 Jinglin Zhao1
Jinglin Zhao1 Li Jiang
Li Jiang Liuli Wu
Liuli Wu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Infect. Microbiol., 06 December 2024
Sec. Clinical Microbiology
Volume 14 - 2024 | https://doi.org/10.3389/fcimb.2024.1484371
Sepsis is a systemic inflammatory response syndrome of multiorgan failure caused by dysregulation of the host response to infection and is a major cause of death in critically ill patients. In recent years, with the continuous development of sequencing technology, the intestinal microecology of this disease has been increasingly studied. The gut microbiota plays a host-protective role mainly through the maintenance of normal immune function and the intestinal barrier. Recent evidence suggests that intestinal flora dysbiosis plays a crucial role in sepsis. Clostridium butyricum (C. butyricum), which has been used as a probiotic in poultry feed since its discovery, has been found to play a potential protective role in intestinal infections, inflammatory bowel disease (IBD), colorectal cancer, and other diseases in recent studies. In this review, we continue to focus on the important role and mechanism of C. butyricum as a probiotic in human diseases, especially intestinal diseases. Additionally, we evaluate the research progress of C. butyricum in treatment of sepsis to identify more therapeutic targets for the clinical treatment of sepsis.
Sepsis is a systemic inflammatory response syndrome caused by various inflammatory factors that can involve multiple organs and has high morbidity and mortality rates (Salomão et al., 2019). It is estimated that sepsis affects approximately 30 million people globally each year, with more than 6 million deaths and a patient mortality rate of up to 30% (Srzić et al., 2022; Oczkowski et al., 2022). The current treatment for sepsis mainly consists of treating the infection with fluids and organ support and controlling the source of infection for appropriate resuscitation (Liu et al., 2022; Zampieri et al., 2023; Vincent, 2022). Although current treatment methods can alleviate the occurrence of septic shock to a certain extent, there is no significant improvement in the morbidity and sepsis remains a major threat to human health.
An increasing number of studies have shown that dysregulation is an important factor in sepsis and further accelerates the transformation of sepsis to septic shock (Xu et al., 2022; Yang et al., 2023). The intestinal microbiota of organisms can be broadly categorized into three groups, the first of which is the commensal flora, which mainly consists of Bacteroides, Clostridium, Bifidobacterium, and Lactobacillus (Yan and Wu, 2023). These bacteria account for more than 99% of the intestinal flora, form a good cooperative relationship with the host, assist in the digestion of a variety of foods, synthesize vitamins to enhance the body’s immunity, and maintain the ecological balance of the intestinal flora (Guo et al., 2020). The second category includes conditionally pathogenic flora, mainly enterococci and enterobacteria (Bowen et al., 2018). The number of this type of flora is not large, but it is unstable in the intestinal tract. When an organism is stimulated or the host’s immunity is low, this type of bacteria can transform into pathogenic bacteria, causing severe damage to the organism (Bowen et al., 2018; Kwiecinski and Horswill, 2020). The third category is pathogenic groups of bacteria, such as Salmonella and pathogenic E. coli. When these pathogenic bacteria enter the body and multiply, they cause a series of intestinal reactions in the body, leading to severe damage (Bäumler and Sperandio, 2016). Studies have shown (Haak and Wiersinga, 2017; Wang et al., 2019) that during sepsis, the intestinal microecosystem is significantly imbalanced, the beneficial flora of the body, such as Bacillus, Clostridium, Bifidobacterium and Lactobacillus, are significantly reduced, and the abundance of pathogenic bacteria is significantly increased (Xiao et al., 2023). This imbalance further exacerbates the damage to the intestinal barrier, causing pathogenic bacteria to enter the bloodstream and lymphatic system and spread throughout the body, leading to multiple organ failure, ultimately resulting in septic shock and death of the body (Chen et al., 2023; Deng et al., 2021) (Figure 1).
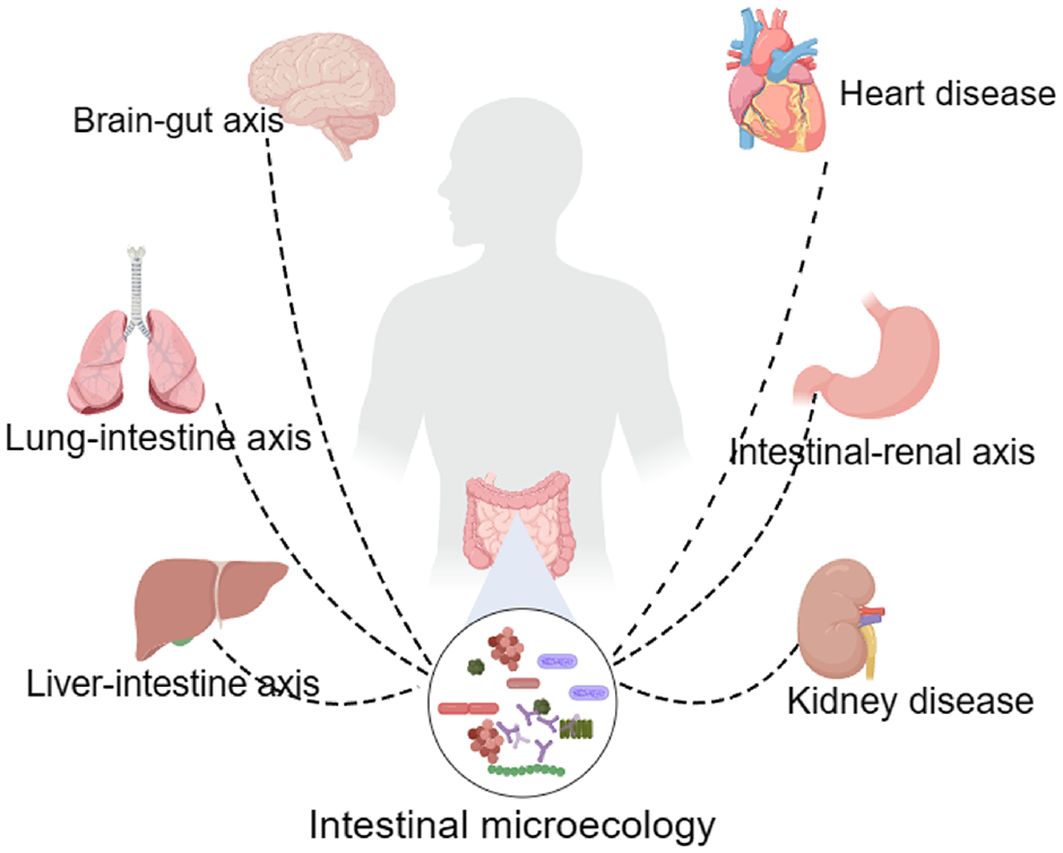
Figure 1. Intestinal microbiota affect multiple tissues and organs in the human body.
C. butyricum, also known as butyric acid bacteria, is an anaerobic bacillus with good tolerance to high temperature, high pressure and bile salts (Stoeva et al., 2021). Since its discovery by Dr. Konji Miyaji in Japan in 1933, research on C. butyricum has been gaining attention (Collins and East, 1998; Kanauchi et al., 2003). As a member of the Clostridium family, C. butyricum has a strong integumentary effect, inhibiting pathogenic bacteria in the intestinal tract and promoting the growth of beneficial bacteria in the intestinal tract, such as bifidobacteria and lactobacilli (Fenicia et al., 2002; Peck, 2009). On the one hand, the metabolites of C. butyricum, such as digestive enzymes, short-chain fatty acids (SCFAS) and amino acids, can protect the stability of the intestinal microecology and improve feed utilization (Cao et al., 2022). On the other hand, C. butyricum produces a large amount of butyric acid through metabolism to protect intestinal health and promote the repair and regeneration of intestinal epithelial cells (Pouillart, 1998). As a probiotic, C. butyricum has the ability to nourish the body, regulate the intestinal microecology, and enhance the immunity of the body; although the specific mechanism has not been fully elucidated, the available evidence suggests its potential as a promising clinical therapeutic candidate. Several studies have shown that C. butyricum has a certain preventive effect on inflammatory bowel disease (IBD), colon cancer (Budu et al., 2022; Liu et al., 2020), obesity (Obanda et al., 2020), diabetes (Lee et al., 2022) and other diseases; among them, the MIYAIRI-588 strain is widely used in Japan, China and other regions for the treatment of antibiotic-associated diarrhoea, and existing C. butyricum live-bacterial microecological preparations (Atenin, Misan, and Changlakang) have achieved some efficacy in the treatment of diarrhoea, IBD, and intestinal tumors (Zhang et al., 2023; Ariyoshi et al., 2020) (Figure 2). Therefore, in this narrative review, we focus on the specific mechanism of C. butyricum regulating intestinal microecology in various diseases and its current research in sepsis and analyze its application prospect in sepsis treatment in the future.
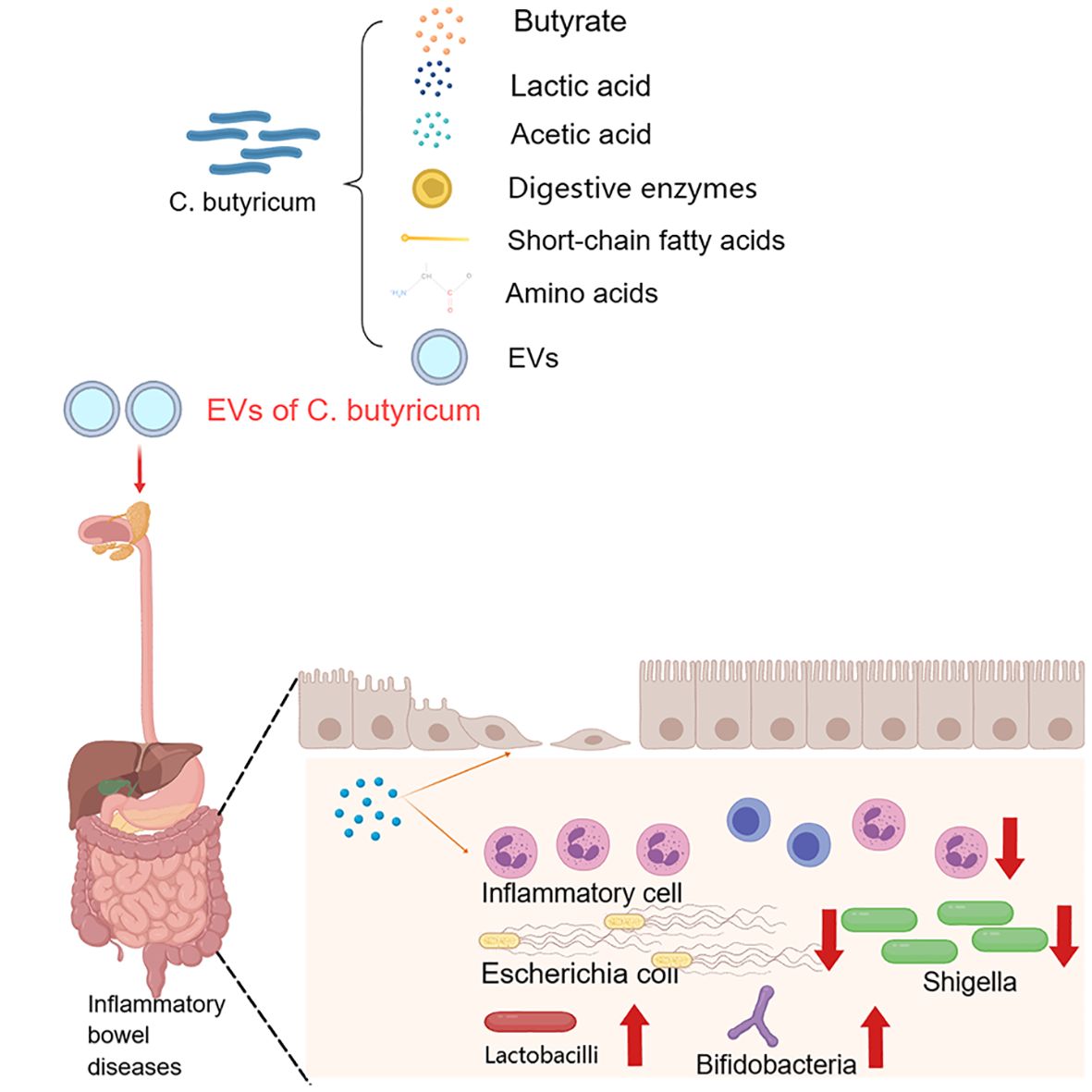
Figure 2. The effect of Clostridium butyricum on intestinal diseases and its possible mechanisms.
Inflammatory bowel diseases (IBD), including ulcerative colitis and Crohn’s disease, are specific inflammatory lesions that occur in the rectum and colon (Adolph et al., 2022). Disruption of the intestinal epithelial barrier leads to increased permeability and infiltration of pathogens and further compromises the body’s immune system, leading to the development of IBD (Basso et al., 2014). An experimental animal study showed (Ma et al., 2022) that extracellular vesicle of C. butyricum-derived metabolites play an important role in alleviating colitis. Administering extracellular vesicles of C. butyricum to mice with IBD resulted in a dual effect; on the one hand, it reduced inflammatory cell infiltration and mucus layer damage in the colon, significantly enhancing the integrity of the intestinal mucus barrier; on the other hand, it modulated the composition of the intestinal microflora, significantly decreasing the abundance of intestinal pathogenic bacteria, such as Shigella and Escherichia coli. Transcriptome sequencing revealed a significant increase in tryptophan content in mice treated with extracellular vesicles, which play an important role in immunity, neuronal function, and intestinal homeostasis regulation. This finding suggested that extracellular vesicles of C. butyricum metabolites may be potentially important targets for the treatment of IBD. Moreover, in another study, C. butyricum was found to inhibit TLR2 signaling and IL-17 secretion and enhance the local mucosal immune system, thus exerting a protective effect against intestinal inflammation (Xie et al., 2020).
An increasing number of studies have also investigated the relationship between intestinal flora dysbiosis and the occurrence and development of intestinal tumors (Ge et al., 2021; Wong and Yu, 2019). In colorectal cancer patients, researchers found that C. butyricum significantly inhibited the development of intestinal tumors in APC-positive mice induced by a high-fat diet (HFD) (Zhou et al., 2017). In addition, mechanistic studies have shown that C. butyricum inhibits the Wnt/β-linker signaling pathway and modulates the composition of the gut microbiota, significantly reducing the abundance of pathogenic bacteria, such as Escherichia coli and Shigella, and increasing the content of probiotics, such as Lactobacillus, and their metabolites, such as free fatty acids (Chen et al., 2020). In another study (Zhou et al., 2022), FISH revealed that C. butyricum was mainly enriched in small intestinal crypts and colonic tumor tissues in a mouse model of colorectal cancer and may inhibit tumor progression by infiltrating into colonic tumor tissues and acting directly on colorectal cancer cells. Moreover, colorectal cancer cell lines were cocultured with C. butyricum to detect the expression of the MyD88 and Nuclear factor kappa-B (NF-κB) genes, and the expression of the two genes in the colorectal cancer cell lines was reduced by C. butyricum treatment, which suggested that C. butyricum may inhibit the progression of colorectal cancer by regulating the expression of the MyD88 and NF-κB genes.
In summary, C. butyricum plays an important role in various intestinal diseases, and its related molecular mechanism has also undergone preliminary exploration. Moreover, as C. butyricum is a probiotic, its corresponding products have been developed for the prevention and treatment of other diseases, such as improving the metabolic capacity of pancreas, adipose tissue, and liver to protect physical health (Li et al., 2023), restored the diminished efficacy of ICB and improved survival in lung cancer patients (Tomita et al., 2022). Resisting type 2 diabetes and reducing obesity (Stoeva et al., 2021). However, whether C. butyricum plays an important role in sepsis, as well as its potential mechanism, must be further investigated.
Sepsis is one of the most common serious illnesses in intensive care units. The pathogenesis of sepsis is extremely complex and mainly involves immune disorders, mitochondrial damage, coagulation disorders, inflammatory dysregulation, endoplasmic reticulum stress and autophagy (Gotts and Matthay, 2016; Jacobi, 2022). The intestinal flora also plays an important role in these processes. According to research reports, healthy intestinal flora can improve the immune function of sepsis patients, enhance host resistance, and prevent the occurrence of septic shock as well as systemic multiorgan failure syndrome (Gong et al., 2019; Sun et al., 2023). At the same time, healthy intestinal flora also produces beneficial metabolites (e.g.,SAFCs), regulate endoplasmic reticulum stress, improve multiorgan functional damage, change the intestinal microenvironment, enhance the body’s intestinal barrier function, prevent bacterial translocation, and so on. Compared with that of healthy people, the intestinal flora of patients with sepsis is significantly changed, mainly manifested by a decrease in the diversity of the intestinal flora, an increase in the abundance of a large number of pathogenic bacteria, such as Escherichia coli and Shigella, and a decrease in the abundance of beneficial bacteria, such as Lactobacillus (Cuna et al., 2021; Haussner et al., 2019). Moreover (Valdés-Duque et al., 2020) dysbiosis of the intestinal flora in sepsis patients further promotes the progression of sepsis to septic shock by decreasing the levels of metabolites, such as free fatty acids, and initiating the inflammatory immune response, which further leads to systemic multiorgan failure.
As a specialized anaerobic gram-positive bacillus, the metabolites of C. butyricum include SCFAS such as butyric acid, acetic acid, lactic acid, B vitamins, digestive enzymes and folic acid, which can regulate the balance of the intestinal flora, promote the proliferation of beneficial intestinal flora, inhibit the propagation of harmful bacteria, and have a strong rectification effect (Gao et al., 2023). Butyric acid, one of the most important metabolites of C. butyricum, can significantly stimulate intestinal epithelial papillary hyperplasia and villus growth and promote intestinal epithelial cell healing, and it has the greatest impact on the physiology and immunity of the human body through a variety of SCFAS (Bruce et al., 2014; Hsiao et al., 2021). Macfarlane CT et al. (Macfarlane and Macfarlane, 2011) showed that an organism is in a hypoxic state when sepsis occurs, and intestinal epithelium ischemia and anoxia lead to cellular damage and mucosal barrier disruption. Kripke et al. (1989). also found that after gavage of 40 mmol/ml butyric acid, the DNA content of the intestinal mucosal tissue was significantly greater in mice than in control mice. The DNA content of the mucosal tissues increased significantly, confirming that butyric acid can stimulate the proliferation of colonic mucosal epithelial cells and ameliorate the damage to intestinal tissues induced by sepsis. In another in vivo study in septic rats (Wang et al., 2015), it was found that compared to that in septic control rats, pathological damage to the intestinal mucosa in septic rats was reduced after butyrate treatment, and the expression of proinflammatory cytokine mRNA was significantly decreased. A further increase in the treatment dose revealed that a high dose of butyric acid significantly activated Caspase-1, elevated the expression of Caspase-1 and its shear bodies P20 and NLRP3, increased apoptosis in the intestinal lamina propria, and ameliorated sepsis-induced intestinal mucosal tissue injury (Figure 3). Moreover, a recent study from Japan also found a significant decrease in the abundance of Clostridiales genus on the first and seventh days of sepsis mouse models, and its corresponding gut microbiota metabolites such as SCFAS and butyric acid also decreased significantly (Muratsu et al., 2022).

Figure 3. Clostridium butyricum plays an intestinal regulatory role in sepsis.
In addition to its intestinal regulatory effects, butyric acid also has protective effects on septic organs. Animal studies have shown that intravenous injection of butyric acid in a rat Cecum Ligation AND Puncture (CLP) sepsis model can significantly reduce the mRNA level of HMGB1 in rat tissues, and the serum alanine aminotransferase (ALT), creatinine (Cre), and pulmonary peroxidase (MPO) levels were significantly lower in the CLP group than in the control group, which confirms that butyric acid can significantly alleviate systemic multiorgan failure caused by sepsis (Zhang et al., 2007). HMGB1, a member of the high mobility family of proteins, can be secreted to the extracellular level to participate in the activation of the inflammatory response and induce endothelial cells to express adhesion molecules for chemotaxis of inflammatory cells, such as tissue extravasation, thus aggravating the systemic inflammatory response syndrome of sepsis (Naglova and Bucova, 2012). Butyric acid, an inhibitor of the enzyme deacetylase (HDAC), can inhibit deacetylation to cause acetylation of the transcription factor NF-κB, thereby attenuating NF-κB-mediated systemic inflammatory response syndrome. NF-κB-mediated transcription of the HGMB1 gene ultimately reduces the mRNA level of HMGB1 and attenuates the systemic inflammatory response syndrome induced by sepsis (Quivy and Van Lint, 2004; Shirasugi et al., 2018). In a mouse model of sepsis brain injury, researchers found that compared with those in the sepsis group, the water maze-generated mice in the sodium butyrate pretreatment group exhibited a shorter escape latency and a greater number of shuttle platforms, which significantly improved neuronal degeneration in the hippocampus of the mice in the sepsis group and attenuated sepsis-induced brain injury in the mice (Bi et al., 2019; Lisman et al., 2017). Moreover, several studies have shown that in sepsis models, intraperitoneal injection of 200 mg/kg sodium butyrate can improve acute lung injury (Zhang et al., 2010), sepsis-induced myocardial inhibition (de Lazari et al., 2020), intestinal barrier function (Rivière et al., 2016), and other functions caused by sepsis. These findings further suggest that C. butyricum plays a modulating role in the treatment of sepsis (Figure 4).
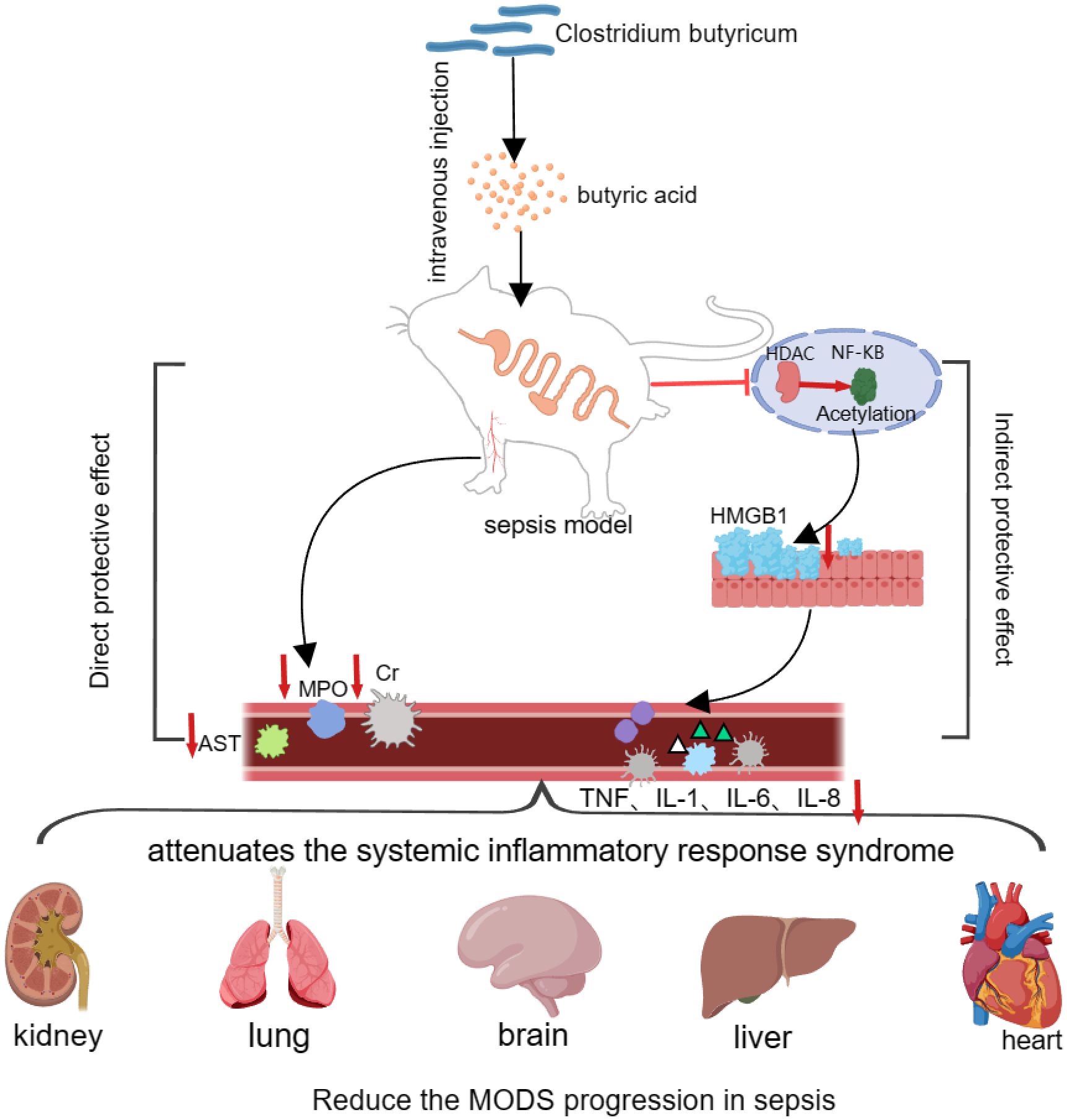
Figure 4. Clostridium butyricum plays an organ-protective role in sepsis.
In addition to its extremely strong rectifying effect, C. butyricum also plays an important role in the immunomodulatory function of sepsis. According to a previous study (Zhang et al., 2016; Xie et al., 2020), C. butyricum can activate dendritic cells and macrophages in the intestinal tract through the Toll-like receptor 2 (TLR2) pathway and promote the secretion of the transforming growth factor β (TGF-β), which further mediates the differentiation of initial T cells to regulatory T cells (Treg), and the interleukin 10 (IL-10) produced by Tregs inhibits the proinflammatory response mediated by Th1 and Th 17, thus exerting anti-inflammatory effects. Similarly, in another animal study, sepsis-related lung injury was established in mice by CLP and treated with butyrate gavage after surgery. The results of the study found that butyrate effectively improved the survival rate and alleviated lung damage in CLP mice, and the main mechanism may be that butyrate increased the number of CD4 + Foxp3 + Tregs and enhanced the barrier function of the gut and lungs (Wei et al., 2024).
This review also has several limitations. According to the literature, the protective effect of C. butyricum on sepsis is still in the basic research stage in animal models. Whether C. butyricum plays a protective role in humans remains to be fully explored in clinical experiments. We look forward to further research on the preventive and protective effects of C. butyricum on sepsis in the future, providing new insights for the treatment of sepsis patients.
Intestinal microecology is a new direction for treatment of sepsis. In view of the important role of C. butyricum in the maintenance of intestinal homeostasis and immune regulation, this review provides theoretical support for the future application of C. butyricum in treatment of sepsis, and we believe that C. butyricum could become a new star in treatment of sepsis in the future.
JZ: Writing – original draft, Writing – review & editing. LJ: Writing – original draft. WH: Writing – review & editing. DH: Writing – review & editing. XY: Writing – review & editing. LW: Writing – original draft, Writing – review & editing. HZ: Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by the Kunming Medical Joint Project (202401AY070001-182) and the High-Quality Professional Youth Introduction Project of Kunming City (None).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
C. butyricum, Clostridium butyricum; IBD, Inflammatory bowel diseases; SCFAS, short-chain fatty acids; CLP, Cecum Ligation and Puncture; ALT, alanine aminotransferase; Cre, creatinine; TLR2, Toll-like receptor 2; TGF-β, Transforming growth factor β; IL-10, interleukin 10; Treg, T cells to regulatory T cells; NF-κB, Nuclear factor kappa-B.
Adolph, T. E., Meyer, M., Schwärzler, J., Mayr, L., Grabherr, F., Tilg, H. (2022). The metabolic nature of inflammatory bowel diseases. Nat. Rev. Gastroenterol. Hepatol. 19, 753–767. doi: 10.1038/s41575-022-00658-y
Ariyoshi, T., Hagihara, M., Eguchi, S., Fukuda, A., Iwasaki, K., Oka, K., et al. (2020). Clostridium butyricum MIYAIRI 588-induced protectin D1 has an anti-inflammatory effect on antibiotic-induced intestinal disorder. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.587725
Basso, D., Zambon, C. F., Plebani, M. (2014). Inflammatory bowel diseases: from pathogenesis to laboratory testing. Clin. Chem. Lab. Med. 52, 471–481. doi: 10.1515/cclm-2013-0588
Bäumler, A. J., Sperandio, V. (2016). Interactions between the microbiota and pathogenic bacteria in the gut. Nature 535, 85–93. doi: 10.1038/nature18849
Bi, W., Lan, X., Zhang, J., Xiao, S., Cheng, X., Wang, H., et al. (2019). USP8 ameliorates cognitive and motor impairments via microglial inhibition in a mouse model of sepsis-associated encephalopathy. Brain Res. 1719, 40–48. doi: 10.1016/j.brainres.2019.05.009
Bowen, W. H., Burne, R. A., Wu, H., Koo, H. (2018). Oral biofilms: pathogens, matrix, and polymicrobial interactions in microenvironments. Trends Microbiol. 26, 229–242. doi: 10.1016/j.tim.2017.09.008
Bruce, T., Leite, F. G., Tschoeke, D. A., Miranda, M., Pereira, N., Jr., Valle, R., et al. (2014). Exploring the genome of a butyric acid producer, clostridium butyricum INCQS635. Genome Announc 2, e01169–14. doi: 10.1128/genomeA.01169-14
Budu, O., Banciu, C., Pinzaru, I., Sarău, C., Lighezan, D., Șoica, C., et al. (2022). A Combination of Two Probiotics, Lactobacillus sporogenes and Clostridium butyricum, Inhibits Colon Cancer Development: An In Vitro Study. Microorganisms 10, 1692. doi: 10.3390/microorganisms10091692
Cao, W., Zheng, C., Xu, X., Jin, R., Huang, F., Shi, M., et al. (2022). Clostridium butyricum potentially improves inflammation and immunity through alteration of the microbiota and metabolism of gastric cancer patients after gastrectomy. Front. Immunol. 13. doi: 10.3389/fimmu.2022.1076245
Chen, D., Jin, D., Huang, S., Wu, J., Xu, M., Liu, T., et al. (2020). Clostridium butyricum, a butyrate-producing probiotic, inhibits intestinal tumor development through modulating Wnt signaling and gut microbiota. Cancer Lett. 469, 456–467. doi: 10.1016/j.canlet.2019.11.019
Chen, Y., Liao, Y., Chen, X., Fan, H., Lin, D., Zheng, T., et al. (2023). Recognition and management of hemorrhaging in combination with emerging enterogenic sepsis during a hepatectomy: a case report. BMC Anesthesiol 23, 186. doi: 10.1186/s12871-023-02108-x
Collins, M. D., East, A. K. (1998). Phylogeny and taxonomy of the food-borne pathogen Clostridium botulinum and its neurotoxins. J. Appl. Microbiol. 84, 5–17. doi: 10.1046/j.1365-2672.1997.00313.x
Cuna, A., Morowitz, M. J., Ahmed, I., Umar, S., Sampath, V. (2021). Dynamics of the preterm gut microbiome in health and disease. Am. J. Physiol. Gastrointest Liver Physiol. 320, G411–g419. doi: 10.1152/ajpgi.00399.2020
de Lazari, M. G. T., Pereira, L. X., Orellano, L. A. A., Scheuermann, K., MaChado, C. T., Vasconcelos, A. C., et al. (2020). Sodium butyrate downregulates implant-induced inflammation in mice. Inflammation 43, 1259–1268. doi: 10.1007/s10753-020-01205-0
Deng, F., Zhao, B. C., Yang, X., Lin, Z. B., Sun, Q. S., Wang, Y. F., et al. (2021). The gut microbiota metabolite capsiate promotes Gpx4 expression by activating TRPV1 to inhibit intestinal ischemia reperfusion-induced ferroptosis. Gut Microbes 13, 1–21. doi: 10.1080/19490976.2021.1902719
Fenicia, L., Da Dalt, L., Anniballi, F., Franciosa, G., Zanconato, S., Aureli, P. (2002). A case if infant botulism due to neurotoxigenic Clostridium butyricum type E associated with Clostridium difficile colitis. Eur. J. Clin. Microbiol. Infect. Dis. 21, 736–738. doi: 10.1007/s10096-002-0816-z
Gao, X., Liu, X., Wang, Y., Wang, T., Fang, D., Hu, K. (2023). Effects of Clostridium butyricum on Intestinal Microflora and Metabolism of Eriocheir sinensis. Int. J. Mol. Sci. 24, 13784. doi: 10.3390/ijms241813784
Ge, Y., Wang, X., Guo, Y., Yan, J., Abuduwaili, A., Aximujiang, K., et al. (2021). Gut microbiota influence tumor development and Alter interactions with the human immune system. J. Exp. Clin. Cancer Res. 40, 42. doi: 10.1186/s13046-021-01845-6
Gong, S., Yan, Z., Liu, Z., Niu, M., Fang, H., Li, N., et al. (2019). Intestinal microbiota mediates the susceptibility to polymicrobial sepsis-induced liver injury by granisetron generation in mice. Hepatology 69, 1751–1767. doi: 10.1002/hep.30361
Gotts, J. E., Matthay, M. A. (2016). Sepsis: pathophysiology and clinical management. Bmj 353, i1585. doi: 10.1136/bmj.i1585
Guo, X. Y., Liu, X. J., Hao, J. Y. (2020). Gut microbiota in ulcerative colitis: insights on pathogenesis and treatment. J. Dig Dis. 21, 147–159. doi: 10.1111/1751-2980.12849
Haak, B. W., Wiersinga, W. J. (2017). The role of the gut microbiota in sepsis. Lancet Gastroenterol. Hepatol. 2, 135–143. doi: 10.1016/s2468-1253(16)30119-4
Haussner, F., Chakraborty, S., Halbgebauer, R., Huber-Lang, M. (2019). Challenge to the intestinal mucosa during sepsis. Front. Immunol. 10. doi: 10.3389/fimmu.2019.00891
Hsiao, Y. P., Chen, H. L., Tsai, J. N., Lin, M. Y., Liao, J. W., Wei, M. S., et al. (2021). Administration of Lactobacillus reuteri Combined with Clostridium butyricum Attenuates Cisplatin-Induced Renal Damage by Gut Microbiota Reconstitution, Increasing Butyric Acid Production, and Suppressing Renal Inflammation. Nutrients 13, 2792. doi: 10.3390/nu13082792
Jacobi, J. (2022). The pathophysiology of sepsis - 2021 update: Part 2, organ dysfunction and assessment. Am. J. Health Syst. Pharm. 79, 424–436. doi: 10.1093/ajhp/zxab393
Kanauchi, O., Mitsuyama, K., Araki, Y., Andoh, A. (2003). Modification of intestinal flora in the treatment of inflammatory bowel disease. Curr. Pharm. Des. 9, 333–346. doi: 10.2174/1381612033391883
Kripke, S. A., Fox, A. D., Berman, J. M., Settle, R. G., Rombeau, J. L. (1989). Stimulation of intestinal mucosal growth with intracolonic infusion of short-chain fatty acids. JPEN J. Parenter Enteral Nutr. 13, 109–116. doi: 10.1177/0148607189013002109
Kwiecinski, J. M., Horswill, A. R. (2020). Staphylococcus aureus bloodstream infections: pathogenesis and regulatory mechanisms. Curr. Opin. Microbiol. 53, 51–60. doi: 10.1016/j.mib.2020.02.005
Lee, J. C., Chiu, C. W., Tsai, P. J., Lee, C. C., Huang, I. H., Ko, W. C., et al. (2022). Clostridium butyricum therapy for mild-moderate Clostridioides difficile infection and the impact of diabetes mellitus. Biosci. Microbiota Food Health 41, 37–44. doi: 10.12938/bmfh.2021-049
Li, Z., Chen, M., Zhang, R., Wang, Z., He, H., Wan, Z., et al. (2023). Clostridium butyricum Ameliorates the Effect of Coprophagy Prevention on Hepatic Lipid Synthesis in Rabbits via the Gut-Liver Axis. Int. J. Mol. Sci. 24, 17554. doi: 10.3390/ijms242417554
Lisman, J., Buzsáki, G., Eichenbaum, H., Nadel, L., Ranganath, C., Redish, A. D. (2017). Viewpoints: how the hippocampus contributes to memory, navigation and cognition. Nat. Neurosci. 20, 1434–1447. doi: 10.1038/nn.4661
Liu, D., Huang, S. Y., Sun, J. H., Zhang, H. C., Cai, Q. L., Gao, C., et al. (2022). Sepsis-induced immunosuppression: mechanisms, diagnosis and current treatment options. Mil Med. Res. 9, 56. doi: 10.1186/s40779-022-00422-y
Liu, M., Xie, W., Wan, X., Deng, T. (2020). Clostridium butyricum modulates gut microbiota and reduces colitis associated colon cancer in mice. Int. Immunopharmacol 88, 106862. doi: 10.1016/j.intimp.2020.106862
Ma, L., Shen, Q., Lyu, W., Lv, L., Wang, W., Yu, M., et al. (2022). Clostridium butyricum and its derived extracellular vesicles modulate gut homeostasis and ameliorate acute experimental colitis. Microbiol. Spectr. 10, e0136822. doi: 10.1128/spectrum.01368-22
Macfarlane, G. T., Macfarlane, S. (2011). Fermentation in the human large intestine: its physiologic consequences and the potential contribution of prebiotics. J. Clin. Gastroenterol. 45 Suppl, S120–S127. doi: 10.1097/MCG.0b013e31822fecfe
Muratsu, A., Ikeda, M., Shimizu, K., Kameoka, S., Motooka, D., Nakamura, S., et al. (2022). Dynamic change of fecal microbiota and metabolomics in a polymicrobial murine sepsis model. Acute Med. Surg. 9, e770. doi: 10.1002/ams2.770
Naglova, H., Bucova, M. (2012). HMGB1 and its physiological and pathological roles. Bratisl Lek Listy 113, 163–171. doi: 10.4149/bll_2012_039
Obanda, D. N., Husseneder, C., Raggio, A. M., Page, R., Marx, B., Stout, R. W., et al. (2020). Abundance of the species Clostridium butyricum in the gut microbiota contributes to differences in obesity phenotype in outbred Sprague-Dawley CD rats. Nutrition 78, 110893. doi: 10.1016/j.nut.2020.110893
Oczkowski, S., Alshamsi, F., Belley-Cote, E., Centofanti, J. E., Hylander Møller, M., Nunnaly, M. E., et al. (2022). Surviving Sepsis Campaign Guidelines 2021: highlights for the practicing clinician. Pol. Arch. Intern. Med. 132, (7–8). doi: 10.20452/pamw.16290
Peck, M. W. (2009). Biology and genomic analysis of Clostridium botulinum. Adv. Microb. Physiol. 55, 183–265,320. doi: 10.1016/s0065-2911(09)05503-9
Pouillart, P. R. (1998). Role of butyric acid and its derivatives in the treatment of colorectal cancer and hemoglobinopathies. Life Sci. 63, 1739–1760. doi: 10.1016/s0024-3205(98)00279-3
Quivy, V., Van Lint, C. (2004). Regulation at multiple levels of NF-kappaB-mediated transactivation by protein acetylation. Biochem. Pharmacol. 68, 1221–1229. doi: 10.1016/j.bcp.2004.05.039
Rivière, A., Selak, M., Lantin, D., Leroy, F., De Vuyst, L. (2016). Bifidobacteria and butyrate-producing colon bacteria: importance and strategies for their stimulation in the human gut. Front. Microbiol. 7. doi: 10.3389/fmicb.2016.00979
Salomão, R., Ferreira, B. L., Salomão, M. C., Santos, S. S., Azevedo, L. C. P., Brunialti, M. K. C. (2019). Sepsis: evolving concepts and challenges. Braz. J. Med. Biol. Res. 52, e8595. doi: 10.1590/1414-431x20198595
Shirasugi, M., Nakagawa, M., Nishioka, K., Yamamoto, T., Nakaya, T., Kanamura, N. (2018). Relationship between periodontal disease and butyric acid produced by periodontopathic bacteria. Inflammation Regener. 38, 23. doi: 10.1186/s41232-018-0081-x
Srzić, I., Nesek Adam, V., Tunjić Pejak, D. (2022). Sepsis definition: what’s new in the treatment guidelines. Acta Clin. Croat 61, 67–72. doi: 10.20471/acc.2022.61.s1.11
Stoeva, M. K., Garcia-So, J., Justice, N., Myers, J., Tyagi, S., Nemchek, M., et al. (2021). Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut Microbes 13, 1–28. doi: 10.1080/19490976.2021.1907272
Sun, S., Wang, D., Dong, D., Xu, L., Xie, M., Wang, Y., et al. (2023). Altered intestinal microbiome and metabolome correspond to the clinical outcome of sepsis. Crit. Care 27, 127. doi: 10.1186/s13054-023-04412-x
Tomita, Y., Goto, Y., Sakata, S., Imamura, K., Minemura, A., Oka, K., et al. (2022). Clostridium butyricum therapy restores the decreased efficacy of immune checkpoint blockade in lung cancer patients receiving proton pump inhibitors. Oncoimmunology 11, 2081010. doi: 10.1080/2162402x.2022.2081010
Valdés-Duque, B. E., Giraldo-Giraldo, N. A., Jaillier-Ramírez, A. M., Giraldo-Villa, A., Acevedo-Castaño, I., Yepes-Molina, M. A., et al. (2020). Stool short-chain fatty acids in critically ill patients with sepsis. J. Am. Coll. Nutr. 39, 706–712. doi: 10.1080/07315724.2020.1727379
Vincent, J. L. (2022). Current sepsis therapeutics. EBioMedicine 86, 104318. doi: 10.1016/j.ebiom.2022.104318
Wang, X., He, G., Peng, Y., Zhong, W., Wang, Y., Zhang, B. (2015). Sodium butyrate alleviates adipocyte inflammation by inhibiting NLRP3 pathway. Sci. Rep. 5, 12676. doi: 10.1038/srep12676
Wang, C., Li, Q., Ren, J. (2019). Microbiota-immune interaction in the pathogenesis of gut-derived infection. Front. Immunol. 10. doi: 10.3389/fimmu.2019.01873
Wei, Y., Li, T., Zhao, D., Sun, T., Ma, C., Zhang, L., et al. (2024). Sodium butyrate ameliorates sepsis-associated lung injury by enhancing gut and lung barrier function in combination with modulation of CD4(+)Foxp3(+) regulatory T cells. Eur. J. Pharmacol. 963, 176219. doi: 10.1016/j.ejphar.2023.176219
Wong, S. H., Yu, J. (2019). Gut microbiota in colorectal cancer: mechanisms of action and clinical applications. Nat. Rev. Gastroenterol. Hepatol. 16, 690–704. doi: 10.1038/s41575-019-0209-8
Xiao, K., Sun, Y., Song, J., Li, L., Mao, W., Jiang, C. (2023). Gut microbiota involved in myocardial dysfunction induced by sepsis. Microb. Pathog. 175, 105984. doi: 10.1016/j.micpath.2023.105984
Xie, Y., Zhou, L., Yao, X., Li, Y. (2020). Protective effects of clostridium butyricum in a murine model of dextran sodium sulfate-induced colitis that involve inhibition of the TLR2 signaling pathway and T helper 17 cells. Am. J. Med. Sci. 360, 176–191. doi: 10.1016/j.amjms.2020.05.021
Xu, Y., Kong, X., Zhu, Y., Xu, J., Mao, H., Li, J., et al. (2022). Contribution of gut microbiota toward renal function in sepsis. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.985283
Yan, X. X., Wu, D. (2023). Intestinal microecology-based treatment for inflammatory bowel disease: Progress and prospects. World J. Clin. cases 11, 47–56. doi: 10.12998/wjcc.v11.i1.47
Yang, X. J., Wang, X. H., Yang, M. Y., Ren, H. Y., Chen, H., Zhang, X. Y., et al. (2023). Exploring choices of early nutritional support for patients with sepsis based on changes in intestinal microecology. World J. Gastroenterol. 29, 2034–2049. doi: 10.3748/wjg.v29.i13.2034
Zampieri, F. G., Bagshaw, S. M., Semler, M. W. (2023). Fluid therapy for critically ill adults with sepsis: A review. Jama 329, 1967–1980. doi: 10.1001/jama.2023.7560
Zhang, L., Jin, S., Wang, C., Jiang, R., Wan, J. (2010). Histone deacetylase inhibitors attenuate acute lung injury during cecal ligation and puncture-induced polymicrobial sepsis. World J. Surg. 34, 1676–1683. doi: 10.1007/s00268-010-0493-5
Zhang, Y., Lu, M., Zhang, Y., Yuan, X., Zhou, M., Xu, X., et al. (2023). Clostridium butyricum MIYAIRI 588 alleviates periodontal bone loss in mice with diabetes mellitus. Ann. N Y Acad. Sci. 1529, 84–100. doi: 10.1111/nyas.15058
Zhang, S. F., Tang, Z. S., Tong, L., Tao, X. X., Suo, Q. F., Xu, X. M. (2016). Effects of clostridium butyricum and bifidobacterium on BTLA expression on CD4(+) T cells and lymphocyte differentiation in late preterm infants. Microb. Pathog. 100, 112–118. doi: 10.1016/j.micpath.2016.09.008
Zhang, L. T., Yao, Y. M., Lu, J. Q., Yan, X. J., Yu, Y., Sheng, Z. Y. (2007). Sodium butyrate prevents lethality of severe sepsis in rats. Shock 27, 672–677. doi: 10.1097/SHK.0b013e31802e3f4c
Zhou, D., Pan, Q., Liu, X. L., Yang, R. X., Chen, Y. W., Liu, C., et al. (2017). Clostridium butyricum B1 alleviates high-fat diet-induced steatohepatitis in mice via enterohepatic immunoregulation. J. Gastroenterol. Hepatol. 32, 1640–1648. doi: 10.1111/jgh.13742
Zhou, M., Yuan, W., Yang, B., Pei, W., Ma, J., Feng, Q. (2022). Clostridium butyricum inhibits the progression of colorectal cancer and alleviates intestinal inflammation via the myeloid differentiation factor 88 (MyD88)-nuclear factor-kappa B (NF-κB) signaling pathway. Ann. Transl. Med. 10, 478. doi: 10.21037/atm-22-1670
Keywords: sepsis, Clostridium butyricum, probiotics, treatment, gut microbiota
Citation: Zhao J, Jiang L, He W, Han D, Yang X, Wu L and Zhong H (2024) Clostridium butyricum, a future star in sepsis treatment. Front. Cell. Infect. Microbiol. 14:1484371. doi: 10.3389/fcimb.2024.1484371
Received: 21 August 2024; Accepted: 19 November 2024;
Published: 06 December 2024.
Edited by:
Gaofeng Wu, Xi’an Jiaotong University, ChinaReviewed by:
Xiaowei Zhang, Peking University, ChinaCopyright © 2024 Zhao, Jiang, He, Han, Yang, Wu and Zhong. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Liuli Wu, d2xsMTMwMTMxNjgxOThAMTYzLmNvbQ==; Haiyan Zhong, emhvbmdoYWl5YW5AZXR5eS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.