
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Cell. Neurosci., 12 May 2022
Sec. Cellular Neuropathology
Volume 16 - 2022 | https://doi.org/10.3389/fncel.2022.866122
This article is part of the Research TopicNew Insights Into the Role of Estrogen in Neurological Disorders: Pre-Clinical StudiesView all 4 articles
 Ivanny Carolina Marchant1,2*
Ivanny Carolina Marchant1,2* Stéren Chabert3,4,5
Stéren Chabert3,4,5 Jonathan Martínez-Pinto6,7,8
Jonathan Martínez-Pinto6,7,8 Ramón Sotomayor-Zárate6,7,8
Ramón Sotomayor-Zárate6,7,8 Ricardo Ramírez-Barrantes9
Ricardo Ramírez-Barrantes9 Lilian Acevedo10
Lilian Acevedo10 Claudio Córdova11
Claudio Córdova11 Pablo Olivero2,11*
Pablo Olivero2,11*Menopause transition can be interpreted as a vulnerable state characterized by estrogen deficiency with detrimental systemic effects as the low-grade chronic inflammation that appears with aging and partly explains age-related disorders as cancer, diabetes mellitus and increased risk of cognitive impairment. Over the course of a lifetime, estrogen produces several beneficial effects in healthy neurological tissues as well as cardioprotective effects, and anti-inflammatory effects. However, clinical evidence on the efficacy of hormone treatment in menopausal women has failed to confirm the benefit reported in observational studies. Unambiguously, enhanced verbal memory is the most robust finding from longitudinal and cross-sectional studies, what merits consideration for future studies aiming to determine estrogen neuroprotective efficacy. Estrogen related brain activity and functional connectivity remain, however, unexplored. In this context, the resting state paradigm may provide valuable information about reproductive aging and hormonal treatment effects, and their relationship with brain imaging of functional connectivity may be key to understand and anticipate estrogen cognitive protective effects. To go in-depth into the molecular and cellular mechanisms underlying rapid-to-long lasting protective effects of estrogen, we will provide a comprehensive review of cognitive tasks used in animal studies to evaluate the effect of hormone treatment on cognitive performance and discuss about the tasks best suited to the demonstration of clinically significant differences in cognitive performance to be applied in human studies. Eventually, we will focus on studies evaluating the DMN activity and responsiveness to pharmacological stimulation in humans.
Menopause is a biological milestone linked to the onset of cognitive impairment, amongst several deleterious systemic effects. There would be a window of opportunity to provide hormone therapy (HT) that would benefit users. Clinical studies show cognitive improvements mainly in verbal memory (Maki, 2000) in women exposed to HT during this period. Studies are, however, hardly comparable due to heterogeneity in menopause origin, HT formulations, neuropsychological tests, and neuroimaging techniques. Experimental studies present differences in methodologies, type, and age of animal injury model that explain the apparent failure of scientific research to show the clinical benefit of estrogens.
Clinical studies have revealed higher activation of fronto-cingulate regions in menopausal women under estrogen treatment by brain functional magnetic resonance imaging (fMRI), although in many cases, no difference in cognitive performance was demonstrated. fMRI studies may also have different appreciations, thus the abnormal activation observed in some individuals who executed adequately the imposed task could be interpreted as a compensatory activation due to sub-clinical cognitive impairment (Comasco and Frokjaer, 2014).
During rest, synchronous hemodynamic activity occurs in different brain networks, the so-called Resting State Networks (RSN) (Ramírez-Barrantes et al., 2019), one of which being the default mode network (DMN). The DMN has been consistently implied as a biomarker of cognitive function and aging-related decline. Estrogen has been implicated in the modulation of DMN in pre and post-menopausal women (Petersen et al., 2014; Weis and Hodgetts, 2019). Resting state fMRI may thus be valuable to explore rapid estrogen effects avoiding confusion related to the application of cognitive tasks. According to the “healthy cell bias of estrogen benefit,” rapid estrogen effects in women with preserved DMN function may provide more insights into the complex nature of responsiveness to estrogen treatment cell bias of estrogen benefit (Brinton, 2008).
Sex hormones have important effects on non-reproductive organs such as the brain. Estrogen associates with the modulation of brain circuits involved in motivated behaviors, emotions, memory, and executive functions (Beyer, 1999; McEwen and Alves, 1999; Bethea et al., 2002; Almey and Milner, 2015). The relationship of plasma estrogen with these behaviors has been modeled in menopausal animals through ovariectomization (OVX) or by the administration of estrogen synthesis inhibitors. Several cognitive tasks modulated by estrogens have been extrapolated to human studies, either in young or post-menopausal women. We will expose examples of cognitive alterations observed in animals and humans and their relationship with estrogen therapy.
Ovariectomization is a safe surgery to produce undetectable serum levels of 17β-estradiol (E2) within 2–4 weeks (Medina-Contreras et al., 2020). Recently, in 5.5-month-old OVX rats, physical and psychological stressors were synergistic to decrease exploration, learning, and memory behaviors (Medina-Contreras et al., 2020). As E2 therapy reverses these alterations, a neuroprotective role of E2 has been suggested in post-menopausal women exposed to chronic stress (Khaleghi et al., 2021).
17β-estradiol effects are less intense in OVX administered in older (12 months) than middle-aged rats, since E2 only reversed anxious behaviors in the open field test without significant changes in the anxiety score in elevated plus maze or memory in novel object recognition test (Renczes et al., 2020). Chronic stress, chronic exposure to obesogenic, and hypercaloric diets can potentiate cognitive and behavioral alterations in OVX rats. In OVX macaques previously fed with a 6-week western diet, the animals treated with E2 for 30 months performed better spatial tasks than animals that received treatment via vehicle (Zimmerman et al., 2020).
4-vinylcyclohexene diepoxide (VCD) produces a gradual loss of ovarian follicles and E2 synthesis (Mayer et al., 2004). The 30-day VCD administration produces neurochemical alterations in monoamine and metabolite contents in the hippocampus, prefrontal cortex, and striatum, all of which express estrogen receptors (Long et al., 2019). E2 was more effective in restoring normal monoamine levels in the surgical model of menopause than in the VCD model (Long et al., 2019) and thus, it would be more effective in restoring cognitive performance in surgical menopause. The intracerebroventricular (icv) administration of the aromatase inhibitor letrozole produces dose-dependent cognitive alterations associated with a reduction in hippocampal E2 and a decrease in the firing rate of pyramidal neurons (Marbouti et al., 2020). Aromatase inhibition increases the expression of estrogen receptor α (Erα) and estrogen receptor β (Erβ) and decreases the expression of G protein-coupled estrogen receptor 1 (GPER) in the hippocampus (Marbouti et al., 2020), probably hampering memory formation. In humans, higher estradiol levels have ben correlated with greater hippocampal volume in men. Hippocampal activity has been reduced by letrozole, while a partially compensating increased prefrontal activity as in AD and aging that might mask estradiol's effect on observable behavior, i.e., memory performance, in some of the studies, can be observed. Subtle memory deficits in women under letrozole for therapeutical reasons may illuminate further analyses on these such E2-abstinence effects.
In 3-month-old rats, icv streptozotocin (STZ) produces similar memory and learning alterations to those observed in animal models of Alzheimer's disease (AD) (Wei et al., 2019). STZ induces oxidative alterations in the prefrontal cortex and hippocampus (Wei et al., 2019). The STZ administration, followed by chronic 21-day treatment with GPER agonist daidzein, increases the navigation time in the target quadrant and decreases the latency time to find the platform in Morris water maze (Wei et al., 2019). This study underlines the therapeutic potential of phytoestrogens in cognitive disorders (Echeverria et al., 2021).
The poly-I:C injection, a Toll-like receptor 3 agonist in pregnant mice, produces cognitive alterations in adult female offspring, which are reversed by the selective E2 receptor modulator raloxifene (Schroeder et al., 2019). Neonatal hypoxia due to carotid artery occlusion for 1 h at post-natal day 7 induces long-term cognitive deficits in rats (Anastacio et al., 2019) that are counteracted by phytoestrogen coumestrol until 3 h of hypoxia, as indicated by cognitive and morphological changes in hippocampus at post-natal day 60 (Hampson, 2018).
The neuroprotective efficacy of HT remains unclarified. Considering that in the next decade, there would be 1.2 billion of menopausal and post-menopausal women (Hampson, 2018; Echeverria et al., 2021), it is necessary to standardize the cognitive task to be carried out in this population along with the pharmacological treatments to use. In humans, cognitive ability can be assessed by the mental rotation task (MRT) which considers perception, identification, orientation, judgment, response, and execution processes (Xue et al., 2017). In normal menstrual cycling women, luteal phase high progesterone levels are associated with better performance in visuospatial tasks in the MRT (Shirazi et al., 2021). In post-menopausal women with schizophrenia cognitive decline, the raloxifene administration did not produce significant changes in cognitive function compared to the group of patients receiving treatment via vehicle (Huerta-Ramos et al., 2020). Rather than associated with a loss of pharmacological HT effect, the diversity of human E2 effects should be correlated with psychiatric or neurodegenerative co-morbid conditions where the degree of cognitive deficit is variable in intensity and not necessarily reversed by neuromodulators.
The ability of estradiol to influence cognition during development, at adulthood, and during aging has been demonstrated years ago (Luine, 2012). E2 enhances in vitro the consolidation of hippocampal memories 5–30 min after treatment through the activation of several cell signaling cascades (Frick, 2015). E2 regulates the hippocampal morphology and function, spine density, neurogenesis, synaptic plasticity, neurotransmission, and gene expression, which are all facilitators of memory consolidation (Patel et al., 2022). It regulates the dendritic spine density in the medial prefrontal cortex, somatosensory cortex, amygdala, and dendritic length in the basal forebrain (Frick, 2015). E2 improves hippocampal-dependent spatial memory and object and spatial recognition memory in OVX rats.
As endocrine contributor, brain synthesizes E2 from androgen precursors by the enzyme aromatase, namely, the so-called neuron derived E2 (NDE2), found at synapses and presynaptic terminals in neurons in both male and female brains of rodents, monkeys, birds, amphibians, and humans. NDE2 regulates sexual differentiation, reproduction, synaptic plasticity, cognition, neuroinflammation, and neuroprotection (Brann et al., 2021). In the rat hippocampus, changing E2 levels across estrous phases suggest that CNS-synthesized E2 may be affected by the estrous cycle in rodents. Besides fluctuating hematic progesterone into the brain and its subsequent conversion into E2 (Kato et al., 2013), neural E2 changes might be related to changes of P450 aromatase depending on fluctuation of kinases related to synaptic plasticity (Hojo, 2018; Tozzi and Bellingacci, 2020).
In rat hippocampal slices treated with aromatase inhibitors, the amplitude of long-term potentiation is reduced with consequently reduced spatial, recognition, and contextual-fear memory, suggesting an important role of NDE2. NDE2 relates to synaptic plasticity and memory via the regulation of actin cytoskeleton polymerization/depolymerization and post-synaptic density dynamics, which is key for spine formation, enhancement of MAPK/ERK and PI3K-AKT signaling, regulation of CREB-BDNF signaling, and mediation by estrogen receptors and SRC-1 (Nelson, 2001; Spencer et al., 2008; Simpkins et al., 2012; Terasawa, 2018).
Rapid E2 mechanisms on hippocampal memory in rodents have been most assessed in spatial tasks, i.e., Morris water maze, radial arm maze, delayed non-match to position, object placement task, and object recognition tasks. Other tools are social learning paradigms like the social transmission of food preferences, female mate choice copying, and social recognition (Zhao et al., 2017; Patel et al., 2022). ERα and ERβ agonists are also good alternatives to depict the molecular rapid E2 mechanisms. To observe rapid actions, E2 or ER agonists must be administered either systemically or intracranially minutes before the cognitive task since the rapid E2 effect expresses on neural plasticity within 1 h, modulating cell signaling, synaptic transmission, and dendritic spine density (Phan et al., 2011). ERα agonist propyl pyrazole triol (PPT) and ERβ agonist diarylpropionitrile (DPN) in vitro have shown to alter cell signaling, synaptic transmission, and long-term depression in hippocampal sections within 1.5 h of application (Phan et al., 2011). E2 can alter the performance in learning tests within 40 min of administration in OVX mice. Similarly, PPT administered to mice at 50 or 75 μg/mice s.c. 15 min before a social discrimination paradigm increased social recognition induced by ERα agonism. In addition, PPT or E2 improved object recognition and object placement learning (Phan et al., 2011, 2012; Gabor et al., 2015). Administered post training, systemically or into the dorsal hippocampus, E2 enhances spatial and object recognition memory consolidation in Morris water maze, spatial memory consolidation in an object location task, and object recognition memory consolidation (Gresack, 2006; Boulware and Heisler, 2013; Patel et al., 2022).
Magnetic resonance images have been useful to explore brain functions for quite some time, as Ogawa et al. (1992) emphasized that the magnetic resonance (MR) signal, called “BOLD signal,” is blood oxygenation level dependent as observed in a task-related imaging set-up. The basis of BOLD signal is the ratio of oxy- vs. deoxy-hemoglobin in local venous blood that results from the rise in oxygen consumption and local blood flow when local neuronal activity increases. As the oxy- vs. deoxy-hemoglobin do not have the same paramagnetic properties, the MR signal is modified through a variation in . It is noteworthy that the peak of signal variation is observed close to 5 s after the stimulus onset (Presa et al., 2020).
These first observations in fMRI are associated to the realization of a specific task of heterogeneous design that relies on patient cooperation with significant variability. A solution, as follows, was proposed by Biswal et al. (1995): with no special instruction to the patient but to just stay still, the time course of low-frequency fluctuation of some regions in the brain were shown to present high temporal correlation. Functional networks of different cortical regions were observed, identifying functionally connected nodes, such as the right and left motor cortex together, or the right and left visual cortex. As no stimulus was presented to the patient, the maps created were denominated resting-state functional MRI (rs-fMRI) (Damoiseaux et al., 2006). In either task-based fMRI or rs-fMRI, the precise physiological mechanisms underlying the temporal synchronous signal fluctuation between cortical areas are not yet clearly defined. The coupling or uncoupling of neuronal activity and vascular reactivity is an active field of study (Rossetti et al., 2021; Stiernman et al., 2021). The RSN is elicited by a wide variety of sensory, motor, and cognitive tasks, representing 20% of the overall energy consumed by a person (Raichle, 2010). It is composed by the salience network, executive network, auditory network, sensory motor network, visuospatial network, and default mode network (Smitha et al., 2017). The DMN is observed in awake individuals, containing areas mainly in the medial prefrontal cortex, and medial temporal lobe, and in the posterior cingulate cortex and angular gyrus (Buckner and Andrews-Hanna, 2008). Observations with other imaging modality, such as glucose imaging using positron emission tomography, supported the consideration that the DMN consists of specific areas connected in a stable network (Buckner and Andrews-Hanna, 2008). Exploration of DMN and its disruption has gained increasing interest in neuropsychiatric disorders as attention deficit hyperactivity disorder or ADHD (Mohan and Roberto, 2016). rs-fMRI provides identification of nodes that are functionally related and allows quantifying correlation, graph analysis, and functional connectivity analysis (Yang and Gohel, 2020). Further cognitive exploration can then be undertaken using different dynamic causal modeling (Friston, 2009). Various processing methods have been proposed in rs-fMRI with still no convergence on a standard one (Yang and Gohel, 2020). Seed-based connectivity analysis looks for correlation between regions-of-interest (ROI) with the associated question on how to define those ROI. Independent component analysis (ICA) can be applied in different manners, with the associated question on how many components must be considered and how to interpret each obtained component. There is also processing using amplitude of low frequency fluctuation (ALFF) or regional homogeneity analysis (ReHo) (Satterthwaite et al., 2012; Yang and Gohel, 2020).
Using task-based fMRI, Dietrich et al. (2001) and Stevens and Clark (2005) stressed the idea that the hemodynamic response function (HRF), whose canonical form lies behind most of the fMRI analysis, is modulated by blood estrogen and by estrogen therapy (Stevens and Clark, 2005). Plasma E2 could thus introduce confusion effects through probable modulation of the vascular compartment and of the neuro-vascular coupling, making it difficult to interpret fMRI results. Rangaprakash et al. (2018) showed that if the HRF variability, between individuals and cortical regions, is not considered in rs-fMRI, then up to 15% of error in functional connectivity estimation could occur and false connectivity detection could be increased. rs-fMRI is without a doubt a powerful tool that still presents some methodological challenges to implement in a robust manner.
The DMN is characterized by the synchronous activation of brain regions as the medial prefrontal cortex, posterior cingulate cortex, precuneus, inferior parietal lobule, and inferolateral temporal cortex (Sood, 2013; Raichle, 2015) without external stimuli. It is presumably related to introspective and self-referential thought processes and becomes attenuated during goal-directed tasks (Ramírez-Barrantes et al., 2019), suggesting that its suppression during task execution favors the goal success (Fox et al., 2005; Hampson et al., 2006; Leech et al., 2011). The activation of DMN associates with processes like creativity and mind wandering, while abnormal activation and deactivation is related to psychiatric and medical conditions like anxiety, major depressive disorder, schizophrenia, and AD (Greicius et al., 2004; Anticevic et al., 2012; Whitfield-Gabrieli, 2012; Sunavsky, 2020).
Despite the important role of the RSN and the DMN in human cognition and wellbeing, little is known about physiological variability of the RSN connectivity across lifespan or between genders. A potential role of sex hormones on differential formation and activation of these RSNs would explain gender differences in cognitive tasks (Weis and Hodgetts, 2019; Pritschet et al., 2020), although the results are still inconsistent probably due to methodological differences between resting state studies. The function of DMN could be modulated across the menstrual cycle phases while it remains stable in men (Weis and Hodgetts, 2019). Predominantly composed by prefrontal areas that are very sensitive to sex hormones, DMN is presumably highly susceptible to menstrual cycle effects. In women, an increased DMN connectivity within the left middle frontal area in the menstrual phase as opposed to the follicular phase has been reported (Weis and Hodgetts, 2019). Other studies indicated that women in the luteal phase had reduced coherence between the left angular gyrus and the remaining network compared to women in the follicular phase. Oral contraceptive users in the active phase of their pill cycle showed reduced coherence between the left angular gyrus and the remaining network than women in follicular phase without contraceptive treatment (Petersen et al., 2014). Altered function of cell cycle or contraceptive methods could thus modify both abstract and self-referential reasoning and contribute to development of disease states involving DMN connectivity.
Estradiol determines central gender dimorphism (Ramírez-Barrantes and Marchant, 2016; Russell and Jones, 2019) and influences the performance in frontally mediated cognitive tasks, such as top-down cognitive control (Hjelmervik et al., 2014; Thimm et al., 2014). Hence, the withdrawal of E2 in the limbic system could be related with changes in mood, behavior, and cognition (Genazzani et al., 2007). In pre-menopausal women, during low E2 menstrual phase, activation of DMN in the left middle frontal area was increased (Weis and Hodgetts, 2019), while greater connectivity was demonstrated during the high-estrogen follicular menstrual phase (Petersen et al., 2014). These associations were observed with both studies having focused their investigation on the anterior frontoparietal network. Wang et al. showed an opposite effect of progesterone and estradiol in DMN (Wang et al., 2020). Estradiol, but not progesterone, could facilitate the medial prefrontal cortex-to-inferior parietal lobule functional connectivity, the posterior component of DMN (Wang et al., 2020). A huge study of the functional reorganization of brain networks during menstrual cycle found that progesterone is associated with negative connection in brain networks, while improved brain functional connectivity is mainly characterized by increased concentration of estradiol throughout the cycle (Pritschet et al., 2020). The activation of DMN probably contributes to memory problems described during the menopause transition. HT has been useful to improve verbal memory and activation of hippocampus in post-menopausal women compared to their counterparts who had never used HT (Ramírez-Barrantes et al., 2020).
Exogenous E2 has been associated with better cognitive performance during aging and reduced risk of AD where DMN is also involved (Henderson, 2006), but this potential effect is still in debate. Oral contraceptive pills have exhibited specific modulation of DMN in women with post-traumatic stress disorders (Wen et al., 2021). Ninety healthy women aged from 18 to 30 years underwent a 3-day fear conditioning and extinction paradigm. They were separated in two groups, namely, oral contraceptive regular users (n = 57) and never users of oral contraceptive pills (n = 33), and their DMN connectivity and attention networks were recorded by fMRI. E2 was beneficial to modulate attention and conscious processes promoting normal fear extinction learning and extinction memory retention. This kind of modulation by exogenous E2 has not been related to a unique network modulation, but to an activation of whole-brain functional connectivity, implying functionally distinct systems as the anterior component of DMN, i.e., regions with high levels of E2 receptors (Wen et al., 2021). Therefore, in addition to endogenous E2 across normal menstrual cycle, exogenous E2 could modulate DMN contributing processes like conscious awareness, affective learning, and memory consolidation in combination with attention network (Higgins et al., 2021).
Different timescales underlie E2 effects (Figure 1). Genomic E2 mechanisms via ERs α and β trigger synthesis, release, and metabolism of neuropeptides and neuroesteroids. Non-genomic E2 actions trigger effects that appear in seconds to minutes (Sbarouni et al., 1997), modulate electrical excitability, and neuronal cell death (Zhao, 2007; Raz et al., 2008; Mukai et al., 2010; Ramírez-Barrantes et al., 2020). Rapid E2 mechanisms can modulate hippocampal memory consolidation within minutes of E2 exposure. In the context of direct acute E2 effect, we have demonstrated that it induces specific mitochondrial-mediated resistance to oxidative stress dependent of the function of membrane channel transient receptor potential cation channel subfamily V member 1 (TRPV1) (Ramírez-Barrantes et al., 2020). Altered mitochondrial proteostasis could thus impede the compensatory mechanisms against cell damage. The rapid modification of the activity of non-classical E2 receptors, such as TRPV1, might be critical for the maintenance of intracerebral functional connectivity during acute E2 stimulation. Future clinical trials focused on these rapid signal changes following estrogen exposure may identify predictors of long-lasting HT effectiveness.
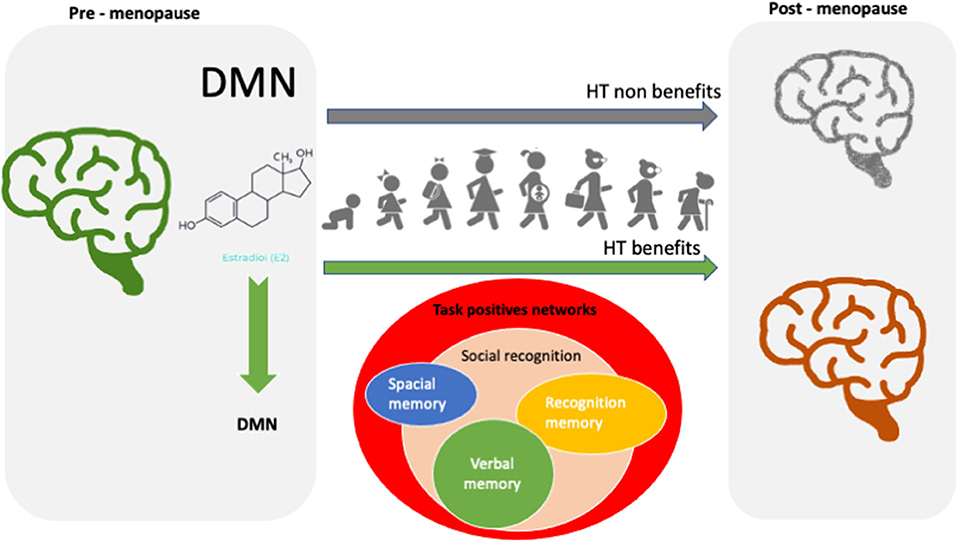
Figure 1. Rapid to long-lasting estrogen effects.
IM and PO conceived the idea and wrote the manuscript. SC, JM-P, RS-Z, and RR-B developed contents in rs-fMRI, animal models, connection with human studies, and DMN, respectively. LA reviewed critically neurological topics. CC developed figures. All authors reviewed critically the final version for intellectual contents.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Almey, A., and Milner, T. A. (2015). Estrogen receptors in the central nervous system and their implication for dopamine-dependent cognition in females. Horm. Behav. 74, 125–138. doi: 10.1016/j.yhbeh.2015.06.010
Anastacio, J. B. R., Sanches, E. F., Nicola, F., Odorcyk, F., Fabres, R. B., and Netto, C. A. (2019). Phytoestrogen coumestrol attenuates brain mitochondrial dysfunction and long-term cognitive deficits following neonatal hypoxia-ischemia. Int. J. Dev. Neurosci. 79, 86–95. doi: 10.1016/j.ijdevneu.2019.10.009
Anticevic, A., Cole, M. W., Murray, J. D., Corlett, P. R., Wang, X.-J., and Krystal, J. H. (2012). The role of default network deactivation in cognition and disease. Trends Cogn. Sci. 16, 584–592. doi: 10.1016/j.tics.2012.10.008
Bethea, C. L., Lu, N. Z., and Gundlah, C. (2002). Diverse actions of ovarian steroids in the serotonin neural system. Front. Neuroendocrinol. 23, 41–100. doi: 10.1006/frne.2001.0225
Beyer, C. (1999). Estrogen and the developing mammalian brain. Anat. Embryol. 199, 379–390. doi: 10.1007/s004290050236
Biswal, B., Zerrin Yetkin, F., and Haughton, V. M. (1995). Functional connectivity in the motor cortex of resting human brain using echo-planar mri. Magn. Reson. Med. 34, 537–541. doi: 10.1002/mrm.1910340409
Boulware, M. I., and Heisler, J. D. (2013). The memory-enhancing effects of hippocampal estrogen receptor activation involve metabotropic glutamate receptor signaling. J. Neurosci. 33, 15184–15194. doi: 10.1523/JNEUROSCI.1716-13.2013
Brann, D. W., Lu, Y., Wang, J., Sareddy, G. R., Pratap, U. P., Zhang, Q., et al. (2021). Neuron-derived estrogenandmdash;a key neuromodulator in synaptic function and memory. Int. J. Mol. Sci. 22, 13242. doi: 10.3390/ijms222413242
Brinton, R. D. (2008). The healthy cell bias of estrogen action: mitochondrial bioenergetics and neurological implications. Trends Neurosci. 31, 529–537. doi: 10.1016/j.tins.2008.07.003
Buckner, R. L., and Andrews-Hanna, J. R. (2008). The brain's default network. Ann. N. Y. Acad. Sci. 1124, 1–38. doi: 10.1196/annals.1440.011
Comasco, E., and Frokjaer, V. G. (2014). Functional and molecular neuroimaging of menopause and hormone replacement therapy. Front. Neurosci. 8, 388. doi: 10.3389/fnins.2014.00388
Damoiseaux, J. S., Rombouts, S. A. R. B., Barkhof, F., Scheltens, P., Stam, C. J., Smith, S. M., et al. (2006). Consistent resting-state networks across healthy subjects. Proc. Nat. Acad. Sci. U. S. A. 103, 13848–13853. doi: 10.1073/pnas.0601417103
Dietrich, T., Krings, T., Neulen, J., Willmes, K., Erberich, S., Thron, A., and Sturm, W. (2001). Effects of blood estrogen level on cortical activation patterns during cognitive activation as measured by functional MRI. Neuroimage 13, 425–432. doi: 10.1006/nimg.2001.0703
Echeverria, V., Echeverria, F., Barreto, G. E., and Echeverria, J. (2021). Estrogenic plants: to prevent neurodegeneration and memory loss and other symptoms in women after menopause. Front. Pharmacol. 12, 644103. doi: 10.3389/fphar.2021.644103
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Essen, D. C. V., and Raichle, M. E. (2005). The human brain is intrinsically organized into dynamic, anticorrelated functional networks. Proc. Nat. Acad. Sci. U. S. A. 102, 9673–9678. doi: 10.1073/pnas.0504136102
Frick, K. M. M. (2015). mechanisms underlying the memory-enhancing effects of estradiol. Horm. Behav. 74, 4–18. doi: 10.1016/j.yhbeh.2015.05.001
Friston, K. (2009). Causal modelling and brain connectivity in functional magnetic resonance imaging. PLoS Biol. 7, e1000033. doi: 10.1371/journal.pbio.1000033
Gabor, C., Lymer, J., and Phan, A. (2015). Rapid effects of the G-protein coupled oestrogen receptor (GPER) on learning and dorsal hippocampus dendritic spines in female mice. Physiol. Behav. 149, 53–60. doi: 10.1016/j.physbeh.2015.05.017
Genazzani, A. R., Pluchino, N., and Luisi, S. (2007). Estrogen, cognition and female ageing. Hum. Reprod. Update 13, 175–187. doi: 10.1093/humupd/dml042
Greicius, M. D., Srivastava, G., and Reiss, A. L. (2004). Default-mode network activity distinguishes Alzheimer's disease from healthy aging: Evidence from functional MRI. Proc. Nat. Acad. Sci. U. S. A.101, 4637–4642. doi: 10.1073/pnas.0308627101
Gresack, J. E. (2006). Post-training estrogen enhances spatial and object memory consolidation in female mice. Pharmacol. Biochem. Behav. 84, 112–119. doi: 10.1016/j.pbb.2006.04.013
Hampson, E. (2018). Estrogens, aging, and working memory. Curr. Psychiatry Rep. 20, 109. doi: 10.1007/s11920-018-0972-1
Hampson, M., Driesen, N. R., Skudlarski, P., and Gore, J. C. (2006). Brain connectivity related to working memory performance. J. Neurosci. 26, 13338–3343. doi: 10.1523/JNEUROSCI.3408-06.2006
Henderson, V. W. (2006). Estrogen-containing hormone therapy and Alzheimer's disease risk: understanding discrepant inferences from observational and experimental research. Neuroscience. 138, 1031–1039. doi: 10.1016/j.neuroscience.2005.06.017
Higgins, C., Liu, Y., Vidaurre, D., Kurth-Nelson, Z., Dolan, R., Behrens, T., et al. (2021). Replay bursts in humans coincide with activation of the default mode and parietal alpha networks. Neuron (2021) 109, 882–893.e7. doi: 10.1101/2020.06.23.166645
Hjelmervik, H., Hausmann, M., Osnes, B., and Westerhausen, R. (2014). Resting states are resting traits - an fMRI study of sex differences and menstrual cycle effects in resting state cognitive control networks. PLoS ONE 9, e103492. doi: 10.1371/journal.pone.0103492
Hojo, Y. (2018). Neurosteroids in adult hippocampus of male and female rodents: biosynthesis and actions of sex steroids. Front. Endocrinol. 9, 183. doi: 10.3389/fendo.2018.00183
Huerta-Ramos, E., Labad, J., Cobo, J., Nez, C., Creus, M., Garca-Pars, G., et al. (2020). Effects of raloxifene on cognition in postmenopausal women with schizophrenia: a 24-week double-blind, randomized, parallel, placebo-controlled trial. Eur. Arch. Psychiatry Clin. Neurosci. 270, 729–737. doi: 10.1007/s00406-019-01079-w
Kato, A., Hojo, Y., Higo, S., Komatsuzaki, Y., Murakami, G., Yoshino, H., et al. (2013). Female hippocampal estrogens have a significant correlation with cyclic fluctuation of hippocampal spines. Front. Neural Circuits 7, 149. doi: 10.3389/fncir.2013.00149
Khaleghi, M., Rajizadeh, M. A., Bashiri, H., Kohlmeier, K. A., Mohammadi, F., Khaksari, M., et al. (2021). Estrogen attenuates physical and psychological stress-induced cognitive impairments in ovariectomized rats. Brain Behav. 11, 2139. doi: 10.1002/brb3.2139
Leech, R., Kamourieh, S., and Beckmann, C. F. (2011). Fractionating the default mode network: distinct contributions of the ventral and dorsal posterior cingulate cortex to cognitive control. J. Neurosci. 31, 3217–3224. doi: 10.1523/JNEUROSCI.5626-10.2011
Long, T., Yao, J. K., Li, J., Kirshner, Z. Z., Nelson, D., Dougherty, G. G., et al. (2019). Estradiol and selective estrogen receptor agonists differentially affect brain monoamines and amino acids levels in transitional and surgical menopausal rat models. Mol. Cell. Endocrinol. 496, 110533. doi: 10.1016/j.mce.2019.110533
Luine, V. N. (2012). Estrogens facilitate memory processing through membrane mediated mechanisms and alterations in spine density. Front. Neuroendocrinol. 33, 388–402. doi: 10.1016/j.yfrne.2012.07.004
Maki, P. M. (2000). Longitudinal effects of estrogen replacement therapy on PET cerebral blood flow and cognition. Neurobiol. Aging 21, 373–383. doi: 10.1016/S0197-4580(00)00123-8
Marbouti, L., Zahmatkesh, M., and Riahi, E. (2020). Inhibition of brain 17beta-estradiol synthesis by letrozole induces cognitive decline in male and female rats. Neurobiol. Learn. Mem. 175, 107300. doi: 10.1016/j.nlm.2020.107300
Mayer, L. P., Devine, P. J., and Dyer, C. A. (2004). The follicle-deplete mouse ovary produces androgen. Biol. Reprod. 71, 130–138. doi: 10.1095/biolreprod.103.016113
McEwen, B. S., and Alves, S. E. (1999). Estrogen actions in the central nervous system. Endocr. Rev. 20, 279–307. doi: 10.1210/er.20.3.279
Medina-Contreras, J., Villalobos-Molina, R., and Zarain-Herzberg, A. (2020). Ovariectomized rodents as a menopausal metabolic syndrome model. A minireview. Mol. Cell. Biochem. 475, 261–276. doi: 10.1007/s11010-020-03879-4
Mohan, A., and Roberto, A. J. (2016). The significance of the default mode network (DMN) in neurological and neuropsychiatric disorders: a review. Yale J. Biol. Med. 89, 49–57.
Mukai, H., Kimoto, T., Hojo, Y., Kawato, S., Murakami, G., Higo, S., et al. (2010). Modulation of synaptic plasticity by brain estrogen in the hippocampus. Biochim. Biophys. Acta Gen. Subj. 1800, 1030–1044. doi: 10.1016/j.bbagen.2009.11.002
Nelson, L. R. (2001). Estrogen production and action. J. Am. Acad. Dermatol. 45, 116–124. doi: 10.1067/mjd.2001.117432
Ogawa, S., Tank, D. W., Menon, R., Ellermann, J. M., Kim, S. G., Merkle, H., et al. (1992). Intrinsic signal changes accompanying sensory stimulation: functional brain mapping with magnetic resonance imaging. Proc. Nat. Acad. Sci. U. S. A. 89, 5951–5955. doi: 10.1073/pnas.89.13.5951
Patel, S. A., Frick, K. M., and Newhouse, P. A. (2022). Estradiol effects on spatial memory in women. Behav. Brain Res. 417, 113592. doi: 10.1016/j.bbr.2021.113592
Petersen, N., Kilpatrick, L. A., and Goharzad, A. (2014). Oral contraceptive pill use and menstrual cycle phase are associated with altered resting state functional connectivity. Neuroimage 90, 24–32. doi: 10.1016/j.neuroimage.2013.12.016
Phan, A., Gabor, C. S., Favaro, K. J., Kaschack, S., Armstrong, J. N., MacLusky, N. J., et al. (2012). Low doses of 17beta-estradiol rapidly improve learning and increase hippocampal dendritic spines. Neuropsychopharmacology 37, 2299–2309. doi: 10.1038/npp.2012.82
Phan, A., Lancaster, K. E., Armstrong, J. N., MacLusky, N. J., and Choleris, E. (2011). Rapid effects of estrogen receptor alpha and beta selective agonists on learning and dendritic spines in female mice. Endocrinology 152, 1492–1502. doi: 10.1210/en.2010-1273
Presa, J. L., Saravia, F., and Bagi, Z. (2020). Vasculo-neuronal coupling and neurovascular coupling at the neurovascular unit: impact of hypertension. Front. Physiol. 11, 584135. doi: 10.3389/fphys.2020.584135
Pritschet, L., Santander, T., Taylor, C. M., Layher, E., Yu, S., Miller, M. B., et al. (2020). Functional reorganization of brain networks across the human menstrual cycle. Neuroimage. 220, 117091. doi: 10.1016/j.neuroimage.2020.117091
Raichle, M. E. T. (2010). views of brain function. Trends Cogn. Sci. 14, 180–190. doi: 10.1016/j.tics.2010.01.008
Raichle, M. E. T. (2015). Brain's default mode network. Annu. Rev. Neurosci. 38, 433–447. doi: 10.1146/annurev-neuro-071013-014030
Ramírez-Barrantes, R., Arancibia, M., Stojanova, J., Asp-Snchez, M., Crdova, C., and Henrquez-Ch, R. A. (2019). Default mode network, meditation, and age-associated brain changes: what can we learn from the impact of mental training on well-being as a psychotherapeutic approach? Neural Plasticity 2019, 7067592. doi: 10.1155/2019/7067592
Ramírez-Barrantes, R., Carvajal-Zamorano, K., Rodriguez, B., Cordova, C., Lozano, C., Simon, F., et al. (2020). TRPV1-estradiol stereospecific relationship underlies cell survival in oxidative cell death. Front. Physiol. 11, 444. doi: 10.3389/fphys.2020.00444
Ramírez-Barrantes, R., and Marchant, I. (2016). TRPV1 may increase the effectiveness of estrogen therapy on neuroprotection and neuroregeneration. Neural Regen. Res. 11, 1204–1207. doi: 10.4103/1673-5374.189162
Rangaprakash, D., Wu, G-. R., Marinazzo, D., and Hu, X. (2018). Hemodynamic response function (HRF) variability confounds resting-state fMRI functional connectivity. Magn. Reson. Med. 80, 1697–1713. doi: 10.1002/mrm.27146
Raz, L., Khan, M. M., Mahesh, V. B., and Vadlamudi, R. K. (2008). Rapid estrogen signaling in the brain. Neurosignals 16, 140–153. doi: 10.1159/000111559
Renczes, E., Borblyov, V., Steinhardt, M., Hpfner, T., Stehle, T., Ostatnkov, D., et al. (2020). The role of estrogen in anxiety-like behavior and memory of middle-aged female rats. Front. Endocrinol. 11, 570560. doi: 10.3389/fendo.2020.570560
Rossetti, G. M., dAvossa, G., Rogan, M., Macdonald, J. H., Oliver, S. J., and Mullins, P. G. (2021). Reversal of neurovascular coupling in the default mode network: evidence from hypoxia. J. Cerebral Blood Flow Metab. 41, 805–818. doi: 10.1177/0271678X20930827
Russell, J. K., and Jones, C. K. (2019). The role of estrogen in brain and cognitive aging. Neurotherapeutics 16, 649–665. doi: 10.1007/s13311-019-00766-9
Satterthwaite, T. D., Wolf, D. H., Loughead, J., Ruparel, K., Elliott, M. A., Hakonarson, H., et al. (2012). Impact of in-scanner head motion on multiple measures of functional connectivity: relevance for studies of neurodevelopment in youth. Neuroimage 60, 623–632. doi: 10.1016/j.neuroimage.2011.12.063
Sbarouni, E., Kyriakides, Z. S., and Antoniadis, A. (1997). Acute hemodynamic effects of estrogen administration in postmenopausal women. Am. J. Cardiol. 80, 532–535. doi: 10.1016/s0002-9149(97)00414-1
Schroeder, A., Nakamura, J. P., Hudson, M., Jones, N. C., Du, X., Sundram, S., et al. (2019). Raloxifene recovers effects of prenatal immune activation on cognitive task-induced gamma power. Psychoneuroendocrinology 110, 104448. doi: 10.1016/j.psyneuen.2019.104448
Shirazi, T. N., Levenberg, K., Cunningham, H., Self, H., Dawood, K., Crdenas, R., et al. (2021). Relationships between ovarian hormone concentrations and mental rotations performance in naturally-cycling women. Horm. Behav. 127, 104886. doi: 10.1016/j.yhbeh.2020.104886
Simpkins, J. W., Singh, M., and Brock, C. (2012). Neuroprotection and estrogen receptors. Neuroendocrinology 96, 119–130. doi: 10.1159/000338409
Smitha, K. A., Raja, K. A., Arun, K., Rajesh, P., Thomas, B., Kapilamoorthy, T., et al. (2017). Resting state fMRI: a review on methods in resting state connectivity analysis and resting state networks. Neuroradiol. J. 30, 305–317. doi: 10.1177/1971400917697342
Sood, A. (2013). On mind wandering, attention, brain networks, and meditation. Explore. 9, 136–141. doi: 10.1016/j.explore.2013.02.005
Spencer, J. L., Waters, E. M., Romeo, R. D., Wood, G. E., Milner, T. A., and McEwen, B. S. (2008). Uncovering the mechanisms of estrogen effects on hippocampal function. Front. Neuroendocrinol. 29, 219–237. doi: 10.1016/j.yfrne.2007.08.006
Stevens, M. C., and Clark, V. P. (2005). Low-dose estradiol alters brain activity. Psychiatry Res. 139, 199–217. doi: 10.1016/j.pscychresns.2005.04.004
Stiernman, L. J., Grill, F., Hahn, A., Rischka, L., Lanzenberger, R., Lundmark, V. P., et al. (2021). Dissociations between glucose metabolism and blood oxygenation in the human default mode network revealed by simultaneous PET-fMRI. Proc. Nat. Acad. Sci. U. S. A. 118, e2021913118. doi: 10.1073/pnas.2021913118
Sunavsky, A. (2020). Neuroimaging predictors of creativity in healthy adults. Neuroimage. 206, 116292. doi: 10.1016/j.neuroimage.2019.116292
Terasawa, E. (2018). Neuroestradiol in regulation of GnRH release. Horm. Behav. 104, 138–145. doi: 10.1016/j.yhbeh.2018.04.003
Thimm, M., Weis, S., and Hausmann, M. (2014). Menstrual cycle effects on selective attention and its underlying cortical networks. Neuroscience. 258, 307–317. doi: 10.1016/j.neuroscience.2013.11.010
Tozzi, A., and Bellingacci, L. (2020). Rapid estrogenic and androgenic neurosteroids effects in the induction of long-term synaptic changes: implication for early memory formation. Front. Neurosci. 14, 964. doi: 10.3389/fnins.2020.572511
Wang, J.-.X., Zhuang, J.-Y., Fu, L., and Lei, Q. (2020). Association of ovarian hormones with mapping concept of self and others in the brain's default mode network. Neuroreport 31, 717–723. doi: 10.1097/WNR.0000000000001477
Wei, J., Yang, F., Gong, C., and Shi, X. (2019). Protective effect of daidzein against streptozotocin-induced Alzheimer's disease via improving cognitive dysfunction and oxidative stress in rat model. J. Biochem. Mol. Toxicol. 33, 22319. doi: 10.1002/jbt.22319
Weis, S., and Hodgetts, S. (2019). Sex differences and menstrual cycle effects in cognitive and sensory resting state networks. Brain Cogn. 131, 66–73. doi: 10.1016/j.bandc.2017.09.003
Wen, Z., Hammoud, M. Z., Scott, J. C., Jimmy, J., Brown, L., Marin, M.-F., et al. (2021). Impact of exogenous estradiol on task-based and resting-state neural signature during and after fear extinction in healthy women. Neuropsychopharmacology 46, 2278–2287. doi: 10.1038/s41386-021-01158-4
Whitfield-Gabrieli, S. (2012). Default mode network activity and connectivity in psychopathology. Annu. Rev. Clin. Psychol. 8, 49–76. doi: 10.1146/annurev-clinpsy-032511-143049
Xue, J., Li, C., Quan, C., Lu, Y., Yue, J., and Zhang, C. (2017). Uncovering the cognitive processes underlying mental rotation: an eye-movement study. Sci. Rep. 7, 10076. doi: 10.1038/s41598-017-10683-6
Yang, J., and Gohel, S. (2020). Current methods and new directions in resting state fMRI. Clin. Imaging 65, 47–53. doi: 10.1016/j.clinimag.2020.04.004
Zhao, L. (2007). Estrogen receptor α and β differentially regulate intracellular Ca2+ dynamics leading to ERK phosphorylation and estrogen neuroprotection in hippocampal neurons. Brain Res. 1172, 48–59. doi: 10.1016/j.brainres.2007.06.092
Zhao, Y., Yu, Y., Zhang, Y., He, L., Qiu, L., Zhao, J., et al. (2017). Letrozole regulates actin cytoskeleton polymerization dynamics in a SRC-1 dependent manner in the hippocampus of mice. J. Steroid Biochem. Mol. Biol. 167, 86–97. doi: 10.1016/j.jsbmb.2016.11.013
Zimmerman, B., Kundu, P., Liu, Z., Urbanski, H. F., Kroenke, C. D., Kohama, S. G., et al. (2020). Longitudinal effects of immediate and delayed estradiol on cognitive performance in a spatial maze and hippocampal volume in menopausal macaques under an obesogenic diet. Front. Neurol. 11, 539. doi: 10.3389/fneur.2020.00539
Keywords: estrogen (17β-estradiol), rapid effects of steroids, cognitive task, magnetic resonance imaging (MRI), cognitive performance, resting state—fMRI, neuroprotection
Citation: Marchant I, Chabert S, Martínez-Pinto J, Sotomayor-Zárate R, Ramírez-Barrantes R, Acevedo L, Córdova C and Olivero P (2022) Estrogen, Cognitive Performance, and Functional Imaging Studies: What Are We Missing About Neuroprotection? Front. Cell. Neurosci. 16:866122. doi: 10.3389/fncel.2022.866122
Received: 30 January 2022; Accepted: 08 March 2022;
Published: 12 May 2022.
Edited by:
Teresa Jover-Mengual, University of Valencia, SpainReviewed by:
Lars Fester, Friedrich-Alexander-Universität Erlangen-Nürnberg, GermanyCopyright © 2022 Marchant, Chabert, Martínez-Pinto, Sotomayor-Zárate, Ramírez-Barrantes, Acevedo, Córdova and Olivero. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ivanny Carolina Marchant, aXZhbm55Lm1hcmNoYW50QHV2LmNs; Pablo Olivero, cGFibG8ub2xpdmVyb0B1di5jbA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.