
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Complex Syst., 18 March 2025
Sec. Multi- and Cross-Disciplinary Complexity
Volume 3 - 2025 | https://doi.org/10.3389/fcpxs.2025.1534330
This article is part of the Research TopicComplex Systems Approaches in EcologyView all articles
 Vasilis Dakos1*
Vasilis Dakos1* Antoine Le Vilain1,2
Antoine Le Vilain1,2 Elisa Thebault2
Elisa Thebault2 Teresa Alcoverro3
Teresa Alcoverro3 Jordi Boada3,4
Jordi Boada3,4 Eduardo Infantes5
Eduardo Infantes5 Dorte Krause-Jensen6
Dorte Krause-Jensen6 Núria Marbà7Oscar Serrano3,8
Núria Marbà7Oscar Serrano3,8 Salvatrice Vizzini9,10Eugenia T. Apostolaki11*
Salvatrice Vizzini9,10Eugenia T. Apostolaki11*Coastal Blue Carbon ecosystems like seagrass meadows are foundation habitats with a capacity to sequester and store organic carbon in their sediments, and their protection and restoration may thereby support climate change mitigation while also supporting biodiversity and many other ecosystem functions. However, seagrass ecosystems are being lost due to human activities, disease and, in some regions, climate change, which may trigger the release of stored carbon into the atmosphere. Yet, we do not fully understand how global change-induced seagrass loss influences sedimentary carbon dynamics. What is even less clear is whether seagrass loss may also result in tipping points, i.e., abrupt and difficult-to-reverse shifts, in carbon flux dynamics turning seagrass ecosystems from net carbon sinks to net carbon sources. Here, we propose that conceptual mechanistic models of coupled ecological and biogeochemical dynamics can help to study the effects of major stressors on seagrass meadows and associated carbon fluxes. We then illustrate one case of such a conceptual model that focuses on anthropogenic induced mortality by physical stress as an example. Our perspective highlights how a modeling approach for understanding the response of carbon fluxes in seagrass ecosystems to global change stressors may be useful in informing coastal seagrass management towards climate change mitigation actions.
Coastal Blue Carbon (BC) ecosystems, which include seagrass meadows, mangrove forests, and tidal marshes, can store carbon in their underlying sediments over centennial to millennial scales, and their protection and restoration could contribute to offsetting greenhouse gas emissions (Nellemann et al., 2009; Duarte et al., 2013; Macreadie et al., 2021). Concurrently, seagrass meadows are biodiversity hotspots, support fisheries, provide natural protection by attenuating wave energy and accreting sediments, so they offer several ecosystem services beyond carbon sequestration,. Therefore, the sustainable management of seagrass meadows and the assurance of their integrity has been recognized as a low-regret Nature-based Solution (NbS) towards mitigation and adaptation to climate change safeguarding coastal resilience (Hoegh-Guldberg et al., 2019; Hoegh-Guldberg et al., 2020; Gattuso et al., 2021).
However, seagrass sensitivity to stressors is acute, and global change impacts have resulted in extensive loss of meadows worldwide (Orth et al., 2006; Unsworth et al., 2022). The loss of at least 20% of the global seagrass area since 1880 (Dunic et al., 2021), with accelerating loss rates from 0.9% yr-1 in the 1940s to 7% y-1 in the 1990s and 2000s (Waycott et al., 2009), highlights a need for further conservation (Unsworth et al., 2022). The loss is attributed to multiple direct pressures and climate change acting synergistically, including coastal eutrophication, mechanical damage, warming and extreme events (de los Santos et al., 2019). Seagrass loss could compromise the climate change mitigation and adaptation potential of seagrass meadows. Yet, there is a significant lack of understanding of the extent to which global change stressors could affect carbon storage in these critical ecosystems. Seagrass loss usually decreases their carbon sequestration capacity (Marbà et al., 2015; Dahl et al., 2016; Thorhaug et al., 2017; Trevathan-Tackett et al., 2017; Githaiga et al., 2019; Salinas et al., 2020; Moksnes et al., 2021), and increases the risk of erosion and subsequent mineralization of historical carbon accumulated in sediments (Marbà et al., 2015; Arias-Ortiz et al., 2018; Roca et al., 2022; Egea et al., 2023), potentially transforming the meadows from sinks to sources of greenhouse gasses (Lovelock et al., 2017). However, the relationship between seagrass ecosystem integrity and carbon storage is not straightforward, and ecological transformations (even dramatic ones such as complete cover loss) may not necessarily threaten the carbon that is already stored in the sediments (Piñeiro-Juncal et al., 2021; Apostolaki et al., 2022). Despite the progress in blue carbon science over the past decade, we still lack an in-depth understanding of how seagrass deterioration and loss affect the functioning of seagrass ecosystems as carbon sinks, and it is not well-understood if there are thresholds to changes in carbon fluxes linked to ecological and/or biogeochemical processes.
Recent studies in seagrass ecosystems have suggested that biogeochemical and biophysical feedbacks (i.e., direct or indirect reciprocal interactions between system parts) can cause seagrass meadows to abruptly and irreversibly shift from a healthy growing state to a deteriorated barren state (Maxwell et al., 2017). Similar shifts due to competition by invasive species have also been modeled (Llabrés et al., 2023; Beca-Carretero et al., 2024). Abrupt shifts can occur at the crossing of thresholds with irreversible changes in ecological functions and services. Such regime shifts might be associated with the presence of tipping points that cause a radical shift to a usually contrasting state propelled by a strong positive feedback (van Nes et al., 2016). Regime shifts have been described for a range of ecosystems like clear lakes turning turbid (Scheffer et al., 1993), coral reefs getting overgrown by macroalgae (Knowlton, 1992), or drylands shifting to desertification (Kéfi et al., 2007), seaweed forests collapsing into urchin barrens (Ling et al., 2015) and seagrasses turning to bare or algae dominated meadows (Unsworth et al., 2015), or degraded dead matte (Rindi et al., 2024). But how likely it is that such nonlinear seagrass responses can also induce tipping responses in the dynamics of carbon storage in seagrass meadows remains unknown.
Although mechanisms and models for describing tipping responses in seagrass ecosystems have been proposed (van der Heide et al., 2007a; Carr et al., 2010; Christianen et al., 2014; Maxwell et al., 2017; Ruiz-Reynés et al., 2017; Adams et al., 2018; Mayol et al., 2022; Minguito-Frutos et al., 2023; Ruiz-Reynés et al., 2023), those have not been directly linked to the loss of carbon storage. This aspect is critical because these abrupt changes can directly influence carbon dynamics by modifying the biogeochemical processes and their potential feedbacks that ultimately will determine the fate of stored carbon. Perhaps the closest modeling approach is based on patch growth models simulating growth and colonization processes under different restoration scenarios and timeframes, by relating cover maps with carbon sequestration rates at local scales (Duarte et al., 2013). Another study modeled CO2 emissions following the degradation of seagrass using decay rates of Corg in above- and below-ground tissues and underlying sediments (Lovelock et al., 2017). For both restoration projects and seagrass degradation scenarios, the most common model used is the Integrated Valuation of Ecosystem Services and Tradeoffs (InVEST) model (Moritsch et al., 2021; González-García et al., 2022). The InVEST model estimates the carbon sequestration potential of seagrass ecosystems by calculating the amount of carbon that is stored in seagrass biomass, as well as the carbon that is stored in the underlying sediment. Yet, to the best of our knowledge, no mechanistic model has so far been developed to evaluate the potential cascading effects of tipping points in seagrass and associated carbon dynamics (see Supplementary Material S1). If abrupt seagrass shifts could also lead to cascading responses in carbon source-sink dynamics, it is important to understand under which conditions and at what scale such an effect takes place.
Here, we propose a minimal modelling perspective for studying the likelihood of tipping point responses in the carbon storage (i.e., from carbon sink to source of CO2) of seagrass ecosystems under global change stressors. We show how conceptual mechanistic models could be built based on expert knowledge by coupling a minimal set of biogeochemical interactions and ecological feedbacks. We then develop one such model to showcase that seagrass meadows can abruptly shift from sink to carbon source in response to anthropogenic induced physical damage. We use this type of anthropogenic induced mortality only as an example, as the mechanistic modelling framework we present could be used to study any other pressure by itself or in combination. We outline limitations and opportunities presented by such minimal modeling approaches towards improving the management of seagrasses to enhance their functioning as carbon storage systems.
We drew a simplified overview of the main processes that govern interactions in the carbon storage of seagrass ecosystems (Figure 1). We distinguished the three pools that make up the carbon stores in seagrass ecosystem: a) the seagrass, b) the upper sediment layer (i.e., the newly deposited organic matter (OM) within the rhizosphere that undergoes faster mineralization), and c) the lower sediment layer (i.e., the more stable OM pool). The origin of organic carbon in the sediment can be autochthonous (i.e., deriving from above- and below-ground tissues) or allochthonous (i.e., deriving from the suspended material trapped by seagrasses), often with similar contributions by the two sources (Kennedy et al., 2022). During the mineralization of OM in the sediment, nutrients are released to pore water and the water column (Figure 1). The factors affecting the different compartments were grouped according to whether they represent environmental settings (i.e., hydrodynamics, nutrient and light availability, and sediment origin, texture and composition), long-term stressors (e.g., warming, eutrophication, siltation, invasive species, herbivory), or acute short-term disturbances (e.g., mechanical damage, storms, heatwaves). Figure 1 summarizes how each of these factors potentially affects the processes that govern carbon dynamics across the three carbon pools in seagrass ecosystems.

Figure 1. Interactions and feedbacks between living seagrass and sedimentary carbon as well as the major factors and stressors affecting their dynamics. Factors affecting seagrass and carbon compartments, numbered and colored according to whether they represent environmental settings, long-term stressors, or acute short-term disturbances. Geophysical factors (green): depth (light availability) (1), hydrodynamics (2), and sediment type (origin, texture and composition) (3). Long-term stressors (red): warming (1), eutrophication (2), herbivory (3), invasive species (4) and siltation (turbidity) (5). Acute short-term disturbances (blue): mechanical damage (1), storms (2), and heatwaves (3).
To build our mechanistic model that couples ecological and biogeochemical dynamics in seagrass meadows we used the processes described above (Figure 1) that were derived through expert knowledge and previous theoretical work. Thus, we did not build nor parameterize a model based on collected ecological data but only developed a conceptual model based on processes. Furthermore, we did not include all described processes (Figure 1), but we focused on the impact of anthropogenic stress – related to physical damage like dredging and anchoring – that could lead to seagrass meadow degradation. There are two dynamical parts in the model: the ecological dynamics that describe seagrass growth (i.e., metabolic capture of carbon) and the biogeochemical dynamics that describe carbon storage in the sediment. Our aim is to show how such a mechanistic model can help understand the qualitative responses of seagrass carbon storage to stressors, even if we cannot fully describe and parameterize the model (Scheffer and Beets, 1994).
We followed the model structure proposed by Maxwell et al. (2017) to describe changes in seagrass biomass (Equation 1). There are three main terms in the model: a logistic growth term of seagrass biomass, and two loss terms which result from the physical (hydrodynamics) and natural mortality. The model includes a positive hydrodynamic feedback where the seagrass canopy can decrease the negative physical stress of water currents on the meadow:
where
We consider two different origins of carbon stored in seagrass sediments: autochthonous carbon,
Details on model analysis and parameter values can be found in the Supplementary Material.
The bifurcation diagrams of Figure 2 show the asymptotic equilibria of seagrass and the three carbon pools within the sediment for a given level of anthropogenic induced mortality mA. The greater the anthropogenic mortality (Figure 2A), the more likely it is that the meadow shifts from a seagrass-vegetated state to a bare state. At a certain threshold of anthropogenic induced mortality mA, the seagrass meadow crosses a tipping point and shifts to a bare state with no standing seagrass biomass, thereby leaving the sediment exposed to faster mineralization and erosion. Looking at the carbon stored in each compartment in relation to the same anthropogenic mortality gradient (Figure 2B), we find that the carbon mass in each compartment responds in a similar way to the seagrass dynamics under different hydrodynamic regimes. Both allochthonous and autochthonous carbon mass decreases with increasing anthropogenic stress. In fact, the absence of standing seagrass biomass results in the loss of
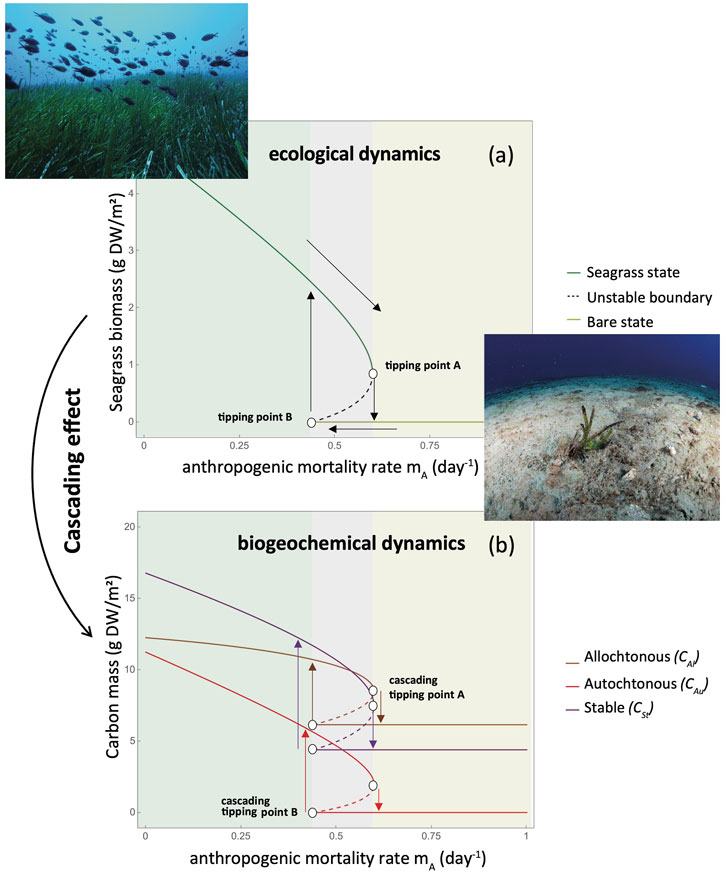
Figure 2. Asymptotic equilibria and tipping points in a coupled seagrass and sedimentary carbon conceptual mechanistic model. Bifurcation diagram of seagrass and sedimentary carbon dynamics along a hypothetical gradient of anthropogenic induced mortality (mA). A seagrass state means that there is one stable equilibrium with the presence of seagrasses. A bare state means that there is only one stable equilibrium with no seagrass. A bistable state means that there are two alternative equilibria, the seagrass and bare state equilibria. (a) Equilibrium states of seagrass biomass (S). Once anthropogenic stress increases, seagrass biomass decreases, until a tipping point A where the seagrass shifts abruptly to a bare state. Seagrass biomass recovers at tipping point B at a lower level of anthropogenic induced mortality compared to tipping point A due to the hysteresis effect. (b) Allochthonous (CAl), autochthonous (CAu) and stable (CS) carbon mass equilibria states along the same anthropogenic stress gradient. The carbon equilibria reflect the equilibrium state changes of living seagrass biomass. Once seagrass crosses the tipping points A or B, it collapses and effect cascades to the carbon compartment, leading to sudden loss of carbon and a low carbon equilibrium state as the sole carbon contribution originates from the allochthonous sources in the water column. [Model parameters used are not specific to a given seagrass species or meadow but constrained by realistic assumptions to showcase the qualitative behavior of the system; see detailed model and parameter description in the Supplementary Material. Photo insets depict a healthy (upper left) and a degraded (lower right) Posidonia oceanica meadow (Photo credits: Julius Glambedakis and Thanos Dailianis).
In this perspective paper, we suggest that there are thresholds beyond which seagrass meadows turn from net carbon sinks to net sources of CO2 due to the crossing of tipping points in seagrass meadows. We described a set of positive and negative interactions at the level of seagrass meadows, as well as between the sediment and the seagrass that altogether affect carbon dynamics in seagrass meadows (Figure 1). We proposed that incorporating this knowledge into a coupled ecological-biogeochemical conceptual model can improve our understanding of the potential response of carbon storage to seagrass meadow degradation. We then built a simplified version of such model to showcase how tipping events in seagrass meadows driven by anthropogenic stress could cascade into the sedimentary carbon dynamics, leading to the eventual loss of stable carbon that implies a transition of the meadow from a net sink to a source of CO2 (Figure 2).
Clearly, our model focusing on a single positive feedback is only a starting point. Seagrass clonal integration, for instance, can be considered among the main reinforcing feedbacks (Nielsen and Pedersen, 2000), as well as increasing light quality through sediment trapping by the canopy, or belowground biomass allocation in response to hydrodynamic and nutrient conditions (Burkholder et al., 2007; Maxwell et al., 2017). Although the choice of the modelled feedback may affect features of the tipping behavior of seagrasses, we do not expect that it will change the overall cascading effect on sedimentary carbon dynamics qualitatively. Instead, it is the approximations and parameters of the feedback functions that are key in predicting carbon storage responses. For instance, in the presented model (Figure 2), the most important parameter that determines the occurrence of tipping points in the system is the half-saturation constant Shh that modulates the feedback effect of seagrass on reducing hydrodynamic intensity. The question is to identify how other feedback functions would be as crucial in modulating tipping responses.
We have assumed the most parsimonious relationships (i.e., linear) in the fluxes between the different compartments, neglecting, for example, priming effects that have been shown to enhance carbon decomposition in seagrass meadows (Trevathan-Tackett et al., 2018). At the same time, we have not considered that decomposition contributes to the pore water nutrient pools (Figure 1) that could create either positive or negative feedbacks in nutrient fluxes from the sediment to the seagrass or other primary producers (de Boer, 2007). A more complete model could integrate light and nutrient limitation as well as temperature dependence of growth and decomposition in order to study the effects of multiple stressors both in isolation and in combination. Future work should also investigate the effect of different time scales, ranging from hours to centuries, involved in the metabolic capture of carbon, and carbon decomposition and preservation dynamics. Processes occurring at different time scales could lead to significant time lags and long transients which may be more relevant than the asymptotic dynamics we focused on here (Hastings et al., 2018). Yet, to achieve all these, robust data on seagrass ecology and carbon biogeochemistry are needed.
Current knowledge gaps in BC science limit our ability to develop and parameterize accurate mechanistic models of carbon fluxes following seagrass deterioration. In our model, we used arbitrary parameter values to explore in general the possibility of tipping responses. The next step is to have species-specific parameter values to estimate the likelihood of tipping responses to occur given the model we use. For example, field experiments could help us to obtain estimates of hydrodynamic shear stress under different depths and canopy densities (e.g., (van der Heide et al., 2007b; Infantes et al., 2009), whereas mortality rates due to anthropogenic disturbances (like anchoring) could be derived from existing surveys (Pergent-Martini et al., 2022).
We also need further empirical evidence of the dynamics between the capture of carbon, its burial and erosion, and its post-depositional transformation to be able to predict the possible change in the carbon sequestration capacity of seagrass ecosystems following disturbance. Long-term laboratory experiments are necessary to quantify the decomposition rates and diagenesis of organic carbon and how these change with the type, intensity and frequency of stressors (Spivak et al., 2019). Spatially larger assessments will allow accounting for the variability in carbon storage caused by large-scale factors (e.g., geomorphic setting, temperature), whereas localized assessments will allow accounting for local variability in carbon budgets, considering the effect of small-scale factors (e.g., sediment typology, hydrodynamics) on carbon fluxes (Mazarrasa et al., 2021; Kennedy et al., 2022).
Conserving the BC capacity of seagrass meadows requires a mechanistic understanding of the most important processes affecting the response of seagrass and associated carbon pools to different environmental, ecological and climate change conditions. Our proposed modeling framework can contribute to such understanding and inspire further model development. For example, by accounting for a combination of stressors, like warming, turbidity or eutrophication, it will be possible to outline potential projections of BC storage while embracing the uncertainty in the model structure and parameter estimates. Or, we can explore possible responses of seagrass meadows and their C-pools under high and low hydrodynamic regimes along a gradient of temperature in the presence of stochastic mechanical damage in hypothetical meadows. Such numerical experiments though require gathering robust ecological data that will eventually allow us to calibrate and validate mechanistic models like the conceptual one we developed here that could be implemented in BC accounting frameworks.
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding authors.
VD: Conceptualization, Funding acquisition, Methodology, Software, Writing–original draft, Writing–review and editing. AL: Conceptualization, Formal Analysis, Writing–review and editing. ET: Conceptualization, Methodology, Writing–review and editing. TA: Conceptualization, Writing–review and editing. JB: Conceptualization, Writing–review and editing. EI: Conceptualization, Writing–review and editing. DK-J: Conceptualization, Writing–review and editing. NM: Conceptualization, Writing–review and editing. OS: Conceptualization, Writing–review and editing. SV: Conceptualization, Writing–review and editing. EA: Conceptualization, Funding acquisition, Writing–original draft, Writing–review and editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the European Marine Research Network through a EuroMarine Foresight workshop “From sink to source: Tipping points of blue carbon storage in Mediterranean seagrass ecosystems” organized in the garrigue (phrygana) outside Montpellier in 2019 and DRESSAGE project (MIS5045792) through the Operational Program ‘Competitiveness, Entrepreneurship and Innovation’ (EPAnEK 2014-2020, NSRF). OS was supported by I + D + i projects RYC 2019-027073-I and PIE HOLOCENO 20213AT014 funded by MCIN/AEI/10.13039/501100011033 and FEDER. DK-J. and NM were supported by OBAMA-NEXT (grant agreement no. 101081642), funded by the European Union under the Horizon Europe program. SV was supported by the project PON03PE_00203_1 Marine Hazard, PON Ricerca e Competitività 2007-2013 MUR. TA was supported by STORM (PID 2020-113745RB-100) of the Spanish Ministry of Science and Innovation (MCIN/AEI/10.13039/501 10000 1 1033). JB acknowledges the support received by the I + D + i project RYC 2021-033650-I and PID 2022-140652NA-I00 MITnPOINT funded by the MCIN/AIEI/10.13039/501100011033 and FEDER and the European Commission–European Union’s Horizon 2020 MSCA – SHIFT2SOLVE-1030591.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcpxs.2025.1534330/full#supplementary-material
Adams, M., Ghisalberti, M., Lowe, R., Callaghan, D., Baird, M., Infantes, E., et al. (2018). Water residence time controls the feedback between seagrass, sediment and light: implications for restoration. Adv. Water Resour. 117, 14–26. doi:10.1016/j.advwatres.2018.04.004
Apostolaki, E. T., Caviglia, L., Santinelli, V., Cundy, A. B., Tramati, C. D., Mazzola, A., et al. (2022). The importance of dead seagrass (Posidonia oceanica) matte as a biogeochemical sink. Front. Mar. Sci. 9. doi:10.3389/fmars.2022.861998
Arias-Ortiz, A., Serrano, O., Masqué, P., Lavery, P. S., Mueller, U., Kendrick, G. A., et al. (2018). A marine heat wave drives massive losses from the world’s largest seagrass carbon stocks. Nat. Clim. Change 8, 338–344. doi:10.1038/s41558-018-0096-y
Beca-Carretero, P., Winters, G., Teichberg, M., Procaccini, G., Schneekloth, F., Zambrano, R. H., et al. (2024). Climate change and the presence of invasive species will threaten the persistence of the Mediterranean seagrass community. Sci. Total Environ. 910, 168675. doi:10.1016/j.scitotenv.2023.168675
Boudsocq, S., Lata, J. C., Mathieu, J., Abbadie, L., and Barot, S. (2009). Modelling approach to analyse the effects of nitrification inhibition on primary production. Funct. Ecol. 23, 220–230. doi:10.1111/j.1365-2435.2008.01476.x
Carr, J., D’Odorico, P., McGlathery, K., and Wiberg, P. (2010). Stability and bistability of seagrass ecosystems in shallow coastal lagoons: role of feedbacks with sediment resuspension and light attenuation. J. Geophys. Res. 115. doi:10.1029/2009JG001103
Christianen, M. J. A., Herman, P. M. J., Bouma, T. J., Lamers, L. P. M., van Katwijk, M. M., van der Heide, T., et al. (2014). Habitat collapse due to overgrazing threatens turtle conservation in marine protected areas. Proc. R. Soc. B Biol. Sci. 281, 20132890. doi:10.1098/rspb.2013.2890
Dahl, M., Deyanova, D., Gütschow, S., Asplund, M. E., Lyimo, L. D., Karamfilov, V., et al. (2016). Sediment properties as important predictors of carbon storage in zostera marina meadows: a comparison of four European areas. PLOS ONE 11, e0167493. doi:10.1371/journal.pone.0167493
de los Santos, C. B., Krause-Jensen, D., Alcoverro, T., Marbà, N., Duarte, C. M., van Katwijk, M. M., et al. (2019). Recent trend reversal for declining European seagrass meadows. Nat. Commun. 10, 3356. doi:10.1038/s41467-019-11340-4
Duarte, C. M., Sintes, T., and Marbà, N. (2013). Assessing the CO2 capture potential of seagrass restoration projects. J. Appl. Ecol. 50, 1341–1349. doi:10.1111/1365-2664.12155
Dunic, J. C., Brown, C. J., Connolly, R. M., Turschwell, M. P., and Côté, I. M. (2021). Long-term declines and recovery of meadow area across the world’s seagrass bioregions. Glob. Change Biol. 27, 4096–4109. doi:10.1111/gcb.15684
Egea, L., Infantes, E., and Jiménez-Ramos, R. (2023). Loss of POC and DOC on seagrass sediments by hydrodynamics. Sci. Total Environ. 901, 165976. doi:10.1016/j.scitotenv.2023.165976
Gattuso, J.-P., Williamson, P., Duarte, C. M., and Magnan, A. K. (2021). The potential for ocean-based climate action: negative emissions technologies and beyond. Front. Clim. 2. doi:10.3389/fclim.2020.575716
Githaiga, M. N., Frouws, A. M., Kairo, J. G., and Huxham, M. (2019). Seagrass removal leads to rapid changes in fauna and loss of carbon. Front. Ecol. Evol. 7. doi:10.3389/fevo.2019.00062
González-García,, A., Arias, M., García-Tiscar, S., Alcorlo, P., and Santos-Martin, F. (2022). National blue carbon assessment in Spain using InVEST: Current state and future perspectives. Ecosyst. Serv. 53, 101397. doi:10.1016/j.ecoser.2021.101397
Hoegh-Guldberg, O., Northrop, E., Ashford, O. S., Chopin, T., Cross, J., Duarte, C., et al. (2020). Updated opportunities for action.
Hoegh-Guldberg, O., Northrop, E., and Lubchenco, J. (2019). The ocean is key to achieving climate and societal goals. Science 365, 1372–1374. doi:10.1126/science.aaz4390
Infantes, E., Terrados, J., Orfila, A., Cañellas, B., and Álvarez-Ellacuria, A. (2009). Wave energy and the upper depth limit distribution of Posidonia oceanica. botm 52, 419–427. doi:10.1515/BOT.2009.050
Kaal, J., Serrano, O., del Río, J. C., and Rencoret, J. (2018). Radically different lignin composition in Posidonia species may link to differences in organic carbon sequestration capacity. Org. Geochem. 124, 247–256. doi:10.1016/j.orggeochem.2018.07.017
Kéfi, S., Rietkerk, M., Alados, C. L., Pueyo, Y., Papanastasis, V. P., ElAich, A., et al. (2007). Spatial vegetation patterns and imminent desertification in Mediterranean arid ecosystems. Nature 449, 213–217. doi:10.1038/nature06111
Kennedy, H., Pagès, J. F., Lagomasino, D., Arias-Ortiz, A., Colarusso, P., Fourqurean, J. W., et al. (2022). Species traits and geomorphic setting as drivers of global soil carbon stocks in seagrass meadows. Glob. Biogeochem. Cycles 36, e2022GB007481. doi:10.1029/2022GB007481
Knowlton, N. (1992). Thresholds and multiple stable states in coral reef community dynamics. Am. Zool. 32, 674–682. doi:10.1093/icb/32.6.674
Ling, S. D., Scheibling, R. E., Rassweiler, A., Johnson, C. R., Shears, N., Connell, S. D., et al. (2015). Global regime shift dynamics of catastrophic sea urchin overgrazing. Philosophical Trans. R. Soc. B Biol. Sci. 370, 20130269. doi:10.1098/rstb.2013.0269
Llabrés, E., Blanco-Magadán, A., Sales, M., and Sintes, T. (2023). Effect of global warming on Western Mediterranean seagrasses: a preliminary agent-based modelling approach. Mar. Ecol. Prog. Ser. 710, 43–56. doi:10.3354/meps14298
Lovelock, C. E., Atwood, T., Baldock, J., Duarte, C. M., Hickey, S., Lavery, P. S., et al. (2017). Assessing the risk of carbon dioxide emissions from blue carbon ecosystems. Front. Ecol. Environ. 15, 257–265. doi:10.1002/fee.1491
Macreadie, P. I., Costa, M. D. P., Atwood, T. B., Friess, D. A., Kelleway, J. J., Kennedy, H., et al. (2021). Blue carbon as a natural climate solution. Nat. Rev. Earth Environ. 2, 826–839. doi:10.1038/s43017-021-00224-1
Marbà, N., Arias-Ortiz, A., Masqué, P., Kendrick, G. A., Mazarrasa, I., Bastyan, G. R., et al. (2015). Impact of seagrass loss and subsequent revegetation on carbon sequestration and stocks. J. Ecol. 103, 296–302. doi:10.1111/1365-2745.12370
Maxwell, P. S., Eklöf, J. S., van Katwijk, M. M., O’Brien, K. R., de la Torre-Castro, M., Boström, C., et al. (2017). The fundamental role of ecological feedback mechanisms for the adaptive management of seagrass ecosystems - a review. Biol. Rev. 92, 1521–1538. doi:10.1111/brv.12294
Mayol, E., Boada, J., Pérez, M., Sanmartí, N., Minguito-Frutos, M., Arthur, R., et al. (2022). Understanding the depth limit of the seagrass Cymodocea nodosa as a critical transition: field and modeling evidence. Mar. Environ. Res. 182, 105765. doi:10.1016/j.marenvres.2022.105765
Mazarrasa, I., Lavery, P., Duarte, C. M., Lafratta, A., Lovelock, C. E., Macreadie, P. I., et al. (2021). Factors determining seagrass blue carbon across bioregions and geomorphologies. Glob. Biogeochem. Cycles 35, e2021GB006935. doi:10.1029/2021GB006935
Minguito-Frutos, M., Adams, M. P., Alcoverro, T., Vilas, M. P., Alonso, D., Mayol, E., et al. (2023). Quantifying the role of photoacclimation and self-facilitation for seagrass resilience to light deprivation. Front. Plant Sci. 14, 1186538. doi:10.3389/fpls.2023.1186538
Moksnes, P.-O., Röhr, E., Holmer, M., Eklöf, J., Eriander, L., Infantes, E., et al. (2021). Major impacts and societal costs of seagrass loss on sediment carbon and nitrogen stocks. Ecosphere 12. doi:10.1002/ecs2.3658
Moritsch, M. M., Young, M., Carnell, P., Macreadie, P. I., Lovelock, C., Nicholson, E., et al. (2021). Estimating blue carbon sequestration under coastal management scenarios. Sci. Total Environ. 777, 145962. doi:10.1016/j.scitotenv.2021.145962
Nellemann, C., Corcoran, E., Duarte, C. M., Valdés, L., De Young, C., Fonseca, L., et al. (2009). “Blue carbon,” in A rapid response assessment. Available at: https://www.grida.
Orth, R. J., Carruthers, T. J. B., Dennison, W. C., Duarte, C. M., Fourqurean, J. W., Heck, K. L., et al. (2006). A global crisis for seagrass ecosystems. BioScience 56, 987–996. doi:10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2
Pergent-Martini, C., Monnier, B., Lehmann, L., Barralon, E., and Pergent, G. (2022). Major regression of Posidonia oceanica meadows in relation with recreational boat anchoring: a case study from Sant’Amanza bay. J. Sea Res. 188, 102258. doi:10.1016/j.seares.2022.102258
Piñeiro-Juncal, N., Kaal, J., Moreira, J. C. F., Martínez Cortizas, A., Lambais, M. R., Otero, X. L., et al. (2021). Cover loss in a seagrass Posidonia oceanica meadow accelerates soil organic matter turnover and alters soil prokaryotic communities. Org. Geochem. 151, 104140. doi:10.1016/j.orggeochem.2020.104140
Rindi, L., Mintrone, C., Ravaglioli, C., and Benedetti-Cecchi, L. (2024). Spatial signatures of an approaching regime shift in Posidonia oceanica meadows. Mar. Environ. Res. 198, 106499. doi:10.1016/j.marenvres.2024.106499
Roca, G., Palacios, J., Ruíz-Halpern, S., and Marbà, N. (2022). Experimental carbon emissions from degraded mediterranean seagrass (Posidonia oceanica) meadows under current and future summer temperatures. J. Geophys. Res. Biogeosciences 127, e2022JG006946. doi:10.1029/2022JG006946
Ruiz-Reynés, D., Gomila, D., Sintes, T., Hernández-García, E., Marba, N., and Duarte, C. (2017). Fairy circle landscapes under the sea. Sci. Adv. 3, e1603262. doi:10.1126/sciadv.1603262
Ruiz-Reynés, D., Mayol, E., Sintes, T., Hendriks, I., Hernández-García, E., Duarte, C., et al. (2023). Self-organized sulfide-driven traveling pulses shape seagrass meadows. Proc. Natl. Acad. Sci. U. S. A. 120, e2216024120. doi:10.1073/pnas.2216024120
Salinas, C., Duarte, C. M., Lavery, P. S., Masque, P., Arias-Ortiz, A., Leon, J. X., et al. (2020). Seagrass losses since mid-20th century fuelled CO2 emissions from soil carbon stocks. Glob. Change Biol. 26, 4772–4784. doi:10.1111/gcb.15204
Scheffer, M., and Beets, J. (1994). Ecological models and the pitfalls of causality. Hydrobiologia 275-276, 115–124. doi:10.1007/bf00026704
Scheffer, M., Hosper, S. H., Meijer, M. L., Moss, B., and Jeppesen, E. (1993). Alternative equilibria in shallow lakes. Trends Ecol. and Evol. 8, 275–279. doi:10.1016/0169-5347(93)90254-M
Spivak, A. C., Sanderman, J., Bowen, J. L., Canuel, E. A., and Hopkinson, C. S. (2019). Global-change controls on soil-carbon accumulation and loss in coastal vegetated ecosystems. Nat. Geosci. 12, 685–692. doi:10.1038/s41561-019-0435-2
Thorhaug, A., Poulos, H. M., López-Portillo, J., Ku, T. C. W., and Berlyn, G. P. (2017). Seagrass blue carbon dynamics in the Gulf of Mexico: stocks, losses from anthropogenic disturbance, and gains through seagrass restoration. Sci. Total Environ. 605–606, 626–636. doi:10.1016/j.scitotenv.2017.06.189
Trevathan-Tackett, S. M., Macreadie, P. I., Sanderman, J., Baldock, J., Howes, J. M., and Ralph, P. J. (2017). A global assessment of the chemical recalcitrance of seagrass tissues: implications for long-term carbon sequestration. Front. Plant Sci. 8, 925. doi:10.3389/fpls.2017.00925
Unsworth, R. K. F., Collier, C. J., Waycott, M., Mckenzie, L. J., and Cullen-Unsworth, L. C. (2015). A framework for the resilience of seagrass ecosystems. Mar. Pollut. Bull. 100, 34–46. doi:10.1016/j.marpolbul.2015.08.016
Unsworth, R. K. F., Cullen-Unsworth, L. C., Jones, B. L. H., and Lilley, R. J. (2022). The planetary role of seagrass conservation. Science 377, 609–613. doi:10.1126/science.abq6923
van der Heide, T., van Nes, E. H., Geerling, G. W., Smolders, A. J. P., Bouma, T. J., and van Katwijk, M. M. (2007a). Positive feedbacks in seagrass ecosystems: implications for success in conservation and restoration. Ecosystems 10, 1311–1322. doi:10.1007/s10021-007-9099-7
van der Heide, T., van Nes, E. H., Geerling, G. W., Smolders, A. J. P., Bouma, T. J., and van Katwijk, M. M. (2007b). Positive feedbacks in seagrass ecosystems: implications for success in conservation and restoration. Ecosystems 10, 1311–1322. doi:10.1007/s10021-007-9099-7
van Nes, E. H., Arani, B. M. S., Staal, A., van der Bolt, B., Flores, B. M., Bathiany, S., et al. (2016). What do you mean, ‘tipping point. Trends Ecol. and Evol. 31, 902–904. doi:10.1016/j.tree.2016.09.011
Keywords: blue carbon, tipping cascade, coastal ecosystem, ecosystem function, regime shift, climate change, biogeochemical model
Citation: Dakos V, Le Vilain A, Thebault E, Alcoverro T, Boada J, Infantes E, Krause-Jensen D, Marbà N, Serrano O, Vizzini S and Apostolaki ET (2025) Carbon storage of seagrass ecosystems may experience tipping points in response to anthropogenic stress - a modeling perspective. Front. Complex Syst. 3:1534330. doi: 10.3389/fcpxs.2025.1534330
Received: 25 November 2024; Accepted: 17 February 2025;
Published: 18 March 2025.
Edited by:
Rosa Maria Benito, Polytechnic University of Madrid, SpainReviewed by:
Murugeswari Durairaj, Thiagarajar College, IndiaCopyright © 2025 Dakos, Le Vilain, Thebault, Alcoverro, Boada, Infantes, Krause-Jensen, Marbà, Serrano, Vizzini and Apostolaki. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vasilis Dakos, dmFzaWxpcy5kYWtvc0B1bW9udHBlbGxpZXIuZnI=; Eugenia T. Apostolaki, ZWFwb3N0QGhjbXIuZ3I=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.