 Xiang Shang
Xiang Shang Huihui Bai2
Huihui Bai2 Linyuan Fan
Linyuan Fan Zhaohui Liu
Zhaohui Liu
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol. , 01 May 2024
Sec. Biofilms
Volume 14 - 2024 | https://doi.org/10.3389/fcimb.2024.1387414
Objective: To determine the optimum biofilm formation ratio of Gardnerella vaginalis (G. vaginalis) in a mixed culture with Escherichia coli (E. coli).
Methods: G. vaginalis ATCC14018, E. coli ATCC25922, as well as five strains of G. vaginalis were selected from the vaginal sources of patients whose biofilm forming capacity was determined by the Crystal Violet method. The biofilm forming capacity of E. coli in anaerobic and non-anaerobic environments were compared using the identical assay. The Crystal Violet method was also used to determine the biofilm forming capacity of a co-culture of G. vaginalis and E. coli in different ratios. After Live/Dead staining, biofilm thickness was measured using confocal laser scanning microscopy, and biofilm morphology was observed by scanning electron microscopy.
Results: The biofilm forming capacity of E. coli under anaerobic environment was similar to that in a 5% CO2 environment. The biofilm forming capacity of G. vaginalis and E. coli was stronger at 106:105 CFU/mL than at other ratios (P<0.05). Their thicknesses were greater at 106:105 CFU/mL than at the other ratios, with the exception of 106:102 CFU/mL (P<0.05), under laser scanning microscopy. Scanning electron microscopy revealed increased biofilm formation at 106:105 CFU/mL and 106:102 CFU/mL, but no discernible E. coli was observed at 106:102 CFU/mL.
Conclusion: G. vaginalis and E. coli showed the greatest biofilm forming capacity at a concentration of 106:105 CFU/mL at 48 hours and could be used to simulate a mixed infection of bacterial vaginosis and aerobic vaginitis in vitro.
Mixed vaginitis is a group of vaginal infections caused by the coexistence of two or more vaginal pathogens, leading to abnormalities in the vaginal microenvironment and causing different signs and symptoms (Paavonen and Brunham, 2018; Xiao et al., 2022). Depending on the vaginal pathogen, they can be further categorized into more than 10 types. The most common of which include bacterial vaginosis (BV), trichomoniasis vaginalis (TV), vulvovaginal candidiasis (VVC), and aerobic vaginitis (AV).
Mixed vaginitis is becoming more common (Maladkar, 2015). The literature reports varying incidence rates across different regions. This variation may be attributed to various factors such as varying levels of pathogen exposure among different races and regions, atypical clinical symptoms, significant cognitive differences, and variations in detection methods and levels (Nyirjesy et al., 2020). High-quality epidemiological data is not yet available, and the incidence varies considerably domestically and internationally, ranging from 4.44% to 56.80% (Rivers et al., 2011; Wang et al., 2016). Depending on the vaginal pathogens, they can be divided into various types, such as BV + AV, BV + VVC, BV + TV, and VVC+AV. According to earlier reports, the most common kind of mixed vaginitis is BV + VVC. Conversely, since the introduction of the concept of AV by Donders et al. in 2002 (Donders et al., 2002) and the increased awareness of AV among clinicians in recent years, the prevalence of BV + AV is currently considered to be the highest among the types of mixed vaginitis (Salinas et al., 2020; Pacha-Herrera et al., 2022), ranging from 37.14% to 55.34% (Fan et al., 2013; Cooperative Group of Infectious Disease, Chinese Society of Obstetrics and Gynecology, Chinese Medical Association, 2021). Both BV and AV can result in serious health complications for women. In addition to the common symptoms of vaginitis, they can also lead to a higher likelihood of pelvic inflammatory disease and HPV infection. When present during pregnancy, the risks of miscarriage, preterm labor, chorioamnionitis, neonatal infections, and other adverse outcomes increase significantly (Feng et al., 2023; Kenfack-Zanguim et al., 2023; Qulu et al., 2023).
The mechanism of mixed vaginitis is unclear, but mixed infections with multiple pathogens often result in the formation of a mixed biofilm that can interact in a variety of ways, either synergistic or antagonistic interactions. This increases the pathogens’ resistance and capacity to elude the host’s immune response (Allison et al., 2016; Castro et al., 2016; Koo et al., 2018; Lohse et al., 2018). Machado et al (MaChado and Cerca, 2015; Salinas et al., 2018) pointed out that the biofilm formation by Gardnerella vaginalis (G. vaginalis) and other anaerobic bacterial plays a key role in the pathogenesis of BV. Therefore, the interaction and influence between multiple pathogens in mixed vaginitis should be clarified during treatment to further adjust the usage and dosage of applicable medications to improve the curative effect. However, studies on mixed infections, especially those involving interspecies interactions between multiple pathogens and the impact of biofilms on diseases, are still at early stages (Schlecht et al., 2015). Therefore, there is an urgent need to study the biofilm state and the interaction between different pathogens in mixed infections and to construct an infection model of mixed vaginitis. This will provide ideas for improving the efficacy of the treatment of mixed vaginitis and reducing the risk of recurrence.
While a number of studies have examined the relationship between G. vaginalis and Candida in BV + VVC in mixed vaginitis (Fox et al., 2014; Allison et al., 2016); relatively few have examined BV + AV (Janulaitiene et al., 2017; Morrill et al., 2020). G. vaginalis is predominant in BV (Salinas et al., 2018), and Escherichia coli (E. coli) is also predominant in patients with AV (Wang et al., 2020; Prasad et al., 2021). Therefore, this study aimed to establish a stable in vitro situation that is most suitable for the growth of G. vaginalis and E. coli, to simulate the mixed infection status of BV and AV. Additionally, it provided a basic in vitro model for the subsequent study of the related mechanism and treatment of BV + AV mixed infection.
The strains of Gardnerella and E. coli in our study included G. vaginalis ATCC14018 (Vaneechoutte et al., 2019), E. coli ATCC25922, and other Gardnerella species isolated from patients with BV. Clinical symptoms and the Nugent score were used to diagnose BV in patients who did not have any other urogenital infections. Antibiotics had not been given to the patients during the preceding week. This study was approved by the Ethics Committee of Beijing Obstetrics and Gynecology Hospital, Capital Medical University (2022-KY-064-01).
G. vaginalis and E. coli were cultured using supplementing with brain-heart infusion (sBHI), which comprised brain-heart infusion broth (AOBOX, Beijing, China), 0.3% starch, and 0.3% glucose (He et al., 2021). The overnight growth of G. vaginalis and E. coli were adjusted to 0.5 McFarland (1.5 × 108 CFU/mL), and diluted to 1 ×106 CFU/mL, and then added to a 96-well plate (Falcon, Corning Inc., NY, USA). Then it was incubated at 37°C in an anaerobic environment (AnaeroPouch-Anaero, C-1, Mitsubishi Gas Chemical CO., INC., Tokyo, Japan). After incubation, the bacterial solution was discarded and the wells washed by 1× phosphate buffer saline (PBS) (P1020, Solarbio, Beijing, China). After staining the samples with 0.2 percent crystal violet (C8470, Solarbio, Beijing, China), they were rinsed with 1× PBS and subsequently decolored with 95% alcohol. The OD value of the eluate was detected using an enzyme marker at 580 nm. The OD cutoff value (ODc) was determined by taking the mean value of the negative controls that contained only sBHI plus three times the standard deviation. The biofilm forming capacity was then calculated by dividing the microtiter wells’ OD value by ODc (Shiroda and Manning, 2020; Dantas-Medeiros et al., 2021). All analyses were repeated three times on different dates.
G. vaginalis is an anaerobic bacterium that requires stringent anaerobic environments. On the other hand, E. coli is a facultative anaerobic bacterium that can be cultured under favorable conditions. It is necessary to conduct preliminary experiments to confirm whether the biofilm forming capacity of E. coli differs in anaerobic and non-anaerobic environments. The biofilm forming capacity of E. coli was determined by the Crystal Violet (CV) method after incubation at 37°C, 5% CO2 and anaerobic environment for 48 hours.
G. vaginalis ATCC14018 and E. coli ATCC25922 were mixed and cultured in sBHI to formulate bacterial volume ratios of 100:106, 101:106, 102:106, 103:106, 104:106, 105:106, 106:106, 106:105, 106:104, 106:103, 106:102, 106:101 and 106:100 CFU/mL, which were added to the 96-well plates and co-cultured for 48 hours at 37°C in an anaerobic environment. Thereafter, using the CV method, the biofilm forming capacity was ascertained.
With some modifications, the Filmtracer Live/Dead biofilm viability kit (L10316, Thermo Fisher Scientific, United States) was used to stain the formed biofilms in accordance with the manufacturer’s instructions (He et al., 2021). After that, Confocal Laser Scanning Microscopy (CLSM) (NIKON ECLIPSE TI, Nikon, Tokyo, Japan) and imaging system (NIKON C2, Nikon, Tokyo, Japan) were used to measure the thickness of biofilm formation. In accordance to method 2.2.2, the samples were incubated anaerobically at 37°C for 48 hours with different bacterial volume ratios. After incubation, the samples were washed three times with sBHI. And then incubated with fluorescent stain, which was prepared by taking 3μL of propidium iodide stain and 3μL of SYTO®9 stain from the Filmtracer Live/Dead biofilm viability kit and adding them to 1mL of sBHI by avoiding light. The samples were then put in a 20×20 mm dish and 1 mL of sBHI was added to cover the surface by 2 mm (He et al., 2021). Claims were made using CLSM, and the excitation wavelengths for green and red light were 488 nm and 561 nm, respectively. In order to obtain a series of images of each layer and combine them, tomography was performed at intervals of 1 μm in the z-axis direction. For each sample, at least four fields of view were chosen in order to detect the thickness of the biofilm (Beaudoin et al., 2017; Ma et al., 2022).
Similar to experimental method 2.3, different proportions of G. vaginalis and E. coli were co-cultured on specialized cell climbing glass (YA0350, Solarbio, Beijing, China) in 24-well plates at 37°C in an anaerobic environment for 48 hours. Biofilm formation was observed by scanning electron microscopy (SEM) (JSM-7900F, JEOL, Japan) at 5 kV after fixation with 1.5% glutaraldehyde (Bulacio et al., 2015; Niu et al., 2017).
Data was analyzed using SPSS version 26.0 (SPSS Statistics for Windows Version 26.0, IBM Corp, Armonk, NY, USA) by the unpaired t-test or non-parametric Wilcoxon signed-rank test. A P value < 0.05 was considered statistical significance.
The Gardnerella strains included G. vaginalis ATCC14018 and five Gardnerella species strains (G. vaginalis 1-G. vaginalis 5) isolated from patients with BV. All Gardnerella strains were identified as G. vaginalis by 16S rRNA sequencing. E. coli ATCC25922 was identified as E. coli.
As shown in Table 1, all strains of G. vaginalis exhibited the strongest biofilm forming capacity at 48 hours, which was stronger than the biofilm forming capacity at 24 hours and 72 hours, with a statistically significant difference (P<0.05). Additionally, G vaginalis ATCC14018 exhibited a stronger biofilm forming capacity at 48 hours compared to other strains, also with a statistically significant difference (P<0.05). Consequently, G. vaginalis ATCC14018 was chosen for further co-culture. The biofilm forming capacity of E. coli at 48 hours was stronger than that at 24 hours, and this difference was statistically significant (P<0.05), whereas the difference was not statistically significant (P>0.05) in contrast to the value after 72 hours. However, 48 hours was chosen as the co-culture observation period because the biofilm forming capacity was stronger at 48 hours than it was at 72 hours.
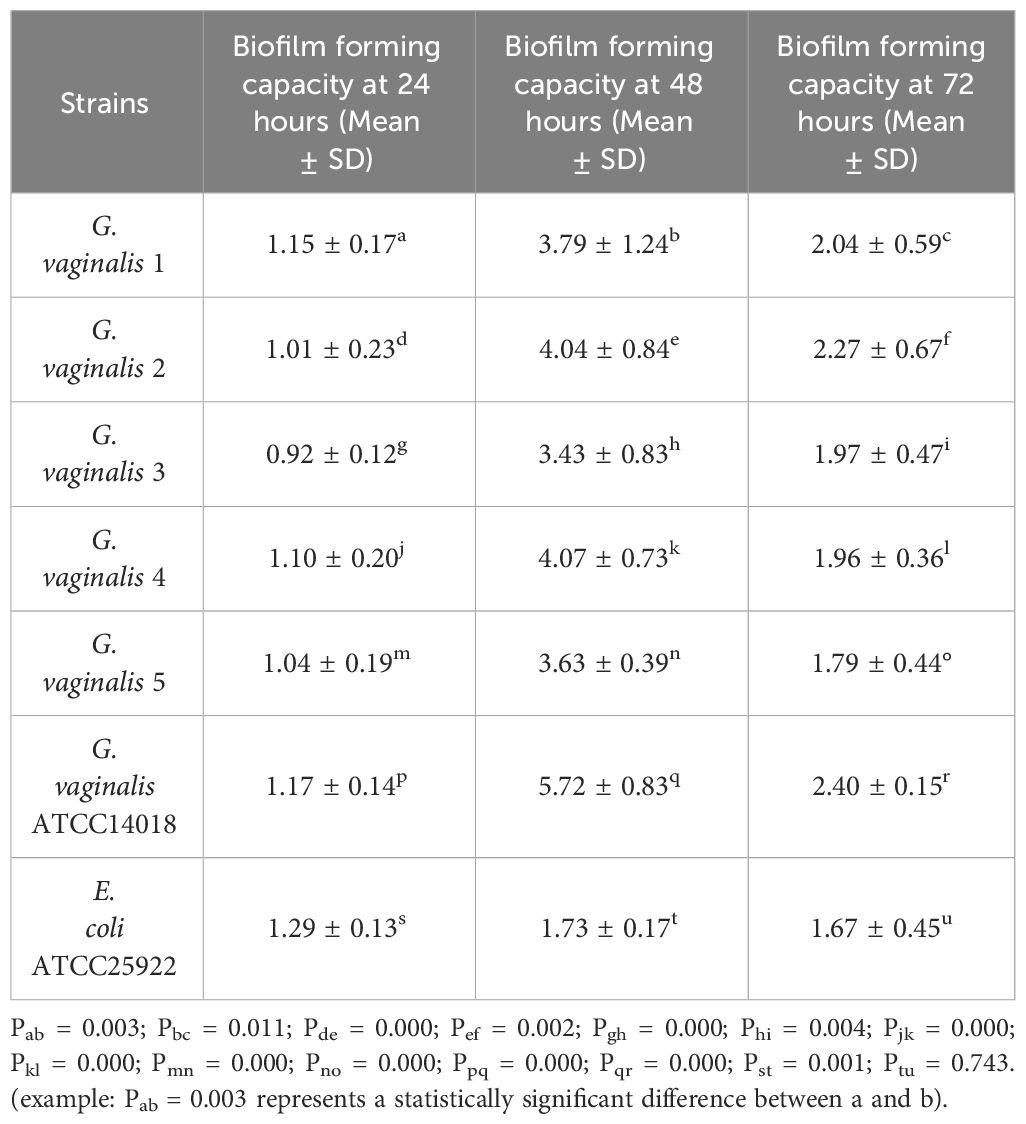
Table 1 Determination of biofilm forming capacity of G. vaginalis and E. coli.
There was no statistically significant difference in the biofilm forming capacity of E. coli between an anaerobic environment and a 5% CO2 environment (P = 0.230, t = -1.277), which is commonly used for culturing E. coli. Therefore, the subsequent co-cultivation assay with G. vaginalis could be performed in an anaerobic environment.
As demonstrated in Figure 1, the biofilm forming capacity of G. vaginalis and E. coli was strong at bacterial volume ratios of 106:105 CFU/mL and 106:102 CFU/mL, with statistically significant differences (P<0.01) when compared to all other ratios and with G. vaginalis and E. coli grown in isolation. The bacterial volume ratio of 106:105 CFU/mL resulted in a stronger biofilm forming capacity compared to 106:102 CFU/mL, and this difference was statistically significant (P<0.05). Therefore, a concentration of 106:105 CFU/mL was chosen for subsequent experiments.
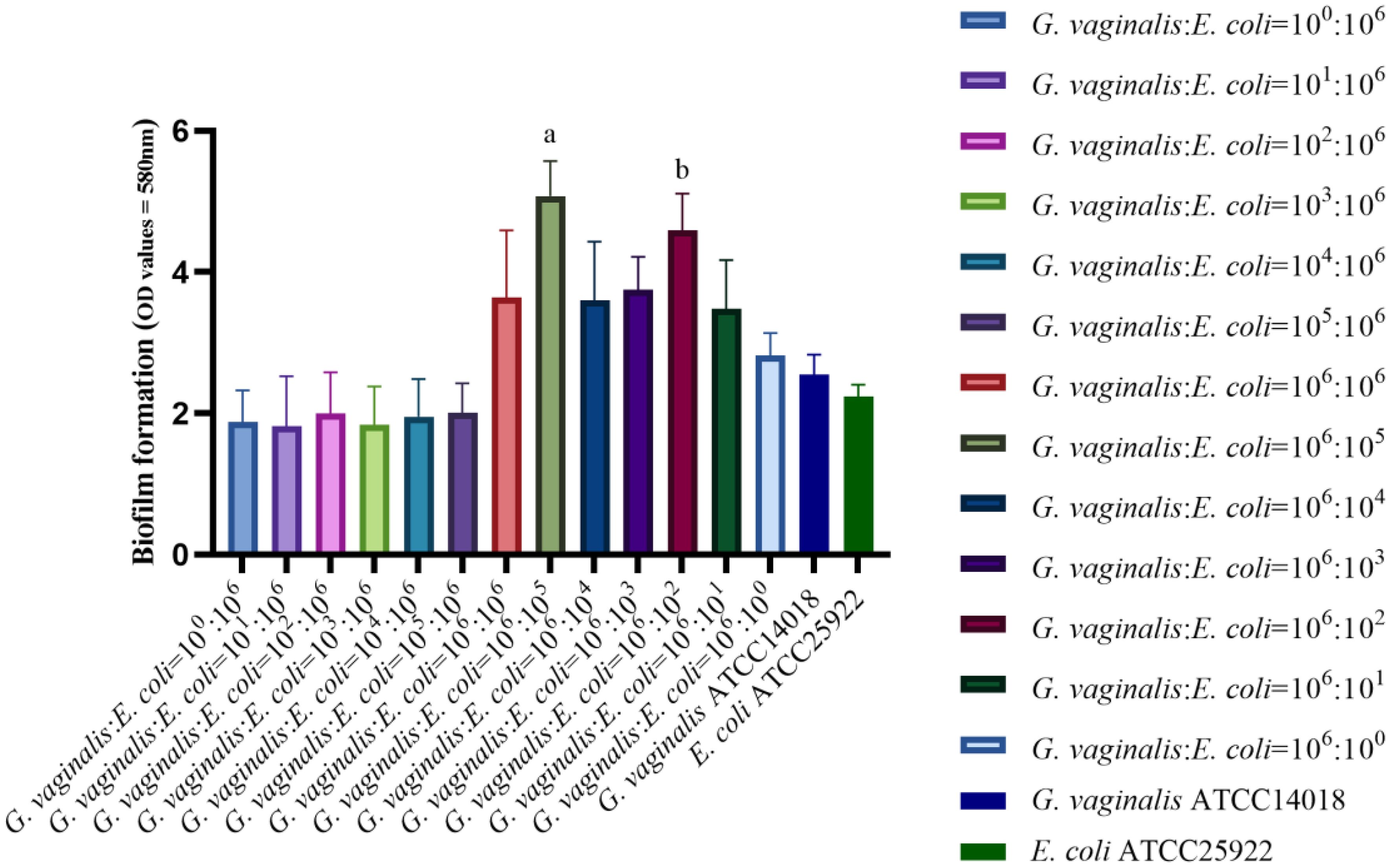
Figure 1 Biofilm forming capacity in co-culture of G. vaginalis and E. coli at different ratios. aThe biofilm forming capacity of G. vaginalis and E. coli at bacterial volume ratios of 106:105 CFU/mL is stronger than other ratios (P<0.05). bThe biofilm forming capacity of G. vaginalis and E. coli at bacterial volume ratios of 106:102 CFU/mL is stronger than other ratios except for 106:105 CFU/mL(P<0.05).
According to the date in Table 2, the thickest biofilm was formed when the volume ratio of G. vaginalis to E. coli was 106:105 CFU/mL. This ratio resulted in a biofilm thickness as measured by CLSM that was significantly greater than all other ratios, except for 106:102 CFU/mL (P<0.05).
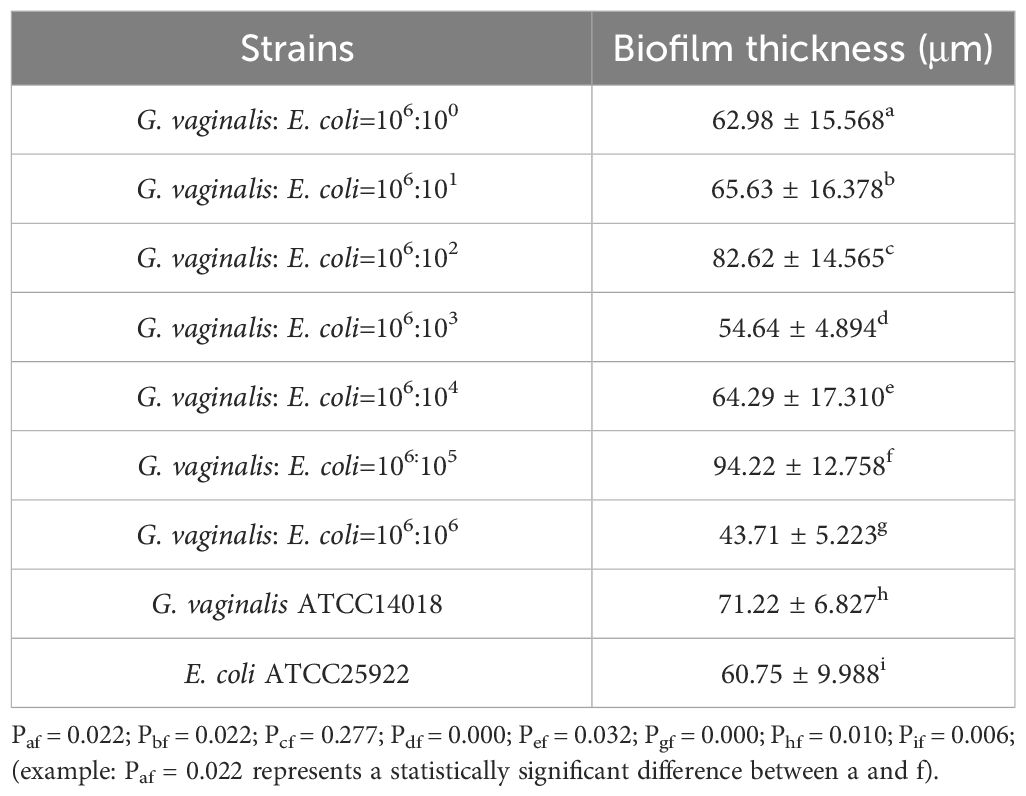
Table 2 Biofilm thickness in co-culture of G. vaginalis and E. coli at different ratios.
Figure 2 shows the performance and 3D images under CLSM after Live/Dead staining and it can be seen that there are more bacteria at the volume ratio of G. vaginalis to E. coli of 106:105 CFU/mL and 106:102 CFU/mL than the other ratio. We could also determine that the biofilm formed at 106:105 CFU/mL and 106:102 CFU/mL was the thickest by looking at the 3D image’s scale lines.
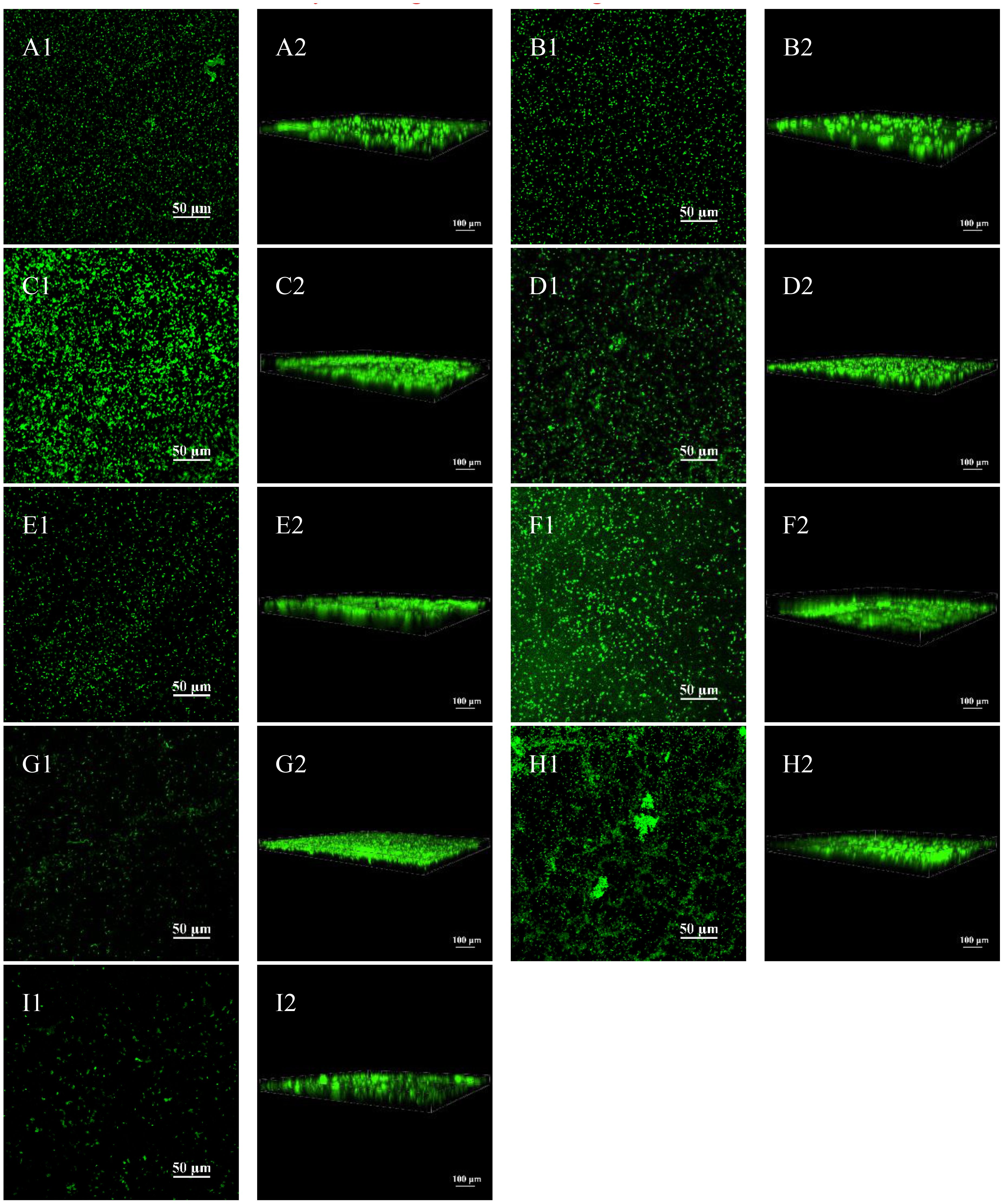
Figure 2 Biofilm formation of G. vaginalis and E. coli via CLSM. (A1) CLSM image at 106:100 CFU/mL; (A2) 3D image at 106:100 CFU/mL; (B1) CLSM image at 106:101 CFU/mL; (B2) 3D image at 106:101 CFU/mL; (C1) CLSM image at 106:102 CFU/mL; (C2) 3D image at 106:102 CFU/mL; (D1) CLSM image at 106:103 CFU/mL; (D2) 3D image at 106:103 CFU/mL; (E1) CLSM image at 106:104 CFU/mL; (E2) 3D image at 106:104 CFU/mL; (F1) CLSM image at 106:105 CFU/mL; (F2) 3D image at 106:105 CFU/mL; (G1) CLSM image at 106:106 CFU/mL; (G2) 3D image at 106:100 CFU/mL; (H1) CLSM image of cultured G. vaginalis only; (H2) 3D image of cultured G. vaginalis only; (I1) CLSM image of cultured E. coli only; (I2) 3D image of cultured G. vaginalis only.
The number of bacteria and biofilm formation were higher when the volume ratio of G. vaginalis to E. coli was 106:105 CFU/mL and 106:102 CFU/mL (Figures 3B, C) than only G. vaginalis was cultured (Figure 3D). As shown in Figure 3A, despite both being rod-shaped, E. coli (yellow arrow) is longer than G. vaginalis (red arrow). Almost no E. coli was seen as demonstrated in Figure 3C.

Figure 3 Biofilm formation of G. vaginalis and E. coli by SEM. (A) SEM image at a bacterial volume ratio of 106:106 CFU/mL; (B) SEM image at a bacterial volume ratio of 106:105 CFU/mL; (C) SEM image at a bacterial volume ratio of 106:102 CFU/mL; (D) SEM image cultured G. vaginalis only. Red arrow: G. vaginalis; yellow arrow: E. coli.
The incidence of mixed vaginitis has been increasing annually in recent years. It also makes diagnosing and treating the infection more difficult due to the combination of several pathogenic bacterial infections, unusual symptoms, and potential interactions between various vaginal pathogens. Imbalances in vaginal flora and biofilm formation have also been linked to an increased risk of infection with human immunodeficiency virus, herpes simplex type 2 and other sexually transmitted diseases like Chlamydia (MaChado et al., 2022). And it also increases the chances of recurrent infections. Our previous study involving 23,181 patients demonstrated that the most common mixed infection pattern was BV + AV in mixed vaginitis (Zong et al., 2023), which is consistent with previous findings (Fan et al., 2013). However, there are a few studies on mixed infection modes and the interaction mechanisms of multiple strains of mixed vaginitis (Benyas and Sobel, 2022; Xiao et al., 2022).
According to recent research, Gardnerella may cause BV through creating biofilms, promoting the attachment of other BV-associated pathogens, promoting the formation of biofilms in a wide range of microorganisms (Hardy et al., 2017; Jung et al., 2017), and generating pro-inflammatory cytokines and other virulence factors (Jang et al., 2017; Cerca, 2019; Shipitsyna et al., 2019). The interaction between a variety of microorganisms, including Gardnerella, is the basis of the pathogenesis of BV. The formation of biofilm is a key pathogenesis and drug resistance mechanism (Africa et al., 2014; Muzny et al., 2019; Castro et al., 2020). Anti-anaerobic medication penetration can be slowed down and bacterial tolerance to the environment can be increased by the physical barrier of a biofilm (Onderdonk et al., 2016). Meanwhile, the expression of the bacterial virulence factor cytolysin reduces the immune response and enhances chronic colonization in women (Castro et al., 2017). It can also reverse the vaginal microenvironment that is conducive to biofilm formation. Antimicrobial resistance has thus grown over the previous few decades and is still growing. There are some treatment options that are currently in use that are becoming less effective (Muñoz-Barreno et al., 2021). The interplay of various pathogen species, the role of biofilms, and whether or not it makes treatment more challenging are urgent problems that need to be solved for mixed vaginitis (Lohse et al., 2018).
AV is a vaginal inflammation that results in congestion and edema of the vaginal mucosa as well as the production of purulent secretions. It is caused by an increase in E. coli, Streptococcus, and other opportunistic pathogens with a decrease in or absence of Lactobacillus. The three main pathogenic bacteria are Staphylococcus aureus, Streptococcus, and E. coli (Tansarli et al., 2013; Rumyantseva et al., 2016; Donders et al., 2017). The development of biofilms contributes significantly to E. coli’s resistance to antibiotic invasion (Sharma et al., 2016). Furthermore, it has also been demonstrated that in urinary tract infection, the presence of Gardnerella induces the proliferation of E. coli, which plays an important role in urinary tract infection (Gilbert et al., 2017). Therefore, in this study, G. vaginalis as well as E. coli were co-cultured to observe biofilm formation and the interactions between various pathogens in order to create a model for further drug trials.
Biofilm formation is a continuous process of adhesion, coaggregation, maturation and dispersion (Jung et al., 2017). In this study, we measured the biofilm forming capacity of a reference strain of G. vaginalis, five G. vaginalis strains from BV patients, and an E. coli reference strain at various points in time. G. vaginalis and E. coli showed the greatest biofilm forming capacity at 48 hours. This indicated that the biofilm was maturing at 48 hours. These results are comparable to those of Castro et al (Castro et al., 2021). Qin et al. also discovered a similar outcome, namely that eight clinical strains of Gardnerella do not spontaneously dissolve during continuous cultivation and instead reach a steady biofilm formation in vitro at 48 hours (Qin et al., 2023). Using the CV method, we also observed that the biofilm forming capacity of G. vaginalis was greater after 48 hours than it after 24 hours. Meanwhile, the biofilm forming capacity of the reference strain of G. vaginalis was greater than that of the clinical strain as measured in this study. Because the most severe scenario of mixed vaginitis infection was simulated in this study using the combination with the strongest biofilm forming capacity in the co-culture, reference strains of G. vaginalis and E. coli were chosen and their co-cultivation was observed for 48 hours to see how their biofilms formed.
G. vaginalis is an anaerobic bacterium that is cultured under harsh conditions (Li et al., 2020), while E. coli is a facultative anaerobic bacterium, who is typically cultured in 5% CO2 environment (Blancarte-Lagunas et al., 2020). There are not any conclusive studies on whether its biofilm forming capacity is different in an anaerobic environment and a 5% CO2 environment. For the first time, we compared the biofilm forming capacity of E. coli in the two environments in this study, and found no statistically significant differences between the two environments. As a result, G. vaginalis and E. coli could be co-cultured in an anaerobic environment.
The most popular and broadly applicable technique for calculating the biofilm forming capacity is the CV method (Magana et al., 2018). However, some studies suggest that the CV method has a risk of unstable measurement, that the biofilm is easily removed by the rinsing process. Mixed infections with multiple pathogens make biofilm formation more complex (Røder et al., 2016; Magana et al., 2018). The ability to form biofilms cannot be determined solely by the CV method. Castro et al (Castro et al., 2022) co-cultured three BV-related anaerobic bacteria, G. vaginalis, Atopobacterium, and Prevotella, and compared biofilm formation using the CV method and fluorescence staining. Every method and culture time produced different results. Therefore, co-cultured biofilms should not be subjected to the CV method alone as an accurate quantitative method. In addition to the CV method, biofilm formation in G vaginalis and E coli co-culture biofilm model was evaluated in this study in a number of ways using biofilm thickness measurement under CLSM following Live/Dead staining, pathogenic bacteria observation, and biofilm morphology analysis under SEM. The benefits of CLSM after Live/Dead staining include clear imaging, the capacity to asses bacterial viability (Freitas et al., 2014; Beaudoin et al., 2017), and the capacity to measure biofilm thickness in the Z-axis direction (He et al., 2021), all of which help to further minimize errors brought on by the use of the CV method. In our study, the volume ratio of 106:105 CFU/mL for G. vaginalis and E. coli produced the thickest biofilm and differed statistically from the other concentrations, with the exception of 106:102 CFU/mL. Biofilm formation was greater under SEM when the ratio of G. vaginalis to E. coli was 106:105 CFU/mL and 106:102 CFU/mL. This was the same as the biofilm forming capacity measured by the CV method and biofilm thickness measured by CLSM. This study verified that, at a ratio of 106:105 CFU/mL, the biofilm forming capacity of G. vaginalis and E. coli was superior to that of other ratios, G. vaginalis by itself, and E. coli alone, employing a variety of techniques like the CV method, measurement of biofilm thickness by CLSM after Live/Dead staining, and SEM observation. Thus, future research on mixed BV and AV infections should take this ratio as a model.
However, as the amount of E. coli decreased, no obvious E. coli was observed under SEM. Actually, E coli was essentially absent when the ratio of G vaginalis to E coli was 106:103 CFU/mL by SEM. On the other hand, the biofilm forming capacity peaked at a bacterial biomass ratio of 106:102 CFU/mL and 106:105 CFU/mL. Research has shown that 92% of Atopobium lost viability when incubated alone for 48 hours without G vaginalis, but that viability increased when Atopobium was co-cultured with G vaginalis or after being treated with sterile supernatants of G vaginalis (Castro et al., 2020). According to an in vitro study, Staphylococcus aureus could infiltrate deep host tissues and contribute to the pathogenic process through the hyphae that carry adherent Candida (Schlecht et al., 2015). Meanwhile, research has demonstrated that the biofilm that Candida forms could shield mixed anaerobic bacteria, enabling them to grow in an oxygen environment (Fox et al., 2014). And mutual promotion of proliferation between E. coli and Streptococcus had been demonstrated in a cross-sectional study (Cools et al., 2016). Furthermore, a strong inhibitory effect on the biofilm of G. vaginalis has been shown for the supernatant of a co-culture of Enterococcus faecalis and Lactobacillus (Zhang et al., 2023). More research is needed to determine whether E. coli acts similarly, whether its products can encourage the formation of Gardnerella biofilms, and the precise mechanism of action.
In conclusion, the ideal ratio of biofilm formation in mixed infections of G. vaginalis and E. coli was identified. This information can be applied to future research on mixed infections of BV and AV and their drug susceptibility. However, this study was limited to in vitro experiments with only two species, G. vaginalis and E. coli. Moreover, as BV and AV are linked to numerous other related bacteria, more research is required to create a mixed infection model of BV and AV in the future. Additionally, more research in the intricate vaginal microenvironment is required to figure out the best way to give medications and increase mixed vaginitis’s effectiveness.
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Approval was granted by the Ethics Committee of Beijing Obstetrics and Gynecology Hospital, Capital Medical University (No.2022-KY-064-01). The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
XS: Conceptualization, Data curation, Investigation, Methodology, Validation, Writing – original draft, Writing – review & editing. HB: Data curation, Methodology, Validation, Writing – original draft. LF: Project administration, Supervision, Validation, Writing – review & editing. XZ: Data curation, Investigation, Writing – original draft. XWZ: Data curation, Investigation, Writing – original draft. ZL: Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by Beijing Obstetrics and Gynecology Hospital, Capital Medical University (XKDTR201803).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Africa, C. W. J., Nel, J., Stemmet, M. (2014). Anaerobes and bacterial vaginosis in pregnancy: virulence factors contributing to vaginal colonization. Int. J. Environ. Res. Public Health 11, 6979–7000. doi: 10.3390/ijerph110706979
Allison, D. L., Willems, H. M. E., Jayatilake, J. a. M.S., Bruno, V. M., Peters, B. M., Shirtliff, M. E. (2016). Candida-bacteria interactions: their impact on human disease. Microbiol. Spectr. 4, 1–6. doi: 10.1128/microbiolspec.VMBF-0030-2016
Beaudoin, T., Yau, Y. C. W., Stapleton, P. J., Gong, Y., Wang, P. W., Guttman, D. S., et al. (2017). Staphylococcus aureus interaction with Pseudomonas aeruginosa biofilm enhances tobramycin resistance. NPJ Biofilms. Microbiomes. 3, 1–9. doi: 10.1038/s41522-017-0035-0
Benyas, D., Sobel, J. D. (2022). Mixed vaginitis due to bacterial vaginosis and candidiasis. J. Low. Genit. Tract. Dis. 26, 68–70. doi: 10.1097/LGT.0000000000000641
Blancarte-Lagunas, M. I., Castro-Escarpulli, G., Navarro-Ocaña, A., Ibáñez-Cervantes, G., Marquez-Valdelamar, L. M., Hernández-Carrillo, J. M., et al. (2020). Commensal and virulent Escherichia coli strains of vaginal origin are reservoirs of resistance cassettes in class 1 integrons. J. Infect. Dev. Ctries. 14, 48–58. doi: 10.3855/jidc.11940
Bulacio, M., de, L. Á., Galván, L. R., Gaudioso, C., Cangemi, R., Erimbaue, M. I. (2015). Enterococcus faecalis biofilm. Formation and development in vitro observed by scanning electron microscopy. Acta Odontol. Latinoam. 28, 210–214.
Castro, J., França, A., Bradwell, K. R., Serrano, M. G., Jefferson, K. K., Cerca, N. (2017). Comparative transcriptomic analysis of Gardnerella vaginalis biofilms vs. planktonic cultures using RNA-seq. NPJ Biofilms. Microbiomes. 3, 3. doi: 10.1038/s41522-017-0012-7
Castro, J., Lima, Â., Sousa, L. G. V., Rosca, A. S., Muzny, C. A., Cerca, N. (2022). Crystal violet staining alone is not adequate to assess synergism or antagonism in multi-species biofilms of bacteria associated with bacterial vaginosis. Front. Cell Infect. Microbiol. 11. doi: 10.3389/fcimb.2021.795797
Castro, J., MaChado, D., Cerca, N. (2016). Escherichia coli and Enterococcus faecalis are able to incorporate and enhance a pre-formed Gardnerella vaginalis biofilm. Pathog. Dis. 74, ftw007. doi: 10.1093/femspd/ftw007
Castro, J., Rosca, A. S., Cools, P., Vaneechoutte, M., Cerca, N. (2020). Gardnerella vaginalis Enhances Atopobium vaginae Viability in an in vitro Model. Front. Cell Infect. Microbiol. 10. doi: 10.3389/fcimb.2020.00083
Castro, J., Rosca, A. S., Muzny, C. A., Cerca, N. (2021). Atopobium vaginae and Prevotella bivia Are Able to Incorporate and Influence Gene Expression in a Pre-Formed Gardnerella vaginalis Biofilm. Pathogens 10, 247. doi: 10.3390/pathogens10020247
Cerca, N. (2019). Could targeting neighboring bacterial populations help combat bacterial vaginosis? Future Microbiol. 14, 365–368. doi: 10.2217/fmb-2019-0045
Cools, P., Jespers, V., Hardy, L., Crucitti, T., Delany-Moretlwe, S., Mwaura, M., et al. (2016). A multi-country cross-sectional study of vaginal carriage of group B streptococci (GBS) and escherichia coli in resource-poor settings: prevalences and risk factors. PloS One 11, e0148052. doi: 10.1371/journal.pone.0148052
Cooperative Group of Infectious Disease, Chinese Society of Obstetrics and Gynecology, Chinese Medical Association (2021). Consensus on the diagnosis and treatment of mixed vaginitis, (2021 edition). Chin. J. Obstet. Gynecol. 56, 15–18. doi: 10.3760/cma.j.cn112141-20200603-00472
Dantas-Medeiros, R., Zanatta, A. C., de Souza, L. B. F. C., Fernandes, J. M., Amorim-Carmo, B., Torres-Rêgo, M., et al. (2021). Antifungal and Antibiofilm Activities of B-Type Oligomeric Procyanidins From Commiphora leptophloeos Used Alone or in Combination With Fluconazole Against Candida spp. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.613155
Donders, G. G. G., Bellen, G., Grinceviciene, S., Ruban, K., Vieira-Baptista, P. (2017). Aerobic vaginitis: no longer a stranger. Res. Microbiol. 168, 845–858. doi: 10.1016/j.resmic.2017.04.004
Donders, G. G. G., Vereecken, A., Bosmans, E., Dekeersmaecker, A., Salembier, G., Spitz, B. (2002). Definition of a type of abnormal vaginal flora that is distinct from bacterial vaginosis: aerobic vaginitis. BJOG.: Internal J. Obs. Gyn. 109, 34–43. doi: 10.1111/j.1471-0528.2002.00432.x
Fan, A., Yue, Y., Geng, N., Zhang, H., Wang, Y., Xue, F. (2013). Aerobic vaginitis and mixed infections: comparison of clinical and laboratory findings. Arch. Gynecol. Obstet. 287, 329–335. doi: 10.1007/s00404-012-2571-4
Feng, F., Hou, Y.-M., Zhang, Y., Wang, L.-Y., Li, P.-P., Guo, Y., et al. (2023). Correlation analysis of vaginal microecology and different types of human papillomavirus infection: a study conducted at a hospital in northwest China. Front. Med. (Lausanne). 10. doi: 10.3389/fmed.2023.1138507
Fox, E. P., Cowley, E. S., Nobile, C. J., Hartooni, N., Newman, D. K., Johnson, A. D. (2014). Anaerobic bacteria grow within Candida albicans biofilms and induce biofilm formation in suspension cultures. Curr. Biol. 24, 2411–2416. doi: 10.1016/j.cub.2014.08.057
Freitas, A. I., Vasconcelos, C., Vilanova, M., Cerca, N. (2014). Optimization of an automatic counting system for the quantification of Staphylococcus epidermidis cells in biofilms. J. Basic. Microbiol. 54, 750–757. doi: 10.1002/jobm.201200603
Gilbert, N. M., O’Brien, V. P., Lewis, A. L. (2017). Transient microbiota exposures activate dormant Escherichia coli infection in the bladder and drive severe outcomes of recurrent disease. PloS Pathog. 13, e1006238. doi: 10.1371/journal.ppat.1006238
Hardy, L., Cerca, N., Jespers, V., Vaneechoutte, M., Crucitti, T. (2017). Bacterial biofilms in the vagina. Res. Microbiol. 168, 865–874. doi: 10.1016/j.resmic.2017.02.001
He, Y., Na, R., Niu, X., Xiao, B., Yang, H. (2021). Lactobacillus rhamnosus and Lactobacillus casei Affect Various Stages of Gardnerella Species Biofilm Formation. Front. Cell. Infect. Microbiol. 11. doi: 10.3389/fcimb.2021.568178
Jang, S.-E., Jeong, J.-J., Choi, S.-Y., Kim, H., Han, M. J., Kim, D.-H. (2017). Lactobacillus rhamnosus HN001 and Lactobacillus acidophilus La-14 Attenuate Gardnerella vaginalis-Infected Bacterial Vaginosis in Mice. Nutrients 9, E531. doi: 10.3390/nu9060531
Janulaitiene, M., Paliulyte, V., Grinceviciene, S., Zakareviciene, J., Vladisauskiene, A., Marcinkute, A., et al. (2017). Prevalence and distribution of Gardnerella vaginalis subgroups in women with and without bacterial vaginosis. BMC Infect. Dis. 17, 394. doi: 10.1186/s12879-017-2501-y
Jung, H.-S., Ehlers, M. M., Lombaard, H., Redelinghuys, M. J., Kock, M. M. (2017). Etiology of bacterial vaginosis and polymicrobial biofilm formation. Crit. Rev. Microbiol. 43, 651–667. doi: 10.1080/1040841X.2017.1291579
Kenfack-Zanguim, J., Kenmoe, S., Bowo-Ngandji, A., Kenfack-Momo, R., Thierry Ebogo-Belobo, J., Kengne-Ndé, C., et al. (2023). Systematic review and meta-analysis of maternal and fetal outcomes among pregnant women with bacterial vaginosis. Eur. J. Obstet. Gynecol. Reprod. Biol. 289, 9–18. doi: 10.1016/j.ejogrb.2023.08.013
Koo, H., Andes, D. R., Krysan, D. J. (2018). Candida-streptococcal interactions in biofilm-associated oral diseases. PloS Pathog. 14, e1007342. doi: 10.1371/journal.ppat.1007342
Li, T., Zhang, Z., Wang, F., He, Y., Zong, X., Bai, H., et al. (2020). Antimicrobial Susceptibility Testing of Metronidazole and Clindamycin against Gardnerella vaginalis in Planktonic and Biofilm Formation. Can. J. Infect. Dis. Med. Microbiol. 2020, 1361825. doi: 10.1155/2020/1361825
Lohse, M. B., Gulati, M., Johnson, A. D., Nobile, C. J. (2018). Development and regulation of single- and multi-species Candida albicans biofilms. Nat. Rev. Microbiol. 16, 19–31. doi: 10.1038/nrmicro.2017.107
Ma, X., Wang, X., Ye, S., Liu, J., Yuan, H., Wang, N. (2022). Biofilm and pathogenic factor analysis of Gardnerella vaginalis associated with bacterial vaginosis in Northeast China. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.1033040
MaChado, A., Cerca, N. (2015). Influence of biofilm formation by gardnerella vaginalis and other anaerobes on bacterial vaginosis. J. Infect. Dis. 212, 1856–1861. doi: 10.1093/infdis/jiv338
MaChado, A., Foschi, C., Marangoni, A. (2022). Editorial: Vaginal dysbiosis and biofilms. Front. Cell Infect. Microbiol. 12. doi: 10.3389/fcimb.2022.976057
Magana, M., Sereti, C., Ioannidis, A., Mitchell, C. A., Ball, A. R., Magiorkinis, E., et al. (2018). Options and limitations in clinical investigation of bacterial biofilms. Clin. Microbiol. Rev. 31, e00084–e00016. doi: 10.1128/CMR.00084-16
Maladkar, M. (2015). Clindamycin, clotrimazole and tinidazole in mixed vaginosis- A “Real world”. Clin. Exp. JGO 3, 49–54. doi: 10.11648/j.jgo.20150303.12
Morrill, S., Gilbert, N. M., Lewis, A. L. (2020). Gardnerella vaginalis as a Cause of Bacterial Vaginosis: Appraisal of the Evidence From in vivo Models. Front. Cell Infect. Microbiol. 10. doi: 10.3389/fcimb.2020.00168
Muñoz-Barreno, A., Cabezas-Mera, F., Tejera, E., MaChado, A. (2021). Comparative effectiveness of treatments for bacterial vaginosis: A network meta-analysis. Antibiot. (Basel). 10, 978. doi: 10.3390/antibiotics10080978
Muzny, C. A., Taylor, C. M., Swords, W. E., Tamhane, A., Chattopadhyay, D., Cerca, N., et al. (2019). An updated conceptual model on the pathogenesis of bacterial vaginosis. J. Infect. Dis. 220, 1399–1405. doi: 10.1093/infdis/jiz342
Niu, X.-X., Li, T., Zhang, X., Wang, S.-X., Liu, Z.-H. (2017). Lactobacillus crispatus Modulates Vaginal Epithelial Cell Innate Response to Candida albicans. Chin. Med. J. (Engl). 130, 273–279. doi: 10.4103/0366-6999.198927
Nyirjesy, P., Banker, W. M., Bonus, T. M. (2020). Physician awareness and adherence to clinical practice guidelines in the diagnosis of vaginitis patients: A retrospective chart review. Population. Health Manage. 23, S13–S21. doi: 10.1089/pop.2020.0258
Onderdonk, A. B., Delaney, M. L., Fichorova, R. N. (2016). The human microbiome during bacterial vaginosis. Clin. Microbiol. Rev. 29, 223–238. doi: 10.1128/CMR.00075-15
Paavonen, J., Brunham, R. C. (2018). Bacterial vaginosis and desquamative inflammatory vaginitis. N. Engl. J. Med. 379, 2246–2254. doi: 10.1056/NEJMra1808418
Pacha-Herrera, D., Erazo-Garcia, M. P., Cueva, D. F., Orellana, M., Borja-Serrano, P., Arboleda, C., et al. (2022). Clustering analysis of the multi-microbial consortium by lactobacillus species against vaginal dysbiosis among Ecuadorian women. Front. Cell Infect. Microbiol. 12. doi: 10.3389/fcimb.2022.863208
Prasad, D., Parween, S., Kumari, K., Singh, N. (2021). Prevalence, etiology, and associated symptoms of vaginal discharge during pregnancy in women seen in a tertiary care hospital in bihar. Cureus 13, e12700. doi: 10.7759/cureus.12700
Qin, H., Liu, Y., Zhai, Z., Xiao, B. (2023). Biofilm-forming capacity and drug resistance of different gardnerella subgroups associated with bacterial vaginosis. Microorganisms 11, 2186. doi: 10.3390/microorganisms11092186
Qulu, W., Mtshali, A., Osman, F., Ndlela, N., Ntuli, L., Mzobe, G., et al. (2023). High-risk human papillomavirus prevalence among South African women diagnosed with other STIs and BV. PloS One 18, e0294698. doi: 10.1371/journal.pone.0294698
Rivers, C. A., Adaramola, O. O., Schwebke, J. R. (2011). Prevalence of bacterial vaginosis and vulvovaginal candidiasis mixed infection in a southeastern american STD clinic. Sexually. Transmitted. Dis. 38, 672–674. doi: 10.1097/OLQ.0b013e31820fc3b8
Røder, H. L., Sørensen, S. J., Burmølle, M. (2016). Studying bacterial multispecies biofilms: where to start? Trends Microbiol. 24, 503–513. doi: 10.1016/j.tim.2016.02.019
Rumyantseva, T. A., Bellen, G., Savochkina, Y. A., Guschin, A. E., Donders, G. G. G. (2016). Diagnosis of aerobic vaginitis by quantitative real-time PCR. Arch. Gynecol. Obstet. 294, 109–114. doi: 10.1007/s00404-015-4007-4
Salinas, A. M., Osorio, V. G., Endara, P. F., Salazar, E. R., Vasco, G. P., Vivero, S. G., et al. (2018). Bacterial identification of the vaginal microbiota in Ecuadorian pregnant teenagers: an exploratory analysis. PeerJ 6, e4317. doi: 10.7717/peerj.4317
Salinas, A. M., Osorio, V. G., Pacha-Herrera, D., Vivanco, J. S., Trueba, A. F., MaChado, A. (2020). Vaginal microbiota evaluation and prevalence of key pathogens in Ecuadorian women: an epidemiologic analysis. Sci. Rep. 10, 18358. doi: 10.1038/s41598-020-74655-z
Schlecht, L. M., Peters, B. M., Krom, B. P., Freiberg, J. A., Hänsch, G. M., Filler, S. G., et al. (2015). Systemic Staphylococcus aureus infection mediated by Candida albicans hyphal invasion of mucosal tissue. Microbiol. (Reading). 161, 168–181. doi: 10.1099/mic.0.083485-0
Sharma, G., Sharma, S., Sharma, P., Chandola, D., Dang, S., Gupta, S., et al. (2016). Escherichia coli biofilm: development and therapeutic strategies. J. Appl. Microbiol. 121, 309–319. doi: 10.1111/jam.13078
Shipitsyna, E., Krysanova, A., Khayrullina, G., Shalepo, K., Savicheva, A., Guschin, A., et al. (2019). Quantitation of all Four Gardnerella vaginalis Clades Detects Abnormal Vaginal Microbiota Characteristic of Bacterial Vaginosis More Accurately than Putative G. vaginalis Sialidase A Gene Count. Mol. Diagn. Ther. 23, 139–147. doi: 10.1007/s40291-019-00382-5
Shiroda, M., Manning, S. D. (2020). Lactobacillus strains vary in their ability to interact with human endometrial stromal cells. PloS One 15, 1–15. doi: 10.1371/journal.pone.0238993
Tansarli, G. S., Kostaras, E. K., Athanasiou, S., Falagas, M. E. (2013). Prevalence and treatment of aerobic vaginitis among non-pregnant women: evaluation of the evidence for an underestimated clinical entity. Eur. J. Clin. Microbiol. Infect. Dis. 32, 977–984. doi: 10.1007/s10096-013-1846-4
Vaneechoutte, M., Guschin, A., Van Simaey, L., Gansemans, Y., Van Nieuwerburgh, F., Cools, P. (2019). Emended description of Gardnerella vaginalis and description of Gardnerella leopoldii sp. nov., Gardnerella piotii sp. nov. and Gardnerella swidsinskii sp. nov., with delineation of 13 genomic species within the genus Gardnerella. Int. J. Syst. Evol. Microbiol. 69, 679–687. doi: 10.1099/ijsem.0.003200
Wang, C., Fan, A., Li, H., Yan, Y., Qi, W., Wang, Y., et al. (2020). Vaginal bacterial profiles of aerobic vaginitis: a case-control study. Diagn. Microbiol. Infect. Dis. 96, 114981. doi: 10.1016/j.diagmicrobio.2019.114981
Wang, Z., Fu, L., Xiong, Z., Qin, Q., Yu, T., Wu, Y., et al. (2016). Diagnosis and microecological characteristics of aerobic vaginitis in outpatients based on preformed enzymes. Taiwanese. J. Obstet. Gynecol. 55, 40–44. doi: 10.1016/j.tjog.2015.06.012
Xiao, B., Disi, A., Qin, H., Mi, L., Zhang, D. (2022). Correlation analysis of vaginal microbiome changes and bacterial vaginosis plus vulvovaginal candidiasis mixed vaginitis prognosis. Front. Cell Infect. Microbiol. 12. doi: 10.3389/fcimb.2022.860589
Zhang, X., Sun, Q., Feng, Y., He, Y., Bai, H., Du, M., et al. (2023). In vitro study on the growth state of intravaginal co-culture of Lactobacillus and Enterococcus faecalis and the inhibitory effect of supernatant on Gardnerella vaginalis. Prog. Obstet. Gynecol. 32, 273–278. doi: 10.13283/j.cnki.xdfckjz.2023.04.032
Zong, X., Feng, Y., Bai, H., Wang, H., Shang, X., Fan, L., et al. (2023). Analysis of vaginal microecology in 23 181 cases of the gynecological female outpatients Analysis of vaginal microecology in 23 181 cases of the gynecological female outpatients. Chin. J. Obstet. Gynecol. 58, 191–197. doi: 10.3760/cma.j.cn112141-20221212-00754
Keywords: Gardnerella vaginalis, Escherichia coli, biofilm, mixed vaginitis, bacterial vaginosis, aerobic vaginitis
Citation: Shang X, Bai H, Fan L, Zhang X, Zhao X and Liu Z (2024) In vitro biofilm formation of Gardnerella vaginalis and Escherichia coli associated with bacterial vaginosis and aerobic vaginitis. Front. Cell. Infect. Microbiol. 14:1387414. doi: 10.3389/fcimb.2024.1387414
Received: 17 February 2024; Accepted: 18 April 2024;
Published: 01 May 2024.
Edited by:
Adline Princy Solomon, SASTRA Deemed to be University, IndiaReviewed by:
Rekha Arya, University of Pittsburgh, United StatesCopyright © 2024 Shang, Bai, Fan, Zhang, Zhao and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhaohui Liu, bGl1emhhb2h1aUBjY211LmVkdS5jbg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.