
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Neurosci. , 14 June 2022
Sec. Cellular Neuropathology
Volume 16 - 2022 | https://doi.org/10.3389/fncel.2022.884813
This article is part of the Research Topic Natural products as sources of innovative approaches in the prevention and treatment of neuronal injuries induced by neurodegenerative diseases and environmental exposures to neurotoxic agents View all 5 articles
 Chirlene Pinheiro Nascimento1†
Chirlene Pinheiro Nascimento1† Luan Oliveira Ferreira2†
Luan Oliveira Ferreira2† Alex Luiz Menezes da Silva1Ana Beatriz Nardelli da Silva1Joao Cleiton Martins Rodrigues2
Alex Luiz Menezes da Silva1Ana Beatriz Nardelli da Silva1Joao Cleiton Martins Rodrigues2 Leonan Lima Teixeira2Julianne Elba Cunha Azevedo1Daniella Bastos de Araujo1
Leonan Lima Teixeira2Julianne Elba Cunha Azevedo1Daniella Bastos de Araujo1 Akira Otake Hamoy1
Akira Otake Hamoy1 Beatriz Holanda Gonçalves1Brenda Hosana De Oliveira Coelho1
Beatriz Holanda Gonçalves1Brenda Hosana De Oliveira Coelho1 Dielly Catrina Favacho Lopes2*Moisés Hamoy1*
Dielly Catrina Favacho Lopes2*Moisés Hamoy1*Epilepsy is one of the most common neurological disorders, which occurs due to the instability in the inhibitory and excitatory synaptic transmissions in the brain. However, many patients develop resistance to the available drugs, which results in cell degeneration caused due to inadequate control of the seizures. Curcumin, Curcuma longa, is known to be effective for the treatment of organic disorders and may prevent seizures, reduce oxidative stress, and decrease brain damage. Given this, the present study evaluated the antiepileptic effects of C. longa in comparison with both the diazepam and the combined application of these two substances, in terms of their effects on the brain activity and the potential histopathological changes in the hippocampus. This study used male Wistar rats (age: 10–12 weeks; weight: 260 ± 20 g), which were pretreated for 4 days with either saline, C. longa, diazepam, or C. longa + diazepam; and on the fifth day, pentylenetetrazol (PTZ) was administered to induce the seizure. In the C. longa group, a significant increase was observed in the latency of the onset of seizure-related behavior. Surprisingly, however, the combined treatment resulted in the best control of the seizure-related behavior, with the greatest latency of the onset of spasms and isolated clonic seizures. This group also obtained the best results in the electroencephalographic trace and seizure control, with a reduction in the frequency and amplitude of the spike-waves. In the saline group, PTZ significantly reduced the number of cells present in the CA1 and CA3 regions of the hippocampus, while the combined treatment obtained the best results in terms of the preservation of the neuron-like cells. These findings indicate that C. longa may contribute to the control of both seizures and the cell damage induced by PTZ, and that its association with diazepam may be a potentially effective option for the treatment of epilepsy in the future.
Epilepsy is one of the most common neurological disorders, affecting approximately 0.7% of the human population worldwide (Fiest et al., 2017). This condition occurs as a result of instability in both the inhibitory synaptic transmission in the brain, which reduces the transmission mediated by the GABA receptors, and the excitatory transmission, which increases the glutamatergic signaling. The available antiepileptic drugs (AEDs) thus act through two pathways, i.e., by either (1) potentiating the inhibitory mechanisms or (2) reducing the excitatory signaling (Sultana et al., 2021).
Although effective medication is available for the control of seizures, approximately one-third of the patients do not respond satisfactorily to the treatment, based on trials involving at least two different AEDs (either individually or in combination) that fail to impede seizures (of all types) in the patients (Sierra et al., 2015; Kalilani et al., 2018). Thus, it is essential to identify additional potential treatments that act on the underlying mechanisms that determine the seizures and have minimal side effects (Sultana et al., 2021).
Epilepsy can also cause neuronal damage in electrically sensitive regions, such as the hippocampus, which means that prolonged seizure activity may lead to increased production of reactive oxygen species, oxidative stress, and mitochondrial dysfunction, eventually leading to severe cerebral damage (Dillioglugil et al., 2010). The oxidative stress and mitochondrial dysfunction provoked by epilepsy disrupt the homeostasis of the intracellular environment, resulting in neuroexcitability and cell death. Oxidative stress damages the mitochondrial respiratory chain and leads to the excessive production of reactive oxygen species, which accumulates to the point of inhibiting the activity of the mitochondrial respiratory chain, eventually resulting in neurodegeneration (Chang and Yu, 2010).
One of the regions most affected by epilepsy is the hippocampus due to its electrical vulnerability. The sensitivity of the hippocampus is due to the presence of a large number of GABAergic neurons in the deeper regions of the dentate gyrus. Glutamate is the principal excitatory neurotransmitter in the hippocampus, and during periods of hyperexcitability, i.e., epilepsy, convulsions occur, which may result in cell death, primarily in the regions rich in glutamatergic receptors, such as the CA1 and CA3 regions (Casillas-Espinosa et al., 2020).
Curcumin is a principal biologically active compound extracted from Curcuma longa, which has been shown to be effective in the treatment of a number of organic disorders (Witkin and Li, 2013; Pricci et al., 2020). Previous studies in several countries have demonstrated the therapeutic value of C. longa as an antioxidant, anti-inflammatory agent, or gut microbiome modulator in in vitro, in vivo, and clinical trials in humans (Yu et al., 2013; Thumann et al., 2019; Rodrigues et al., 2021). Mehla et al. (2010) found that curcumin is effective as an anticonvulsant, with the potential to prevent seizures, reduce oxidative stress, and decrease brain damage. These findings indicate that curcumin may have potent antiepileptic effects, in particular by delaying the onset of seizures, although the exact mechanisms through which it achieves these results are still unclear (Mehla et al., 2010).
In this context, the present study evaluated the antiepileptic effects of C. longa in relation to brain activity in comparison with diazepam (DZP) and the combined treatment (C. longa + DZP), based on the decomposition of brain waves using electroencephalograms (EEGs) in a pentylenetetrazol (PTZ)-induced seizure model. The effects of the different treatments were also evaluated in terms of the results of an electromyogram (EMG) and histopathological changes in the hippocampus.
The present study used male Wistar rats (n = 72 animals) aged 10–12 weeks and weighing 260 g (± 20 g). These animals were housed in standard cages in a controlled environment (22 ± 2°C; 12/12 h light/dark cycle, 55 ± 10% relative humidity) with ad libitum access to food and water. The experimental procedures were approved by the relevant Brazilian federal agencies and were in accordance with the Brazilian National Council for the Control of Animal Experimentation and the Ethics Committee on Use of Animals of the Biological Sciences Institute at the Federal University of Pará (CEUA/UFPA no. 9149220321). The data presented here were also collected in compliance with the ARRIVE (Animal Research: Reporting In Vivo Experiments) guidelines. All necessary precautions were taken to prevent animal suffering and distress.
The animals in this study were maintained in the research facility for at least 7 days prior to the experiment, for adaptation and acclimation (Figure 1), with the electrodes being implanted in the cortex 1 day prior to the application of the treatments. During the experiment, the rats were pretreated for 4 days with either saline, C. longa, DZP, or C. longa + DZP via the orogastric route (gavage) at 24-h intervals. On the fifth day (24 h after the last application), seizures were induced by a single dose of PTZ, intraperitoneally (Agarwal et al., 2013), with electroencephalographic and EMG records being collected over the subsequent 15 min. The rats were monitored for the subsequent 7 days (follow-up) prior to being euthanized. The brain was then extracted, sectioned, and stained with cresyl violet for cell counting. All these procedures were conducted strictly between 08:00 and 11:00 a.m.
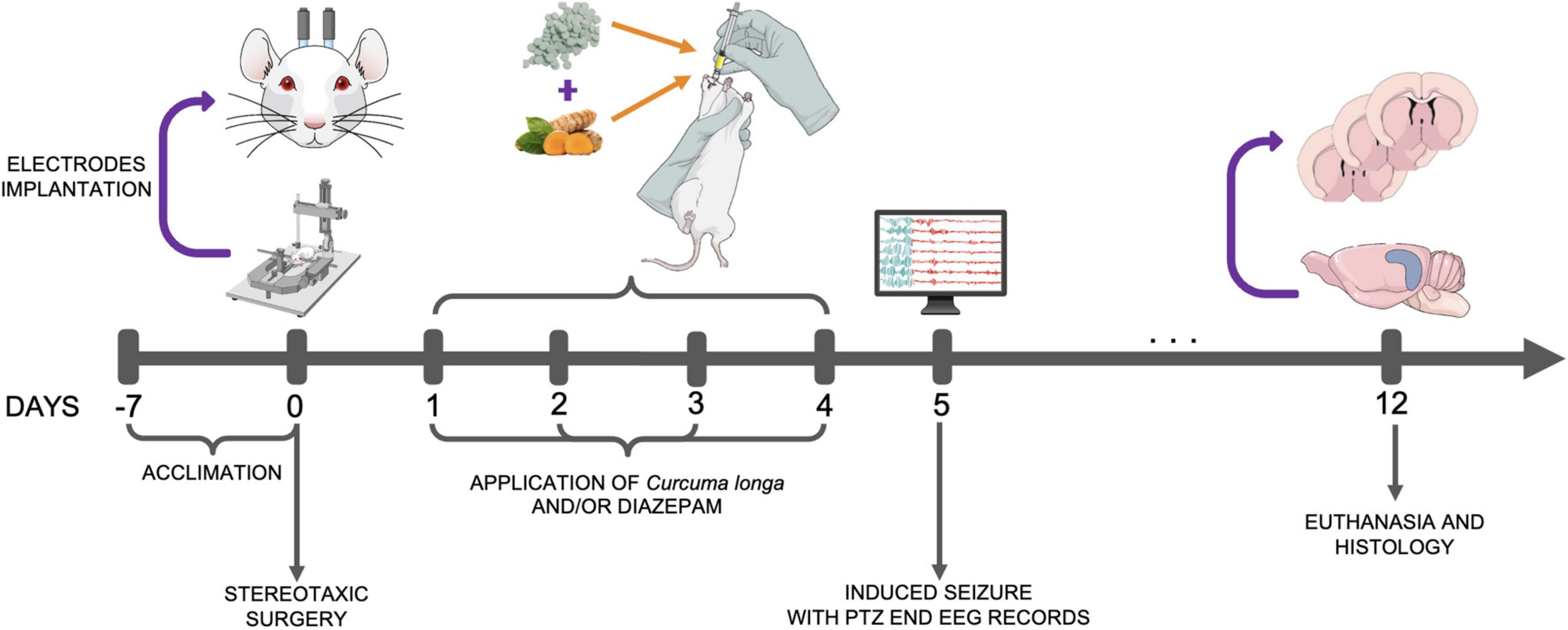
Figure 1. Experimental design. PTZ, pentylenetetrazol; EEG, electroencephalograph.
The animals were divided into eight groups (each containing nine animals): (i) saline + saline (SAL); (ii) C. longa (CL) + saline; (iii) DZP + saline; (iv) CL/DZP + saline; (v) SAL + PTZ; (vi) CL + PTZ; (vii) DZP + PTZ; (viii) CL/DZP + PTZ. The seizure behavior was recorded after the application of PTZ.
In addition to the two drugs evaluated in the present study, three types of anesthetics were also used for handling the rats. C. longa was applied in the form of purified pharmaceutical grade curcumin, supplied as 250-mg capsules containing 50 mg of curcumin together with excipients (Aché Laboratórios Farmacêuticos S.A., Brazil). The curcumin was administered via oral gavage at a dose of 80 mg/kg (Akula and Kulkarni, 2014); while the DZP, 10 mg/2 ml (União Química, Embu-Guaçu, SP, Brazil), was administered at a dose of 5 mg/kg (V.O.). Three different types of anesthetic were used in the present study. Ketamine hydrochloride was obtained from the Köing Laboratory (Santana de Parnaíba, SP, Brazil) and xylazine hydrochloride was acquired from the Vallée Laboratory (Montes Claros, MG, Brazil), while the local anesthetic lidocaine, which was used to implant the electrodes, was obtained from the Hipolabor Laboratory (Sabará, MG, Brazil). The PTZ was obtained from Sigma Chemical Co. (St. Louis, MO, United States).
The EEGs were recorded as described by Estumano et al. (2019). For this, the animals were anesthetized and placed in a stereotaxic apparatus for the implantation of stainless-steel electrodes (exposed tip 1.0 mm in diameter) on the dura mater above the pre-frontal cortex at the bregma coordinates − 0.96 mm and ± 1.0 mm lateral, and were fixed with dental acrylic cement. The data were registered with the help of the electrodes using a digital data acquisition system composed of a high impedance amplifier (Grass Technologies, P511, United States), an oscilloscope (Protek, 6510, United States), and a data acquisition and digitalization board (National Instruments, Austin, TX, United States). Data were collected continuously at 1 kHz, at a low pass of 3 kHz, and a high pass of 0.3 Hz. During the recording sessions, the animals were confined to acrylic boxes (20 cm × 45 cm × 15 cm), and the EEG activity was recorded for 15 min immediately after the application of the PTZ or saline solution. The records collected using the digital data acquisition system were analyzed offline. The analyses were run at frequencies of up to 40 Hz, and then split into four bands, that is, the delta (1–4 Hz), theta (4–8 Hz), alpha (8–12 Hz), beta (12–28 Hz), and gamma (28–40 Hz) bands (Aminov et al., 2017).
The behavior of the animals was monitored during the seizures and compared with the latency patterns of the behaviors observed in the PTZ group. Latency was measured in relation to the onset of the following behaviors: (i) generalized tremor; (ii) spasms of the forelimbs; (iii) isolated clonic seizures with no loss of the posture reflex; (iv) generalized clonic seizures with transient loss of the posture reflex; and (v) tonic-clonic seizures with total loss of the posture reflex.
Electrodes were implanted in parallel to the masseter muscle, 5 mm above their point of insertion in the jaw to record the muscle activity during seizures in the PTZ groups. As for the EEG, the data were recorded for 15 min (Santos et al., 2021).
Once euthanized, the rats were perfused transcardially with phosphate-buffered saline (PBS, pH 7.4) at 4°C, followed by 4% formaldehyde (pH 7.4). The brain was extracted post-reperfusion, fixed in 4% formaldehyde for 72 h, cryoprotected in 30% sucrose for 24 h, and then cut into serial coronal sections (40 μm) and stained with Nissl (0.3% cresyl violet acetate). The number of cells of six coronal sections of the hippocampus (CA1 and CA3) of each rat were counted to provide a mean count for each group (n = 9 rats per group). The cell counts were based on the inspection of a field of 50 μm × 50 μm in each region (Wang et al., 2021). The counted (treatment-blind) cells were observed using the ImageJ digital imaging software (NIH, Bethesda, MD, United States).
The normality of the data variances was verified using the Kolmogorov–Smirnov test. All the data are presented as the mean and standard deviation (SD), and the F and p-values are included where pertinent. A p < 0.05 significance level was considered for all the analyses. The significance of differences between the pairs of groups was verified using Student’s t-test, while the variation among three or more groups was evaluated using an Analysis of Variance (ANOVA), either one-way or two-way, followed by Tukey’s test for pairwise multiple comparisons. The analyses were run in GraphPad Prism, version 9 (Graph-Pad Software Inc., San Diego, CA, United States).
The behavior of the rats was assessed to determine the evolution of the seizures (Table 1). The animals pretreated with saline that received PTZ progressed quickly to tonic-clonic seizures with the loss of the postural reflex after a mean interval of less than 5 min. Latency prior to the onset of seizure increased significantly in the group pretreated with C. longa, although the evolution to tonic-clonic seizure with loss of postural reflex was not interrupted.
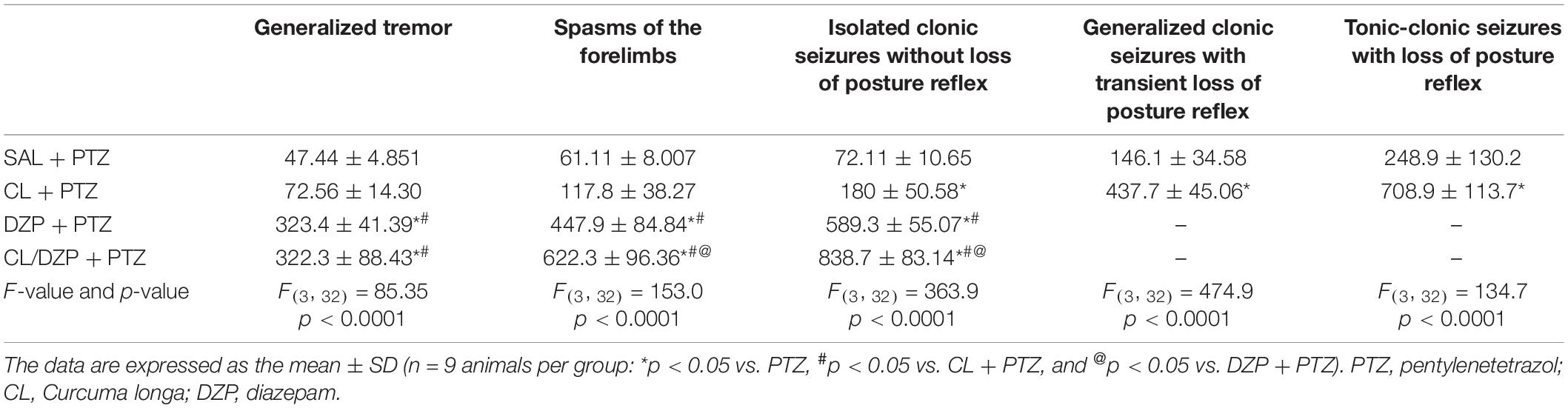
Table 1. Description of the seizure-related behavior of animals treated with Curcuma longa and/or diazepam.
The group pretreated with DZP that received PTZ presented a greater latency to the onset of seizures in comparison with the C. longa group, in addition to the stabilization of the symptoms, given that the rats presented only isolated clonic seizures with no loss of the postural reflex. Surprisingly, the combined pretreatment (C. longa + DZP) resulted in even better control of the seizure-related behavior, in comparison with the DZP + PTZ group, with the greatest latency to the onset of the spasms and only isolated clonic seizures. These results indicate that the association of C. longa and DZP may provide effective control and prevent the evolution of the seizure.
The EEGs were first obtained from the four saline groups (i–iv), that is, the animals that were pretreated with saline, C. longa, DZP, and C. longa + DZP, respectively, and then received saline on the fifth day (+ SAL). This provided a baseline for the verification of the possible effects of the pretreatment on brain activity. The animals pretreated with saline (group i) had amplitudes below 0.02 mV (Figure 2A), and the spectrogram reveals energy concentrations of below 10 Hz. None of the animals of the other groups (ii–iv) presented any significant difference in the brain activity (Figures 2B–D) in comparison with the control (i), which indicates that none of the pretreatments alter this activity.
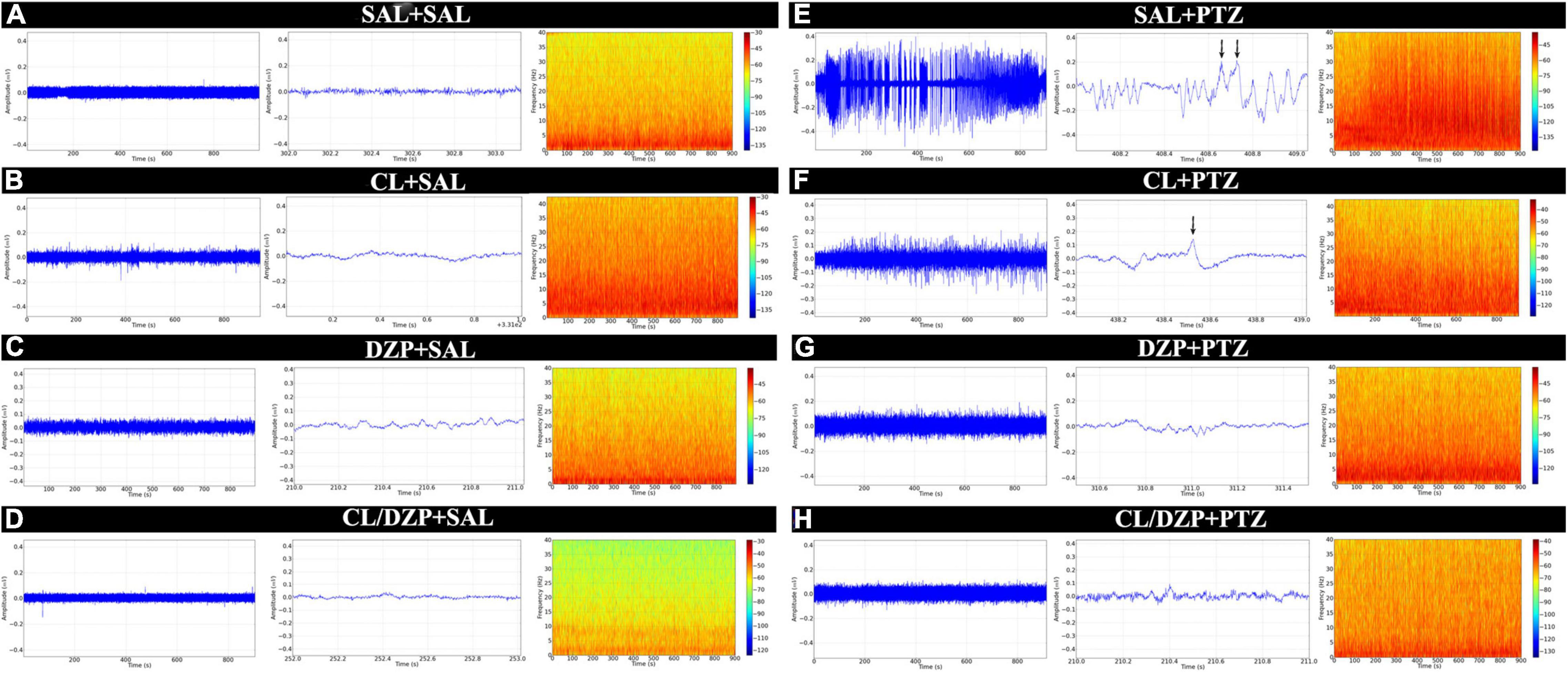
Figure 2. Electroencephalographic recordings of animals submitted to the PTZ-induced seizure model and treated with Curcuma longa and/or diazepam. The linear trace was obtained by electroencephalography (left). Representative 1 s sequence of the EEG trace (center). Spectrogram frequency (right). (A) Animals that received no treatment (SAL-SAL). (B) Animals pretreated with Curcuma longa but not injected with PTZ (CL-SAL). (C) Animals pretreated with diazepam but not injected with PTZ (DZP-SAL). (D) Animals pretreated with Curcuma longa + diazepam not injected with PTZ (CL/DZP-SAL). (E) Animals pretreated with saline and injected with PTZ (SAL-PTZ). (F) Animals pretreated with Curcuma longa and injected with PTZ (CL-SAL). (G) Animals pretreated with diazepam and injected with PTZ (DZP-PTZ). (H) Animals pretreated with Curcuma longa + diazepam and injected with PTZ (CL/DPZ-PTZ). SAL, saline; CL, Curcuma longa; DZP, diazepam; PTZ, pentylenetetrazol.
In contrast, group v (SAL + PTZ) presented significant changes in the EEG trace, with peaks of amplitude of over 0.3 mV, and activity characterized by constant levels of spike-waves with a high frequency and amplitude (black arrow, Figure 2E). In group vi (CL + PTZ), amplitude varied up to 0.2 mV, while the frequency and amplitude of the spike-waves decreased (black arrow, Figure 2F). In group vii (DZP + PTZ), the changes in the EEG trace were less intense than in groups v and vi, that is, close to 0.1 mV (Figure 2G), which indicates control of the seizure. Finally, the combined pretreatment (CL/DZP + PTZ) obtained the best results in terms of seizure control, with an amplitude of 0.08 mV, and a reduction in the frequency and amplitude of the spike-waves (Figure 2H).
In addition, the total power did not vary significantly among the saline groups, i.e., groups i–iv [F(3, 32) = 0.2671; p = 0.8486; Figure 3A]. The administration of PTZ to the saline group (v) resulted in a significant increase in the total power in comparison with group i (SAL + SAL: 0.1985 ± 0.0740 mV2/Hz × 10–3 vs. SAL + PTZ: 5.509 ± 0.9856 mV2/Hz × 10–3; p < 0.0001; Figure 3B). Significant variation [F(3, 32) = 78.75; p < 0.0001; Figure 3C] was also found among the other PTZ groups (vi–viii), with all the different pretreatments reducing the total power of the PTZ-induced seizures. The mean total power of group vi (CL + PTZ) was 2.942 ± 0.5694 mV2/Hz × 10–3, which was significantly lower (p < 0.0001) than the PTZ group (v). The mean total power of group vii (DZP + PTZ) was 2.066 ± 0.2846 mV2/Hz × 10–3, significantly lower than that recorded for either group v (p < 0.0001: DZP + PTZ vs. SAL + PTZ) or vi (p = 0.0239: DZP + PTZ vs. CL + PTZ). However, the combined treatment (CL/DZP) resulted in the lowest total power of all (1.348 ± 0.3624 mV2/Hz × 10–3), which was significantly lower than that recorded for the groups v–vii (p < 0.0001 in both cases), indicating that this treatment is the most effective one for the control of PTZ-triggered seizures.
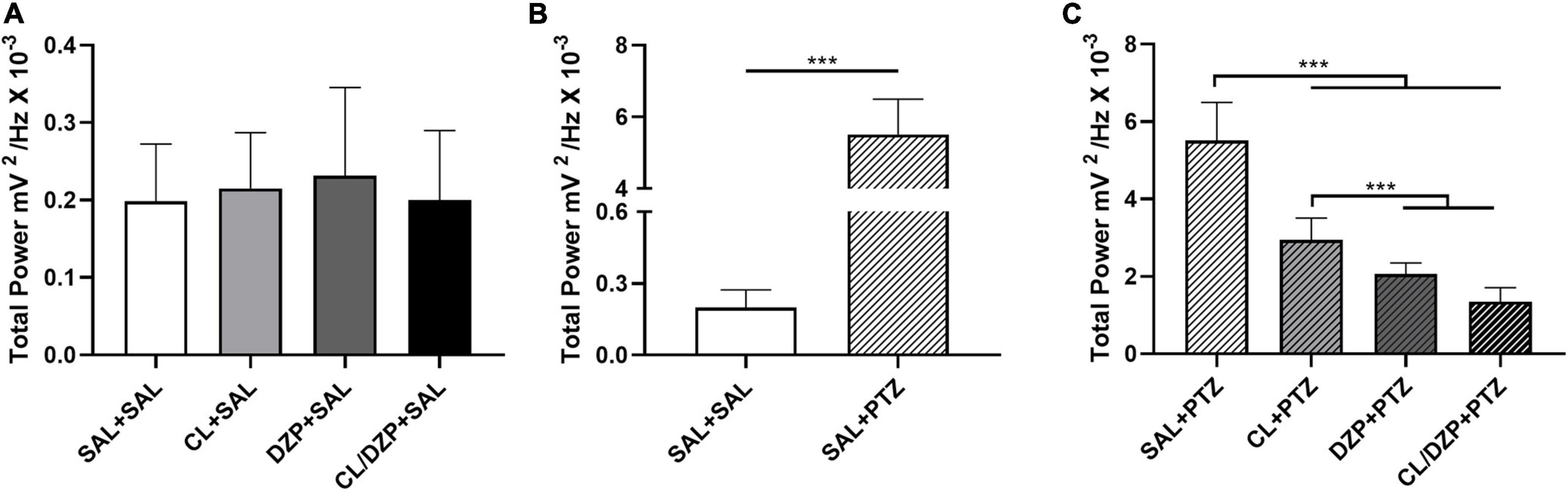
Figure 3. Total linear brainwave power recorded by electroencephalography. (A) Quantitative distribution of the total linear power of the brainwaves of the animals that received saline on the fifth day. (B) Quantitative distribution of the animals that were pretreated with saline and received saline or pentylenetetrazol on the fifth day. (C) Quantitative distribution of the total linear power of the brainwaves of the animals that received pentylenetetrazol on the fifth day. The data are expressed as the mean ± SD (n = 9 per group); ***p < 0.001. SAL, saline; CL, Curcuma longa; DZP, diazepam; PTZ, pentylenetetrazol.
The decomposition of the brainwaves was analyzed only for the PTZ groups (v–viii). In the case of the low-frequency waves, a significant increase (group v) was recorded in the bandpower of the delta waves [F(3, 32) = 80.49; p < 0.0001; Figure 4A]. The animals that received PTZ presented brainwave patterns consistent with disorganized brain activity. However, pretreatment with C. longa (group vi) attenuated the effects of PTZ significantly (p < 0.0001 vs. SAL + PTZ), which indicates beneficial properties for the reduction of seizures. The attenuation of the delta waves in the two DZP groups (vii and viii) was also significantly greater in comparison with the animals pretreated only with C. longa (p < 0.0001 for DZP + PTZ and CL/DZP + PTZ vs. CL + PTZ).
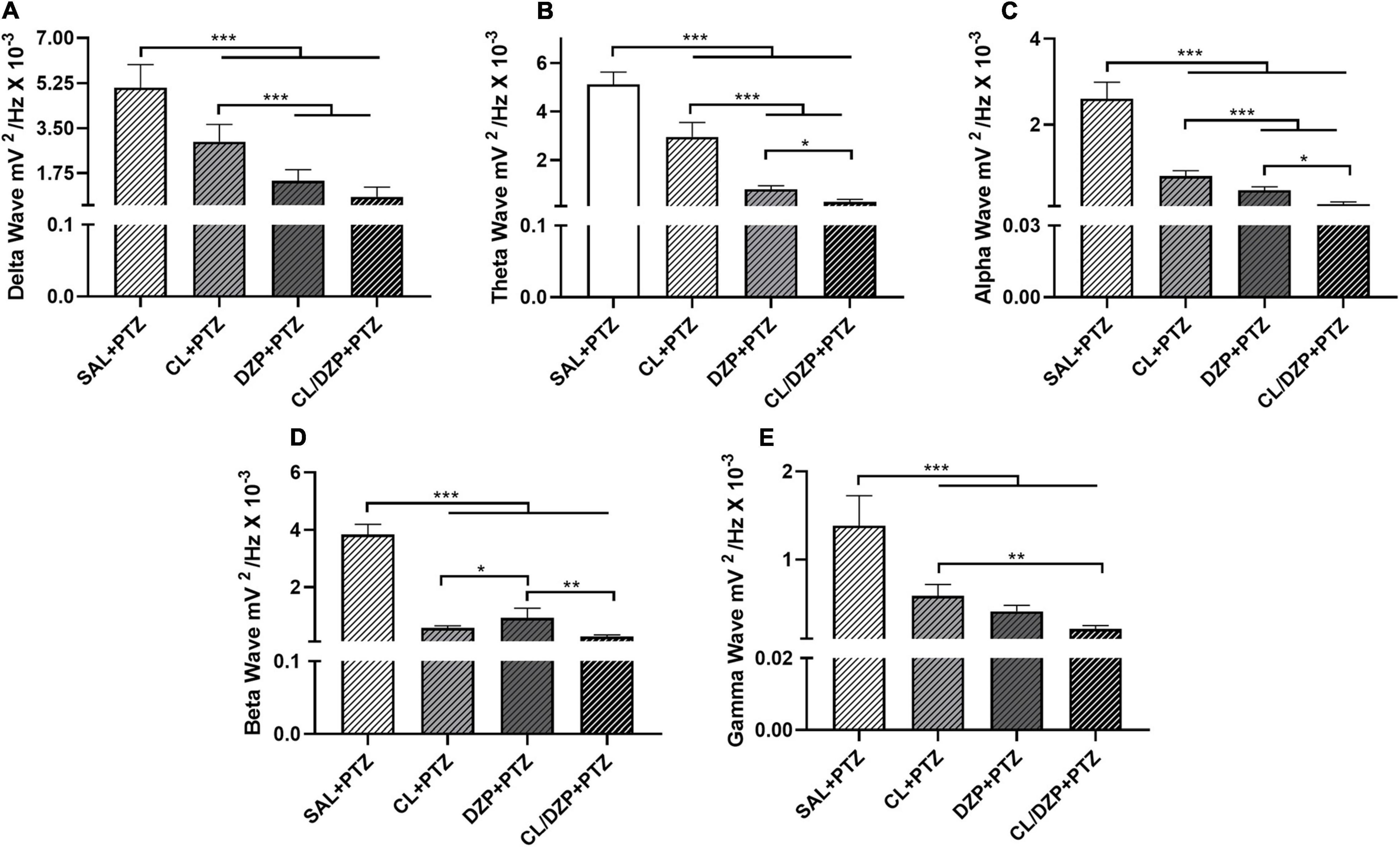
Figure 4. Relative bandpower of the brainwaves (1–40 Hz) of the animals that received pentylenetetrazol on the fifth day (PTZ groups). Quantitative electroencephalographic data on the relative bandpower of the (A) delta waves; (B) theta waves; (C) alpha waves; (D) beta waves; (E) gamma waves. The data are expressed as the mean ± SD (n = 9 animals per group); *p < 0.05, **p < 0.01, ***p < 0.001. SAL, saline; CL, Curcuma longa; DZP, diazepam; PTZ, pentylenetetrazol.
A similar pattern was observed in the case of the theta waves (Figure 4B), with significant attenuation in the pretreatment groups (vi–viii) in comparison with the saline + PTZ group [F(3, 32) = 275.0; p < 0.0001]. While C. longa alone significantly mitigated the effects of PTZ on the brain (p < 0.0001), the pretreatment with DZP (groups vii and viii) was significantly more effective (p < 0.0001).
The administration of PTZ also altered the alpha bandpower (Figure 4C) significantly [F(3, 32) = 244.8; p < 0.0001]. While all the different pretreatments had significant beneficial effects (p < 0.0001, for all comparisons with SAL + PTZ), CL/DZP was the most effective (CL + PTZ vs. CL/DZP + PTZ: p < 0.0001; DZP + PTZ vs. CL/DZP + PTZ: p = 0.0134), which indicates, once again, that the combination of C. longa and DZP provides better control than each compound on its own.
Similar results were also obtained for the higher-frequency waves, that is, the beta and gamma waves. As in the case of the alpha wave, all the different pretreatments had a positive effect on the beta wave (Figure 4D), thus reducing the oscillations significantly [F(3, 32) = 388.0; p < 0.0001]. In this case, however, the C. longa treatments (CL or CL/DZP) reduced bandpower significantly more than the group that was pretreated only with DZP (CL + PTZ vs. DZP + PTZ: p = 0.0264; CL/DZP + PTZ vs. DZP + PTZ: p < 0.01). This indicates that C. longa may be especially beneficial for seizure control, in terms of the mechanisms that trigger the beta waves.
A similar pattern was recorded in the case of the gamma wave (Figure 4E), where all the pretreatments reduced the gamma wave bandpower significantly [F(3, 32) = 69.62; p < 0.0001]. While the combined application of C. longa and DZP provided a better control for seizures than C. longa alone (p = 0.0008), it was no different from pure DZP (p = 0.1202).
As orofacial movement (chewing) is a diagnostic trait of PTZ-induced seizures, conjugated electrodes were implanted in the masseter muscle to evaluate its activity during the seizures. Following the application of PTZ, the seizures caused intense muscle contractions, with oscillations in amplitude of up to 0.5 mV in the electromyographic trace (Figure 5A). However, pretreatment with C. longa relieved the muscle contractions during the seizure, as revealed by a reduction and stabilization of this amplitude, and a significant reduction in the total power [F(3, 32) = 93.35; p < 0.0001; SAL + PTZ vs. CL + PTZ: p = 0.0003; Figure 5B]. The use of DZP also resulted in intense myorelaxation, with a significant reduction in the total power in the EMG (Figure 5B; p < 0.0001). In the combined pretreatment (CL/DZP), the trace was less altered (Figure 5A), with significant improvement in the EMG in comparison with the SAL + PTZ group (Figure 5B, p < 0.0001), but not in comparison with the group treated with DZP alone (p = 0.8498).
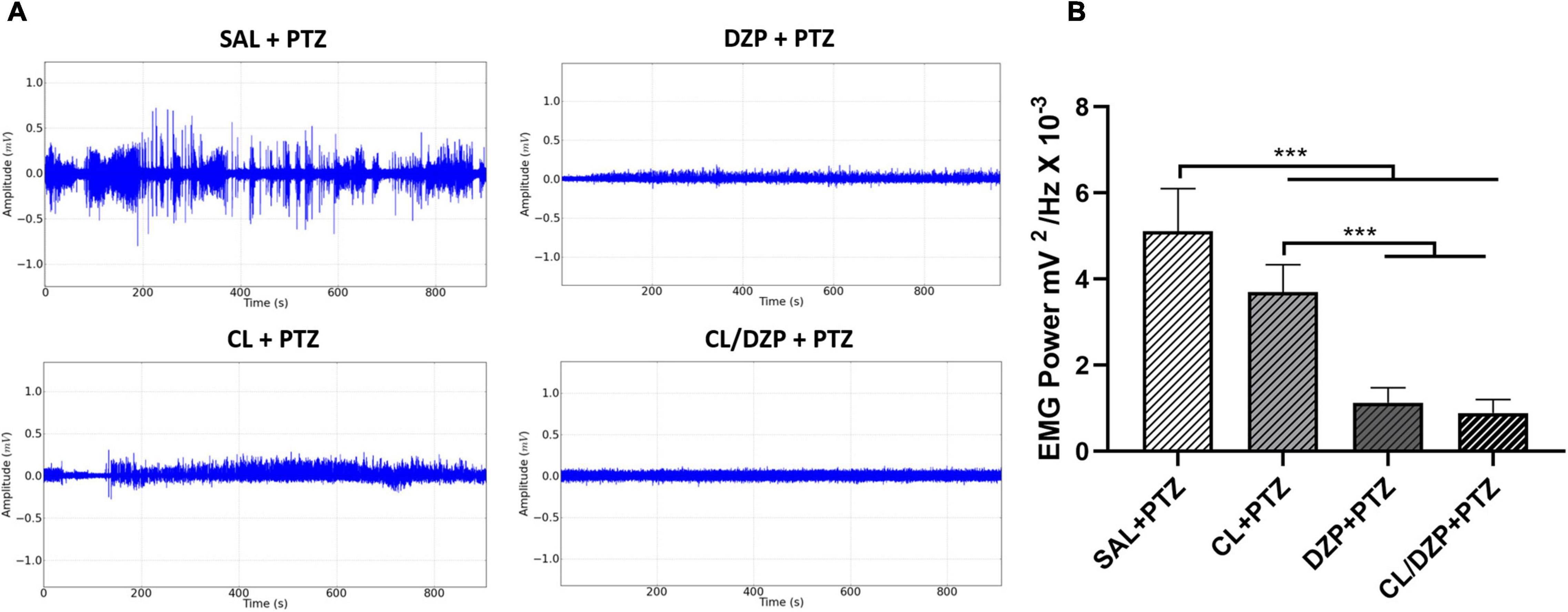
Figure 5. Surface electromyographic recordings of PTZ-induced seizures pretreated with Curcuma longa and/or diazepam. (A) Electromyographic linear trace. (B) Quantitative linear distribution of the power of the muscle contraction. The data are expressed as the mean ± SD (n = 9 per group); ***p < 0.001. SAL, saline; CL, Curcuma longa; DZP, diazepam; PTZ, pentylenetetrazol.
The quantification of the Nissl-stained neuron-like cells in the CA1 and CA3 regions of the hippocampus (Figure 6A) indicated that the animals pretreated with saline that received PTZ on the fifth day suffered a significant reduction in the number of neuron-like cells in the CA1 region [F(7, 64) = 226.9; p < 0.0001; SAL + (SAL and CL and DZP and CL + DZP) vs. SAL + PTZ: p < 0.0001; Figure 6B]. Although pretreatment with both C. longa (SAL + PTZ vs. CL + PTZ: p < 0.0001) and DZP (SAL + PTZ vs. CL + PTZ: p < 0.0001) resulted in the significant preservation of these cells, and C. longa performed significantly better than DZP (p < 0.0001), the combination of these two substances (CL/DZP) was the best pretreatment for the preservation of the neuron-like cells (SAL + PTZ vs. CL/DZP + PTZ: p < 0.0001; CL/DZP + PTZ vs. CL + PTZ: p < 0.0001; CL/DZP + PTZ vs. CL + PTZ: p < 0.0001). This indicates a protective effect on the cells in the CA1 region of the hippocampus 7 days after the seizures.
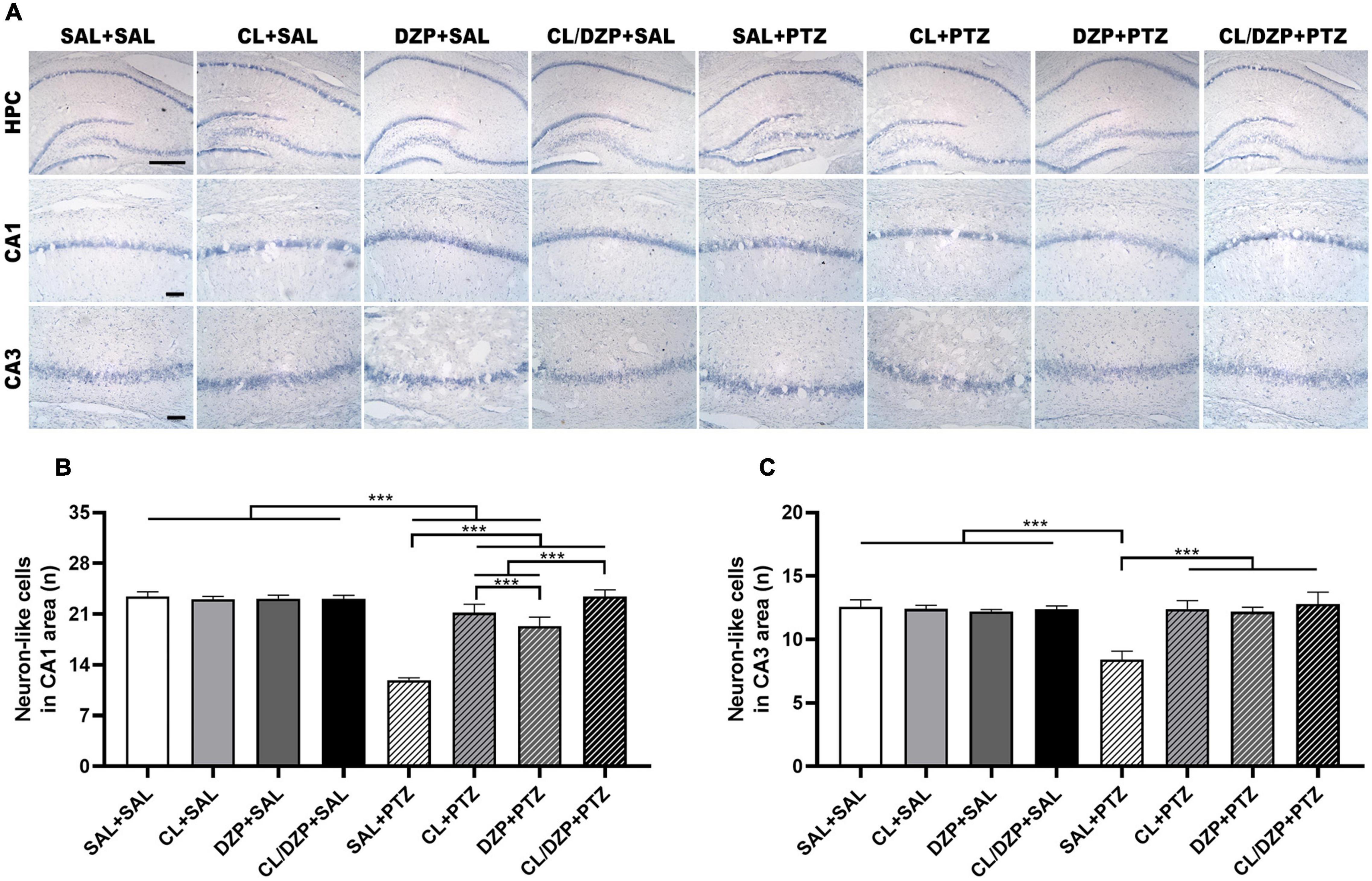
Figure 6. Nissl staining of the rat hippocampus in PTZ-induced seizures pretreated with Curcuma longa and/or diazepam. (A) Representative images. (B) Quantitative data on the number of neuron-like cells in the CA1 region. (C) Quantitative data for the number of neuron-like cells in the CA3 region. Data are presented as the mean ± SD (n = 9 per group); ***p < 0.001. SAL, saline; CL, Curcuma longa; DZP, diazepam; PTZ, pentylenetetrazol; HPC, hippocampus. Scale bar = 200 μm. CA1 and CA3: scale bar = 50 μm.
In the case of the CA3 region of the hippocampus (Figure 6C), a significant change was observed only in the saline group that received PTZ on the fifth day [F(7, 64) = 62.69; p < 0.0001], which indicates that a lack of adequate preventive treatment for seizures may result in the damage of this layer. All the pretreatments tested in the present study provided significant prevention of the loss of the neuron-like cells (p < 0.001; for all comparisons).
The results of the present study demonstrate that C. longa has anticonvulsant properties that are effective for the attenuation of PTZ-induced seizures. The data also showed that the combined application of C. longa with DZP decreased the seizure threshold and prevented the behavioral progression of the seizure, while also reducing the neuronal damage it causes.
Epilepsy is one of the most common disorders of the central nervous system that, when treated incorrectly or when the patient is resistant to the available medication, can impact the quality of life significantly (Sultana et al., 2021). Previous studies have shown that the recurrence of epileptic events may eventually have major degenerative effects that are also associated with a cognitive and behavioral decline. Some antiepileptic drugs may also cause harmful changes in the brain, which reinforces the need for the development of new treatments that can reduce the brain damage and minimize the side effects.
C. longa is widely cultivated in Asia, where it is a part of the traditional approach for the treatment of a variety of health problems, including gastrointestinal disorders, pain, and even epilepsy (Touhidi et al., 2018). While its mechanism of action is still unclear, some studies have found evidence of the modulation of the GABA receptors, which increases the synthesis of this neurotransmitter, reduces the activity of acetylcholinesterase, and inhibits the catecholaminergic and oxidative stress mechanisms (Aboul Ezz et al., 2011; Reeta et al., 2011; Vijayakumar et al., 2018). Other studies have also shown that C. longa reduces the activity of the glutamate receptors and contributes to the intracellular homeostasis of calcium (Noor et al., 2012).
Despite the evidence of its protective effects, the therapeutic potential of curcumin is limited by its poor bioavailability, given its reduced absorption and limited passage through the blood–brain barrier (BBB) (Tsai et al., 2011). These authors demonstrated that purified curcumin crosses the BBB at lower concentrations than when transported by nanoparticles. Technologies that facilitate the transport of curcumin to the brain, including nanocarriers and polymeric nanoparticles, are currently under investigation (Tsai et al., 2011; Askarizadeh et al., 2020). Given this, one of the limitations of the present study is the lack of the definition of the amount of curcumin that crossed the BBB.
The present study showed that pretreatment for 4 days with C. longa alone or in combination with DZP was able to reduce the duration of seizures. Saha et al. (2016) and Haghighizad et al. (2017) obtained similar results showing that treatment for at least 2 weeks with a minimum dose of 100 mg/kg of C. longa delayed the onset time and duration of tonic-clonic PTZ-induced seizures. Other studies have also corroborated these findings. Mehla et al. (2010) showed that curcumin caused a significant increase in the latency to the onset of seizures and reduced the mortality caused by the seizures induced by the repeated administration of a subconvulsant dose of PTZ. The present study obtained similar results through pretreatment with curcumin, even after the administration of only one dose of PTZ. This indicates that pretreatment or continuous treatment with curcumin may help shorten the duration of seizures, and it may be represented as a potential option for the treatment of epilepsy.
Some previous studies have also shown that the combination of C. longa with other antiepileptic drugs, such as sodium valproate, at a lower dose, may have a similar effect to the drug when administered alone (Aboul Ezz et al., 2011; Reeta et al., 2011; Noor et al., 2012). The present study showed that C. longa associated with DZP elicited a better response than either drug administered alone. These findings are extremely important, because the combination of C. longa with an AED may permit the reduction of the dose, which may, in turn, reduce its side effects.
The EEG trace of the seizures induced by PTZ had an amplitude of 0.3 mV, with high amplitude spike-waves, which were attenuated by the administration of C. longa. Orellana-Paucar et al. (2012) and Jiang et al. (2015) obtained similar results in which the use of curcumin, the principal biologically active component of C. longa, reduced the abnormal brain activity induced by the seizure. It is important to note that PTZ-induced seizures that can be extremely harmful and can cause hippocampal damage, especially in the CA1 and CA3 regions, may result in short- or long-term cognitive deficits (Kaur et al., 2014; Hashemian et al., 2017). The present study showed that the pretreatment, either with C. longa alone or in combination with DZP, reduces the damage in the hippocampus of rats, which is consistent with the previous studies that have demonstrated the potential protective properties of this substance.
It is interesting to note that epileptiform activity can be observed in almost 100% of surface EEGs, which can thus be used to predict possible brain injuries (Janszky et al., 2005; Singla et al., 2020). Although only a single pair of electrodes was used in the present study, which may be vulnerable to the influence of early motor signals, as well as the scalp and cerebrospinal fluid (Beleza and Pinho, 2011), which may limit spatial accuracy in comparison with multichannel systems, Johnstone et al. (2012) and Hemington and Reynolds (2014) validated this approach for EEG recording and diagnosis.
An increase in the delta and beta bandpower may reflect electrical alterations in the temporal and extratemporal lobes (Rosenow et al., 2015), and may also be present in other vascular diseases of the central nervous system (Ferreira et al., 2021). The present study showed that all three pretreatments (C. longa, DZP, or C. longa/DZP) reduced the bandpower of the delta and beta waves, which indicates that the seizure was controlled and brain damage was reduced. This indicates that C. longa may play a protective role, in particular, in the cells of the hippocampus that are highly sensitive to electrical and inflammatory disorders, and may become atrophied moderately or severely if left untreated.
Even so, the exact mechanisms through which the anti-inflammatory properties of C. longa are implemented are still unknown, although some authors have reported that it upregulates genes related to the anti-inflammatory cytokines and reduces the expression of pro-inflammatory cytokines, such as IL-1β and TNF-α (Hashemian et al., 2017; Yin et al., 2018). One other potential mechanism, described by Peng et al. (2021), is the inhibition of the expression of the iNOS gene by C. longa that interferes with the nitric oxide synthase pathway. Other studies have demonstrated the potential of C. longa for the protection of the hippocampal cells against electrical disturbances (Kaur et al., 2015; Hashemian et al., 2017), which is consistent with the findings of the present study, given the observed attenuation of the damage caused by PTZ in the CA1 and CA3 regions. These authors have also reported that C. longa inhibits the activation of astrocytes and microglia during electrical disturbances (Kaur et al., 2015; Hashemian et al., 2017).
Overall, the results of the present study indicate that C. longa has considerable potential for the control of the seizures and cell damage induced by PTZ, and that the association of this substance with DZP may represent a valuable approach for the treatment of epilepsy, thereby increasing the therapeutic options available to the patients. However, further research will be needed to better define the signaling pathways that determine the protective properties of C. longa.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The animal study was reviewed and approved by the Ethics Committee on Use of Animals.
CN and LF performed the experiment and drafted the manuscript. ALMS, ABNS, JR, and LT conducted the bioinformatic analysis and interpreted the results. JA, DA, AH, BG, and BC performed the histological analyses. MH and DL reviewed and edited the manuscript. All authors contributed to the manuscript revision, and read and approved the submitted version.
This work was partly funded by PROPESP-UFPA.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
We are grateful to João Neto for his help with the photographs of the hippocampus.
Aboul Ezz, H. S., Khadrawy, Y. A., and Noor, N. A. (2011). The neuroprotective effect of Curcumin and Nigella sativa oil against oxidative stress in the pilocarpine model of epilepsy: a comparison with valproate. Neurochem. Res. 36, 2195–2204. doi: 10.1007/s11064-011-0544-9
Agarwal, N. B., Jain, S., Nagpal, D., Agarwal, N. K., Mediratta, P. K., and Sharma, K. K. (2013). Liposomal formulation of curcumin attenuates seizures in different experimental models of epilepsy in mice. Fundam. Clin. Pharmacol. 27, 169–172. doi: 10.1111/j.1472-8206.2011.01002.x
Akula, K. K., and Kulkarni, S. K. (2014). Effect of curcumin against pentylenetetrazol-induced seizure threshold in mice: possible involvement of adenosine A 1 receptors. Phyther. Res. 28, 714–721. doi: 10.1002/ptr.5048
Aminov, A., Rogers, J. M., Johnstone, S. J., Middleton, S., and Wilson, P. H. (2017). Acute single channel EEG predictors of cognitive function after stroke. PLoS One 12:e0185841. doi: 10.1371/journal.pone.0185841
Askarizadeh, A., Barreto, G. E., Henney, N. C., Majeed, M., and Sahebkar, A. (2020). Neuroprotection by curcumin: a review on brain delivery strategies. Int. J. Pharm. 585, 119476. doi: 10.1016/j.ijpharm.2020.119476
Beleza, P., and Pinho, J. (2011). Frontal lobe epilepsy. J. Clin. Neurosci. 18, 593–600. doi: 10.1016/j.jocn.2010.08.018
Casillas-Espinosa, P. M., Ali, I., and O’Brien, T. J. (2020). Neurodegenerative pathways as targets for acquired epilepsy therapy development. Epilepsia Open 5, 138–154. doi: 10.1002/epi4.12386
Chang, S.-J., and Yu, B.-C. (2010). Mitochondrial matters of the brain: mitochondrial dysfunction and oxidative status in epilepsy. J. Bioenerg. Biomembr. 42, 457–459. doi: 10.1007/s10863-010-9317-4
Dillioglugil, M. O., Kir, H. M., Demir, C., Ilbay, G., Sahin, D., Dillioglugil, O., et al. (2010). Effect of pentylenetetrazole and sound stimulation induced single and repeated convulsive seizures on the MDA, GSH and NO levels, and SOD activities in rat liver and kidney tissues. Brain Res. Bull. 83, 356–359. doi: 10.1016/j.brainresbull.2010.09.007
Estumano, D. P., Ferreira, L. O., Bezerra, P. A. L., da Silva, M. C. P., Jardim, G. C., Santos, G. F. S., et al. (2019). Alteration of testosterone levels changes brain wave activity patterns and induces aggressive behavior in rats. Front. Endocrinol. (Lausanne) 10:654. doi: 10.3389/fendo.2019.00654
Ferreira, L. O., Mattos, B. G., Jóia de Mello, V., Martins-Filho, A. J., da Costa, E. T., Yamada, E. S., et al. (2021). Increased relative delta bandpower and delta indices revealed by continuous qEEG monitoring in a rat model of ischemia-reperfusion. Front. Neurol. 12:645138. doi: 10.3389/fneur.2021.645138
Fiest, K. M., Sauro, K. M., Wiebe, S., Patten, S. B., Kwon, C.-S., Dykeman, J., et al. (2017). Prevalence and incidence of epilepsy. Neurology 88, 296–303. doi: 10.1212/WNL.0000000000003509
Haghighizad, H., Touhidi, A., Pourmotabbed, A., Moradpour, F., Nedaei, S. E., and Pourmotabbed, T. (2017). Curcumin improves chronic stress induced potentiated seizure activity in experimental model of epilepsy. J. Neurol. Sci. 34, 76–85. doi: 10.1016/j.pbb.2014.08.001
Hashemian, M., Anissian, D., Ghasemi-Kasman, M., Akbari, A., Khalili-Fomeshi, M., Ghasemi, S., et al. (2017). Curcumin-loaded chitosan-alginate-STPP nanoparticles ameliorate memory deficits and reduce glial activation in pentylenetetrazol-induced kindling model of epilepsy. Prog. Neuro Psychopharmacol. Biol. Psychiatry 79, 462–471. doi: 10.1016/j.pnpbp.2017.07.025
Hemington, K. S., and Reynolds, J. N. (2014). Electroencephalographic correlates of working memory deficits in children with Fetal Alcohol Spectrum Disorder using a single-electrode pair recording device. Clin. Neurophysiol. 125, 2364–2371. doi: 10.1016/j.clinph.2014.03.025
Janszky, J., Hoppe, M., Clemens, Z., Janszky, I., Gyimesi, C., Schulz, R., et al. (2005). Spike frequency is dependent on epilepsy duration and seizure frequency in temporal lobe epilepsy. Epileptic Disord. 7, 355–359.
Jiang, Z., Guo, M., Shi, C., Wang, H., Yao, L., Liu, L., et al. (2015). Protection against cognitive impairment and modification of epileptogenesis with curcumin in a post-status epilepticus model of temporal lobe epilepsy. Neuroscience 310, 362–371. doi: 10.1016/j.neuroscience.2015.09.058
Johnstone, S. J., Blackman, R., and Bruggemann, J. M. (2012). EEG from a single-channel dry-sensor recording device. Clin. EEG Neurosci. 43, 112–120. doi: 10.1177/1550059411435857
Kalilani, L., Sun, X., Pelgrims, B., Noack-Rink, M., and Villanueva, V. (2018). The epidemiology of drug-resistant epilepsy: a systematic review and meta-analysis. Epilepsia 59, 2179–2193. doi: 10.1111/epi.14596
Kaur, H., Bal, A., and Sandhir, R. (2014). Curcumin supplementation improves mitochondrial and behavioral deficits in experimental model of chronic epilepsy. Pharmacol. Biochem. Behav. 125, 55–64. doi: 10.1016/j.pbb.2014.08.001
Kaur, H., Patro, I., Tikoo, K., and Sandhir, R. (2015). Curcumin attenuates inflammatory response and cognitive deficits in experimental model of chronic epilepsy. Neurochem. Int. 89, 40–50. doi: 10.1016/j.neuint.2015.07.009
Mehla, J., Reeta, K. H., Gupta, P., and Gupta, Y. K. (2010). Protective effect of curcumin against seizures and cognitive impairment in a pentylenetetrazole-kindled epileptic rat model. Life Sci. 87, 596–603. doi: 10.1016/j.lfs.2010.09.006
Noor, N. A., Aboul Ezz, H. S., Faraag, A. R., and Khadrawy, Y. A. (2012). Evaluation of the antiepileptic effect of curcumin and Nigella sativa oil in the pilocarpine model of epilepsy in comparison with valproate. Epilepsy Behav. 24, 199–206. doi: 10.1016/j.yebeh.2012.03.026
Orellana-Paucar, A. M., Serruys, A.-S. K., Afrikanova, T., Maes, J., De Borggraeve, W., Alen, J., et al. (2012). Anticonvulsant activity of bisabolene sesquiterpenoids of Curcuma longa in zebrafish and mouse seizure models. Epilepsy Behav. 24, 14–22. doi: 10.1016/j.yebeh.2012.02.020
Peng, Y., Ao, M., Dong, B., Jiang, Y., Yu, L., Chen, Z., et al. (2021). Anti-inflammatory effects of curcumin in the inflammatory diseases: status, limitations and countermeasures. Drug Des. Devel. Ther. 15, 4503–4525. doi: 10.2147/DDDT.S327378
Pricci, M., Girardi, B., Giorgio, F., Losurdo, G., Ierardi, E., and Di Leo, A. (2020). Curcumin and colorectal cancer: from basic to clinical evidences. Int. J. Mol. Sci. 21, 2364. doi: 10.3390/ijms21072364
Reeta, K. H., Mehla, J., Pahuja, M., and Gupta, Y. K. (2011). Pharmacokinetic and pharmacodynamic interactions of valproate, phenytoin, phenobarbitone and carbamazepine with curcumin in experimental models of epilepsy in rats. Pharmacol. Biochem. Behav. 99, 399–407. doi: 10.1016/j.pbb.2011.05.011
Rodrigues, H. C. N., Martins, T. F. P., Santana, N. C. F. e. S., Braga, C. C., Silva, M. A. C., da Cunha, L. C., et al. (2021). Antioxidant and anti-inflammatory response to curcumin supplementation in hemodialysis patients: a randomized, double-blind, placebo-controlled clinical trial. Clin. Nutr. ESPEN 44, 136–142. doi: 10.1016/j.clnesp.2021.06.006
Rosenow, F., Klein, K. M., and Hamer, H. M. (2015). Non-invasive EEG evaluation in epilepsy diagnosis. Expert Rev. Neurother. 15, 425–444. doi: 10.1586/14737175.2015.1025382
Saha, L., Chakrabarti, A., Kumari, S., Bhatia, A., and Banerjee, D. (2016). Antiapoptotic and neuroprotective role of Curcumin in Pentylenetetrazole (PTZ) induced kindling model in rat. Indian J. Exp. Biol. 54, 133–141.
Santos, G. F. S., Ferreira, L. O., Gerrits Mattos, B., Fidelis, E. J., Souza, A. S., Batista, P. S., et al. (2021). Electrocorticographic description of the effects of anticonvulsant drugs used to treat lidocaine-induced seizures. Brain Behav. 11:e01940. doi: 10.1002/brb3.1940
Sierra, A., Gröhn, O., and Pitkänen, A. (2015). Imaging microstructural damage and plasticity in the hippocampus during epileptogenesis. Neuroscience 309, 162–172. doi: 10.1016/j.neuroscience.2015.04.054
Singla, S., Garcia, G. E., Rovenolt, G. E., Soto, A. L., Gilmore, E. J., Hirsch, L. J., et al. (2020). Detecting seizures and epileptiform abnormalities in acute brain injury. Curr. Neurol. Neurosci. Rep. 20:42. doi: 10.1007/s11910-020-01060-4
Sultana, B., Panzini, M.-A., Veilleux Carpentier, A., Comtois, J., Rioux, B., Gore, G., et al. (2021). Incidence and prevalence of drug-resistant epilepsy. Neurology 96, 805–817. doi: 10.1212/WNL.0000000000011839
Thumann, T. A., Pferschy-Wenzig, E.-M., Moissl-Eichinger, C., and Bauer, R. (2019). The role of gut microbiota for the activity of medicinal plants traditionally used in the European Union for gastrointestinal disorders. J. Ethnopharmacol. 245:112153. doi: 10.1016/j.jep.2019.112153
Touhidi, A., Haghighizad, H., and Pourmotabbed, A. (2018). Effect of curcumin on passive avoidance learning disorders induced by seizure activity under chronic restraint stress in rats. Neurol. Sci. Neurophysiol. 35, 77–83. doi: 10.5152/NSN.2018.10203
Tsai, Y.-M., Chien, C.-F., Lin, L.-C., and Tsai, T.-H. (2011). Curcumin and its nano-formulation: the kinetics of tissue distribution and blood–brain barrier penetration. Int. J. Pharm. 416, 331–338. doi: 10.1016/j.ijpharm.2011.06.030
Vijayakumar, S., Kasthuri, G., Prabhu, S., Manogar, P., and Parameswari, N. (2018). Screening and identification of novel inhibitors against human 4-aminobutyrate-aminotransferase: a computational approach. Egypt. J. Basic Appl. Sci. 5, 210–219. doi: 10.1016/j.ejbas.2018.05.008
Wang, L., Ding, J., Zhu, C., Guo, B., Yang, W., He, W., et al. (2021). Semaglutide attenuates seizure severity and ameliorates cognitive dysfunction by blocking the NLR family pyrin domain containing 3 inflammasome in pentylenetetrazole-kindled mice. Int. J. Mol. Med. 48:219. doi: 10.3892/ijmm.2021.5052
Witkin, J., and Li, X. (2013). Curcumin, an active constiuent of the ancient medicinal herb Curcuma longa L.: some uses and the establishment and biological basis of medical efficacy. CNS Neurol. Disord. Drug Targets 12, 487–497. doi: 10.2174/1871527311312040007
Yin, Y. H., Ahmad, N., Azmi, N., and Makmor-Bakry, M. (2018). Curcumin: the molecular mechanisms of action in inflammation and cell death during kainate-induced epileptogenesis. Indian J. Pharm. Educ. Res. 52, 32–41. doi: 10.5530/ijper.52.1.4
Keywords: Curcuma longa, seizure, diazepam, neurodegenaration, hippocampus
Citation: Nascimento CP, Ferreira LO, Silva ALMd, Silva ABNd, Rodrigues JCM, Teixeira LL, Azevedo JEC, Araujo DBd, Hamoy AO, Gonçalves BH, Coelho BHDO, Lopes DCF and Hamoy M (2022) A Combination of Curcuma longa and Diazepam Attenuates Seizures and Subsequent Hippocampal Neurodegeneration. Front. Cell. Neurosci. 16:884813. doi: 10.3389/fncel.2022.884813
Received: 27 February 2022; Accepted: 16 May 2022;
Published: 14 June 2022.
Edited by:
Hasan Turkez, Atatürk University, TurkeyReviewed by:
Gabriele Ruffolo, Sapienza University of Rome, ItalyCopyright © 2022 Nascimento, Ferreira, Silva, Silva, Rodrigues, Teixeira, Azevedo, Araujo, Hamoy, Gonçalves, Coelho, Lopes and Hamoy. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dielly Catrina Favacho Lopes, ZGNsb3Blc0B1ZnBhLmJy; Moisés Hamoy, aGFtb3l1ZnBhQGdtYWlsLmNvbQ==
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.