
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Conserv. Sci., 07 June 2021
Sec. Animal Conservation
Volume 2 - 2021 | https://doi.org/10.3389/fcosc.2021.665000
This article is part of the Research TopicConservation CriminologyView all 7 articles
 Raphaël Arlettaz1*
Raphaël Arlettaz1* Guillaume Chapron2
Guillaume Chapron2 Marc Kéry3Elisabeth Klaus1Stéphane Mettaz1
Marc Kéry3Elisabeth Klaus1Stéphane Mettaz1 Stefanie Roder1
Stefanie Roder1 Sergio Vignali1
Sergio Vignali1 Fridolin Zimmermann4
Fridolin Zimmermann4 Veronika Braunisch1,5
Veronika Braunisch1,5Illegal hunting represents a major threat to the conservation of predators, but its impact remains difficult to assess as there are strong incentives to conceal this criminal activity. Attributing declines of carnivores to poaching is therefore an important conservation challenge. We present a case study of the Eurasian lynx (Lynx lynx) in the Swiss Alps (Valais) where the current distribution range is smaller than in the recent past and population density is by ≥80% lower than in the adjacent Swiss Prealps. We tested four hypotheses to explain this lower density: (1) a too low density of camera-traps deployed for lynx surveys in Valais compared to the Prealps (methodological artifact); (2) less favorable environmental conditions around the camera-trap sites; (3) lower densities of the main prey; and (4) poaching. We estimated lynx and ungulate densities and environmental conditions at trail camera sites and could clearly reject the first three hypotheses because: (1) the survey protocol was similarly effective; (2) environmental conditions around the trapping sites in Valais were even more favorable for lynx detection than in the Prealps; and (3) prey supply was even larger in Valais. Concerning hypothesis 4, we discovered a network of illegal lynx traps (neck snares) in the main immigration corridor into Valais from the thriving adjacent lynx population in the Prealps, suggesting intense local poaching. Our findings substantiate the suspicions of long-lasting lynx poaching as a threat to the establishment and survival of the Valais population. The fact that instances of poaching were publicly known since 1995 but remained unabated for at least two decades, until the first conviction occurred, questions the commitment of local authorities to address this case of wildlife crime. Our study demonstrates the need for inquiries about poaching of top predators to be carried out at the highest levels of jurisdiction to avoid any risk of collusion between law enforcement agents and poachers.
Protection of large predators in human dominated landscapes represents an important challenge for contemporary conservation. In particular, large predators may interfere with economic activities, especially animal husbandry, leisure and commercial hunting (Treves and Karanth, 2003), making a smooth coexistence with humans difficult to achieve (Chapron and López-Bao, 2016; López-Bao et al., 2017). Although large carnivores are declining globally (Ripple et al., 2014), some regions of the world have seen these species making an often unforeseen comeback (Chapron et al., 2014). An essential driver of such recoveries, besides an increase of prey species and more favorable public attitudes, is the legal protection of predators (Chapron et al., 2014). In many countries, it is a crime to kill large carnivores outside strictly defined conditions approved by the relevant authorities. For example, in the European Union, the Habitats Directive imposes limitations on legal killing of large carnivores (Epstein and Chapron, 2018).
However, illegal activities remain a persistent threat and recent research has demonstrated that illegal killing of large carnivores can be much more frequent and widespread than it is generally thought. In Scandinavia, Liberg et al. (2012) found that a half of wolf (Canis lupus) mortality was due to poaching and that two thirds of this poaching mortality was not directly observable. Andrén et al. (2006) found that poaching accounted for 46% of the mortality cases of adult Eurasian lynx (Lynx lynx) in Norway and Sweden. In Croatia, Sindičić et al. (2016) investigated mortality causes of lynx and found that poaching accounted for 60% of all recorded fatalities from 1999 to 2013. Heurich et al. (2018) found that the source-sink dynamics of reintroduced lynx populations in Central Europe was essentially driven by poaching. A survey of Czech hunters revealed that more than one third of them knew of instances of lynx poaching, with 10% even anonymously claiming to be the actual perpetrators (Cervený et al., 2002).
As poaching can be such a substantial source of mortality in lynx populations, documenting its exact magnitude is essential for conservation planning. However, since poaching is illegal by nature, there are strong incentives to hide it from law enforcement authorities, making it very difficult to document and quantify. In addition, sometimes it remains unclear whether law enforcement forces are passively complicit in poaching by failing to prosecute poachers when evidence is available or even by advising poachers on how to evade legal prosecution (Liberg et al., 2012).
In this paper, we report long suspected (but until recently unpunished) poaching activities in what is presumably the key lynx dispersal corridor into the southwestern, inner Swiss Alps (Valais). This provides evidence that illegal hunting threatens the population that was reintroduced to Switzerland from the early 1980s and was still thriving in Valais until three to four decades ago. Although suspicions of poaching already existed in that region of Switzerland since the time of lynx reintroduction (Haller, 1992), the first public evidence of lynx poaching emerged in 1995 when a photograph of a hunter portrayed with a rifle and two dead lynx was broadcast by the media (Breitenmoser and Breitenmoser-Würsten, 2008; Arlettaz et al., 2019). However, a local court then accepted the hunter's claims that he had merely found the animals that were already dead, and acquitted him. A more substantial evidence of poaching in this area emerged in 2011 with the discovery (by the authors) of the first active lynx traps. This occurred at the start of our long-term research programme aimed at developing simple and cost-effective methods for investigating the spatio-temporal relationships between large carnivores and their ungulate prey, especially in the context of wolf recolonization into the Swiss Alps. In this programme, we deployed a network of camera-traps to assess the spatial distribution and population densities of large carnivores. This monitoring effort led us to discover unexpectedly low densities of lynx in Valais, reaching <20% of the densities observed in other areas of Switzerland where environmental conditions are fairly similar and where standard lynx monitoring schemes have been carried out for many years (Molinari-Jobin et al., 2001, 2006, 2012; Zimmermann et al., 2011). In addition, our programme revealed an almost complete absence of reproduction in Valais (Biollaz et al., 2016). This low density compromised the research project from the onset by providing too small sample sizes to allow drawing inferences on predator-prey interactions, but it also raised questions about the possible factors causing it.
In this study, we evaluate four plausible hypotheses that could potentially explain the extremely low density of the lynx population in Valais—representing only 12–20% of what might reasonably be expected—as framed in the previous assessment of its status (Biollaz et al., 2016): methodological limitations and caveats linked to the lynx monitoring techniques themselves, suboptimal ecological conditions for the lynx in Valais, and high poaching pressure. Elucidating the causes of this very low lynx density in Valais is of major conservation relevance for the wider distribution of this felid in the Alps because the Valais lynx population could act as a key stepping-stone for further recolonisation of the Italian and French Alps where the species is still rare (Haller, 1992; Molinari et al., 2001; Biollaz et al., 2016).
Four hypotheses offer the most plausible explanations for the restricted distribution range and low lynx density observed in Valais (Biollaz et al., 2016):
1) The first hypothesis proposes that a less dense trail camera-trap network in Valais compared to other areas of Switzerland where lynx monitoring is routinely carried out might lead to the underestimation of local lynx population densities in Valais.
2) The second hypothesis states that there are fewer obligate passages for traveling lynx in the landscape configuration of Valais compared to the Prealps. This might induce a reduced probability of lynx photo-captures at camera-trap sites in Valais.
3) The third hypothesis is that lower local densities of the two main prey species—roe deer Capreolus capreolus and chamois Rupicapra rupicapra—of lynx in Valais (Haller, 1992) preclude lynx densities from reaching values as high as in other, healthy populations in Switzerland.
4) The fourth hypothesis relates to more intense poaching in Valais compared to adjacent regions in Switzerland where lynx thrive. Discussed earlier (Biollaz et al., 2016; Biollaz and Arlettaz, 2017), this is a legitimate hypothesis to investigate because there is some evidence suggesting that poaching may be not uncommon. This includes a hunter posing with two dead lynx (Supplementary Figure 1) and a senator representing Valais in the Swiss parliament officially and merrily claiming in 2010 that the Valais lynx population “can be managed and regulated in total transparency” (Fournier, 2010) although the lynx is a strictly protected species in Switzerland and no permits for lynx culling or regulation had ever been issued to the Valais authorities by the relevant Swiss federal governmental agency.
We conducted this study between 2011 and 2016 in two regions of the Swiss Alps with widely different lynx densities: the Valais Alps (hereafter Valais) in the southwestern Swiss Alps and the northwestern Swiss Alps (hereafter Prealps) situated on the northern outer margin of the Alpine range, immediately adjacent to the northwest of Valais (Gonseth et al., 2001). The Prealps study area comprises parts of the cantons of Bern and Vaud, including Simmental, Diemtigtal, Saanenland and the Pays d'Enhaut, whereas the Valais region is separated by the deep Rhône river valley into the Bernese Alps in the north and the Pennine Alps in the south (Figure 1).
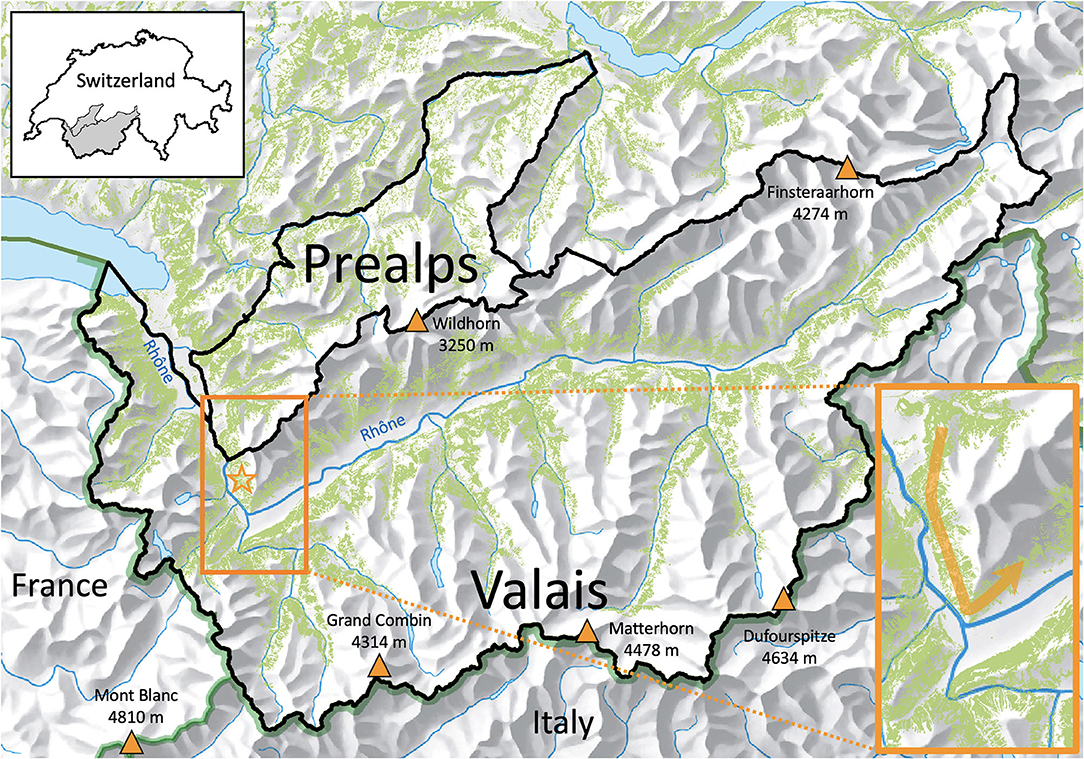
Figure 1. The forest cover (in green) continuum of southwestern Switzerland (forest cover not shown for France and Italy) depicting the distribution of main lynx habitat. Valais is enclaved within high mountain ranges culminating at more than 4,000 m a.s.l., both in the north and south. This isolates the Valais lynx population, with little connectivity to the north (where dense lynx populations occur), east and south (Italy). The forest continuum is higher in western Valais, with an important dispersal corridor linking to the Prealps, where lynx thrive, and France, south of the Rhône river, where lynx are rare. The main dispersal corridor (orange arrow) into the Upper Rhône valley (Valais) from the thriving lynx population in the Prealps is shown in the inset. The star indicates the location of the network of 17 illegal neck-snare traps found in the present study (see also Figure 6).
Although immediate neighbors, the two study areas differ in climate, topography, elevation and forest cover. Valais is characterized by a vast altitudinal gradient, from 372 m to 4,634 m a.s.l., with high elevation areas receiving much precipitation, whereas the valley bottoms are particularly arid (ca 550 mm annual precipitation) due to surrounding high peaks. With a total area of 5,224 km2, 22.2% is covered by forests, the major lynx habitat, distributed in a belt-like shape along the slopes of the main valley and its numerous tributaries, with a timberline at 1,800–2,300 m a.s.l. (Biollaz et al., 2016). Due to effects of topography, forested areas within Valais are well-connected, offering a continuous woodland belt, but are almost totally disconnected from the forested areas situated in the north (where denser lynx populations occur), east or south (Figure 1). The only forested connection with areas outside Valais is in the northwest (canton of Vaud, with a thriving lynx population, Zimmermann et al., 2016)—constituting the main dispersal corridor for lynx locally—and in the southwest, in the Haute-Savoie in France, which has only scarce lynx presence. Six species of wild ungulates are present in the area. Red deer (Cervus elaphus), roe deer and chamois (the latter two species being the main prey of Swiss lynx; Breitenmoser and Haller, 1987) are abundant and distributed in the whole area, whereas populations of ibex (Capra ibex), wild boars (Sus scrofa), and non-native mouflons (Ovis orientalis musimon) occur more locally.
In the Prealps, the topo-climatic conditions are less extreme, with a larger amount of precipitation more evenly distributed across the altitudinal gradient. This study area encompasses 1,080 km2, covering an altitudinal range of 450–2,900 m a.s.l., and is covered by forests that are mostly interspersed with grasslands, with a timberline at 1,700–1,800 m a.s.l. (Breitenmoser and Haller, 1987). The ungulate community is similar to that in Valais, with the exception of the mouflon which is absent. For a typical forest-dwelling species such as lynx, the Prealps thus offer a continuum of suitable (i.e., forested) habitat (Figure 1). Hence, in Valais there is a comparatively less isotropic forest matrix, simply because the Upper Rhône valley is surrounded by the highest peaks of Western Europe (Figure 1). This has clear implications for lynx dispersal: while the Prealps offer only limited barriers to dispersal, the topography of Valais limits any dispersal opportunities into and from the Upper Rhône valley. Except from the aforementioned connectivity with the Prealps in the northwest, dispersal would only be possible through high-alpine passes above the timberline, but these are rarely crossed by lynx (Breitenmoser and Breitenmoser-Würsten, 2008).
In both areas, we restricted our surveys to an elevational range from the lowest foothills at 500 m up to 2,000 m a.s.l., i.e., excluding the valley bottoms with dense human population and the mountain peaks devoid of forests. This resulted in survey areas of 2,555 km2 for Valais and 957 km2 for the Prealps.
Lynx data were obtained from two different, ongoing monitoring schemes and research programmes. Whereas lynx presence and density in the Prealps (and only recently in Valais, e.g., Zimmermann et al., 2014) were assessed every second to fourth year by the Carnivore Ecology and Wildlife Management Foundation (KORA), lynx density estimates for Valais were obtained mostly from the camera-trap network deployed by the Conservation Biology Division of the University of Bern since 2011/2012 as part of a study of the spatio-temporal relationships between large carnivores and their ungulate prey (Supplementary Figure 2A).
In the Prealps, 79 trail camera-trapping sites were selected by KORA in a stratified manner in winter, placing a trapping site in every second cell of a sampling grid, with each square cell measuring 2.7 km in width, which resulted in a trap density of 4.7/100 km2 (Zimmermann et al., 2016). At each location, two camera-traps equipped with white flash (Cuddeback, Inc. Green Bay, Wisconsin, USA) were installed (not symmetrically facing each other for avoiding flash interference) in order to facilitate individual lynx identification by assessing the fur pattern on both body flanks (Zimmermann and Foresti, 2016).
In Valais, the forested area below 2,000 m altitude corresponding roughly to two thirds (~3,700 km2) of the cantonal territory was divided into 34 grid cells of 100 km2 size (10 km side-length), each cell being attributed to one of seven topographically defined landscape compartments (Figure 2; see also Figure 2 in Biollaz et al., 2016). In each of these 34 cells, three camera-traps fitted with a movement and heat detector and an infrared flash (Reconyx PC900 Hyperfire Professional, Inc. Reconyx, Wisconsin, USA) were placed by the University of Bern, resulting in a total of 102 camera-traps, i.e., a camera-trap density of 3/100 km2 (Biollaz et al., 2016). Yet, contrary to the KORA approach the camera-traps were used singly; simultaneous photography of both flanks of the lynx body was thus not possible.
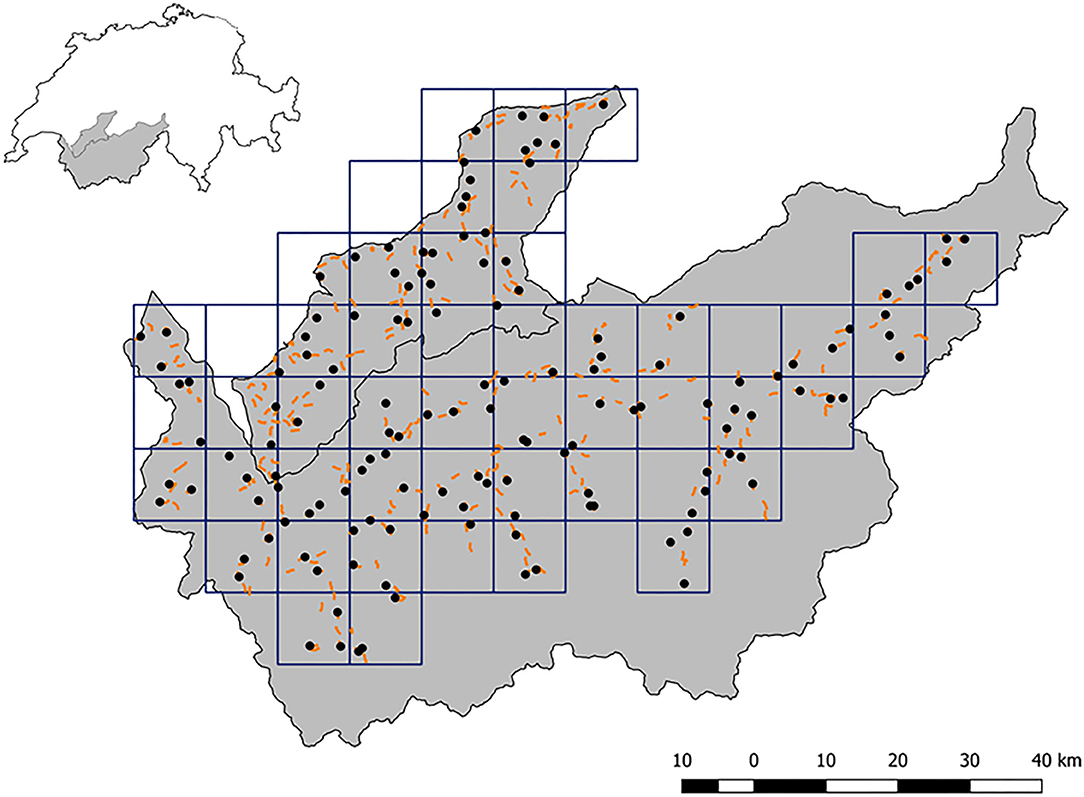
Figure 2. The study areas in the Prealps (upper perimeter) and Valais (lower perimeter; see Figure 1) and their location within Switzerland (upper left inset). The grid (10 × 10 km) indicates the raster used for sampling stratification and orange lines show the 313 1-km long transects along which snowtracking of wild ungulates was performed. The black dots depict camera-trap locations.
To allow a direct comparison of the camera survey outcomes, the same monitoring scheme as in the Prealps was applied during the winter 2013/2014 to the northwestern part of Valais by KORA, in parallel to the ongoing survey scheme by the University of Bern. A comparison of the two simultaneous surveys carried out by the two research groups in the same area delivered information about the influence of camera-trap density on the number of individual lynx identified, and therefore on lynx density estimates (see Biollaz et al., 2016).
In both study areas, camera-traps were placed along forest roads, trails or game paths. Obligate passages, i.e., narrow paths toward which the movements of animals are strongly directed by topographic circumstances or other natural obstacles, notably gaps among rocky cliffs and dense vegetation formations, were chosen in priority wherever possible (Biollaz et al., 2016). The camera-traps were operational between mid-November and late March (University of Bern) or January-March (KORA).
Ungulate data were collected using snowtracking along 1-km transects (Valais: n = 218, Prealps: n = 95) in winter 2015/16. In order to place transects in a stratified manner, the two study areas were divided into grid squares with a side-length of 10 km (Figure 2; 34 and 16 squares in Valais and in the Prealps, respectively) similar to the camera-trap sampling design. In each square, on average six transects (range: 1–10) were placed so as to represent the altitudinal and environmental gradients while accounting for accessibility, topography and human safety with respect to avalanches. Transects were visited twice during each winter (December to March). All tracks found in the snow of the two main prey species of lynx (roe deer and chamois) were recorded (Supplementary Data Sheet 3). Based on imprint size and track configuration, we estimated the minimum number of individuals present during each visit. Multiple individuals were counted if tracks of different sizes (i.e., from different sexes or age classes) or individuals traveling together in a group could be distinguished, using a conservative approach: when in doubt, we always recorded the lower number of individuals (for details see Roder et al., 2020).
We recorded 14 variables to determine the factors influencing detection probability at camera-trap sites. Eleven variables were mapped in the field on a local scale, around the camera-trap sites, while the remaining three were assessed on a landscape scale from extant geographical data (Table 1; Supplementary Data Sheet 2).
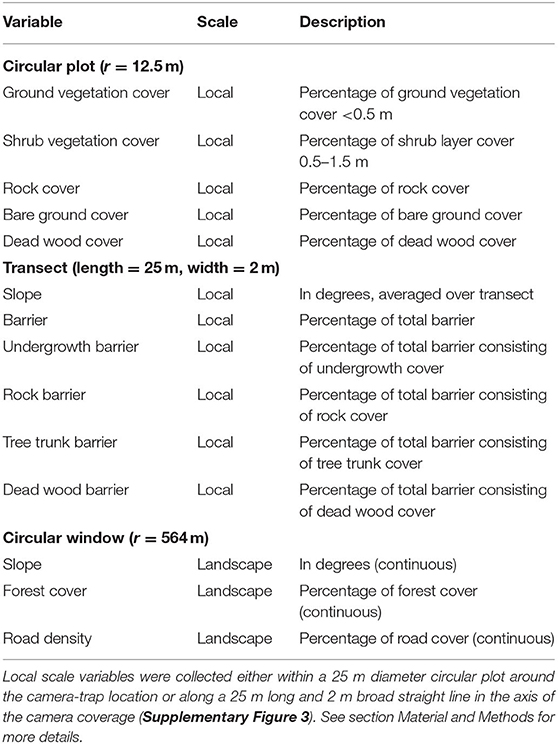
Table 1. Variables describing the surroundings of the camera-trap sites, obtained from field mapping (local scale) or extant databases (landscape scale).
Topographic and vegetation conditions at all camera-trap sites were assessed between June and August 2016. Variables were acquired within a circular plot (12.5 m radius) directly around the camera-trap location or from a 25 m long (matching the plot diameter) and 2 m wide transect perpendicular to the expected lynx trajectory (along a road or path), i.e., exactly in the axis of the camera coverage (Supplementary Figure 3). Within the plot, we estimated the vegetation cover of both ground (<0.5 m) and shrub (0.5–1.5 m) layers, rocks, bare ground and deadwood. Along the transect, we estimated the total percentage of barriers defined as any obstacle that would constrain lynx movement at a height of 0.3–1 m above ground. In addition, we separately recorded the percentage of dense undergrowth, rocks, dead wood and tree trunks, as well as streams wider than 1 m. Slope was measured using a smartphone app (Slope Angle, Radislav 2015). On a landscape scale, average slope, percentage of forest cover and forest road density were assessed within a radius of 564 m around the camera trapping sites, which corresponds to an area of 1 km2. Spatial analyses were performed using QGIS v. 2.18.9 (QGIS Development Team, 2017).
The environmental variables we used for predicting area-wide relative ungulate densities (Table 2) were extracted from existing digital information, so as to form basic grid layers with a 25 × 25 m resolution (Supplementary Data Sheet 3). We obtained data on topography (altitude, roughness, slope, topographic position index) from the Swiss national digital elevation model (DEM). Land cover data (forest, rock, scree, waterbodies, anthropogenic areas, roads and railways, ski lifts) were derived from vector25, a digital vector-map regularly updated by the Federal Office for Topography Swisstopo (https://www.swisstopo.admin.ch), except for grassland cover which was obtained from the Arealstatistics of Switzerland (https://www.bfs.admin.ch). Forest type information originating from Landsat5 data was provided by the Swiss Federal Office of Statistics (https://www.bfs.admin.ch). Winter ambient temperature and precipitation came from the Worldclim dataset (http://www.worldclim.org), which was downscaled from a 1-km2 raster to the resolution of 100 x 100 m based on the SRTM-V4 digital elevation model following the method described in Zimmermann and Roberts (2001). The densities of sheep and goats, as potential competitors of wild ungulates, were estimated by dividing the number of these domestic animals held in each community by the area of alpine pastures present on the territory of the same community. Finally, information about national wildlife reserves (where hunting is legally banned) was obtained from the Federal Office for the Environment (FOEN). All variables were assessed within a 200 m buffer around the transect (corresponding to an area of 50 ha, which approximates the average home range size in winter for the two main ungulate prey species (Hamr, 1985; Mysterud, 1999) by calculating the proportion (boolean variables) or the average (continuous variables) within this area. The preparation of the variables was performed using QGIS v. 2.18.9 (QGIS Development Team, 2017) and ArcGIS Release 10 (ESRI, 2011).
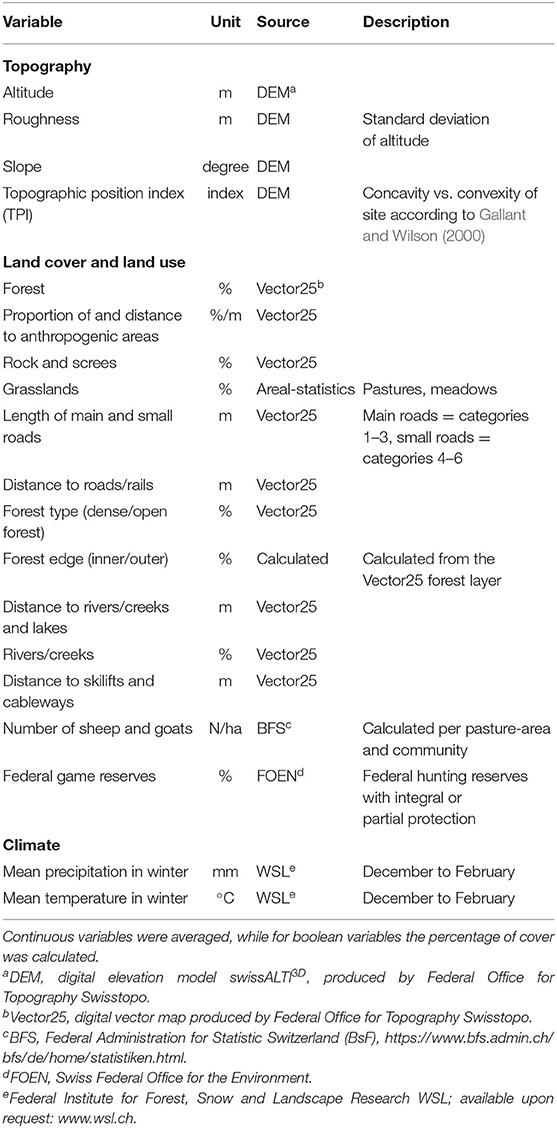
Table 2. Environmental variables used to predict relative ungulate densities, extracted from buffer zones around the transects or circular moving windows of 50 ha (see section Material and Methods for more details).
We modeled the detection probability of ungulates as a function of survey conditions during the transect walks and used several covariates of snow status (Table 3). Snow depth, snow condition and the percentage of snow cover in open and forested areas were recorded directly during snowtracking. In addition, the daily amount of fresh snow at all weather stations in the study areas (56 in Valais and 30 in the Prealps) was obtained from the Swiss Institute for Snow and Avalanche Research (WSL/SLF). From these data, we calculated the number of days elapsed between the last snowfall and the date of the transect survey (days since last snowfall, Table 3), assigning each transect to the nearest weather station. We also transformed this continuous variable into a categorical variable describing snow age. Finally, we recorded the amount of fresh snow that had fallen on the last day with snowfall (Supplementary Data Sheet 3).
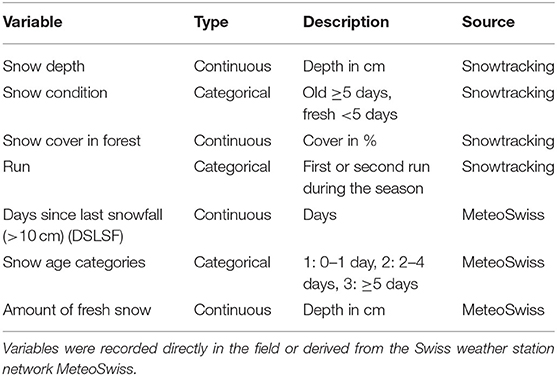
Table 3. Snow variables used for predicting the detection probability of ungulates based on snowtracking survey data.
The serendipitous discovery of active lynx traps during the winter 2011/2012 triggered an intensive search of the whole Valais study area in the subsequent years to evaluate the magnitude of this poaching activity. Several traps were operational until 2015. Such contemporary traps had never ever been observed outside Valais, neither in the Prealps nor elsewhere in Switzerland, despite decades of lynx research throughout Switzerland, notably by KORA. Our search consisted of systematically inspecting trees along the 313 1-km long transects (218 in Valais and 95 in the Prealps; Figure 2) for evidence of current and recent historical lynx trap installation. In effect, lynx trapping systems deployed in Valais are mostly neck snares typically placed in an obligate passage, i.e., in a bottleneck of microtopographic configurations, such as between big boulders and/or dense vegetation formations, or next to a rock cliff. There, a unique possible path is created by accumulating tree branches in the surroundings so as to form a sort of funnel that channels lynx movement (Figure 3). A quasi-circular neck snare is placed vertically at lynx head height, constituting the only possible way through the vegetation, forcing the lynx to pass its head through the snare. When the trap is armed, the snare is connected via a long wire to a large stone that is positioned 2 m above the ground along a tree trunk. Between the snare and the stone, on the trunk surface, there is a hook (often a simple, but thick, folded nail) through which the wire passes. This piece is positioned much higher than the stone itself, which therefore puts the whole system under tension when armed. When a lynx enters its head into the snare and then pulls at it, it triggers the system, which ejects the individual into the air and strangles it (see the contorted lynx heads in Supplementary Figure 1). Our quest focused principally on the presence of folded nails, hooks, pulleys and wires on tree trunks, which are often the only parts of a trap system that occur at inactive trapping sites.

Figure 3. An active neck-snare trap discovered in 2015 in the lynx entrance corridor into Valais from the Prealps. (1) This illegal capture system has been installed in an obligate passage along a wildlife trail; branches have been added in the surroundings to augment the channeling effect, i.e., to steer the lynx toward the snare (a bike brake wire) located within the orange circle; (2) a close-up view of the neck snare itself (cable visible within the orange circle); (3) the trapping system viewed from the other side: the position of the snare is again indicated by the orange circle; the cable bending the system goes through a hook positioned high on the thin tree trunk and goes down toward the right; the system is put under tension with a large stone (highlighted in orange on the right); (4) at an inactive trapping site, sometimes only the middle hook positioned high above the ground is visible. A video showing another active system can be obtained from the authors upon request.
To compare trapping efficiency with respect to camera-trap density, we compared the number of distinct individual lynx identified by the two survey operations of KORA and University of Bern, both conducted in the northwestern part of Valais in the same winter 2013/2014 (Biollaz et al., 2016). To identify individuals and avoid double-counting, all recorded lynx photographs were compared with each other, as well as with the photographs of all identified lynx stored in the KORA database since 1998. Individuals could be identified in most cases through specific fur patterns, even when only one flank of a lynx was photographed. Individuals that could not be identified due to insufficient photograph quality were dropped from the analyses. The number of identified lynx was then related to the total area of suitable lynx habitat in the two studies (as modeled by Zimmermann, 2004) to estimate lynx density per 100 km2 of suitable habitat, given that their study areas slightly differed in size. Due to the high density of camera-traps, long duration of surveys and the very small size of the lynx population in Valais (see Results), we assumed that detection error was negligible. On the one hand, Zimmermann and Foresti (2016) had calculated a cumulative probability of detection of 0.92 for a much larger part of the Swiss lynx population that had been camera-trap surveyed by KORA based on a similar design as deployed in the present study. On the other hand, perfect detectability has been corroborated by KORA for the northwestern Valais population: a regular capture-recapture approach yielded an estimated population size of 5 lynx (95% confidence interval: 5–5; Zimmermann et al., 2014), which was also the observed number of unique animals. We therefore did not apply capture-recapture methods for assessing lynx population size based on the data collected by the University of Bern, which photographed the same lynx as KORA (see section Results). Instead we merely relied on the number of identified lynx as a measure of population size.
In our comparison of environmental conditions prevailing at camera-trap locations in normal (Prealps) vs. extremely low (Valais) lynx density areas, we first tested for differences in topographic and vegetation conditions (Table 1) using a Wilcoxon rank sum test. Second, we compared topographic and vegetation conditions between camera-traps with and without lynx events to identify the conditions that affected lynx detection in the two study areas. For that analysis, we used data from two winters (2013/2014 and 2015/2016) for the Prealps and three consecutive winters (2013/2014, 2014/2015 and 2015/2016) for Valais. This choice was based on the fact that camera-traps with lynx captures in Valais were much scarcer than in the Prealps. Each camera-trap with at least one lynx detection within this period was classified as “detection” (“non-detection” otherwise), resulting in 26 out of 102 camera-trap sites with lynx detections (25%) in Valais, compared with 56 cameras-traps out of 79 (71%) in the Prealps (Supplementary Data Sheet 2). We then modeled the probability of lynx detection as a function of environmental conditions at camera-trap sites using generalized linear mixed models (GLMM, R package lme4; Bates et al., 2014) with the study region as random factor. In a first step, we tested for correlations between explanatory variables using the Spearman's rank correlation coefficient. Out of the pairs or groups of correlated variables (|rs| > 0.5), only the one with the better fit in univariate models (according to Akaike's Information Criterion AIC; Sakamoto et al., 1986) was retained for subsequent analyses. Variables that were significant in a univariate analysis were then included into a multivariate analysis. To identify the most parsimonious model, we tested all possible variable combinations using the dredge function in the R package MuMIn (Bartón, 2014) and selected the AIC-based best model.
To obtain detection-corrected relative abundance (hereafter called “abundance”) estimates of roe deer and chamois along the transects, N-mixture models were fitted with the R package unmarked (Fiske et al., 2017), taking into account the sampling occasion (i.e., first or second survey as described in Roder et al., 2020), with snow predictors as detection variables (Table 3) and environmental predictors (Table 2) as habitat variables (Roder et al., 2020; Supplementary Data Sheet 3). We used Poisson models for roe deer abundance and zero-inflated Poisson models for chamois abundance. We started calibrating the models using only detection variables. First, among the pairs or groups of correlated variables (Spearman's |rs| > 0.5) significantly influencing detection probability, we chose the one that performed best in univariate models, according to AIC corrected for small sample sizes (Akaike, 1974). Then, models including all possible combinations of variables significantly affecting detection probability were tested and ranked according to their AICc using the dredge function of the R package MuMIn (Bartón, 2014). Once the best variable combination for detection probability was identified, we additionally included habitat variables into subsequent modeling, adopting the same procedure as above. The best model for each species was then used to predict relative densities (hereafter called “densities,” calculated as the minimum number of individuals per 50 ha) across the whole study area. For this purpose, the proportion or average of the relevant environmental predictors (Table 2) were calculated within a circular moving windows of 50 ha (radius of 399 m) around each pixel of the study area. We restricted our predictions to the areas within the same altitudinal range (500–2,000 m a.s.l.) and with a similar amount of forest cover (≥14.1%) as found in the 200 m buffer zones around the transects. Additionally, we predicted overall ungulate abundance (number/50 ha) and overall prey biomass (kg/50 ha), assuming average species body masses as reported by the Valais hunting statistics (https://www.vs.ch): 17.5 kg for roe deer (years 2012–2015) and 21.5 kg for chamois (year 2005; both after evisceration). To compare prey densities between the two study areas, a regular grid of 1-km side length, based on the Swiss grid coordinate system, was applied to the whole study area. For each grid cell, we calculated the average densities of roe deer and chamois separately, of both species pooled, and of their pooled average biomass. Then, the densities per cell in both regions were compared using a Wilcoxon rank sum test (Wilcoxon, 1945). All statistical analyses were performed in R v. 3.3.4 (R-Core-Team, 2014).
All signs of recent historical and contemporary presence of lynx “snare-ejecting-strangling traps” (hereafter “neck snares”) encountered along our 313 1-km survey transects were documented with precise GPS coordinates and photographs (Supplementary Data Sheet 4). In the area where evidence of illegal trapping systems was found, we intensified the search for other locations of these snares by walking the majority of trails around the spot, progressing away from the neck-snaring site until no hints for further illegal trapping sites were found over a maximum of 2 km along any such trail.
The two independent surveys carried out during the winter 2013/2014 by the University of Bern and KORA photographed six different lynx in total (Supplementary Data Sheet 1; details in Biollaz et al., 2016 and Zimmermann et al., 2014). Each team photo-trapped five distinct individuals (Bern: B395, B396, B397, B398, B408; KORA: B395, B396, B397, B398, B399). The lynx B399 portraited only by KORA was also photographed several times by the University of Bern in the same winter but outside the reference area of the present study, in the southwest (Supplementary Data Sheet 1). The lynx B408, photographed by the University of Bern but not by KORA, was known to the latter from another monitoring scheme in the northwest of the area considered in this study. The estimated lynx densities obtained by the University of Bern and KORA were comparable (and extremely low!), with 0.83 and 0.92 individuals per 100 km2 of suitable habitat, respectively. Thus, the higher density of the camera-trap network deployed by KORA had no noticeable effect on the number of photo-captured individual lynx, which rejects hypothesis 1. Therefore, large differences in lynx density between the Prealps and Valais cannot be explained by the large methodological differences in the survey schemes.
Camera-trap locations at Valais study sites were characterized by steeper slopes, both near the camera-trap (local scale: Valais: 42.6°; Prealps: 37.2°; W = 3,073, p = 0.006) and in wider surroundings (landscape scale: Valais: 30.0°; Prealps: 26.9°; W = 2,656, p < 0.001), higher proportions of rocks (Valais: 9.8%; Prealps: 3.0%; W = 3,104, p = 0.003), obstacles and barriers (Valais: 42.3%; Prealps: 31.6%; W = 2,812, p < 0.001) on a local scale, and by higher forest cover (Valais: 62.1%; Prealps: 55.7%; W = 3129, p = 0.01) and density of forest roads (Valais: 6.9 m/km2; Prealps: 5.7 m/km2; W = 2,826, p < 0.001) on a landscape scale. Lynx detection probability at a camera-trap site was only correlated with slope on the landscape scale, with steeper terrain associated with higher detection probabilities (GLMM, intercept = −3.536; SE = 1.683, p = 0.036; slope = 0.136, SE = 0.041, p < 0.001). These results reveal that the expected detection probability should actually be higher in Valais than in the Prealps. This refutes the hypothesis that different settings for camera-trapping could be the reason for a lower lynx density observed in Valais compared with the Prealps.
Average per-individual detection probability for roe deer during snowtracking was 49%. It peaked when the snow was <5 days old and fresh snow was not too deep. Roe deer abundance was best explained by an intermediate forest cover, low winter precipitation and the vicinity of streams and creeks (Table 4). Detection-corrected predictions of (minimum) roe deer abundance at the transect sites ranged from 0.23 to 11.11 (mean: 3.81) individuals. Predicted roe deer densities across both study areas ranged between 0.44 and 6.61 individuals per 50 ha (Figure 4A).
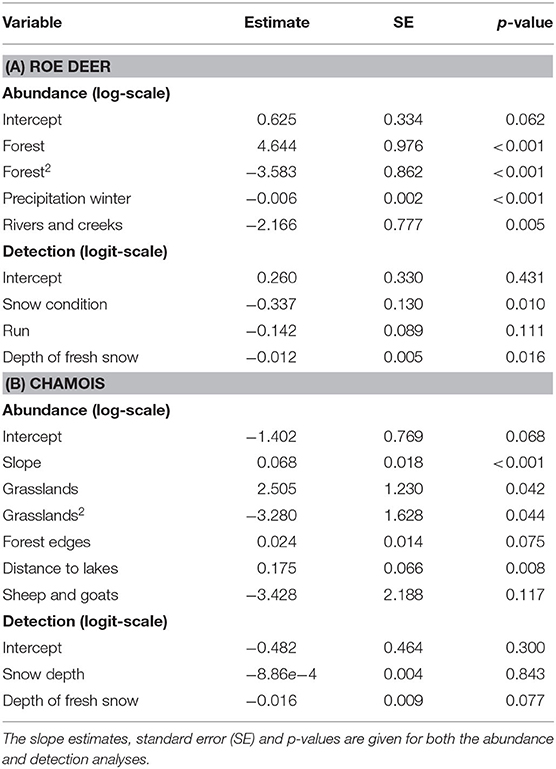
Table 4. The best N-mixture models for predicting relative densities of roe deer (A) and chamois (B).
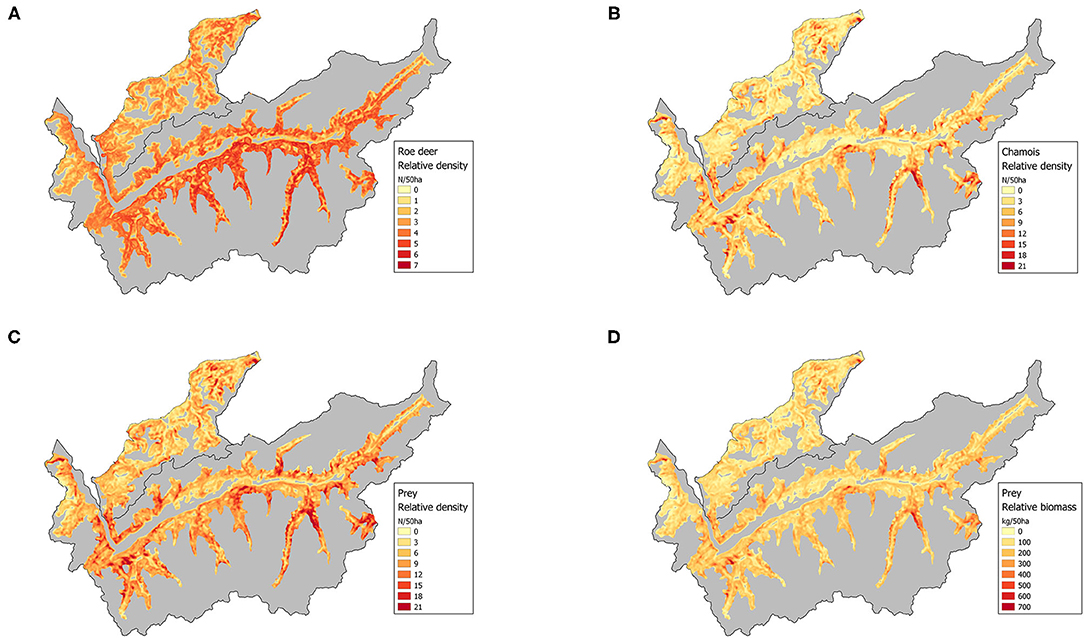
Figure 4. Maps of predicted relative densities of (A) roe deer (ranging from 0.44 to 6.61 individuals per 50 ha); (B) chamois (0–37.76 per 50 ha); (C) both species grouped (0.47–42.59 per 50 ha) and (D) their biomass (8.38–896.39 kg per 50 ha). Predictions were restricted to the colored area.
For chamois, mean per-individual detection probability was 30%. The depth of fresh snow had a negative, but non-significant effect on detection probability. Chamois abundance was positively influenced by slope, intermediate grassland cover and by distance to lakes (Table 4). Detection-corrected predictions of abundance along the transects ranged from 0 to 10.03 (average: 1.41) individuals, while predicted chamois densities in the two study areas ranged from 0 to 37.8 individuals per 50 ha (Figure 4B).
Estimated densities for both species combined ranged from 0.5 to 42.6 individuals per 50 ha and their total estimated biomass per 50 ha from 8.4 to 896.4 kg (Figures 4C,D). Overall prey densities differed between the two study areas. With an average of 2.41 individuals per 50 ha in the Prealps compared to 3.29 per 50 ha in Valais (Wilcoxon test: W = 679,280, p < 0.001), roe deer densities were significantly higher in Valais (Figure 5A). A similar pattern existed for chamois densities (Wilcoxon test: W = 888,420, p < 0.001), with an average of 3.44 individuals per 50 ha in the Prealps and 4.75 individuals per 50 ha in Valais (Figure 5B). Total prey density (Prealps: 5.85; Valais: 8.04) and biomass per 50 ha (Prealps: 116.06 kg; Valais: 159.61 kg) were also substantially higher in Valais (Wilcoxon test: W = 764,310, p < 0.001; W = 779,900, p < 0.001, respectively; Figures 5C,D). Thus, the third hypothesis that a lower lynx density in Valais could be caused by lower prey supply in this area in comparison with the Prealps was rejected.
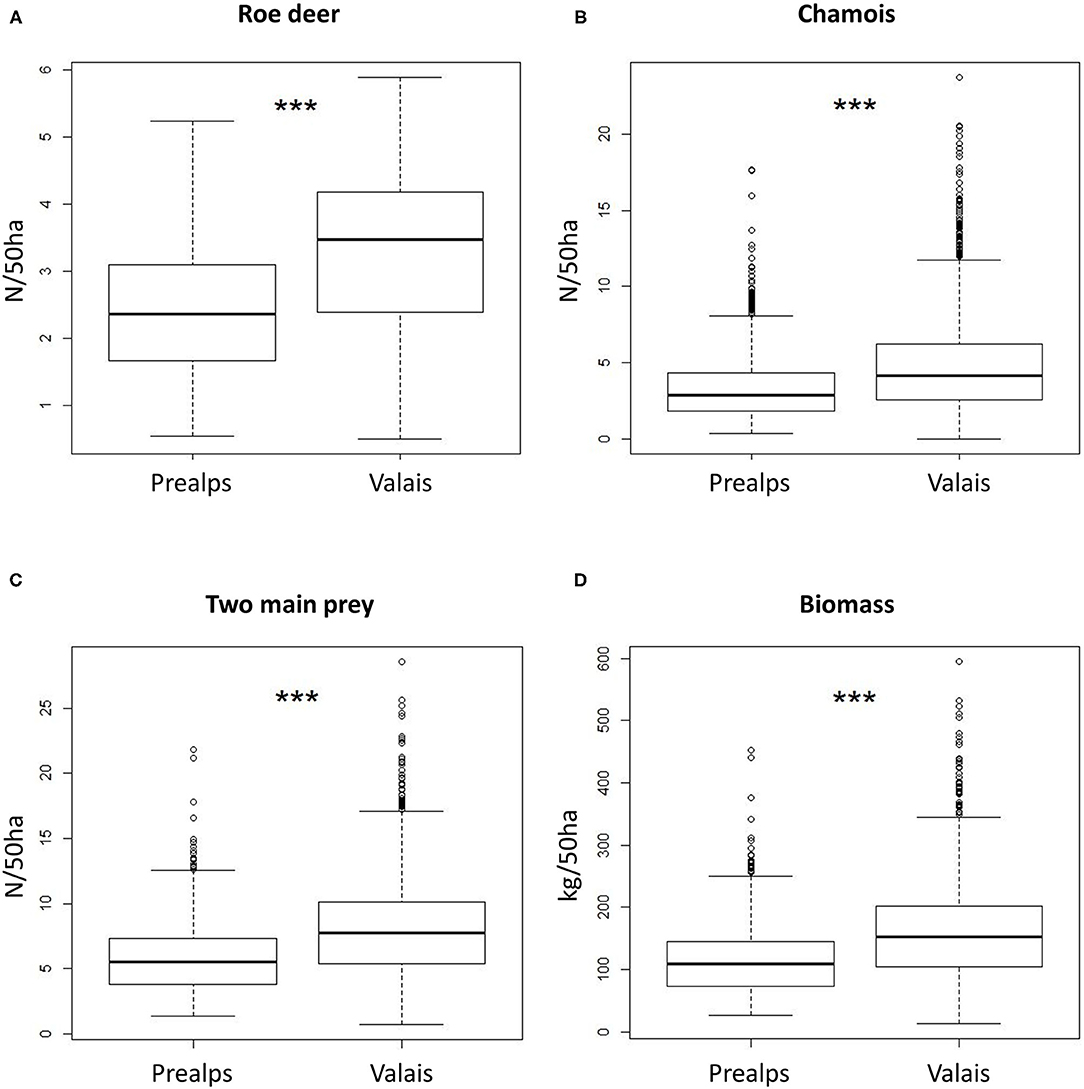
Figure 5. Statistical comparison of wild ungulate relative densities and biomass between the northwestern Prealps and Valais: (A) roe deer density; (B) chamois density; (C) density of these two main prey species grouped and (D) their biomass in number of individuals and kg per 50 ha, respectively. ***P < 0.001 (Wilcoxon test).
We found evidence of current and recent use of lynx traps only in a specific area surrounding the village of Champex d'Alesses above Dorénaz (postcode, 1905). A more in-depth survey in 2015 along 39 km of trails over a total area of 15 km2 around the village enabled us to discover the presence of 17 different neck-snaring sites, with evidence of their set-up in the past and present (Figure 3; Supplementary Data Sheet 4). The 17 trapping systems were concentrated in an area of ca 4 km2, thus at a density of ca 4 neck snares per km2 (Figure 6). Three traps were armed on the very day when we discovered them, while the other 14 were inactive, with their most recent years of deployment unknown. We dismantled all active traps and delivered them to the cantonal police. In contrast, no signs of similar trap systems were discovered along any of the other 1-km transects in the two study areas, suggesting that snaring was a fairly local activity around Champex d'Alesses. However, the network of traps covered exactly the main immigration corridor for lynx into Valais from the Prealps, i.e., from the only healthy lynx population adjacent to Valais from where immigration might reasonably take place (Figures 1, 6). It appears likely that the strategic location of this “entrance gate of Valais” against immigrating lynx may have leveraged its substantial impact by greatly reducing the immigration of lynx into Valais. We therefore failed to reject the hypothesis that poaching activities reduce lynx densities in Valais.

Figure 6. The Google EarthTM views (upper projection from the southwest; lower projection from the northwest; compare to Figure 1) of the entrance corridor into Valais from the lynx-populated Prealps showing 39 km of trails (orange lines) explored for neck-snare traps, with an indication of the 17 neck-snaring sites (orange dots) discovered (two neck-snare traps are so close that they appear as a simple dot). The forest belt and the barriers for dispersal shaped by high mountain chains are obvious.
Our study aimed to elucidate the reason why the current lynx density in Valais is only about 20% of the expected one, with an almost complete absence of reproduction for many years (Biollaz et al., 2016). Our analysis rejected all methodological and biological hypotheses but provided evidence in support of intense local and strategically placed trapping pressure as a plausible explanation for the currently restricted distribution and extremely low density of lynx in Valais. Although evidence for poaching of lynx in this study exclusively proves neck-snaring in a key specific location, illegal hunting in other parts of Valais, probably with other methods such as guns, has also been documented (Breitenmoser and Breitenmoser-Würsten, 2008).
First, we found that the lower density (by 36%) of camera-traps in Valais compared to the Prealps did not materially affect the detection probability for individual lynx. This means that the two monitoring schemes operated in the Prealps and Valais by KORA and the University of Bern, respectively, yielded comparable estimates of lynx population size (Biollaz et al., 2016). Second, we found that the environmental conditions surrounding the camera-trap sites (which were preferentially placed on obligate passages) should have made it even more likely for lynx to be photo-captured in Valais than in the Prealps. This suggests that the probability of detecting the presence of a lynx should be higher in Valais than in the Prealps. Third, the density and total biomass of the two key prey species, roe deer and chamois, were substantially higher in Valais than in the Prealps, indicating that a reduced prey base cannot explain the much lower lynx population density in Valais (Zengaffinen, 2016). In contrast, the fourth hypothesis on the effect of poaching was supported by the discovery of a dense network of neck-snare traps in the principal immigration corridor into the Upper Rhône valley (Valais) from the dense lynx population occurring in the Prealps to the north. Poaching of lynx thus remains as the only plausible limitation of lynx survival in Valais.
Forest corridors are essential for lynx dispersal. Contrary to other top carnivores such as wolf and bear (Ursus arctos), which are regularly observed in open alpine habitat well above the timberline, lynx are typical forest-dwellers even during long-distance dispersal. There is only one radio-documented case of a lynx dispersing into Valais directly from the north over a high Alpine pass, out of several dozen radio-tracked dispersal routes (Biollaz et al., 2016). The almost exclusive possibility for lynx to enter Valais is therefore through the aforementioned forested corridor (Figure 1 and Biollaz et al., 2016). It is precisely in this unique narrow and steep wooded corridor where the dense network of illegal traps was located. With a very high density of four neck snares per km2, it appears highly likely that this illegal trapping effort could have a major effect on lynx population dynamics in the entire Valais, although the precise magnitude of its impact remains unknown.
Biollaz et al. (2016) documented the existence of a gradual spatial decrease in lynx density from that entry corridor in the west toward the east and south of Valais (Figure 2 in Biollaz et al., 2016), with some of the main tributaries of the Pennine Alps (south of the Rhône), such as the valleys of Hérens and Anniviers, being almost devoid of any lynx today (Biollaz et al., 2016). This has recently been corroborated by further camera-trap surveys carried out by KORA (Zimmermann et al., 2019) and contrasts sharply with the situation in the 1980s, when the species occurred over a much wider area of Valais (Haller, 1992), notably south of the river Rhône and within the aforementioned side valleys. The question that arises then is whether the mortality generated by the newly revealed local neck-snare network could have reduced the overall Valais lynx population to the extremely low density observed at present, which contrasts so strongly with the recent past situation (Haller, 1992), with the projections of the habitat suitability model by Zimmermann (2004) and with expectations based on the highly abundant prey supply as evidenced in the present study. While evidence for illegal hunting of lynx in this study stems exclusively from neck-snaring in a specific area, poaching of lynx in other parts of Valais has already been documented from stuffed lynx (Breitenmoser and Breitenmoser-Würsten, 2008). In effect, some Swiss hunters publicly claim to regulate lynx because they believe hunting is threatened by the return of top predators (Breitenmoser, 1998; Ceza et al., 2001; Lüchtrath and Schraml, 2015). Therefore, our interpretation of the above-mentioned lynx density gradient is that poaching, possibly using methods other than neck snares (e.g., guns), is taking place throughout Valais at some unknown level, with lynx steadily trying to colonize the Rhône valley from the thriving, better protected populations in the Prealps. A fraction of these potential immigrants would have been continuously removed by the neck-snare network situated in the main entry corridor (Filliez, 2013), impeding immigration to compensate for the probable poaching-related losses within Valais. Hence, if illegal lynx trapping may possibly not be the only cause of the extremely low lynx population density at present, it certainly contributes to it.
A remarkable aspect of the illegal poaching activity using snares is that it had been officially recognized since at least 1995 by the cantonal game wardens and administrative authorities, as assessed by the NGOs Pro Natura (Ceza et al., 2001) and WWF (WWF Valais, in litt.) and even by Swiss national television (Moser, 2016). In that year, a licensed Valais hunter (hereafter X) had been portrayed, not far from his place of residence and from the neck-snare trap network we discovered, proudly posing with a gun and two dead lynx, which had obviously been killed by strangulation, based on the contorted posture of their heads (Supplementary Figure 1, see also Breitenmoser and Breitenmoser-Würsten, 2008; Arlettaz et al., 2019). The photograph was sent anonymously to the federal wildlife service which reported to have forwarded it to the Valais judiciary (Table 5). In 2013, X claimed in an interview to a Swiss magazine that he had “trapped to death 10 lynx with snares” (Filliez, 2013), thus corroborating our hypothesis of a possible population-level impact of the neck-snare trap network. Following this broadcast, the Game and Fishery Service of Valais, together with WWF and Pro Natura, filed a charge against X, but the court dismissed the case in 2014, arguing that the investigations by local police and game wardens could not provide evidence of the existence of any operational traps while the hunter claimed that the 1995 photograph displayed two road-killed lynx (WWF Valais, in litt.). One may question the quality of the inquiry in 2014 given that in 2015 we discovered a network of 17 snaring systems, none being of recent installation. Among them, three were armed, i.e., active in operation. We dismantled the traps and delivered them to the cantonal criminal police, which is the highest regional judicial authority, while filing a formal complaint against X who was eventually convicted for use of illegal hunting devices after his DNA was found on traps (Figure 3, Table 4; Ministère public du Canton du Valais, 2015).
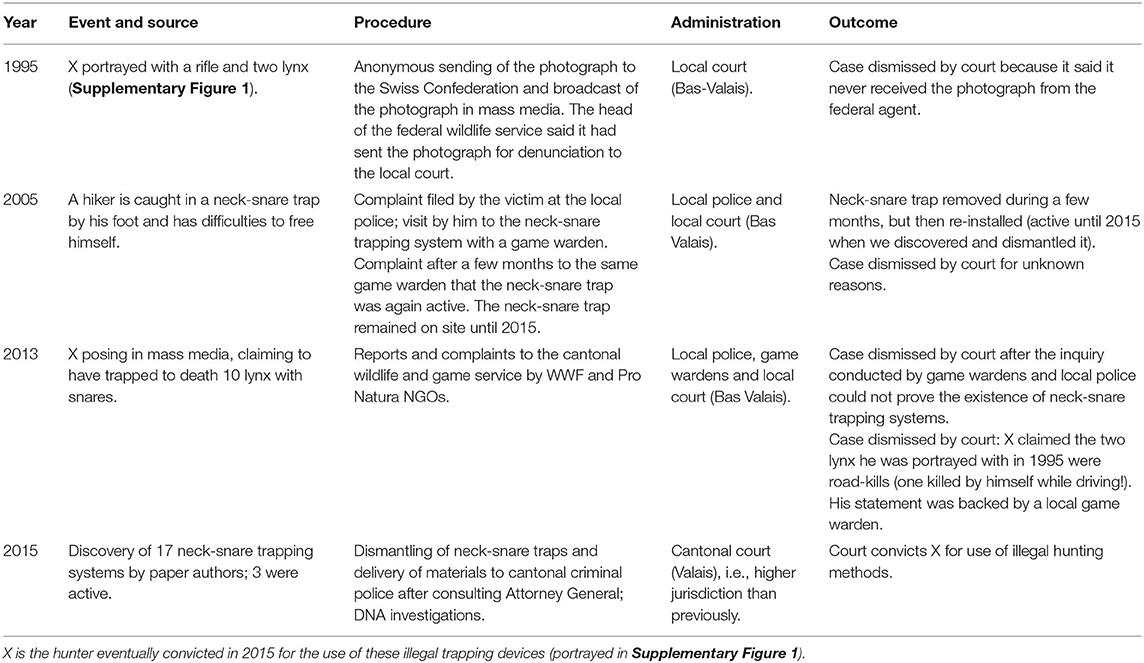
Table 5. History of lynx poaching with neck-snare traps in Valais, southwestern Switzerland, based on media broadcast and justice decisions.
To understand the broader context, it is worth mentioning that in 2005 a hiker was caught by the foot in one of these neck-snare traps (Moser, 2016). He filed a lawsuit at a local police station and was accompanied by a cantonal game warden who took photographs of the trapping system on the spot. Although this resulted in the temporary removal of the trap, the victim found the same neck-snare trap operational again only a few months later, where it remained in situ until we dismantled it in 2015. This suggests that the same trap network may have been operating for years, if not decades. It was clear that the intervention of local game wardens was not sufficient to stop these illegal activities during at least a full decade.
Our study, as well as other facts including those provided above and reported by media and the judiciary since 1995, supports the hypothesis that lynx poaching had long taken place in Valais, and that—if strategically placed—illegal activities of perhaps a single person may represent a serious threat to a small and highly isolated population (Biollaz et al., 2016). It is almost certain that lynx trapping continued unabated until we approached the cantonal Attorney General in 2015. From that very moment, it was the cantonal criminal police, instead of local law enforcement agents, which was mandated for conducting inquiries about carnivore trap-poaching (Ministère public du Canton du Valais, 2015). This seems to have triggered a series of serious investigations (including the decisive use of DNA analysis) for the first time, which fairly rapidly led to a conviction (Ministère public du Canton du Valais, 2015).
Poaching of large carnivores may be a source of additive mortality that can have severe consequences, especially for small populations and even more so when they are highly isolated. In Valais, poaching is likely to represent a real threat to lynx as vast zones that had been recolonised in as early as the 1980s are now devoid of this felid (Haller, 1992; Biollaz et al., 2016). Other causes (e.g., diseases or inbreeding depression) can be ruled out in our opinion because other Swiss populations would then have shown signs of such impacts on demography as well. In light of all the evidence presented in this study, long-lasting poaching with the use of neck snares and possibly other methods appears to be the most plausible cause of the extremely low lynx density found in Valais today. In Valais, poaching of legally protected species has also been confirmed for bearded vulture (Gypaetus barbatus) and wolf, with recurrent events for the latter species in the recent times, indicating a certain “frontier mentality” and a disrespect for biodiversity conservation law among some inhabitants, including state agents. In effect, the history of confirmed, but unaddressed, poaching events raises a question of whether some level of proximity between poachers and local law enforcement agents may have allowed poaching to stay unabated. By not taking sufficient measures against neck-snare deployment, even after a person was injured, local law enforcement authorities appear to have sent a signal of tacit approval to lynx poaching. This alone may explain why the lynx trapper even felt emboldened to appear in mass media with a belief of total impunity (Filliez, 2013).
Collusions between game wardens, police forces and poachers are known from other European countries. In Norway in 2016, a police chief inspector conducting investigations on wolf poaching sent text messages to an accused hunter, explaining him how to answer questions from the criminal police so as to have the case dismissed (Supreme Court of Norway, 2018). This case reached the Supreme Court of Norway, which ruled in 2018 that the breach of the inspector's service obligations was serious because his advice to the poacher could reduce the trust among the public regarding the ability and willingness of the police to investigate criminal matters (Supreme Court of Norway, 2018). Also, in Norway in 2015, a retired policeman was found guilty of having helped conceal evidence of illegal wolf hunting (Supreme Court of Norway, 2015). It is worth noting that this occurred in a country that is comparable to Switzerland (World Justice Project, 2019) in being consistently ranked among the nations with high legal compliance and with a quasi-absence of corruption. Informal discussions we have had with other large carnivore researchers in northern Europe indicate that informing local police about large carnivore poaching may indeed merely amount to informing poachers about the visibility of their illegal activity. In effect, in many instances, local police forces seem to treat poaching, especially of large carnivores, as being a socially acceptable crime. It is clear that such a mentality will be a serious impediment to proper judicial investigations.
In the Swiss context, poachers and official cantonal game wardens often belong to the same local social network. In effect, in the Swiss federal political system, police forces are mostly appointed locally at the cantonal level. This is in contrast to France and Italy, for instance, where law enforcement agents in the countryside are usually not appointed in the area they originate from (Houte, 2011), but are re-allocated across the country to prevent corruption, which has led to strong charges against lynx poachers (Ceza et al., 2001). The belief among poachers that they are immune from prosecution likely emboldens them to continue their activity. If a coherent strategy aimed at reducing poaching of large carnivores needs to be integrative in all respects, from prevention to legal prosecution (Schraml, 2019), the risk of a possible tolerance of police forces to poaching must be addressed head-on. In other terms, efforts to control poaching may therefore include policing the police, as the second century Roman poet Juvenal wrote in his Satires “Quis custodiet ipsos custodes.” In this respect, we would recommend that the authorities of Switzerland and other countries where poaching is suspected of threatening top predators to undertake investigations at the highest possible administrative level or establish a centralized police force tasked with investigating major environmental crimes, including illegal hunting of large carnivores.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethical review and approval was not required for the animal study because the study was entirely based on non-invasive sampling (i.e., camera trapping and snow-tracking). Written informed consent was not obtained from the deceased hunter pictured in Supplementary Figure 1 but from the photographer of the scene (©Jean-Claude Tornay).
RA and VB conceived the study. EK, FZ, SM, SR, and SV collected the data. RA, EK, FZ, SR, and VB did the analyses. RA, MK, and VB supervised the analyses. RA, GC, and VB led the writing. All authors commented on the manuscript.
The research programme was funded by the Swiss Federal Office for the Environment (Bundesamt für Umwelt BAFU), Switzerland.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank all the people who made this study possible, in particular Dr. Reini Schnidrig-Petrig, Swiss Federal Office for the Environment, who provided financial support to our research programme. François Biollaz participated in the field surveys while James Hale revised the English. Particular thanks to the attorney general and the criminal police for their inquiry into the perpetator of lynx trap installation and recurrent damages to our trail camera-trap network.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcosc.2021.665000/full#supplementary-material
Akaike, H. (1974). A new look at the statistical model identification. IEEE Trans. Autom. Control 19, 716–723. doi: 10.1109/TAC.1974.1100705
Andrén, H., Linnell, J. D. C., Liberg, O., Andersen, R., Danell, A., Karlsson, J., et al. (2006). Survival rates and causes of mortality in Eurasian lynx (Lynx lynx) in multi-use landscapes. Biol. Conserv. 131, 23–32. doi: 10.1016/j.biocon.2006.01.025
Arlettaz, R., Imstepf, R., Jacot, A., Oggier, P.-A., Posse, B., Pradervand, J.-N., et al. (2019). Oiseaux et biodiversité du Valais: comment les préserver. Sempach: Station ornithologique suisse.
Bartón, K. (2014). “MuMIn: Multi-model interference. Version 1.15.6,” in R package. 1.15.6 ed. Available online at : https://cran.r-project.org/web/packages/MuMIn/index.html (accessed February 28, 2017).
Bates, D., Maechler, M., Bolker, B., Walker, S., Christensen, R. H. B., Singmann, H., et al. (2014). “lme4: linear mixed-effects models using Eigen and S4,” in R package. 1.1-7 ed. doi: 10.18637/jss.v067.i01
Biollaz, F., and Arlettaz, R. (2017, February 23). Le problème du Valais avec les superprédateurs. Le Temps.
Biollaz, F., Mettaz, S., Zimmermann, F., Braunisch, V., and Arlettaz, R. (2016). Statut du Lynx en Valais quatre décennies après son retour: suivi ou moyen de pièges photographiques. Bull. Murithienne 133, 29–44.
Breitenmoser, U. (1998). Large predators in the Alps: the fall and rise of man's competitors. Biol. Conserv. 83, 279–289. doi: 10.1016/S0006-3207(97)00084-0
Breitenmoser, U., and Breitenmoser-Würsten, C. (2008). Der Luchs: ein Grossraubtier in der Kulturlandschaft. Wohlen, Bern: Salm Verlag.
Breitenmoser, U., and Haller, H. (1987). Zur Nahrungsökologie des Luchses (Lynx lynx) in den schweizerischen Nordalpen. Z. Säugetierkunde 52, 168–191.
Cervený, J., Koubek, P., and Bufka, L. (2002). Eurasian Lynx (Lynx Lynx) and its chance for survival in Central Europe: the case of the Czech Republic. Acta Zool. Lituanica 12, 428–432. doi: 10.1080/13921657.2002.10512534
Ceza, B., Kessler, R., Marti, K., Rochat, N., and Tester, U. (2001). Wer tötet den Luchs? Tatsachen, Hintergründe und Indizien zu illegalen Luchstötungen in der Schweiz. Basel: Pro Natura.
Chapron, G., Kaczensky, P., Linnell, J. D. C., von Arx, M., Huber, D., Andrén, H., et al. (2014). Recovery of large carnivores in Europe's modern human-dominated landscapes. Science 346:1517. doi: 10.1126/science.1257553
Chapron, G., and López-Bao, J. V. (2016). Coexistence with large carnivores informed by community ecology. Trends Ecol. Evol. 31, 578–580. doi: 10.1016/j.tree.2016.06.003
Epstein, Y., and Chapron, G. (2018). The hunting of strictly protected species: the tapiola case and the limits of derogation under article 16 of the habitats directive. Eur. Energy Environ. Law Rev. 27, 78–87.
Filliez, X. (2013). Les lynx et les loups, on le bousille, un point c'est tout. L'illustré 13, 20–23.
Fiske, I., Chandler, R., Miller, D., Royle, A., Kery, M., Hostetler, J., et al. (2017). “Unmarked: models for data from unmarked animals. Version 0.11-0,” in R package. 0.12-2 ed. Available online at: https://cran.r-project.org/web/packages/unmarked/index.html (accessed February 28, 2017).
Fournier, J.-R. (2010). Motion Jean-René Fournier dans le cadre de la Révision de l'article 22 de la Convention de Berne. Bulletin officiel du Conseil des Etats, Parlement suisse. Motion 10.3264.
Gallant, J. C., and Wilson, J. P. (2000). “Primary topographic attributes,” in Terrain Analysis: Principles and Applications, eds P. Wilson, and J. C. Gallant (Hoboken: John Wiley and Sons), 51–85.
Gonseth, Y., Wohlgemuth, T., Sansonnens, B., and Buttler, A. (2001). Die biogeographischen Regionen der Schweiz. Erläuterungen und Einteilungsstandard. Bern: Bundesamt für Umwelt (BAFU).
Haller, H. (1992). Zur Ökologie des Luchses Lynx lynx im Verlauf seiner Wiederansiedlung in den Walliser Alpen. Hamburg: Verlag Paul Parey.
Hamr, J. (1985). “Seasonal home range size and utilisation by female chamois (Rupicapra rupicapra) in Northern Tyrol,” in The Biology and Management of Mountain Ungulates, eds S. Lovari (London: Croom Helm), 106–116.
Heurich, M., Schultze-Naumburg, J., Piacenza, N., Magg, N., Cervený, J., Engleder, T., et al. (2018). Illegal hunting as a major driver of the source-sink dynamics of a reintroduced lynx population in Central Europe. Biol. Conserv. 224, 355–365. doi: 10.1016/j.biocon.2018.05.011
Houte, A.-D. (2011). Les mutations de gendarmes depuis le XIXe siècle, entre contrainte institutionnelle et liberté individuelle. Trav. emploi 127, 29–39. doi: 10.4000/travailemploi.5313
Liberg, O., Chapron, G., Wabakken, P., Pedersen, H. C., Thompson Hobbs, N., and Sand, H. (2012). Shoot, shovel and shut up: cryptic poaching slows restoration of a large carnivore in Europe. Proc. R. Soc. B 279, 910–915. doi: 10.1098/rspb.2011.1275
López-Bao, J. V., Bruskotter, J., and Chapron, G. (2017). Finding space for large carnivores. Nat. Ecol. Evol. 1:140. doi: 10.1038/s41559-017-0140
Lüchtrath, A., and Schraml, U. (2015). The missing lynx – understanding hunters' opposition to large carnivores. Wildlife Biol. 21, 110–119. doi: 10.2981/wlb.00068
Ministère public du Canton du Valais (2015). Ordonnance pénale du 12 novembre 2015 contre M. Lini Paccolat. MPB 15:1387.
Molinari, P., Rotelli, L., Catello, M., and Bassano, B. (2001). Present status and distribution of the Eurasian lynx (Lynx lynx) in the Italian Alps. Hystrix 12, 3–9. doi: 10.4404/hystrix-12.2-4172
Molinari-Jobin, A., Wölfl, S., Marboutin, E., Molinari, P., Wölfl, M., Kos, I., et al. (2012). Monitoring the Lynx in the Alps. Hystrix 23, 49–53. doi: 10.4404/hystrix-23.1-4553
Molinari-Jobin, A., Zimmermann, F., Angst, C., Breitenmoser-Würsten, C., Capt, S., and Breitenmoser, U. (2006). Status and distribution of the lynx in the Swiss Alps 2000–2004. Acta Biol. Slov. 49, 3–11.
Molinari-Jobin, A., Zimmermann, F., Breitenmoser-Würsten, C., Capt, S., and Breitenmoser, U. (2001). Present status and distribution of the lynx in the Swiss Alps. Hystrix 12, 17–27. doi: 10.4404/hystrix-12.2-4174
Moser, A. (2016). Rehbock, ledig, sucht … Program Netznatur. Zürich: Swiss German Television (SRG1).
Mysterud, A. (1999). Seasonal migration pattern and home range of roe deer (Capreolus capreolus) in an altitudinal gradient in southern Norway. J. Zool. 247, 479–486. doi: 10.1111/j.1469-7998.1999.tb01011.x
QGIS Development Team (2017). Geographic Information System. Version 2.18.9. Las Palmas. Open Source Geospatial Foundation Project. Available online at: http://www.qgis.org (accessed February 4, 2021).
R-Core-Team (2014). R: A Language and Environment for Statistical Computing. Version 3.3.4. Available online at: https://www.r-project.org/ (accessed February 4, 2021).
Ripple, W. J., Estes, J. A., Beschta, R. L., Wilmers, C. C., Ritchie, E. G., Hebblewhite, M., et al. (2014). Status and ecological effects of the world's largest carnivores. Science 343:1241484. doi: 10.1126/science.1241484
Roder, S., Biollaz, F., Mettaz, S., Zimmermann, F., Manz, R., Kéry, M., et al. (2020). Deer density drives habitat use of establishing wolves in the Western European Alps. J. Appl. Ecol. 57, 995–1008. doi: 10.1111/1365-2664.13609
Sakamoto, Y., Ishiguro, M., and Kitagawa, G. (1986). Akaike Information Criterion Statistics. Dordrecht: Springer.
Schraml, U. (2019). “Wildtiermanagement für Menschen,” in Wolf, Luchs und Bär in der Kulturlandschaft. Konflikte, Chancen, Lösungen im Umgang mit grossen Beutegreifern, ed M. Heurich (Stuttgart: Eugen Ulmer KG), 113–148.
Sindičić, M., Gomerčić, T., Kusak, J., Slijepčević, V., Huber, Ð., and Frković, A. (2016). Mortality in the Eurasian lynx population in Croatia over the course of 40 years. Mamm. Biol. 81, 290–294. doi: 10.1016/j.mambio.2016.02.002
Supreme Court of Norway (2015). Straffutmåling for å ha hjulpet til med å skjule en ulovlig felt ulv. HR-2015-01134-A.
Treves, A., and Karanth, K. U. (2003). Human-carnivore conflict and perspectives on carnivore management worldwide. Conserv. Biol. 17, 1491–1499. doi: 10.1111/j.1523-1739.2003.00059.x
Wilcoxon, F. (1945). Individual comparisons by ranking methods. Biom. Bull. 1, 80–83. doi: 10.2307/3001968
Zimmermann, F. (2004). Conservation of the Eurasian Lynx (Lynx lynx) in a fragmented landscape - habitat models, dispersal and potential distribution (Ph.D. thesis). University of Lausanne, Lausanne, Switzerland.
Zimmermann, F., Dulex, N., Foresti, D., Breitenmoser-Würsten, C., and Breitenmoser-Würsten, U. (2014). Abundanz und Dichte des Luchses im Wallis Nord: Fang-Wiederfang-Schätzung mittels Fotofallen im K-VII im Winter 2013/14. Muri: KORA.
Zimmermann, F., and Foresti, D. (2016). “Capture-recapture methods for density estimation,” in Camera Trap-Ping for Wildlife Research, eds F. Rovero and F. Zimmermann (Exeter: Pelagic Publishing), 95–141.
Zimmermann, F., Kunz, F., Rhein, K., Shepherd, M., Tschanz, P., Breitenmoser-Würsten, C., et al. (2016). Abondance et densité du lynx dans le Nord-Ouest des Alpes: estimation par capture-recapture photographique dans l'aire de référence étendue au canton de Vaud dans le C-VI durant l'hiver 2015/16. Muri: KORA.
Zimmermann, F., Küttel, J., Breitenmoser-Würsten, C., Breitenmoser, U., and Kunz, F. (2019). Monitoring déterministe du lynx avec les pièges-photos dans le Sud du Bas-Valais IVd durant l'hiver 2018/19. Muri: KORA.
Zimmermann, F., Molinari-Jobin, A., Ryser, A., Breitenmoser-Würsten, C., Pesenti, E., and Breitenmoser, U. (2011). Status and distribution of the lynx (Lynx lynx) in the Swiss Alps 2005–2009. Acta Biol. Slov. 54, 74–84.
Keywords: camera trap survey, large carnivores, Lynx lynx, N-mixture models, poaching, ungulate densities, prey availability, illegal hunting
Citation: Arlettaz R, Chapron G, Kéry M, Klaus E, Mettaz S, Roder S, Vignali S, Zimmermann F and Braunisch V (2021) Poaching Threatens the Establishment of a Lynx Population, Highlighting the Need for a Centralized Judiciary Approach. Front. Conserv. Sci. 2:665000. doi: 10.3389/fcosc.2021.665000
Received: 06 February 2021; Accepted: 30 April 2021;
Published: 07 June 2021.
Edited by:
Yonggang Nie, Institute of Zoology, Chinese Academy of Sciences (CAS), ChinaCopyright © 2021 Arlettaz, Chapron, Kéry, Klaus, Mettaz, Roder, Vignali, Zimmermann and Braunisch. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raphaël Arlettaz, cmFwaGFlbC5hcmxldHRhekBpZWUudW5pYmUuY2g=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.