 Alexandre Bouron
Alexandre Bouron Laurence Aubry1
Laurence Aubry1 Desirée Loreth
Desirée Loreth Marie-Odile Fauvarque
Marie-Odile Fauvarque- 1Université Grenoble Alpes, Inserm, CEA, UA13, BGE, Grenoble, France
- 2Institute of Cellular and Integrative Physiology, University Medical Center Hamburg-Eppendorf, Hamburg, Germany
1. Introduction: Identification of UCH-L1 (PGP9.5)
High-resolution two-dimensional electrophoresis was employed to detect soluble protein gene products (PGPs) in aqueous extracts from human brain and twelve other organs (Jackson and Thompson, 1981). This permitted the identification of four specific brain PGPs, one of them termed PGP9.5. Its molecular identity was unknown at the time but PGP9.5 was described as a “soluble nervous system-specific protein” (Jackson and Thompson, 1981) albeit found at much lower levels in other organs like kidney, large intestine, prostate and testis. The PGP9.5 protein was shown to display COOH-terminal hydrolase activity (Wilkinson et al., 1989) and finally turned out to correspond to ubiquitin carboxyl-terminal hydrolase L1 (UCH-L1), an enzyme previously isolated from cytosolic fractions of bovine calf thymus (Duerksen-Hughes et al., 1989; Mayer and Wilkinson, 1989). Immunoperoxidase staining of human cerebral cortex sections showed the restricted presence of UCH-L1 (PGP9.5) in neurons (Doran et al., 1983). Its brain concentration was at least 50 times greater than in other organs like kidney, prostate, or testis. Doran et al. (1983) introduced UCH-L1 as a “new neurone-specific cytoplasmic marker” thought to represent 1–2% of total brain soluble proteins. Additional studies extended these observations by showing that UCH-L1 was not only present in neurons of the central nervous system but also in neurons of the peripheral nervous system and in cells of the neuroendocrine system (Thompson et al., 1983). UCH-L1 (PGP9.5) (mRNA and protein) was detected at very early stages (e.g., 10 days p.c.) of neural tube formation (Schofield et al., 1995) in progenitor cells and neurons (Kent and Clarke, 1991).
2. UCH-L1 subcellular localization
Although initially described as a major protein of the cytosol, nearly 30% of neuronal UCH-L1 is membrane-associated (Bishop et al., 2014). It is now clear that a significant portion of UCH-L1 is associated to membranes of the endoplasmic reticulum (ER) (Liu et al., 2009; Gao et al., 2020). Immunogold labeling experiments on rat proximal caput epididymidis had previously revealed a mitochondrial localization (Martin et al., 1995). The association of UCH-L1 with mitochondria has however been questioned. For instance, the presence of UCH-L1 in mitochondrial-enriched fractions isolated from brain was proposed to result from a contamination with other fractions such as synaptosomes (Van Laar et al., 2009). Subsequent studies provided novel arguments in favor of a link between UCH-L1 and mitochondria. (i) A truncated variant of UCH-L1 (NT-UCH-L1) identified in the brain was found to associate with mitochondria (Kim et al., 2014). (ii) Low levels of the enzyme were detected in mitochondrial fractions prepared from soleus, a skeletal muscle that preferentially contains slow oxidative muscle fibers (Gao et al., 2020). (iii) Immunofluorescence stainings showed a close association of UCH-L1 with ER and mitochondria in muscle and neuroblastoma SH-SY5Y cells (Cerqueira et al., 2020; Gao et al., 2020). (iv) The DUB was shown to influence the morphology and respiratory functions of mitochondria in skeletal muscle (Gao et al., 2020). Altogether, these experimental data point toward the existence of a link between UCH-L1 and the vital organelles mitochondria.
3. Molecular and cellular functions of UCH-L1
UCH-L1 is a 223 amino acids protein (25 kDa) belonging to the Ub carboxyl-terminal hydrolases (UCHs) subfamily of DUBs that comprises three other members: UCH-L3, UCH-L5 (UCH37) and BAP1 (BRCA1-associated protein 1). So far, three main functions have been associated to this protein (Mi and Graham, 2023). UCH-L1 can generate free Ub species, thereby contributing to the recycling of Ub. Intriguingly, it also exhibits a Ub E3 ligase activity (Liu et al., 2002). However, the main cellular function of UCH-L1 is thought to be related to its binding to monomeric Ub that prevents the proteasomal degradation of the polypeptide. Thus, by maintaining a pool of available Ub within cells, UCH-L1 is a key regulator of the multiple cellular Ub-dependent biological processes. In addition, altered protein expression levels of UCH-L1 have been observed in a wide range of tumors (Jara et al., 2013; Sharma et al., 2020).
Originally introduced as a specific neuronal brain protein, mutations in the human UCH-L1 protein, animal models and pharmacological studies indicate that the loss of UCH-L1 or the impairment of its enzymatic activity is associated with neurodegenerative phenotypes (Mi and Graham, 2023). UCH-L1 seems required to maintain the integrity of synaptic structures as shown in the hippocampus (Cartier et al., 2009) and at the neuromuscular junction (Chen et al., 2010). UCH-L1 has also been introduced as a promising serum biomarker of neuronal cell injury in patients with traumatic brain injury (Papa et al., 2016). However, a detailed molecular understanding of the neuronal functions of UCH-L1 in health and disease is still lacking.
4. UCH-L1 and mitochondrial proteins and functions
As mentioned above, a fraction of UCH-L1 has been shown to localize to mitochondria. In parallel, several studies have documented a functional link between the DUB and these organelles. Studies conducted on neuroblastoma SH-SY5Y and immortalized INS1 beta cells showed that silencing of UCH-L1 expression (via a shRNA strategy) induces significant changes in mitochondrial morphology and reduces interactions (tethering) between mitochondria and the ER by increasing the physical distance between the two organelles (Cerqueira et al., 2020). In addition, UCH-L1 was shown to associate with mitochondrial proteins. Immunoprecipitation experiments identified the mitochondrial chaperone protein HSP60 as a partner of UCH-L1 in skeletal muscle from mice (Gao et al., 2020). Tandem affinity purification coupled to mass spectrometry showed that UCH-L1 can also form a complex with Parkin, an E3 Ub ligase involved in mitophagy, when overexpressed in HEK-293 cells (Davison et al., 2009), suggesting that UCH-L1 may participate to the clearance of dysfunctional mitochondria.
Another interesting feature of UCH-L1 is its impact on the abundance of various mitochondrial proteins. For instance, diminishing and increasing UCH-L1 expression have been shown to reduce and enhance mitofusin protein levels, respectively (Cerqueira et al., 2020). However, UCH-L1 has been proposed to regulate mitochondrial physiology beyond mitophagy (Cerqueira et al., 2020). Deletion of UCH-L1 in the brain of mice increases the protein levels of the manganese (Mn) superoxide dismutase (MnSOD) and the ATP synthase subunit beta (ATPB), two mitochondrial matrix proteins that exert antioxidant and ATP production functions, respectively (Reinicke et al., 2019). In skeletal muscles from mice, the loss of UCH-L1 decreases the abundance of SBHD (component of complex II) and UQCRC2 (component of complex III) proteins, whereas it increases the protein abundance of NDUFB8 (a protein of complex I) (Gao et al., 2020), all three proteins being components of the mitochondrial respiratory chain. Of note, deletion of the Uch-l1 gene (dUch) in Drosophila brain elevates the mRNA levels of the cytosolic and mitochondrial SOD (Huynh et al., 2022), an observation supporting a conserved functional relationship between UCH-L1 and mitochondrial proteins in fly and mammals.
Direct measurements of mitochondrial activity have demonstrated that UCH-L1 interferes with mitochondrial bioenergetics. In SH-SY5Y and immortalized INS1 beta cells, UCH-L1 silencing augments the activity of the mitochondrial complex I, the mitochondrial ATP production and oxygen consumption (Cerqueira et al., 2020). In mouse muscle cells, the lack of UCH-L1 reduces the activity of the mitochondrial electron transfer chain complex II/III (Gao et al., 2020), which is likely to perturb the mitochondrial production of ATP. In Drosophila brains, deletion of dUch causes a severe reduction (>50%) of the ATP levels without modifying the mitochondrial membrane potential (Huynh et al., 2022). In contrast, the pharmacological inhibition of UCH-L1 activity with LDN-57444 has been shown to reduce the mitochondrial membrane potential of oocytes (Yuan et al., 2020).
A pathophysiological connection between UCH-L1 and mitochondria has been established in Alzheimer's and Parkinson's diseases, two prominent neurodegenerative disorders. A significant fraction of UCH-L1 colocalizes with Parkin and mitochondria in vivo in Alzheimer's disease brains as well as in transgenic Alzheimer's disease mice (Corsetti et al., 2015). In association with Parkin, UCH-L1 promotes neuronal death by causing an extensive mitochondrial clearance (Corsetti et al., 2015). Nakamura et al. (2021) demonstrated another role of UCH-L1 in Alzheimer's disease: the DUB can function as an upstream signaling molecule participating in an intracellular cascade involving Cdk5 and leading to the transfer of nitric oxide (NO) to the mitochondrial GTPase Drp1 to form NO-Drp1. This non-canonical pathway has been proposed to stimulate mitochondrial fragmentation and bioenergetics dysregulation leading to synaptic insults (Nakamura et al., 2021). The UCH-L1-dependent transnitrosylation pathway could play a role in Alzheimer's disease-related synapse loss and in the pathogenesis of neurodegenerative disorders (Nakamura et al., 2021). Additionally, Ham et al. (2021) showed that UCH-L1 could play a role in Parkinson's disease by (indirectly) controlling mitochondrial homeostasis. UCH-L1 loss promotes mitophagy in Drosophila mutants and mammalian cell lines, probably via a process involving pyruvate kinase, a key glycolytic enzyme. Indeed, depletion of UCH-L1 strongly decreases the amount of pyruvate kinase by favoring its proteasomal degradation. Altogether, UCH-L1 seems to exert a house-keeping role in mitochondrial homeostasis that depends on the cytosolic energy status (Ham et al., 2021). Moreover, the authors showed that loss of UCH-L1 rescues some defects related to Parkinson's disease. Taken together, these data indicate that UCH-L1 could participate in the regulation of glucose metabolism, mitochondrial homeostasis, and Parkinson's disease pathogenesis (Ham et al., 2021). This link between mitochondria and UCH-L1 has been reinforced by Roy et al. (2022) who showed another aspect of this complex regulation: the pesticide rotenone inhibits the electron transport chain complex I of neuroblastoma N2A cells, enhancing the ROS production while decreasing cellular ATP levels and collapsing the mitochondrial membrane potential. Importantly, rotenone strongly diminishes UCH-L1 mRNA and protein levels (Roy et al., 2022). This observation suggests the intriguing possibility of the existence of a bidirectional regulation of mitochondria and UCH-L1: UCH-L1 influences mitochondrial activity while mitochondria regulate the expression of UCH-L1.
5. Conclusion
Altogether, these studies point toward a complex and multifaceted evolutionary conserved role of UCH-L1 in mitochondrial functions (Figure 1). Understanding the molecular mechanisms at work is a promising and exciting avenue for future studies. They should bring important information on the pathophysiological relevance of this DUB in neuronal physiology and health and disease states.
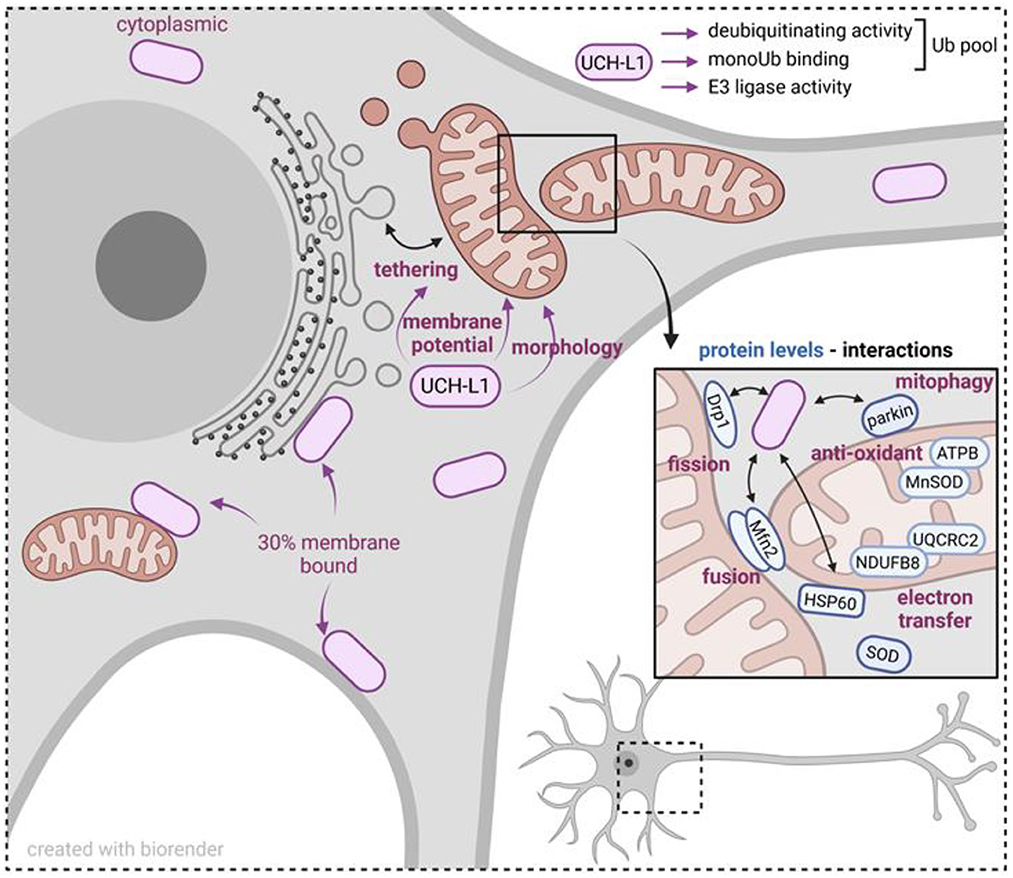
Figure 1. Scheme summarizing mitochondrial proteins and mitochondrial functions influenced by UCH-L1. UCH-L1 is the most abundant deubiquitinating enzyme within neurons, predominantly in the cytoplasm, and 30% as a membrane bound protein. Biochemically, UCH-L1 regulates the cytoplasmic pool of mono ubiquitin (Ub) and exerts an E3 ligase activity. UCH-L1 localizes to the mitochondrial membrane and is associated with modifying mitochondrial-ER tethering, mitochondrial membrane potential and morphology. UCH-L1 impacts the abundance of mitochondrial cytoplasmic and matrix proteins and by this influences mitochondrial fusion and fission processes, electron transfer, and removal of defective mitochondria by mitophagy. In summary, UCH-L1 affects mitochondrial homeostasis and cellular energy status.
Author contributions
AB, LA, DL, M-OF, and CM-S: bibliographic search, manuscript writing, and editing. All authors contributed to the article and approved the submitted version.
Acknowledgments
The authors wish to acknowledge support from the Center National de la Recherche Scientifique (CNRS), Commissariat à l'Energie Atomique et aux Energies Alternatives (CEA), Université de Grenoble Alpes (UGA), and Institut National de la Santé et de la Recherche Médicale (Inserm).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Bishop, P., Rubin, P., Thomson, A. R., Rocca, D., and Henley, J. M. (2014). The ubiquitin C-terminal hydrolase L1 (UCH-L1) C terminus plays a key role in protein stability, but its farnesylation is not required for membrane association in primary neurons. J. Biol. Chem. 289, 36140–36149. doi: 10.1074/jbc.M114.557124
Cartier, A. E., Djakovic, S. N., Salehi, A., Wilson, S. M., Masliah, E., and Patrick, G. N. (2009). Regulation of synaptic structure by ubiquitin C-terminal hydrolase L1. J. Neurosci. 29, 7857–7868. doi: 10.1523/JNEUROSCI.1817-09.2009
Cerqueira, F. M., von Stockum, S., Giacomello, M., Goliand, I., Kakimoto, P., Marchesan, E., et al. (2020). A new target for an old DUB: UCH-L1 regulates mitofusin-2 levels, altering mitochondrial morphology, function and calcium uptake. Redox. Biol. 37, 101676. doi: 10.1016/j.redox.2020.101676
Chen, F., Sugiura, Y., Myers, K. G., Liu, Y., and Lin, W. (2010). Ubiquitin carboxyl-terminal hydrolase L1 is required for maintaining the structure and function of the neuromuscular junction. Proc. Natl. Acad. Sci. U. S. A. 107, 1636–1641. doi: 10.1073/pnas.0911516107
Corsetti, V., Florenzano, F., Atlante, A., Bobba, A., Ciotti, M. T., Natale, F., et al. (2015). NH2-truncated human tau induces deregulated mitophagy in neurons by aberrant recruitment of Parkin and UCHL-1: implications in Alzheimer's disease. Hum. Mol. Genet. 24, 3058–3081. doi: 10.1093/hmg/ddv059
Davison, E. J., Pennington, K., Hung, C. C., Peng, J., Rafiq, R., Ostareck-Lederer, A., et al. (2009). Proteomic analysis of increased Parkin expression and its interactants provides evidence for a role in modulation of mitochondrial function. Proteomics 9, 4284–4297. doi: 10.1002/pmic.200900126
Doran, J. F., Jackson, P., Kynoch, P. A., and Thompson, R. J. (1983). Isolation of PGP 9.5, a new human neurone-specific protein detected by high-resolution two-dimensional electrophoresis. J. Neurochem. 40, 1542–1547. doi: 10.1111/j.1471-4159.1983.tb08124.x
Duerksen-Hughes, P. J., Williamson, M. M., and Wilkinson, K. D. (1989). Affinity chromatography using protein immobilized via arginine residues: purification of ubiquitin carboxyl-terminal hydrolases. Biochemistry 28, 8530–8536. doi: 10.1021/bi00447a039
Gao, H., Antony, R., Srinivasan, R., Wu, P., Wang, X., and Li, Y. (2020). UCHL1 regulates oxidative activity in skeletal muscle. PLoS ONE 15, e0241716. doi: 10.1371/journal.pone.0241716
Ham, S. J., Lee, D., Xu, W. J., Cho, E., Choi, S., Min, S., et al. (2021). Loss of UCHL1 rescues the defects related to Parkinson's disease by suppressing glycolysis. Sci. Adv. 7, 4574. doi: 10.1126/sciadv.abg4574
Huynh, T. K. T., Mai, T. T. T., Huynh, M. A., Yoshida, H., Yamaguchi, M., and Dang, T. T. P. (2022). Crucial roles of ubiquitin carboxy-terminal hydrolase L1 in motor neuronal health by drosophila model. Antioxid. Redox. Signal 37, 257–273. doi: 10.1089/ars.2021.0057
Jackson, P., and Thompson, R. J. (1981). The demonstration of new human brain-specific proteins by high-resolution two-dimensional polyacrylamide gel electrophoresis. J. Neurol. Sci. 49, 429–438. doi: 10.1016/0022-510X(81)90032-0
Jara, J. H., Frank, D. D., and Ozdinler, P. H. (2013). Could dysregulation of UPS be a common underlying mechanism for cancer and neurodegeneration? Lessons from UCHL1. Cell Biochem. Biophys. 67, 45–53. doi: 10.1007/s12013-013-9631-7
Kent, C., and Clarke, P. J. (1991). The immunolocalisation of the neuroendocrine specific protein PGP9.5 during neurogenesis in the rat. Brain Res. Dev. Brain Res. 58, 147–150. doi: 10.1016/0165-3806(91)90248-H
Kim, H. J., Kim, H. J., Jeong, J. E., Baek, J. Y., Jeong, J., Kim, S., et al. (2014). N-terminal truncated UCH-L1 prevents Parkinson's disease associated damage. PLoS ONE 9, e99654. doi: 10.1371/journal.pone.0099654
Liu, Y., Fallon, L., Lashuel, H. A., Liu, Z., and Lansbury, P. T. Jr. (2002). The UCH-L1 gene encodes two opposing enzymatic activities that affect alpha-synuclein degradation and Parkinson's disease susceptibility. Cell 111, 209–218. doi: 10.1016/S0092-8674(02)01012-7
Liu, Z., Meray, R. K., Grammatopoulos, T. N., Fredenburg, R. A., Cookson, M. R., Liu, Y., et al. (2009). Membrane-associated farnesylated UCH-L1 promotes alpha-synuclein neurotoxicity and is a therapeutic target for Parkinson's disease. Proc. Natl. Acad. Sci. U. S. A. 106, 4635–4640. doi: 10.1073/pnas.0806474106
Martin, R., Santamaria, L., Fraile, B., Paniagua, R., and Polak, J. M. (1995). Ultrastructural localization of PGP 9.5 and ubiquitin immunoreactivities in rat ductus epididymidis epithelium. Histochem. J. 27, 431–439.
Mayer, A. N., and Wilkinson, K. D. (1989). Detection, resolution, and nomenclature of multiple ubiquitin carboxyl-terminal esterases from bovine calf thymus. Biochemistry 28, 166–172. doi: 10.1021/bi00427a024
Mi, Z., and Graham, S. H. (2023). Role of UCHL1 in the pathogenesis of neurodegenerative diseases and brain injury. Ageing Res. Rev. 86, 101856. doi: 10.1016/j.arr.2023.101856
Nakamura, T., Oh, C. K., Liao, L., Zhang, X., Lopez, K. M., Gibbs, D., et al. (2021). Non-canonical transnitrosylation network contributes to synapse loss in Alzheimer's disease. Science 371, 843. doi: 10.1126/science.aaw0843
Papa, L., Brophy, G. M., Welch, R. D., Lewis, L. M., Braga, C. F., Tan, C. N., et al. (2016). Time Course and diagnostic accuracy of glial and neuronal blood biomarkers GFAP and UCH-L1 in a large cohort of trauma patients with and without mild traumatic brain injury. JAMA Neurol. 73, 551–560. doi: 10.1001/jamaneurol.2016.0039
Reinicke, A. T., Laban, K., Sachs, M., Kraus, V., Walden, M., Damme, M., et al. (2019). Ubiquitin C-terminal hydrolase L1 (UCH-L1) loss causes neurodegeneration by altering protein turnover in the first postnatal weeks. Proc. Natl. Acad. Sci. U. S. A. 116, 7963–7972. doi: 10.1073/pnas.1812413116
Roy, T., Chatterjee, A., and Swarnakar, S. (2022). Rotenone induced neurodegeneration is mediated via cytoskeleton degradation and necroptosis. Biochim. Biophys. Acta Mol. Cell Res. 1870, 119417. doi: 10.1016/j.bbamcr.2022.119417
Schofield, J. N., Day, I. N., Thompson, R. J., and Edwards, Y. H. (1995). PGP9.5, a ubiquitin C-terminal hydrolase; pattern of mRNA and protein expression during neural development in the mouse. Brain Res. Dev. Brain Res. 85, 229–238. doi: 10.1016/0165-3806(94)00217-N
Sharma, A., Liu, H., Tobar-Tosse, F., Chand Dakal, T., Ludwig, M., Holz, F. G., et al. (2020). Ubiquitin carboxyl-terminal hydrolases (UCHs): potential mediators for cancer and neurodegeneration. Int. J. Mol. Sci. 21, 3910. doi: 10.3390/ijms21113910
Thompson, R. J., Doran, J. F., Jackson, P., Dhillon, A. P., and Rode, J. (1983). PGP 9.5: a new marker for vertebrate neurons and neuroendocrine cells. Brain Res. 278, 224–228. doi: 10.1016/0006-8993(83)90241-X
Van Laar, V. S., Mishizen, A. J., Cascio, M., and Hastings, T. G. (2009). Proteomic identification of dopamine-conjugated proteins from isolated rat brain mitochondria and SH-SY5Y cells. Neurobiol. Dis. 34, 487–500. doi: 10.1016/j.nbd.2009.03.004
Wilkinson, K. D., Lee, K. M., Deshpande, S., Duerksen-Hughes, P., Boss, J. M., and Pohl, J. (1989). The neuron-specific protein PGP 9.5 is a ubiquitin carboxyl-terminal hydrolase. Science 246, 670–673. doi: 10.1126/science.2530630
Keywords: mitochondria, brain, deubiquitinating enzymes, neurodegeneration, pathology, UCH-L1
Citation: Bouron A, Aubry L, Loreth D, Fauvarque M-O and Meyer-Schwesinger C (2023) Role of the deubiquitinating enzyme UCH-L1 in mitochondrial function. Front. Cell. Neurosci. 17:1149954. doi: 10.3389/fncel.2023.1149954
Received: 23 January 2023; Accepted: 10 March 2023;
Published: 23 March 2023.
Edited by:
Angela M. Mabb, Georgia State University, United StatesReviewed by:
Di Hu, Case Western Reserve University, United StatesCopyright © 2023 Bouron, Aubry, Loreth, Fauvarque and Meyer-Schwesinger. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexandre Bouron, YWxleGFuZHJlLmJvdXJvbkBjZWEuZnI=