
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol. , 26 March 2024
Sec. Bacteria and Host
Volume 14 - 2024 | https://doi.org/10.3389/fcimb.2024.1376358
 Zeinab Razooqi1†‡
Zeinab Razooqi1†‡ Ingeborg Tjellström1†Carola Höglund Åberg1‡
Ingeborg Tjellström1†Carola Höglund Åberg1‡ Francis Kwamin2Rolf Claesson1Dorte Haubek3‡
Francis Kwamin2Rolf Claesson1Dorte Haubek3‡ Anders Johansson1‡
Anders Johansson1‡ Jan Oscarsson1*‡
Jan Oscarsson1*‡The Gram-positive bacterium, Filifactor alocis is an oral pathogen, and approximately 50% of known strains encode a recently identified repeat-in-toxin (RTX) protein, FtxA. By assessing a longitudinal Ghanaian study population of adolescents (10-19 years of age; mean age 13.2 years), we recently discovered a possible correlation between deep periodontal pockets measured at the two-year follow-up, presence of the ftxA gene, and a high quantity of F. alocis. To further understand the contribution of F. alocis and FtxA in periodontal disease, we used qPCR in the present study to assess the carriage loads of F. alocis and the prevalence of its ftxA gene in subgingival plaque specimens, sampled at baseline from the Ghanaian cohort (n=500). Comparing these results with the recorded clinical attachment loss (CAL) longitudinal progression data from the two-year follow up, we concluded that carriers of ftxA-positive F. alocis typically exhibited higher loads of the bacterium. Moreover, high carriage loads of F. alocis and concomitant presence of the ftxA gene were two factors that were both associated with an enhanced prevalence of CAL progression. Interestingly, CAL progression appeared to be further promoted upon the simultaneous presence of F. alocis and the non-JP2 genotype of Aggregatibacter actinomycetemcomitans. Taken together, our present findings are consistent with the notion that F. alocis and its ftxA gene promotes CAL during periodontal disease.
Periodontitis is a bacterial-induced oral inflammatory disease, which over time degrades the periodontal tissues, ultimately causing tooth loss (Pihlstrom et al., 2005). Research is indicating that when the natural balance among the resident bacterial species is disturbed, causing a dysbiotic environment, some of the bacteria might exhibit potential pathogenic characteristics, leading to an inflammatory response in the periodontal tissues, which together with factors related to the host susceptibility promotes the degradation (Darveau, 2010). Periodontal diseases affecting adolescents and young individuals typically have been typically referred to as aggressive periodontitis and are at present defined as grade C indicating a rapid progress (Tonetti et al., 2018). There is a wide geographical spread (Bouziane et al., 2020). Among adolescents from the western and northern parts of Africa, the disease has been observed to have a high incidence (Kissa et al., 2022; Yoshida et al., 2021). A highly leukotoxin (LtxA)-producing genotype of the Gram-negative bacterium, Aggregatibacter actinomycetemcomitans, JP2, is associated with the periodontal disease progress in these geographical regions (Haubek et al., 2008; Höglund Åberg et al., 2014). It carries a typical deletion of 530 base pairs (bp) in the ltxCABD promoter (Brogan et al., 1994). To the best of our knowledge, few studies executed in the northern and western geographical regions of Africa have been reported on regarding the relation between periodontitis and the progression of this disease in young individuals, and concomitantly focusing on bacterial species other than A. actinomycetemcomitans (Yoshida et al., 2021; Dahlén et al., 2014), thus including the Gram-positive, anaerobic bacterium Filifactor alocis. This species has recently been identified in the oral microbiome by the method called high throughput DNA sequencing. F. alocis is culturable and is regarded as an emerging oral pathogen with significant roles in the etiology of periodontal (Aruni et al., 2014; Greenwood et al., 2020), peri-implantitis (Sanz-Martin et al., 2017), and endodontic (Zehnder et al., 2017) infections. It is a potential biomarker for active disease in young children (Aruni et al., 2014). Interestingly, A. actinomycetemcomitans and F. alocis seem to have a synergistic relationship in active periodontal disease (Fine et al., 2013; Wang et al., 2013). However, A. actinomycetemcomitans appears thereafter to be outcompeted by F. alocis in the deeper pockets (Dahlén et al., 2014). This suggests that F. alocis has mechanisms to be able to adapt and drive the disease process forward in the absence of A. actinomycetemcomitans. Mechanisms for this are yet unknown.
In the present work, we have assessed longitudinal data from a study population of adolescents in Ghana. This study population was followed up after two years (Höglund Åberg et al., 2014; Höglund Åberg et al., 2012). In the previous studies, cultivation and PCR were used to analyse the pooled subgingival plaque samples, which revealed a 54.2% prevalence of A. actinomycetemcomitans at baseline (Höglund Åberg et al., 2012). Among the individuals, it was also found that 8.8% in the studied population carried the JP2 genotype of A. actinomycetemcomitans (Höglund Åberg et al., 2012). At baseline, an association between clinical attachment loss (CAL) and carriage of A. actinomycetemcomitans was observed, and in addition an increased extent regarding individuals carrying the JP2 genotype (Höglund Åberg et al., 2012).
F. alocis was recently found to encode and express a putative repeats-in-toxin (RTX) protein, FtxA (Bao et al., 2022; Oscarsson et al., 2020). Interestingly, only approximately 50% of the isolated F. alocis strains appeared to carry the ftxA gene (Oscarsson et al., 2020), suggesting the presence of a potentially more virulent genotype of this species. In a recent study (Razooqi et al., 2022), a possible correlation between deep periodontal pockets, presence of the ftxA gene, and a high quantity of F. alocis in the Ghanaian study population could be seen. Moreover, there seemed to be an association between the presence of the ftxA gene, the JP2 genotype of A. actinomycetemcomitans, high levels of F. alocis, and deep periodontal pockets (Razooqi et al., 2022).
To further understand the potential contribution of F. alocis and its ftxA gene in periodontal disease, we aimed at determining the loads of carried F. alocis at baseline among the individuals in the Ghanaian study cohort, and the prevalence of ftxA, and relate these data to the presence and/or absence, respectively of periodontal attachment loss progress observed at the two-year follow-up.
All clinical specimens used in the present work were earlier collected from adolescents (10-19 years of age; mean age 13.2 years) going to 11 Ghanaian schools, and consisted of subgingival plaque samples from nine different sites of periodontal pockets collected with paper points as described (Höglund Åberg et al., 2012). These samples were divided into three groups, referred to as A, B and C. Plaque in group A originated from mesial pockets of the first permanent molars, group B from the distal pockets of the permanent central incisors, and in group C plaque was sampled from one specific individually chosen site. A total of 500 adolescents were included at baseline (Höglund Åberg et al., 2012). At the two-year-follow-up, 397 of the same 500 adolescents could be included, and the clinical recordings were the same as for baseline, i.e., including CAL. Individuals with at least one site with CAL 3 mm or more were classified as CAL-positive. If CAL increased 3 mm or more in at least one site from baseline to the two-year follow-up observations, the individual was classified as ‘CAL progression positive’ (Höglund Åberg et al., 2014). Presence of A. actinomycetemcomitans and its JP2 genotype, respectively in the baseline samples were also determined earlier (Höglund Åberg et al., 2014).
The plaque samples, A, B and C, were pooled, i.e., mixed in equal proportions and subject to genomic DNA isolation using a GXT NA Extraction Kit® (Hain Lifesience, GmBH, Nehren, Germany) and an Arrow extraction instrument (Diasorin, Dublin, Ireland) using procedures described earlier (Razooqi et al., 2022).
For this procedure, DNA isolated from the subgingival plaque samples, A, B and C, were mixed in equal proportions, and served as templates in qPCR for each of the individuals (n=500), respectively. The KAPA SYBR® FAST qPCR Kit (Kapa Biosystems, Wilmington, MA, USA) was used, with cycling conditions as described previously (Razooqi et al., 2022), and with a forward (5’-AGGCAGCTTGCCATACTGCG-3’), and a reverse (5’-ACTGTTAGCAACTACCGATGT-3’) oligonucleotide primer, respectively, targeting the F. alocis 16s rRNA gene (Siqueira and Rocas, 2003). Loads of F. alocis were determined as numbers of cells/ml sample as described (Razooqi et al., 2022).
A forward (5′-GGCTCAGATACCTACTTCTTC-3′) and a reverse (5′-GAAGGCTATGATTTGATTGTTTCC-3′) oligonucleotide primer were used to amplify a 798-base pair (bp) internal fragment of the ftxA gene, as described previously (Oscarsson et al., 2020).
Data analyses were performed using SPSS 22.0 (SPSS Inc., Chicago, IL, USA), or Microsoft Excel (version 16.80). Statistical parameters included those earlier recorded regarding the presence or non-presence of (i) CAL for the individuals at baseline, (ii) progression of CAL recorded at the two-year follow up, (iii) A. actinomycetemcomitans, and (iv) the JP2 genotype of A. actinomycetemcomitans, respectively. Statistical parameters also included the microbial data at baseline deduced in the present work, i.e., presence or non-presence of F. alocis (v), the ftxA gene (vi), and load of F. alocis (vii). Significant differences between sample groups were examined with the Mann–Whitney U test or t-test. Results were estimated by an odds ratio (OR) with 95% confidence interval (CI).
The studies in the present work were ethically approved by Medical Research, University of Ghana (IRB 000 1276), and from the local Ethical committee of Umeå University, Sweden (Dnr 2010-188-31M).
A flow chart summarizing the outline of the present study is shown in Figure 1. For each of the individuals (n=500), a pooled subgingival plaque sample collected at baseline was analyzed with regards to F. alocis loads by qPCR. At baseline, F. alocis was found to be carried by 486 out of the 500 analyzed individuals, i.e., a prevalence of 97.2%. Absence of F. alocis (i.e., F. alocis-negative) was based on a detection level <100 cells/ml sample. Cut off values of 10,000 F. alocis cells/ml sample was then used to divide the individuals as carriers of “high” (≥10,000), and “low” (100<10,000) loads of F. alocis per ml sample, respectively. Out of the 486 individuals carrying F. alocis, 322 (66.3%) were revealed to carry high, and 164 (33.7%) low levels of this bacterium, respectively (Table 1).
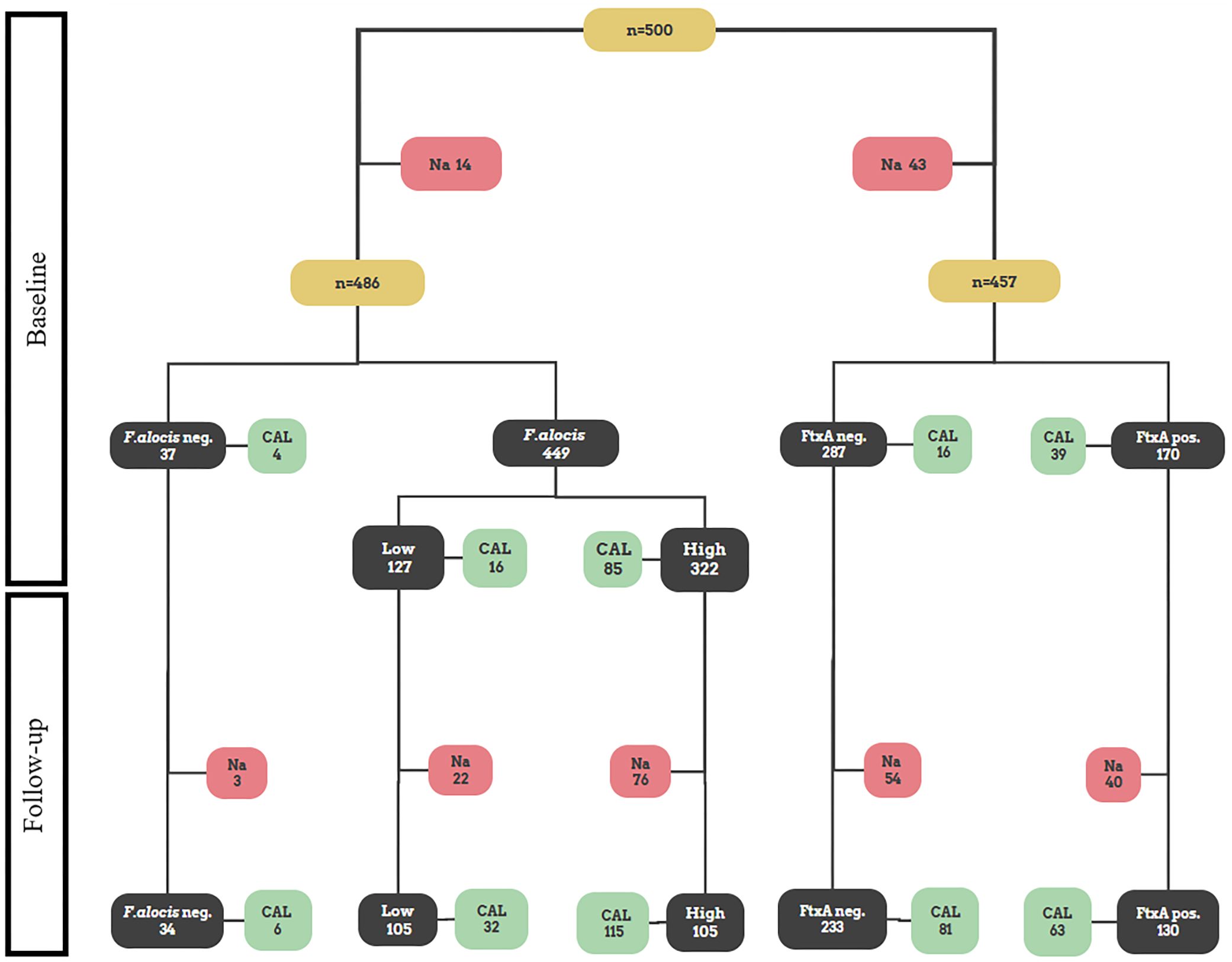
Figure 1 Flowchart illustrating a schematic outline of the present study. All figures in the flowchart represent numbers of patients. Pooled subgingival plaque samples from 500 Ghanaian adolescents were originally sampled at baseline for subsequent analysis of microbial data, and CAL was measured (Höglund Åberg et al., 2012). Out of those, 486 were assessed in the present work regarding presence of high and low levels of F. alocis, respectively, and 457 for presence or absence of the ftxA gene, respectively. Thereafter, at the two-year follow up, the re-examined individuals (n=385) were assessed regarding CAL, and for CAL progression relative to baseline (Höglund Åberg et al., 2014). Indicated are also numbers of non-analyzed samples (Na), which represent samples that were not analyzed due to lack of material.
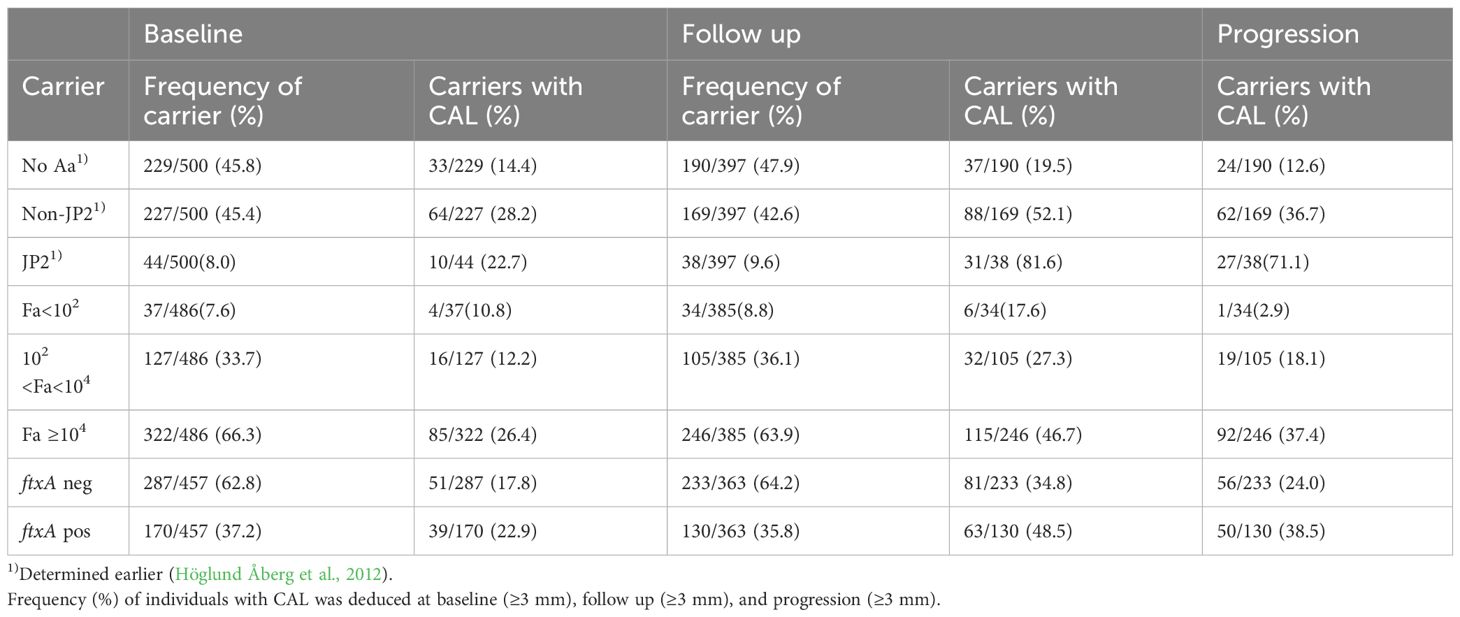
Table 1 Frequency (%) of carriers with CAL among the individuals carrying A. actinomycetemcomitans (Aa) and its JP2 or non-JP2 genotype, and F. alocis (Fa) at low and high loads, and Fa that is ftxA (positive; pos) or without ftxA (negative; neg), respectively.
As summarized in Table 1, at the follow up (n=385), there were 246 (63.9%) individuals who carried high levels of F. alocis at the baseline examination. Of these 246, 46.7% exhibited CAL, and 37.4% a progressed CAL at the follow up relative to baseline. The prevalence of CAL progression was also gradually enhanced relative to increased carriage loads of F. alocis (Figure 2). As summarized in Table 2, relative to the low-level and non-carriers of F. alocis as grouped together (<104 cells/ml sample), those carrying high loads (≥104 cells/ml sample) exhibited a significantly increased prevalence of CAL both at baseline (OR = 2.58; p<0.001), and at follow up (OR = 2.33; p<0.001), and also regarding progression of CAL (OR = 3.56; p<0.001).
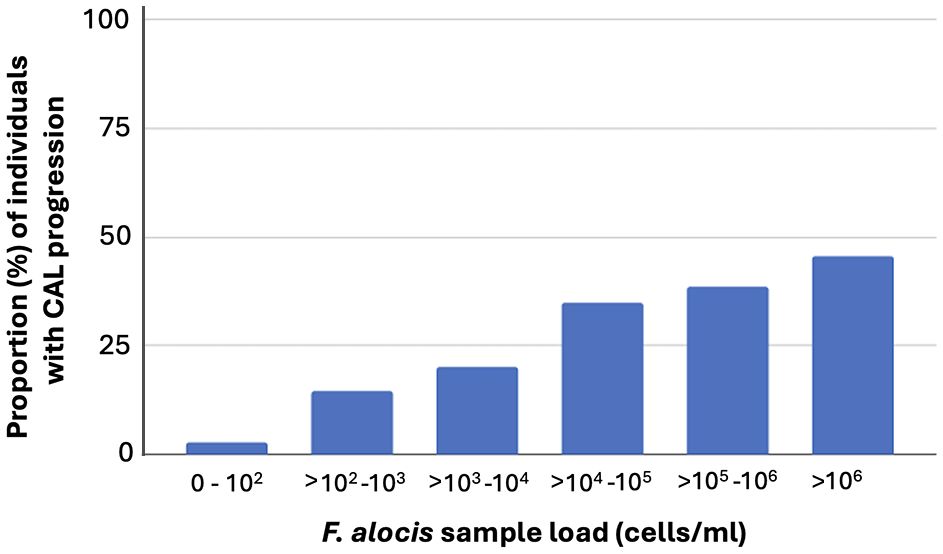
Figure 2 Prevalence (%) of CAL progression among the 385 individuals assessed at the follow up, related to their carriage levels of F. alocis, determined at baseline, in pooled subgingival plaque samples.
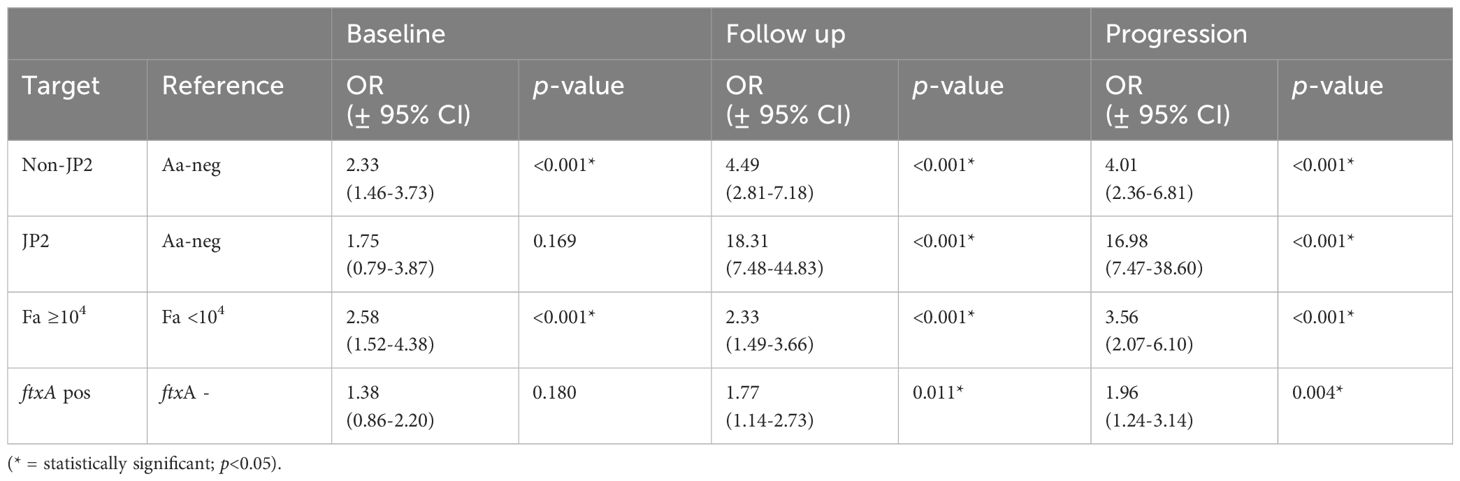
Table 2 Sample group comparisons (target versus reference) as outlined below, related to odds ratio for CAL at baseline (BL ≥3 mm), follow up (FU ≥ 3 mm), and progression (BL-FU ≥3 mm).
The prevalence of the ftxA gene in the analyzed samples at baseline showed a total of 37.2% (i.e., of those 486 that were F. alocis-positive; ≥102 cells/ml sample) (Table 1). Interestingly, the samples analyzed positive for the ftxA gene were revealed to have significantly (p<0.001) higher loads of F. alocis cells/ml in the pooled subgingival plaque samples compared to those that were F. alocis-positive but were detected as ftxA-negative (Figure 3). The proportion of ftxA-positive relative to ftxA-negative F. alocis was also markedly enhanced upon gradually increasing loads of this bacterium present in the samples (Figure 4). Taken together this supports the notion that F. alocis carrying ftxA may exhibit enhanced fitness in the subgingival environment.
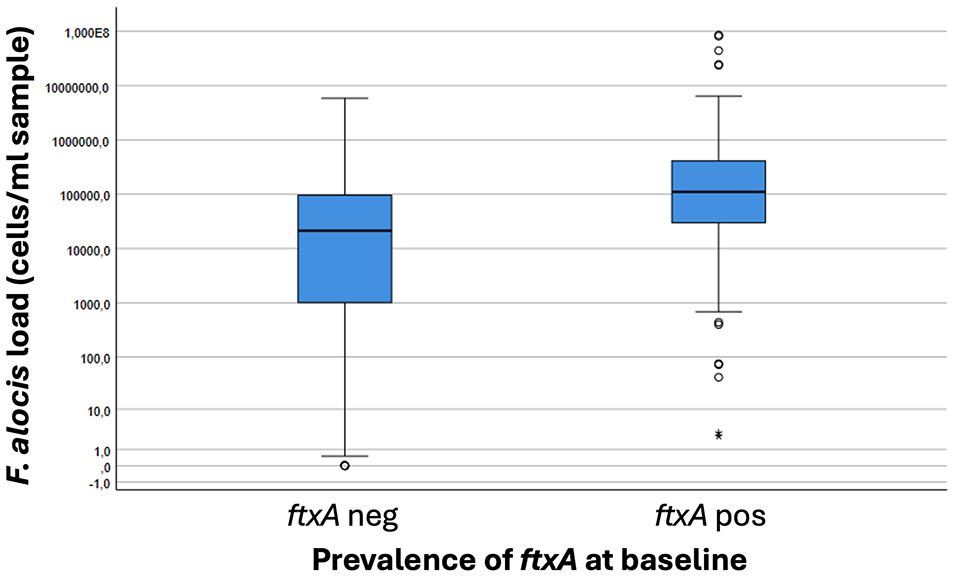
Figure 3 Boxplot illustrating loads of F. alocis in pooled subgingival plaque samples from carriers of this bacterium (n=457) correlated to presence (positive; pos) or absence (negative; neg) of the ftxA gene. P<0.001, ftxA pos vs ftxA neg.
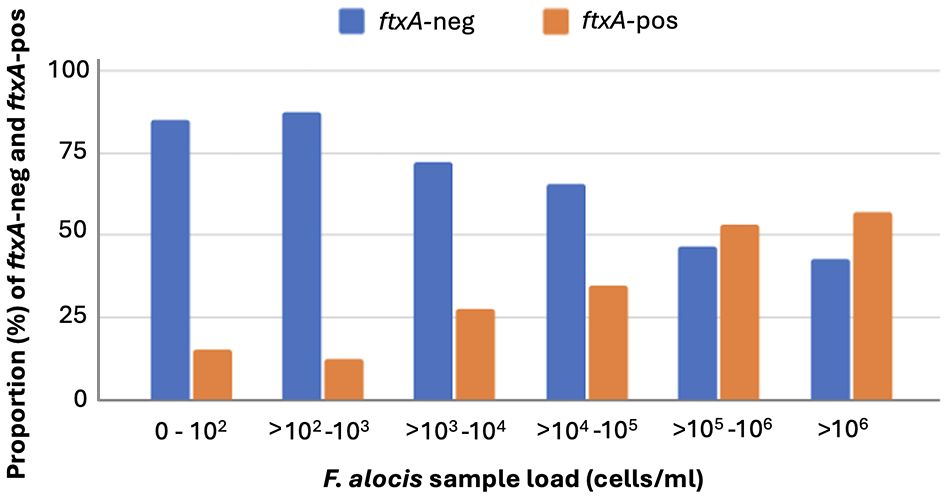
Figure 4 Enhanced proportion of ftxA-positive (-pos) compared to ftxA-negative (-neg) F. alocis upon gradually increasing loads (cells/ml) of this bacterium present in the subgingival plaque samples.
As summarized in Table 1 of the ftxA-positive samples, 39 out of 170, i.e., 22.9% exhibited CAL. At the follow up, 130 (35.8%) were positive for ftxA, and 48.5% of these exhibited CAL. Of the 130 individuals positive for ftxA, 38.5% also showed progression of CAL. Moreover, as summarized in Table 2, carriers of ftxA-positive F. alocis showed a significantly increased prevalence of CAL at follow up (OR=1.77; p=0.011), but not at baseline (OR=1.38; p<0.180). There was also a significant association between progression of CAL and presence of ftxA (OR= 1.96; p<0.04).
To assess possible synergism between F. alocis (Fa) and A. actinomycetemcomitans (Aa), we compared the proportions of individuals with CAL progression between carriers of Fa, but not Aa, and carriers of Fa and non-JP2 Aa, respectively (Figure 5). This revealed that in the absence of A. actinomycetemcomitans, F. alocis was required to be present in high quantities to promote CAL progression. Interestingly, on the other hand, if F. alocis-positive individuals also carried the non-JP2 genotype of A. actinomycetemcomitans, there was a synergistic effect enabling the advancement of CAL even at low loads of these bacterial species. Moreover, there was an apparent distinctive influence of the JP2 genotype of A. actinomycetemcomitans, consistant with an ability to drive CAL progression regardless of if F. alocis was present or not (Figure 5).
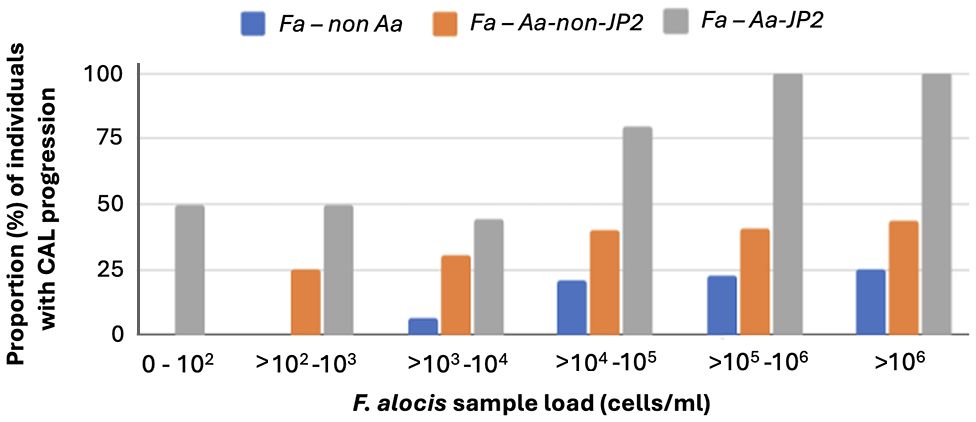
Figure 5 Synergism between F. alocis and non-JP2 genotype of A. actinomycetemcomitans in promoting CAL progression also at low sample loads. The diagram illustrates the proportion (%) of individuals with CAL progression related to F. alocis sample load (cells/ml), and whether the individuals also carried (non-JP2 red bars; JP2 grey bars) or were not carrying (blue bars) A. actinomycetemcomitans, respectively.
In the present work, we have used qPCR and PCR, respectively, to characterize the prevalence and carriage loads of the emerging oral pathogen F. alocis, and its RTX-toxin encoding ftxA gene, in subgingival plaque specimens, sampled at baseline from a longitudinal cohort of Ghanaian adolescents (n=500) (Höglund Åberg et al., 2014; Höglund Åberg et al., 2012). The obtained results were thereafter compared with the recorded CAL and CAL progression data, collected at baseline and at the two-year follow up of the Ghanaian cohort, respectively (Höglund Åberg et al., 2014; Höglund Åberg et al., 2012). The rationale for using this cohort in the present study was that this collection is unique due to being linked to clinical longitudinal data showing attachment loss progression over a two-year time, and it was therefore very suitable for analyzing associations between F. alocis carriage, presence of the ftxA gene, and periodontal disease progression. Our results revealed that carriers of ftxA-positive F. alocis typically exhibited higher loads of the bacterium. Moreover, high carriage loads of F. alocis, and presence of the ftxA gene were two factors both associated with an enhanced prevalence of CAL progression.
The high proportion of individuals with CAL, and CAL progression at the follow up, in the Ghanaian cohort can be partly explained by the presence of A. actinomyctemcomitans at baseline, especially the high proportion (8.8%) of individuals carrying the highly leukotoxic JP2 genotype (Höglund Åberg et al., 2014). However, A. actinomyctemcomitans cannot itself explain all cases of CAL, and CAL progression in this cohort. One reason for assessing F. alocis and its ftxA in relation to CAL in the present work was that there appears to be a synergism between F. alocis and A. actinomycetemcomitans in active periodontal disease, based on observations, such as that A. actinomycetemcomitans could stimulate F. alocis accumulation, albeit that this effect appeared to be dependent on the strain of F. alocis it interacted with (Fine et al., 2013; Wang et al., 2013). Evidently, isolates of A. actinomycetemcomitans from the oral cavity are also heterogeneous with respect to their expression levels of LtxA and production of fimbriae (Haubek, 2010; Rylev et al., 2011), and as suggested (Wang et al., 2013), different strains of A. actinomycetemcomitans might therefore also exhibit different patterns of interaction with F. alocis.
Indeed, the results from our present work would support a scenario where CAL progression is further promoted upon simultaneous presence of F. alocis and a non-JP2 genotype of A. actinomycetemcomitans. This would be consistent with the notion that there may occur genotype-specific synergy between F. alocis and A. actinomycetemcomitans. Mechanism(s) for such interbacterial interactions between these two species, and whether it could be a dependence on FtxA, are yet unknown, but could potentially include interplay based on reduction of the oxidative environment, which evidently was conducted via F. alocis in synergy between this organism and Porphyromonas gingivalis (Mishra et al., 2024). In contrast, CAL progression was not further promoted upon simultaneous presence of F. alocis and the highly leukotoxic JP2 genotype of A. actinomycetemcomitans, which most likely was a result of an apparent distinctive influence of this genotype to drive CAL progression itself, regardless F. alocis was present or not. This would be consistent with the strong leukotoxic activity of the JP2 genotype, as evidenced by the observation that only one bacterial cell was sufficient to lyse the majority of macrophage cells in in vitro experiments (Kelk et al., 2008).
Interestingly, A. actinomycetemcomitans seems to be outcompeted by F. alocis in the deeper periodontal pockets (Dahlén et al., 2014), suggesting that this bacterium can have the capability to adapt and drive the disease process forward in later stages, in the absence of A. actinomycetemcomitans. Concomitantly, we recently discovered a possible correlation between deep periodontal pockets at the two-year follow up of the Ghanaian cohort, presence of the ftxA gene, and a high quantity of F. alocis (Razooqi et al., 2022). As the relatively newly discovered FtxA toxin of F. alocis belongs to the same family of RTX proteins as the A. actinomycetemcomitans leukotoxin, LtxA (Oscarsson et al., 2020; Linhartova et al., 2010), we hypothesized that high F. alocis levels, and carriage of F. alocis encoding ftxA, might correlate to CAL and to CAL progression in the cohort. Indeed, our present results supported the idea that carriers of ftxA-positive F. alocis typically exhibited higher loads of the bacterium. Whether this depends on mechanism(s) directly executed by FtxA is not known, but as presence of the ftxA gene as well as high carriage loads of F. alocis were two factors both associated with an enhanced prevalence of CAL progression, we suggest that FtxA represents a factor promoting the disease progression, at least in younger individuals in these ages.
Carriers of ftxA-positive F. alocis typically exhibited higher loads of the bacterium. Moreover, high carriage loads of F. alocis and the presence of the ftxA gene were two factors that were both associated with an enhanced prevalence of CAL progression. Interestingly, CAL progression appeared to be further promoted upon simultaneous presence of F. alocis and non-JP2 genotype of Aggregatibacter actinomycetemcomitans. Taken together, our present findings are consistent with the notion that F. alocis and its ftxA gene promote CAL during periodontal disease. Hence, ftxA appears to have the potential to be used as a PCR-based gene marker for the identification of F. alocis that are more prone to promote disease progress. This finding might be of importance in the risk assessment in young individuals and is therefore a relevant discovery for development of new diagnostic tools and/or treatment strategies.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
The studies involving humans were approved by Medical Research, University of Ghana (IRB 000 1276), and from the local Ethical committee of Umeå University, Sweden (Dnr 2010-188-31M). The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from primarily isolated as part of your previous study for which ethical approval was obtained. Written informed consent for participation was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and institutional requirements.
ZR: Formal analysis, Investigation, Methodology, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. IT: Formal analysis, Investigation, Validation, Writing – original draft, Writing – review & editing. CHÅ: Writing – review & editing, Resources, Supervision. FK: Resources, Writing – review & editing. RC: Conceptualization, Formal analysis, Investigation, Methodology, Resources, Validation, Writing – review & editing. DH: Resources, Writing – review & editing. AJ: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. JO: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by TUA grants from the County Council of Region Västerbotten, Sweden (to JO; grant number 7003766, and to AJ; 7003193), and by grants from the Medical Faculty of Umeå University (Insamlingsstiftelsen to JO and AJ, respectively.
We are grateful to Björn Tavelin, statistician at the Department of Radiation Sciences, Umeå University, for help with the statistical calculations. We also wish to thank Carina Öhman, Jessica Radu, and Karin Lilja for valuable technical assistance.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Aruni, W., Chioma, O., Fletcher, H. M. (2014). Filifactor alocis: the newly discovered kid on the block with special talents. J. Dent. Res. 93, 725–732. doi: 10.1177/0022034514538283
Bao, K., Claesson, R., Gehrig, P., Grossmann, J., Oscarsson, J., Belibasakis, G. N. (2022). Proteomic characterization of the oral pathogen filifactor alocis reveals key inter-protein interactions of its RTX toxin: ftxA. Pathogens 11, 590. doi: 10.3390/pathogens11050590
Bouziane, A., Hamdoun, R., Abouqal, R., Ennibi, O. (2020). Global prevalence of aggressive periodontitis: A systematic review and meta-analysis. J. Clin. Periodontol 47, 406–428. doi: 10.1111/jcpe.13266
Brogan, J. M., Lally, E. T., Poulsen, K., Kilian, M., Demuth, D. R. (1994). Regulation of Actinobacillus actinomycetemcomitans leukotoxin expression: analysis of the promoter regions of leukotoxic and minimally leukotoxic strains. Infect. Immun. 62, 501–508. doi: 10.1128/iai.62.2.501-508.1994
Dahlén, G., Claesson, R., Åberg, C. H., Haubek, D., Johansson, A., Kwamin, F. (2014). Subgingival bacteria in Ghanaian adolescents with or without progression of attachment loss. J. Oral. Microbiol. 6, 23977. doi: 10.3402/jom.v6.23977
Darveau, R. P. (2010). Periodontitis: a polymicrobial disruption of host homeostasis. Nat. Rev. Microbiol. 8, 481–490. doi: 10.1038/nrmicro2337
Fine, D. H., Markowitz, K., Fairlie, K., Tischio-Bereski, D., Ferrendiz, J., Furgang, D., et al. (2013). A consortium of Aggregatibacter actinomycetemcomitans, Streptococcus parasanguinis, and Filifactor alocis is present in sites prior to bone loss in a longitudinal study of localized aggressive periodontitis. J. Clin. Microbiol. 51, 2850–2861. doi: 10.1128/JCM.00729-13
Greenwood, D., Afacan, B., Emingil, G., Bostanci, N., Belibasakis, G. N. (2020). Salivary microbiome shifts in response to periodontal treatment outcome. Proteomics Clin. Appl. 14, e2000011. doi: 10.1002/prca.202000011
Haubek, D. (2010). The highly leukotoxic JP2 clone of Aggregatibacter actinomycetemcomitans: evolutionary aspects, epidemiology and etiological role in aggressive periodontitis. APMIS. Supplementum 130, 1–53. doi: 10.1111/j.1600-0463.2010.02665.x
Haubek, D., Ennibi, O. K., Poulsen, K., Vaeth, M., Poulsen, S., Kilian, M. (2008). Risk of aggressive periodontitis in adolescent carriers of the JP2 clone of Aggregatibacter (Actinobacillus) actinomycetemcomitans in Morocco: a prospective longitudinal cohort study. Lancet 371, 237–242. doi: 10.1016/S0140-6736(08)60135-X
Höglund Åberg, C., Kwamin, F., Claesson, R., Dahlen, G., Johansson, A., Haubek, D. (2014). Progression of attachment loss is strongly associated with presence of the JP2 genotype of Aggregatibacter actinomycetemcomitans: a prospective cohort study of a young adolescent population. J. Clin. Periodontol 41, 232–241. doi: 10.1111/jcpe.12209
Höglund Åberg, C., Kwamin, F., Claesson, R., Johansson, A., Haubek, D. (2012). Presence of JP2 and non-JP2 genotypes of Aggregatibacter actinomycetemcomitans and attachment loss in adolescents in Ghana. J. Periodontol 83, 1520–1528. doi: 10.1902/jop.2012.110699
Kelk, P., Claesson, R., Chen, C., Sjöstedt, A., Johansson, A. (2008). IL-1beta secretion induced by Aggregatibacter (Actinobacillus) actinomycetemcomitans is mainly caused by the leukotoxin. Int. J. Med. Microbiol. 298, 529–541. doi: 10.1016/j.ijmm.2007.06.005
Kissa, J., El Houari, B., Amine, K., Chemlali, S., Khlil, N., Mikou, S., et al. (2022). Prevalence of periodontal disease in young Moroccans: A national survey. J. Periodontol 93, 1867–1877. doi: 10.1002/JPER.22-0103
Linhartova, I., Bumba, L., Masin, J., Basler, M., Osicka, R., Kamanova, J., et al. (2010). RTX proteins: a highly diverse family secreted by a common mechanism. FEMS Microbiol. Rev. 34, 1076–1112. doi: 10.1111/j.1574-6976.2010.00231.x
Mishra, A., Dou, Y., Wang, C., Fletcher, H. M. (2024). Filifactor alocis enhances survival of Porphyromonas gingivalis W83 in response to H2O2-induced stress. Mol. Oral. Microbiol. 39, 12–26. doi: 10.1111/omi.12445
Oscarsson, J., Claesson, R., Bao, K., Brundin, M., Belibasakis, G. N. (2020). Phylogenetic analysis of filifactor alocis strains isolated from several oral infections identified a novel RTX toxin, ftxa. Toxins (Basel) 12, 687. doi: 10.3390/toxins12110687
Pihlstrom, B. L., Michalowicz, B. S., Johnson, N. W. (2005). Periodontal diseases. Lancet 366, 1809–1820. doi: 10.1016/S0140-6736(05)67728-8
Razooqi, Z., Höglund Åberg, C., Kwamin, F., Claesson, R., Haubek, D., Oscarsson, J., et al. (2022). Aggregatibacter actinomycetemcomitans and Filifactor alocis as Associated with Periodontal Attachment Loss in a Cohort of Ghanaian Adolescents. Microorganisms 10, 2511. doi: 10.3390/microorganisms10122511
Rylev, M., Bek-Thomsen, M., Reinholdt, J., Ennibi, O. K., Kilian, M. (2011). Microbiological and immunological characteristics of young Moroccan patients with aggressive periodontitis with and without detectable Aggregatibacter actinomycetemcomitans JP2 infection. Mol. Oral. Microbiol. 26, 35–51. doi: 10.1111/j.2041-1014.2010.00593.x
Sanz-Martin, I., Doolittle-Hall, J., Teles, R. P., Patel, M., Belibasakis, G. N., Hammerle, C. H. F., et al. (2017). Exploring the microbiome of healthy and diseased peri-implant sites using Illumina sequencing. J. Clin. Periodontol 44, 1274–1284. doi: 10.1111/jcpe.12788
Siqueira, J. F., Jr., Rocas, I. N. (2003). Detection of Filifactor alocis in endodontic infections associated with different forms of periradicular diseases. Oral. Microbiol. Immunol. 18, 263–265. doi: 10.1034/j.1399-302X.2003.00073.x
Tonetti, M. S., Greenwell, H., Kornman, K. S. (2018). Staging and grading of periodontitis: Framework and proposal of a new classification and case definition. J. Periodontol 89 Suppl 1, S159–S172. doi: 10.1002/JPER.18-0006
Wang, Q., Wright, C. J., Dingming, H., Uriarte, S. M., Lamont, R. J. (2013). Oral community interactions of Filifactor alocis in vitro. PloS One 8, e76271. doi: 10.1371/journal.pone.0076271
Yoshida, A., Bouziane, A., Erraji, S., Lakhdar, L., Rhissassi, M., Miyazaki, H., et al. (2021). Etiology of aggressive periodontitis in individuals of African descent. Jpn Dent. Sci. Rev. 57, 20–26. doi: 10.1016/j.jdsr.2020.12.001
Keywords: Filifactor alocis, FtxA, RTX toxin, Aggregatibacter actinomycetemcomitans, JP2, periodontitis, clinical attachment loss (CAL)
Citation: Razooqi Z, Tjellström I, Höglund Åberg C, Kwamin F, Claesson R, Haubek D, Johansson A and Oscarsson J (2024) Association of Filifactor alocis and its RTX toxin gene ftxA with periodontal attachment loss, and in synergy with Aggregatibacter actinomycetemcomitans. Front. Cell. Infect. Microbiol. 14:1376358. doi: 10.3389/fcimb.2024.1376358
Received: 25 January 2024; Accepted: 18 March 2024;
Published: 26 March 2024.
Edited by:
Li Zhang, University of New South Wales, AustraliaReviewed by:
Gunnar Dahlen, University of Gothenburg, SwedenCopyright © 2024 Razooqi, Tjellström, Höglund Åberg, Kwamin, Claesson, Haubek, Johansson and Oscarsson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jan Oscarsson, amFuLm9zY2Fyc3NvbkB1bXUuc2U=
†These authors have contributed equally to this work
‡ORCID: Zeinab Razooqi, orcid.org/0000-0001-9715-8039
Carola Höglund Åberg, orcid.org/0000-0003-0765-6435
Dorte Haubek, orcid.org/0000-0001-9983-8767
Anders Johansson, orcid.org/0000-0002-8069-8263
Jan Oscarsson, orcid.org/0000-0002-7948-9464
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.