 Oriana E. Chafe1,2,3†
Oriana E. Chafe1,2,3† Adrian P. Broz4,5*†
Adrian P. Broz4,5*† Eric S. Levenson1†
Eric S. Levenson1† Michael D. Farinacci1†Riley O. Anderson2†
Michael D. Farinacci1†Riley O. Anderson2† Lucas C. R. Silva1,2,3
Lucas C. R. Silva1,2,3- 1Environmental Studies Program, University of Oregon, Eugene, OR, United States
- 2Department of Biology, University of Oregon, Eugene, OR, United States
- 3Institute of Ecology and Evolution, University of Oregon, Eugene, OR, United States
- 4Department of Earth, Atmospheric and Planetary Science, Purdue University, West Lafayette, IN, United States
- 5Department of Earth Sciences, University of Oregon, Eugene, OR, United States
Natural climate solutions have been proposed as a way to mitigate climate change by removing CO2 and other greenhouse gases from the atmosphere and increasing carbon storage in ecosystems. The adoption of such practices is required at large spatial and temporal scales, which means that local implementation across different land use and conservation sectors must be coordinated at landscape and regional levels. Here, we describe the spatiotemporal domains of research in the field of climate solutions and, as a first approximation, we use the Pacific Northwest (PNW) of the United States as a model system to evaluate the potential for coordinated implementations. By combining estimates of soil organic carbon stocks and CO2 fluxes with projected changes in climate, we show how land use may be prioritized to improve carbon drawdown and permanence across multiple sectors at local to regional scales. Our consideration of geographical context acknowledges some of the ecological and social challenges of climate change mitigation efforts for the implementation of scalable solutions.
Introduction
Natural Climate Solutions (NCS) are a set of strategies that involve the conservation, restoration, and sustainable management of ecosystems to enhance their ability to capture and store carbon dioxide from the atmosphere. These solutions leverage the natural processes of ecosystems to mitigate climate change by sequestering carbon and reducing greenhouse gas emissions. Natural climate solutions consist of land conservation, restoration, and management efforts that have potential to mitigate climate change by removing CO2 and other greenhouse gasses from the atmosphere, as well as by increasing carbon (C) storage in terrestrial and nearshore ecosystems (e.g., Griscom et al., 2017). NCS can also achieve mitigation by avoiding emissions from changes in management or land use (e.g., manure acidification to avoid methane release, or protecting forests from conversion to avoid emissions from logging and subsequent decomposition of organic material) (Cook-Patton et al., 2021). Examples of NCS practices include reforestation and other forest management practices, regenerative agricultural practices such as cover cropping (Drever et al., 2021), biochar application, and ecosystem restoration (Graves et al., 2020). In the USA, many land-based NCS techniques are readily deployable and could mitigate about 21% of the annual atmospheric CO2 emitted nationwide (Fargione et al., 2018).
However, the risks and benefits of NCS for people and ecosystems have yet to be fully assessed at local (tens of km2) to regional (hundreds to thousands of km2) scales. Simply put, these techniques will not work everywhere (Anderson et al., 2019), and in some cases, they may cause unanticipated release of CO2 or a decrease in the mean residence time of carbon (Silva et al., 2022). Furthermore, many of the social and ecological factors that shape ecosystem dynamics and determine the overall success of potential NCS have yet to be quantified and incorporated into NCS frameworks (Silva, 2022). To advance research in this field, we explore the interactions between plants, soils and climate change. To enhance our understanding of the mechanisms that control carbon storage and release across ecosystems, we show how the majority of previously published studies of NCS often fail to address local and regional spatial scales as well as drawdown and storage over long (>100 years) temporal scales. Our objective is to build on existing knowledge to address data gaps and create an adaptable framework for prioritizing areas for NCS research and implementation.
Successfully implementing NCS requires a framework that is consistent at multiple spatial and temporal scales. Such a framework must consider local social and ecological factors that can be missed within large-scale efforts. For example, not all landscapes may be appropriate for NCS projects. Some have competing socioeconomic uses (e.g., forestry, agriculture, and/or indigenous land) (Hasegawa et al., 2018), some are projected to become too dry to sustain growing plant biomass (Marvin et al., 2023), and others are already rich in soil organic carbon (SOC) (Bossio et al., 2020). The variation in land use and resource availability constraints both the technical and realizable potential for C drawdown and permanence because no single NCS method is appropriate for all bioclimatic conditions (Baldocchi et al., 2018). Thus, scalable projects would require coordination of land use, conservation, and restoration efforts (Bossio et al., 2020; Silva, 2022). Here, we present a geographic-based framework for guiding NCS implementation using soil C stocks (a proxy for belowground sequestration) and projected climate change (a control on future drawdown potential) across multiple ecoregions in the Pacific Northwest (PNW) of the United States. In accordance with a “protect-manage-restore” approach to NCS implementation (Cook-Patton et al., 2021) that also considers future changes in climate (Marvin et al., 2023), the ecoregions considered here provide examples of how sector-specific NCS activities (e.g., forestry and agriculture) can be ranked relative to conservation and thus prioritized to inform decisions.
Within the exceptionally diverse bioclimatic, social, and ecological settings that exist in the Pacific Northwest, we chose ecosystems with exceptional NCS potential in the states of Oregon and Washington to serve as examples. We build on other examples of landscape prioritization for C sequestration (Cook-Patton et al., 2021; Law et al., 2022) and socioecological benefits through carbon markets (Aoyama et al., 2022; Bomfim et al., 2022), conservation and management that influence how carbon drawdown efforts affect the water cycle (Maxwell et al., 2018) as well as plant-soil feedbacks that influence nutrient use and carbon stabilization (Silva and Lambers, 2020) and lessons from other model regions where “fundamental mechanisms can be identified, understood, and tested” (Vitousek and Loope, 1987). The resulting NCS priority areas identified below serve as a delineation of high-priority NCS locations, as a first approximation toward a framework for implementation at regional to local scales.
Methods
Inspired by previous studies of spatial and temporal domains in ecology (Estes et al., 2018), we set out to conduct a systematic meta-analysis using data from a literature review that covers most NCS studies published through 2023. To assess the general representation of spatial and temporal scales that have been considered in the NCS literature, we gathered data from previously published studies about NCS (Figure 1). We reviewed 55 studies published from 2011 to 2023 that contained the keywords “Natural Climate Solutions” or “Nature-based Solutions”. These studies used either a field-based and/or a modeling approach for estimating CO2 sequestration potential of NCS, which allowed for approximating the spatial and temporal extent considered in each study (Supplementary material). The frequencies of spatial and temporal observations are displayed as histograms to illustrate the general scales that have been traditionally considered in theoretical and experimental approaches to NCS implementation (Figure 1). If no specific spatial scale was mentioned by the original study authors, we approximated the spatial and temporal scales of the study by (a) estimating the spatial extent of the modeled study area as categorical data (e.g., global, national, regional, or local) which were then binned into the approximate area of proposed NCS implementation (km2). Temporal scales of published studies, if not author-identified, were approximated by using boundary conditions of the study. For example, if a study estimated the climate mitigation potential in 2020 and 2100 (GtCO2 eq. yr−1) of a NCS, we approximated the temporal extent of the study to be 20–100 years. As many studies reported C permanence on temporal scales of ~20, 50, or 100 years (e.g., predictions for the years 2040, 2070, or 2100), we considered a bin of 20–100 years to accommodate this range. It should be noted that this approach considers a large temporal scale (e.g., 80 years) that has large variations in mitigation potential and associated uncertainties (e.g., Griscom et al., 2017).
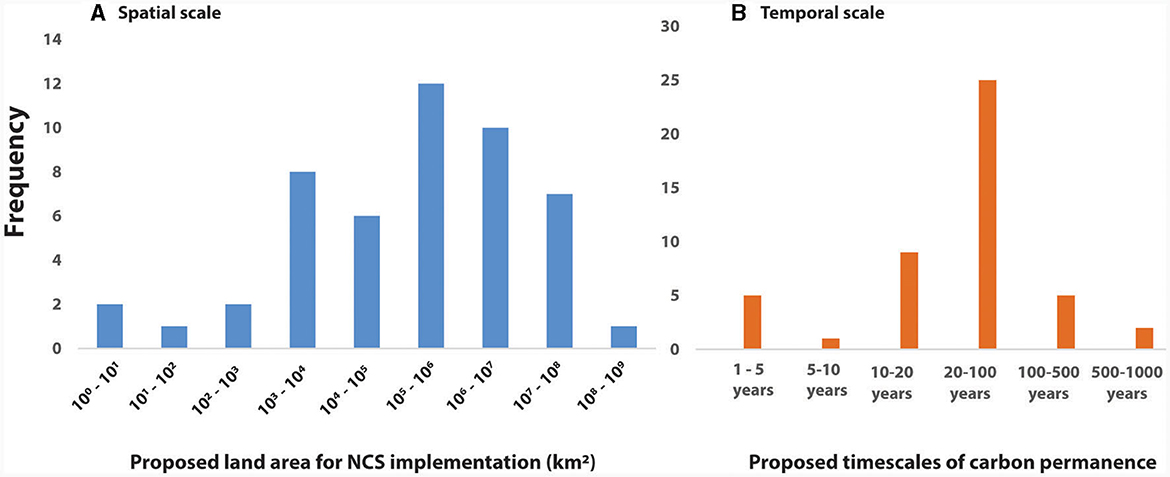
Figure 1. Data from previously published work showing the spatial and temporal domains of natural climate solutions (NCS) research. (A, B) Histograms of (A) the spatial scale of NCS implementation and (B) temporal scales of carbon permanence collectively gathered from a survey of 30 studies focused on natural climate solutions. Local scales are defined here as ~101-102 km2 and regional scales are ~102-103 km2.
A large proportion of the papers reviewed propose various NCS at a national or global scale, and thus precise estimates on the scales of NCS areas have large associated uncertainties. For example, many NCS papers cover a range of practices that vary widely in their spatial extent. We dealt with this variability by considering the spatial scales of all of the proposed NCS in each paper, not for each individual/specific practice (e.g., the modeling of GHG drawdown from NCS was at a national or global level).
We next used available geospatial data that caters to local and/or regional scales to prioritize areas for NCS implementation or conservation, ensuring a replicable approach (Figure 2). Here, we consider conservation as distinct from NCS, which include management and restoration. We acknowledge that most literature on NCS considers “conservation” as a type of NCS, but the ecoregions considered here provide examples of how sector-specific NCS activities (e.g., forestry and agriculture) can be ranked relative to conservation and thus prioritized to inform decisions.
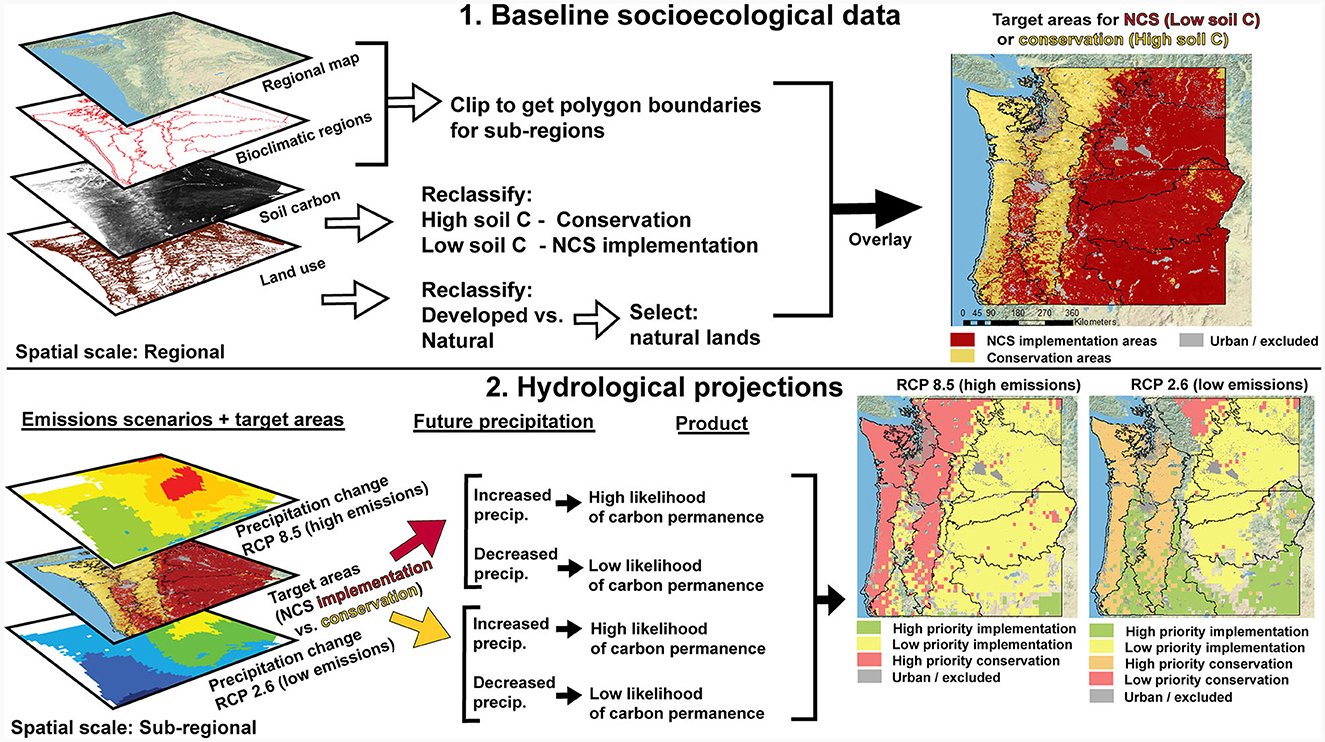
Figure 2. A proposed framework for prioritization of natural climate solutions (NCS) implementation areas or conservation across regional and local spatial scales. Step 1 uses baseline data of a socio-ecological context including bioclimatic zones, soil carbon stocks, and land use to identify target areas for NCS implementation and conservation, respectively. Step 2 combines those target areas with future projections of precipitation using both high and low emission scenarios to rank areas by priority.
Bioclimatic regions are defined here by Washington and Oregon state Level III ecoregions, defined by the EPA as “areas of general similarity in ecosystems and in the type of quality, and quantity of environmental resources”. These regions are necessary for the proper structure and execution of ecosystem management strategies across agencies and non-governmental organizations accountable for the natural resources within a geographic area (U. S. EPA, 2020). Terrestrial baseline soil carbon was based on 250 m spatial resolution soil data from the World Soil Information Service database and accessed via SoilGridsTM version 2.0. Tidal wetlands carbon was derived from the North American Carbon Project (soilgrids.com, Hinson et al., 2019).
Land use data was categorized as developed versus natural lands and was derived from the Washington State Department of Ecology 2010 State Land use dataset and the Oregon Development Project–2014 dataset, originated by the USFS Pacific Northwest Research Station. The projected precipitation data was collected via the Downscaled CMIP3 and CMIP5 Climate and Hydrology Projections (https://gdo-dcp.ucllnl.org/downscaled_cmip_projections/), which is a collaborative archive containing fine spatial resolution climate projections over the contiguous United States.
Finally, we propose a combination of biophysical and land use maps that consider existing carbon baseline data into opportunity maps for drawdown and permanence under future climates. To this end, we used the downscaled version of CMIP5 multi-model ensemble output, which utilizes 1/8° resolution bias corrected precipitation projections (Reclamation, 2013) that were produced by running period analyses using the MIROC5 (Model for Interdisciplinary Research on Climate), applying RCP2.6 as an emission scenario that leads to low greenhouse gas concentrations, or RCP8.5 which is a baseline scenario in the absence of a climate change policy. The RCP2.6 scenario assumes substantial changes in energy use, but a large increase in global cropland area. In contrast, the RCP8.5 scenario represents a business-as-usual scenario. MIROC5 is an atmosphere-ocean general circulation model that was selected for its wide-use with improved results especially found in precipitation (Watanabe et al., 2010), created by the Atmosphere and Ocean Research Institute (The University of Tokyo), National Institute for Environmental Studies, and Japan Agency for Marine-Earth Science and Technology.
Results and discussion
Observational scales of natural climate solutions
A compilation of previously published literature shows that spatial and temporal approaches to examining NCS have two broad distributions (Figure 1). First, the spatial extent of NCS research has largely been focused at the regional, national and global scales. For example, many studies that take a modeling approach to assessing NCS feasibility are focused on large (national and global) scales. The temporal scale of NCS implementation, e.g., the duration of carbon permanence, has a normal distribution, with most studies considering temporal scales of 20–100 years (Figure 1) for carbon permanence. A focus on such large spatial scales and may fail to consider local and regional challenges to NCS (Mu et al., 2019). Similarly, the focus of carbon permanence on short temporal scales (e.g., 20 years) may not adequately consider the long-term C sequestration potential of agricultural NCS such as biochar application or enhanced mineral weathering, which may encourage C permanence over geologic time scales (e.g., 500–1,000 years), further discussed in the “Carbon Permanence” Section (Section Discussion).
It should be noted that our results for approximating scales of NCS were potentially influenced by several method-based issues which may have potential biases and uncertainty in quantification of scales. Several studies that modeled C sequestration potential of NCS did not precisely state the observational scales used, and therefore we had to estimate the scale values for most observations, especially those focused on the national or global scale. Our estimation errors are therefore incomplete and should be expanded in future studies. Despite this limitation, our analysis provides a general frame of reference to interpret the most common observations in the consideration of NCS implementation and monitoring. Results in Figure 1 highlight the need for a focus on local to regional scales that consider carbon permanence on time scales longer than 100 years. In the following sections, we address the opportunities and tradeoffs for NCS implementation across these spatial and temporal scales.
Guiding tradeoffs for NCS prioritization
This geographical framework for NCS implementation is focused on addressing two challenges within the process of research and implementation: (1) delineating locations that present opportunities for carbon drawdown, and (2) identifying areas that would benefit more from conservation than management for NCS implementation (Figure 2). Our suggestion for developing NCS strategies is guided by the tradeoffs that are inherent in ecosystem mining of CO2 from the atmosphere (Baldocchi et al., 2018). We use the carbon-water relationship (Figure 3) as an example of a tradeoff that applies across locations and spatial scales and can be leveraged to describe the site-specific implications of NCS.
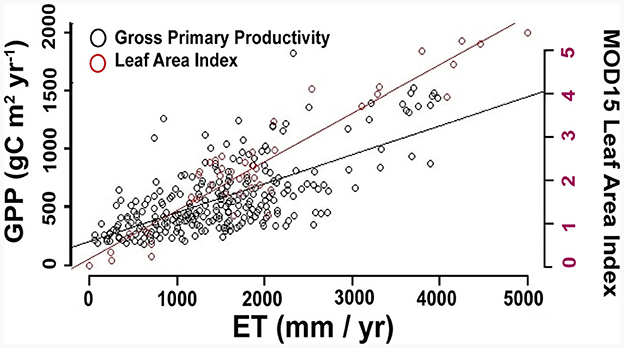
Figure 3. Relationships between evapotranspiration (ET), gross primary productivity (GPP), and leaf area index (LAI) (the “carbon-water tradeoff”). Data for the GPP and ET relationship were adapted from Baldocchi and Penuelas (2019). The GPP and ET values represent annual averages of eddy covariance measurements from the FLUXNET 2015 dataset (Pastorello et al., 2020) that includes 155 global flux towers (Supplementary Table 1) (Chu et al., 2021). The resulting linear regression is ET = 213.4 + 0.247 (GPP). Data for the LAI and ET relationship are from Hashimoto et al. (2012). LAI values represent annual averages from 2001 to 2008 from MODIS product cutouts surrounding 21 FLUXNET towers (Supplementary Table S2), and include tree LAI for agricultural settings, represented by the equation LAI = 0.76 + 0.00153 (ET).
The assimilation of carbon through vegetation is physically and biologically tied to water use and plant morphology (e.g., Silva and Lambers, 2020). Gross primary production (GPP) scales with evapotranspiration (ET) and Leaf Area Index (LAI) (Figure 3) quasi-linearly, showing that on average evapotranspiration increases by 0.247 millimeters for each increase in GPP per gram of carbon per square meter in a year [ET = 213.4+0.247(GPP)]. This is consistent with previous studies comparing annual estimates of CO2 and water loss through ET (Liles et al., 2019; Maxwell and Silva, 2020; Segura et al., 2021; Law et al., 2022). The scaling of GPP with ET, which varies in tandem with LAI within a given range of vapor pressure deficit, suggests that ecosystem functions relevant to NCS are strongly tied to ecosystem structure composition and climate (Hudiburg et al., 2013).
The significant variability about the mean regression line (Figure 3) is an important indicator of differences in ecosystem scale water use efficiency toward carbon drawdown. A reasonable interpretation of this variability is that sites or ecosystems represented by points falling below the regression line are more efficient with respect to water use than points falling above the regression line. Species composition and ecosystem structure are primary sources of variation in the relationship between GPP and ET (e.g., Migliavacca et al., 2021), and the regression between LAI and GPP in part explains the role and water cost of assimilating CO2 in diverse communities and ecosystems. However, this relationship does not fully explain the variability in the ET—GPP data. We suggest that future studies should prioritize characterizing the sources of this variability to better support efforts to optimize the balance between water loss and carbon gain through NCS implementation. Further, we emphasize that successful implementation of NCS management techniques will require optimizing CO2 fixation per unit of water lost to evapotranspiration, which relies upon characterizing the sources of variation in the carbon-water tradeoff for different ecosystems to inform conservation, management, or restoration decisions.
The role of plant species composition, morphology, and ecosystem structure in defining carbon-water tradeoffs can be extended to multiple spatiotemporal scales. For example, the evolution of vein density in leaves and wood transport structure can be used to reconstruct past and predict future canopy conductance and ecosystem responses to vapor pressure deficit and climate variability (e.g., Boyce et al., 2009; Earles et al., 2016; Sperling et al., 2017).
Within the Pacific Northwest, the primary forest types that are considered in NCS plans occur in mesic and dry landscapes. Mesic forests throughout the western Cascade and Coast Mountain Ranges are typically dominated by Douglas-fir (Pseudotsuga menziesii), while dry forests dominated by ponderosa pine (Pinus ponderosa) cover the eastern Cascades and interior mountain ranges (Figure 4). Ecophysiological differences cause inter- and intraspecific variation in growth rates throughout these forests (e.g., Hudiburg et al., 2013). In general, trees growing in mesic conditions have higher GPP and lower WUE (water-use efficiency) than their more arid counterparts (Ruzicka et al., 2017). Driven by biotic and abiotic conditions, water use efficiency varies across species, space, and time and as a result, these forests have higher carbon sequestration capacities but also higher rates of water loss through transpiration. In contrast, trees growing in more arid conditions have less of a water cost for carbon assimilation.
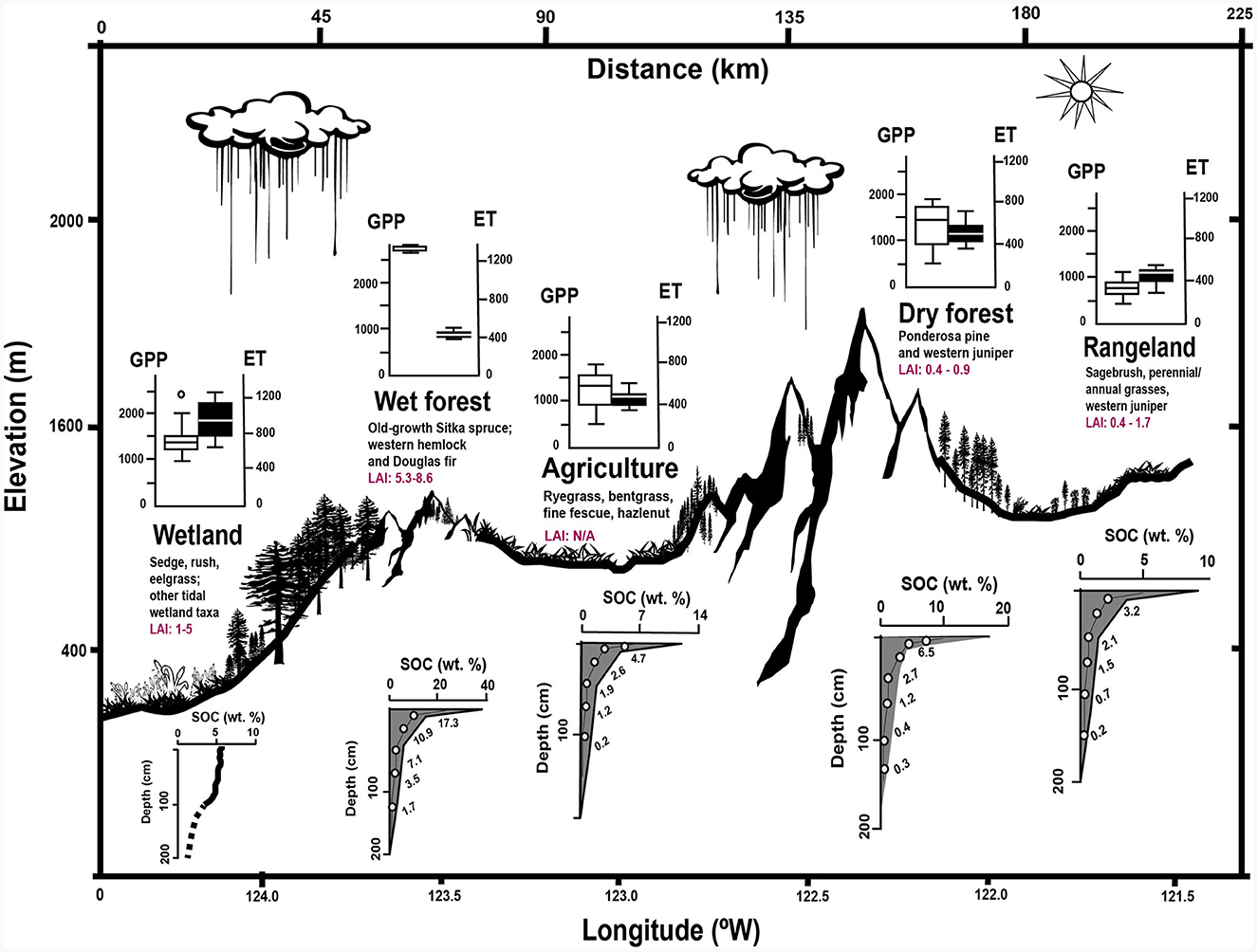
Figure 4. Carbon-water tradeoffs across a Pacific Northwest transect. Boxplots show the carbon-water tradeoff (Gross Primary Productivity [GPP] and evapotranspiration [ET]) across ecosystems (data from Pacific Northwest eddy covariance sites in Baldocchi and Penuelas, 2019 and Kwon et al., 2018 [wet forest data]). Leaf area index (LAI) data were from Bonan (2015); agricultural LAI was defined as N/A because of large seasonal LAI variations typical of agricultural settings (e.g., Du et al., 2022). Modeled soil organic carbon (SOC) data were derived from SoilGridsTM (soilgrids.org) by randomly selecting a point within each ecosystem (except wetlands) across a transect from Cape Perpetua, OR to Burns, OR. Mean derived SOC values are listed next to each data point. Modeled mean SOC from soilgrids.org was computed using a default random forests algorithm in the R package “ranger”. The 0.05 and 0.95 quantiles (gray shaded area) present the lower and upper boundaries of a 90% prediction interval. Wetland SOC is after Hinson et al. (2017) using National Wetlands Inventory data and the U.S. Soil Survey Geographic Database to model the area-weighted wetland SOC to 100 cm depth across all U.S. west coast tidal wetlands; SOC is estimated and not modeled from 100 to 200 cm (dotted line).
Agricultural areas in the PNW and elsewhere present additional uncertainties for estimating the carbon-water tradeoff. Agricultural areas typically exhibit low Leaf Area Index (LAI) for trees or shrubs, except in cases like orchards, silvopastoral systems, or agroforestry (e.g., He et al., 2020). However, crops in these areas often have high LAI for non-woody species (Du et al., 2022). A key overall distinction is the significant seasonal fluctuation of agricultural LAI, which perhaps limits its role as a stable and substantial carbon sink.
This variation in water-carbon relations is of significant consequence to the applicability of NCS implementation at a regional scale (Figure 4). In Oregon, the GPP of a mesic Douglas fir-dominated wet forest is 2,590 gCm−2yr−1 vs. 1,631 gCm−2yr−1 in dry forests of ponderosa pine approximately 160 km to the east (Kwon et al., 2018). Critically, the intrinsic WUE of semi-arid ponderosa pine forests is estimated to be approximately four times greater than in mesic Douglas-fir forests (Kwon et al., 2018).
Tree ring analysis of Douglas-fir sites along a ~160 km North-South transect in western Oregon show greater growth in wetter sites vs. higher intrinsic WUE in drier sites (Ruzicka et al., 2017). While moisture availability is primary control on plant growth, additional parameters such as geomorphological setting, soil nutrient availability, growing season length further constrain NCS potential (Maxwell et al., 2018). Our preliminary maps (Figure 5) point to areas where NCS implementation is desirable, such as across the Willamette Valley in Oregon, but there are additional considerations that we believe will ultimately determine the effectiveness of NCS. These include considerations of past and current soil conditions and land cover type, which has implications for baseline conditions and for increasing carbon permanence across landscapes, especially across PNW agricultural areas (e.g., Waldo et al., 2016) where precipitation is modeled to increase under both RCP pathways (Karimi et al., 2017; Mu et al., 2019). Prioritization of conservation or management for NCS across several ecosystems of the PNW “natural laboratory” (Figures 4, 5) are discussed in the following paragraphs.
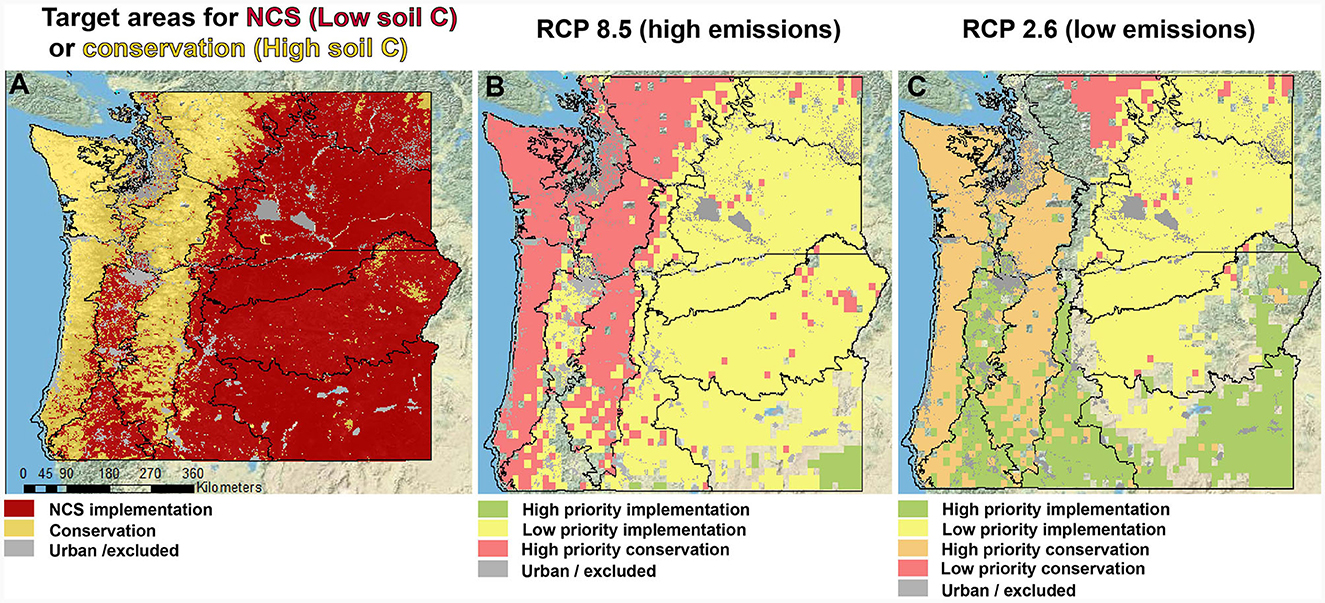
Figure 5. Data products for NCS prioritization areas in the Pacific Northwest considering a carbon-water tradeoff under different emissions scenarios. (A) Consideration of bioclimatic zones, soil carbon stocks, and land use to identify target areas for NCS implementation, where soil C is low (<5 wt. % soil organic carbon [SOC]) or high (<5 wt. % SOC); (B, C) combine target areas (red) from (A) and incorporate future projections of precipitation using both high and low emission scenarios to rank areas by priority, where “high priority implementation” areas have low SOC and projected increases in precipitation.
Wet forest
Globally, carbon- and water-rich habitats are often co-located and spatiotemporally correlated (REF). Consistent with this global pattern, some of Earth's most carbon-rich forests lay within the wet temperate zone in the humid PNW (Figures 4, 5). The measured carbon densities from PNW forests are higher than all other types of vegetation across the globe (Smithwick et al., 2002); however, with warming, these forests could become carbon sources, as increasing temperatures impact plant and soil respiration rates and the inter-annual variability of net ecosystem exchanges (Baldocchi et al., 2018). Climate projections suggest an increased warming and increased precipitation for the region, where droughts are likely to occur between intense precipitation storms (IPCC, 2014). The projected warming and fluctuations in wetness for this region will likely impact productivity rates because the photosynthetic response of mid- and high-latitude plants to increased warming depends upon soil moisture, where the effect will be positive when moisture is sufficient and negative were soils are drier (Reich et al., 2018). This is one example of the trade-offs that occur between water and carbon that have implications for this ecosystem to sequester carbon.
In Figure 4 we show that coastal forests, as one example of wet forests, have a similar range of GPP and ET values, where the cost of maintaining a high level of GPP requires more ET than other ecosystems. This pattern is not ubiquitous for all wet forests, however, and it varies upon management history and microclimate. The coastal forest ecoregion in the PNW has been more intensely harvested in comparison to the cascades due to its largely fragmented patchwork of ownership with differing management goals (Creutzburg et al., 2017). The climate of the two ecoregions can differ considerably from the interaction of maritime and continental air masses that impacts precipitation amounts, timing and whether it falls as snow or rain (Waring and Franklin, 1979). NCS that consider these geographic constraints are more likely to be successful, as conserving ecoregions with high levels of baseline carbon stocks will be dependent upon water-carbon tradeoffs. The spatial extent of NCS conservation areas across wet forests of western Oregon presented here (Figure 5) overlap with previously published regional frameworks for maximizing C storage and preserving biodiversity through the establishment of strategic forest reserves in Oregon (Law et al., 2022).
With the goal of increasing carbon drawdown and permanence, we suggest that NCS in coastal PNW forests of Oregon and Washington (Figure 5) should be focused on conserving existing carbon in the form soil organic carbon and, whenever possible, expanding ecological restoration and forestry for sustainable timber management and extended rotations. Ecological forestry favors the removal of smaller amounts of trees and longer harvest cycles promote carbon sequestration and overall forest health relative to clearcutting (Franklin et al., 2018). Our suggestion is consistent with the guiding principles of Ecological Forestry, which has been favored by the Northwest Forest Plan (NWFP) as a way to reduce harvesting of old-growth forests, while also allowing for clearcutting and thinning in certain contexts. Furthermore, NCS recommendations for conservation of wet forests with high baseline aboveground and soil C (Figures 4, 5) are consistent with previous work in the PNW calling for protection of large trees (>51 cm diameter), which dominate aboveground carbon storage in both wet and dry forests (Mildrexler et al., 2020). Such trees can account for just 3% of all stems in PNW forests but hold 42% of total aboveground carbon (Mildrexler et al., 2023).
Our analysis also suggests that large scale shifts from mature/old forests toward dense tree plantations might be unsustainable under future climates because it could compromise the provision of water for people and ecosystems, increasing the risk of catastrophic wildfire and related carbon emissions, as already observed in some regions within the PNW and elsewhere (Perry and Jones, 2017; Jerrett et al., 2022). To address this issue, we suggest a greater emphasis on integrated landscape management practices intended to preserve and increase baseline carbon stocks. These may include longer forestry rotations with riparian buffers (Segura et al., 2021), which have some of the highest carbon sequestration potential in the region (Graves et al., 2020), with the possibility of including community-led prescribed fires to decrease the risk of wildfire (Lake et al., 2017).
Dry forest
The dry forests of the eastern Cascades, Klamath, and interior mountains are dominated by ponderosa pine (Figure 4) and a frequent, low to mixed-severity fire regime (Halofsky et al., 2020). In this region, projected changes include increasing temperatures and hydrologic cycle intensification resulting in increased drought frequency and severity. These changes are anticipated as moisture loss during the prolonged summer dry season is amplified by warmer temperatures while precipitation increases are restricted to winter months. Increases in the annual number of large fires in the western United States that were consistent with increased drought severity and reflect long-term patterns that are to be expected with climatic changes (Dennison et al., 2014). Both plant species composition and ecosystem productivity are projected to change due to changes in growing-season length, precipitation, and climatic water deficits (Westerling, 2016). As a result, fire regimes are expected to be impacted by interacting effects of short-term variations in productivity and long-term vegetation shifts. These shifts will not only impact NCS capacity, but also the potential permanence of carbon sequestered going forward (Liang et al., 2018).
East of the Cascades, landscapes tend to support less productive, drier conifer forests (Figure 4), which are typically subject to frequent fires of low to mixed severity. Due to the variation in forest productivity, land use, and historical fire patterns, differences in Non-Industrial Private Forest (NIPF) management and owner responses are expected across the ecoregions. This expectation led to the necessity of using a stratified approach to prioritize landscape sampling and design of NCS projects.
Fuel is the most significant control on fire severity in these forests, followed by fire weather, climate, and topography, respectively (Parks et al., 2018). As a consequence, land management decisions have critical implications for the amount of carbon released during a fire event as well as the trajectory of ecosystems post-fire. This is especially important given evidence that the frequency of extreme fire weather is increasing (Parks et al., 2018; Davis et al., 2019). Additionally, patterns of repeat fires suggest that fine fuel accumulation is a strong control on disturbance frequency intervals. This pattern is spatially dependent (coastal areas with higher productivity can re-burn sooner than drier interior areas), indicating that the structure of forest vegetation is an important component of fire regime frequency and severity (Buma et al., 2020).
In this way, fuels and fire management planning is the Natural Climate Solution. All NCS activities must consider the preexisting fire regime to ensure the long-term effectiveness of C drawdown, favoring riparian restoration and conservation efforts over expanding tree plantations within landscapes that may be primed to burn at high severity.
Managed burning (the alternative to active fire suppression) under non-extreme fire weather conditions has been suggested as a pathway to decrease fire severity over the next century (Parks et al., 2016) and is applicable to our study area in the PNW (Figure 4). A small proportion of fires are responsible for ~95% of the annual area burned. By aggressively suppressing low-severity fires, we paradoxically select for high-severity fires by increasing fuel on the landscape (Dunn et al., 2017). As a result, when fires do escape they burn at higher severity and release more carbon (Parks et al., 2018). The combination of high fuel loads resulting from a century of fire suppression activities, warmer climatic conditions and extreme fire weather interact to increase the frequency of high-severity, stand-killing fires (Taylor et al., 2016). Our analysis suggests that current bioclimatic conditions and modeled changes in precipitation (Figure 5) could serve as a blueprint to understanding and predicting the success of NCS projects under predominant fire regime and help to inform appropriate landscape specific conservation and/or management actions (Figure 2). This approach is aligned with delineations at local to regional scales of dry forest areas in Oregon and Washington where fire could be reintroduced to thin understory vegetation (Law et al., 2023).
Tidal wetlands
Tidal wetlands are valuable carbon sinks present within the Coast Range ecoregion (Figure 4). The wetlands have been reduced for agriculture and pastureland via implementation of dikes (Boule and Bierly, 1987), yet legislative protections in the Pacific Northwest have limited conversions, and therefore the potential for reducing GHGs is estimated to be limited to ~5,200 ha of degraded area (Graves et al., 2020). Despite the limited geographic extent, tidal wetlands hold the highest carbon per unit area (Griscom et al., 2017) and the Pacific Northwest coast contains some 1.4–1.6 million tons of carbon (Hinson et al., 2019) (Figure 4). Albeit less influenced by precipitation changes than other ecosystems, net CO2 emissions from restored tidal wetland is dependent on initial greenhouse gas inputs, rainfall, and elevation (Negandhi et al., 2019). Tidal reinstatement in higher elevation sites during flood events can result in lower CO2 emissions and unchanged CH4 emissions (Negandhi et al., 2019).
The carbon-water tradeoff for NCS implementation in tidal wetlands is distinct from other ecosystems considered in this work (Figure 4). For example, high ET may not be as detrimental to this aquatic landscape as it is to other ecosystems in our model region (Figure 4). Instead, anthropogenic activities such as dredging, nutrient loading in the watershed via logging or runoff, coastal development, and introduction of invasive species are more serious threats to tidal wetlands and seagrass establishment with the potential to convert even reinstated tidal wetlands from carbon sinks to sources (Short and Wyllie-Echeverria, 1996). This “coastal squeeze” in which coastal ecosystems like seagrass meadows and salt marshes, both of which enhance carbon storage through aquatic vegetation, are stuck between sea level rise, increasing sea temperatures, and coastal infrastructure, is a prominent threat to tidal wetlands (Mills et al., 2016).
Varying soil organic carbon stock in PNW estuaries also provides opportunities and challenges for prioritizing NCS implementation (Figure 5). In general, carbon storage in estuaries varies by latitude and estuary size (Hinson et al., 2019). However, conservation and management decisions can greatly improve NCS potential in these systems. For example, a spatial analysis combining extensive sampling across salinity gradients and management regimes (disturbed, restored, and reference) in 22 wetland sites across two Oregon estuaries, demonstrates that it is possible to estimate GHG fluxes in both existing and former wetlands in the region, from a small set of environmental data combined with past and current land use data (Schultz et al., 2023). This requires collecting environmental data that accurately captures the variability of the site and spans a sufficient time frame to account for seasonal changes (e.g., Pham et al., 2019), ideally covering at least a full year and employing a widespread carbon flux sampling method that utilizes machine-learning. Adopting this method could offer a cost-effective way to assess the viability of wetland restoration as a strategy for climate mitigation. Smaller tidal wetland zones are more sensitive to dredging, the “coastal squeeze”, and poor water quality. Seagrass presence can increase carbon storage and can delineate where conservation vs. implementation should take place within these “blue carbon” zones. In other words, tidal wetlands with dense seagrass meadows in the PNW should be preserved, whereas areas that are bare but capable of hosting native species, such as eelgrass (Zostera marina), should be targeted for conservation and restoration.
Similarly, seagrass conservation and restoration efforts can be used to increase carbon capture with multiple benefits. Prioritization based on pre- and post-event disturbance data collection has been proposed to leverage long-term monitoring with new machine learning approaches for developing funding streams to support flexible decisions and prepare for extreme future conditions (Aoki et al., 2022). Although seagrass restoration is generally thought to have low success rates in the PNW (Fonseca et al., 1988; Stamey, 2004), compelling results have been obtained in areas where removing dikes to reinstate tidal wetlands can be implemented to encourage seagrass expansion and eradication of invasive species (Herrick and Wolf, 2005). Indeed restoration and conservation of tidal wetlands has many benefits when compared with business-as-usual use of such ecosystems, including implications for net carbon storage following disturbance events, but many questions remain with respect to carbon source and permanence in tidal wetlands. How much of the carbon is sequestered in situ vs. deposited from other systems? How long will the sequestered carbon be stored? We suggest that these and other questions should be considered in future efforts to prioritize NCS implementation especially across areas containing significant terrestrial and aquatic carbon sinks.
Carbon permanence
The long-term success of NCS ultimately depends on carbon permanence, defined as the mean residence time of sequestered carbon across landscapes (Figure 1). How long will carbon (C) be sequestered after NCS implementation? What use are NCS if sequestered C is rapidly released? We use carbon permanence here to describe C sequestration over timescales relevant to the current carbon budget outlined by Goldstein et al. (2020) to limit warming to 1.5°C above preindustrial levels. Natural climate solutions have the potential to sequester between 100 and 550 Gt CO2 between 2018 and 2100 at a cost of < $100 per ton carbon (Griscom et al., 2017), but ensuring that sequestered C is sufficiently “permanent” will determine the ultimate success of NCS. For example, previously proposed approaches to increase carbon stocks on land through afforestation, even when considering soil type (Bastin et al., 2019) generally do not emphasize baseline soil organic carbon stocks, which in some cases can be higher in unmanaged savannas and grasslands than in planted forests. Thus, if implemented through uniform tree plantings across landscapes, NCS efforts could result in unintentional liberation of C from areas with high baseline soil C (Figure 5A). Inherent differences in soil C stocks across ecosystems demand landscape-specific solutions for encouraging long-term soil C sequestration. For example, geomorphological and ecological context can be used for better predicting baseline conditions as well as projecting the potential for additional landscape carbon capture (Roering et al., 2023).
On land, soil organic matter (SOM) is the largest terrestrial pool of organic C, containing more organic C than global vegetation and the atmosphere combined (Lehmann and Kleber, 2015). Spatial prioritization of NCS (Figure 5) requires a baseline assessment of SOM: how old is soil C, and how much variation is there on C residence time across ecosystems? A multitude of environmental and biological controls determine C stability in soils (Schmidt et al., 2011) and sediments (Nelson and Baldock, 2005), including those of the PNW study region (Figure 4). Sorption of organic molecules to the surfaces of minerals and amorphous colloids influences the biochemical stability of SOM (Kleber et al., 2005; Plante et al., 2011). Organisms in the rhizosphere decompose soil C to progressively smaller organic compounds which encourages reactivity with mineral surfaces and/or formation of aggregates, which promotes soil C permanence (Schmidt et al., 2011). This and other factors ultimately lead to large differences in residence time of soil C across ecosystems in the PNW and elsewhere (Lehmann and Kleber, 2015) and many mysteries remain to be solved with respect to the stability of different SOM fractions, their potential sources, and permanence.
The molecular composition of SOM alone does not determine carbon permanence (Schmidt et al., 2011), but molecular structure can be a proxy for biochemical stability of soil C (Lehmann and Kleber, 2015). Upwards of 90% of all biomolecules in the SOM pool are proteins, lignin, carbohydrates, lipids, and aliphatic and carbonyl compounds (Nelson and Baldock, 2005), and the proportion of each differs across ecosystems, soil types, and climate regimes. Plant and microbial lipids are more recalcitrant relative to cellulose (Silva et al., 2015) and can persist in buried, ancient soils (paleosols) for tens of thousands of years or longer (Marin-Spiotta et al., 2014), suggesting that NCS management strategies that promote increased abundances of soil lipids may effectively increase carbon permanence in soils. Similarly, pyrogenic C (char) exhibits biochemical stability over thousands of years in early Holocene paleosols (Marin-Spiotta et al., 2014) and is found in much older paleosols (Retallack, 2019) from the late Jurassic (~180 million years old) (Matthewmann et al., 2019) and late Permian (~290 million years old) (Miller et al., 1996).
The persistence of pyrogenic C in paleosols suggests that pyrolysis of organic matter could encourage carbon permanence over long (e.g., geological) timescales (Broz, 2020). For this reason, NCS activities for agricultural areas in the PNW (Waldo et al., 2016) can include application of biochar (pyrogenic C) to cropping systems, which can provide climate mitigation potential on par with the widespread restoration of coastal wetlands (Griscom et al., 2017). Such techniques could potentially increase carbon permanence by two to three orders of magnitude relative to additions of more labile forms of soil organic matter (e.g., cellulose). Addition of biochar to natural and agricultural soils such as those in the Willamette Valley in Oregon (Figure 5) may therefore be an effective technique for increasing carbon permanence in the PNW region and elsewhere.
A next phase of understanding
In this study we explore the potential of existing baseline data combined with climate projections to orient NCS through coordinated land conservation or management. We use example ecosystems across the PNW to illustrate how local and regional decisions might reduce risks and improve benefits of NCS activities. We caution that concrete implementation plans must go beyond the general examples proposed here to consider social impacts and to evaluate ecologically appropriate contexts under current and projected bioclimatic conditions, ideally using a co-production of knowledge approach to quantify risks and benefits of widespread adoption in vulnerable communities. The recommended approaches for managing or conserving landscapes for NCS would require a substantial commitment of finances and resources, which goes beyond the scope of ecosystem processes discussed here. Recent advances in ecological monitoring may serve to address this obstacle and allow for the next phase of understanding in which bottom-up community-led NCS adoption can effectively increase carbon drawdown and permanence at local and regional scales.
Adoption of this framework for prioritizing NCS implementation yields three primary benefits: (1) By balancing NCS potential with carbon permanence potential, estimates of carbon sequestration take into consideration both the short-term and long-term dynamics of NCS implementation within the context of climatic change. (2) Adopting a scalable approach to parameterizing NCS potential reduces uncertainty in national sector-based estimates of carbon sequestration by considering variability in sequestration rates introduced by local geographic features, climatic conditions, and plant assemblages. (3) The flexibility of this framework aids prioritization of NCS implementation at a regional scale, while also enabling land managers to consider the suitability and potential tradeoffs of NCS implementation at the local and landscape-scales.
Future efforts should also include consideration of inorganic carbon sinks, such as enhanced weathering projects, which could have greater carbon sequestration potential than the biological processes described above globally. For example, agriculture could sequester 4.1 Pg CO2 eq. year−1; ecological restoration (e.g., protection and sustainable management of forests, savannas, and grasslands) could sequester 7.3 Pg CO2 eq. year−1, whereas enhanced weathering could sequester 1–100 Pg CO2 eq. year−1 although with much larger uncertainties (Shukla et al., 2019). Tailoring global NCS estimates and implementation efforts to specific regions with diverse social and ecological contexts is a challenge that requires further interdisciplinary research. Thus, future implementation efforts should involve stakeholders and researchers from diverse disciplines, sectors, and socioeconomic backgrounds.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
OC: Conceptualization, Data curation, Formal analysis, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing. AB: Conceptualization, Methodology, Visualization, Writing – original draft, Writing – review & editing. EL: Conceptualization, Methodology, Validation, Visualization, Writing – original draft, Writing – review & editing. MF: Conceptualization, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing. RA: Conceptualization, Visualization, Writing – original draft, Writing – review & editing. LS: Formal analysis, Methodology, Supervision, Validation, Visualization, Writing – original draft.
Funding
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
Acknowledgments
We acknowledge the World Climate Research Programme's Working Group on Coupled Modeling, which is responsible for CMIP and we thank the climate modeling groups for producing and making available their model output. For CMIP the U.S. Department of Energy's Program for Climate Model Diagnosis and Intercomparison provides coordinating support and led development of software infrastructure in partnership with the Global Organization for Earth System Science Portals.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fclim.2024.1273632/full#supplementary-material
References
Anderson, C. M. R. S, DeFries, R., Litterman, P. A., Matson, D. C., Nepstad, S., et al. (2019), Field, natural climate solutions are not enough. Science 363, 933–934. doi: 10.1126/science.aaw2741.
Aoki, L. R., Brisbin, M. M., Hounshell, A. G., Kincaid, D. W., Larson, E. I., Sansom, B. J., et al. (2022). Preparing aquatic research for an extreme future: call for improved definitions and responsive, multidisciplinary approaches. BioScience 72, 508–520. doi: 10.1093/biosci/biac020
Aoyama, L., Bartolome, J. W., Silva, L., and Silver, W. L. (2022). Using ecological site descriptions to make ranch-level decisions about where to manage for soil organic carbon. California Agricult. 2022, 1–8. doi: 10.3733/ca.2022a0007
Baldocchi, D., Chu, H., and Reichstein, M. (2018). Inter-annual variability of net and gross ecosystem carbon fluxes: a review. Agricult. Forest Meteorol. 249, 520–33. doi: 10.1016/j.agrformet.2017.05.015
Baldocchi, D., and Penuelas, J. (2019). The physics and ecology of mining carbon dioxide from the atmosphere by ecosystems. Global Change Biol. 25, 1191–1197. doi: 10.1111/gcb.14559
Bastin, J. F., Finegold, Y., Garcia, C., Mollicone, D., Rezende, M., Routh, D., et al. (2019). The global tree restoration potential. Science 365, 76–79. doi: 10.1126/science.aax0848
Bomfim, B., Pinag,é, E. R., Emmert, F., and Kueppers, L. M. (2022). Improving sustainable tropical forest management with voluntary carbon markets. Plant Soil. 479, 53–60. doi: 10.1007/s11104-021-05249-5
Bonan, G. (2015). Ecological Climatology: Concepts and Applications. 3rd Edn. Cambridge: Cambridge University Press.
Bossio, D. A., Cook-Patton, S. C., Ellis, P. W., Fargione, J., Sanderman, J., Smith, P., et al. (2020). The role of soil carbon in natural climate solutions. Nat. Sustainab. 3, 391–398. doi: 10.1038/s41893-020-0491-z
Boule, M. E., and Bierly, K. F. (1987). History of estuarine wetland development and alteration-what have we wrought. Northwest Environm. J. 3, 43–61.
Boyce, C. K., Brodribb, T. J., Feild, T. S., and Zwieniecki, M. A. (2009). Angiosperm leaf vein evolution was physiologically and environmentally transformative. Proc. Royal Soc. B: Biol. Sci. 276, 1771–1776. doi: 10.1098/rspb.2008.1919
Broz, A. P. (2020). Organic matter preservation in ancient soils of earth and mars. Life 10, 113. doi: 10.3390/life10070113
Buma, B., Weiss, S., Hayes, K., and Lucash, M. (2020). Wildland fire reburning trends across the US West suggest only short-term negative feedback and differing climatic effects. Environ Res. Lett. 15, 034026. doi: 10.1088/1748-9326/ab6c70
Chu, H., Luo, X., Quyang, Z., Chan, W. S., Dengel, S., and Biraud, S. C. (2021), Representativeness of eddy-covariance flux footprints for areas surrounding AmeriFlux sites. Agricult. Forest Meteorol. 301–302, 108350. doi: 10.1016/j.agrformet.2021.108350.
Cook-Patton, S. C., Drever, C. R., Griscom, B. W., Hamrick, K., Hardman, H., Kroeger, T., et al. (2021). Protect, manage and then restore lands for climate mitigation. Nat. Clim. Chang. 11, 1027–1034. doi: 10.1038/s41558-021-01198-0
Creutzburg, M. K., Scheller, R. M., Lucash, M. S., LeDuc, S. D., and Johnson, M. G. (2017). Forest management scenarios in a changing climate: trade-offs between carbon, timber, and old forest. Ecol. Appl. 27, 503–518. doi: 10.1002/eap.1460
Davis, K. T., Dobrowski, S. Z., Higuera, P. E., Holden, Z. A., Veblen, T. T., Rother, M. T., et al. (2019). Wildfires and climate change push low-elevation forests across a critical climate threshold for tree regeneration. Proc. Natl. Acad. Sci. USA. 116, 6193–6198. doi: 10.1073/pnas.1815107116
Dennison, P. E., Brewer, S. C., Arnold, J. D., and Moritz, M. A. (2014). Large wildfire trends in the western United States, 1984–2011. Geophys. Res. Lett. 41, 2928–2933. doi: 10.1002/2014GL059576
Drever, C. R., Cook-Patton, S. C., Akhter, F., Badiou, P. H., Chmura, G. L., Davidson, S. J., et al. (2021). Natural climate solutions for Canada. Sci. Adv. 7:eabd6034. doi: 10.1126/sciadv.abd6034
Du, L., Yang, H., Song, X., Wei, N., Yu, C., Wang, W., et al. (2022). Estimating leaf area index of maize using UAV-based digital imagery and machine learning methods. Sci. Rep. 12, 15937 doi: 10.1038/s41598-022-20299-0
Dunn, C. J., Thompson, M. P., and Calkin, D. E. (2017). A framework for developing safe and effective large-fire response in a new fire management paradigm. For. Ecol. Manage. 404, 184–196. doi: 10.1016/j.foreco.2017.08.039
Earles, J., Sperling, O., Silva, L. C. R., McElrone, A. J., Brodersen, C. R., North, M. P., et al. (2016). Bark water uptake promotes localized hydraulic recovery in coastal redwood crown. Plant Cell Environ. 39, 320–328. doi: 10.1111/pce.12612
Estes, L., Elsen, P. R., Treuer, T., Ahmed, L., Caylor, T., Chang, J., et al. (2018). The spatial and temporal domains of modern ecology. Nat. Ecol. Evol. 2, 819–826. doi: 10.1038/s41559-018-0524-4
Fargione, J. E., Bassett, S., Boucher, T., Bridgham, S. D., Conant, R. T., Cook-Patton, S. C., et al. (2018). Natural climate solutions for the United States. Sci. Adv. 4, 1–15. doi: 10.1126/sciadv.aat1869
Fonseca, M., Thayer, G. W., and Kenworthy, W. J. (1988). Restoration and management of seagrass systems: a review. Ecol. Manage. Wetlands 6, 401–420. doi: 10.1007/978-1-4615-9807-7_38
Franklin, J. F., Norman Johnson, K., and Johnson, D. L. (2018). Ecological Forest Management. Long Grove, IL: Waveland Press,
Goldstein, A., Turner, W. R., Spawn, S. A., Anderson-Teixeira, K. J., Cook-Patton, S., Fargione, J., et al (2020), Protecting irrecoverable carbon in Earth's ecosystems. Nat. Clim. Change 10, 287–295. doi: 10.1038/s41558-020-0738-8.
Graves, R. A., Haugo, R. D., Holz, A., Nielsen-Pincus, M., Jones, A., Kellogg, B., et al. (2020). Potential greenhouse gas reductions from natural climate solutions in Oregon, USA. PLoS ONE 15, e0230424. doi: 10.1371/journal.pone.0230424
Griscom, B. W., Adams, J., Ellis, P. W., Houghton, R. A., Lomax, G., Miteva, D. A., et al. (2017). Natural climate solutions. Proc. Nat. Acad. Sci. 114, 11645–11650. doi: 10.1073/pnas.1710465114
Halofsky, J. E., Peterson, D. L., and Harvey, B. J. (2020). Changing wildfire, changing forests: the effects of climate change on fire regimes and vegetation in the Pacific Northwest, USA. Fire Ecol. 16, 8. doi: 10.1186/s42408-019-0062-8
Hasegawa, P., Fujimori, S., Havlík, P., Valin, H., Bodirsky, B. L., Doelman, J. C., et al. (2018). Risk of increased food insecurity under stringent global climate change mitigation policy. Nat. Clim. Chang. 8, 699–703. doi: 10.1038/s41558-018-0230-x
Hashimoto, H., Wang, W., Milesi, C., White, M. A., Ganguly, S., Gamo, M., et al. (2012). Exploring simple algorithms for estimating gross primary production in forested areas from satellite data. Remote Sens. 4, 303–326. doi: 10.3390/rs4010303
He, L., Ren, X., Wang, Y., Liu, B., Zhang, H., Liu, W., et al. (2020). Comparing methods for estimating leaf area index by multi-angular remote sensing in winter wheat. Sci. Rep. 10, 13943 doi: 10.1038/s41598-020-70951-w
Herrick, B. M., and Wolf, A. T. (2005). Invasive plant species in diked vs. undiked Great Lakes wetlands. J. Great Lakes Res. 31, 277–287. doi: 10.1016/S0380-1330(05)70259-8
Hinson, A. L., Feagin, R. A., and Eriksson, M. (2019). Tidal Wetlands Soil Organic Carbon and Estuarine Characteristics, USA, 1972-2015. Tennessee, USA: ORNL DAAC.
Hinson, A. L., Feagin, R. A., Eriksson, M., Najjar, R. G., Herrmann, M., Bianchi, T. S., et al. (2017). The spatial distribution of soil organic carbon in tidal wetland soils of the continental United States. Global Change Biol. 23, 5468–5480. doi: 10.1111/gcb.13811
Hudiburg, T. W., Law, B. E., and Thornton, P. E. (2013). Evaluation and improvement of the Community Land Model (CLM4) in Oregon forests. Biogeosciences 10, 453–470. doi: 10.5194/bg-10-453-2013
IPCC (2014). “Climate Change 2014: Synthesis Report,” in Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change, eds. R. K. Pachauri, and L. A. Meyer (Geneva, Switzerland: IPCC), 151.
Jerrett, M., Jina, A. S., and Marlier, M. E. (2022). Up in smoke: California's greenhouse gas reductions could be wiped out by 2020 wildfires. Environ. Pollut. 310:119888. doi: 10.1016/j.envpol.2022.119888
Karimi, T., Stöckle, C.O., Higgins, S. S., Nelson, R. L., and Huggins, D. (2017). Projected dryland cropping system shifts in the Pacific northwest in response to climate change. Front. Ecol. Evol. 5:20. doi: 10.3389/fevo.2017.00020
Kleber, M., Mikutta, R., Torn, M., and Jahn, R. (2005). Poorly crystalline mineral phases protect organic matter in acid subsoil horizons. Eur. J. Soil Sci. 56, 717–725. doi: 10.1111/j.1365-2389.2005.00706.x
Kwon, H., Law, B. E., Thomas, C. K., and Johnson, B. G. (2018). The influence of hydrological variability on inherent water use efficiency in forests of contrasting composition, age, and precipitation regimes in the Pacific Northwest. Agric. For. Meteorol. 249, 488–500. doi: 10.1016/j.agrformet.2017.08.006
Lake, F. K., Wright, V., Morgan, P., McFadzen, M., McWethy, D., and Stevens-Rumann, C. (2017). Returning fire to the land: celebrating traditional knowledge and fire. J. Forest. 115, 343–353. doi: 10.5849/jof.2016-043R2
Law, B. E., Berner, L. T., Mildrexler, D. J., Bloemers, R. O., and Ripple, W. J. (2022). Strategic reserves in Oregon's forests for biodiversity, water, and carbon to mitigate and adapt to climate change. Front. For. Global Change 5, 1–19. doi: 10.3389/ffgc.2022.1028401
Law, B. E., Bloemers, R. O., Colleton, N., and Allen, M. (2023). Redefining the wildfire problem and scaling solutions to meet the challenge. Bull. Atomic Scient. 79, 377–384. doi: 10.1080/00963402.2023.2266941
Lehmann, J., and Kleber, M. (2015). The contentious nature of soil organic matter. Nature 528, 60–68. doi: 10.1038/nature16069
Liang, S., Hurteau, M. D., and Westerling, A. L. (2018). Large-scale restoration increases carbon stability under projected climate and wildfire regimes. Front. Ecol. Environ. 16, 207–212. doi: 10.1002/fee.1791
Liles, G. C., Maxwell, T. M., Silva, L. C. R., Zhang, J. W., and Horwath, W. R. (2019). Two decades of experimental manipulation reveal potential for enhanced biomass accumulation and water use efficiency in ponderosa pine plantations across climate gradients. J. Geophys. Res. 124, 2321–2334. doi: 10.1029/2019JG005183
Marin-Spiotta, E., Chaopricha, N. T., Plante, A. F., Diefendorf, A. F., Mueller, C. W., Grandy, A. S., et al. (2014). Long-term stabilization of deep soil carbon by fire and burial during early Holocene climate change. Nat. Geosci. 7, 428–432. doi: 10.1038/ngeo2169
Marvin, D. C., Sleeter, B. M., Cameron, D. R., Nelson, E., and Plantinga, A. J. (2023). Natural climate solutions provide robust carbon mitigation capacity under future climate change scenarios. Sci. Rep. 13, 19008 doi: 10.1038/s41598-023-43118-6
Matthewmann, R., Cotton, L. J., Martins, Z., and Sephton, M. A. (2019). Organic geochemistry of late Jurassic paleosols (Dirt Beds) of Dorset, UK. Mar. Petrol. Geol. 37, 4152. doi: 10.1016/j.marpetgeo.2012.05.009
Maxwell, T. M., and Silva, L. C. R. (2020). A state factor model for ecosystem carbon - water relations. Trends Plant Sci. 25, 652–660. doi: 10.1016/j.tplants.2020.02.007
Maxwell, T. M., Silva, L. C. R., and Horwath, W. R. (2018). Integrating effects of species composition and soil properties to predict shifts in montane forest carbon–Water Relations. Proc. Nat. Acad. Sci. 115, E4219–E4226. doi: 10.1073/pnas.1718864115
Migliavacca, M., Musavi, T., Mahecha, M. D., Nelson, J. A., Knauer, J., Baldocchi, D. D., et al. (2021). The three major axes of terrestrial ecosystem function. Nature 598, 468–472. doi: 10.1038/s41586-021-03939-9
Mildrexler, D. J., Berner, L. T., Law, B. E., Birdsey, R. A., and Moomaw, W. R. (2020). Large trees dominate carbon storage in forests east of the cascade crest in the United States Pacific Northwest. Front. For. Global Change 3, 1–15. doi: 10.3389/ffgc.2020.594274
Mildrexler, D. J., Berner, L. T., Law, B. E., Birdsey, R. A., and Moomaw, W. R. (2023). Protect large trees for climate mitigation, biodiversity, and forest resilience. Conserv. Sci. Pract. 5, e12944. doi: 10.1111/csp2.12944
Miller, K. B., McCahon, T. J., and West, R. R. (1996). Lower Permian (Wolfcampian) Paleosol-bearing cycles of the U.S. Midcontinent; evidence of climatic cyclicity. J. Sediment. Res. 66, 71–84. doi: 10.1306/D42682B6-2B26-11D7-8648000102C1865D
Mills, M., Leon, J. X., Saunders, M. I., Bell, J., Liu, Y., O'Mara, J., et al. (2016). Reconciling development and conservation under coastal squeeze from rising sea level. Conservat. Lett. 9, 361–368. doi: 10.1111/conl.12213
Mu, J. E., Antle, J. M., and Abatzoglou, J. T. (2019). Representative agricultural pathways, climate change, and agricultural land uses: an application to the Pacific Northwest of the USA. Mitig. Adapt. Strateg. Glob. Change 24, 819–837. doi: 10.1007/s11027-018-9834-8
Negandhi, K., Edwards, G., Kelleway, J. J., Howard, D., Safari, D., and Saintilan, N. (2019). Blue carbon potential of coastal wetland restoration varies with inundation and rainfall. Sci. Rep. 9, 1–9. doi: 10.1038/s41598-019-40763-8
Nelson, P. N., and Baldock, J. A. (2005). Estimating the molecular composition of a diverse range of natural organic materials from solid-state CNMR and elemental analyses. Biogeochemistry 72, 1–34. doi: 10.1007/s10533-004-0076-3
Parks, S. A., Holsinger, L. M., Panunto, M. H., Jolly, W. M., Dobrowski, S. Z., and Dilion, G. (2018). High-severity fire: evaluating its key drivers and mapping its probability across western US forests. Environ. Res. Lett. 13, 44037. doi: 10.1088/1748-9326/aab791
Parks, S. A., Miller, C., Abatzoglou, J. T., Holsinger, L. M., Parisien, M., and Dobrowski, S. Z. (2016). How will climate change affect wildland fire severity in the western US? Environ. Res. Lett. 11, 035002. doi: 10.1088/1748-9326/11/3/035002
Pastorello, G., Trotta, C., Canfora, E., Chu, H., Christianson, H., Cheah, Y., et al. (2020). The FLUXNET2015 dataset and the ONEFlux processing pipeline for eddy covariance data. Sci. Data 7, 225. doi: 10.1038/s41597-020-0534-3
Perry, T. D., and Jones, J. A. (2017). Summer streamflow deficits from regenerating Douglas-fir forest in the Pacific Northwest, USA. Ecohydrol. 10, e1790. doi: 10.1002/eco.1790
Pham, T. D., Xia, J., Ha, N. T., Bui, D. T., Le, N. N., and Tekeuchi, W. (2019). A review of remote sensing approaches for monitoring blue carbon ecosystems: mangroves, seagrasses and salt marshes during 2010–2018. Sensors 19, 1933. doi: 10.3390/s19081933
Plante, A. F., Fernández, J. M., Haddix, M. L., Steinweg, J. M., and Conant, R. T. (2011). Biological, chemical and thermal indices of soil organic matter stability in four grassland soils. Soil Biol. Biochem. 43, 1051–1058 doi: 10.1016/j.soilbio.2011.01.024
Reclamation (2013). “Downscaled CMIP3 and CMIP5 Climate and Hydrology Projections: Release of Downscaled CMIP5 Climate Projections, Comparison with preceding Information, and Summary of User Needs”, in U.S. Department of the Interior, Bureau of Reclamation, Technical Services Center (Denver, Colorado: Reclamation), 47.
Reich, P. B., Sendall, K. M., Stefanski, A., Rich, R. L., Hobbie, S. E., and Montgomery, R. A. (2018). Effects of climate warming on photosynthesis in boreal tree species depend on soil moisture. Nature 562, 263–267. doi: 10.1038/s41586-018-0582-4
Retallack, G. J. (2019). Soils of the Past: An Introduction to Paleopedology (Third Edition). Chichester: Wiley, 537.
Roering, J. J., Hunter, B. D., Ferrier, K. L., Chadwick, O. A., Yoo, K., Wackett, A., et al. (2023). Quantifying erosion rates and weathering pathways that maximize soil organic carbon storage. Biogeochemistry 164, 319–333 doi: 10.1007/s10533-023-01054-7
Ruzicka, K. J., Puettmann, K. J., and Brooks, J. R. (2017). Cross-scale interactions affect tree growth and intrinsic water use efficiency and highlight the importance of spatial context in managing forests under global change. J. Ecol. 105, 1425–1436. doi: 10.1111/1365-2745.12749
Schmidt, M. W. I., Torn, M. S., Abiven, S., Dittmar, T., Guggenberger, G., Janssens, I. A., et al. (2011). Persistence of soil organic matter as an ecosystem property. Nature 478, 49–56. doi: 10.1038/nature10386
Schultz, M. A., Janousek, C. N., Brophy, L. S., Schmitt, S. D., and Bridgham, S. (2023). How management interacts with environmental drivers to control greenhouse gas fluxes from Pacific Northwest coastal wetlands. Biogeochemistry 165, 165–190 doi: 10.1007/s10533-023-01071-6
Segura, C., Baldon, K. D., Hatten, J. A., Jones, J. A., Hale, V. C., and Ice, G. G. (2021). Long-term effects of forest harvesting on summer low flow deficits in the Coast Range of Oregon. J. Hydrol. 585:124749. doi: 10.1016/j.jhydrol.2020.124749
Short, F. T., and Wyllie-Echeverria, S. (1996). Natural and human- induced disturbance of seagrasses. Environ. Conserv. 23, 17–27. doi: 10.1017/S0376892900038212
Shukla, P.R., Skeg, J., Buendıa, E., Masson-Delmotte, V., Pörtner, H., Roberts, D. C., et al. (2019). Climate Change and Land: An IPCC Special Report on Climate Change, Desertification, Land Degradation, Sustainable Land Management, Food Security, and Greenhouse Gas Fluxes in Terrestrial Ecosystems. Available online at: https://www.ipcc.ch/site/assets/uploads/sites/4/2019/12/Climate-Change-and-Land-Errata_12-DEC_2019.pdf
Silva, L. C. R. (2022). Expanding the scope of biogeochemical research to accelerate atmospheric carbon capture. Biogeochemistry 161, 19–40 doi: 10.1007/s10533-022-00957-1
Silva, L. C. R., and Lambers, H. (2020). Soil-plant-atmosphere interactions: structure, function, and predictive scaling for climate change mitigation. Plant Soil. 461, 5–27. doi: 10.1007/s11104-020-04427-1
Silva, L. C. R., Pedroso, G., Doane, T. A., Mukome, F. N. D., and Horwath, W. R. (2015). Beyond the cellulose: oxygen isotope composition of plant lipids as a proxy for terrestrial water balance. Geochemical Perspect. Lett. 1, 33–42. doi: 10.7185/geochemlet.1504
Silva, L. C. R., Wood, M. C., Johnson, B. R., Coughlan, M., Brinton, H., McGuire, K., et al. (2022). A generalizable framework for enhanced natural climate solutions. Plant Soil 479, 3–24. doi: 10.1007/s11104-022-05472-8
Smithwick, E. A. H., Harmon, M. E., Remillard, S. M., Acker, S. A., and Franklin, J. F. (2002). Potential upper bounds of carbon stores in forests of the pacific northwest. Ecol. Appl. 12, 1303–1317. doi: 10.1890/1051-0761(2002)0121303:PUBOCS2.0.CO
Sperling, O., Silva, L. C. R., Tixier, A., Théroux-Rancourt, G., and Zwieniecki, M. A. (2017). Temperature gradients assist carbohydrate allocation within trees. Sci. Rep. 7, 3265 doi: 10.1038/s41598-017-03608-w
Stamey, M. (2004). An Analysis of Eelgrass Transplantation Performance in Puget Sound, WA, 1990–2000 (Thesis). University of Washington, Seattle, WA, United States.
Taylor, A. H., Trouet, V., Skinner, C. N., and Stephens, S. (2016). Socioecological transitions trigger fire regime shifts and modulate fire-climate interactions in the Sierra Nevada, USA, 1600-2015 CE. Proc Natl Acad Sci. U. S. A. 113, 13684–13689. doi: 10.1073/pnas.1609775113
U. S. EPA (2020). Western Ecology Division, Ecoregion Maps and GIS Resources. Available online at: http://www.epa.gov/eco-research/ecoregions.htm (accessed 31 May, 2020).
Vitousek, P., and Loope, C. (1987). Stone Introduced species in Hawaii. biological effects and opportunities for ecological research. Trends Ecol. Evol. 2, pp. 224–227. doi: 10.1016/0169-5347(87)90026-7
Waldo, S., Chi, J., Pressley, S. N., O'Keeffe, P., Pan, W. L., Brooks, E. S., et al. (2016). Assessing carbon dynamics at high and low rainfall agricultural sites in the inland Pacific Northwest US using the eddy covariance method. Agric. For. Meteorol. 218–219, 25–36. doi: 10.1016/j.agrformet.2015.11.018
Waring, R. H., and Franklin, J. F. (1979). Evergreen coniferous forests of the Pacific Northwest. Science 204, 1380–1386. doi: 10.1126/science.204.4400.1380
Watanabe, M., Suzuki, T., O'ishi, R., Komuro, Y., Watanabe, S., Emori, S., et al. (2010). Improved climate simulation by MIROC5: mean states, variability, and climate sensitivity. J. Clim. 23, 6312–6335. doi: 10.1175/2010JCLI3679.1
Keywords: natural climate solutions, nature-based solutions, soil organic carbon, climate change, carbon persistence
Citation: Chafe OE, Broz AP, Levenson ES, Farinacci MD, Anderson RO and Silva LCR (2024) The spatiotemporal domains of natural climate solutions research and strategies for implementation in the Pacific Northwest, USA. Front. Clim. 6:1273632. doi: 10.3389/fclim.2024.1273632
Received: 16 August 2023; Accepted: 22 January 2024;
Published: 14 February 2024.
Edited by:
Sirkku Juhola, University of Helsinki, FinlandReviewed by:
Charles Drever, Nature United, CanadaTara Hudiburg, University of Idaho, United States
Copyright © 2024 Chafe, Broz, Levenson, Farinacci, Anderson and Silva. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Adrian P. Broz, YWJyb3pAdW9yZWdvbi5lZHU=
†These authors have contributed equally to this work