 Qinrui Li
Qinrui Li Youyu Gu
Youyu Gu Jingjing Liang1,2
Jingjing Liang1,2 Zhixian Yang
Zhixian Yang Jiong Qin
Jiong Qin
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Neurosci., 26 June 2024
Sec. Cellular Neuropathology
Volume 18 - 2024 | https://doi.org/10.3389/fncel.2024.1386205
This article is part of the Research TopicRole of Microbiota in Neurocognitive Disorders: A Developmental Origin PerspectiveView all 9 articles
Epilepsy is a common neurological disorder that affects approximately 10.5 million children worldwide. Approximately 33% of affected patients exhibit resistance to all available antiseizure medications, but the underlying mechanisms are unknown and there is no effective treatment. Increasing evidence has shown that an abnormal gut microbiota may be associated with epilepsy. The gut microbiota can influence the function of the brain through multiple pathways, including the neuroendocrine, neuroimmune, and autonomic nervous systems. This review discusses the interactions between the central nervous system and the gastrointestinal tract (the brain–gut axis) and the role of the gut microbiota in the pathogenesis of epilepsy. However, the exact gut microbiota involved in epileptogenesis is unknown, and no consistent results have been obtained based on current research. Moreover, the target that should be further explored to identify a novel antiseizure drug is unclear. The role of the gut microbiota in epilepsy will most likely be uncovered with the development of genomics technology.
Epilepsy is a chronic disease of the central nervous system (CNS) and is characterized by recurrent and unprovoked seizures (Fisher et al., 2017). Epilepsy affects more than 50–70 million people worldwide, including 10.5 million children under 15 years of age who have active epilepsy (Thijs et al., 2019; Singh and Sander, 2020). Approximately 80% of patients with epilepsy live in low- and middle-income countries (Saxena and Li, 2017; Singh and Sander, 2020). The majority of these countries are in Africa, with Uganda being affected the most(Newton and Garcia, 2012). A systematic review reported that the prevalence of epilepsy in children and young adults is approximately two-fold greater than that in middle-aged patients (Benamer and Grosset, 2009). Approximately 33% of patients suffer from refractory epilepsy, which places a tremendous burden on global health and finances (Dalic and Cook, 2016). Therefore, additional and more effective antiseizure medications are urgently needed. Many studies have shown that the gut microbiota is associated with epilepsy (Xie et al., 2017; Peng et al., 2018, 2023; Zhang et al., 2018; Huang et al., 2019, 2022; Lindefeldt et al., 2019; Gong et al., 2020, 2022; Lee et al., 2020, 2021; Safak et al., 2020; Cui et al., 2021; Xu et al., 2021; Bertuccioli et al., 2022; Dahlin et al., 2022; Dai et al., 2022; Ouyang et al., 2022; Zhou et al., 2022, 2023; Turay et al., 2023; Wan et al., 2024); however, it is difficult to determine whether the gut microbiota is the cause of epilepsy or whether changes in the microbiota are only a result of epilepsy. Therefore, a long journey lies ahead in discovering drugs that target the gut microbiota for epilepsy treatment.
The human gut hosts approximately 100 trillion microorganisms, which are collectively referred to as the gut microbiota (Saad et al., 2016). The gut microbiota defines the genes and genomes of the microbiota, as well as the products of the microbiota and the host environment (Whiteside et al., 2015). However, the association between the microbiota and heredity has not been determined. Acquired factors, such as diet and drugs, have a considerable influence on the composition of the gut microbiota (Rothschild et al., 2018). Additionally, the gut microbiota fluctuates over the lifetime of a person and changes rapidly before 3 years of age, especially during the weaning period (Ling et al., 2022).
In the human gastrointestinal tract, the most dominant bacterial phyla are Firmicutes, Bacteroidetes, Actinobacteria, Proteobacteria, Fusobacteria, Verrucomicrobia, and Cyanobacteria; among which, Bacteroidetes and Firmicutes constitute >90% of the total bacteria (Ding et al., 2021). The gut microbiota regulates many physiological functions, including digestion (El et al., 2014), absorption (Zeng et al., 2017), excretion (Karl et al., 2017), vitamin synthesis, and immune function (Cheng et al., 2019). Gastrointestinal symptoms, such as abdominal pain or diarrhea, might be the first or only obvious symptoms in individuals with epilepsy and can sometimes even lead to incorrect diagnoses in clinical practice (Ozkara et al., 2009; Yunus et al., 2016). Additionally, patients with inflammatory bowel disease have a greater risk of suffering from epilepsy (De Caro et al., 2019). According to these clinical findings, the role of the microbiota and the gut–brain axis in epilepsy should be noted. This article focuses on the gut microbiota and reviews the relationship between the gut microbiota and epilepsy and the underlying mechanisms involved. In addition, this review describes the shortcomings and limitations of related studies, such as methodology and confounding factors, and discusses the challenges and limitations of epilepsy treatment from the perspective of the intestinal flora.
Recently, a new concept named the “brain–gut axis,” which refers to the gut microbiota, was proposed to influence the CNS (Quigley, 2017). Millions of neurons are present in the gastrointestinal mucosa. These cells constitute the enteric nervous system (ENS), regulate gastrointestinal functions, and influence the CNS mainly via the vagus nerve. Thus, the gut is considered a “second brain.” The gut microbiota acts as an important medium in the bidirectional system of communication between the brain and the gut (Quigley, 2017). This communication needs to occur through multiple systems, including the neuroendocrine, neuroimmune, and autonomic nervous systems (Martin et al., 2018). Because of the brain–gut axis, abnormal gut microbiota may disrupt the homeostasis of the CNS and may be involved in the pathogenesis of Alzheimer’s disease (Shen and Ji, 2019), Parkinson’s disease in humans and animals (Scheperjans et al., 2015; Sampson et al., 2016; Pietrucci et al., 2019; Zhang et al., 2023), stroke in humans and mice (Winek et al., 2016; Meng et al., 2023; Chen et al., 2024), autism spectrum disorder (ASD) in humans and animals (Li et al., 2017; Kong et al., 2021), and epilepsy in humans and animals (Olson et al., 2018). The oral microbiota has also been reported to be altered during epilepsy (Lian et al., 2023). Moreover, a ketogenic diet (KD) is a non-pharmacological therapy for drug-resistant epileptic patients and has good curative effects (van der Louw et al., 2016; Imdad et al., 2022). Additionally, the intestinal microbiota of infants is different from that of older children. Therefore, infants may not have generalized tonic–clonic seizures (Korff and Nordli, 2005). Here, we discuss the prototypical relationships between the gut microbiota and epilepsy through several systems.
In human studies, it is difficult to control confounding factors, such as diet, habitat, living environment, and life stress, in gut microbiota. Therefore, the results from human studies are at risk of being inconsistent. First, the α diversity of the gut microbiota in epileptic patients is not consistent. Three studies showed that the microbiota diversity of patients with epilepsy was lower than that of people without epilepsy (Xie et al., 2017; Gong et al., 2020; Cui et al., 2021); however, Peng et al. reported that the diversity of the microbiota was significantly greater in patients with drug-resistant epilepsy than in patients with drug-sensitive epilepsy and healthy controls (Peng et al., 2018). Second, the gut microbiota is altered in epileptic patients, but the results are inconsistent. Most studies have shown that epilepsy patients have an increased abundance of the phylum Firmicutes and a decreased abundance of the phylum Bacteroidetes (Xie et al., 2017; Peng et al., 2018; Lee et al., 2020). In contrast, Zhou et al. reported that Actinobacteria, Bacteroidetes, and Proteobacteria increased and Firmicutes decreased in adults with epilepsy (Zhou et al., 2023). For instance, studies on the gut microbiota in epileptic patients have yielded different results. A study showed that the abundances of the genera Campylobacter, Delftia, Haemophilus, Lautropia, and Neisseria among the Proteobacteria phylum were significantly greater in patients with epilepsy than in people without epilepsy (Safak et al., 2020). The Fusobacteria phylum could be detected in only 10.6% of patients with epilepsy (Safak et al., 2020). In addition, compared to those in the control group, the bacteria in the epilepsy group had increased Proteobacteria and Actinobacteria and decreased Bacteroidetes. Moreover, the bacteria in the epilepsy group were enriched in Faecalibacterium, Escherichia-Shigella, Subdoligranulum, and unclassified Enterobacteriaceae and decreased in Bacteroides, Megamonas, Prevotella, unclassified Lachnospiraceae, and Blautia (Cui et al., 2021). Dahlin et al. reported that the levels of Bifidobacteria increased after KD treatment and were associated with an antiepileptic response (Dahlin et al., 2022). These patients had structural, genetic, and unknown information, but the relationships between the etiology and the gut microbiota were not compared (Dahlin et al., 2022). Proteobacteria and Cronobacter predominate in refractory epileptic infants of unknown cause (Xie et al., 2017). Furthermore, patients with drug-resistant epilepsy had increased levels of the bacterial taxa Actinobacteria, Verrucomicrobia, and Nitrospirae and the genera Blautia, Bifidobacterium, Subdoligranulum, Dialister, and Anaerostipes (Gong et al., 2020). In children with intractable epilepsy, Bacteroidetes levels were lower and Actinobacteria levels were higher. The ATP-binding cassette (ABC) transporter-associated microbiota included the Enterococcus faecium group, the Bifidobacterium longum group, and the Eggerthella lenta, which are likely biomarkers of intractable epilepsy (Lee et al., 2020). However, in another study by Lee et al., the abundances of Bacteroides finegoldii and Ruminococcus_g2 increased in the drug-responsive group, and the relative abundance of Negativicutes (belonging to Firmicutes) increased in the drug-resistant group (Lee et al., 2021). After KD therapy, seizures decreased significantly, electroencephalogram improved, α-diversity and the abundance of Firmicutes decreased, and the abundance of Bacteroidetes increased. In addition, Clostridiales, Ruminococcaceae, Rikenellaceae, Lachnospiraceae, and Alistipes were likely non-responsive patient biomarkers (Zhang et al., 2018). Compared with patients who have more than four seizures per year, patients with fewer than four seizures per year had an increase in the abundance of Bifidobacteria and Lactobacillus (Peng et al., 2018). Additionally, antibiotic treatment also reduced seizure frequency in some patients (Ghanizadeh and Berk, 2015; Cheraghmakani et al., 2021). Mu et al. also showed that manipulating the gut microbiota attenuated seizures in an animal model of infantile spasm syndrome by KD (Mu et al., 2022).
In summary, all these studies indicated that the gut microbiota is associated with epilepsy and therapy, but the causal mechanism of these associations has not yet been determined. However, these studies had several limitations. First, all these studies had small sample sizes, which impacted the results and conclusions. Second, the follow-up time of the studies was short. Third, most of the patients enrolled in the studies had taken antiseizure drugs, which could also influence the gut microbiota (Gong et al., 2022; Ilhan et al., 2022). Fourth, not all these studies controlled for confounding factors, such as diet, antibiotic discontinuation time, or age, which could also influence the alteration of the gut microbiota. Fifth, these studies did not draw the same conclusion about the exact bacterial effect on epileptogenesis. However, all these studies demonstrated a relationship between the gut microbiota and epilepsy. The human gut also contains fungi, viruses, parasites, and archaea; however, there were no studies that investigated their role in epilepsy. Therefore, future investigations are needed to better understand the role of these microbiota in epilepsy.
With the development of methods for identifying the microbiota, we can obtain more information about the gut microbiota and its metabolites. Next-generation sequencing (NGS) is the most used method for identifying the microbiota in humans. The key NGS methods include 16S ribosomal RNA (16S rRNA) and shotgun metagenomic sequencing. 16S rRNA sequencing is the most used method for bacterial identification. It is targeted for polymerase chain reaction amplification of the bacterial 16S ribosomal RNA (rRNA) gene and cannot detect fungi, parasites, or viruses (Durazzi et al., 2021). Moreover, 16S rRNA has a lower risk of host contamination (Liu et al., 2021). However, 16S rRNA sequencing has less sensitivity and cannot detect strain-level changes. In addition, 16S rRNA sequencing cannot provide a direct functional profile because the method only characterizes sequences from one essential gene. Shotgun metagenomics is more expensive than 16S rRNA sequencing but offers far broader species- and strain-level resolution, more accurate functional profiling, and possibly the identification of unknown species and strains of microbes (Jovel et al., 2016).
Sequencing platforms have also been developed that are classified by maximum output, reads per run, accuracy, run time, number of nucleic acids, and read length (Illumina, Ion Torrent, Sequencing by Oligonucleotide Ligation and Detection (SOLiD), PacBio, and Nanopore). Illumina sequencing technology is a method that allows for the simultaneous sequencing of millions of fragments and is a significant advancement over DNA sequencing due to its speed and cost (Bentley et al., 2008). Ion Torrent sequencing technology uses semiconductor chips to detect hydrogen ions released during DNA polymerization but is not widely used for detecting gut microbiota (Guerra et al., 2018). The SOLiD system has high-throughput sequencing capabilities and accuracy, particularly in the context of identifying single-nucleotide polymorphisms (SNPs) and other genetic variations, and can generate large amounts of data and detect both sequence and structural variations in the genome (McKernan et al., 2009). PacBio and nanopores can enable the sequencing of much longer DNA fragments than short-read sequencing platforms such as Illumina. These sequencing platforms improved the resolution of complex genomic regions, structural variations, and repetitive sequences. Nanopores can test nucleic acids, including both native DNA and RNA molecules, and have the advantages of rapid readout, high accuracy, low cost, and portability (Dorey and Howorka, 2024). It has been reported that nanopore sequences are promising for polypeptide identification and sequencing, capturing folded proteins, and achieving peptide threading (Dorey and Howorka, 2024). With the progress of sequencing technologies, we will learn more about the gut microbiota and epilepsy.
Currently, an increasing number of studies have shown that antiseizure medications probably influence the gut microbiota, which will confound our understanding of the relationship between the gut microbiota and epilepsy. Gong et al. reported that valproate could increase the ratio of the phyla Firmicutes to Bacteriodetes in children with focal epilepsy (Gong et al., 2022). Ilhan et al. reported that antiseizure medications (carbamazepine, lamotrigine, and topiramate) reduced the growth of more than 10 bacterial strains, and antiseizure medications with syrup excipient and artificial sweeteners reduced or stimulated microbiota growth in HT-29 cells, but they did not report the effects of the patients’ symptoms (Ilhan et al., 2022). In addition, the gut microbiota can also affect the efficacy of drugs. Akkermansia muciniphila likely reduces the expression of drug-resistance genes in HT-29 cells, and Bifidobacterium longum may reduce the cytotoxic effects of carbamazepine and lamotrigine in cells (Ilhan et al., 2022). Thai et al. reported that topiramate significantly increased Lactobacillus johnsonii in naïve mice (Ilhan et al., 2022). Valproate prevents peritoneal adhesion following abdominal injury through chymase inhibition and decreases intestinal inflammation in individuals with IBS (Liu et al., 2020; Felice et al., 2021). Phenobarbital treats postnatal hyperbilirubinemia through its effects on the hepatic enzymatic elimination of bilirubin (Chawla and Parmar, 2010). Wan et al. also reported that non-responsive children with infantile epileptic spasm syndrome had upregulated Clostridioides and Peptoclostridium_phage_p630P2 and downregulated Lachnospiraceae and Phascolarctobacterium after adrenocorticotropic hormone (ACTH) treatment (Fairlie et al., 2020). In summary, these antiseizure medications were only found to affect the gut microbiota. It is important to note that no research has yet explored the potential correlation between the efficacy of antiseizure medications and alterations in gut microbiota. Additional studies have confirmed that alterations in the gut microbiota can be influenced by antiseizure medications and their ingredients.
The gut-brain signaling pathways primarily rely on vagal receptors located in the gastrointestinal mucosa, which are capable of detecting changes in the gut microbiota and associated metabolites and transmitting these signals to the CNS (de Lartigue et al., 2011). Various factors, such as gut peptides, inflammatory molecules, and food or drug components, can influence the composition of gut microbiota and its metabolites (Sen et al., 2017). Metabolomics has revealed that microbiota-mediated metabolites, including short-chain fatty acids (SCFAs), phenolic compounds, and free amino acids, are implicated in epilepsy (Brandsma et al., 2019; Boeri et al., 2022). Dietary fibers are fermented and subsequently produce SCFAs by some gut microbiota, including Akkermansia municiphilla, Ruminococcus bromii, and Faecalibacterium prausnitzii (Morrison and Preston, 2016). Circulating SCFAs can cross the blood–brain barrier (BBB) as signaling molecules within the gut–brain axis. They directly or indirectly provide energy to neurons while maintaining glucose and energy homeostasis and regulate neurotransmitters’ function (Hu et al., 2018). Notably, drug-resistant epilepsy often coincides with energy deficiency (Araujo et al., 2014), suggesting that SCFAs might play an important role in the occurrence and progression of epilepsy. Importantly, a KD, which is a treatment for drug-resistant epilepsy, can reduce seizure frequency, probably through the production of SCFAs (Gong et al., 2021).
Phenolic compounds produced by the gut microbiota principally consist of serotonin, dopamine, γ-aminobutyric acid (GABA), and norepinephrine (NE) (Maia et al., 2017; Uchida et al., 2017). Furthermore, elevated endogenous noradrenergic transmission causes an increase in NE release, which is involved in some cases of epilepsy (Fitzgerald, 2010).
The relationship between free amino acids (FAAs) and the gut microbiota is also bidirectional (Kumar et al., 2018). Through metabolomic sequencing, Zhou et al. discovered that tryptophan and kynurenine levels were decreased in adults with epilepsy, suggesting their potential as biomarkers for epilepsy. Additionally, they observed positive associations between Providencia, Candidatus Marithrix, Ulvibacter, Methanospirillum, and Gaetbulibacter and serum tryptophan levels. Furthermore, Providencia, Mesotoga, and Desulfurispora exhibited positive correlations with serum kynurenine (Zhou et al., 2023). It has been reported that tryptophan can be metabolized to kynurenine, which regulates neuroendocrine and intestinal immune effects (Gao et al., 2020). Moreover, high concentrations of kynurenine have been shown to possess neuroprotective effects and regulate glutamate function (Rho, 2004). Few studies have explored the relationship between epilepsy and FAAs. This could be a future research direction.
The gut communicates with the brain through immunological pathways. The gut microbiota can influence the development and function of microglia, which are crucial resident immune cells in the CNS involved in CNS disorders (Maneu et al., 2016). Microglia activation occurs during epileptogenesis in both rat and human brains affected by epilepsy (Yankam et al., 2017). Probiotics reduce microglia activation in the hippocampus via the vagus nerve through the microbiota–gut brain axis (Liu et al., 2021). In the mouse, in the case of hepatitis virus-induced nerve injury, the gut microbiota can enhance the ability of microglia to respond to infection and prevent neurological diseases (Brown et al., 2019). B. longum can mediate tryptophan metabolism to ameliorate microglial activity in the cerebellum (Brown et al., 2019). Microglia are activated by microbial metabolites, including cytokines and hormones, and inhibited by SCFAs and tryptophan derivatives (Zheng et al., 2023). Braniste et al. reported that abnormal gut microbiota in mice is directly associated with reduced expression of tight junction proteins, including occludin and claudin-5, leading to an increase in BBB permeability (Braniste et al., 2014). During systemic inflammation, increases in the levels of inflammatory cytokines, such as interleukin-1β (IL-1β), interleukin-6 (IL-6), and tumor necrosis factor-α (TNF-α), can accelerate the disruption of the BBB (Geng et al., 2018; Kim et al., 2022). In patients with temporal lobe epilepsy, chronic inflammation is very common in brain parenchyma (Gales and Prayson, 2017) and may lead to mitochondrial dysfunction (Volmering et al., 2016).
To date, there are no completely effective or safe therapies for refractory epilepsy. However, a systematic review reported that manipulating the gut microbiota through the use of a fecal microbiota transplant (FMT), antibiotic treatment, a ketogenic diet, or prebiotic treatment could improve dysbiosis and ameliorate epilepsy-induced brain injury to some extent (Arulsamy et al., 2020). Thus, in this section, the important potential treatments for epilepsy are summarized.
A KD contains a high fat content, low carbohydrate content, appropriate protein levels, and other nutrients (Rekdal and Balskus, 2018). Many recent studies have shown that a KD has a positive effect on many disorders, such as Alzheimer’s disease (Lim et al., 2022), malignant tumors (Cohen et al., 2018; Ok et al., 2018; Iyikesici, 2019), obesity (Moreno et al., 2016; Yuan et al., 2022), diabetes (Saslow et al., 2017), and epilepsy (Cohen et al., 2018; Fan et al., 2019; Iyikesici, 2019). Recently, several studies have shown that the abundances of Akkermansia and Parabacteroides increase after KD treatment (Newell et al., 2016; Xie et al., 2017; Olson et al., 2018; Lindefeldt et al., 2019). Previous studies have shown that the beneficial effects of Akkermansia muciniphila are associated with SCFAs following KD consumption (Paoli et al., 2019; Attaye et al., 2021). Additionally, the abundance of Bifidobacteria decreased after KD treatment, and these bacteria displayed antiseptic effects (Lindefeldt et al., 2019; Dahlin et al., 2022). Some studies have shown that α-diversity decreases, the abundance of the phylum Bacteroidetes increases, and the abundances of the phyla Proteobacteria and Firmicutes decrease in children with refractory epilepsy after KD treatment (Xie et al., 2017; Zhang et al., 2018). Thus, scientists speculate that the antiepileptic effect of KD therapy might depend on the gut microbiota. As shown by Olson et al., germ-free mice with epilepsy are resistant to KD treatment, while epileptic mice treated with Akkermansia and Parabacteroides exhibit a satisfactory response (Olson et al., 2018). This study confirmed that the gut microbiota is an essential link for the antiepileptic effect of a KD. In summary, a KD is a traditional but effective therapy for epilepsy and most likely works through alteration of the gut microbiota.
Probiotics are beneficial to host health when they are supplied appropriately. These bacteria mainly consist of Lactobacillus, Bifidobacterium, Lactococcus, Saccharomycetes, and several enzymes. Many researchers have reported that probiotics can prevent or treat many disorders, such as allergies and infectious diseases (Ishaque et al., 2018; Trick et al., 2018). Because of the relationship between the gut microbiota and the brain, probiotics are used to prevent and treat CNS-related diseases. For example, treatment with L. plantarum MTCC1325 can significantly improve cognition, ameliorate brain acetylcholine levels, and resolve histopathological lesions in albino rats with D-galactose-induced Alzheimer’s disease (Nimgampalle and Kuna, 2017). A recent observational study showed that, compared with healthy controls, newborns with rotaviral infection are more likely to suffer from neonatal seizures and that treatment with probiotics immediately after birth can reduce this risk (Yeom et al., 2019). Additionally, the use of probiotics as a supplementary treatment can obviously improve seizure control and quality of life in patients with drug-resistant epilepsy (Gomez-Eguilaz et al., 2018). Oral probiotic treatment dramatically reduces seizure severity and improves spatial learning and memory in Pentylenetetrazol-induced kindling in rats (Bagheri et al., 2019). These results indicate that probiotic therapy is promising for the prevention and treatment of epilepsy. Prebiotics are non-digestible food ingredients that selectively stimulate the growth and activity of one or a few gut microbes, such as Bifidobacteria and Lactobacillus, and improve the intestinal microenvironment (Chen and Quigley, 2014). A recent systematic review and meta-analysis of randomized controlled trials (RCTs) confirmed that prebiotics can increase the abundance of Bifidobacteria in patients with IBS or other functional bowel disorders (Wilson et al., 2019). Prebiotics are also effective for treating several diseases of the CNS. For children with ASD, prebiotic intervention can significantly improve psychological traits (Grimaldi et al., 2018). An RCT by Barichella et al. showed that fermented milk containing multiple probiotics and prebiotic fiber can improve constipation in patients with Parkinson’s disease (Barichella et al., 2016). However, although the gut microbiota is closely related to epilepsy, no studies have focused on the effect of prebiotics on this disease. This might be a research direction in the future.
Fecal microbiota transplant (FMT) is a therapeutic approach aimed at rebuilding the gut microbiota. By transferring an intestinal microbiota obtained from the feces of a healthy donor into the patient’s gastrointestinal tract, FMT has proven to be an efficacious treatment for recurrent Clostridium difficile infection (Kelly et al., 2016; Hvas et al., 2019). FMT is also used for the treatment of inflammatory bowel disease or irritable bowel syndrome and can ameliorate the depression and anxiety symptoms induced by IBS (Kurokawa et al., 2018). Only two studies have evaluated FMT for the treatment of epilepsy. He et al. studied a 22-year-old girl with Crohn’s disease who had a history of epilepsy for 17 years and who received FMT treatment. Following the FMT, she was seizure-free without antiseizure medication for 20 months and experienced remission of intestinal symptoms (He et al., 2017), indicating that FMT has good prospects for epilepsy treatment. However, the safety of FMT needs to be further explored. DeFilipp et al. reported that two patients suffered from severe extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli bacteremia after FMT, and one patient died from this bacteremia (DeFilipp et al., 2019). In both cases, the bacteremia was caused by ESBL-producing Escherichia coli from the donor feces, and it was subsequently transmitted through FMT. Identifying healthy and eligible donors is essential (Kassam et al., 2019).
We can easily identify gut microbes and mechanisms in animal models; however, it is difficult to translate these findings to humans. In humans, observational studies have shown differences in the gut microbiota between patients and healthy people, but causal relationships have not been assessed. Furthermore, the findings regarding gut microbiota in various studies have demonstrated inconsistencies due to variations in subjects’ ages, races, and dietary habits. In addition, Dahlin et al. found that Bifidobacteria decreased after KD treatment; however, Zhang et al. reported that Firmicutes and Actinobacteria both decreased after KD therapy and Bacteroidetes increased after KD therapy (Zhang et al., 2018; Dahlin et al., 2022). The alterations in the gut microbiota in different studies are displayed in Table 1. The composition of the gut microbiota was assessed using next-generation sequencing of 16S ribosomal RNA genes or whole-genome shotgun sequencing based on DNA-associated methods, which also allows the inference of microbiota functions (Ranjan et al., 2016; Vogtmann et al., 2016). Recently, metabonomics has been used to identify metabolic products of the gut microbiota in stool and serum, which provides hope for the future study of the gut microbiota (Zhao et al., 2017).
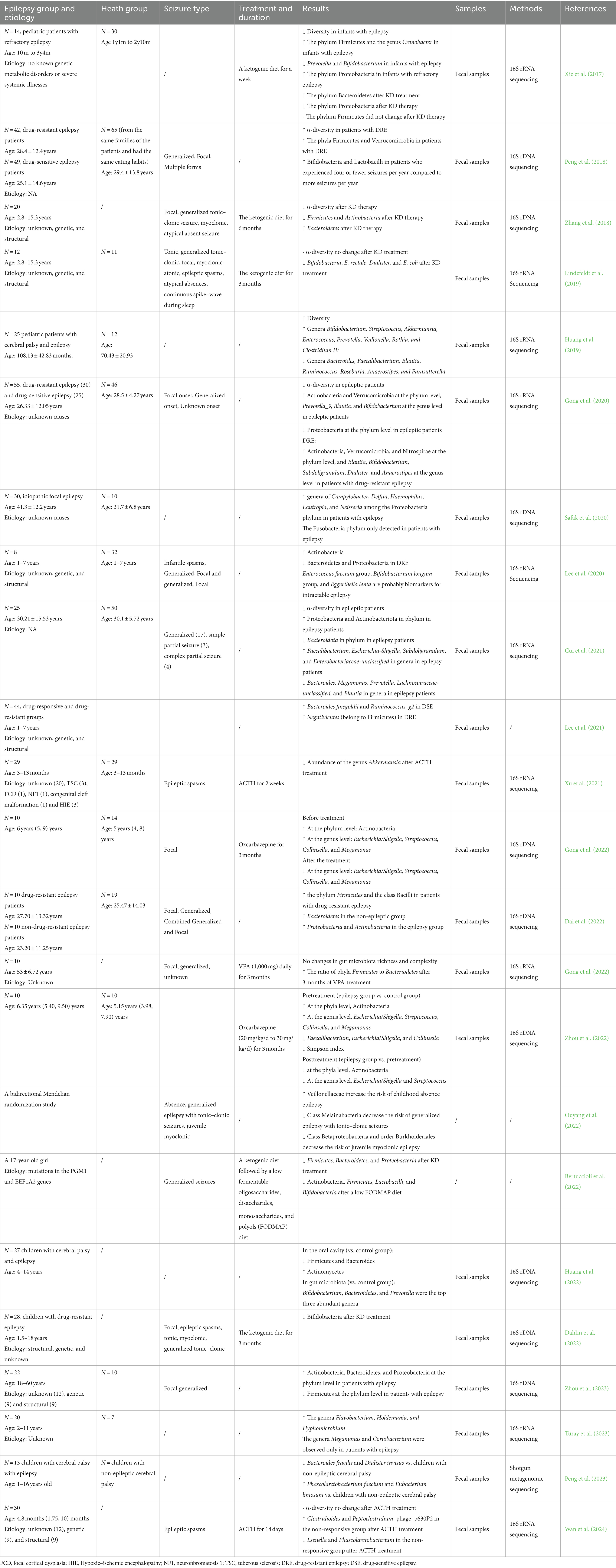
Table 1. Focus on the gut microbiota and epilepsy in human studies.
In this review, we present a summary of human studies investigating the relationship between gut microbiota and epilepsy and discuss several potential therapeutic approaches for modulating gut microbiota in individuals with epilepsy, including KD, FMT, and probiotics or prebiotics interventions. The relationship between gut microbiota and epilepsy is probably related to heritability. Zhernakova et al. reported that host genetics regulate the genetic diversity of gut microbiota (Zhernakova et al., 2024). Furthermore, a bidirectional Mendelian randomization study demonstrated that the composition of the gut microbiota is associated with seizure type; for example, the family Veillonellaceae is associated with a greater risk of childhood absence epilepsy, the class Melainabacteria is associated with a lower risk of generalized epilepsy with tonic–clonic seizures, and the class Betaproteobacteria and the order Burkholderiales are associated with a lower risk of juvenile myoclonic epilepsy. These findings indicated that genetic factors or seizure types are likely involved in the relationship between the gut microbiota and epilepsy. Therefore, the alteration of the gut microbiota is probably the result of the individual genetic background and environment. Future studies should focus on the etiology of epilepsy and identifying genetic mutations associated with gut microbiota in order to expand our understanding of these gene functions. With advancements in genomics technology, elucidating both associations and causal relationships will undoubtedly unravel the intricate interplay between gut microbiota and epilepsy while simultaneously identifying novel drug targets from a microbial perspective.
QL: Conceptualization, Writing – original draft. YG: Writing – original draft. JL: Writing – review & editing. ZY: Writing – review & editing. JQ: Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This study was supported by the National Natural Science Foundation of China, Grant no.: 82101516; the 2018 Beijing Clinical Key Specialty Construction Project-Pediatrics Foundation (2199000726); and the Peking University People’s Hospital R&D Fund (RDJP2022-22).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Araujo, B., Torres, L., Stein, M., Cabral, F. R., Herai, R., Okamoto, O., et al. (2014). Decreased expression of proteins involved in energy metabolism in the hippocampal granular layer of rats submitted to the pilocarpine epilepsy model. Neurosci. Lett. 561, 46–51. doi: 10.1016/j.neulet.2013.12.040
Arulsamy, A., Tan, Q. Y., Balasubramaniam, V., O'Brien, T. J., and Shaikh, M. F. (2020). Gut microbiota and epilepsy: a systematic review on their relationship and possible therapeutics. ACS Chem. Neurosci. 11, 3488–3498. doi: 10.1021/acschemneuro.0c00431
Attaye, I., van Oppenraaij, S., Warmbrunn, M. V., and Nieuwdorp, M. (2021). The role of the gut microbiota on the beneficial effects of ketogenic diets. Nutrients 14:191. doi: 10.3390/nu14010191
Bagheri, S., Heydari, A., Alinaghipour, A., and Salami, M. (2019). Effect of probiotic supplementation on seizure activity and cognitive performance in ptz-induced chemical kindling. Epilepsy Behav. 95, 43–50. doi: 10.1016/j.yebeh.2019.03.038
Barichella, M., Pacchetti, C., Bolliri, C., Cassani, E., Iorio, L., Pusani, C., et al. (2016). Probiotics and prebiotic fiber for constipation associated with parkinson disease: an rct. Neurology 87, 1274–1280. doi: 10.1212/WNL.0000000000003127
Benamer, H. T., and Grosset, D. G. (2009). A systematic review of the epidemiology of epilepsy in Arab countries. Epilepsia 50, 2301–2304. doi: 10.1111/j.1528-1167.2009.02058.x
Bentley, D. R., Balasubramanian, S., Swerdlow, H. P., Smith, G. P., Milton, J., Brown, C. G., et al. (2008). Accurate whole human genome sequencing using reversible terminator chemistry. Nature 456, 53–59. doi: 10.1038/nature07517
Bertuccioli, A., Cardinali, M., Di Pierro, F., Zonzini, G. B., and Matera, M. R. (2022). Ketogenic and low fodmap diet in therapeutic management of a young autistic patient with epilepsy and dysmetabolism poorly responsive to therapies: clinical response and effects of intestinal microbiota. Int. J. Mol. Sci. 23:8829. doi: 10.3390/ijms23158829
Boeri, L., Donnaloja, F., Campanile, M., Sardelli, L., Tunesi, M., Fusco, F., et al. (2022). Using integrated meta-omics to appreciate the role of the gut microbiota in epilepsy. Neurobiol. Dis. 164:105614. doi: 10.1016/j.nbd.2022.105614
Brandsma, E., Kloosterhuis, N. J., Koster, M., Dekker, D. C., Gijbels, M., van der Velden, S., et al. (2019). A proinflammatory gut microbiota increases systemic inflammation and accelerates atherosclerosis. Circ.Res. 124, 94–100. doi: 10.1161/CIRCRESAHA.118.313234
Braniste, V., Al-Asmakh, M., Kowal, C., Anuar, F., Abbaspour, A., Toth, M., et al. (2014). The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 6:263ra158. doi: 10.1126/scitranslmed.3009759
Brown, D. G., Soto, R., Yandamuri, S., Stone, C., Dickey, L., Gomes-Neto, J. C., et al. (2019). The microbiota protects from viral-induced neurologic damage through microglia-intrinsic TLR signaling. eLife 8:8. doi: 10.7554/eLife.47117
Chawla, D., and Parmar, V. (2010). Phenobarbitone for prevention and treatment of unconjugated hyperbilirubinemia in preterm neonates: a systematic review and meta-analysis. Indian Pediatr. 47, 401–407. doi: 10.1007/s13312-010-0075-5
Chen, J., Gao, X., Liang, J., Wu, Q., Shen, L., Zheng, Y., et al. (2024). Association between gut microbiota dysbiosis and poor functional outcomes in acute ischemic stroke patients with covid-19 infection. mSystems :e0018524. doi: 10.1128/msystems.00185-24
Chen, W. C., and Quigley, E. M. (2014). Probiotics, prebiotics & synbiotics in small intestinal bacterial overgrowth: opening up a new therapeutic horizon! Indian J. Med. Res. 140, 582–584
Cheng, H. Y., Ning, M. X., Chen, D. K., and Ma, W. T. (2019). Interactions between the gut microbiota and the host innate immune response against pathogens. Front. Immunol. 10:607. doi: 10.3389/fimmu.2019.00607
Cheraghmakani, H., Rezai, M. S., Valadan, R., Rahimzadeh, G., Moradi, M., Jahanfekr, V., et al. (2021). Ciprofloxacin for treatment of drug-resistant epilepsy. Epilepsy Res. 176:106742. doi: 10.1016/j.eplepsyres.2021.106742
Cohen, C. W., Fontaine, K. R., Arend, R. C., Soleymani, T., and Gower, B. A. (2018). Favorable effects of a ketogenic diet on physical function, perceived energy, and food cravings in women with ovarian or endometrial cancer: a randomized, controlled trial. Nutrients 10:1187. doi: 10.3390/nu10091187
Cui, G., Liu, S., Liu, Z., Chen, Y., Wu, T., Lou, J., et al. (2021). Gut microbiome distinguishes patients with epilepsy from healthy individuals. Front. Microbiol. 12:696632. doi: 10.3389/fmicb.2021.696632
Dahlin, M., Singleton, S. S., David, J. A., Basuchoudhary, A., Wickstrom, R., Mazumder, R., et al. (2022). Higher levels of bifidobacteria and tumor necrosis factor in children with drug-resistant epilepsy are associated with anti-seizure response to the ketogenic diet. EBioMedicine 80:104061. doi: 10.1016/j.ebiom.2022.104061
Dai, Y., Wang, M., Zhong, D., and Xu, X. (2022). Bacillus subtilis plays a role in the inhibition of transporter abcb1 in caco-2 cells. Epilepsy Res. 183:106925. doi: 10.1016/j.eplepsyres.2022.106925
Dalic, L., and Cook, M. J. (2016). Managing drug-resistant epilepsy: challenges and solutions. Neuropsychiatr. Dis. Treat. 12, 2605–2616. doi: 10.2147/NDT.S84852
De Caro, C., Leo, A., Nesci, V., Ghelardini, C., di Cesare, M. L., Striano, P., et al. (2019). Intestinal inflammation increases convulsant activity and reduces antiepileptic drug efficacy in a mouse model of epilepsy. Sci. Rep. 9:13983. doi: 10.1038/s41598-019-50542-0
de Lartigue, G., de La Serre, C. B., and Raybould, H. E. (2011). Vagal afferent neurons in high fat diet-induced obesity; intestinal microflora, gut inflammation and cholecystokinin. Physiol. Behav. 105, 100–105. doi: 10.1016/j.physbeh.2011.02.040
Defilipp, Z., Bloom, P. P., Torres, S. M., Mansour, M. K., Sater, M., Huntley, M. H., et al. (2019). Drug-resistant e. coli bacteremia transmitted by fecal microbiota transplant. N. Engl. J. Med. 381, 2043–2050. doi: 10.1056/NEJMoa1910437
Ding, M., Lang, Y., Shu, H., Shao, J., and Cui, L. (2021). Microbiota-gut-brain axis and epilepsy: a review on mechanisms and potential therapeutics. Front. Immunol. 12:742449. doi: 10.3389/fimmu.2021.742449
Dorey, A., and Howorka, S. (2024). Nanopore dna sequencing technologies and their applications towards single-molecule proteomics. Nat. Chem. 16, 314–334. doi: 10.1038/s41557-023-01322-x
Durazzi, F., Sala, C., Castellani, G., Manfreda, G., Remondini, D., and De Cesare, A. (2021). Comparison between 16s rrna and shotgun sequencing data for the taxonomic characterization of the gut microbiota. Sci. Rep. 11:3030. doi: 10.1038/s41598-021-82726-y
El, K. A., Armougom, F., Raoult, D., and Henrissat, B. (2014). Gut microbiota and digestion of polysaccharides. M S-Med. Sci. 30, 259–265. doi: 10.1051/medsci/20143003013
Fairlie, W. D., Tran, S., and Lee, E. F. (2020). Crosstalk between apoptosis and autophagy signaling pathways. Int. Rev. Cell Mol. Biol. 352, 115–158. doi: 10.1016/bs.ircmb.2020.01.003
Fan, Y., Wang, H., Liu, X., Zhang, J., and Liu, G. (2019). Crosstalk between the ketogenic diet and epilepsy: from the perspective of gut microbiota. Mediat. Inflamm. 2019:8373060. doi: 10.1155/2019/8373060
Felice, C., Lewis, A., Iqbal, S., Gordon, H., Rigoni, A., Colombo, M. P., et al. (2021). Intestinal inflammation is linked to hypoacetylation of histone 3 lysine 27 and can be reversed by valproic acid treatment in inflammatory bowel disease patients. Cell. Mol. Gastroenterol. Hepatol. 11, 889–891.e6. doi: 10.1016/j.jcmgh.2020.11.009
Fisher, R. S., Cross, J. H., French, J. A., Higurashi, N., Hirsch, E., Jansen, F. E., et al. (2017). Operational classification of seizure types by the international league against epilepsy: position paper of the ilae commission for classification and terminology. Epilepsia 58, 522–530. doi: 10.1111/epi.13670
Fitzgerald, P. J. (2010). Is elevated norepinephrine an etiological factor in some cases of epilepsy? Seizure 19, 311–318. doi: 10.1016/j.seizure.2010.04.011
Gales, J. M., and Prayson, R. A. (2017). Chronic inflammation in refractory hippocampal sclerosis-related temporal lobe epilepsy. Ann. Diagn. Pathol. 30, 12–16. doi: 10.1016/j.anndiagpath.2017.05.009
Gao, K., Mu, C. L., Farzi, A., and Zhu, W. Y. (2020). Tryptophan metabolism: a link between the gut microbiota and brain. Adv. Nutr. 11, 709–723. doi: 10.1093/advances/nmz127
Geng, J., Wang, L., Zhang, L., Qin, C., Song, Y., Ma, Y., et al. (2018). Blood-brain barrier disruption induced cognitive impairment is associated with increase of inflammatory cytokine. Front. Aging Neurosci. 10:129. doi: 10.3389/fnagi.2018.00129
Ghanizadeh, A., and Berk, M. (2015). Beta-lactam antibiotics as a possible novel therapy for managing epilepsy and autism, a case report and review of literature. Iran. J. Child Neurol. 9, 99–102
Gomez-Eguilaz, M., Ramon-Trapero, J. L., Perez-Martinez, L., and Blanco, J. R. (2018). The beneficial effect of probiotics as a supplementary treatment in drug-resistant epilepsy: a pilot study. Benef. Microbes 9, 875–881. doi: 10.3920/BM2018.0018
Gong, X., Cai, Q., Liu, X., An, D., Zhou, D., Luo, R., et al. (2021). Gut flora and metabolism are altered in epilepsy and partially restored after ketogenic diets. Microb. Pathog. 155:104899. doi: 10.1016/j.micpath.2021.104899
Gong, X., Liu, X., Chen, C., Lin, J., Li, A., Guo, K., et al. (2020). Alteration of gut microbiota in patients with epilepsy and the potential index as a biomarker. Front. Microbiol. 11:517797. doi: 10.3389/fmicb.2020.517797
Gong, X., Liu, Y., Liu, X., Li, A. Q., Guo, K. D., Zhou, D., et al. (2022). Analysis of gut microbiota in patients with epilepsy treated with valproate: results from a three months observational prospective cohort study. Microb. Pathog. 162:105340. doi: 10.1016/j.micpath.2021.105340
Gong, S. Z., Qiu, J., Wu, L. W., and Tan, L. H. (2022). Change in intestinal flora after treatment in children with focal epilepsy. Zhongguo Dang Dai Er Ke Za Zhi 24, 290–296. doi: 10.7499/j.issn.1008-8830.2109134
Gonzalez-Bosch, C., Boorman, E., Zunszain, P. A., and Mann, G. E. (2021). Short-chain fatty acids as modulators of redox signaling in health and disease. Redox Biol. 47:102165. doi: 10.1016/j.redox.2021.102165
Grimaldi, R., Gibson, G. R., Vulevic, J., Giallourou, N., Castro-Mejia, J. L., Hansen, L. H., et al. (2018). A prebiotic intervention study in children with autism spectrum disorders (asds). Microbiome 6:133. doi: 10.1186/s40168-018-0523-3
Guerra, S. G., Chong, W., Brown, C. J., and Navarrete, C. V. (2018). Evaluation of ion torrent sequencing technology for rapid clinical human leucocyte antigen typing. Int. J. Immunogenet. 45, 230–235. doi: 10.1111/iji.12378
He, Z., Cui, B. T., Zhang, T., Li, P., Long, C. Y., Ji, G. Z., et al. (2017). Fecal microbiota transplantation cured epilepsy in a case with crohn's disease: the first report. World J. Gastroenterol. 23, 3565–3568. doi: 10.3748/wjg.v23.i19.3565
Hu, J., Lin, S., Zheng, B., and Cheung, P. (2018). Short-chain fatty acids in control of energy metabolism. Crit. Rev. Food Sci. Nutr. 58, 1243–1249. doi: 10.1080/10408398.2016.1245650
Huang, C., Chu, C., Peng, Y., Zhang, N., Yang, Z., You, J., et al. (2022). Correlations between gastrointestinal and oral microbiota in children with cerebral palsy and epilepsy. Front. Pediatr. 10:988601. doi: 10.3389/fped.2022.988601
Huang, C., Li, Y., Feng, X., Li, D., Li, X., Ouyang, Q., et al. (2019). Distinct gut microbiota composition and functional category in children with cerebral palsy and epilepsy. Front. Pediatr. 7:394. doi: 10.3389/fped.2019.00394
Hvas, C. L., Dahl, J. S., Jorgensen, S. P., Storgaard, M., Lemming, L., Hansen, M. M., et al. (2019). Fecal microbiota transplantation is superior to fidaxomicin for treatment of recurrent clostridium difficile infection. Gastroenterology 156, 1324–1332.e3. doi: 10.1053/j.gastro.2018.12.019
Ilhan, Z. E., Brochard, V., Lapaque, N., Auvin, S., and Lepage, P. (2022). Exposure to anti-seizure medications impact growth of gut bacterial species and subsequent host response. Neurobiol. Dis. 167:105664. doi: 10.1016/j.nbd.2022.105664
Imdad, K., Abualait, T., Kanwal, A., Alghannam, Z. T., Bashir, S., Farrukh, A., et al. (2022). The metabolic role of ketogenic diets in treating epilepsy. Nutrients 14:5074. doi: 10.3390/nu14235074
Ishaque, S. M., Khosruzzaman, S. M., Ahmed, D. S., and Sah, M. P. (2018). A randomized placebo-controlled clinical trial of a multi-strain probiotic formulation (bio-kult(r)) in the management of diarrhea-predominant irritable bowel syndrome. BMC Gastroenterol. 18:71. doi: 10.1186/s12876-018-0788-9
Iyikesici, M. S. (2019). Feasibility study of metabolically supported chemotherapy with weekly carboplatin/paclitaxel combined with ketogenic diet, hyperthermia and hyperbaric oxygen therapy in metastatic non-small cell lung cancer. Int. J. Hyperth. 36, 445–454. doi: 10.1080/02656736.2019.1589584
Jovel, J., Patterson, J., Wang, W., Hotte, N., O'Keefe, S., Mitchel, T., et al. (2016). Characterization of the gut microbiome using 16s or shotgun metagenomics. Front. Microbiol. 7:459. doi: 10.3389/fmicb.2016.00459
Karl, J. P., Meydani, M., Barnett, J. B., Vanegas, S. M., Barger, K., Fu, X., et al. (2017). Fecal concentrations of bacterially derived vitamin k forms are associated with gut microbiota composition but not plasma or fecal cytokine concentrations in healthy adults. Am. J. Clin. Nutr. 106, 1052–1061. doi: 10.3945/ajcn.117.155424
Kassam, Z., Dubois, N., Ramakrishna, B., Ling, K., Qazi, T., Smith, M., et al. (2019). Donor screening for fecal microbiota transplantation. N. Engl. J. Med. 381, 2070–2072. doi: 10.1056/NEJMc1913670
Kelly, C. R., Khoruts, A., Staley, C., Sadowsky, M. J., Abd, M., Alani, M., et al. (2016). Effect of fecal microbiota transplantation on recurrence in multiply recurrent clostridium difficile infection: a randomized trial. Ann. Intern. Med. 165, 609–616. doi: 10.7326/M16-0271
Kim, S., Park, S., Choi, T. G., and Kim, S. S. (2022). Role of short chain fatty acids in epilepsy and potential benefits of probiotics and prebiotics: targeting "health" of epileptic patients. Nutrients 14:2982. doi: 10.3390/nu14142982
Kong, X. J., Liu, J., Liu, K., Koh, M., Sherman, H., Liu, S., et al. (2021). Probiotic and oxytocin combination therapy in patients with autism spectrum disorder: a randomized, double-blinded, placebo-controlled pilot trial. Nutrients 13:1552. doi: 10.3390/nu13051552
Korff, C., and Nordli, D. J. (2005). Do generalized tonic-clonic seizures in infancy exist? Neurology 65, 1750–1753. doi: 10.1212/01.wnl.0000187125.87414.f3
Kumar, M., Ji, B., Babaei, P., Das, P., Lappa, D., Ramakrishnan, G., et al. (2018). Gut microbiota dysbiosis is associated with malnutrition and reduced plasma amino acid levels: lessons from genome-scale metabolic modeling. Metab. Eng. 49, 128–142. doi: 10.1016/j.ymben.2018.07.018
Kurokawa, S., Kishimoto, T., Mizuno, S., Masaoka, T., Naganuma, M., Liang, K. C., et al. (2018). The effect of fecal microbiota transplantation on psychiatric symptoms among patients with irritable bowel syndrome, functional diarrhea and functional constipation: an open-label observational study. J. Affect. Disord. 235, 506–512. doi: 10.1016/j.jad.2018.04.038
Lee, K., Kim, N., Shim, J. O., and Kim, G. H. (2020). Gut bacterial dysbiosis in children with intractable epilepsy. J. Clin. Med. 10:5. doi: 10.3390/jcm10010005
Lee, H., Lee, S., Lee, D. H., and Kim, D. W. (2021). A comparison of the gut microbiota among adult patients with drug-responsive and drug-resistant epilepsy: an exploratory study. Epilepsy Res. 172:106601. doi: 10.1016/j.eplepsyres.2021.106601
Li, Q., Han, Y., Dy, A., and Hagerman, R. J. (2017). The gut microbiota and autism spectrum disorders. Front. Cell. Neurosci. 11:120. doi: 10.3389/fncel.2017.00120
Lian, X., Liu, Z., Wu, T., Lou, J., Chen, Y., Liu, S., et al. (2023). Oral microbiome alterations in epilepsy and after seizure control. Front. Microbiol. 14:1277022. doi: 10.3389/fmicb.2023.1277022
Lim, J. M., Letchumanan, V., Tan, L. T., Hong, K. W., Wong, S. H., Ab, M. N., et al. (2022). Ketogenic diet: a dietary intervention via gut microbiome modulation for the treatment of neurological and nutritional disorders (a narrative review). Nutrients 14:3566. doi: 10.3390/nu14173566
Lindefeldt, M., Eng, A., Darban, H., Bjerkner, A., Zetterstrom, C. K., Allander, T., et al. (2019). The ketogenic diet influences taxonomic and functional composition of the gut microbiota in children with severe epilepsy. NPJ Biofilms Microbomes 5:5. doi: 10.1038/s41522-018-0073-2
Ling, Z., Liu, X., Cheng, Y., Yan, X., and Wu, S. (2022). Gut microbiota and aging. Crit. Rev. Food Sci. Nutr. 62, 3509–3534. doi: 10.1080/10408398.2020.1867054
Liu, S., Liu, L., Jin, D., Zhang, Q., and Takai, S. (2020). The novel mechanism of valproate to prevent peritoneal adhesion formation. Surg. Today 50, 1091–1098. doi: 10.1007/s00595-020-01979-8
Liu, Y. X., Qin, Y., Chen, T., Lu, M., Qian, X., Guo, X., et al. (2021). A practical guide to amplicon and metagenomic analysis of microbiome data. Protein Cell 12, 315–330. doi: 10.1007/s13238-020-00724-8
Liu, Y., Sanderson, D., Mian, M. F., Mcvey, N. K., and Forsythe, P. (2021). Loss of vagal integrity disrupts immune components of the microbiota-gut-brain axis and inhibits the effect of lactobacillus rhamnosus on behavior and the corticosterone stress response. Neuropharmacology 195:108682. doi: 10.1016/j.neuropharm.2021.108682
Rekdal, V. M., and Balskus, E. P. (2018). Gut microbiota: rational manipulation of gut bacterial metalloenzymes provides insights into dysbiosis and inflammation. Biochemistry 57, 2291–2293. doi: 10.1021/acs.biochem.8b00340
Maia, G. H., Brazete, C. S., Soares, J. I., Luz, L. L., and Lukoyanov, N. V. (2017). Serotonin depletion increases seizure susceptibility and worsens neuropathological outcomes in kainate model of epilepsy. Brain Res. Bull. 134, 109–120. doi: 10.1016/j.brainresbull.2017.07.009
Maneu, V., Noailles, A., Gomez-Vicente, V., Carpena, N., Cuenca, N., Gil, M. L., et al. (2016). Immunosuppression, peripheral inflammation and invasive infection from endogenous gut microbiota activate retinal microglia in mouse models. Microbiol. Immunol. 60, 617–625. doi: 10.1111/1348-0421.12405
Martin, C. R., Osadchiy, V., Kalani, A., and Mayer, E. A. (2018). The brain-gut-microbiome axis. Cell. Mol. Gastroenterol. Hepatol. 6, 133–148. doi: 10.1016/j.jcmgh.2018.04.003
Mckernan, K. J., Peckham, H. E., Costa, G. L., Mclaughlin, S. F., Fu, Y., Tsung, E. F., et al. (2009). Sequence and structural variation in a human genome uncovered by short-read, massively parallel ligation sequencing using two-base encoding. Genome Res. 19, 1527–1541. doi: 10.1101/gr.091868.109
Meng, C., Deng, P., Miao, R., Tang, H., Li, Y., Wang, J., et al. (2023). Gut microbiome and risk of ischaemic stroke: a comprehensive mendelian randomization study. Eur. J. Prev. Cardiol. 30, 613–620. doi: 10.1093/eurjpc/zwad052
Moreno, B., Crujeiras, A. B., Bellido, D., Sajoux, I., and Casanueva, F. F. (2016). Obesity treatment by very low-calorie-ketogenic diet at two years: reduction in visceral fat and on the burden of disease. Endocrine 54, 681–690. doi: 10.1007/s12020-016-1050-2
Morrison, D. J., and Preston, T. (2016). Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 7, 189–200. doi: 10.1080/19490976.2015.1134082
Mu, C., Nikpoor, N., Tompkins, T. A., Choudhary, A., Wang, M., Marks, W. N., et al. (2022). Targeted gut microbiota manipulation attenuates seizures in a model of infantile spasms syndrome. JCI Insight 7. doi: 10.1172/jci.insight.158521
Newell, C., Bomhof, M. R., Reimer, R. A., Hittel, D. S., Rho, J. M., and Shearer, J. (2016). Ketogenic diet modifies the gut microbiota in a murine model of autism spectrum disorder. Mol. Autism. 7:37. doi: 10.1186/s13229-016-0099-3
Newton, C. R., and Garcia, H. H. (2012). Epilepsy in poor regions of the world. Lancet 380, 1193–1201. doi: 10.1016/S0140-6736(12)61381-6
Nimgampalle, M., and Kuna, Y. (2017). Anti-alzheimer properties of probiotic, lactobacillus plantarum mtcc 1325 in alzheimer's disease induced albino rats. J. Clin. Diagn. Res. 11:KC01–KC05. doi: 10.7860/JCDR/2017/26106.10428
Ok, J. H., Lee, H., Chung, H. Y., Lee, S. H., Choi, E. J., Kang, C. M., et al. (2018). The potential use of a ketogenic diet in pancreatobiliary cancer patients after pancreatectomy. Anticancer Res. 38, 6519–6527. doi: 10.21873/anticanres.13017
Olson, C. A., Vuong, H. E., Yano, J. M., Liang, Q. Y., Nusbaum, D. J., and Hsiao, E. Y. (2018). The gut microbiota mediates the anti-seizure effects of the ketogenic diet. Cell 173, 1728–1741.e13. doi: 10.1016/j.cell.2018.04.027
Ouyang, Y., Chen, Y., Wang, G., Song, Y., Zhao, H., Xiao, B., et al. (2022). Genetically proxied gut microbiota, gut metabolites with risk of epilepsy and the subtypes: a bi-directional mendelian randomization study. Front. Molec. Neurosci. 15:994270. doi: 10.3389/fnmol.2022.994270
Ozkara, C., Benbir, G., and Celik, A. F. (2009). Misdiagnosis due to gastrointestinal symptoms in an adolescent with probable autonomic status epilepticus and panayiotopoulos syndrome. Epilepsy Behav. 14, 703–704. doi: 10.1016/j.yebeh.2009.02.013
Paoli, A., Mancin, L., Bianco, A., Thomas, E., Mota, J. F., and Piccini, F. (2019). Ketogenic diet and microbiota: friends or enemies? Genes 10:534. doi: 10.3390/genes10070534
Peng, Y., Chiu, A., Li, V., Zhang, X., Yeung, W. L., Chan, S., et al. (2023). The role of the gut-microbiome-brain axis in metabolic remodeling amongst children with cerebral palsy and epilepsy. Front. Neurol. 14:1109469. doi: 10.3389/fneur.2023.1109469
Peng, A., Qiu, X., Lai, W., Li, W., Zhang, L., Zhu, X., et al. (2018). Altered composition of the gut microbiome in patients with drug-resistant epilepsy. Epilepsy Res. 147, 102–107. doi: 10.1016/j.eplepsyres.2018.09.013
Pietrucci, D., Cerroni, R., Unida, V., Farcomeni, A., Pierantozzi, M., Mercuri, N. B., et al. (2019). Dysbiosis of gut microbiota in a selected population of parkinson's patients. Parkinsonism Relat. Disord. 65, 124–130. doi: 10.1016/j.parkreldis.2019.06.003
Quigley, E. (2017). Microbiota-brain-gut axis and neurodegenerative diseases. Curr. Neurol. Neurosci. Rep. 17:94. doi: 10.1007/s11910-017-0802-6
Ranjan, R., Rani, A., Metwally, A., Mcgee, H. S., and Perkins, D. L. (2016). Analysis of the microbiome: advantages of whole genome shotgun versus 16s amplicon sequencing. Biochem. Biophys. Res. Commun. 469, 967–977. doi: 10.1016/j.bbrc.2015.12.083
Rho, J. M. (2004). Basic science behind the catastrophic epilepsies. Epilepsia 45, 5–11. doi: 10.1111/j.0013-9580.2004.05001.x
Rothschild, D., Weissbrod, O., Barkan, E., Kurilshikov, A., Korem, T., Zeevi, D., et al. (2018). Environment dominates over host genetics in shaping human gut microbiota. Nature 555, 210–215. doi: 10.1038/nature25973
Saad, M. J., Santos, A., and Prada, P. O. (2016). Linking gut microbiota and inflammation to obesity and insulin resistance. Physiology 31, 283–293. doi: 10.1152/physiol.00041.2015
Safak, B., Altunan, B., Topcu, B., and Eren, T. A. (2020). The gut microbiome in epilepsy. Microb. Pathog. 139:103853. doi: 10.1016/j.micpath.2019.103853
Sampson, T. R., Debelius, J. W., Thron, T., Janssen, S., Shastri, G. G., Ilhan, Z. E., et al. (2016). Gut microbiota regulate motor deficits and neuroinflammation in a model of parkinson's disease. Cell 167, 1469–1480.e12. doi: 10.1016/j.cell.2016.11.018
Saslow, L. R., Mason, A. E., Kim, S., Goldman, V., Ploutz-Snyder, R., Bayandorian, H., et al. (2017). An online intervention comparing a very low-carbohydrate ketogenic diet and lifestyle recommendations versus a plate method diet in overweight individuals with type 2 diabetes: a randomized controlled trial. J. Med. Internet Res. 19:e36. doi: 10.2196/jmir.5806
Saxena, S., and Li, S. (2017). Defeating epilepsy: a global public health commitment. Epilepsia Open 2, 153–155. doi: 10.1002/epi4.12010
Scheperjans, F., Aho, V., Pereira, P. A., Koskinen, K., Paulin, L., Pekkonen, E., et al. (2015). Gut microbiota are related to parkinson's disease and clinical phenotype. Mov. Disord. 30, 350–358. doi: 10.1002/mds.26069
Sen, T., Cawthon, C. R., Ihde, B. T., Hajnal, A., Dilorenzo, P. M., de La Serre, C. B., et al. (2017). Diet-driven microbiota dysbiosis is associated with vagal remodeling and obesity. Physiol. Behav. 173, 305–317. doi: 10.1016/j.physbeh.2017.02.027
Shen, L., and Ji, H. F. (2019). Associations between gut microbiota and alzheimer's disease: current evidences and future therapeutic and diagnostic perspectives. J. Alzheimers Dis. 68, 25–31. doi: 10.3233/JAD-181143
Singh, G., and Sander, J. W. (2020). The global burden of epilepsy report: implications for low- and middle-income countries. Epilepsy Behav. 105:106949. doi: 10.1016/j.yebeh.2020.106949
Thijs, R. D., Surges, R., O'Brien, T. J., and Sander, J. W. (2019). Epilepsy in adults. Lancet 393, 689–701. doi: 10.1016/S0140-6736(18)32596-0
Trick, W. E., Sokalski, S. J., Johnson, S., Bunnell, K. L., Levato, J., Ray, M. J., et al. (2018). Effectiveness of probiotic for primary prevention of clostridium difficile infection: a single-center before-and-after quality improvement intervention at a tertiary-care medical center. Infect. Control Hosp. Epidemiol. 39, 765–770. doi: 10.1017/ice.2018.76
Turay, S., Cangur, S., Kahraman, G., Kayabasi, E., Cetiner, O. F., Aydin, B., et al. (2023). Can the gut microbiota serve as a guide to the diagnosis and treatment of childhood epilepsy? Pediatr. Neurol. 145, 11–21. doi: 10.1016/j.pediatrneurol.2023.04.006
Uchida, T., Lossin, C., Ihara, Y., Deshimaru, M., Yanagawa, Y., Koyama, S., et al. (2017). Abnormal gamma-aminobutyric acid neurotransmission in a kcnq2 model of early onset epilepsy. Epilepsia 58, 1430–1439. doi: 10.1111/epi.13807
van der Louw, E., van den Hurk, D., Neal, E., Leiendecker, B., Fitzsimmon, G., Dority, L., et al. (2016). Ketogenic diet guidelines for infants with refractory epilepsy. Eur. J. Paediatr. Neurol. 20, 798–809. doi: 10.1016/j.ejpn.2016.07.009
Vogtmann, E., Hua, X., Zeller, G., Sunagawa, S., Voigt, A. Y., Hercog, R., et al. (2016). Colorectal cancer and the human gut microbiome: reproducibility with whole-genome shotgun sequencing. PLoS One 11:e0155362. doi: 10.1371/journal.pone.0155362
Volmering, E., Niehusmann, P., Peeva, V., Grote, A., Zsurka, G., Altmuller, J., et al. (2016). Neuropathological signs of inflammation correlate with mitochondrial dna deletions in mesial temporal lobe epilepsy. Acta Neuropathol. 132, 277–288. doi: 10.1007/s00401-016-1561-1
Wan, L., Shi, X., Yan, H., Liang, Y., Liu, X., Zhu, G., et al. (2024). Abnormalities in clostridioides and related metabolites before acth treatment may be associated with its efficacy in patients with infantile epileptic spasm syndrome. CNS Neurosci. Ther. 30:e14398. doi: 10.1111/cns.14398
Whiteside, S. A., Razvi, H., Dave, S., Reid, G., and Burton, J. P. (2015). The microbiome of the urinary tract--a role beyond infection. Nat. Rev. Urol. 12, 81–90. doi: 10.1038/nrurol.2014.361
Wilson, B., Rossi, M., Dimidi, E., and Whelan, K. (2019). Prebiotics in irritable bowel syndrome and other functional bowel disorders in adults: a systematic review and meta-analysis of randomized controlled trials. Am. J. Clin. Nutr. 109, 1098–1111. doi: 10.1093/ajcn/nqy376
Winek, K., Engel, O., Koduah, P., Heimesaat, M. M., Fischer, A., Bereswill, S., et al. (2016). Depletion of cultivatable gut microbiota by broad-spectrum antibiotic pretreatment worsens outcome after murine stroke. Stroke 47, 1354–1363. doi: 10.1161/STROKEAHA.115.011800
Xie, G., Zhou, Q., Qiu, C. Z., Dai, W. K., Wang, H. P., Li, Y. H., et al. (2017). Ketogenic diet poses a significant effect on imbalanced gut microbiota in infants with refractory epilepsy. World J. Gastroenterol. 23, 6164–6171. doi: 10.3748/wjg.v23.i33.6164
Xu, L., Chen, D., Zhao, C., Jiang, L., Mao, S., Song, C., et al. (2021). Decreased abundance of akkermansia after adrenocorticotropic hormone therapy in patients with west syndrome. BMC Microbiol. 21:126. doi: 10.1186/s12866-021-02189-z
Yankam, N. J., Costes, N., Bouillot, C., Bouvard, S., Fieux, S., Becker, G., et al. (2017). Quantitative longitudinal imaging of activated microglia as a marker of inflammation in the pilocarpine rat model of epilepsy using [(11)c]-(r)-pk11195 pet and mri. J. Cereb. Blood Flow Metab. 37, 1251–1263. doi: 10.1177/0271678X16653615
Yeom, J. S., Park, J. S., Kim, Y. S., Kim, R. B., Choi, D. S., Chung, J. Y., et al. (2019). Neonatal seizures and white matter injury: role of rotavirus infection and probiotics. Brain and Development 41, 19–28. doi: 10.1016/j.braindev.2018.07.001
Yuan, W., Lu, W., Wang, H., Wu, W., Zhou, Q., Chen, Y., et al. (2022). A multiphase dietetic protocol incorporating an improved ketogenic diet enhances weight loss and alters the gut microbiome of obese people. Int. J. Food Sci. Nutr. 73, 238–250. doi: 10.1080/09637486.2021.1960957
Yunus, Y., Sefer, U., Dondu, U. U., Ismail, O., and Yusuf, E. (2016). Abdominal epilepsy as an unusual cause of abdominal pain: a case report. Afr. Health Sci. 16, 877–879. doi: 10.4314/ahs.v16i3.32
Zeng, H., Huang, C., Lin, S., Zheng, M., Chen, C., Zheng, B., et al. (2017). Lotus seed resistant starch regulates gut microbiota and increases short-chain fatty acids production and mineral absorption in mice. J. Agric. Food Chem. 65, 9217–9225. doi: 10.1021/acs.jafc.7b02860
Zhang, X., Tang, B., and Guo, J. (2023). Parkinson's disease and gut microbiota: from clinical to mechanistic and therapeutic studies. Transl. Neurodegener. 12:59. doi: 10.1186/s40035-023-00392-8
Zhang, Y., Zhou, S., Zhou, Y., Yu, L., Zhang, L., and Wang, Y. (2018). Altered gut microbiome composition in children with refractory epilepsy after ketogenic diet. Epilepsy Res. 145, 163–168. doi: 10.1016/j.eplepsyres.2018.06.015
Zhao, L., Ni, Y., Su, M., Li, H., Dong, F., Chen, W., et al. (2017). High throughput and quantitative measurement of microbial metabolome by gas chromatography/mass spectrometry using automated alkyl chloroformate derivatization. Anal. Chem. 89, 5565–5577. doi: 10.1021/acs.analchem.7b00660
Zheng, H., Zhang, C., Zhang, J., and Duan, L. (2023). "sentinel or accomplice": gut microbiota and microglia crosstalk in disorders of gut-brain interaction. Protein Cell 14, 726–742. doi: 10.1093/procel/pwad020
Zhernakova, D. V., Wang, D., Liu, L., Andreu-Sanchez, S., Zhang, Y., Ruiz-Moreno, A. J., et al. (2024). Host genetic regulation of human gut microbial structural variation. Nature 625, 813–821. doi: 10.1038/s41586-023-06893-w
Zhou, C., Gong, S., Xiang, S., Liang, L., Hu, X., Huang, R., et al. (2022). Changes and significance of gut microbiota in children with focal epilepsy before and after treatment. Front. Cell. Infect. Microbiol. 12:965471. doi: 10.3389/fcimb.2022.965471
Keywords: epilepsy, gut microbiota, brain–gut axis, probiotics, ketogenic diet
Citation: Li Q, Gu Y, Liang J, Yang Z and Qin J (2024) A long journey to treat epilepsy with the gut microbiota. Front. Cell. Neurosci. 18:1386205. doi: 10.3389/fncel.2024.1386205
Edited by:
Hayet Sellami, University of Sfax, TunisiaReviewed by:
Mingzi Zhang, University of Southern California, United StatesCopyright © 2024 Li, Gu, Liang, Yang and Qin. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiong Qin, cWluamlvbmdAcGt1cGguZWR1LmNu
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.