 Greta Ponsecchi1,2
Greta Ponsecchi1,2 Tommaso Olimpieri1
Tommaso Olimpieri1 Noemi Poerio1
Noemi Poerio1 Alberto Antonelli3,4,5
Alberto Antonelli3,4,5 Marco Coppi3,4,5
Marco Coppi3,4,5 Gustavo Di Lallo1
Gustavo Di Lallo1 Mariangela Gentile6
Mariangela Gentile6 Eugenio Paccagnini6
Eugenio Paccagnini6 Pietro Lupetti6
Pietro Lupetti6 Claudio Lubello7
Claudio Lubello7 Gian Maria Rossolini3,4,5
Gian Maria Rossolini3,4,5 Maurizio Fraziano1
Maurizio Fraziano1 Marco Maria D’Andrea1*
Marco Maria D’Andrea1*- 1Department of Biology, University of Rome “Tor Vergata”, Rome, Italy
- 2PhD Program in Evolutionary Biology and Ecology, Department of Biology, University of Rome “Tor Vergata”, Rome, Italy
- 3Department of Experimental and Clinical Medicine, University of Florence, Florence, Italy
- 4Microbiology and Virology Unit, Florence Careggi University Hospital, Florence, Italy
- 5Nutrition, combating infection and Antibiotic Resistance in Rehabilitation (NARR) Joint Laboratory for Antimicrobial Resistance Research and Control, University of Florence-Istituto di Ricovero e Cura a Carattere Scientifico (IRCCS) Don Gnocchi Foundation, Florence, Italy
- 6Department of Life Sciences, University of Siena, Siena, Italy
- 7Department of Civil and Environmental Engineering (DICEA), University of Florence, Florence, Italy
The global dissemination of multi-drug resistant (MDR) pathogenic bacteria requires the rapid research and development of alternative therapies that can support or replace conventional antibiotics. Among MDR pathogens, carbapenem-resistant Klebsiella pneumoniae (CR-Kp) are of particular concern due to their extensive resistance profiles, global dissemination in hospital environments, and their major role in some life-threatening infections. Phages, or some of their components, are recognized as one of the potential alternatives that might be helpful to treat bacterial infections. In this study, we have isolated and characterized four lytic bacteriophages targeting K. pneumoniae strains of Sequence Type (ST) 307 or ST147, two predominant high-risk clones of CR-Kp. Phages, designated vB_KpS_GP-1, vB_KpP_GP-2, vB_KpP_GP-4, and vB_KpP_GP-5, were isolated from sewage wastewater samples. The vB_KpS_GP-1 phage was a siphovirus unable to establish lysogeny with its host, while the other three were podoviruses. While 85.7% of K. pneumoniae strains of ST307 were selectively lysed by the phages vB_KpS_GP-1 or vB_KpP_GP-5, the other two phages were able to lyse all the tested strains of ST147 (n = 12). Phages were stable over a broad pH and temperature range and were characterized by burst sizes of 10–100 plaque forming units and latency periods of 10–50 minutes. Genome sequencing confirmed the absence of antibiotic resistance genes, virulence factors or toxins and revealed that two phages were likely members of new genera. Given their strictly lytic nature and high selectivity towards two of the major high-risk clones of K. pneumoniae, cocktails of these phages could represent promising candidates for further evaluation in in vivo experimental models of K. pneumoniae infection.
1 Introduction
The spread of multi-drug resistant (MDR) bacterial pathogens in hospital environments represents one of the leading contemporary public health threats that requires global and coordinated actions (Centers for Disease Control and Prevention, 2019; World Health Organization, 2021). Indeed, infections caused by MDR bacteria are frequently associated to high mortality rates, prolonged hospitalizations, and increased healthcare-associated costs, factors that translate, for example, to estimates of a global burden of 1.27 million deaths in 2019 alone (Giske et al., 2007; Murray et al., 2022).
Among MDR Gram negative bacteria, carbapenem-resistant Klebsiella pneumoniae (CR-Kp) are one of the major clinically relevant pathogens (Navon-Venezia et al., 2017; Karampatakis et al., 2023), reaching rates higher than 20% in several countries (Di Pilato et al., 2024). For these reasons, CR-Kp are considered as a critical priority antibiotic-resistant pathogens for which new antibiotics are urgently needed (Tacconelli et al., 2018; World Health Organization, 2024).
The rapid spread of CR-Kp in clinical settings is largely attributed to the global dissemination of a restricted number of successful clones, which have been the cause of several outbreaks at global level (Navon-Venezia et al., 2017; Dong et al., 2022; Karampatakis et al., 2023). These clones, part of the so-called “high-risk clones” (HiRiCs), belong to few different Clonal Groups (CG), including CG11/CG258, CG15, CG37, CG101 and, more recently, ST395, CG307 and CG147 (David et al., 2019; Wyres et al., 2019; Peirano et al., 2020; Shaidullina et al., 2023). HiRiCs pose a significant threat to human health, considering their large dissemination and the extended antibiotic resistance levels, which can also include drugs recently approved for clinical uses. Due to the limited availability of effective antimicrobial compounds and the slow progress in the development of new antibiotics, novel therapeutic strategies to address this growing crisis are therefore urgently required.
Bacteriophages have emerged as a promising approach to tackle antibiotic resistant bacteria. Phage therapy is an old antimicrobial strategy proposed more than a century ago that currently aims at using lytic bacteriophages for the treatment of bacterial infections (Gordillo Altamirano and Barr, 2019; Kortright et al., 2019; Strathdee et al., 2023). In the context of the global dissemination of a restricted number of MDR clones, as it has been observed with CR-Kp, bacteriophages appear of particular interest as a strategy to selectively target and kill bacterial pathogens.
The current study reports on the isolation and characterization of four newly discovered lytic bacteriophages active against K. pneumoniae isolates of Sequence Type (ST) 147 and ST307.
2 Materials and methods
2.1 Bacterial strains
Four K. pneumoniae clinical strains representative of two major HiRiCs, namely ST147 and ST307, were used as indicators for the isolation of lytic bacteriophages. Three out of four isolates were carbapenem resistant due to the production of different carbapenemases, while the remainder was a member of ST307 negative for such enzymes (Table 1). The ability of the isolated phages to target different clones of K. pneumoniae was assessed by using a collection of CR-Kp clinical strains previously characterized in Italian and European surveillance surveys on the spread of carbapenem-resistant K. pneumoniae (Cannatelli et al., 2013; Giani et al., 2013; D’Andrea et al., 2014; David et al., 2019; Di Pilato et al., 2021; Henrici De Angelis et al., 2021; Coppi et al., 2022; Di Pilato et al., 2022). Bacteria in this collection have been selected with the aim to choose clinical isolates belonging to high-prevalent STs, producing epidemiologically relevant carbapenemases and characterized by the maximum genetic distance as resulting by whole-genome comparisons.
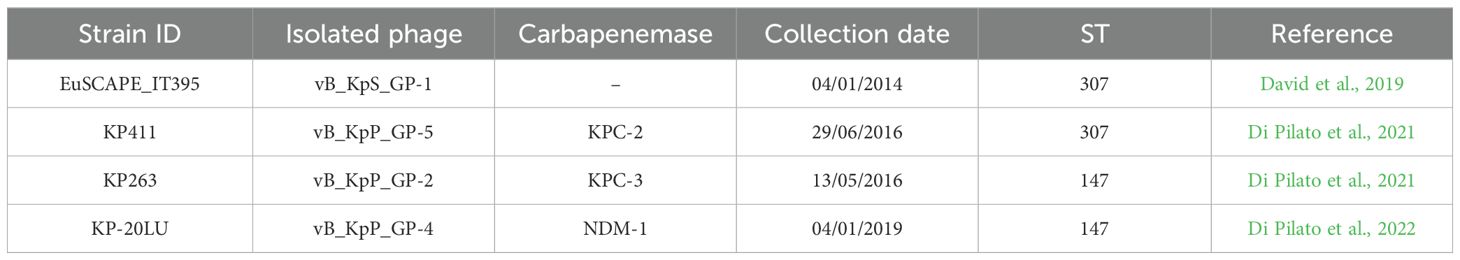
Table 1. Characteristics of the clinical strains used for the isolation of the four bacteriophages.
2.2 Phages isolation
Bacteriophages were isolated from samples of untreated wastewater collected near three hospitals located in central Italy using standard enrichment followed by double-layer agar overlay methods. Briefly, an aliquot of 4.8 ml of each sample was mixed with 5 ml of double-concentrated Lysogeny Broth medium (LB; tryptone 10 g/L, NaCl 10 g/L, yeast extract 5 g/L, pH=7.0) and with 200 µl of a bacterial inoculum of the indicator strain grown overnight (O/N) in LB. The suspension was subsequently incubated O/N at 37°C under shaking and centrifuged at 1,500 x g for 20 min. The obtained supernatant was then filtered through 0.45 µm and 0.22 µm Minisart® filters (Sartorius Minisart, Sarstedt, Italy) to remove bacterial cells and sample debris and used for screening the presence of bacteriophage by spot-test, as previously described (Kutter, 2009). Briefly, 200 μl of a O/N culture of each indicator strain was mixed with 5 ml of soft agar (LB + 0.5% w/v agar), then poured onto LBA plates (LB + 1.5% w/v agar) to form a bacterial lawn. Five microliters of tenfold serial dilutions of each enriched suspension were spotted on the lawn and plates were incubated at 37°C O/N. After incubation, spots having plaques with clear lysis zones were considered positive for the presence of bacteriophages. Well-isolated plaques were then picked and suspended in SM buffer (100 mM NaCl, 8 mM MgSO4•7H2O, 50 mM Tris HCl pH = 7.5). For each phage, three rounds of infection followed by single-plaque isolation have been performed to obtain pure bacteriophage suspensions.
2.3 Large-scale production of bacteriophage suspensions and purification
Propagation of isolated phages obtained by using the EuSCAPE_IT395, KP411, and KP-20LU strains was performed using double-layer agar overlay as reported previously (D’Andrea et al., 2017), while phage selected on the KP263 strain was obtained in liquid. In the latter case, an isolated colony of the indicator strain was inoculated in 5 ml of LB and incubated at 37°C under shaking to an OD600 of 0.4 (≈4x108 CFU/ml). Subsequently, an aliquot of phage suspension was added to obtain a multiplicity of infection (MOI), i.e., a phage/host ratio, of 0.01. This suspension was incubated at 37°C in aerobic conditions for 4 hours and then centrifuged at 1,500 x g for 15 min. The supernatant was filtered at 0.22 µm to remove bacterial debris and titrated by the double-layer agar overlay method as described above. Phage suspensions were subjected to purification by adding 5 ml of polyethylene glycol solution (20% PEG-8000, 3.3 M NaCl) to an iso-volume of phage lysate. The mixture was incubated O/N at 4°C, centrifuged at 12,000 × g for 60 min at 4°C and then the pellet was resuspended in 1 ml of SM buffer and stored at 4°C. For electron microscopy analysis, these phage suspensions were further purified from PEG and concentrated (2.5-5 X) by using Amicon Ultra-4 (10 kDa cut off) centrifugal filters (Merck KGaA, Darmstadt, Germany).
2.4 Transmission electron microscopy
Purified bacteriophages suspensions were subjected to transmission electron microscopy (TEM) analysis to investigate their morphological features. Samples were diluted 1:10 in distilled water, then processed by negative stain. Briefly, 3.5 μl of each phage suspension (≈109-1011 PFU/ml) was laid down on a carbon-coated 300 mesh electron microscopy grid, let to adsorb for 2 minutes and then stained with 1.5% uranyl acetate for 15 seconds. The staining solution was then blotted by filter paper, grid was air-dried, inspected and imaged by TEM. Micrographs were obtained with a FEI Tecnai G2 Spirit TEM microscope (FEI, Eindhoven, The Netherlands) fitted with a TVIPS TemCam F216 CMOS camera (TVIPS, Gilching, German).
2.5 Determination of host-range
The host-range of the isolated bacteriophages was evaluated by spot-test using the four indicator strains used for phages isolation plus other 32 clinical isolates of K. pneumoniae (ST307, n = 12; ST147, n = 10; ST258/512, n = 4; ST101, n = 1; ST395, n = 5). In these experiments, a well-isolated colony of each bacterial strain was inoculated into 500 μl of LB and incubated under shaking at 37°C for 30 min. Aliquots of 10 μl of each inoculum were spotted onto an LBA plate, allowed to absorb and then challenged with 5 μl of phage suspensions (108 PFU/ml) by deposition with partial overlap to the bacterial spot. Results were recorded after 5-6 hours of incubation at 37°C by observation or not of a lysis halo in the overlapped area. Each strain was finally classified as “sensitive (++)”, if the lysis zone was clear, “partially sensitive (+/-)”, if the lysis zone was present but not well defined, and “resistant (-)”, in absence of lysis.
2.6 Efficiency of plating
The ability of the isolated phages to produce productive infections on isolates displaying a “sensitive” or “partially sensitive” phenotype by spot-test, was assessed through the determination of the efficiency of plating (EOP) using the double-layer agar overlay method, as previously described (Khan Mirzaei and Nilsson, 2015). In brief, 5 μl of tenfold serial dilution of phage suspensions (range 108–103 PFU/ml) were spotted on a bacterial lawn of each tested strain. Plates were incubated O/N at 37°C and the number of plaque-forming units (PFU) was finally enumerated. Assays were performed in triplicate for each phage/strain combination and results were computed as the mean of three observations. The EOP was calculated as the ratio between PFU/ml on the tested strains and the PFU/ml on the corresponding indicator strain and used to rank lytic efficiency as “high productive” (EOP ≥ 0.5), “medium productive” (0.1 ≤ EOP < 0.5), “low productive” (0.001 ≤ EOP < 0.1), “inefficient” (EOP < 0.001), and “lysis from without” (in case of observation of lysis but lacking of distinct lysis plaques in any tested concentrations).
2.7 One-step growth curve
The latency period and the burst size of the isolated phages were investigated by using the one-step growth curve technique, as previously described (Kropinski, 2018). Briefly, a single well-isolated colony of each indicator strain was inoculated into 5 ml of LB and incubated under aerobic conditions at 37°C to OD600 = 0.3-0.4. One ml of each culture was centrifuged at 16,873 x g for 5 min and the pellet resuspended in 1 ml of SM buffer. Bacterial suspensions were mixed with phage lysates to achieve a MOI of 0.01, let stand for 5 or 10 min (depending on the phage and according to preliminary tests) at 37°C statically in a water-bath to allow phage absorption, and then centrifuged at 16,873 x g for 2 min at RT to remove the non-absorbed phages. The pellet was then suspended in 1 ml of SM buffer, diluted 1:1000 in 10 ml of LB medium and incubated at 37°C under shaking. Aliquots of 100 µl were sampled every 5 or 10 min up to 50 or 120 minutes. Each aliquot was mixed with 5 ml of soft agar, poured onto an LBA plate and incubated O/N at 37°C. The latency period was defined as the interval between the onset of infection (excluding the pretreatment period) and the beginning of the release of phage progeny. The burst size of each phage was calculated as the ratio between the average number of released phage particles at plateau and the average number of infected bacterial cells during the latency period. Each experiment was performed three times, and results are reported as mean values ± standard deviation.
2.8 Effects of pH and temperature on phage stability
To evaluate the stability to variations of environmental conditions, phages were incubated at different temperatures or pH. Thermal stability was assessed by incubating 1 ml of phage suspensions (range 108-109 PFU/ml) at 37°C, 50°C, 60°C, and 70°C for 10, 20, 40 and 60 minutes. Stability to pH was evaluated by diluting 10 μl of each phage suspension (range 107-1010 PFU/ml) 1:100 in 1 ml of SM buffer previously adjusted to pH from 2.0 to 12.0 with intervals of 1 unit. Mixtures were then incubated statically at 37°C for 60 min. In both tests, at each time point serial dilutions of phages were titrated using the corresponding indicator strain by double-layer agar assay. Both experiments were performed in triplicate and results are reported as mean values ± standard deviation.
2.9 Phage DNA extraction and genome sequencing
The Wizard® DNA Clean-Up System (Promega, Madison, WI, USA) was used for extraction of bacteriophage genomic DNA, following the protocol provided by the manufacturer. Before DNA extraction phage lysates were treated by adding 0.8 μl of DNase I (2 U/μl) and 1 μl of RNase A (100 mg/ml) and incubated at RT for 15 min to remove host nucleic acids. The obtained DNA preparations were quantified by using a NanoDrop Spectrophotometer (Nanodrop Technologies Inc., Wilmington, USA) and visualized by 0.8% agarose gel electrophoresis to check the integrity of the genome. Whole genome sequencing was performed by Illumina NovaSeq 6000 platform with a paired end (2 x 150 bp) approach by using the Illumina DNA Prep kit (Illumina Inc., San Diego, CA, USA).
2.10 Bioinformatics analysis
Raw sequencing reads were quality checked with the software FastQC (version 0.11.9) and assembled de novo by using the Shovill pipeline (version 1.1.0) and SPAdes (version 3.15.5) as assembler. The obtained contigs were analyzed with PhageTerm (version 1.0.11) to derive information on each genome topology, tentatively define each phage DNA termini and DNA packaging models (Garneau et al., 2017). Search for Open Reading Frames (ORFs) and their annotation were performed by using the Rapid Annotation using Subsystem Technologies (RAST) web-service, Prokka (version 1.14.5) and Prodigal (version 2.6.3) (Aziz et al., 2008; Hyatt et al., 2010; Seemann, 2014), followed by manual curation. When different start codons have been proposed, each putative ORFs was manually assessed by searching homologues of the E. coli ribosomal binding site (5’- AGGAGG -3’) upstream of the predicted start codons. Putative functions of each ORFs were further verified by using HHpred, HMMER (version 3.3.2), InterPro (version 98.0) and BLASTp (NCBI database) (Finn et al., 2015; Zimmermann et al., 2018; Paysan-Lafosse et al., 2023). Rho-independent transcriptional terminators were identified by using ARNold, TransTermHP and PePPER web-software (Kingsford et al., 2007; Naville et al., 2011; de Jong et al., 2012), while putative promoters were searched by using PhagePromoter (Sampaio et al., 2019). Only transcriptional terminators detected by at least two tools and promoters displaying a score >0.8 and not fully located on ORFs were considered. Search of polysaccharide depolymerase encoding genes was performed by using DepoScope (Concha-Eloko et al., 2024). The BLASTn tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi) was used to search for the closest homologs of each isolated phage among those deposited in databases of the International Nucleotide Standard Database Collaboration (INSDC). Genomes displaying >70% nucleotide identity (computed as % identity multiplied by % coverage) over their full genome length were retrieved and used for phylogenetic analysis and taxonomic classification (Turner et al., 2021). The VICTOR webserver and VIRIDIC were employed to create phylogenetic trees and classify the viruses, respectively (Meier-Kolthoff and Göker, 2017; Moraru et al., 2020).
3 Results
3.1 Isolation of K. pneumoniae phages
Four distinct bacteriophages, designated as vB_KpS_GP-1, vB_KpP_GP-5, vB_KpP_GP-2, and vB_KpP_GP-4, were isolated from wastewater samples using K. pneumoniae strains EuSCAPE_IT395 (ST307), KP411 (ST307), KP263 (ST147), and KP20LU (ST147), respectively. The four phages, named in accordance with directions proposed by Turner et al., are hereafter referred to as GP-1, GP-5, GP-2, and GP-4 for short (Turner et al., 2021). All the bacteriophages formed small, transparent, and well-evident plaques with varying diameters on their respective host strains. Plaques of all phages exhibited a lysis zone with a diameter of ≈ 1 mm. In addition, plaques formed by GP-1, GP-2, and GP-5 were surrounded by an evident fuzzy halo of varying opacity and diameters (Figure 1).

Figure 1. Morphology of lysis plaques formed by the four bacteriophages on their indicator strains. Plaques of GP-1 on K. pneumoniae EuSCAPE_IT395 (A), GP-5 on K. pneumoniae KP411 (B), GP-2 on K. pneumoniae KP263 (C) and GP-4 on K. pneumoniae KP20-LU (D). The bar indicates 1 cm.
3.2 Phage host range and EOP
The host range of the isolated phages was assessed by testing a collection of 32 MDR K. pneumoniae strains plus the four indicator strains used for phage isolation (Table 2). Results showed that GP-1 was able to selectively lyse, although with different efficiency, 10/13 (77%) strains of K. pneumoniae ST 307 (excluding its indicator strain) and that GP-5 was able to perform a productive infection on two K. pneumoniae ST307 strains both resistant to the infection by GP-1. Interestingly, GP-5 was also able to infect, even if at a low productivity, a capsule-deficient strain of K. pneumoniae ST258 clade II resistant to the φBO1E phage (D’Andrea et al., 2017; Henrici De Angelis et al., 2021). A similar association between ST and phage host-range was observed with GP-4, which efficiently lysed 10/11 (91%) strains of K. pneumoniae ST147 (excluding its indicator strain), while GP-2 was able to productively infect 3 ST147 strains, including the strain resistant to GP-4. In addition, the GP-2 phage was also able to infect with low or inefficient productivity 9/14 (64%) strains of ST307 (Table 2). Notably, except for GP-5, the four phages were not able to infect strains of ST different from ST147 or ST307.
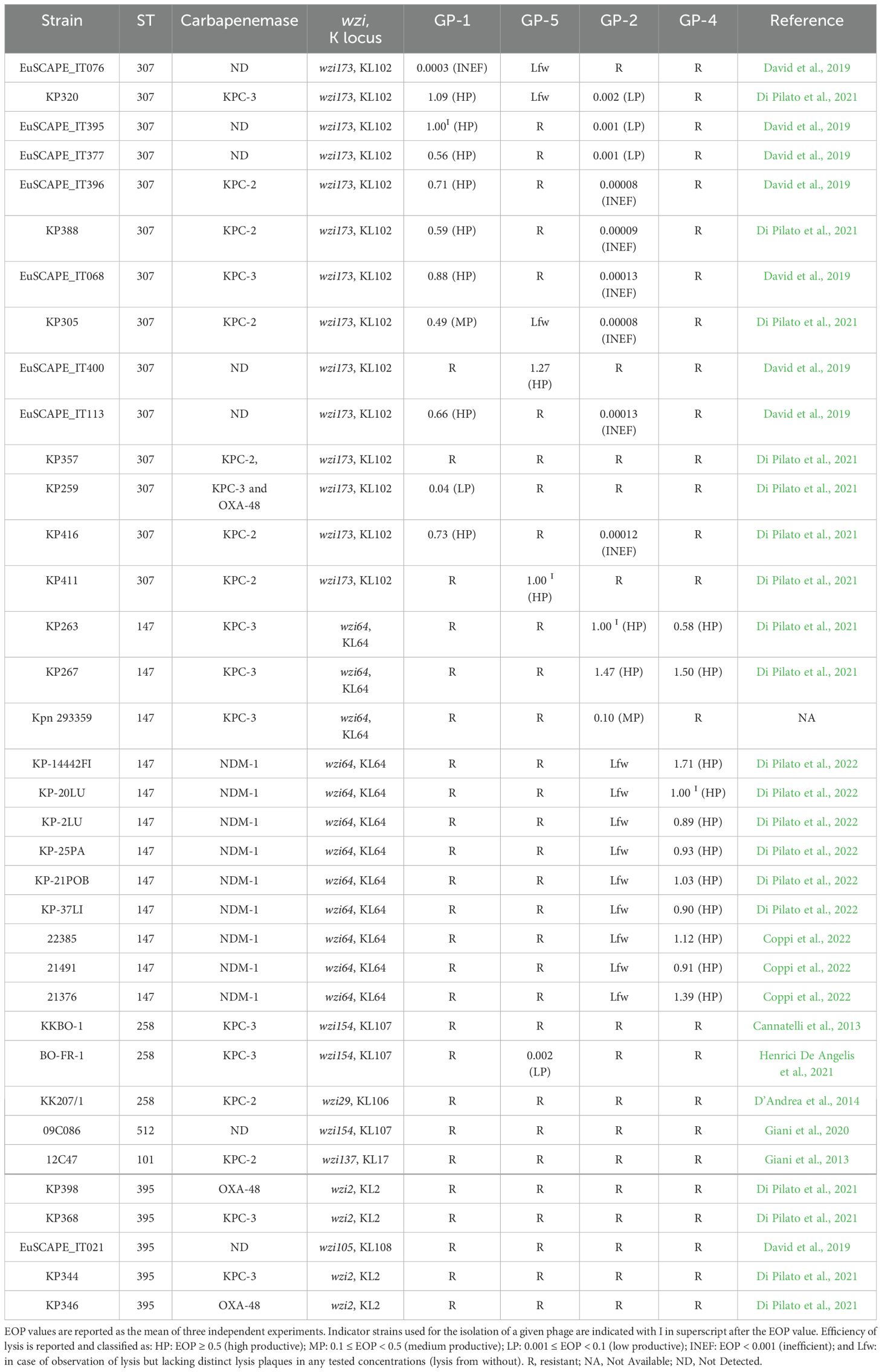
Table 2. Host range of the four phages on 36 MDR K. pneumoniae strains as determined by spot-test and EOP analysis.
3.3 One-step curve experiments
The assessment of the replicative cycle of the four phages was investigated by one-step growth curve. Results from these experiments showed that phage GP-1 exhibited the longest replicative cycle, approximately 120 minutes, which was characterized by a latent period of ≈50 minutes, a release stage of ≈50 minutes and a burst size of ≈150 PFU/Infected cell. On the contrary, the GP-2, GP-4, and GP-5 phages displayed shorter replicative cycles (from 30 to 40 minutes), were similar to each other in terms of latency periods (from 10 to 15 minutes), and showed burst sizes of 9, 53, and 87, respectively (Figure 2).
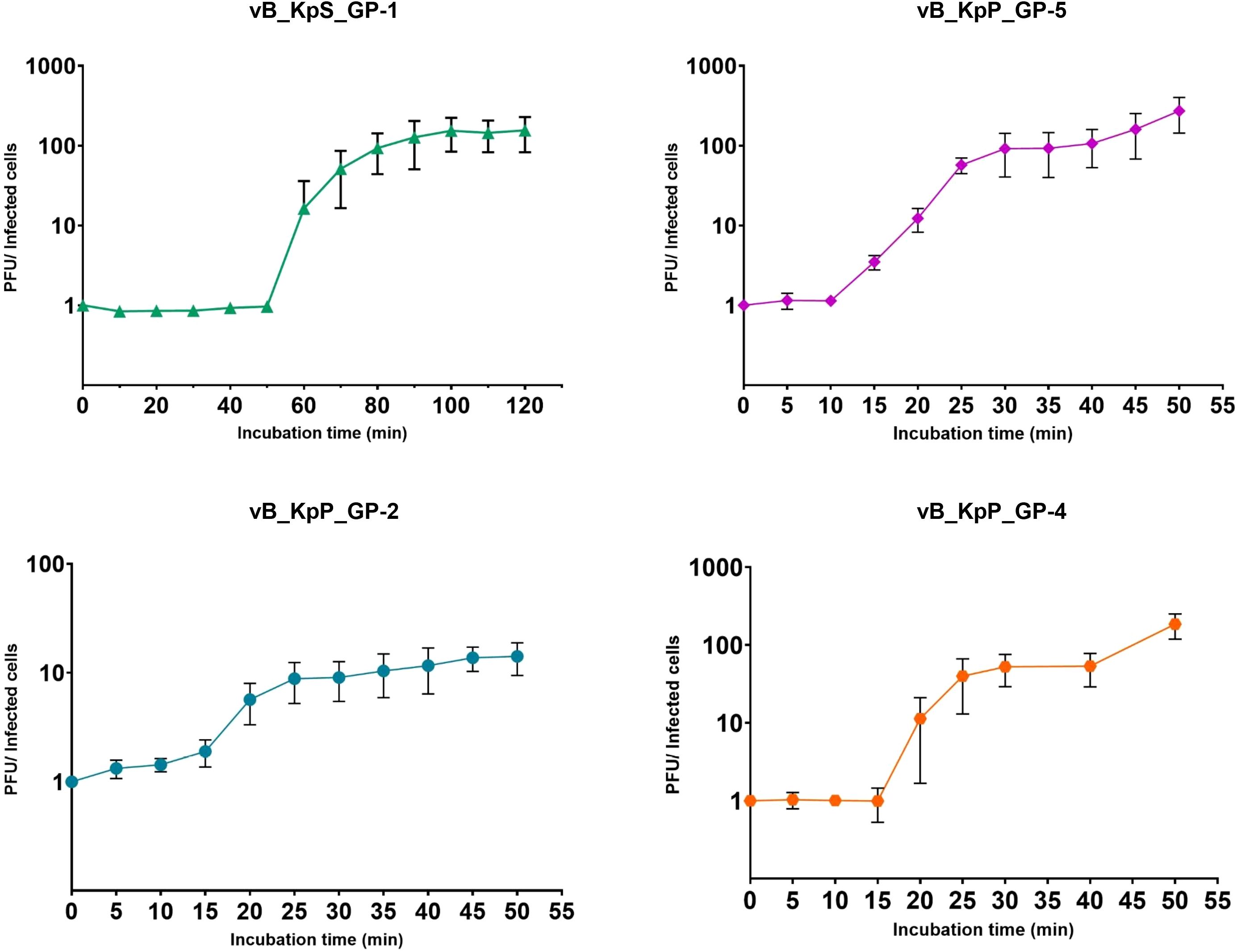
Figure 2. One-step growth curves of the four isolated bacteriophages. The ratios between PFU and the number of infected bacterial cells at different times are shown. Data are the mean from three independent experiments. Vertical black bars indicate one standard deviation.
3.4 Sensitivity to temperature and pH variations
Results from experiments performed to assess the physical and chemical stability of the four phages, showed that GP-1 and GP-5 were stable after 1-hour incubation at temperatures ≤60°C, with a notable decrease of phage titers only after incubation at 70°C. In detail, 30 minutes at 70°C were able to reduce the GP-1 infectivity to zero, while GP-5 demonstrated a higher stability, showing a ≈6-log loss of infectivity after 60 minutes of incubation at the same temperature. On the contrary, a notable decrease of the infective capacity (≥ 3 log) was observed for GP-2 and GP-4 after incubation for 1 hour at 60°C, with a total loss of infectivity following 20- or 10-minutes incubation at 70°C, respectively (Figure 3).
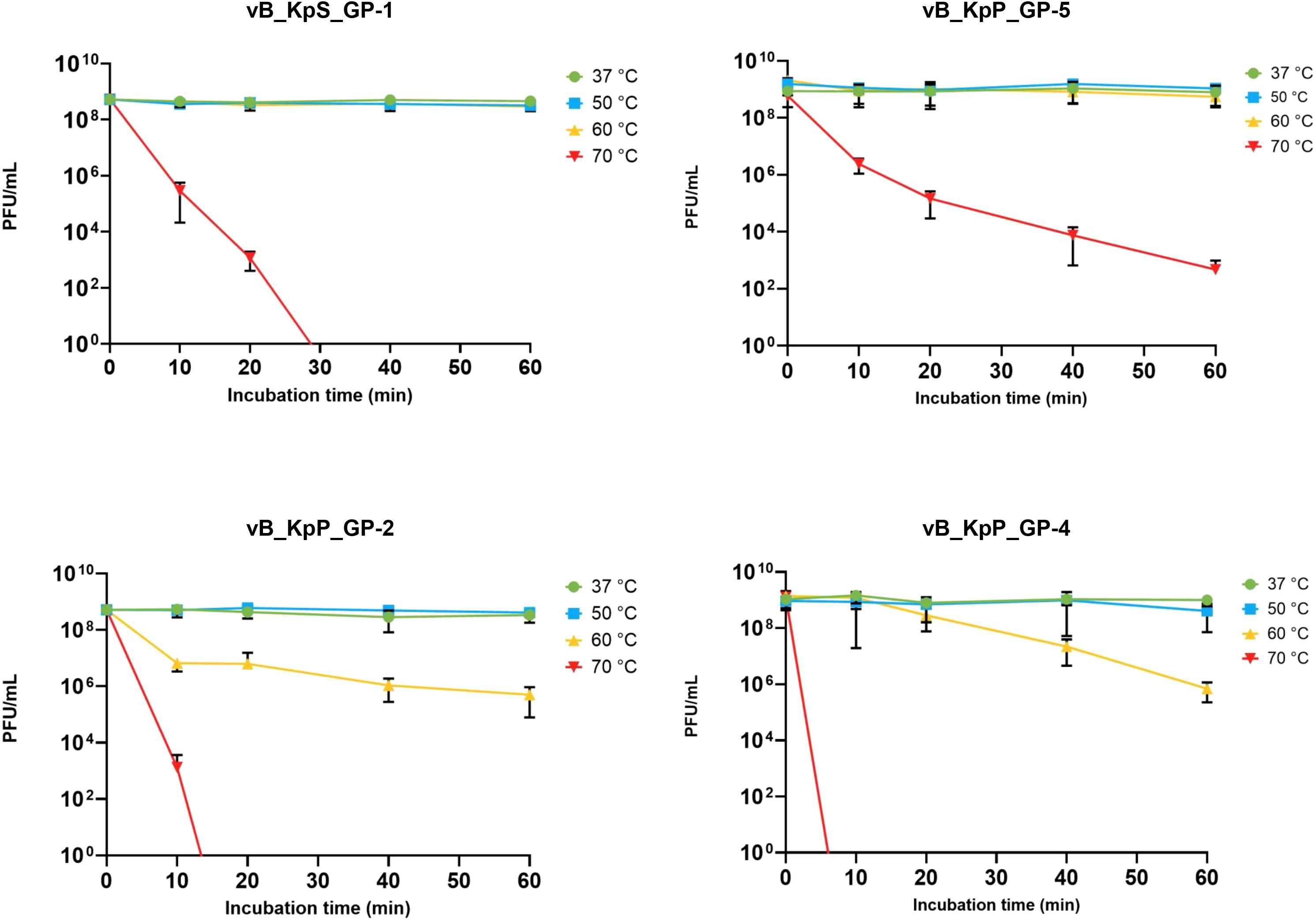
Figure 3. Effect of temperature on infectivity of the four isolated phages. Data represents the mean of three independent experiments. Vertical black bars indicate one standard deviation.
Experiments to assess the effects of pH on phage infectivity demonstrated that all four bacteriophages exhibited comparable stability with limited loss of infective ability within the pH range of 4 to 11. In addition, GP-2 was overall stable also after 1-hour incubation at pH 3 and GP-1 displayed the highest pH stability range, revealing a notable loss of phage infectivity only after 60 minutes of incubation at pH 12 (Figure 4).
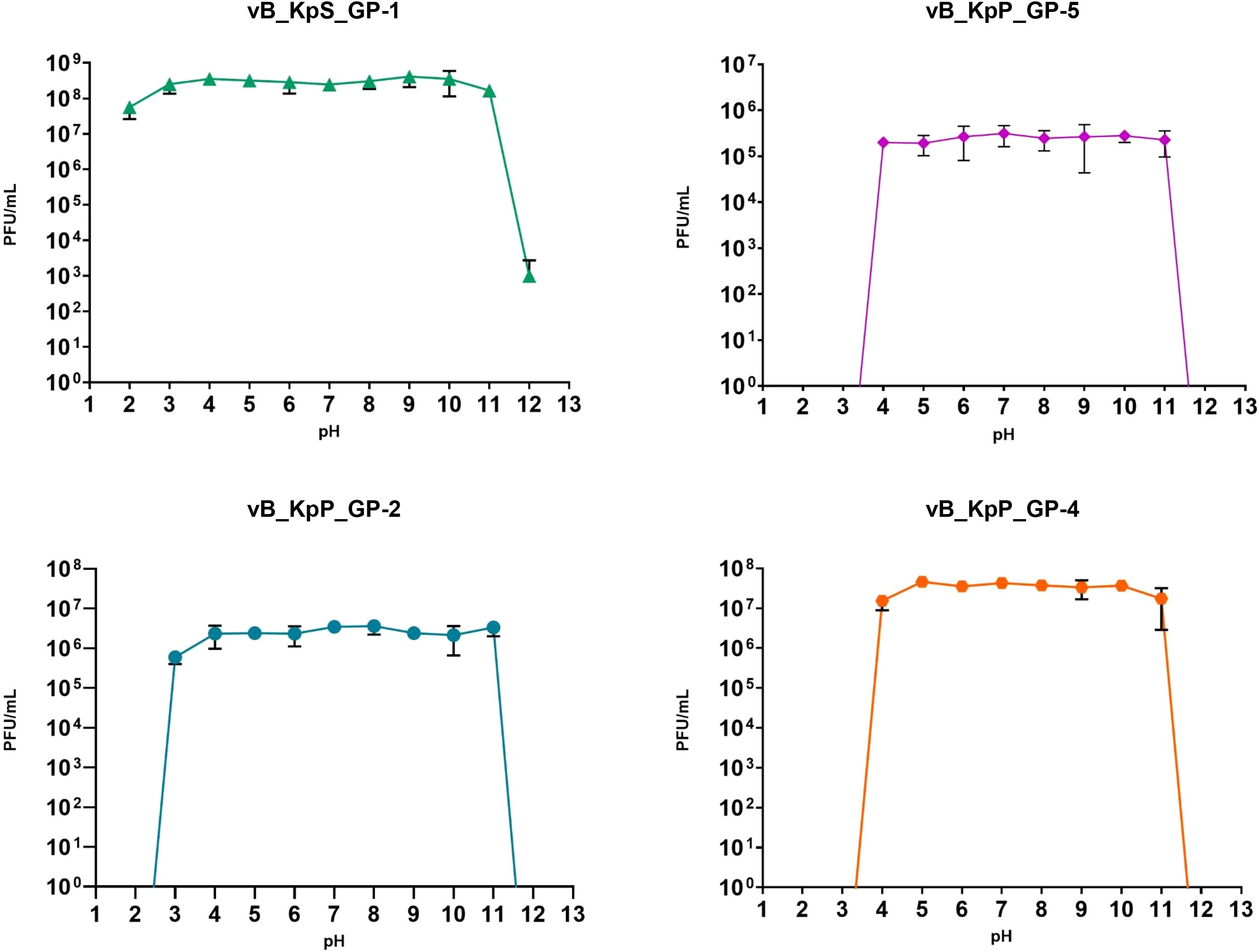
Figure 4. Influence of pH towards phages infectivity. Data are the mean of three independent experiments. Vertical black bars represent one standard deviation.
3.5 Transmission electron microscopy
TEM analysis showed that all phages belonged to the Caudoviricetes class. In detail, GP-1 exhibited typical features of siphovirus, composed of an icosahedral capsid with a diameter of ≈54 nm and a long, non-contractile and flexible tail of ≈130 nm. On the contrary, GP-2, GP-4, and GP-5 were characterized by a short, non-contractile tail connected to an icosahedral capsid with a diameter of ≈54 nm, ≈59 nm, and ≈60 nm, respectively. Overall, the observed morphological features allowed the classification of the latter three phages as podoviruses (Figure 5).

Figure 5. Transmission electron microscopy micrograph of phages vB_KpS_GP-1 (A), vB_KpP_GP-5 (B), vB_KpP_GP-2 (C), and vB_KpP_GP-4 (D). The bar indicates 50 nm.
3.6 Bioinformatics analysis
The whole genomes of the isolated phages were obtained by using a Next Generation Sequencing approach. Results from de novo assembly, followed by PhageTerm analysis, revealed that all the four genomes were composed by double-stranded DNA and showed similar sizes and GC content (Table 3). A total of 48-50 ORFs, all encoded on the same strand, were identified in the three podoviruses (GP-2, GP-4, and GP-5), while 81 ORFs, encoded on either strand, were present in the siphovirus GP-1. Results from the pipeline employed for genome annotation allowed to infer a putative function for 39.1% (GP-1), 84.0% (GP-2), 58.3% (GP-4), and 84.0% (GP-5) ORFs. The genomes of the three podoviruses were characterized by direct terminal repeats located at the 5′- and 3′-ends, with sizes of 179 bp (GP-2), 239 bp (GP-5), and 584 bp (GP-4), while the genome of GP-1 was circularly permuted, suggesting a headful-packaging mechanism in which cutting at different positions of linear DNA concatemers occurs during phage assembly until the capsid is entirely filled with genetic material.
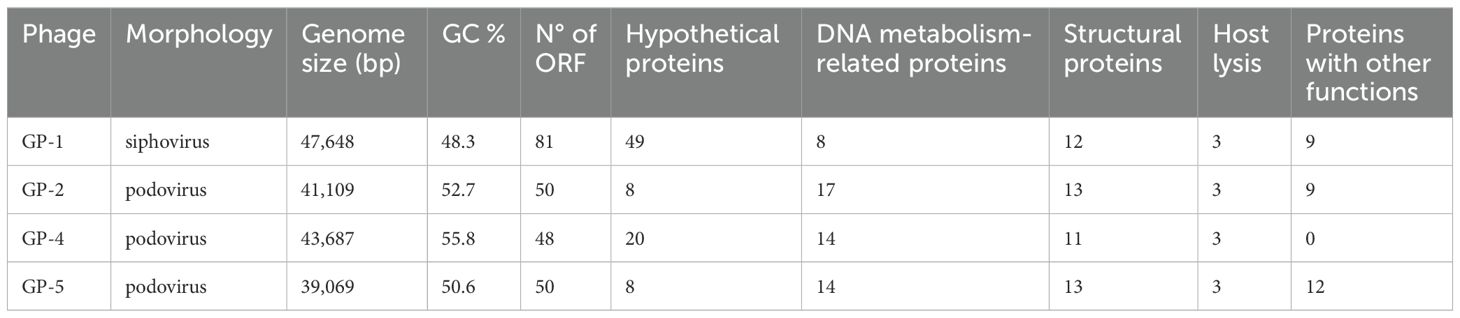
Table 3. Morphology and genomic features of the isolated phages.
Search for regulatory elements revealed the presence of two putative rho-independent terminators in the genomes of GP-2, GP-4, and GP-5, while ten were identified in GP-1. In addition, one (GP-4) to five (GP-5) host promoters have been detected and 14 and 15 phage promoters, showing high similarity to the E. coli phage T7 and T3 RNA polymerases binding sites, were annotated in GP-2 and GP-5, respectively (Supplementary Table 1; Supplementary Figures 1, 2).
No genes associated with virulence factors, antibiotic resistance, or lysogeny were found. The lack of ability to lysogenize its indicator strain was also experimentally confirmed by a PCR approach for the siphovirus GP-1 by using ten different phage-resistant derivatives obtained after infections at high MOI (data not shown). Search for polysaccharide depolymerase-encoding genes suggested the presence of one hit in the GP-1, GP-4, and GP-5 phages (CDS 58, CDS 35, and CDS 39, respectively) and of two hits in the GP-2 genome (CDS 40 and CDS 45).
3.7 Phylogenetic analysis
Results from phylogenetic analysis suggested that GP-1 was a member of a novel genus of the Caudoviricetes class. Indeed, BLAST analysis showed that the GP-1 genome displayed values >70% only with the VLCpiS13a (NC_071152.1) phage (Beamud et al., 2023) and VIRIDIC confirmed these findings. Similar results were obtained for GP-4, for which both BLAST and VIRIDIC analysis suggested that it was part of a new genus together with the vB_Kpl_K59PH2 (OY757063.1), 6937 (OL362270.1), and KYP (ON755176.1) phages. Conversely, results of taxonomic analysis of GP-2 and GP-5 obtained from the two approaches suggested that these phages were members of the Przondovirus and Teetrevirus genus, respectively. Phylogenetic trees obtained by VICTOR analysis are reported in Supplementary Figures 3-6.
4 Discussion
The increasing prevalence of antibiotic-resistant bacterial strains in clinical environments (including both acute care and long-term care rehabilitation facilities) poses significant and worrying public health issues on a global scale. In particular, the rapid dissemination of high-risk CR-Kp clones in these settings strongly limits the available therapeutic options. Therefore, the development of alternative therapies that can replace or complement the use of conventional antibiotics is a major and unmet need. In this context, lytic bacteriophages, or their components, may represent effective tools to address this problem.
Here we report on the isolation and characterization of four newly discovered lytic bacteriophages targeting two major clones of CR-Kp (ST147 and ST307) circulating at a worldwide scale. In detail, to develop a phage cocktail able to lyse the majority of clinical isolates of K. pneumoniae ST307 and ST147, the strains KP411 and KP20LU, resistant to the lytic activity of GP-1 and GP-2, were used as hosts for the isolation of GP-5 and GP-4, respectively. Our results demonstrated that, in combination of two, the described bacteriophages displayed selective lytic capacity against almost all tested clinical isolates belonging to the two clones of interest, with remarkable efficiency. Resistance to phage lysis by some tested strains could be due to the extensive intraclone diversification of the capsular polysaccharide genetic loci commonly observed in K. pneumoniae clinical isolates, as previously reported for members of the CG258 (Wyres et al., 2015).
All the characterized phages exhibit a strictly lytic nature and feature a very narrow host range, aspects that can be particularly useful to select candidates for targeted phage-cocktails. Indeed, with the exception of GP-2 and to a lesser extent GP-5, each phage was able to give productive infections only on strains characterized by a given capsular polysaccharide genetic locus, suggesting that the bacterial target exploited at the initial stages of phage infection is a component of the capsular polysaccharide. In line with this hypothesis, the ability of GP-2 to lyse with low productivity strains of ST307 could be related to the presence in its genome of two capsular depolymerase encoding genes. Moreover, three phages produce plaques surrounded by prominent and fuzzy halos, thus underscoring the likely production of polysaccharide depolymerases. The selectivity to lyse only pathogens responsible of an infection, while leaving the common microbial flora unaffected, is one of the main advantages of using phage therapy or phage-based drugs. Capsular depolymerases could be components of novel biotechnological drugs able to degrade biofilm or enhance the action of host innate immunity against K. pneumoniae in a selective manner, as previously demonstrated (Domingo-Calap et al., 2020; Henrici De Angelis et al., 2021). On the other hand, the lytic ability of GP-5 against strains not recognized nor lysed by the other phages (including the previously characterized capsule-deficient derivative of a strain ST258), suggests that the primary receptor for this phage may not be a component of the bacterial capsule. This hypothesis is supported by a bioinformatics analysis, which revealed a high similarity of the entire genomic sequence (95.87%) of GP-5 with the Klebsiella phage mtp4 (OX335434.1), which is selective for capsular-deficient strains of K. pneumoniae and, similarly to GP-5, exhibits a broader host spectrum (Lourenço et al., 2023). Another point supporting this speculation is given by a genome analysis of the KP411 and EuSCAPE_IT400 strains, in both of which a disruption of the wbaP gene, which encodes a glycosyltransferase involved in the first step of the biosynthesis of the capsular polysaccharide, has been detected. Anyway, additional studies on the mechanisms of interaction of the four phages with their hosts through the characterization of phage-resistant derivatives are currently underway.
The phylogenetic and genetic analysis indicated that three of four phages (GP-2, GP-4 and GP-5) are podoviruses from different genera within the Autographiviridae family of the class Caudoviricetes. Indeed, the genomes of these three phages have some features typical of this family, including: i) the presence of direct terminal repeats at genome ends and genome sizes (39 - 43 Kbp), values similar to those found in the literature for other phages of this family and comparable with the values estimated by the ICTV (Pan et al., 2019; Tan et al., 2019; Li et al., 2021); ii) the presence of genes encoding for RNA polymerase, exploited for an efficient transcription of phage genes; iii) a genetic organization into specific functional modules, with genes encoded on the same DNA strand; and iv) the presence of functional domains typical of polysaccharide depolymerase enzymes in genes of tail fibers. Assignment of GP-2 and GP-5 in the Przondovirus and Teetrevirus genera, respectively, has been made given their high identity with other phages included in the ICTV. On the other hand, the absence of genomes exhibiting enough similarity to GP-4 or GP-1, together with bioinformatics analysis, suggest that these phages are member of two novel genera. In conclusion, results from this work represent solid basis for further in vivo investigations in animal models aimed to elucidate the complex phage/bacterium/host interactions. These preclinical studies will also contribute to evaluate the safety and therapeutic value of the herein reported bacteriophages.
Data availability statement
The nucleotide sequences of vB_KpS_GP-1, vB_KpP_GP-2, vB_KpP_GP-4, and vB_KpP_GP-5 were deposited in the GenBank database under accession numbers PP454752, PP454753, PP454754, and PP454755, respectively.
Author contributions
GP: Conceptualization, Formal analysis, Investigation, Methodology, Visualization, Writing – original draft. TO: Investigation, Methodology, Writing – review & editing. NP: Investigation, Methodology, Writing – review & editing. AA: Data curation, Visualization, Writing – review & editing. MC: Data curation, Visualization, Writing – review & editing. GDL: Data curation, Methodology, Writing – review & editing. MG: Data curation, Methodology, Visualization, Writing – review & editing. EP: Data curation, Methodology, Visualization, Writing – review & editing. PL: Data curation, Visualization, Writing – review & editing. CL: Methodology, Supervision, Writing – review & editing. GMR: Conceptualization, Supervision, Writing – review & editing. MF: Conceptualization, Supervision, Writing – review & editing. MMD: Conceptualization, Formal analysis, Funding acquisition, Investigation, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was partially supported by a research grant financed by the European Union -Next Generation EU -PRIN 2022 (20224T3X8K) by the Italian Ministry of Education, University and Research to MMD, by the ECS17 Tuscany Health Ecosystem (THE) to CL, and by the PAN-HUB 2021 project and by the PNRR project 3138/2021 CN_3: National Center for Gene Therapy and Drugs based on RNA Technology to GMR.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
The handling editor PG declared a past co-authorship with the authors GMR, AA, MC and MMD.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2024.1473668/full#supplementary-material
References
Aziz, R. K., Bartels, D., Best, A. A., DeJongh, M., Disz, T., Edwards, R. A., et al. (2008). The RAST server: Rapid annotations using subsystems technology. BMC Genom. 9, 75. doi: 10.1186/1471-2164-9-75
Beamud, B., García-González, N., Gómez-Ortega, M., González-Candelas, F., Domingo-Calap, P., Sanjuan, R. (2023). Genetic determinants of host tropism in Klebsiella phages. Cell Rep. 42, 2. doi: 10.1016/j.celrep.2023.112048
Cannatelli, A., D’Andrea, M. M., Giani, T., Di Pilato, V., Arena, F., Ambretti, S., et al. (2013). In vivo emergence of colistin resistance in Klebsiella pneumoniae producing KPC-type carbapenemases mediated by insertional inactivation of the PhoQ/PhoP mgrB regulator. Antimicrob. Agents Chemother. 57, 5521–5526. doi: 10.1128/aac.01480-13
Centers for Disease Control and Prevention. (2019). Antibiotic Resistance Threats in the United States 2019 Vol. 2019 (Atlanta, GA: Centers for Disease Control and Prevention). doi: 10.15620/cdc:82532
Concha-Eloko, R., Stock, M., De Baets, B., Briers, Y., Sanjuan, R., Domingo-Calap, P., et al. (2024). DepoScope: accurate phage depolymerase annotation and domain delineation using large language models. PLoS Comput. Biol. 20, e1011831. doi: 10.1371/journal.pcbi.1011831
Coppi, M., Antonelli, A., Niccolai, C., Bartolini, A., Bartolini, L., Grazzini, M., et al. (2022). Nosocomial outbreak by NDM-1-producing Klebsiella pneumoniae highly resistant to cefiderocol, Florence, Italy, August 2021 to June 2022. Euro Surveill. 27, pii=2200795. doi: 10.2807/1560-7917.ES.2022.27.43.2200795
D’Andrea, M. M., Amisano, F., Giani, T., Conte, V., Ciacci, N., Ambretti, S., et al. (2014). Diversity of capsular polysaccharide gene clusters in Kpc-producing Klebsiella pneumoniae clinical isolates of sequence type 258 involved in the Italian epidemic. PloS One 9, e96827. doi: 10.1371/journal.pone.0096827
D’Andrea, M. M., Marmo, P., Henrici De Angelis, L., Palmieri, M., Ciacci, N., Di Lallo, G., et al. (2017). φBO1E, a newly discovered lytic bacteriophage targeting carbapenemase-producing Klebsiella pneumoniae of the pandemic Clonal Group 258 clade II lineage. Sci. Rep. 7, 2614. doi: 10.1038/S41598-017-02788-9
David, S., Reuter, S., Harris, S. R., Glasner, C., Feltwell, T., Argimon, S., et al. (2019). Epidemic of carbapenem-resistant Klebsiella pneumoniae in Europe is driven by nosocomial spread. Nat. Microbiol. 4, 1919–1929. doi: 10.1038/S41564-019-0492-8
de Jong, A., Pietersma, H., Cordes, M., Kuipers, O. P., Kok, J. (2012). PePPER: a webserver for prediction of prokaryote promoter elements and regulons. BMC Genom. 13, 299. doi: 10.1186/1471-2164-13-299
Di Pilato, V., Errico, G., Monaco, M., Giani, T., Del Grosso, M., Antonelli, A., et al. (2021). The changing epidemiology of carbapenemase-producing Klebsiella pneumoniae in Italy: toward polyclonal evolution with emergence of high-risk lineages. J. Antimicrob. Chemother. 76, 355–361. doi: 10.1093/jac/dkaa431
Di Pilato, V., Henrici De Angelis, L., Aiezza, N., Baccani, I., Niccolai, C., Parisio, E. M., et al. (2022). Resistome and virulome accretion in an NDM-1-producing ST147 sublineage of Klebsiella pneumoniae associated with an outbreak in Tuscany, Italy: a genotypic and phenotypic characterisation. Lancet Microbe 3, e224–e234. doi: 10.1016/S2666-5247(21)00268-8
Di Pilato, V., Pollini, S., Miriagou, V., Rossolini, G. M., D’Andrea, M. M. (2024). Carbapenem-resistant Klebsiella pneumoniae: the role of plasmids in emergence, dissemination, and evolution of a major clinical challenge. Expert Rev. Anti Infect. Ther. 22, 25–43. doi: 10.1080/14787210.2024.2305854
Domingo-Calap, P., Beamud, B., Mora-Quilis, L., González-Candelas, F., Sanjuán, R. (2020). Isolation and characterization of two Klebsiella pneumoniae phages encoding divergent depolymerases. Int. J. Mol. Sci. 21, 3160. doi: 10.3390/ijms21093160
Dong, N., Yang, X., Chan, E. W. C., Zhang, R., Chen, S. (2022). Klebsiella species: Taxonomy, hypervirulence and multidrug resistance. EBioMedicine 79, 103998. doi: 10.1016/j.ebiom.2022.103998
Finn, R. D., Clements, J., Arndt, W., Miller, B. L., Wheeler, T. J., Schreiber, F., et al. (2015). HMMER web server: 2015 Update. Nucleic Acids Res. 43, W30–W38. doi: 10.1093/nar/gkv397
Garneau, J. R., Depardieu, F., Fortier, L. C., Bikard, D., Monot, M. (2017). PhageTerm: A tool for fast and accurate determination of phage termini and packaging mechanism using next-generation sequencing data. Sci. Rep. 7, 8292. doi: 10.1038/s41598-017-07910-5
Giani, T., Antonelli, A., Sennati, S., Di Pilato, V., Chiarelli, A., Cannatelli, A., et al. (2020). Results of the Italian infection-Carbapenem Resistance Evaluation Surveillance Trial (iCREST-IT): activity of ceftazidime/avibactam against Enterobacterales isolated from urine. J. Antimicrob. Chemother. 75, 979–983. doi: 10.1093/jac/dkz547
Giani, T., Pini, B., Arena, F., Conte, V., Bracco, S., Migliavacca, R., et al. (2013). Epidemic diffusion of KPC carbapenemase-producing Klebsiella pneumoniae in Italy: Results of the first countrywide survey, 15 may to 30 june 2011. Euro Surveill. 18, pii=20489. doi: 10.2807/ese.18.22.20489-en
Giske, C. G., Monnet, D. L., Cars, O., Carmeli, Y., ReAct-Action on Antibiotic Resistance (2007). Clinical and economic impact of common multidrug-resistant gram-negative bacilli. Antimicrob. Agents Chemother. 52, 813–821. doi: 10.1128/aac.01169-07
Gordillo Altamirano, F. L., Barr, J. J. (2019). Phage therapy in the postantibiotic era. Clin. Microbiol. Rev. 32, e00066–e00018. doi: 10.1128/cmr.00066-18
Henrici De Angelis, L., Poerio, N., Di Pilato, V., De Santis, F., Antonelli, A., Thaller, M. C., et al. (2021). Phage resistance is associated with decreased virulence in KPC-producing Klebsiella pneumoniae of the Clonal Group 258 Clade II Lineage. Microorganisms 9, 762. doi: 10.3390/microorganisms9040762
Hyatt, D., Chen, G. L., Locascio, P. F., Land, M. L., Larimer, F. W., Hauser, L. J. (2010). Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 11, 119. doi: 10.1186/1471-2105-11-119
Karampatakis, T., Tsergouli, K., Behzadi, P. (2023). Carbapenem-resistant Klebsiella pneumoniae: virulence factors, molecular epidemiology and latest updates in treatment options. Antibiotics 12, 234. doi: 10.3390/antibiotics12020234
Khan Mirzaei, M., Nilsson, A. S. (2015). Isolation of phages for phage therapy: a comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PloS One 10, e0118557. doi: 10.1371/journal.pone.0118557
Kingsford, C. L., Ayanbule, K., Salzberg, S. L. (2007). Rapid, accurate, computational discovery of Rho-independent transcription terminators illuminates their relationship to DNA uptake. Genome Biol. 8, R22. doi: 10.1186/gb-2007-8-2-r22
Kortright, K. E., Chan, B. K., Koff, J. L., Turner, P. E. (2019). Phage therapy: a renewed approach to combat antibiotic-resistant bacteria. Cell Host Microbe 25, 219–232. doi: 10.1016/j.chom.2019.01.014
Kropinski, A. M. (2018). Practical advice on the one-step growth curve. Methods Mol. Biol. 1681, 41–47. doi: 10.1007/978-1-4939-7343-9_3
Kutter, E. (2009). Phage host range and efficiency of plating. Methods Mol. Biol. 501, 141–149. doi: 10.1007/978-1-60327-164-6_14
Li, N., Zeng, Y., Bao, R., Zhu, T., Tan, D., Hu, B. (2021). Isolation and characterization of novel phages targeting pathogenic Klebsiella pneumoniae. Front. Cell Infect. Microbiol. 11. doi: 10.3389/fcimb.2021.792305
Lourenço, M., Osbelt, L., Passet, V., Gravey, F., Megrian, D., Strowig, T., et al. (2023). Phages against Noncapsulated Klebsiella pneumoniae: broader host range, slower resistance. Microbiol. Spectr. 11, 4. doi: 10.1128/spectrum.04812-22
Meier-Kolthoff, J. P., Göker, M. (2017). VICTOR: genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 33, 3396–3404. doi: 10.1093/bioinformatics/btx440
Moraru, C., Varsani, A., Kropinski, A. M. (2020). VIRIDIC - A novel tool to calculate the intergenomic similarities of prokaryote-infecting viruses. Viruses 12, 1268. doi: 10.3390/v12111268
Murray, C. J., Ikuta, K. S., Sharara, F., Swetschinski, L., Robles Aguilar, G., Gray, A., et al. (2022). Global burden of bacterial antimicrobial resistance in 2019: a systematic analysis. Lancet 399, 629–655. doi: 10.1016/S0140-6736(21)02724-0
Naville, M., Ghuillot-Gaudeffroy, A., Marchais, A., Gautheret, D. (2011). ARNold: A web tool for the prediction of rho-independent transcription terminators. RNA Biol. 8, 11–13. doi: 10.4161/rna.8.1.13346
Navon-Venezia, S., Kondratyeva, K., Carattoli, A. (2017). Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 41, 252–275. doi: 10.1093/femsre/fux013
Pan, Y. J., Lin, T. L., Chen, Y. Y., Lai, P. H., Tsai, Y. T., Hsu, C. R., et al. (2019). Identification of three podoviruses infecting Klebsiella encoding capsule depolymerases that digest specific capsular types. Microb. Biotechnol. 12, 472–486. doi: 10.1111/1751-7915.13370
Paysan-Lafosse, T., Blum, M., Chuguransky, S., Grego, T., Pinto, B. L., Salazar, G. A., et al. (2023). InterPro in 2022. Nucleic Acids Res. 51, D418–D427. doi: 10.1093/nar/gkac993
Peirano, G., Chen, L., Kreiswirth, B. N., Pitouta, J. D. D. (2020). Emerging antimicrobial-resistant high-risk Klebsiella pneumoniae Clones ST307 and ST147. Antimicrob. Agents Chemother. 64, 10. doi: 10.1128/aac.01148-20
Sampaio, M., Rocha, M., Oliveira, H., Dias, O. (2019). Predicting promoters in phage genomes using PhagePromoter. Bioinformatics 35, 5301–5302. doi: 10.1093/bioinformatics/btz580
Seemann, T. (2014). Prokka: Rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Shaidullina, E. R., Schwabe, M., Rohde, T., Shapovalova, V. V., Dyachkova, M. S., Matsvay, A. D., et al. (2023). Genomic analysis of the international high-risk clonal lineage Klebsiella pneumoniae sequence type 395. Genome Med. 15, 9. doi: 10.1186/s13073-023-01159-6
Strathdee, S. A., Hatfull, G. F., Mutalik, V. K., Schooley, R. T. (2023). Phage therapy: From biological mechanisms to future directions. Cell 186, 17–31. doi: 10.1016/j.cell.2022.11.017
Tacconelli, E., Carrara, E., Savoldi, A., Harbarth, S., Mendelson, M., Monnet, D. L., et al. (2018). Discovery, research, and development of new antibiotics: the WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 18, 318–327. doi: 10.1016/S1473-3099(17)30753-3
Tan, D., Zhang, Y., Cheng, M., Le, S., Gu, J., Bao, J., et al. (2019). Characterization of Klebsiella pneumoniae ST11 isolates and their interactions with lytic phages. Viruses 11, 1080. doi: 10.3390/v11111080
Turner, D., Kropinski, A. M., Adriaenssens, E. M. (2021). A roadmap for genome-based phage taxonomy. Viruses 13, 506. doi: 10.3390/v13030506
World Health Organization. (2021). Antimicrobial resistance Vol. 2021 (Geneva: World Health Organization). Available at: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance.
World Health Organization. (2024). WHO bacterial priority pathogens list 2024: bacterial pathogens of public health importance to guide research, development and strategies to prevent and control antimicrobial resistance Vol. 2024 (Geneva: World Health Organization). Available at: https://iris.who.int/bitstream/handle/10665/376776/9789240093461-eng.pdf?sequence=1.
Wyres, K. L., Gorrie, C., Edwards, D. J., Wertheim, H. F., Hsu, L. Y., Van Kinh, N., et al. (2015). Extensive capsule locus variation and large-scale genomic recombination within the Klebsiella pneumoniae Clonal Group 258. Genome Biol. evolution. 7, 1267–1279. doi: 10.1093/gbe/evv062
Wyres, K. L., Wick, R. R., Judd, L. M., Froumine, R., Tokolyi, A., Gorrie, C. L., et al. (2019). Distinct evolutionary dynamics of horizontal gene transfer in drug resistant and virulent clones of Klebsiella pneumoniae. PLoS Genet. 15, e1008114. doi: 10.1371/journal.pgen.1008114
Keywords: phage, Klebsiella pneumoniae, multi-drug resistance, phage-therapy, Klebsiella pneumoniae ST147, Klebsiella pneumoniae ST307, carbapenem-resistance
Citation: Ponsecchi G, Olimpieri T, Poerio N, Antonelli A, Coppi M, Di Lallo G, Gentile M, Paccagnini E, Lupetti P, Lubello C, Rossolini GM, Fraziano M and D’Andrea MM (2024) Characterization of four novel bacteriophages targeting multi-drug resistant Klebsiella pneumoniae strains of sequence type 147 and 307. Front. Cell. Infect. Microbiol. 14:1473668. doi: 10.3389/fcimb.2024.1473668
Received: 31 July 2024; Accepted: 06 September 2024;
Published: 04 October 2024.
Edited by:
Paolo Gaibani, IRCCS University Hospital of Bologna Sant Orsola Polyclinic, ItalyReviewed by:
Erica Diani, University of Verona, ItalyAntonio Curtoni, University Hospital of the City of Health and Science of Turin, Italy
Virginia Lotti, University of Verona, Italy
Copyright © 2024 Ponsecchi, Olimpieri, Poerio, Antonelli, Coppi, Di Lallo, Gentile, Paccagnini, Lupetti, Lubello, Rossolini, Fraziano and D’Andrea. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marco Maria D’Andrea, bWFyY28uZGFuZHJlYUB1bmlyb21hMi5pdA==