
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol. , 03 March 2025
Sec. Fungal Pathogenesis
Volume 15 - 2025 | https://doi.org/10.3389/fcimb.2025.1547800
This article is part of the Research Topic Evolutionary adaptation in human-infecting fungi: Ecological traits and pathogenicity View all articles
 Sara Mina1*
Sara Mina1* Hajar Yaakoub2,3
Hajar Yaakoub2,3 Bienvenue Razafimandimby2
Bienvenue Razafimandimby2 Elske Dwars4Méline Wéry5
Elske Dwars4Méline Wéry5 Nicolas Papon2Wieland Meyer4
Nicolas Papon2Wieland Meyer4 Jean-Philippe Bouchara2
Jean-Philippe Bouchara2Background: Scedosporium species are filamentous fungi causing a wide spectrum of infections in healthy and debilitated individuals. Despite their clinical significance, the ecology of Scedosporium species remains understudied, particularly in the Middle East.
Methods: In this context, we conducted an environmental study to elucidate the distribution and ecological preferences of Scedosporium species in the North of Lebanon. One hundred and fifty-five soil samples were collected from different environmental areas and analyzed for several chemical parameters. Scedosporium isolates were then selected for species identification and genotyping.
Results: Overall, 39 (25.16%) were positive for Scedosporium species, with a predominance of S. apiospermum (80.56%). Soil analysis revealed associations between the fungal presence and pH, nitrogen, phosphorus, and organic matter content. Moreover, genotyping analysis using MultiLocus Sequence Typing identified five major clusters. Interestingly, a number of Lebanese isolates formed an Asian-specific cluster (V) with one clinical Chinese isolate, whereas two clusters (II and III) showed a close association with German isolates, and clusters (I and IV) contained isolates with a global distribution.
Conclusion: These findings provide new insights into the ecology of Scedosporium species, bridging a gap in our knowledge of their distribution on the Asian continent and laying the groundwork for future clinical investigations. Future international collaborations are essential to trace the origin of S. apiospermum.
Scedosporium species are emerging opportunistic pathogens causing a wide variety of infections in humans. Depending principally on the mode of transmission and the host immunity, infections can range from focal infections to severe disseminated infections (Ramirez-Garcia et al., 2018; Bouchara et al., 2020). Over the past two decades, these filamentous fungi have been recognized as serious agents in patients with cystic fibrosis (CF) (Cimon et al., 2000; Bouchara and Papon, 2019). All pertinent prevalence studies consistently demonstrate that Scedosporium species occupy the second rank among fungal agents responsible for airway colonization following Aspergillus fumigatus (Schwarz et al., 2018; Bouchara and Papon, 2019; Delhaes et al., 2019). Although usually well tolerated, colonization of the CF airways may lead to pulmonary exacerbations requiring hospitalization and to allergic broncho-pulmonary mycoses. In addition, they may be responsible for severe disseminated infections that may be fetal in the case of immunodeficiency (Bouchara and Papon, 2019). Difficulty in treating these infections is compounded by the fungal therapy-refractory nature and possession of miscellaneous virulence factors in favor of fungal invasion (Ramirez-Garcia et al., 2018; Bouchara and Papon, 2019; Mello et al., 2019). Therefore, enhanced understanding of the ecological niches of these fungi is a prerequisite for identifying the potential sources of contamination/infection and to prevent the airway colonization in predisposed patients.
Scedosporium species are saprophytic fungi frequently isolated from anthropogenic areas but rarely from natural environments (Soussi Abdallaoui et al., 2007; Rougeron et al., 2018; Hallouti et al., 2020; Mouhajir et al., 2020; Huang et al., 2023). The isolation of these fungi has been reported in several types of poorly aerated and nutrient-rich environments. One example is contaminated industrial soil with hydrocarbons. The abundance of these fungi in such environments is based on their ability to break down aromatic pollutants (April et al., 1998; Płaza et al., 1998; Naranjo et al., 2007; Janda-Ulfig et al., 2008; Covino et al., 2015), to tolerate heavy metals, such as cadmium (Płaza et al., 1998), as well as to produce fungistatic substances (Ko et al., 2010). Likewise, Scedosporium species have been reported in polluted water, like sewage sludge and swamps (Ulfig, 2003; Rainer and De Hoog, 2006), as well as in soil of parks and playgrounds in urban areas (Kaltseis et al., 2009; Harun et al., 2010; Pakshir et al., 2013; Rougeron et al., 2015; Luplertlop et al., 2016), thus underscoring the influence of anthropogenic factors on the ecology of these fungi. Agricultural soils rich in nitrogen are also potential biotopes for Scedosporium species favored by their ability to assimilate ammonium salts (Ajello, 1956; Babu et al., 2014; Nascimento Mdo et al., 2014; Alvarez and Sanhueza, 2016; Javidnia et al., 2022). Moreover, their tolerance to high NaCl concentrations explains their occurrence in salty water (Soussi Abdallaoui et al., 2007; Lan et al., 2014; Han et al., 2021). Additionally, studies have shown evidence of their occasional presence in animal droppings and bat guano (Rougeron et al., 2018). These findings suggested not only the possible use of these fungi as biomarkers of ecosystem exposure to human activities (Al-Yasiri et al., 2017), but also in bioremediation (Prenafeta-Boldú et al., 2006). Accordingly, researchers have pointed the use of fungal inoculum composed of Scedosporium species in the abatement of environmental pollutants (Naranjo et al., 2007). Otherwise, Scedosporium species have been very rarely reported from indoor environments (Rougeron et al., 2018). The only indoor reservoirs identified so far are potted plants. These fungi have been recovered by cultures from the soil of potted plants in a Canadian hospital (Summerbell et al., 1989) and also at the homes of patients with CF (Bouchara et al., 2020).
Presently, the genus Scedosporium encompasses 15 distinct species (Zhang et al., 2021) exhibiting varying clinical significance. Only seven of these species have been reported in the clinic, including S. apiospermum, S. boydii, and S. aurantiacum, and to a lesser extent S. americanum (Abrantes et al., 2021), S. ellipsoideum, S. minutisporum, and S. dehoogii. Several ecological studies have been performed across the continents, comprising Europe (Kaltseis et al., 2009; Rougeron et al., 2015), Africa (Nweze and Okafor, 2010; Mouhajir et al., 2020), America (Alvarez and Sanhueza, 2016; Elizondo-Zertuche et al., 2017), Australia (Harun et al., 2010) and Asia (Luplertlop et al., 2016; Luplertlop et al., 2019; Zhang et al., 2021; Huang et al., 2023), which revealed differences in the relative distribution of these species. For instance, S. apiospermum predominated in Bangkok (Luplertlop et al., 2016; Luplertlop et al., 2019), Taiwan (Huang et al., 2023), Austria and The Netherlands (Kaltseis et al., 2009), Mexico (Elizondo-Zertuche et al., 2017), and Morocco (Mouhajir et al., 2020). Conversely, S. dehoogii was the predominant species found in Western France (Rougeron et al., 2015), whereas S. aurantiacum was more prevalent in Australia (Harun et al., 2010).
Considering the absence of environmental surveys on Scedosporium species in the Middle East, we initiated a comprehensive study in this region. Here, we present the findings of an environmental study conducted in North Lebanon. In parallel to cultivation of the soil samples on a Scedosporium-selective culture medium, we assessed various soil chemical properties to define the ecophysiological characteristics of Scedosporium species. Additionally, genotyping using MultiLocus Sequence Typing (MLST) analysis was conducted to compare Lebanese isolates with isolates recovered from other geographical areas.
One hundred and fifty-five sites were selected according to various levels of human activities and natural environments. The areas exhibiting high human activity included: urban parks (n = 13), plant beds (n = 4), gardens (n = 8), port sites (n = 2), petrol stations (n = 25), residential areas (n = 19), roadsides (n = 12), agricultural fields (n = 28), pigsties (n = 3), refugee’s camps (n = 3), recreational areas (n = 6), landfill (n = 3) and industrial areas (n = 9). Natural regions, supposed to hold the lowest human impact, were represented by seashores (n = 5), forests (n = 10), and the Al Araneb island, a remote island part of the Palm Islands Nature Reserve located 5.5 km off the northwest coast of Tripoli (n = 5). After removing rough materials, such as plant debris and stones, soil samples were collected by mixing soil from 3-4 spots within a square meter, up to a depth of 15 cm, into sterile plastic bottles.
Soil samples were inoculated on agar plates according to Kaltseis et al (Kaltseis et al., 2009). with some adjustments. Briefly, 10 g of soil samples were suspended in 40 mL of sterile water and the resulting suspensions were shaken for 1 h at 37°C on a horizontal shaker (300 rpm). Afterward, 200-µL aliquots of the obtained suspensions were plated in triplicate onto Scedosporium selective culture medium (Scedo-Select III), which contains: 4-hydroxybenzoate 0.9 g/L; ammonium sulfate 5 g/L; potassium dihydrogen phosphate 1.25 g/L; magnesium sulfate 0.625 g/L; ferrous sulphate 0.01 g/L; dichloran 0.002 g/L; benomyl 0.008 g/L; chloramphenicol 0.5 g/L; and agar 20 g/L (Pham et al., 2015). Suspected colonies were stained with methylene blue and investigated microscopically on glass slides after 5-7 days of incubation at 35°C. Colonies of Scedosporium species were then counted and values of colony-forming unit (CFU) per gram of soil were determined for each sample. Isolates were then subcultured onto yeast extract-peptone-dextrose agar (YPDA) (yeast extract 5 g/L; peptone 10 g/L; glucose 20 g/L; agar 20 g/L; and chloramphenicol 0.5 g/L) plates which were incubated at 35°C.
Soil samples were air-dried and ground. Eight grams of soil were dispersed in 20 mL distilled water and throuroughly shaken for 2 min. The obtained mixture was then kept for 15 min at room temperature until further agitation. pH of the soil suspensions was measured using a pH meter. The phosphorus content was determined spectrophotometrically according to the Olsen method (Ryan et al., 2001). Total nitrogen content was determined by the Kjeldahl method (Bremner, 1960). The total organic matter (OM) content was determined by the reduction of potassium dichromate according to the Walkley-Black method (Walkley et al., 1934). The electrical conductivity was measured in a 1:5 soil-to-water suspension (Ryan et al., 2001). Potassium content was determined by flame photometry.
The mycelium from 9-day-old cultures on YPDA plates was collected by scraping the agar plates in distilled water. After centrifugation of the obtained suspension, the pellet was ground with glass beads in a mini-bead beater and then suspended by gentle vortexing in lysis buffer (10 mM Tris-HCl; 1 mM EDTA; 2% Triton X-100; 1% SDS; 0.1 M NaCl, pH 8). Genomic DNA was purified by phenol-chloroform method (Sigma-Aldrich, USA) and ethanol precipitation, then treated with RNase A (0.2 mg/mL) and stored at 4°C.
Molecular identification was performed by PCR amplification of a part of the ß-tubulin gene (BT2) (Gilgado et al., 2005). For S. aurantiacum identification, the internal transcribed spacer (ITS) 1 and 2 regions of the ribosomal DNA (rDNA) were amplified (Gilgado et al., 2005). PCR amplicons were purified using the NucleoSpin Extract II kit (Macherey-Nagel, France) before sequencing. Similarities between sequences were identified via BLASTn (Basic Local Alignment Search Tool) searches against the ITS and the BT2 sequences of the different Scedosporium species in the NCBI database, based on more than 97% sequence similarity. The obtained sequences were issued GenBank accession numbers provided in Supplementary Table S1.
MLST was conducted as previously described by Bernhardt et al (Bernhardt et al., 2013). Different pairs of primers were used to partially amplify the following genetic loci: calmodulin (CAL, exon 3–4), actin (ACT), the second largest subunit of RNA polymerase II gene (RPB2), ß-tubulin (TUB, exon 2–4) and the manganese superoxide dismutase (SOD2). The NucleoSpin Extract II kit (Macherey-Nagel) was used to purify the amplified products. They were then commercially sequenced in both directions and analyzed using the BioEdit sequence Alignment Editor (version 7.1.11). The sequences were trimmed as previously reported by Bernhardt et al (Bernhardt et al., 2013). All the obtained sequences were deposited in the GenBank database (Supplementary Table S2). Allele (AT) and sequence types (ST) were assigned according to the S. apiospermum MLST database at mlst.mycologylab.org. New ATs and STs were sequentially added to the database.
For phylogenetic analysis, multiple alignments were generated using ClustalOmega (v1.2.3). Nucleotide sequences were trimmed using trimAl (v1.4.rev15 build[2013-12-17]) (Capella-Gutiérrez et al., 2009). The concatenated DNA sequences of five MLST loci (ACT, CAL, RPB2, SOD2, TUB) from the different STs identified in this study were aligned using the MUSCLE algorithm in MEGA11 (version 11.0.13) along with the other S. apiospermum STs publicly available (Bernhardt et al., 2013; Wang et al., 2015; Matray et al., 2016; Chen et al., 2022) (Supplementary Table S2). MEGA11 was then used to construct a dendrogram with the best-fit model chosen according to the Bayesian information criterium (BIC). For the S. apiospermum STs, the Tamura 3-parameter with gamma distribution and invariant sites (T92+G+I) was used to calculate the dendrogram with a bootstrap analysis using 1,000 replications. The tree was obtained with MEGA11 and labeled using Adobe Illustrator 2024 (version 28.5).
Soil samples were classified into two groups: Scedosporium-positive and Scedosporium-negative samples. GraphPad Prism version 9.5.1 (GraphPad Software Inc., USA) was used to perform bivariate analysis. The mean, median, first and third quartiles were determined for each chemical parameter investigated. According to the D’Agostino-Pearson normality and Kolmogorov-Smirnov tests, all the chemical parameter distributions did not follow a normal distribution. The non-parametric Mann-Whitney test was then used to compare these data, with p < 0.05 for significance.
Results were also analyzed by Pearson’s correlation and non-metric multidimensional scaling (NMDS). Pearson’s correlation matrix (Heatmap) was carried out using IBM SPSS Statistics 27.0 to analyze the relationship between the presence of Scedosporium spp. in soil samples and each of the soil chemical properties. NMDS biplot was generated using XLSTAT 2024 (Lumivero, Denver, USA) to visualize the relationships between the presence of Scedosporium species and soil parameters using a nonparametric, distance-based approach.
Of the 155 soil samples, 39 (25.16%) showed growth of Scedosporium species. The percentage of positive samples was higher in the following environmental areas: pigsties (100%); refugee’s camps (66.67%); recreational areas, port sites, and plant beds (50%); industrial areas, agricultural areas, and the landfill (44.44%, 35.71%, and 33.33% respectively). Lower frequencies of Scedosporium species were reported from parks (23.08%), forests (20%), residential areas (15.79%), gardens (12.5%), petrol stations (12%), and roadsides (8.33%). In addition, no Scedosporium species were isolated from all samples collected from the Palm Islands Nature Reserve and seashores (Figure 1A).
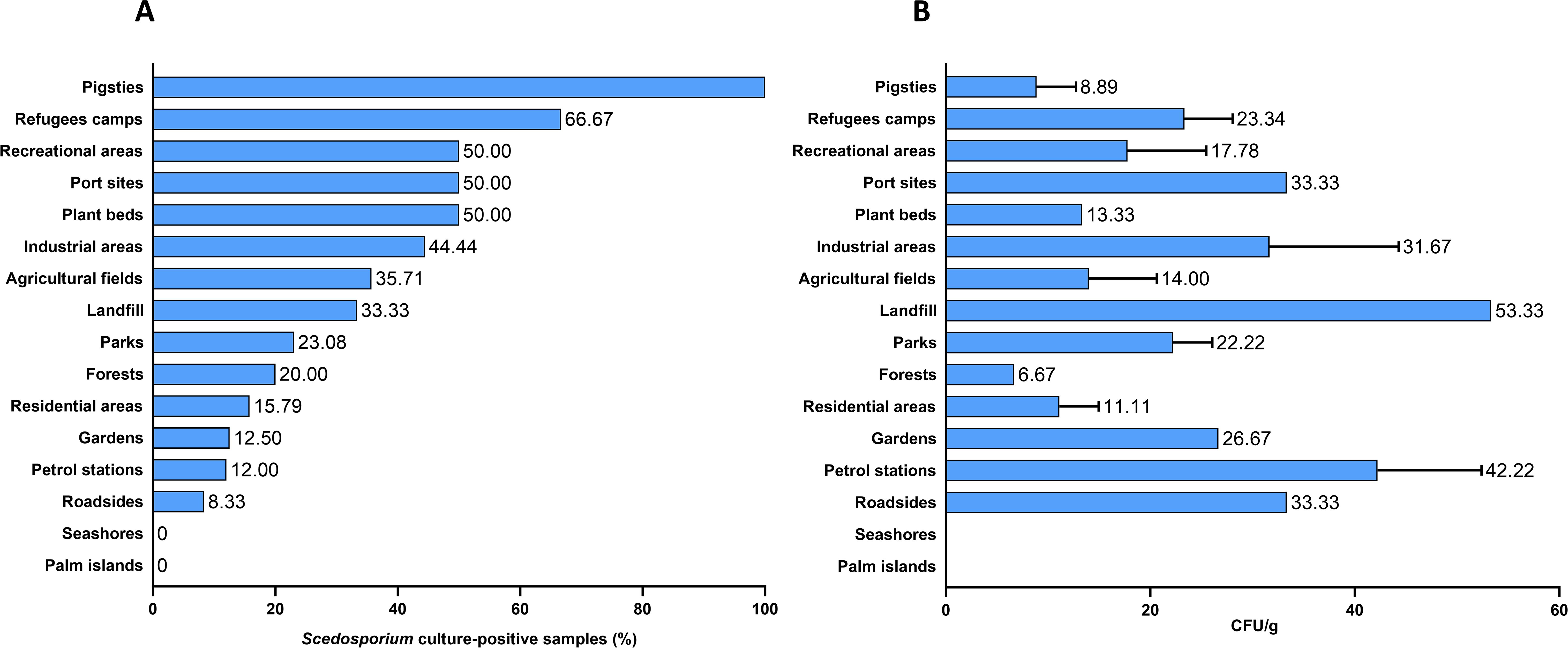
Figure 1. Frequency (A) and abundance (B) of Scedosporium species in human-impacted areas.
The average fungal burden (CFU) varied across samples. The highest CFU value of 53.33 CFU/g was observed in the landfill sample, followed by an average of 42.22 CFU/g in petrol station samples. Port sites, roadsides, and industrial areas showed average CFU values between 31.67 and 33.33 CFU/g. Forest samples exhibited the lowest fungal density, with an average value of 6.67 CFU/g (Figure 1B).
Total OM, pH, nitrogen, calcium, and phosphorus amounts were determined for all samples (Supplementary Table S3). Considering the pH, Scedosporium culture-positive samples exhibited a pH ranging from 6.2 to 7.87 (mean value: 6.98), whereas Scedosporium culture-negative samples collected from the Palm Islands and seashores exhibited an alkaline pH (> 8.19). As for the nutrients, Scedosporium isolates were found in soils with high nitrogen amounts (mean value: 0.21%), and very rich in phosphorus (mean value: 109.5 ppm) and potassium (mean value: 241.4 ppm). The calcium content of positive soil samples was highly variable, from 70 to 8,755 mg/L. In addition, Scedosporium culture-positive samples showed high OM content.
For each parameter, data distributions were compared between the two groups of soil samples using bivariate analysis (Figure 2). Statistical analysis revealed significant differences between the two groups for the pH, nitrogen, phosphorus, and organic matter content (p < 0.0001). Conversely, no differences were observed for potassium (p = 0.5798) and calcium (p = 0.2340) amounts, and electrical conductivity (p = 0.0812) between the two groups.
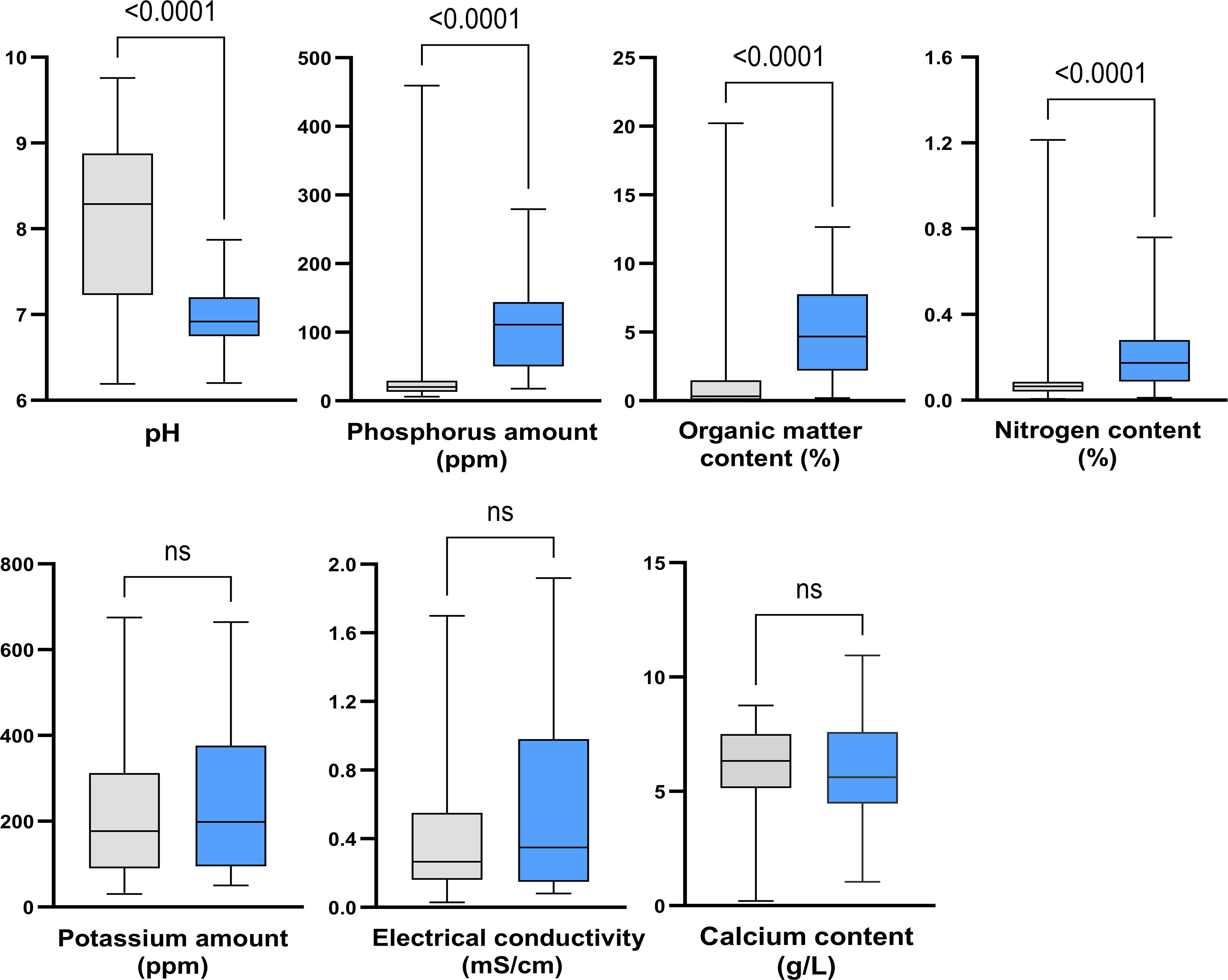
Figure 2. Impact of soil properties on the abundance of Scedosporium species. Grey, culture-negative samples; Blue, culture-positive samples. ns: not significant.
The correlation matrix highlighted a strong negative correlation between the presence of Scedosporium species and the absolute pH of soil samples (r= -0.5). In addition, moderate positive correlations were shown between the fungal growth and OM (r= 0.46), phosphorus (r= 0.43), EC (r= 0.26), and nitrogen (r= 0.3). Weaker correlations were found with calcium (r= 0.05) and potassium (r= 0.04) (Figure 3). Likewise, the two-dimension NMDS biplot (stress value =0.123) also revealed the influence of OM, phosphorus and nitrogen on the presence of Scedosporium species (Supplementary Figure S1).
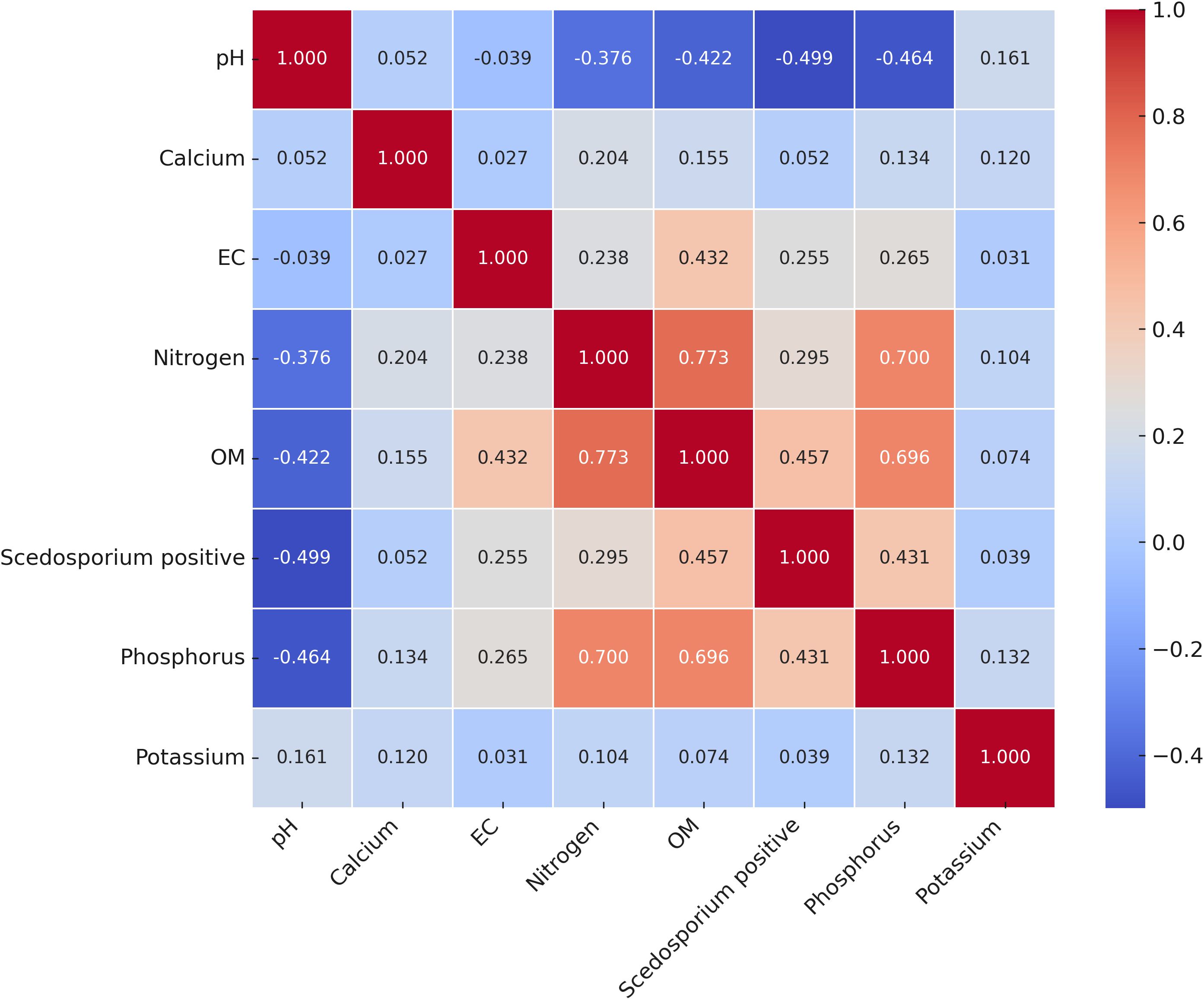
Figure 3. Pearson's correlation matrix (Heatmap) showing the relationship between soil chemical properties and the presence of Scedosporium species in soil samples. The scale with color gradient indicates the Pearson correlation coefficient values. Cooler colors represent negative correlations and warmer colors denote positive correlations.
Single culture fungal isolation was not possible for colonies obtained from 11 culture-positive samples because of the simultaneous growth of extensively growing fungi on the primary plates. Thirty-six colonies recovered from 28 soil samples, collected from urban and rural areas in North Lebanon during the dry (May-September) and wet seasons (October-December), were re-isolated for identification at species level (Table 1). S. apiospermum was the most abundant Scedosporium species (80.56%; 29/36 isolates). Moreover, S. apiospermum was recovered from all culture-positive samples except for refugee camps and port sites. S. boydii was recovered only once from a port site. Three isolates (8.33%) from different sites (recreational area, pigsty, and refugee camp) were identified as S. aurantiacum. Interestingly, BT2 sequences of three isolates recovered from agricultural fields could not be assigned to a particular Scedosporium species (Table 1).
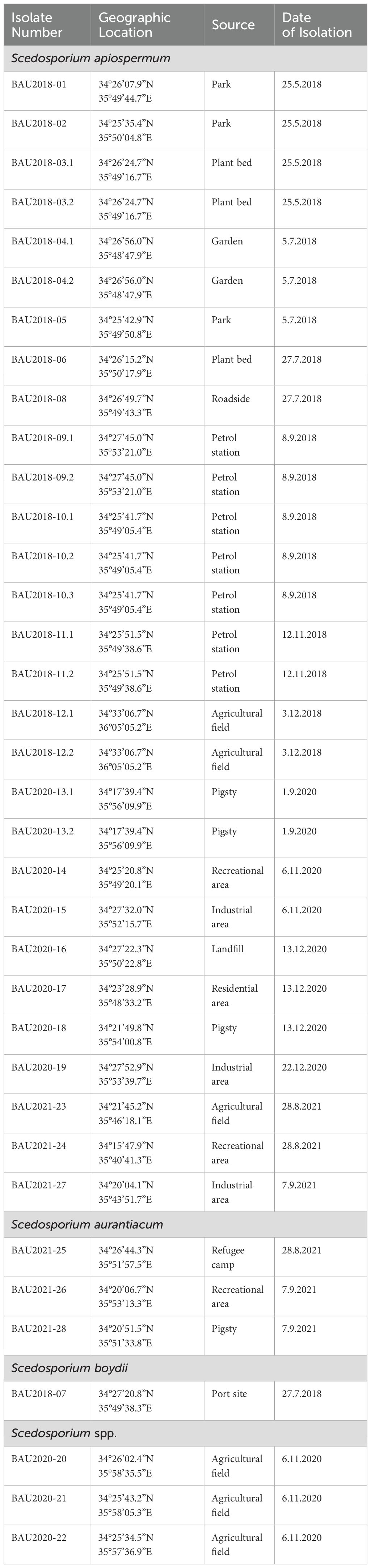
Table 1. List of Lebanese Scedosporium isolates, including geographic origin, source, and date of isolation.
Analysis of the combined sequences of the five MLST loci identified a high genetic diversity amongst S. apiospermum isolates. One new allele was obtained for both the ACT and CAL loci (Table 2). Three new alleles were identified for the RPB2 locus, seven for SOD2, and five for the TUB locus. Twenty-three new allelic combinations were identified, corresponding to new STs (ST44-ST66). Only two STs (ST13 and ST39) had been previously described. Moreover, two isolates from the same sampling site shared the same ST (ST56 for BAU2020-13.1 and BAU2020-13.2, or ST61 for BAU2018-10.2 and BAU2018-10.3). By contrast, isolates belonging to distinct STs sometimes were recovered from the same soil sample (for example, BAU2018-12.1 and BAU2018-12.2, which belonged to ST52 and ST55, respectively), whereas some isolates (BAU2020-16 and BAU2020-27) from different sampling sites exhibited the same ST (ST63).
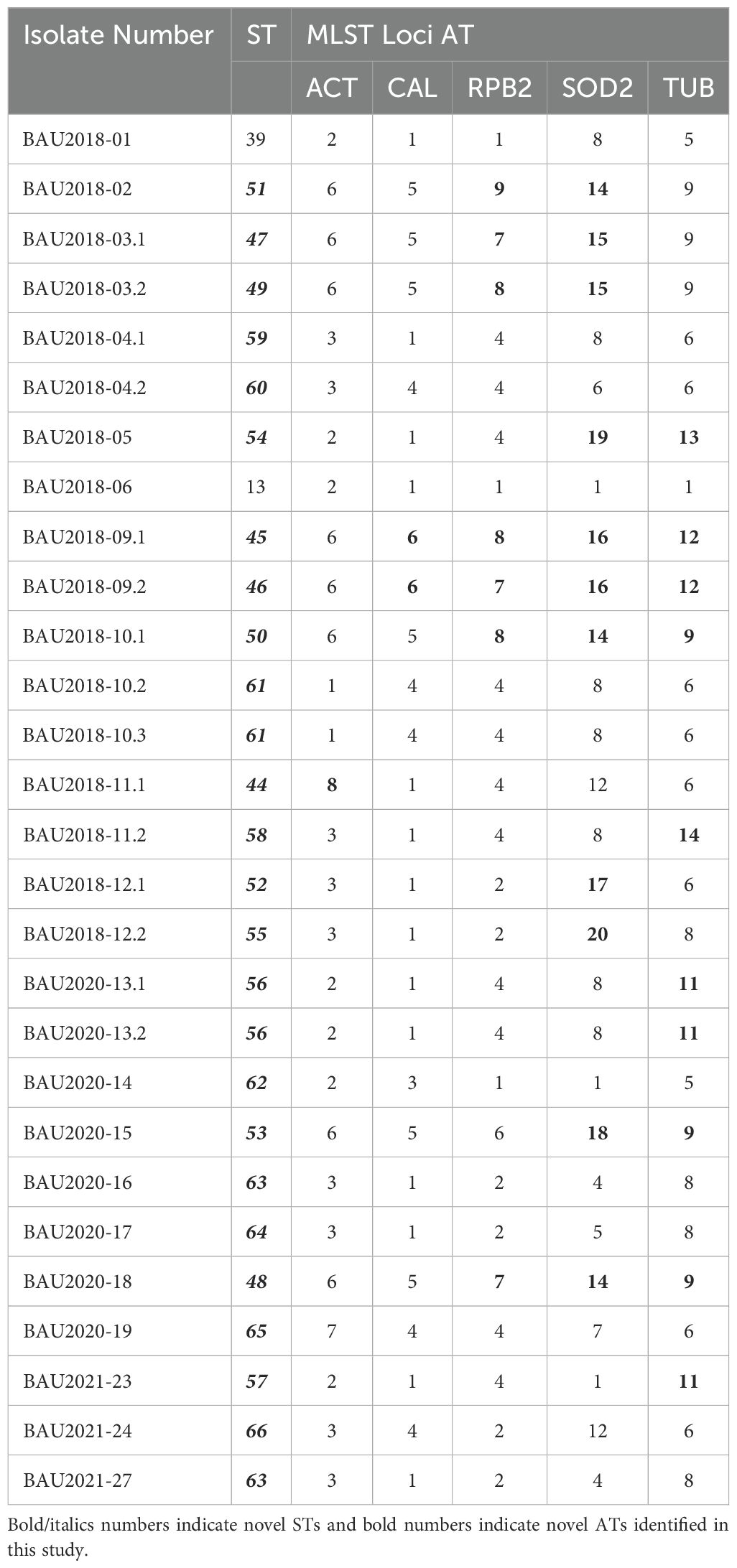
Table 2. List of MLST sequence types (ST), and allele types (AT) of Lebanese Scedosporium apiospermum isolates.
To place the herein identified STs, 72 isolates from 6 countries harboring different STs were included in the genetic relatedness analysis (Supplementary Table S2). The obtained dendrogram separated the isolates into five major clusters confirming the high genetic diversity observed amongst the Lebanese isolates (Figure 4). Cluster I comprised isolates from all the studied countries except Brazil, while cluster IV included a Brazilian isolate. Interestingly, clusters II and III comprised subsets of Lebanese isolates intermixed in both cases with one German isolate, and eight Lebanese isolates clustered together with one clinical isolate from China in cluster V, suggesting some geographical clustering. In addition, comparison of MLST data from Lebanese environmental isolates with those available for S. apiospermum isolates in the MLST database suggested some clustering with clinical isolates predominating in clusters I and IV. Nevertheless, MLST analysis should be conducted on a larger set of clinical and environmental S. apiospermum isolates from different continents and countries for definitive conclusions.
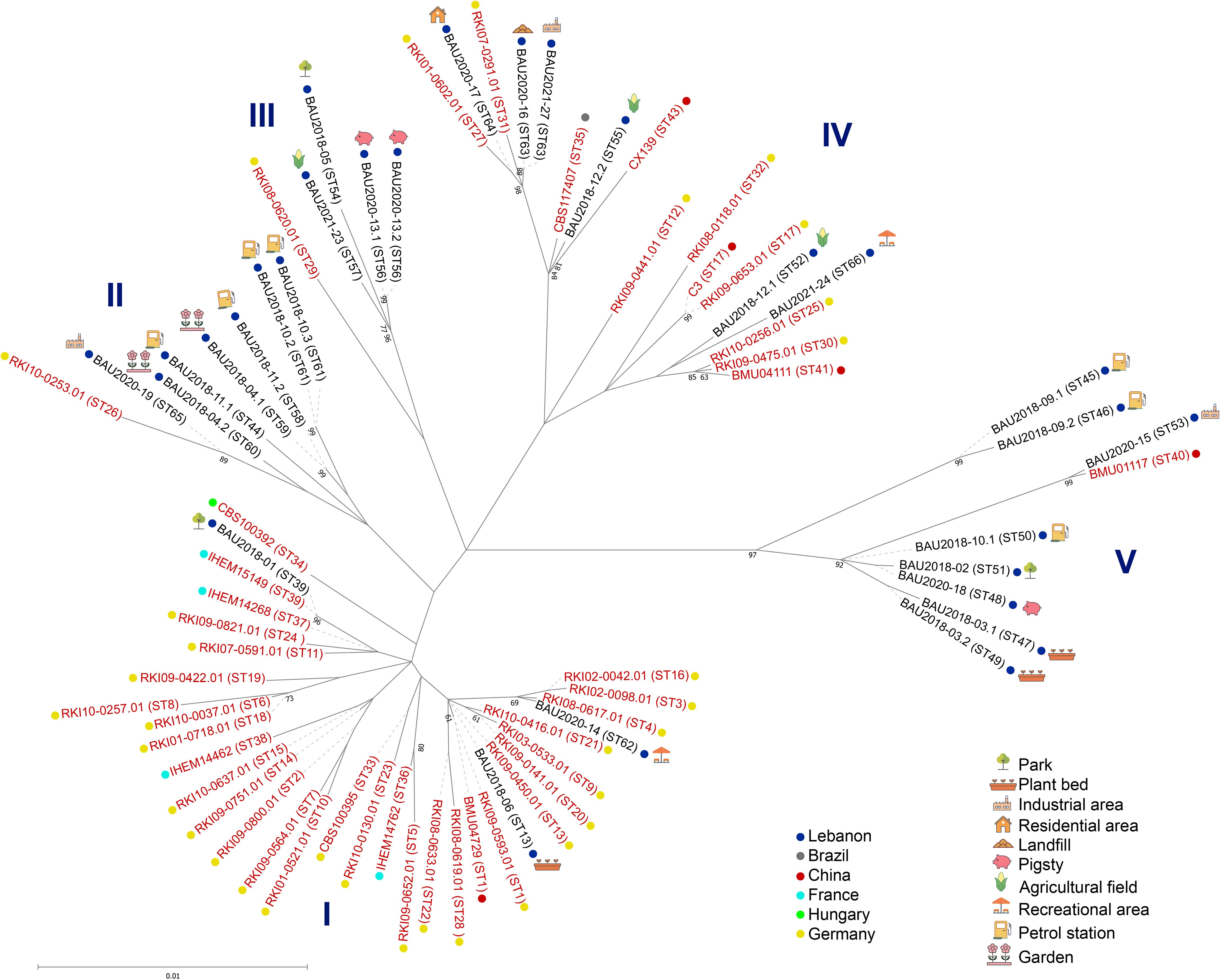
Figure 4. Maximum-likelihood tree based on concatenated MLST gene loci sequences of S. apiospermum isolates. Label colors indicate the clinical (red), environmental (black), or missing data (grey) origin of the isolates, and different dot colors indicate their geographical origin. ST: Sequence Type.
Scedosporium species have been isolated from different countries, with some geographical discrepancies in the distribution pattern of the species (Kaltseis et al., 2009; Harun et al., 2010; Rougeron et al., 2015), but no information on the distribution and environmental niches of Scedosporium species are available from the Middle East. We report herein for the first time data on the environmental distribution of Scedosporium species in North Lebanon. In the current study, 25.16% of soil samples were Scedosporium-culture positive, a frequency lower than those reported in Northwestern France, Taiwan, Morocco, and Thailand (40.33%, 48.4%, 48.5%, and 60.29% respectively) (Rougeron et al., 2015; Luplertlop et al., 2016; Mouhajir et al., 2020; Huang et al., 2023).
The observed CFU values of Scedosporium species indicate that human activity influences the abundance of these fungi in the environment. Indeed, samples from wild regions (seashores and Palm Islands) were negative for Scedosporium. This is in line with previous reports showing little occurrence of Scedosporium species in natural habitats or suburban areas (Soussi Abdallaoui et al., 2007; Harun et al., 2010; Hallouti et al., 2020; Mouhajir et al., 2020; Huang et al., 2023). Likewise, the highest values of fungal density were detected in samples obtained from petrol stations, indicating that elevated levels of petroleum hydrocarbons contribute to the proliferation of this fungus. This observation aligns with the findings of Kaltseis et al (Kaltseis et al., 2009), who noted the highest CFU counts in busy roads and petrol stations. Our findings support the hypothesis that Scedosporium species can serve as a marker for human activity.
In agreement with previous reports from Austria (Kaltseis et al., 2009), France (Rougeron et al., 2015), Thailand (Luplertlop et al., 2016; Rougeron et al., 2018), and Morocco (Mouhajir et al., 2020), Scedosporium species were mostly present in samples exhibiting a neutral pH (between 6.2 and 7.87). This preference for neutral pH values may also explain the absence of Scedosporium species in soil samples from seashores and Palm Islands which exhibited a higher pH. Among samples collected from forests, we also found two Scedosporium-culture positive samples, though they exhibited lower fungal burden. One culture-positive sample had previously been reported from forest samples in Morocco (Mouhajir et al., 2020). In Lebanon, access to forests in protected areas or natural reserves is controlled to limit human activities. Nevertheless, one cannot disregard totally a human impact as forests may be influenced by nearby agricultural practices, pollution, and recreational activities. These activities may introduce nutrients and alter environmental conditions, creating niches suitable for the growth of Scedosporium. Moreover, some Scedosporium strains were isolated from French Guiana termites suggesting the forests as natural habitat (Sorres et al., 2022). Additionally, the correlation matrix and NMDS analysis showed a correlation between the presence of Scedosporium species and three chemical parameters. Scedosporium presence in the soil was mainly correlated with increasing phosphorus amount, followed by elevated OM content, and nitrogen amount. Interestingly, our findings showed that culture-positive samples collected from agricultural fields and gardens contained high concentrations of OM. The use of fertilizers was extensively demonstrated in the literature as an important factor in the regulation of soil fungal communities; however, their impact on the distribution of fungal communities is still poorly recognized (Du et al., 2022). Another positive sample collected from Tripoli port, the second major seaport in Lebanon, showed increased concentrations of OM. This can be correlated with the presence of polychlorinated biphenyls (PCBs) previously detected in the sediments of Tripoli harbor (Merhaby et al., 2015). In addition, a strong negative correlation was seen between the presence of Scedosporium species and the pH of soil samples, with alkaline pH inhibiting the fungal growth which is line with a previous report from Mouhajir et al (Mouhajir et al., 2020).
Several molecular methods have been described to distinguish between Scedosporium species, including M13 PCR fingerprinting, quantitative PCR (qPCR), PCR-based reverse line blotting (PCR-RLB), and restriction fragment length polymorphism analysis after PCR amplification of the ITS regions (ITS-RFLP) (Delhaes et al., 2008; Lu et al., 2011). Nevertheless, sequencing methods remain the gold standard for accurate species identification, especially with the recent recognition of new Scedosporium species (Chen et al., 2022). In this study, the ITS 1 and 2 regions of rDNA and part of the β-tubulin gene were amplified to identify the isolates at the species level. S. apiospermum was the predominant species (80.55%) as previously found in Morocco (Mouhajir et al., 2020), Bangkok (Alastruey-Izquierdo et al., 2007), Mexico (Elizondo-Zertuche et al., 2017), Austria, The Netherlands (Kaltseis et al., 2009), and Taiwan (Huang et al., 2023). S. aurantiacum was recovered from 8.33% of the samples, which contrasts with its high frequency in Australia (Harun et al., 2010), and to a lesser extent in France (Rougeron et al., 2015), and Morocco (Mouhajir et al., 2020) (54.6%, 21.6%, and 17%, respectively). Finally, S. boydii was the least abundant species (2.7%), similarly to previous reports from Australia (2.1%) (Harun et al., 2010). Unlike previous ecological surveys conducted to investigate the distribution of Scedosporium species in different parts of the world, S. dehoogii was not retrieved from our samples. The most notable finding from this first ecological survey was the three unidentified Scedosporium isolates that are very close to the Thai undetermined isolates when considering the exons 5 and 6 of the β-tubulin gene (Luplertlop et al., 2016). Whole genome sequencing of these isolates should be conducted to determine if they could belong to a new species.
The five-loci MLST scheme is a highly reproducible and discriminatory tool for genotyping S. apiospermum species (Bernhardt et al., 2013). The herein reported study derived 25 distinct STs from 28 isolates by combining the five MLST loci sequences. Thus, nearly each isolate investigated showed an individual ST, exhibiting a high genetic diversity. Similar findings have been reported in other genotyping studies of S. aurantiacum (Harun et al., 2021). In addition, this MLST scheme was used to characterize the population structure using concatenated sequences from our isolates and other strains isolated from different countries. Interestingly, we found 23 novel STs, carrying novel alleles for the herein studied genetic loci, that can be crucial for future genomic and surveillance studies to describe strains encountered in clinical settings. Another notable finding is the close relationship between Lebanese isolates and a Chinese isolate in a specific Asian cluster (V), while two clusters (II and III) showed a close association between Lebanese and German isolates (Figure 3). In addition, this comparison suggested some clustering according to the source of the isolates, with clinical isolates predominating in clusters 1 and 4. However, this needs to be confirmed on a larger set of clinical and environmental isolates since it may be expected, as previously reported in a genotyping study of S. aurantiacum strains (Harun et al., 2021), a close relationship between environmental and clinical S. apiospermum isolates. Likewise given that the global population structure of S. apiospermum is still poorly understood, further studies expanding the number of S. apiospermum isolates of different origins are warranted to identify the origin of this species.
In this first ecological survey of the distribution and abundance of Scedosporium species in North Lebanon, we demonstrated that the occurrence of these fungi is associated with areas with high human activity and a neutral pH. These findings highlight the potential use of these fungi as environmental pollution indicators and as cost-effective bioremediators for polluted sites. We also showed that S. apiospermum emerged as the predominant species, with isolates displaying notable genetic diversity and geographical clustering alongside foreign clinical isolates. These results raise awareness of the exposure of susceptible individuals to these fungi and set the scene for further studies highlighting a possible correlation between environmental sources and clinical infections. Future studies, including extended MLST genotyping in other Asian countries, will be crucial to deepen our understanding of the global distribution and ecological impact of S. apiospermum, as previously achieved for S. aurantiacum (Harun et al., 2021).
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
SM: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Validation, Writing – original draft, Writing – review & editing. HY: Data curation, Formal Analysis, Writing – original draft, Writing – review & editing. BR: Investigation, Writing – review & editing. ED: Data curation, Formal Analysis, Writing – review & editing. MW: Formal Analysis, Writing – review & editing. NP: Resources, Validation, Writing – review & editing. WM: Methodology, Validation, Writing – review & editing. J-PB: Conceptualization, Data curation, Investigation, Methodology, Project administration, Resources, Validation, Writing – original draft, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Lebanese National Council for Scientific Research (CNRS-L) and Beirut Arab University (BAU) (Grant number: 12-05-18).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2025.1547800/full#supplementary-material
Supplementary Figure 1 | Non-metric Multidimensional Scaling (NMDS) biplot showing the relationship between soil parameters and the presence of Scedosporium species in soil samples. OM: Organic Matter; EC: Electrical Conductivity.
Abrantes, R. A., Refojo, N., Hevia, A. I., Fernández, J., Isla, G., Córdoba, S., et al. (2021). Scedosporium spp. from Clinical Setting in Argentina, with the Proposal of the New Pathogenic Species Scedosporium americanum. J. Fungi (Basel). 7, 160. doi: 10.3390/jof7030160
Ajello, L. (1956). Soil as natural reservoir for human pathogenic fungi. Science. 123, 876–879. doi: 10.1126/science.123.3203.876
Alastruey-Izquierdo, A., Cuenca-Estrella, M., Monzón, A., Rodriguez-Tudela, J. L. (2007). Prevalence and susceptibility testing of new species of Pseudallescheria and Scedosporium in a collection of clinical mold isolates. Antimicrob. Agents Chemother. 51, 748–751. doi: 10.1128/AAC.01177-06
Alvarez, E., Sanhueza, C. (2016). New record of Scedosporium dehoogii from Chile: Phylogeny and susceptibility profiles to classic and novel putative antifungal agents. Rev. Iberoam Micol. 33, 224–229. doi: 10.1016/j.riam.2016.03.007
Al-Yasiri, M. H., Normand, A. C., Mauffrey, J. F., Ranque, S. (2017). Anthropogenic impact on environmental filamentous fungi communities along the Mediterranean littoral. Mycoses. 60, 477–484. doi: 10.1111/myc.12612
April, T. M., Abbott, S. P., Foght, J. M., Currah, R. S. (1998). Degradation of hydrocarbons in crude oil by the ascomycete Pseudallescheria boydii (Microascaceae). Can. J. Microbiol. 44, 270–278. doi: 10.1139/w97-152
Babu, A. G., Kim, S. W., Yadhav, D. R., Adhikari, M., Kim, C., Lee, H. B., et al. (2014). A new record of Pseudallescheria boydii isolated from crop field soil in Korea. Mycobiology. 42, 397–400. doi: 10.5941/MYCO.2014.42.4.397
Bernhardt, A., Sedlacek, L., Wagner, S., Schwarz, C., Würstl, B., Tintelnot, K. (2013). Multilocus sequence typing of Scedosporium apiospermum and Pseudallescheria boydii isolates from cystic fibrosis patients. J. Cyst Fibros. 12, 592–598. doi: 10.1016/j.jcf.2013.05.007
Bouchara, J. P., Le Govic, Y., Kabbara, S., Cimon, B., Zouhair, R., Hamze, M., et al. (2020). Advances in understanding and managing Scedosporium respiratory infections in patients with cystic fibrosis. Expert Rev. Respir. Med. 14, 259–273. doi: 10.1080/17476348.2020.1705787
Bouchara, J. P., Papon, N. (2019). Scedosporium apiospermum. Trends Microbiol. 27, 1045–1046. doi: 10.1016/j.tim.2019.07.003
Bremner, J. M. (1960). Determination of nitrogen in soil by the Kjeldahl method. J. Agric. Sci. 55, 11–33. doi: 10.1017/S0021859600021572
Capella-Gutiérrez, S., Silla-Martínez, J. M., Gabaldón, T. (2009). trimAl: a tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 25, 1972–1973. doi: 10.1093/bioinformatics/btp348
Chen, M., Zhu, X., Cong, Y., Chen, H., Hou, Q., Hong, N., et al. (2022). Genotypic diversity and antifungal susceptibility of Scedosporium species from clinical settings in China. Mycoses. 65, 1159–1169. doi: 10.1111/myc.13507
Cimon, B., Carrère, J., Vinatier, J. F., Chazalette, J. P., Chabasse, D., Bouchara, J. P. (2000). Clinical significance of Scedosporium apiospermum in patients with cystic fibrosis. Eur. J. Clin. Microbiol. Infect. Dis. 19, 53–56. doi: 10.1007/s100960050011
Covino, S., D’Annibale, A., Stazi, S. R., Cajthaml, T., Čvančarová, M., Stella, T., et al. (2015). Assessment of degradation potential of aliphatic hydrocarbons by autochthonous filamentous fungi from a historically polluted clay soil. Sci. Total Environ. 505, 545–554. doi: 10.1016/j.scitotenv.2014.10.027
Delhaes, L., Harun, A., Chen, S. C., Nguyen, Q., Slavin, M., Heath, C. H., et al. (2008). Molecular typing of Australian Scedosporium isolates showing genetic variability and numerous S. aurantiacum. Emerg. Infect. Dis. 14, 282–290. doi: 10.3201/eid1402.070920
Delhaes, L., Touati, K., Faure-Cognet, O., Cornet, M., Botterel, F., Dannaoui, E., et al. (2019). Prevalence, geographic risk factor, and development of a standardized protocol for fungal isolation in cystic fibrosis: Results from the international prospective study “MFIP. J. Cyst Fibros. 18, 212–220. doi: 10.1016/j.jcf.2018.10.001
Du, S., Trivedi, P., Wei, Z., Feng, J., Hu, H. W., Bi, L., et al. (2022). The proportion of soil-borne fungal pathogens increases with elevated organic carbon in agricultural soils. mSystems. 7, e0133721. doi: 10.1128/msystems.01337-21
Elizondo-Zertuche, M., de-J-Treviño-Rangel, R., Robledo-Leal, E., Luna-Rodríguez, C. E., Martínez-Fierro, M. L., Rodríguez-Sánchez, I. P., et al. (2017). Molecular identification and in vitro antifungal susceptibility of Scedosporium complex isolates from high-human-activity sites in Mexico. Mycologia. 109, 874–881. doi: 10.1080/00275514.2017.1416260
Gilgado, F., Cano, J., Gené, J., Guarro, J. (2005). Molecular phylogeny of the Pseudallescheria boydii species complex: proposal of two new species. J. Clin. Microbiol. 43, 4930–4942. doi: 10.1128/JCM.43.10.4930-4942.2005
Hallouti, A., Ait Hamza, M., Zahidi, A., Ait Hammou, R., Bouharroud, R., Ait Ben Aoumar, A., et al. (2020). Diversity of entomopathogenic fungi associated with Mediterranean fruit fly (Ceratitis capitata (Diptera: Tephritidae)) in Moroccan Argan forests and nearby area: impact of soil factors on their distribution. BMC Ecol. 20, 64. doi: 10.1186/s12898-020-003342
Han, Y., Huang, X., Wang, Y., Du, J., Ma, K., Chen, Y., et al. (2021). Fungal community and biodeterioration analysis of Hull wood and its storage environment of the Nanhai No. 1 shipwreck. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.609475
Harun, A., Gilgado, F., Chen, S. C., Meyer, W. (2010). Abundance of Pseudallescheria/Scedosporium species in the Australian urban environment suggests a possible source for scedosporiosis including the colonization of airways in cystic fibrosis. Med. Mycol. 48 Suppl 1, S70–S76. doi: 10.3109/13693786.2010.515254
Harun, A., Kan, A., Schwabenbauer, K., Gilgado, F., Perdomo, H., Firacative, C., et al. (2021). Multilocus sequence typing reveals extensive genetic diversity of the emerging fungal pathogen Scedosporium aurantiacum. Front. Cell Infect. Microbiol. 11. doi: 10.3389/fcimb.2021.761596
Huang, Y. T., Hung, T. C., Fan, Y. C., Chen, C. Y., Sun, P. L. (2023). The high diversity of Scedosporium and Lomentospora species and their prevalence in human-disturbed areas in Taiwan. Med. Mycol. 61, myad041. doi: 10.1093/mmy/myad041
Janda-Ulfig, K., Ulfig, K., Cano, J., Guarro, J. (2008). A study of the growth of Pseudallescheria boydii isolates from sewage sludge and clinical sources on tributyrin, rapeseed oil, biodiesel oil and diesel oil. Ann. Agric. Environ. Med. 15, 45–49.
Javidnia, J., Badali, H., Haghani, I., Abastabar, M. (2022). A new record of Scedosporium dehoogii isolated from paddy field soil in Iran: Phylogeny and antifungal susceptibility profiles. Curr. Med. Mycol. 8, 27–31. doi: 10.32598/CMM.2023.1368
Kaltseis, J., Rainer, J., De Hoog, G. S. (2009). Ecology of Pseudallescheria and Scedosporium species in human-dominated and natural environments and their distribution in clinical samples. Med. Mycol. 47, 398–405. doi: 10.1080/13693780802585317
Ko, W. H., Tsou, Y. J., Ju, Y. M., Hsieh, H. M., Ann, P. J. (2010). Production of a fungistatic substance by Pseudallescheria boydii isolated from soil amended with vegetable tissues and its significance. Mycopathologia. 169, 125–131. doi: 10.1007/s11046-009-9237-1
Lan, W. J., Liu, W., Liang, W. L., Xu, Z., Le, X., Xu, J., et al. (2014). Pseudaboydins A and B: novel isobenzofuranone derivatives from marine fungus Pseudallescheria boydii associated with starfish Acanthaster planci. Mar. Drugs 12, 4188–4199. doi: 10.3390/md12074188
Lu, Q., Gerrits van den Ende, A. H., Bakkers, J. M., Sun, J., Lackner, M., Najafzadeh, M. J., et al. (2011). Identification of Pseudallescheria and Scedosporium species by three molecular methods. J. Clin. Microbiol. 49, 960–967. doi: 10.1128/JCM.01813-10
Luplertlop, N., Muangkaew, W., Pumeesat, P., Suwanmanee, S., Singkum, P. (2019). Distribution of Scedosporium species in soil from areas with high human population density and tourist popularity in six geographic regions in Thailand. PloS One 14, e0210942. doi: 10.1371/journal.pone.0210942
Luplertlop, N., Pumeesat, P., Muangkaew, W., Wongsuk, T., Alastruey-Izquierdo, A. (2016). Environmental screening for the Scedosporium apiospermum species complex in public parks in bangkok, Thailand. PloS One 11, e0159869. doi: 10.1371/journal.pone.0159869
Matray, O., Mouhajir, A., Giraud, S., Godon, C., Gargala, G., Labbé, F., et al. (2016). Semi-automated repetitive sequence-based PCR amplification for species of the Scedosporium apiospermum complex. Med. Mycol. 54, 409–419. doi: 10.1093/mmy/myv080
Mello, T. P., Bittencourt, V. C. B., Liporagi-Lopes, L. C., Aor, A.C., Branquinha, M.H., Santos, A.L.. (2019). Insights into the social life and obscure side of Scedosporium/Lomentospora species: ubiquitous, emerging and multidrug-resistant opportunistic pathogens. Fungal Biol. Rev. 33, 16–46. doi: 10.1016/j.fbr.2018.07.002
Merhaby, D., Net, S., Halwani, J., Ouddane, B. (2015). Organic pollution in surficial sediments of Tripoli harbour, Lebanon. Mar. pollut. Bull. 93, 284–293. doi: 10.1016/j.marpolbul.2015.01.004
Mouhajir, A., Poirier, W., Angebault, C., Rahal, E., Bouabid, R., Bougnoux, M. E., et al. (2020). Scedosporium species in soils from various biomes in Northwestern Morocco. PloS One 15, e0228897. doi: 10.1371/journal.pone.0228897
Naranjo, L., Urbina, H., De Sisto, A., Leon, V. (2007). Isolation of autochthonous non-white rot fungi with potential for enzymatic upgrading of Venezuelan extra-heavy crude oil. Biocatal Biotransformation. 25, 341–349. doi: 10.1080/10242420701379908
Nascimento Mdo, D., Leitão, V. M., Neto Silva, M. A., Maciel, L. B., Filho Muniz, W. E., Viana, G. M., et al. (2014). Eco-epidemiologic study of emerging fungi related to the work of babaçu coconut breakers in the State of Maranhão, Brazil. Rev. Soc. Bras. Med. Trop. 47, 74–78. doi: 10.1590/0037-8682-0237-2013
Nweze, E. I., Okafor, J. I. (2010). In vitro activity of conventional antifungal agents against Scedosporium apiospermum isolates recovered from clinical and environmental samples in Nigeria. Acta Microbiol. Immunol. Hung. 57, 209–214. doi: 10.1556/AMicr.57.2010.3.5
Pakshir, K., Ghiasi, M. R., Zomorodian, K., Gharavi, A. R. (2013). Isolation and molecular identification of keratinophilic fungi from public parks soil in Shiraz, Iran. BioMed. Res. Int. 2013, 619576. doi: 10.1155/2013/619576
Pham, T., Giraud, S., Schuliar, G., Rougeron, A., Bouchara, J. P. (2015). Scedo-Select III: a new semi-selective culture medium for detection of the Scedosporium apiospermum species complex. Med. Mycol. 53, 512–519. doi: 10.1093/mmy/myv015
Płaza, G., Lukasik, W., Ulfig, K. (1998). Effect of cadmium on growth of potentially pathogenic soil fungi. Mycopathologia. 141, 93–100. doi: 10.1023/a:1006991306756
Prenafeta-Boldú, F. X., Summerbell, R., Sybren de Hoog, G. (2006). Fungi growing on aromatic hydrocarbons: biotechnology’s unexpected encounter with biohazard? FEMS Microbiol. Rev. 30, 109–130. doi: 10.1111/j.1574-6976.2005.00007.x
Rainer, J., De Hoog, G. S. (2006). Molecular taxonomy and ecology of Pseudallescheria, Petriella and Scedosporium prolificans (Microascaceae) containing opportunistic agents on humans. Mycol Res. 110, 151–160. doi: 10.1016/j.mycres.2005.08.003
Ramirez-Garcia, A., Pellon, A., Rementeria, A., Buldain, I., Barreto-Bergter, E., Rollin-Pinheiro, R., et al. (2018). Scedosporium and Lomentospora: an updated overview of underrated opportunists. Med. Mycol 56, 102–125. doi: 10.1093/mmy/myx113
Rougeron, A., Giraud, S., Alastruey-Izquierdo, A., Cano-Lira, J., Le Gal, S., Nevez, G., et al. (2018). Ecology of Scedosporium species: knowledge and future research. Mycopathologia. 183, 185–200. doi: 10.1007/s11046-017-0200-2
Rougeron, A., Schuliar, G., Leto, J., Sitterlé, E., Landry, D., Bougnoux, M. E., et al. (2015). Human-impacted areas of France are environmental reservoirs of the Pseudallescheria boydii/Scedosporium apiospermum species complex. Environ. Microbiol. 17, 1039–1048. doi: 10.1111/1462-2920.12472
Ryan, J., Estefan, G., Rashid, A. (2001). Soil and plant analysis laboratory manual (Jodhpur: Scientific Publishers (India).
Schwarz, C., Bouchara, J. P., Buzina, W., Chrenkova, V., Dmeńska, H., de la Pedrosa, E. G. G., et al. (2018). Organization of patient management and fungal epidemiology in cystic fibrosis. Mycopathologia. 183, 7–19. doi: 10.1007/s11046-017-0205-x
Sorres, J., Hebra, T., Elie, N., Leman-Loubière, C., Grayfer, T., Grellier, P., et al. (2022). Antiparasitic ovalicin derivatives from Pseudallescheria boydii, a mutualistic fungus of French Guiana termites. Molecules. 27, 1182. doi: 10.3390/molecules27041182
Soussi Abdallaoui, M., Boutayeb, H., Guessous-Idrissi, N. (2007)Flore fongique du sable de deux plages à Casablanca (Maroc). Med. Mal. Infect. 17, 58–62. doi: 10.1016/j.mycmed.2006.12.001
Summerbell, R. C., Krajden, S., Kane, J. (1989). Potted plants in hospitals as reservoirs of pathogenic fungi. Mycopathologia. 106, 13–22. doi: 10.1007/BF00436921
Ulfig, K. (2003). Studies of keratinolytic and keratinophilic fungi in sewage sludge by means of a multi-temperature hair baiting method. Pol. J. Environ. Stud. 12, 461–466.
Walkley, A., Black, I., Armstrong, I. (1934). An examination of the Degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Science. 37, 29–37. doi: 10.1097/00010694-193401000-00003
Wang, H., Wan, Z., Li, R., Lu, Q., Yu, J. (2015). Molecular identification and susceptibility of clinically relevant Scedosporium spp. in China. BioMed. Res. Int. 2015, 109656. doi: 10.1155/2015/109656
Keywords: Scedosporium, ecology, MLST, genotypes, Lebanon
Citation: Mina S, Yaakoub H, Razafimandimby B, Dwars E, Wéry M, Papon N, Meyer W and Bouchara J-P (2025) First environmental survey of Scedosporium species in Lebanon. Front. Cell. Infect. Microbiol. 15:1547800. doi: 10.3389/fcimb.2025.1547800
Received: 18 December 2024; Accepted: 13 February 2025;
Published: 03 March 2025.
Edited by:
Rhys Alexander Farrer, University of Exeter, United KingdomReviewed by:
Çağrı Ergin, Pamukkale University, TürkiyeCopyright © 2025 Mina, Yaakoub, Razafimandimby, Dwars, Wéry, Papon, Meyer and Bouchara. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sara Mina, cy5taW5hQGJhdS5lZHUubGI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.