University of Kent, Canterbury, United Kingdom.
Pheasey, H., Matechou, E., Griffiths, R. A., and Roberts, D. L. (2021). Trade of legal and illegal marine wildlife products in markets: integrating shopping list and survival analysis approaches. Anim. Conserv. doi: 10.1111/acv.12675
Pheasey, H., Roberts, D. L., Rojas-Cañizales, D., Mejías-Balsalobre Griffiths, R. A., and Williams-Guillen, K. (2020). Using GPS-enabled decoy turtle eggs to track illegal trade. Curr. Biol. 30, R1066–R1068. doi: 10.1016/j.cub.2020.08.065
Pires, S. F., and Moreto, W. D. (2011). Preventing wildlife crimes: solutions that can overcome the ‘Tragedy of the Commons'. Eur. J. Crime Policy Res. 17, 101–123. doi: 10.4324/9781315084589-25
R Core Team (2019). R: A Language and Environment for Statistical Computing. Available online at: https://www.R-project.org (accessed July 21, 2021).
Robinson, J. E., and Sinovas, P. (2018). Challenges of analyzing the global trade in CITES-listed wildlife. Conserv. Biol. 32, 1203–1206. doi: 10.1111/cobi.13095
Rosen, G. E., and Smith, K. F. (2010). Summarizing the evidence on the international trade in illegal wildlife. Ecohealth 7, 24–32. doi: 10.1007/s10393-010-0317-y
Saborío Rodríguez, G. (2017). Informe de valoracion economica del daño ambiental del caso del señor Pedro Vega Araya y otros. Heredia: SINAC-MINAE.
Scheffers, B. R., Oliveira, B. F., Lamb, I., and Edwards, D. P. (2019). Global wildlife trade across the tree of life. Science 366, 71–76. doi: 10.1126/science.aav5327
Schwarz, G. E. (1978). Estimating the dimension of a model. Ann. Stat. 6, 461–464. doi: 10.1214/aos/1176344136
Seminoff, J. A., Allen, C. D., Balazs, G. H., Dutton, P. H., Eguchi, T., Haas, H. L., et al. (2015). Status Review of the Green Turtle (Chelonia mydas) Under the US Endangered Species Act. NOAA Tech Memo NOAA-NMFS-SWFSC-539. Available online at: https://swfsc.noaa.gov/publications/TM/SWFSC/NOAA-TM-NMFS-SWFSC-539.pdf (accessed July 21, 2021).
Senko, J., Mancini, A., and Seminoff, J. A. (2014). Bycatch and directed harvest drive high green turtle mortality at Baja California Sur, Mexico. Biol. Conserv. 169, 24–30. doi: 10.1016/j.biocon.2013.10.017
SINAC (Sistema Nacional de Áreas de Conservación) (2019). Propuesta de Plan General de Manejo del Refugio Nacional de Vida Silvestre Dr. Archie Carr. Área de Conservación Tortuguero. San José, CA: Costa Rica.
Tomillo, P. S., Saba, V. S., Piedra, R., Paladino, F. V., and Spotila, J. R. (2008). Effects of illegal harvest of eggs on the population decline of leatherback turtles in Las Baulas Marine National Park, Costa Rica. Conserv. Biol. 22, 1216–1224. doi: 10.1111/j.1523-1739.2008.00987.x
Troëng, S., Chacón, D., and Dick, B. (2004). Possible decline in leatherback turtle Dermochelys coriacea nesting along the coast of Caribbean Central America. Oryx 38, 395–403.
Troëng, S., and Rankin, E. (2005). Long-term conservation efforts contribute to positive green turtle Chelonia mydas nesting trend at Tortuguero, Costa Rica. Biol. Conserv. 121, 111–116. doi: 10.1016/j.biocon.2004.04.014
Wallace, B., and Eckert, K. (2018). Northwest Atlantic Leatherback Turtle (Dermochelys coriacea) Status Assessment. Conservation Science Partners and the Wider Caribbean Sea Turtle Conservation Network (WIDECAST). WIDECAST Technical Report No. 16. Godfrey, IL: WIDECAST.
Wallace, B. P., Tiwari, M., and Girondot, M. (2013). Dermochelys coriacea. The IUCN Red List of Threatened Species 2013: e.T6494A43526147. Available online at: https://dx.doi.org/10.2305/IUCN.UK.2013-2.RLTS.T6494A43526147.en (accessed July 21, 2021).
WIDECAST (2008). Programa de Conservación de las Tortugas Marinas en Playa Gandoca. Available online at: http://latinamericanseaturtles.com/library.php?d=64 (accessed July 21, 2021).
Wilson, L., and Boratto, R. (2020). Conservation, wildlife crime, and tough-on-crime policies: Lessons from the criminological literature. Biol. Conserv. 251:108810. doi: 10.1016/j.biocon.2020.108810
Wilson-Wilde, L. (2010). Combatting wildlife crime. Forensic. Sci. Med. Pathol. 6, 149–150. doi: 10.1007/s12024-010-9179-4
Wittemyer, G., Northrupa, J. M., Blanc, J., Douglas-Hamilton, I., Omondif, P., and Burnhama, K. P. (2014). Illegal killing for ivory drives global decline in African elephants. PNAS 111, 13117–13121. doi: 10.1073/pnas.1403984111
Wood, S. N. (2017). Generalise Additive Models. An Introduction in R. 2nd Edn. Boca Raton, FL: CRC Press.
Woods, C. S. (2015). Addressing prison overcrowding in Latin America: a comparative analysis of the necessary precursors to reform. ILSA J. Int. Comp. Law 22, 533–561. Available online at: https://nsuworks.nova.edu/cgi/viewcontent.cgi?referer=https://scholar.google.co.uk/&httpsredir=1&article=1913&context=ilsajournal/
Keywords: conservation, green criminology, illegal wildlife trade, poaching, sea turtles, situational crime prevention, supply-side trade, Tortuguero
Citation: Pheasey H, Glen G, Allison NL, Fonseca LG, Chacón D, Restrepo J and Valverde RA (2021) Quantifying Illegal Extraction of Sea Turtles in Costa Rica. Front. Conserv. Sci. 2:705556. doi: 10.3389/fcosc.2021.705556
Received: 05 May 2021; Accepted: 13 July 2021;
Published: 23 August 2021.
Edited by:
Meredith L. Gore, University of Maryland, College Park, United StatesReviewed by:
Jessica Bell Rizzolo, Michigan State University, United StatesMatthew Grainger, Norwegian Institute for Nature Research (NINA), Norway
Copyright © 2021 Pheasey, Glen, Allison, Fonseca, Chacón, Restrepo and Valverde. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Helen Pheasey, aGVsZW5waGVhc2V5QGdtYWlsLmNvbQ==