
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Infect. Microbiol., 06 March 2024
Sec. Virus and Host
Volume 14 - 2024 | https://doi.org/10.3389/fcimb.2024.1360586
This article is part of the Research TopicMpox: Understanding the scientific gaps to combat the threatView all 7 articles
 Emmanuel Alakunle1
Emmanuel Alakunle1 Daniel Kolawole1
Daniel Kolawole1 Diana Diaz-Cánova2
Diana Diaz-Cánova2 Faith Alele3
Faith Alele3 Oyelola Adegboye4
Oyelola Adegboye4 Ugo Moens2*
Ugo Moens2* Malachy Ifeanyi Okeke1*
Malachy Ifeanyi Okeke1*Monkeypox virus (MPXV) is the etiological agent of monkeypox (mpox), a zoonotic disease. MPXV is endemic in the forested regions of West and Central Africa, but the virus has recently spread globally, causing outbreaks in multiple non-endemic countries. In this paper, we review the characteristics of the virus, including its ecology, genomics, infection biology, and evolution. We estimate by phylogenomic molecular clock that the B.1 lineage responsible for the 2022 mpox outbreaks has been in circulation since 2016. We interrogate the host-virus interactions that modulate the virus infection biology, signal transduction, pathogenesis, and host immune responses. We highlight the changing pathophysiology and epidemiology of MPXV and summarize recent advances in the prevention and treatment of mpox. In addition, this review identifies knowledge gaps with respect to the virus and the disease, suggests future research directions to address the knowledge gaps, and proposes a One Health approach as an effective strategy to prevent current and future epidemics of mpox.
Monkeypox virus (MPXV) is the etiological agent of a zoonotic disease called monkeypox (mpox). It is a double-stranded DNA (dsDNA) virus belonging to Orthopoxvirus (OPXV) genus within the Poxviridae family and Chordopoxvirinae as the subfamily (Alakunle et al., 2020). Other members of this genus include Variola virus (VARV), Cowpox virus (CPXV), Vaccinia virus (VACV), Camelpox virus (CMLV), Taterapox virus (TATV) and Ectromelia virus (ECTV). MPXV is divided into Clade I and Clade II, with Clade II subclassified as Clade IIa and IIb (Happi et al., 2022). For five decades, MPXV was endemic in West and Central Africa (Earl et al., 2012), and exportation of the virus to non-endemic regions was rare (Alakunle and Okeke, 2022). However, the incidence (since 2017) of mpox outside endemic regions has increased, and the epidemiological profile of the disease within endemic regions has changed (Grothe et al., 2022). This may have led to the MPXV emergence and re-emergence in endemic countries in 2022 (Alakunle and Okeke, 2022). This paper will cover the current state of knowledge on the characteristics of MPXV and mpox, the infection biology, molecular pathogenesis, and evolution of MPXV as well as the clinical features, diagnosis, epidemiology, and therapeutic options against mpox. In addition, the review will critically interrogate and evaluate the contributions of viral, host, and anthropogenic factors to the emergence and reemergence of mpox across the globe.
Before 1970, there was no documented report of human MPXV infection, although the virus had previously caused infections in monkeys and apes (Arita and Henderson, 1968). Infections in monkeys were reported in laboratory/captive animals and were first identified in captive monkeys in Denmark in 1958. The first human mpox case emerged in a 9-month-old boy in the Democratic Republic of the Congo (DRC) in August 1970 (Ladnyj et al., 1972). Subsequently, six additional mpox cases were identified between September 1970 and April 1971 in Liberia, Sierra Leone and Nigeria (Lourie et al., 1972). Since then, MPXV has been reported in several countries and is endemic in Benin, Cameroon, the Central African Republic, the DRC, Gabon, Ivory Coast, Liberia, Nigeria, the Republic of the Congo, Sierra Leone, and South Sudan (Bass et al., 2013; World Health Organization, 2022a).
Figure 1 displays the global mpox outbreak timeline. Between 1970 and 2021, the cases have been sporadic and geographically limited within endemic regions (Brown and Leggat, 2016; Titanji et al., 2022). Notably, the DRC is the only country that continuously reports yearly cases of mpox with tropical rainforest regions accounting for 98.7% of all cases pre-2022 (Brown and Leggat, 2016; Durski et al., 2018). In Nigeria, sporadic cases were reported in the 1970s; however, re-emergence of the disease started in 2017 with an eleven-year-old boy as the index case in Bayelsa state (Yinka-Ogunleye et al., 2018). At the end of 2017, Nigeria recorded 88 cases, and during this outbreak, travel-related cases in non-endemic countries were reported, including the United Kingdom (UK), the United States of America (USA), Israel, and Singapore, between 2018 and 2021 (Adegboye et al., 2022).
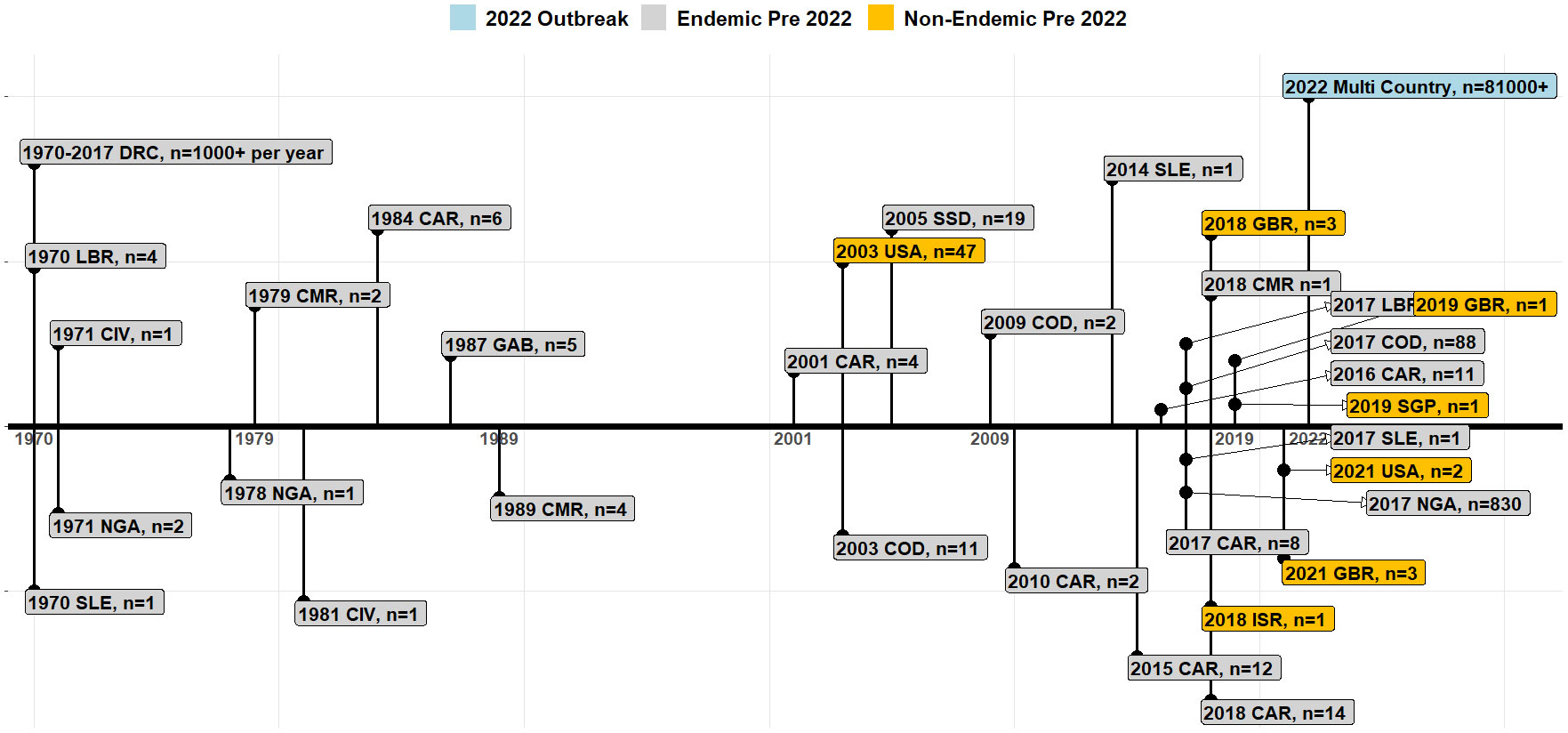
Figure 1 Timeline of MPXV emergence and re-emergence in endemic regions and globally. Each event timeline indicates the pre-2022 outbreak in endemic and non-endemic countries and the 2022 mpox outbreak (Durski et al., 2018; World Health Organization, 2022). Note: Cameroon (CMR), the Central African Republic (CAR), Cote d’Ivoire (CIV), the Democratic Republic of the Congo (DRC), Gabon (GAB), the United Kingdom (GBR), Liberia (LBR), Nigeria (NGA), Israel (ISR), Sierra Leone (SLE), Singapore (SGP), the Republic of the Congo (COD), South Sudan (SSD), the United States of America (USA).
The first mpox outbreak in a non-endemic country was reported in 2003 in the USA linked to importation of rodents from Ghana (Figure 1) (Anderson et al., 2003; Centre for Disease Control, 2003; Croft et al., 2007). By the end of the outbreak, 47 people had been infected (10 probable and 37 confirmed cases) (Centre for Disease Control, 2003; Center for Diseases Control and Prevention (CDC), 2023). There were no other travel-related cases reported until 2018. Between 2018 to 2021, 11 travel-related mpox cases were recorded in the UK, Singapore, Israel, and the USA (Figure 1). Of these, four resulted in secondary cases: one healthcare worker in the UK was infected by contaminated bedding, an adult and a child from a family from the UK had a travel history to Nigeria, and one traveler to Israel who had visited Nigeria in 2018. Between 2019 and 2021, a total of seven mpox outbreaks occurred outside Africa in Singapore, the UK and the USA (Figure 1). All travel related cases originated in Nigeria, with high-throughput sequencing confirming it as Clade II (Vaughan et al., 2018; Erez et al., 2019; Fang et al., 2020; Hobson et al., 2021; Bunge et al., 2022). Between 2017 and October 30, 2022, a total of 830 cases were recorded in 33 out of 36 states in Nigeria (Nigeria Centre for Disease Control (NCDC), 2022).
The current global mpox outbreak started in May 2022 (Alakunle et al., 2020; Adegboye et al., 2022) and was declared a public health emergency of international concern on July 23, 2022 (Nuzzo et al., 2022). As of August 02, 2023, there were a total of 88,600 laboratory-confirmed cases and 152 deaths (case-fatality rate, 0.17%) across 113 countries including 106 countries that have not historically reported mpox (Figures 2A-C) (World Health Organization, 2022). The Americas recorded the highest number of cases during the 2022 mpox outbreak, with the USA (n = 29,513) and Brazil (n = 10,168) accounting for 48.32% of the total cases (Figure 2C). Other notable affected countries include Spain (n = 7,408), France (n = 4,110), Colombia (n = 3,880), the UK (n = 3,730), Germany (n = 3,673), Peru (n = 3,561), Mexico (n = 3,455), and Canada (n = 1,459). In Africa, Nigeria has the highest mpox cases with 634 cases.
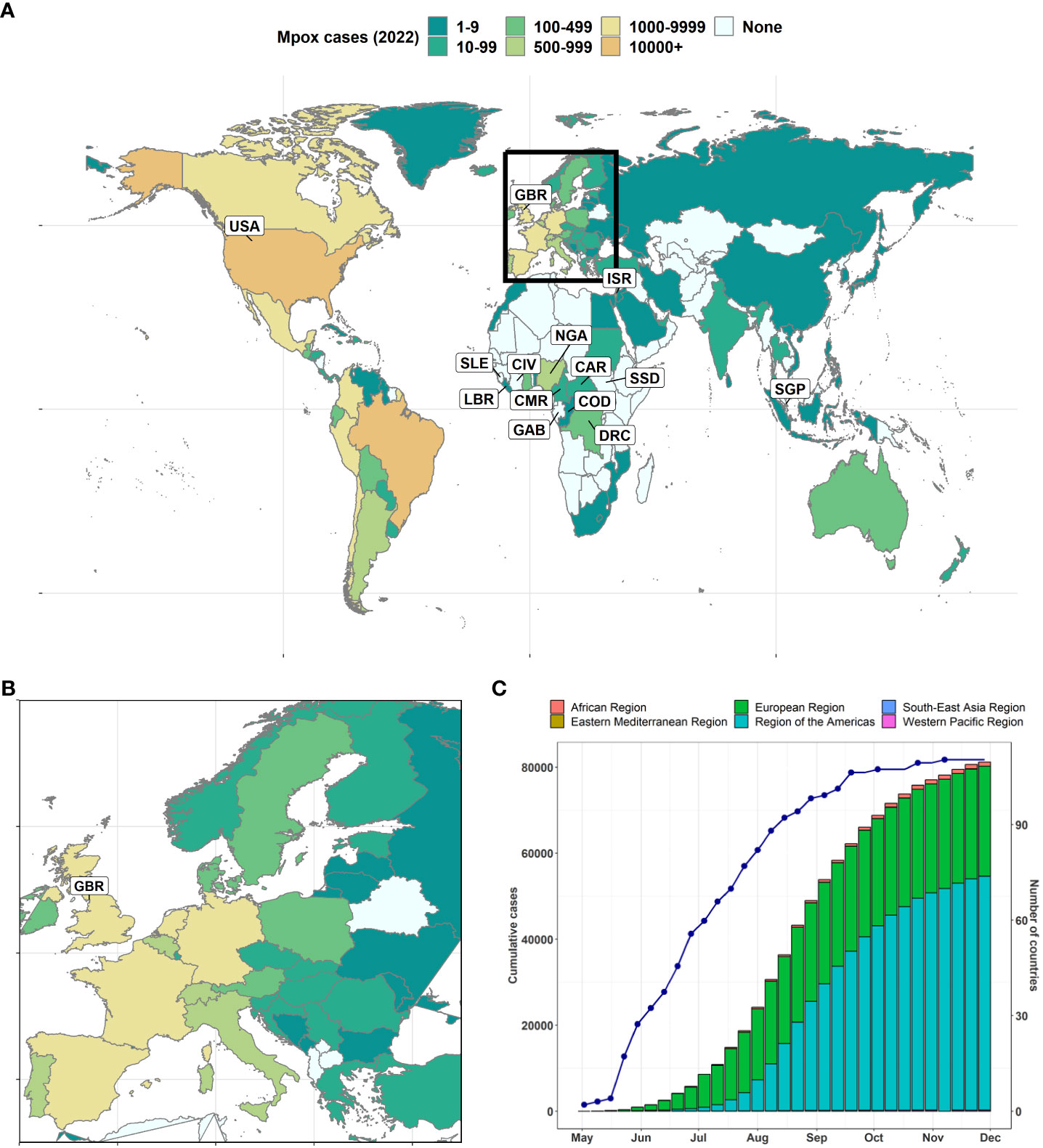
Figure 2 Timeline of MPXV re-emergence and global spread. (A) Global map of 2022 mpox of the geographical distribution of the outbreak as of December 5, 2022 (Durski et al., 2018; World Health Organization, 2022). The names of countries with at least one case pre-2022 are labelled. (B) Global maps zoom on Europe. (C) Weekly cumulative number of cases reported to World Health Organization (WHO) stacked by WHO region. The line/dot represents the cumulative number of countries affected. Countries: Cameroon (CMR), the Central African Republic (CAR), Cote d’Ivoire (CIV), the Democratic Republic of the Congo (DRC), Gabon (GAB), the United Kingdom (GBR), Liberia (LBR), Nigeria (NGA), Israel (ISR), Sierra Leone (SLE), Singapore (SGP), the Republic of the Congo (COD), South Sudan (SSD), the United States of America (USA).
Despite the name, monkeypox, monkeys are not the genuine reservoir of MPXV. Several animals can naturally or experimentally be infected with MPXV (Table 1) (Li et al., 2023), but the natural host reservoir remains elusive. While specific host-cell receptors are responsible for cell tropism, the specificity of MPXV is yet to be determined. Factors like the monkeypox inhibitor of complement enzymes (MOPICE) and complement control protein (CCP) can influence the viral cellular and tissue tropism (Hudson et al., 2012). Nonetheless, a wide spectrum of tissue and host tropism is expected, which may explain the possibility of MPXV establishing animal reservoirs in non-endemic regions (Kmiec and Kirchhoff, 2022). Organs such as ovaries, kidneys, heart, brain, pancreas, liver, and lung have been identified as some of the tissue tropism for MPXV (Arthur et al., 2022). However, specific virus ligands remain unidentified. The inability to identify specific virus ligands and cognate host receptors for MPXV tropism suggests that the virus uses many alternative ligands to successfully invade host cells or the host receptors have functional redundancy to the virus ligand. Spillover to humans (zoonotic transmission) might arise from the disruptions of the natural habitats of wild animals (Domán et al., 2022). This could occur via various routes, including aerosol, direct contact, and fomite transmission (Walker, 2022). It is believed that the MPXV outbreaks in Africa prior to 2022 occurred as a result of a spillover from animals to humans (Faye et al., 2018; Kabuga and El Zowalaty, 2019; Petersen et al., 2019b; Happi et al., 2022; Riopelle et al., 2022). Thus, there is a likelihood of MPXV being sustained in the spillover due to the wide geographical coverage of the MPXV hosts (Tu, 2015).
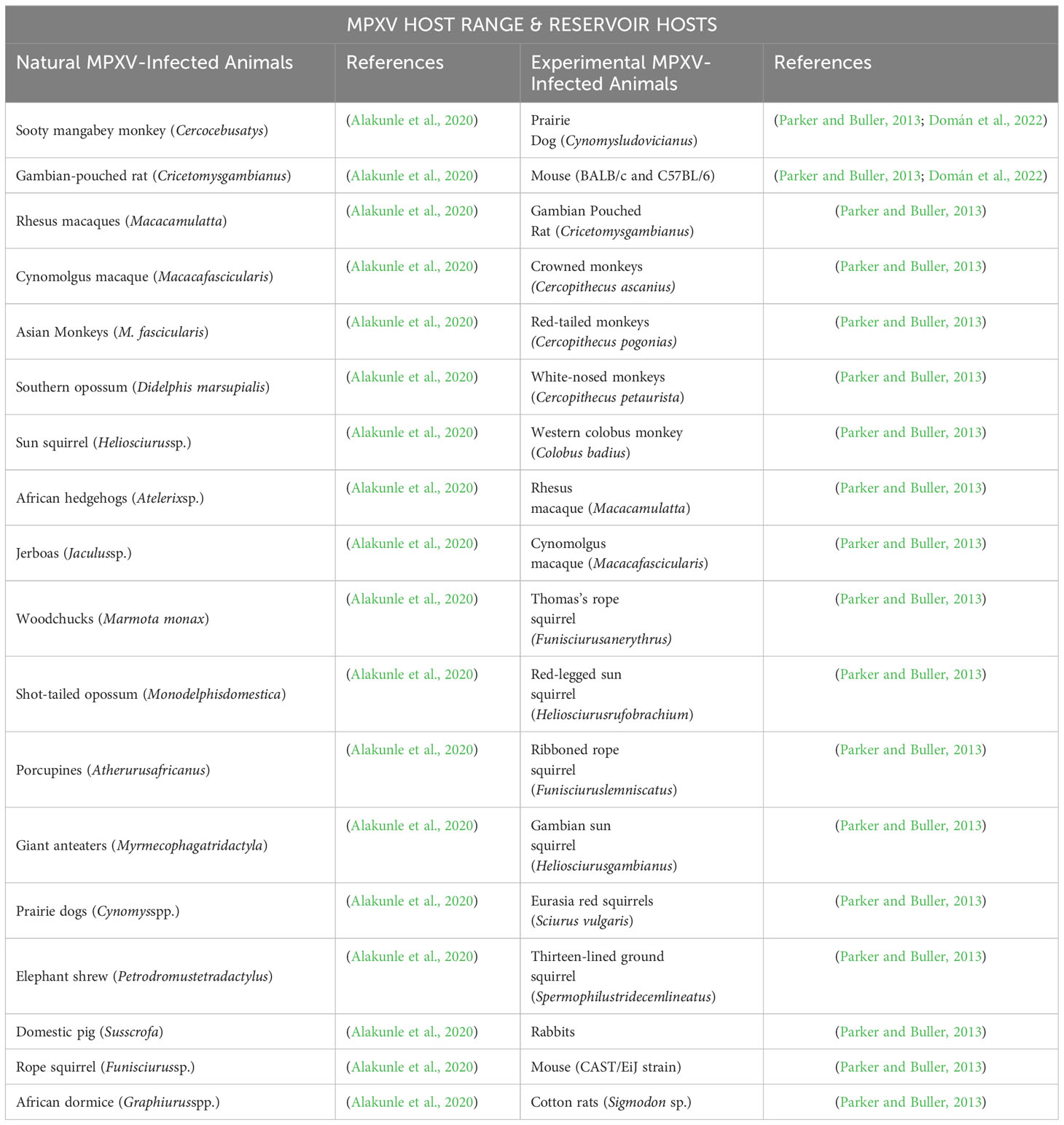
Table 1 List of natural MPXV-infected animals and experimental MPXV-infected animals.
MPXV has a long and complex genome of 196 Kbp - 211 Kbp with a conserved central region and variable inverted terminal repeats (ITR) (Figure 3A). Within MPXV clades, Clade I isolates have more uniform genome length (196 Kbp - 199 Kbp) than Clade II isolates (196 Kbp - 211 Kbp). The length of MXPV ITR varies between 6.5 Kbp to 17.5 Kbp (Shchelkunov et al., 2002; Likos et al., 2005; Nakazawa et al., 2013). Our genomic comparison of MPXV clades (Clade I, n=23; Clade IIa, n=13 and Clade IIb, n=161) indicated similar gene synteny (excluding the ITR), but slight differences in the gene content among the clades due to some genes that are missing or truncated in either one or two clades. The results are consistent with a previous study (Forni et al., 2022a) which indicates that four genes (D14L, D15L, D16L and D17L) are missing and three genes (D4L, B14L and B15L) are truncated in Clade II. Furthermore, the homologue of VACV-Cop E5R is only absent in Clade I and three genes (K1R, homologues of VACV-Cop A47L and VACV-Cop B11R) are truncated. Another major difference is the gene content of their left terminal in which Clade IIa contains four genes (N3R, N2R, N1R and R1R) that are absent in Clade I and IIb. However, these four genes are present on the right terminal of all clades. Furthermore, Clade I and IIb have theD2L gene. Within Clade IIb, gene content of the lineage A and B.1 are similar.
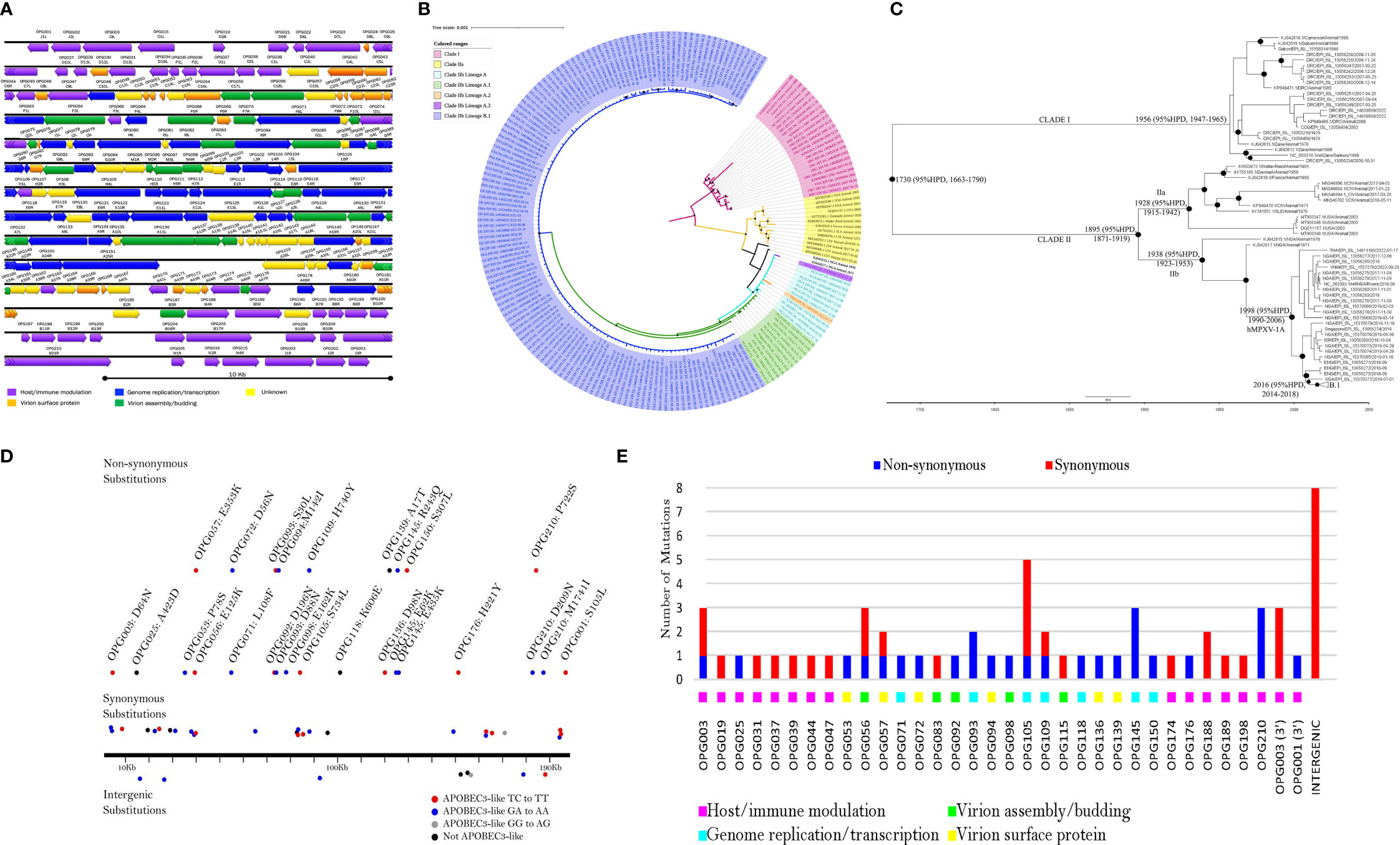
Figure 3 Genome annotation, phylogenomic tree, maximum clade credibility tree, and mutation map of MPXV. (A) Schematic presentation of MPXV genome. Annotation is given for MPXV reference strain NC_063383.1. The orientation of ORFs is given by the direction of arrow heads. ORFs are named according to the nomenclature of orthopoxvirus genes (example: OPG001) (Senkevich et al., 2021) and VACV Western Reserve nomenclature (example: J1L). MPXV genome consists of a conserved central region (OPG048 to OPG151) flanked by variable terminal regions, which contain inverted terminal repeats (ITR) (Shchelkunov et al., 2001; Shen-Gunther et al., 2023). The central region encodes genes for genome replication, essential enzymes, and structural proteins. Conversely, the variable terminal regions contain mainly virulence and host-range genes (Shchelkunov et al., 2001). MPXV genome encompasses >190 nonoverlapping open reading frames (ORFs) (Shchelkunov et al., 2001; Hendrickson et al., 2010; Shen-Gunther et al., 2023) and at least 4 ORFs are located in ITR (Shchelkunov et al., 2002; Likos et al., 2005; Nakazawa et al., 2013). ORFs are colored based on their function. (B) Bayesian Inference phylogenetic tree of concatenated 62 non-recombinant conserved genes from 197 MPXV genomes retrieved from GenBank and GISAID. Recombination detection program 4 (RDP4) (Martin et al., 2015) was used to detect recombination in the 62 conserved genes (Diaz-Cánova et al., 2022a) and the phylogenetic tree was reconstructed using MrBayes v3.2.7 (Ronquist et al., 2012), as previously published (Diaz-Cánova et al., 2022a). Black squares at the nodes indicate posterior probabilities ≥ 0.95. The scale bar represents expected substitutions per site. (C) Time-Scaled Bayesian Inference phylogenetic tree of concatenated 62 non-recombinant conserved genes from 197 MPXV strains. The presence of a temporal signal within the dataset was examined by regression of genetic divergence (root-to-tip genetic distance) and the sampling date using TempEst v.1.5.3 (Rambaut et al., 2016). The Maximum likelihood tree of 62 non-recombinant conserved genes built as described previously (Diaz-Cánova et al., 2022a) was used for TempEst. The maximum-clade-credibility (MCC) tree was generated using BEAST 1.10.4 (Suchard et al., 2018), using a log-normal strict clock, constant population size, and HKY substitution model. Markov Chain Monte Carlo (MCMC) chains were run until reaching convergence. The convergence of MCMC chains was checked by the effective sample size (ESS) values >200 for each parameter (after burn-in) using Tracer v1.7.1 (Rambaut et al., 2018). The maximum-clade-credibility (MCC) tree was generated using TreeAnnotator v1.10.4. Black circles at the nodes indicate posterior probabilities ≥ 0.9. The scale bar represents expected substitutions per site. (D) Mutation map showing all 60 consensus substitutions. 51 of the 60 consensus nucleotide substitutions possessing APOBEC3 like pattern of mutation (GA > AA, GG > AG, and TC > TT). Twenty-eight of these were GA > AA substitutions, two were GG > AG (this mutational pattern is a product of APOBEC3G), twenty-one were TC > TT, and the remaining nine substitutions were not typical of APOBEC3 editing. (E) Synonymous and non-synonymous mutational count.
Our Bayesian phylogenetic (BI) analysis of 62 non-recombinant conserved genes (Diaz-Cánova et al., 2022a) from 197 MPXV isolates resolved MPXV into three monophyletic clades, namely Clade I (Congo Basin Clade), Clade IIa (West Africa Clade) and Clade IIb consisting majorly of human MPXV (hMPXV) isolated between 2017-2022 (Figure 3B). Clade IIb was further divided into lineages: A (n=11), A.1 (n=11), A.2 (n=1), A.3 (n=1), and B.1 (n=135) as assigned by the GISAID (Global Initiative on Sharing All Influenza Data) (https://gisaid.org/). MPXV tree topology reported here is similar to that of the trees reported previously (Gigante et al., 2022; Isidro et al., 2022; Luna et al., 2022; Wang et al., 2022). Isidro et al. reported that the transition from A.1/A.1.1 to B.1 is characterized by a long, divergent branch (Isidro et al., 2022) which suggests accelerated microevolution. The findings of this current study (Figure 3B) agree with the suggestion.
Lineage A (hMPXV-1A) corresponds to the 2017–2019 outbreak, although it contains strains isolated after this time frame (Figure 3B) and this observation has been reported by others (Gigante et al., 2022). A recent study showed that new Nigerian hMPXV genomes isolated in 2019-2020 were identified as belonging to the lineage A (Ndodo et al., 2023). Lineage B.1 contains most hMPXV genomes from 2022 (Figure 3B). It is poorly resolved, and its sub-lineages cannot be unequivocally assigned (Figure 3B). The low clade supports are probably due to very low genetic variability among isolates (Scarpa et al., 2022). Polytomy within lineage B.1 could be an indication of uncertainty in the relatedness of isolates or the belief that those isolates evolved independently from a single origin (Slowinski, 2001; Phylogenetic pitchforks - Understanding Evolution, 2023), although recombination cannot be excluded (Yeh et al., 2022).
Furthermore, molecular dating analysis was carried out on the 62 non-recombinant conserved genes of 197 MPXV isolates to estimate the evolutionary rate and the time of the Most Recent Ancestor (tMRCA) (Figure 3C). TempEst analysis showed temporal signal in the dataset (R=0.65). Maximum Clade Credibility (MCC) tree (Figure 3C) demonstrated that MPXV emerged at 1730 (95% high posterior density interval (HPD), 1663 – 1790) with a mean evolutionary rate estimated to be 5.68 × 10−6 (subs/site/year), with 95% HPD of 4.53 × 10−6 − 6.86 × 10−6 subs/site/year. A recent publication estimated the substitution rate to be 5 x 10-6 (Dumonteil et al., 2023) which is in agreement with our result, but previous estimations of the substitution rates of Clades I and IIa were smaller (Gigante et al., 2022). Clades I, IIa, and IIb were estimated to have emerged in 1956, 1928, and 1938, respectively which is earlier by 33, 47, and 97 years as estimated by Forni et al. (Forni et al., 2022b). Lineage A and B.1 were estimated to have a tMRCA of 1998 (95% HPD, 1990 - 2006) and 2016 (95% HPD, 2014 – 2018), respectively. Our dating analysis put the emergence of the B.1 lineage six years earlier than that estimated by Luna et al. (Luna et al., 2022) and Nextclade (https://clades.nextstrain.org/). This discrepancy may be explained by molecular dating methods used (ML-TSP versus BI-MCC). We hypothesize that the emergence of lineage A about 1998 allowed enough time for lineage A evolution and that may explain the long divergent branch length from lineage A to B.1. Similarly, the emergence of lineage B.1 in 2016 in tandem with clustering of some 2022 isolates within the lineage A (2017-2019) support cryptic transmission of the B.1 viruses prior to the current 2022 outbreak in multiple non-endemic countries (Alakunle and Okeke, 2022; Dumonteil et al., 2023). A recent molecular clock analysis based on accumulation of APOBEC3 (apolipoprotein B mRNA editing enzyme, catalytic polypetide 3)-type mutations inferred that MPXV with APOBEC3 editing has been in circulation since at least 2016 (O’Toole et al., 2023), and this is in agreement with our molecular dating reported herein although their interpretation differs from ours. While we infer that B.1 lineage emerged in 2016, O`Toole et al. concluded that APOBEC3-type mutations, a predictor of human-to-human transmission emerged in 2016. The close phylogenetic relationship between A.1 isolates from Nigeria (in particular EPI_ISL_15370077) with B.1 suggests a single origin for lineage B.1.
Our mutational analysis revealed that lineage B.1 has 66-86 nucleotide substitutions (60 consensus) and 28-39 amino acid substitutions (26 consensus) compared to reference genome NC_063383.1, MPXV from 2018 (Figures 3D, E). Consensus substitutions predominantly affect genes responsible for host/immune modulation and viral replication/transcription (Figure 3E). B21R (OPG210) codes for a T cell suppressor; our analysis demonstrated that this open reading frame (ORF) contained three consensus amino acid substitutions (D209N, P722S, and M1741I). A18R (OPG145) contained three substitutions (E62K, R243Q, and E435K). Excluding G8R (OPG093) with two substitutions, all the other affected ORFs contained one consensus amino acid substitution each (Figure 3E). The impact of these substitutions on the pathogenicity, transmissibility, immune evasion, and host specificity of MPXV remains unknown. The E353K substitution within F13L (OPG057) affects the target for the antiviral agent, tecovirimat; however, functional studies have not revealed any effect on the efficacy of the drug (Gigante et al., 2022). Wang et al. also reported three mutations in OPG210 protein in MPXV 2022 outbreak strains (Wang et al., 2022). Additionally, they showed that this protein together with other nine proteins (D2L‐like, OPG023, OPG047, OPG071, OPG105, OPG109, A27L‐like, OPG153, and OPG188 proteins) have more mutations compared to the other MPXV proteins (Wang et al., 2022).
In contrast to our study, previous mutational studies have reported fewer consensus substitutions (Isidro et al., 2022; Wang et al., 2022; Wassenaar et al., 2022). They reported 46 shared mutations in MPXV genomes from 2022 outbreak compared with NC_063383 (Chen et al., 2005; Isidro et al., 2022). However the number of mutations identified here and in other studies is higher than one would expect for MPXV (Chen et al., 2005; Isidro et al., 2022; Ndodo et al., 2023), based on the low substitution rate of OPXV (Firth et al., 2010). The accelerated evolution of MPXV has been attributed to the human APOBEC3 (Isidro et al., 2022; O’Toole and Rambaut, 2022; Wang et al., 2022). Since most mutations identified here (Figure 3D) and elsewhere were GA>AA and TC>TT mutations, which are compatible with APOBEC3 activity. APOBEC3 is a cytidine deaminase known to play important functions in innate anti-viral immunity (Harris and Dudley, 2015; Salter et al., 2016). Evidence for the accumulation of APOBEC3 enriched substitutions in MPXV isolates has been reported since 2017, which corresponds to the detection of lineage A (Gigante et al., 2022). Ever since, every year, there has been an increase in APOBEC3-like mutations in MPXV (Ndodo et al., 2023). This mutational pattern has become more pronounced as suggested from more recent isolates (B.1) (Gigante et al., 2022; Ndodo et al., 2023), (Figure 3D). The recent pattern of upsurge of APOBEC3 derived mutations may be an indication of a change in virus-host interaction such as sustained human-human transmission (Ndodo et al., 2023) or a new route of infection (Gigante et al., 2022). Gigante et al. (Gigante et al., 2022) suggested the possibility of recombination in MPXV following observation of three sequences that showed a chimeric pattern in their genomes, although assembly errors could be a plausible explanation. Recently, tandem repeats and linkage disequilibrium analysis provided evidence of natural recombination in lineage B.1 (Yeh et al., 2022).
Poxvirus mature particles are ovoid or brick-shaped with surface tubules and have a characteristic dumbbell-shaped nucleoprotein core containing the viral genome (Shchelkunov et al., 2001). MPXV virions are ~200 nm in diameter and ~300 nm in length (Wenner et al., 1968). Like other OPXVs, MPXV forms three distinct infectious virus particles: intracellular mature virus (IMV), cell associated enveloped virus (CEV) and extracellular enveloped virus (EEV) although CEV and EEV are morphologically and structurally indistinguishable. Morphogenesis and transmission of IMV, CEV, and EEV are described in Figure 4A.
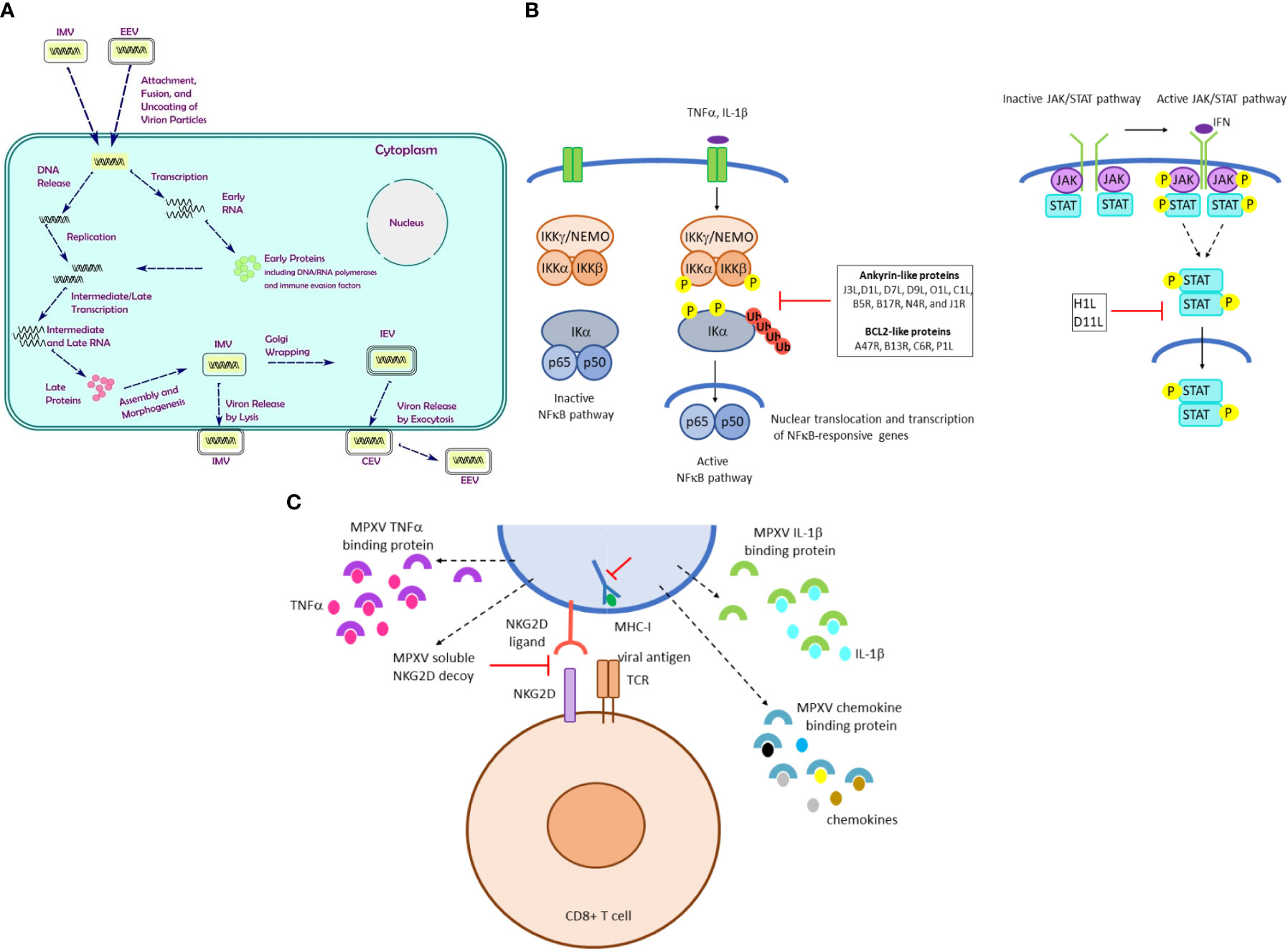
Figure 4 MPXV morphogenesis, signaling and immune evasion strategies. (A) The MPXV replication cycle. After attachment of the virion to the host cell membrane, the viral genome is released by uncoating followed by transcription and duplication of the viral genome. Translation and subsequent assembly results in IMV. MPXV virions can exist as intracellular mature virus (IMV), cell associated enveloped virus (CEV) and extracellular-enveloped virus (EEV). IMV assembles in the cytoplasm and consists of a core particle wrapped in a membrane. IMV particles egress from the infected cells by lysis, whereas some IMVs are transported through microtubules and wrapped by an intracellular membrane to produce an intracellular enveloped virus (IEV), which can further fuse with the cell membrane and be released to form EEV (Schmidt and Mercer, 2012; Sivan et al., 2016; Realegeno et al., 2020). Some EEVs remain attached to the cell surface (CEV) and are responsible for cell-to-cell spread, whereas EEV that detaches from the infected cells play a role in long-range dissemination within the host (Blasco and Moss, 1992). IMV particles are thought to be the form responsible for inter-host viral transmission. In contrast, EEV are known to be important for intra-host viral dissemination (Payne, 1980). (B) MPXV strategies to activate signaling pathways. Left panel: The canonical NFκB pathway requires activation of the cytoplasmic p65/p50 dimer, which is anchored into the cytoplasm through its interaction with IKα. The trimer IKKα/IKKβ/IKKγ will upon activation phosphorylate IKα, which is subsequently ubiquitinated and probed for proteasomal degradation. This results in release of p65/p50, which translocate to the nucleus and can induce transcription of NFκB target genes. Several MPXV proteins can perturb the NFκB pathway. Right panel: The JAK/STAT pathway consist of the tyrosine kinases JAK that phosphorylate and activate the transcription factor STAT. The MPXV proteins encoded by the genes H1L and D11L can inhibit activation of the JAK/STAT pathway. (C) MPXV avoids detection of virus-infection cells by cytotoxic CD8+T cells. Infected host cells will present viral peptide fragments by MHC-I molecules. These will be recognized by T cell receptors (TCR). Co-recognition of NKG2D ligand on the MPXV-infected cell and the NKG2D receptor on the CD8+ T cell is required. MPXV will prevent the latter interaction by expressing soluble NKG2D. Moreover, the cytokine gradient produced upon MPXV infection will be disturbed by MPXV-encoded proteins that will bind these cytokines such as TNFα, IL-1β.
Observations of cynomolgus monkeys infected with MPXV gave the first indications of the pathogenic role of this virus with intramuscular injection of the virus leading to an intense inflammatory immune response (Wenner et al., 1968, 1969). The pathogenesis of MPXV has also been studied in other animals, including rabbits, rodents and prairie dogs (Hutson et al., 2009; Bunge et al., 2022). Studies in monkeys, mice, and prairie dogs demonstrated that Clade I viruses are more virulent than Clade II strains, which reflects the situation in humans (Saijo et al., 2009; Hutson et al., 2010; Bunge et al., 2022; Americo et al., 2023; Falendysz et al., 2023). Except for disease severity, the clinical features of the two clades are similar (Kipkorir et al., 2022).
MPXV infection has an incubation period of 5–21 days and the most common symptoms for the 2022 human mpox outbreak in non-endemic regions based on 48,622 patients were skin lesions (95%), fever (58%), lymphadenopathy (53%), fatigue/asthenia (39%), myalgia (31%), and headache (30%) (Liu et al., 2023). Regarding skin lesions, anogenital lesions were most frequent (66%), followed by lesions on the trunk/torso (48%), face/head (39%), and extremities (~30%) (Liu et al., 2023). This is in contrast with previous human mpox outbreaks from 1980-2022 where skin lesions were most common on the face and extremities (Pourriyahi et al., 2023). Lymphadenopathy, which typically occurs 1-2 days before rash, is a distinct feature of MPXV which is used to distinguish it from smallpox and chickenpox (Altindis et al., 2022; McCarthy, 2022). The morphological progression of the rash is macular, popular, vesicular, and pustular lesions. The crust formed by pustules desquamate after 1-2 weeks (Altindis et al., 2022; McCarthy, 2022; Nakhaie et al., 2022). In the current outbreak, inguinal lymphadenopathy was more frequent than cervical and axillary lymphadenopathy (Liu et al., 2023), whereas in previous outbreaks in endemic countries submandibular, cervical and axillary lymphadenopathy were more frequent (Damon, 2011). The differences in clinical symptoms between human mpox infections before 2022 and the current outbreak are probably the result of different virus strain (Clade I versus Clade II, respectively) and patient group (both female and male youngsters versus mainly adult men having sex with men). In men having sex with men (MSM), atypical clinical symptoms such as genital lesions and anal ulcers were observed (Mitjà et al., 2022; Bragazzi et al., 2023).
MPXV can enter the host via the oral/respiratory tract, infecting the oral and respiratory tract mucosae, with the upper, middle and lower airway epithelium as main targets for primary infection (Giulio and Eckburg, 2004; Damon, 2011; Kipkorir et al., 2022; Mitjà et al., 2022). MPXV can directly infect damaged skin, and replicate in keratinocytes, fibroblasts, and endothelial cells (Mitjà et al., 2022). From the initial infection sites, virus can spread to draining lymph nodes, where the virus can replicate. MPXV can subsequently reach the tonsils, the spleen, and the liver. Replication in these organs results in a second viraemia wave, enabling the virus to access distant organs such as the lung, kidneys, intestines, and skin and causing recognizable clinical manifestation (Giulio and Eckburg, 2004; Damon, 2011; Kipkorir et al., 2022; Mitjà et al., 2022).
Comparison of the Clade I and II strains has provided an indication which viral gene products may be responsible for their difference in virulence. The D14L gene which encodes valosin-containing protein (VCP)-MPXV, also known as CCP or MOPICE, is the orthologue of VACV secreted complement C3b/C4b-binding protein VACV-Cop C3L (Chen et al., 2005). This protein is known to inhibit the complement and to contribute to virulence (Kotwal et al., 1990; Isaacs et al., 1992). Compared to the VACV protein, VCP-MPXV/MOPICE is truncated due to a single nucleotide deletion in the D14L gene leading to a stop codon (Uvarova and Shchelkunov, 2001). The gene is absent from Clade II strains due to ~10 kbp deletion (Likos et al., 2005). Deletion of the C3L gene caused VACV attenuation in infected animal models (Isaacs et al., 1992; DeHaven et al., 2010; Girgis et al., 2011). Studies with Clade I MPXV with deleted D14L showed that intranasal infection of prairie dogs resulted in decreased morbidity and mortality. However, ablation of D14L did not significantly affect virus replication compared to animals infected with control Clade I virus. On the other hand, Clade II virus with inserted D14L gene did not have virulence compared to Clade I virus and no apparent effect on disease-associated mortality compared to control Clade II virus was observed (Hudson et al., 2012). So, other factors besides VCP-MPXV/MOPICE are necessary to explain the difference in virulence between Clade I and II MPXV.
Other candidate virulence genes include BR-203, BR-209, and the OPXV major histocompatibility complex class I–like protein orthologue (OMCP or N3R) (Chen et al., 2005; Likos et al., 2005). BR-203 protein is an orthologue to the myxoma virus M-T4, which plays a role in avoiding apoptosis of infected lymphocytes, hence promoting viral spread within the host (Barry et al., 1997; Weaver and Isaacs, 2008). Myxoma virus expressing C-terminal truncated M-T4 caused increased inflammatory response compared to rabbits infected with wildtype virus, whereas challenging with virus lacking M-T4 resulted in disease attenuation (Barry et al., 1997; Hnatiuk et al., 1999). Thus, BR-203 may have a dual function in protecting infected lymphocytes from apoptosis and in modulating the inflammatory response to virus infection.
The BR-209 gene encodes a 326 aa interleukin-1β (IL-1β) binding protein, which prevents IL-1β from interacting with the IL-1 receptor. Mice intranasally infected with VACV lacking IL-1β binding protein developed a more severe illness than wildtype virus (Alcami and Smith, 1992). Perturbing the IL-1 signaling pathway dampens the innate and acquired immunity, explaining the virulent action of the IL-1β binding protein (Dinarello, 2018). Interestingly, the BR-209 gene of Clade I strains has two ORFs that can encode a putative N-terminal protein fragment of 210 aa and a C-terminal protein fragment of 126 aa, whereas the Clade II strains encode a putative N-terminal 163 aa polypeptide and a C-terminal 132 aa fragment (Weaver and Isaacs, 2008). It is unknown if any of the fragments function in a way similar to the full-length protein, nor if the differences in the length of the N-terminal fragments of Clade I versus Clade II strains of MPXV contribute to the differences in virulence (Weaver and Isaacs, 2008).
The OMCP gene (N3R) from Clade I strains isolated in the DRC carries a 628 bp deletion, which removes the 5’region of the N3R gene (which encodes the OMCP) and the N2R gene (encoding a 73 aa polypeptide with unknown function) (Kugelman et al., 2014). OMPC functions as soluble natural killer group 2, member D receptor (NKG2D) ligand, possibly representing a strategy to avoid NK-cell-mediated killing (Campbell et al., 2007; Lazear et al., 2013). Based on the clinical metadata, it was suggested that the deletion is associated with increased human-to-human transmission and pathogenicity compared to Clade IIa and Clade IIb isolates (Forni et al., 2022a).
The Clade I strain also contains truncated orthologues of VACV-Cop E3L and VACV-Cop K3L, two proteins that function in interferon (IFN) resistance (Weaver and Isaacs, 2008). Clade I and Clade II genomes differ also in VACV-CopH5R (the VACV orthologue encodes a late transcription factor, which plays a role in viral replication, transcription and morphogenesis (Van Vliet et al., 2009), VACV-Cop A9L (VACV orthologue encodes a morphogenesis factor) (Van Vliet et al., 2009), VACV-Cop A50R (VACV orthologue encodes a DNA ligase) (Van Vliet et al., 2009)), and VACV-Cop A36R (playing role in actin tail formation) (Wolffe et al., 1998)). The exact mechanisms by which these proteins may also contribute to differences in pathogenicity between Clade I and II strains remains elusive.
Manipulation of signaling pathways promotes viral replication and determines disease outcomes, mainly by targeting cell growth and immune responses (Krajcsi and Wold, 1998; Greber, 2002). MPXV infection suppresses expression of host genes whose products are implicated in regulation of histone expression, cytoskeletal rearrangements, cell cycle progression, IFN-associated genes, and signaling pathways such as the nuclear factor kappa B (NFκB), the mitogen-activated protein kinase (MAPK), and metabolic pathways (Alkhalil et al., 2010; Rubins et al., 2011; Xuan et al., 2022).
The NFκB pathway plays pivotal roles in inflammation and immunity (Shao-Cong Sun, 2017). NFκB is typically found as a cytosolic trimer of p65/p50 and the inhibitor protein IκB. Phosphorylation of IκB by the trimer IKKα/IKKβ/IKKγ results in the release and activation of p65/p50, which translocate to the nucleus and act as a transcription factor (Hayden and Ghosh, 2008) (Figure 4B). Ankyrin proteins from OPXVs can inhibit the NFκB pathway by interfering with different components of this pathway (Shisler and Jin, 2004; Chen et al., 2008; Mohamed and McFadden, 2009; Ember et al., 2012; Mansur et al., 2013; Herbert et al., 2015). MPXV genome encodes the ankyrin-like proteins J3L, D1L, D7L, D9L, O1L, C1L, B5R, B17R, N4R, and J1R (Shchelkunov et al., 2002; Lum et al., 2022), but their exact function remains to be determined. In addition, the MPXV BCL2-like proteins A47R, B13R, C6R, and P1L can prevent activation of the NFκB pathway (Lum et al., 2022). The MPXV B1R gene encodes the VACV Kelch-like protein A55, which inhibits the NFκB pathway and stimulates CD8+ T cell proliferation (Lysakova-Devine et al., 2010). However, it remains to be determined whether B1R exerts a similar function.
The Janus kinase (JAK) and signal transducer and activator of transcription (STAT) pathway mediates cellular responses to cytokines and growth factors (Mythology, 1990). H1L inactivates STAT1, and C6R blocks STAT2 (Mann et al., 2008; Stuart et al., 2016). Whether the MPXV H1L and D11L orthologues act similarly has not been investigated (Figure 4B).
The VACV epidermal growth factor homologue VGF (C11R) usurps the epidermal growth factor receptor (EGFR) pathway to provoke cell proliferation and to stimulate efficient virus spread and pathogenesis (Buller et al., 1988; Beerli et al., 2019). The D3R gene (OPG019) encodes an EGF homologue that might activate the EGFR pathway and enhance viral dissemination. VACV-Cop F11L also promotes viral spreading by inhibiting the RhoA signaling pathway (Beerli et al., 2019). It is not known whether the MPXV orthologue C17L possesses the same function.
Circumventing apoptosis is used by viruses to achieve productive replication. VACV F1L and N1L proteins can prevent apoptosis by inhibiting pro-apoptotic proteins BAK, BID, BAD, and BAX (Aoyagi et al., 2006; Cooray et al., 2007). It is unknown whether the MPXV C7L and P1L isologues have similar functions. Other MPXV genes that prevent apoptosis include B12R, B19R, D5R, and F3L (Lum et al., 2022). B12 is a serine protease and its VACV orthologue B13R has been shown to be a caspase inhibitor (Li and Beg, 2000).
Signal transduction is often mediated through a cascade of phosphorylation events (Denhardt, 1996). Kindrachuk et al. compared the phosphorylation pattern of host cell proteins after Clade I or Clade II MPXV infection (Kindrachuk et al., 2012). They found that Clade I MPXV infection down-regulated pathways related to cell proliferation and apoptosis as compared with Clade II MPXV. Differences in MPXV-induced posttranslational modification may explain the differences in virulence between MPXV clades.
Human infection with MPXV is associated with increased levels of ILs, C-C motif chemokine ligand 2 (CCL2) and CCL5 (Johnston et al., 2015), and a significant decrease in tumor necrosis factor alpha (TNF-α), IFN-γ, and IL-2 and IL-12 (Li et al., 2022). MPXV infection provokes IgM and IgG antibodies, long-term persistence of residual IgG-memory B cells, and a rapid expansion of activated effector CD4+ and CD8+ T cells followed by a decrease over time (Agrati et al., 2022; Mitjà et al., 2022). MPXV also interferes with adaptive immune responses of antiviral CD8 + and CD4 + T cell responses via inhibiting T cell receptor-mediated T cell activation (Hammarlund et al., 2008).
Because of the high homology between the MPXV genes and the corresponding OPXV orthologues, it is assumed and, in some cases, demonstrated that MPXV applies similar strategies to evade the host immune defense system. The Toll-like receptor (TLR) family functions as pattern recognition receptors, which recognize damage-associated molecular patterns such as viral dsRNA. Binding of viral dsRNA to specific TLR members triggers the expression of proinflammatory molecules involved in host anti-viral responses and subsequent activation of the adaptive immune defense system (Mogensen, 2009). The VAVC A46 protein inhibits the TLR4 signaling pathway (Lysakova-Devine et al., 2010). However, it is unknown whether the MPXV orthologue A47 has the same property, but the A47 protein has structural similarities to VACV protein A52R, which can inhibit the TLR3 and TLR4 signaling pathways (Bowie et al., 2000; Harte et al., 2003), underscoring a role for MPXV A47 in interfering with TLR signaling. MPXV produces low levels of dsRNA intermediates (Arndt et al., 2016), but whether these are recognized by TLR3 has not been investigated, although transcriptome analysis of MPXV-infected cells revealed repression of TLR3 target genes (Rubins et al., 2011). dsRNA can also activate kinase R (PKR), which mediates phosphorylation of eIF2α, resulting in the inhibition of viral and cellular mRNA translation (Pears, 1995). VACV E3 and K3 are inhibitors of PKR, allowing VACV to evade an antiviral response (Seet et al., 2003; Deng et al., 2008). The N-terminal domain of VACV E3 protein is absolutely required for the interaction with dsRNA (White and Jacobs, 2012). MPXV F3 protein is the VACV E3 protein homologue with a 37 aa truncation at the amino terminus (Arndt et al., 2015), suggesting that MPXV F3 does not bind dsRNA. However, MPXV can inhibit host immune responses, although a recombinant VACV expressing the MPXV F3L gene did not inhibit host PKR activation (Arndt et al., 2015), suggesting that MPXV has evolved to encode for yet undiscovered proteins that compensate for the missing N-terminal amino acids of F3 in limiting host antiviral activities. As previously mentioned, MPXV CCP (D14L gene) is completely absent in Clade II isolates, whereas the Clade I strains are predicted to express CCP, albeit with a truncated fourth short consensus repeat (Yeh and Contreras, 2022). Loss of expression of CCP/MOPICE limited the adaptive immune response against MPXV infection in rhesus macaques (Estep et al., 2011).
IFNs are main effectors of the innate immune response and can inhibit virus replication (Seet et al., 2003; Randall and Goodbourn, 2008). The MPXV B16 protein (VACV B19 orthologue) is a secreted type I IFN inhibitor and suppresses the antiviral type I IFN-induced signaling pathway (Fernández de Marco et al., 2010). VACV K7 abrogates IFN signaling by destabilizing IFN-regulated factor 3 (IRF3) and inhibits NFκB activation, whereas VACV H1 can block IFN signaling (Benfield et al., 2013; Liu and Moss, 2018). Whether the MPXV D9 and H1 orthologues have the same function remains to be proven, but both proteins are predicted to interact with several cellular proteins of the immune system (Mann et al., 2008; Kumar et al., 2023). VACV E3 perturbs IFN signaling by binding host Z-DNA binding protein 1 (ZBP1). The MPXV E3 orthologue (F3) perturbs the IFN pathway, but whether in a ZBP1-dependent manner needs to be established. The DNA-sensing receptor pathway cGAS/STING (cyclic GMP-AMP synthase/stimulator of interferon genes) can activate the IFN and NFκB pathways (Motwani et al., 2019). The VACV B16 and B8 proteins block IFN signaling by operating as soluble IFN-α and IFN-γ receptors, respectively (Alcamí and Smith, 1995; Colamonici et al., 1995). The MPXV ORF B16R and B9R encode functional homologues (Shchelkunov et al., 2002). OPXVs encode poxins that degrade 2′,3′ cGAMP and thereby inhibit cGAS/STING signaling (Eaglesham et al., 2019). Poxin is conserved in MPXV where it is fused to an additional C-terminal domain previously noted to have homology with human schlafen proteins (VACV B2R = MPXV B4R) (Eaglesham et al., 2019), hence MPXV may evade the immune system by targeting the cGAS/STING pathway.
MPXV can dodge the immune system by targeting the antiviral cytokine TNFα and other immunomodulating molecules. The MPXV-encoded cytokine response-modifying protein B (CrmB; J2L or OPG002) functions as a decoy receptor for TNFα (Gileva et al., 2006). The C-terminal domain of VACV CrmB can bind CCL28, CCL25, CXC motif chemokine ligand 12 (CXCL12), CXCL13 and CXCL14 (Alejo et al., 2006). It is unknown whether MPXV CrmB interacts with these chemokines, but the amino acid sequence of the corresponding domain in MPXV CrmB differs significantly (Gileva et al., 2006).
Natural killer (NK) cells and cytotoxic T cells (CTL) play a crucial role in eliminating viral infections. Although the number of NK cells expand significantly in peripheral blood and lymph nodes in MPXV-infected rhesus macaques, their migrating capacity was reduced, and several functions such as expression of chemokine receptors (including CCR5, CCR6 and CXCR3) and secretion of IFNγ and TNFα were impaired (Song et al., 2013). The importance of NK cells in controlling MPXV viral load was demonstrated in the highly vulnerable to OPXV infection CAST/EiJ mouse strain owing to low numbers of NK cells. IL-15 treatment, known to increase the numbers of IFNγ-secreting NK cells and CD8+ T cells, protected CAST/EiJ mice from lethal MPXV infection even when both CD4+ and CD8+ T cells were depleted. This implies that the expanded NK cells were responsible for the protective effect (Al-musa et al., 2022; Lum et al., 2022).
MPXV protein B10 avoids detection of virus-infected cells by CTL by impairing peptide loading and MHC-I trafficking within the endoplasmic reticulum. However, NK cells continually screen cells via NKG2D for the absence of MHC-I, thereby ensuring that the MHC system is not compromised (Lum et al., 2022). MPXV-infected cells with downregulated MHC-I expression overcomes detection by NK cells by secreting the OCMP protein that binds NKG2D and suppress the typical NKG2D-dependent NK cell lysis of cells that do not express MHC-I (Lum et al., 2022) (Figure 4C). IL-18, an IFNγ-inducing factor, stimulates the synthesis of various cytokines and chemokines, regulates Th1 and Th2 cell responses, and activates NK and CTL (Seet et al., 2003). OPXVs block IL-18 activities by producing an IL-18 binding protein (Born et al., 2000). The MPXV D6L gene encodes such IL-18 binding protein (Shchelkunov et al., 2002).
The MPXV J3R and A41L genes encode chemokine binding proteins (Shchelkunov et al., 2002), which are assumed to destroy the chemokine concentration gradient resulting in decreased neutrophil migration in tissues infected with MPXV and thus reducing viral virulence and inflammatory response (Bahar et al., 2008).
The complement system, which forms an essential part of the innate immune system is activated early in MPXV infection in mice, and is crucial for viral control (Moulton et al., 2008). MPXV MOPICE which from studies with VARV and VACV MOPICE was found to inhibit activation of the complement pathway (Liszewski et al., 2006). MOPICE modulates the antiviral immune response as observed by enhanced viral replication in vivo and dampened adaptive immune response in rhesus macaques infected with MPXV lacking MOPICE expression (Estep et al., 2011; Hudson et al., 2012). Because of the absence of the D14L gene in the Clade II, the virus and virus-infected cells would be predicted to be susceptible to the host complement attack. However, the expression of MOPICE in Clade II did not increase its virulence (Hudson et al., 2012), demonstrating that MOPICE is not the sole determinant of differences in viral virulence between the two clades.
Less is known how MPXV may interfere with the adaptive immune system. MPXV interferes with adaptive immune responses of antiviral CD8 + and CD4 + T cell responses via inhibiting T cell receptor-mediated T cell activation (Hammarlund et al., 2008).VACV A35 blocks immune priming of T lymphocytes by interfering with MHC class II-restricted antigen presentation. Moreover, infection studies in cells with MPXV lacking the A35R gene demonstrated that A35 inhibits production of cytokines and chemokines (Rehm et al., 2011). MPXV A35 analogue, A37, might suppress presentation of viral antigens to immune cells and help the virus to evade the host immune defense system. However, MPXV does not seem to interfere with MHC expression or intracellular transport of MHC molecules (Hammarlund et al., 2008).
Surveillance data on mpox in endemic countries during different periods between 1970-2015 showed that 71%-83% of the disease occurred in children (<10 years of age) and 51%-67% in males (Breman et al., 1980; Heymann et al., 1998; Rimoin et al., 2011). In contrast, the median age for the 2017-2018 outbreak in Nigeria was 29 years, with males accounting for 64% of the cases (Yinka-Ogunleye et al., 2019). However, for the 2022 multi-country outbreak, males accounted for 96.8% of the cases, and the median age was 34 years (Interquartile range: 29 – 41) (World Health Organization., 2022). In the African region, children (0-9 years of age) accounted for 23.08% of mpox cases compared to <1% in Europe and the Americas. Notably, the male-to-female ratio in Africa is also markedly lower than in other regions (Figures 5A-C) (World Health Organization, 2022).
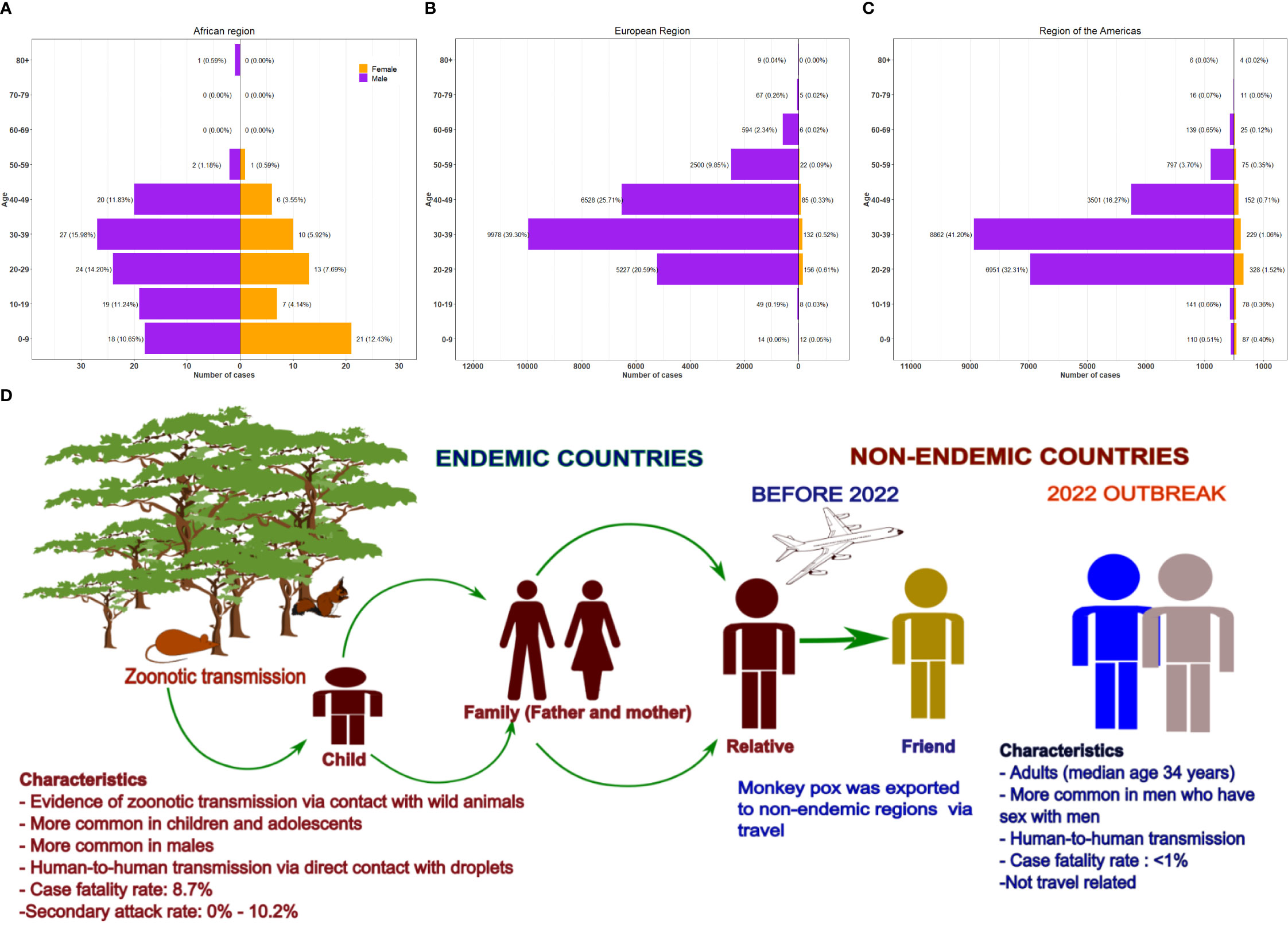
Figure 5 Demographic characteristics and transmission routes of human mpox. Demographic characteristics of 2022 human mpox cases according to sex and age in three WHO regions based on data from 4727328. (A) African region, (B) European region and (C) region of Americas. The pyramid plots show the number of cases and percentage of overall cases. (D)Transmission of mpox in endemic and non-endemic regions pre-2022 and 2022 outbreak. The characteristics of outbreaks in the different regions were highlighted. Prior to 2022, mpox was limited to the endemic regions and cases outside the region were usually travel related. However, the 2022 outbreak in non-endemic region was not travel related and has been reported in several countries.
Before the 2022 outbreak, the overall pooled estimated case fatality rate (CFR) was 8.7% and varied by the clade of the virus (Bunge et al., 2022). The pooled CFR for Clade II was 3.6% and 10.6% for Clade I (Bunge et al., 2022). The 2022 outbreak has an estimated CFR of 0.08% (World Health Organization, 2022). The low CFR in the current outbreak may be related to several factors, including the fact that the Clade II has a CFR of <1%, active surveillance, early diagnosis, and treatment (World Health Organization, 2022b).
Over the past five decades, the secondary attack rate for mpox has been stable and ranges from 0% to 10.2% (Breman et al., 1977; Merouze and Lesoin, 1983; Tchokoteu et al., 1991; McCollum et al., 2014; Besombes et al., 2019; Ye et al., 2019). Higher estimates were reported in the 2013 outbreak in the DRC (50% among 16 households) and Nigeria (71% in the 2017- 2018 outbreak from a single household) (Nolen et al., 2015; Beer and Bhargavi Rao, 2019; Bunge et al., 2022). However, it is imperative to note that it is unclear how the high estimates were obtained and thus may likely be overestimated (Beer and Bhargavi Rao, 2019).
Reproduction number (R0) is the number of secondary cases anticipated to develop from a single primary case in a naive population (Beer and Bhargavi Rao, 2019). While historical data on R0 is limited, published evidence from DRC’s active surveillance between 1980 and 1984 estimated R0 to be 0.8 (Beer and Bhargavi Rao, 2019), implying that transmission is ineffective as a human-to-human epidemic is likely to die out (Beer and Bhargavi Rao, 2019). However, early estimations of the R0 in the 2022 outbreak in non-endemic countries showed a high variability ranging from 1.54 (Belgium) to 3.62 (Germany), with a median of 2.44 (Branda et al., 2023). This suggests sustained human-to-human transmission, possibly due to several contributing factors, including different social and/or sexual behaviors, different MPXV variants, population density or other unknown causes (Branda et al., 2023). While the early estimations indicated sustained transmission, the outbreak has slowed down with case reductions. This could be due to behavior change, infection induced immunity, and vaccinations.
Transmission of MPXV can occur from animal-to-human (zoonotic) and from human-to-human (interhuman). Zoonotic transmission usually happens through contact with an infected animal’s bodily fluid or through a bite or scratch (Reynolds et al., 2007). Exposure to animal reservoirs, especially in regions with deforestation enhancing animal-human contact, and uncooked meat products are major risk factors for zoonotic transmission (Kipkorir et al., 2022). During the 2003 USA outbreak, exposure was classified as “non-invasive” (touching an infected animal) or “complex” (invasive bite from an ill animal; non-invasive exposure i.e., any exposure that did not break the skin) (Reynolds et al., 2018). Patients with complex exposures were more likely to develop systemic illness compared to those with non-invasive exposure (Reynolds et al., 2018). Dewitt et al. predicted the MPXV mode of transmission in most 2022 studies to be caused by inter-human transmission (Dewitt et al., 2022) (Figure 5D). Large respiratory droplets, bodily fluids, contaminated fomites, and viral shedding through feces are also considered risk factors for viral transmission (Rimoin et al., 2010; Sklenovská and Van Ranst, 2018; El Eid et al., 2022). Airborne transmission of MPXV between animals in experimental settings has also been reported and MPXV was detected in upper respiratory samples, suggesting that interhuman transmission of MPXV via the airborne route may be possible. However, epidemiological observations do not support airborne transmission as the primary route of transmission (Riopelle et al., 2022). The virus can also cross the placenta, suggesting vertical transmission (Mbala et al., 2017; Cuerel et al., 2022). At least 12 pregnant women have been infected during the 2022 outbreak, but vertical transmission was not observed in any case (Khalil et al., 2020).
MPXV has also been detected in human semen (Antinori et al., 2022; Noe et al., 2022), and in archival testes tissue of crab-eating macaque (Liu et al., 2022), suggesting potential sexual transmission of the virus. Recent outbreak suggests that MSM subpopulation may also be at an increased risk (Kipkorir et al., 2022).
In addition, waning immunity against smallpox has been considered another potential risk factor for the disease (Rimoin et al., 2010). Evidence from early outbreaks in the 1980s showed that previous smallpox vaccination provided 85% protection against mpox (Rimoin et al., 2010). Most cases reported were among those born after vaccination ceased in 1980, and herd immunity has significantly decreased (Rimoin et al., 2010).
Evidence from the literature suggests that those who are immunocompromised due to HIV and other underlying conditions are at increased risk of severe mpox disease (Center for Diseases Control and Prevention (CDC), 2022b) (Yinka-Ogunleye et al., 2019). A high proportion of patients with mpox in the 2022 outbreak had concurrent HIV infection and sexually transmitted infections (STI) (Fischer et al., 2022; Ghaffar and Shahnoor, 2022). Mpox patients with HIV infection were more likely to be hospitalized than those without HIV infection (Curran et al., 2022). However, the evidence on the reason for hospitalization is limited, and it is unknown if it reflects a more severe mpox illness (Curran et al., 2022).
Furthermore, co-infection with other conditions with rashes, such as a varicella-zoster virus (VZV), can occur (Hughes et al., 2021; Stephen et al., 2022). This herpesvirus causes chickenpox and shingles and is frequently misdiagnosed as mpox in regions where both diseases are endemic (Hoff et al., 2017; Hughes et al., 2021). Chickenpox is exclusive to humans and more common in younger age groups, and co-infection with mpox has been reported more commonly among children (Leung et al., 2019; Hughes et al., 2021).
Electron microscopy, immunohistochemical detection of MPXV proteins, and detectable levels of anti-OPXV IgM antibody during the period of 4 to 56 days after rash onset can be used to diagnose MPXV infection, but all methods are not specific (Center for Diseases Control and Prevention (CDC), 2022a). According to the Centers for Disease Control and Prevention (CDC) (Center for Diseases Control and Prevention (CDC), 2022a), mpox cases should be confirmed by real-time polymerase chain reaction (qPCR) or Next-Generation sequencing and isolation of MPXV in culture from a clinical specimen. The F3L, E9L, B6R and J2R genes are all target of qPCR in MPXV diagnosis (Li et al., 2010; Alakunle et al., 2020; Altindis et al., 2022; Cheema et al., 2022; Nakhaie et al., 2022). The suspected mpox cases are characterized by fulfilling one of the epidemiological criteria (within 21 days of illness onset), as outlined by CDC (Center for Diseases Control and Prevention (CDC), 2022a).
One of the crucial ways of controlling the spread of mpox is contact tracing (Harapan et al., 2022; Kalyar et al., 2022). Individuals exposed to MPXV should be monitored for 21 days checking mpox symptoms, and those with suspected or confirmed mpox cases should be isolated to avoid infecting others (Titanji et al., 2022). Velavan et al. predicted the mpox outbreak would not last provided that cases are well isolated through the contact tracing (Velavan and Meyer, 2022). Although much attention is given to human-human contact tracing, great efforts need to be put into animal-animal and animal-human contact tracing especially due to non-specificity of reservoir hosts for MPXV (Petersen et al., 2019a). Surveillance cannot be undermined in curtailing mpox as surveillance would provide more insight into the epidemiology of the disease (Riopelle et al., 2022). In Nigeria, the Outbreak Response Management and Analysis System (SORMAS) for mpox surveillance across portions of 8 states was implemented in November 2017 for the mpox outbreak. The use of the system increased the quantity of epidemiological data collected and the communication of aggregate case data (Mauldin et al., 2022).
Although there is no specific vaccine for MPXV (Hobson et al., 2021; Sah et al., 2022a), the smallpox vaccines have been reported to give 85% cross-immunity against MPXV due to shared antigenic features (Alakunle et al., 2020).
ACAM2000™ is a replication-competent vaccinia virus vaccine licensed by Food and Drug Administration (FDA) in August 2007 for smallpox prevention, and it is derived from a single clonal viral isolate from Dryvax (Gruber, 2022; Poland et al., 2022) which is a first-generation smallpox vaccine. As a second-generation attenuated vaccinia virus vaccine (Gessain et al., 2022), ACAM2000™ has been recommended as MPXV post-exposure prophylaxis (Aden et al., 2022; McCarthy, 2022). Although high level of protection against mpox in animal models has been recorded, the safety of ACAM2000™ in humans is still of great concern as cardiac complications, and extremely painful and uncomfortable cutaneous reaction at the injection site have been associated with the vaccine (Chandran et al., 2022). Therefore, the vaccine is no longer licensed by European Union (Luo and Han, 2022).
JYNNEOS (Imvamune or Imvanex) was approved by FDA in September 2019 for prevention of smallpox and mpox in adults aged >18years (Alakunle et al., 2020). JYNNEOS is the brand name of Modified Vaccinia virus Ankara Bavarian Nordic (MVA-BN) vaccine (Kmiec and Kirchhoff, 2022), a non-replicating third-generation attenuated vaccine. JYNNEOS is considered safer (with proven efficacy in animals and humans) than ACAM2000™ (Sah et al., 2022a). The Advisory Committee on Immunization Practices has recommended JYNNEOS as an alternative to ACAM2000™ (Harapan et al., 2022). Nonetheless, both vaccines (JYNNEOS and ACAM2000™) have been recommended for MPXV high-risk groups (Petersen et al., 2022).
LC16m8 is another potential vaccine for MPXV which is obtained by subjecting VACV lister to 36 serial passages at low temperature (30°C) in primary rabbit kidney cells (Domán et al., 2022; Poland et al., 2022; Schnierle, 2022). As a third-generation attenuated vaccine (Gessain et al., 2022), LC16m8 has been shown to be protective against MPXV in animal models with lower neurotoxicity (Domán et al., 2022; Schnierle, 2022). The frameshift mutation in LC16m8’s major extracellular enveloped virion antigen (B5R) contributes to the vaccine replication competence and low virulence (Domán et al., 2022). Presently, LC16m8 is only licensed in Japan (Domán et al., 2022; Schnierle, 2022).
Although there are no specific antivirals for mpox, some antivirals (tecovirimat, brincidofovir, cidofovir) have been explored (Riopelle et al., 2022). Tecovirimat (ST-246 or TPOXX®), 4-trifluoromethylphenol derivative, was approved (for smallpox) by FDA in 2018 and approved by European Medicines Agency in January 2022 for treatment of smallpox and cowpox. Tecovirimat inhibits VP37 (p37) protein of VACV by targeting the viral F13L gene (Frenois-Veyrat et al., 2022) and the CPXV homolog V016 gene (Siegrist and Sassine, 2022). VP37, a highly conserved protein in OPXV genus, is required for viral maturation and release from the infected cell. Inhibition of VP37 prevents viral spread within an infected animal models (Rizk et al., 2022). Goyal etal. recommended tecovirimat to be administered as first line of mpox treatment in pregnant and breastfeeding patients (Goyal et al., 2022). A tecovirimat analogue (synthesized by the State Research Center of Virology and Biotechnology, Russia) has been highlighted as a promising antivirals against OPXV infections (Singhal et al., 2022).
Cidofovir (CDV or Vistide®) prodrug is an acyclic nucleoside phosphate (Alakunle et al., 2020) that was approved by FDA in 1996 for the treatment of retinitis (caused by cytomegalovirus) in AIDS patients (Harapan et al., 2022). The efficacy of CDV has been identified during the in vitro studies in MPXV-infected animals, but the clinical data of CDV efficacy against mpox in human are not available. Brincidofovir (CMX001 or hexadecyloxypropyl-cidofovir), a CDV derivative, was approved for smallpox treatment in 2021 by FDA, and it has lesser toxic effects than CDV (Ortiz-Saavedra et al., 2022). Evaluation of CMX001 efficacy and safety in human mpox through the clinical trials is needed (Ortiz-Saavedra et al., 2022; Siegrist and Sassine, 2022).
Due to inadequate genome surveillance data particularly in endemic regions, little information about MPXV recombination is available (Yeh et al., 2022). However, there are some evidences of recombination between coinfecting or superinfecting OPXVs both in a laboratory setting and in nature (Estep et al., 2011; Goff et al., 2011; Brennan et al., 2022; Diaz-Cánova et al., 2022b, 2022a; Gigante et al., 2022).
MPXV circulating in non-endemic regions where other OPXVs are endemic, for instance, CPXV in Europe and VACV-like in south America, and vaccination against mpox with JYNNEOUS or ACAM 2000 are scenarios for coinfection and superinfection between different species of OPXV. Thus, there remains a potential risk of recombination between MPXV and other OPXVs that may result in MPXVs with mosaic genomes and altered biological characteristics.
Although there is insufficient evidence of MPXV being used for bioterrorism at the moment (Hosseini-Shokouh et al., 2022), many scientists have expressed concerns over its potential use for bioterrorism because there is a report that there was an attempt by the former Soviet Union to use MPXV as a bioweapon (Nalca et al., 2005; Duraffour et al., 2007; Kindrachuk et al., 2012; Hosseini-Shokouh et al., 2022; Makkar, 2022). The possibility of MPXV as a potential bioweapon due to its global reemergence and its clinical similarities with VARV has placed the virus on the global public health agenda (Ellis et al., 2012; Ihekweazu et al., 2020). For example, the USA has made preparation for the possibility of smallpox virus as a potential bioterror biological by storing smallpox vaccines and antivirals after the 9/11 attack (Franz, 2004; Xiang and White, 2022). Smallpox virus is in category A of the CDC list of bioterrorism agents (Hosseini-Shokouh et al., 2022). The risk of bioterrorism is heightened knowing that MPXV genomes can be synthesized from publicly available sequence data and life viruses (with or without further genetic modification) can be re-constituted. This has already been demonstrated with horsepox virus (HPXV), which shares the same genus as MPXV (Noyce et al., 2018). Hence, a strict dual-use policy must be agreed upon and implemented in all laboratories across the globe.
Sexual orientation (especially LGBQTI+ community) (Sousa et al., 2022) and racial (Happi et al., 2022) are two stigmas associated with MPXV. The persistent narrative of the media alongside many scientists linking the mpox 2022 outbreak to Africa/West Africa/Nigeria is worrisome (Sousa et al., 2022). Despite the mpox 2022 outbreak, which occurred outside Africa, the nomenclature and the geographical labels of MPXV strains still reference West African, even though the origin of this outbreak is still unresolved. Furthermore, high mpox cases were reported among MSM (Happi et al., 2022; Malta et al., 2022; Sousa et al., 2022; Sah et al., 2022b). This narrative portrays these men contracting MPXV because they engaged in sexual intercourse with fellow men, meanwhile the spread of the virus can occur regardless the sex. Vaccine inequality affects lower- and middle-income countries (LMICs). An unknown number of mpox cases in LMICs are not captured due to a shortage of resources like limited testing and surveillance capacity (Malta et al., 2022). As highlighted by Malta et al., the MPXV vaccines are presently accessible only in high-income countries (Canada, the USA, and the UK) (Malta et al., 2022).
MPXV has emerged and re-emerged for over five decades and yet not much is known about its virological profile and the characteristics of the disease it causes. In particular, the reservoir host of MPXV remains unknown, the viral, host and environmental factors that modulate the virus maintenance in the wild, animal-to-animal transmission, zoonotic transmission and reverse spillover are still a mystery. Neither are we closer to reliable prognostication of virus emergence and accurate modelling of the disease outcomes. Current and future studies should prioritize understanding the molecular basis of MPXV infection to develop effective drugs and vaccines against mpox as well as functional mutational studies that will shed insight into the dynamics of MPXV transmission across hosts. To improve tracking of MPXV, laboratories particularly in resource-poor countries should be equipped with genome-based surveillance capacity and capability. Lastly, MPXV and mpox affect human, animal, and ecosystem health. Thus, a One Health strategy is indispensable to the prevention and treatment of current and future outbreaks.
MO: Conceptualization, Data curation, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing. EA: Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. DK: Formal analysis, Methodology, Writing – original draft. DD: Data curation, Formal analysis, Investigation, Writing – original draft. FA: Investigation, Writing – original draft. OA: Formal analysis, Investigation, Writing – original draft. UM: Conceptualization, Investigation, Methodology, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The APC was covered by funds from The University of Tromsø—The Arctic University of Norway.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Adegboye, O. A., Castellanos, M. E., Alele, F. O., Pak, A., Ezechukwu, H. C., Hou, K., et al. (2022). Travel-related monkeypox outbreaks in the era of COVID-19 pandemic: are we prepared? Viruses 14, 1–7. doi: 10.3390/v14061283
Aden, T.A., Blevins, P., York, S.W., Rager, S., Balachandran, D., Huston, C.L., et al (2022). Rapid diagnostic testing for response to the monkeypox outbreak — Laboratory response network, United States, may 17–june 30, 2022. MMWR Morb. Mortal Wkly. Rep. 71, 904–907. doi: 10.15585/mmwr.mm7128e1
Agrati, C., Cossarizza, A., Mazzotta, V., Grassi, G., Casetti, R., Biasi, S., et al. (2022). Immunological signature in human cases of monkeypox infection in 2022 outbreak: an observational study. Lancet Infect. Dis. 23, 320–330. doi: 0.1016/S1473-3099(22)00662-4
Alakunle, E., Moens, U., Nchinda, G., Okeke, M. I. (2020). Monkeypox virus in Nigeria: infection biology, epidemiology, and evolution. Viruses 12, 1–29. doi: 10.3390/v12111257
Alakunle, E. F., Okeke, M. I. (2022). Monkeypox virus : a neglected zoonotic pathogen spreads globally. Nat. Rev. Microbiol. 20, 507–508. doi: 10.1038/s41579-022-00776-z
Alcami, A., Smith, G. L. (1992). A soluble receptor for interleukin-1β encoded by vaccinia virus: A novel mechanism of virus modulation of the host response to infection. Cell 71, 153–167. doi: 10.1016/0092-8674(92)90274-G
Alcamí, A., Smith, G. L. (1995). Vaccinia, cowpox, and camelpox viruses encode soluble gamma interferon receptors with novel broad species specificity. J. Virol. 69, 4633–4639. doi: 10.1128/jvi.69.8.4633-4639.1995
Alejo, A., Ruiz-Argüello, M. B., Ho, Y., Smith, V. P., Saraiva, M., Alcami, A. (2006). A chemokine-binding domain in the tumor necrosis factor receptor from variola (smallpox) virus. Proc. Natl. Acad. Sci. U. S. A. 103, 5995–6000. doi: 10.1073/pnas.0510462103
Alkhalil, A., Hammamieh, R., Hardick, J., Ichou, M. A., Jett, M., Ibrahim, S. (2010). Gene expression profiling of monkeypox virus-infected cells reveals novel interfaces for host-virus interactions. Virology 7, 1–19. doi: 10.1186/1743-422X-7-173
Al-musa, A., Chou, J., Labere, B. (2022). The resurgence of a neglected orthopoxvirus: Immunologic and clinical aspects of monkeypox virus infections over the past six decades. Clin. Immunol. 243, 109108. doi: 10.1016/j.clim.2022.109108
Altindis, M., Puca, E., Shapo, L. (2022). Diagnosis of monkeypox virus – An overview. Travel Med. Infect. Dis. 50, 102459. doi: 10.1016/j.tmaid.2022.102459
Americo, J. L., Earl, P. L., Moss, B. (2023). Virulence differences of mpox (monkeypox) virus clades I, IIa, and IIb.1 in a small animal model. Proc. Natl. Acad. Sci. U. S. A. 120, 4–9. doi: 10.1073/pnas.2220415120
Anderson, M. G., Frenkel, L. D., Homann, S., Guffey, J. (2003). A case of severe monkeypox virus disease in an American child: Emerging infections and changing professional values. Pediatr. Infect. Dis. J. 22, 1093–1096. doi: 10.1097/01.inf.0000101821.61387.a5
Antinori, A., Mazzotta, V., Vita, S., Carletti, F., Tacconi, D., Lapini, L. E., et al. (2022). Epidemiological, clinical and virological characteristics of four cases of monkeypox support transmission through sexual contact, Italy, May 2022. Eurosurveillance 27, 1–6. doi: 10.2807/1560-7917.ES.2022.27.22.2200421
Aoyagi, M., Zhai, D., Jin, C., Aleshin, A. E., Stec, B., Reed, J. C., et al. (2006). Vaccinia virus N1L protein resembles a B cell lymphoma-2 (Bcl-2) family protein. Protein Sci. 16, 118–124. doi: 10.1110/ps.062454707
Arita, I., Henderson, D. A. (1968). Smallpox and monkeypox in non-human primates. Bull. World Health Organ. 39, 277–283.
Arndt, W. D., Cotsmire, S., Trainor, K., Harrington, H., Hauns, K., Kibler, K. V., et al. (2015). Evasion of the innate immune type I interferon system by monkeypox virus. J. Virol. 89, 10489–10499. doi: 10.1128/JVI.00304-15
Arndt, W. D., White, S. D., Johnson, B. P., Huynh, T., Liao, J., Harrington, H., et al. (2016). Monkeypox virus induces the synthesis of less dsRNA than vaccinia virus, and is more resistant to the anti-poxvirus drug, IBT, than vaccinia virus. Virology 497, 125–135. doi: 10.1016/j.virol.2016.07.016
Arthur, R., Upadhayay, S., Arthur, R., Soni, D., Yadav, P., Navik, U. (2022). Monkeypox infection : The past , present , and future. Int. Immunopharmacol. 113, 109382. doi: 10.1016/j.intimp.2022.109382
Bahar, M. W., Kenyon, J. C., Putz, M. M., Abrescia, N. G. A., Pease, J. E., Wise, E. L., et al. (2008). Structure and function of A41, a vaccinia virus chemokine binding protein. PLoS Pathog. 4, 0055–0068. doi: 10.1371/journal.ppat.0040005
Barry, M., Hnatiuk, S., Mossman, K., Lee, S. F., Boshkov, L., McFadden, G. (1997). The myxoma virus M-T4 gene encodes a novel RDEL-containing protein that is retained within the endoplasmic reticulum and is important for the productive infection of lymphocytes. Virology 239, 360–377. doi: 10.1006/viro.1997.8894
Bass, J., Tack, D. M., McCollum, A. M., Kabamba, J., Pakuta, E., Malekani, J., et al. (2013). Enhancing health care worker ability to detect and care for patients with monkeypox in the democratic republic of the Congo. Int. Health 5, 237–243. doi: 10.1093/inthealth/iht029
Beer, E. M., Bhargavi Rao, V. (2019). A systematic review of the epidemiology of human monkeypox outbreaks and implications for outbreak strategy. PloS Negl. Trop. Dis. 13, 1–20. doi: 10.1371/journal.pntd.0007791
Beerli, C., Yakimovich, A., Kilcher, S., Reynoso, G. V., Fläschner, G., Müller, D. J., et al. (2019). Vaccinia virus hijacks EGFR signalling to enhance virus spread through rapid and directed infected cell motility. Nat. Microbiol. 4, 216–225. doi: 10.1038/s41564-018-0288-2
Benfield, C. T. O., Ren, H., Lucas, S. J., Bahsoun, B., Smith, G. L. (2013). Vaccinia virus protein K7 is a virulence factor that alters the acute immune response to infection. J. Gen. Virol. 94, 1647–1657. doi: 10.1099/vir.0.052670-0
Besombes, C., Gonofio, E., Konamna, X., Selekon, B., Gessain, A., Berthet, N., et al. (2019). Intrafamily transmission of monkeypox virus, Central African Republi. Emerg. Infect. Dis. 25, 1602–1604. doi: 10.3201/eid2508.190112
Blasco, R., Moss, B. (1992). Role of cell-associated enveloped vaccinia virus in cell-to-cell spread. Am. Soc Microbiol. 66, 4170–4179. doi: 10.1128/jvi.66.7.4170-4179.1992
Born, T. L., Morrison, L. A., Esteban, D. J., VandenBos, T., Thebeau, L. G., Chen, N., et al. (2000). A poxvirus protein that binds to and inactivates IL-18, and inhibits NK cell response. J. Immunol. 164, 3246–3254. doi: 10.4049/jimmunol.164.6.3246
Bowie, A., Kiss-Toth, E., Symons, J. A., Smith, G. L., Dower, S. K., O’Neill, L. A. J. (2000). A46R and A52R from vaccinia virus are antagonists of host IL-1 and toll-like receptor signaling. Proc. Natl. Acad. Sci. U.S.A. 97, 10162–10167. doi: 10.1073/pnas.160027697
Bragazzi, N. L., Kong, J. D., Mahroum, N., Tsigalou, C., Khamisy-Farah, R., Converti, M., et al. (2023). Epidemiological trends and clinical features of the ongoing monkeypox epidemic: A preliminary pooled data analysis and literature review. J. Med. Virol. 95, e27931. doi: 10.1002/jmv.27931
Branda, F., Pierini, M., Mazzoli, S. (2023). Monkeypox: Early estimation of basic reproduction number R0 in Europe. J. Med. Virol. 95, e28270. doi: 10.1002/jmv.28270
Breman, J. G., Nakano, J. H., Coffi, E., Godfrey, H., Gautun, J. C. (1977). Human poxvirus disease after smallpox eradication. Am. J. Trop. Med. Hyg. 26, 273–281. doi: 10.4269/ajtmh.1977.26.273
Breman, J. G., Ruti, K., Steniowski, M. V. (1980). Human monkeypox 1970-79. Bull. World Health Organ. 58, 165–182.
Brennan, G., Stoian, A. M. M., Yu, H., Rahman, M. J., Banerjee, S., Stroup, J. N., et al. (2022). Molecular mechanisms of poxvirus evolution. Am. Soc Microbiol. 14, e0152622. doi: 10.1128/mbio.01526-22
Brown, K., Leggat, P. A. (2016). Human monkeypox: Current state of knowledge and implications for the future. Trop. Med. Infect. Dis. 1, 1–13. doi: 10.3390/tropicalmed1010008
Buller, R. M. L., Chakrabarti, S., Moss, B., Fredricksont, T. (1988). Cell proliferative response to vaccinia virus is mediated by VGF. Virology 164, 182–192. doi: 10.1016/0042-6822(88)90635-6
Bunge, E. M., Hoet, B., Chen, L., Lienert, F., Weidenthaler, H., Baer, L. R., et al. (2022). The changing epidemiology of human monkeypox—A potential threat? A systematic review. PloS Negl. Trop. Dis. 16, 1–20. doi: 10.1371/journal.pntd.0010141
Campbell, J. A., Trossman, D. S., Yokoyama, W. M., Carayannopoulos, L. N. (2007). Zoonotic orthopoxviruses encode a high-affinity antagonist of NKG2D. J. Exp. Med. 204, 1311–1317. doi: 10.1084/jem.20062026
Center for Diseases Control and Prevention (CDC). (2022a). Case Definitions† for Use in the 2022 Mpox Response | Mpox | Poxvirus | CDC. (United States of America: Center for Diseases Control and Prevention (CDC)). Available at: https://www.cdc.gov/poxvirus/monkeypox/clinicians/case-definition.html (Accessed February 4, 2023).
Center for Diseases Control and Prevention (CDC). (2022b). Severe Manifestations of Monkeypox among People who are Immunocompromised Due to HIV or Other Conditions.
Center for Diseases Control and Prevention (CDC). (2023). Past U.S. Cases and Outbreaks | Mpox | Poxvirus | CDC. (United States of America). Available at: https://www.cdc.gov/poxvirus/monkeypox/outbreak/us-outbreaks.html (Accessed February 2, 2023).
Centre for Disease Control. (2003). Multistate Outbreak of Monkeypox— Illinois, Indiana, and Wisconsi (United States of America: American Medical Association (AMA)). doi: 10.1001/jama.290.1.30
Chandran, D., Dhama, K., K, M. A. M., Chakraborty, S., Mohapatra, R. K., Yatoo, M. I., et al. (2022). Monkeypox : an update on current knowledge and research advances. J. Exp. Biol. Agric. Sci. 10, 679–688. doi: 10.18006/2022.10(4)
Cheema, A. Y., Ogedegbe, O. J., Munir, M., Alugba, G., Ojo, T. K. (2022). Monkeypox : A review of clinical features , diagnosis , and treatment. Cureus 14, 14–17. doi: 10.7759/cureus.26756
Chen, N., Li, G., Liszewski, M. K., Atkinson, J. P., Jahrling, P. B., Feng, Z., et al. (2005). Virulence differences between monkeypox virus isolates from West Africa and the Congo basin. Virology 340, 46–63. doi: 10.1016/j.virol.2005.05.030
Chen, R. A. J., Ryzhakov, G., Cooray, S., Randow, F., Smith, G. L. (2008). Inhibition of IκB kinase by vaccinia virus virulence factor B14. PLoS Pathog. 4, e22. doi: 10.1371/journal.ppat.0040022
Colamonici, O. R., Domanski, P., Sweitzer, S. M., Larner, A., Buller, R. M. L. (1995). Vaccinia virus B18R gene encodes a type I interferon-binding protein that blocks interferon α transmembrane signaling. J. Biol. Chem. 270, 15974–15978. doi: 10.1074/jbc.270.27.15974
Cooray, S., Bahar, M. W., Abrescia, N. G. A., McVey, C. E., Bartlett, N. W., Chen, R. A. J., et al. (2007). Functional and structural studies of the vaccinia virus virulence factor N1 reveal a Bcl-2-like anti-apoptotic protein. J. Gen. Virol. 88, 1656–1666. doi: 10.1099/vir.0.82772-0
Croft, D. R., Sotir, M. J., Williams, C. J., Kazmierczak, J. J., Wegner, M. V., Rausch, D., et al. (2007). Occupational risks during a monkeypox outbreak, Wisconsi. Emerg. Infect. Dis. 13, 1150–1157. doi: 10.3201/eid1308.061365
Cuerel, A., Favre, G., Vouga, M., Pomar, L. (2022). Monkeypox and pregnancy : latest updates. Viruses 14, 1–12. doi: 10.3390/v14112520
Curran, K. G., Eberly, K., Russell, O. O., Snyder, R. E., Phillips, E. K., Tang, E. C., et al. (2022). HIV and sexually transmitted infections among persons with monkeypox — Eight U.S. Jurisdictions, may 17–july 22, 2022. MMWR. Morb. Mortal. Wkly. Rep. 71, 1141–1147. doi: 10.15585/mmwr.mm7136a1
Damon, I. K. (2011). Status of human monkeypox: Clinical disease, epidemiology and research. Vaccine 29, D54–D59. doi: 10.1016/j.vaccine.2011.04.014
DeHaven, B. C., Girgis, N. M., Xiao, Y., Hudson, P. N., Olson, V. A., Damon, I. K., et al. (2010). Poxvirus complement control proteins are expressed on the cell surface through an intermolecular disulfide bridge with the viral A56 protein. J. Virol. 84, 11245–11254. doi: 10.1128/JVI.00372-10
Deng, L., Dai, P., Parikh, T., Cao, H., Bhoj, V., Sun, Q., et al. (2008). Vaccinia virus subverts a mitochondrial antiviral signaling protein-dependent innate immune response in keratinocytes through its double-stranded RNA binding protein, E3. J. Virol. 82, 10735–10746. doi: 10.1128/JVI.01305-08
Denhardt, D. T. (1996). Signal-transducing protein phosphorylation cascades mediated by Ras/Rho proteins in the mammalian cell: the potential for multiplex signalling. Biochem. J. 318, 729–747. doi: 10.1042/bj3180729
Dewitt, M. E., Polk, C., Williamson, J., Shetty, A. K., Passaretti, C. L., Mcneil, C. J. (2022). Global monkeypox case hospitalisation rates : A rapid systematic review and meta-analysis. eClinicalMedicine 54, 101710. doi: 10.1016/j.eclinm.2022.101710
Diaz-Cánova, D., Mavian, C., Brinkmann, A., Nitsche, A., Moens, U., Okeke, M. I. (2022a). Genomic sequencing and phylogenomics of cowpox virus. Viruses 14, 1–16. doi: 10.3390/v14102134
Diaz-Cánova, D., Moens, U. L., Brinkmann, A., Nitsche, A., Okeke, M. I. (2022b). Genomic sequencing and analysis of a novel human cowpox virus with mosaic sequences from North America and old world orthopoxvirus. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.868887
Dinarello, C. A. (2018). Overview of the IL-1 family in innate inflammation and acquired immunity. Immunol. Rev. 281, 8–27. doi: 10.1111/imr.12621
Domán, M., Fehér, E., Varga-Kugler, R., Jakab, F., Bányai, K. (2022). Animal models used in monkeypox research. Microorganisms 10 2192. doi: 10.3390/microorganisms10112192
Dumonteil, E., Herrera, C., Sabino-Santos, G. (2023). Monkeypox virus evolution before 2022 outbreak. Emerg. Infect. Dis. 29, 451–453. doi: 10.3201/eid2902.220962
Duraffour, S., Ishchenko, A. A., Saparbaev, M., Crance, J. M., Garin, D. (2007). Substrate specificity of homogeneous monkeypox virus uracil-DNA glycosylase. Biochemistry 46, 11874–11881. doi: 10.1021/bi700726a
Durski, K. N., McCollum, A. M., Nakazawa, Y., Petersen, B. W., Reynolds, M. G., Briand, S., et al. (2018). Emergence of monkeypox — West and Central Africa 1970–2017. Morb. Mortal. Wkly. Rep. 67, 306–310. doi: 10.15585/mmwr.mm6710a5
Eaglesham, J. B., Pan, Y., Kupper, T. S., Kranzusch, P. J. (2019). Viral and metazoan poxins are cGAMP-specific nucleases that restrict cGAS–STING signalling. Nature 566, 259–263. doi: 10.1038/s41586-019-0928-6
Earl, P. L., Americo, J. L., Moss, B. (2012). Lethal monkeypox virus infection of CAST/eiJ mice is associated with a deficient gamma interferon response. J. Virol. 86, 9105–9112. doi: 10.1128/JVI.00162-12
El Eid, R., Allaw, F., Haddad, S. F., Kanj, S. S. (2022). Human monkeypox: A review of the literature. PloS Pathog. 18, e1010768. doi: 10.1371/journal.ppat.1010768
Ellis, C. K., Carroll, D. S., Lash, R. R., Townsend Peterson, A., Damon, I. K., Malekani, J., et al. (2012). Ecology and geography of human monkeypox case occurrences across Africa. J. Wildl. Dis. 48, 335–347. doi: 10.7589/0090-3558-48.2.335
Ember, S. W. J., Ren, H., Ferguson, B. J., Smith, G. L. (2012). Vaccinia virus protein C4 inhibits NF-κB activation and promotes virus virulence. J. Gen. Virol. 93, 2098–2108. doi: 10.1099/vir.0.045070-0
Erez, N., Achdout, H., Milrot, E., Schwartz, Y., Wiener-Well, Y., Paran, N., et al. (2019). Diagnosis of imported monkeypox, Israe. Emerg. Infect. Dis. 25, 980–983. doi: 10.3201/eid2505.190076
Estep, R. D., Messaoudi, I., O’Connor, M. A., Li, H., Sprague, J., Barron, A., et al. (2011). Deletion of the monkeypox virus inhibitor of complement enzymes locus impacts the adaptive immune response to monkeypox virus in a nonhuman primate model of infection. J. Virol. 85, 9527–9542. doi: 10.1128/JVI.00199-11
Falendysz, E. A., Lopera, J. G., Rocke, T. E., Osorio, J. E. (2023). Monkeypox virus in animals: current knowledge of viral transmission and pathogenesis in wild animal reservoirs and captive animal models. Viruses 15, 905. doi: 10.3390/v15040905
Fang, E. S. Y., Ng, O. T., Marc, J. Z. H., Mak, T. M., Marimuthu, K., Vasoo, S., et al. (2020). Imported monkeypox, Singapore. Emerg. Infect. Dis. 26, 1826–1830. doi: 10.3201/eid2608.191387
Faye, O., Pratt, C. B., Faye, M., Fall, G., Chitty, J. A., Diagne, M. M., et al. (2018). Genomic characterisation of human monkeypox virus in Nigeria. Lancet Infect. Dis. 18, 246. doi: 10.1016/S1473-3099(18)30043-4
Fernández de Marco, M., del, M., Alejo, A., Hudson, P., Damon, I. K., Alcami, A. (2010). The highly virulent variola and monkeypox viruses express secreted inhibitors of type I interferon. FASEB J. 24, 1479–1488. doi: 10.1096/fj.09-144733
Firth, C., Kitchen, A., Shapiro, B., Suchard, M. A., Holmes, E. C., Rambaut, A. (2010). Using time-structured data to estimate evolutionary rates of double-stranded DNA viruses. Mol. Biol. Evol. 27, 2038–2051. doi: 10.1093/molbev/msq088
Fischer, F., Mehrl, A., Kandulski, M., Schlosser, S., Müller, M., Schmid, S. (2022). Monkeypox in a patient with controlled HIV infection initially presenting with fever , painful pharyngitis , and tonsillitis. Medicina (B. Aires). 58, 1–6. doi: 10.3390/medicina58101409
Forni, D., Cagliani, R., Molteni, C., Clerici, M., Sironi, M. (2022a). Monkeypox virus: The changing facets of a zoonotic pathogen. Infect. Genet. Evol. 105, 105372. doi: 10.1016/j.meegid.2022.105372
Forni, D., Molteni, C., Cagliani, R., Sironi, M. (2022b). Geographic structuring and divergence time frame of monkeypox virus in the endemic region. J. Infect. Dis. 1, 1–10. doi: 10.1093/infdis/jiac298
Franz, M. (2004). Development and Characterization of Monoclonal Antibodies Directed Against Tanapox Virus. (Kalamazoo, Michigan: Western Michigan University).
Frenois-Veyrat, G., Gallardo, F., Gorgé, O., Marcheteau, E., Ferraris, O., Baidaliuk, A., et al. (2022). Tecovirimat is effective against human monkeypox virus in vitro at nanomolar concentrations. Nat. Microbiol. 7, 1951–1955. doi: 10.1038/s41564-022-01269-8
Gessain, A., Nakoune, E., Yazdanpanah, Y. (2022). Monkeypox. N. Engl. J. Med. 19, 1–11. doi: 10.1056/NEJMra2208860
Ghaffar, R. A., Shahnoor, S. (2022). Increased prevalence of HIV among Monkeypox patients – An alarming update. New Microbes New Infect. 49–50, 101039. doi: 10.1016/j.nmni.2022.101039
Gigante, C. M., Korber, B., Seabolt, M. H., Wilkins, K., Davidson, W., Rao, A. K., et al. (2022). Multiple lineages of monkeypox virus detected in the United States 2021–2022. Science 378, 560–565. doi: 10.1126/science.add4153
Gileva, I. P., Nepomnyashchikh, T. S., Antonets, D. V., Lebedev, L. R., Kochneva, G. V., Grazhdantseva, A. V., et al. (2006). Properties of the recombinant TNF-binding proteins from variola, monkeypox, and cowpox viruses are different. Biochim. Biophys. Acta - Proteins Proteomics 1764, 1710–1718. doi: 10.1016/j.bbapap.2006.09.006
Girgis, N. M., DeHaven, B. C., Xiao, Y., Alexander, E., Viner, K. M., Isaacs, S. N. (2011). The vaccinia virus complement control protein modulates adaptive immune responses during infection. J. Virol. 85, 2547–2556. doi: 10.1128/JVI.01474-10
Giulio, D.B.D., Eckburg, P. B. (2004). Human monkeypox: an emerging zoonosis. Lancet Infect. Dis. 4, 15–25. doi: 10.1016/s1473-3099(03)00856-9
Goff, A., Mucker, E., Raymond, J., Fisher, R., Bray, M., Hensley, L., et al. (2011). Infection of cynomolgus macaques with a recombinant monkeypox virus encoding green fluorescent protein. Arch. Virol. 156, 1877–1881. doi: 10.1007/s00705-011-1065-1
Goyal, L., Ajmera, K., Pandit, R., Pandit, T. (2022). Prevention and treatment of monkeypox: A step-by-step guide for healthcare professionals and general population. Cureus 14, 1–13. doi: 10.7759/cureus.28230
Greber, U. F. (2002). Signalling in viral entry. Cell. Mol. Life Sci. 58, I. doi: 10.1007/s00018-002-8453-3
Grothe, J. H., Cornely, O. A., Salmanton-García, J. (2022). Monkeypox diagnostic and treatment capacity at epidemic onset: A VACCELERATE online survey. J. Infect. Public Health 15, 1043–1046. doi: 10.1016/j.jiph.2022.08.008
Gruber, M. F. (2022). Current status of monkeypox vaccines. NPJ Vaccines 7, 1–3. doi: 10.1038/s41541-022-00527-4
Hammarlund, E., Dasgupta, A., Pinilla, C., Norori, P., Früh, K., Slifka, M. K. (2008). Monkeypox virus evades antiviral CD4+ and CD8+ T cell responses by suppressing cognate T cell activation. Proc. Natl. Acad. Sci. U.S.A. 105, 14567–14572. doi: 10.1073/pnas.0800589105
Happi, C., Adetifa, I., Mbala, P., Njouom, R., Nakoune, E., Happi, A., et al. (2022). Urgent need for a non-discriminatory and non-stigmatizing nomenclature for monkeypox virus. PLoS Biol. 20, 1–6. doi: 10.1371/journal.pbio.3001769
Harapan, H., Ophinni, Y., Megawati, D., Frediansyah, A., Mamada, S. S. (2022). Monkeypox : A comprehensive review. Viruses 14, 1–26. doi: 10.3390/v14102155
Harris, R. S., Dudley, J. P. (2015). APOBECs and virus restriction. Virology 479–480, 131–145. doi: 10.1016/j.virol.2015.03.012
Harrison, D.A (1990). The JAK/STAT pathway: fact sheet. Cold Spring Harb. Perspect. Biol. 4, a011205.
Harte, M. T., Haga, I. R., Maloney, G., Gray, P., Reading, P. C., Bartlett, N. W., et al. (2003). The poxvirus protein A52R targets toll-like receptor signaling complexes to suppress host defense. J. Exp. Med. 197, 343–351. doi: 10.1084/jem.20021652
Hayden, M. S., Ghosh, S. (2008). Shared principles in NF-κB signaling. Cell 132, 344–362. doi: 10.1016/j.cell.2008.01.020
Hendrickson, R. C., Wang, C., Hatcher, E. L., Lefkowitz, E. J. (2010). Orthopoxvirus genome evolution: The role of gene loss. Viruses 2, 1933–1967. doi: 10.3390/v2091933
Herbert, M. H., Squire, C. J., Mercer, A. A. (2015). Poxviral ankyrin proteins. Viruses 7, 709–738. doi: 10.3390/v7020709
Heymann, D. L., Szczeniowski, M., Esteves, K. (1998). Re-emergence of monkeypox in Africa: A review of the past six years. Br. Med. Bull. 54, 693–702. doi: 10.1093/oxfordjournals.bmb.a011720
Hnatiuk, S., Barry, M., Zeng, W., Liu, L., Lucas, A., Percy, D., et al. (1999). Role of the C-terminal RDEL motif of the myxoma virus M-T4 protein in terms of apoptosis regulation and viral pathogenesis. Virology 263, 290–306. doi: 10.1006/viro.1999.9946
Hobson, G., Adamson, J., Adler, H., Firth, R., Gould, S., Houlihan, C., et al. (2021). Family cluster of three cases of monkeypox imported from Nigeria to the United Kingdom, May 2021. Eurosurveillance 26, 1–4. doi: 10.2807/1560-7917.ES.2021.26.32.2100745
Hoff, N. A., Morier, D. S., Kisalu, N. K., Johnston, S. C., Doshi, R. H., Hensley, L. E., et al. (2017). Varicella coinfection in patients with active monkeypox in the democratic republic of the congo. Ecohealth 14, 564–574. doi: 10.1007/s10393-017-1266-5
Hosseini-Shokouh, S.-J., Gholami, M., Barati, M., Naghoosi, H. (2022). Monkeypox : A re-emerging viral zoonotic infection. Ann. Mil. Heal. Sci. Res. 20, e129475. doi: 10.5812/amh
Hudson, P. N., Self, J., Weiss, S., Braden, Z., Xiao, Y., Girgis, N. M., et al. (2012). Elucidating the role of the complement control protein in monkeypox pathogenicity. PloS One 7, e35086. doi: 10.1371/journal.pone.0035086
Hughes, C. M., Liu, L., Davidson, W. B., Radford, K. W., Wilkins, K., Monroe, B., et al. (2021). A tale of two viruses : coinfections of monkeypox and varicella zoster virus in the Democratic Republic of Congo. Am. J. Trop. Med. Hyg. 104, 604–611. doi: 10.4269/ajtmh.20-0589
Hutson, C. L., Carroll, D. S., Self, J., Weiss, S., Hughes, C. M., Braden, Z., et al. (2010). Dosage comparison of Congo Basin and West African strains of monkeypox virus using a prairie dog animal model of systemic orthopoxvirus disease. Virology 402, 72–82. doi: 10.1016/j.virol.2010.03.012
Hutson, C. L., Olson, V. A., Carroll, D. D., Abel, J. A., Hughes, C. M., Braden, Z. H., et al. (2009). A prairie dog animal model of systemic orthopoxvirus disease using west African and Congo Basin strains of Monkeypox virus. J. Gen. Virol. 90, 323–333. doi: 10.1099/vir.0.005108-0
Ihekweazu, C., Yinka-Ogunleye, A., Lule, S., Ibrahim, A. (2020). Importance of epidemiological research of monkeypox: is incidence increasing? Expert Rev. Anti Infect. Ther. 18, 389–392. doi: 10.1080/14787210.2020.1735361
Isaacs, S. N., Kotwal, G. J., Moss, B. (1992). Vaccinia virus complement-control protein prevents antibody-dependent complement-enhanced neutralization of infectivity and contributes to virulence. Proc. Natl. Acad. Sci. U.S.A. 89, 628–632. doi: 10.1073/pnas.89.2.628
Isidro, J., Borges, V., Pinto, M., Sobral, D., Santos, J. D., Nunes, A., et al. (2022). Phylogenomic characterization and signs of microevolution in the 2022 multi-country outbreak of monkeypox virus. Nat. Med. 28, 1569–1572. doi: 10.1038/s41591-022-01907-y
Johnston, S. C., Johnson, J. C., Stonier, S. W., Lin, K. L., Kisalu, N. K., Hensley, L. E., et al. (2015). Cytokine modulation correlates with severity of monkeypox disease in humans. J. Clin. Virol. 63, 42–45. doi: 10.1016/j.jcv.2014.12.001
Kabuga, A. I., El Zowalaty, M. E. (2019). A review of the monkeypox virus and a recent outbreak of skin rash disease in Nigeria. J. Med. Virol. 91, 533–540. doi: 10.1002/jmv.25348
Kalyar, F., Chen, X., Kunasekaran, M., Moa, A., Chughtai, A., Maclntyre, R. (2022). An unprecedented global resurgence of monkeypox. (Kalamazoo, Michigan: Global Biosecurity). doi: 10.31646/gbio.170
Khalili, A., Samara, A., O’Brien, P., Coutinho, C. M., Duarte, G., Quintana, S. M., et al. (2020). Monkeypox in pregnancy: update on current outbreak. Lancet 22, 2020–2022. doi: 10.1016/s1473-3099(03)00856-9
Kindrachuk, J., Arsenault, R., Kusalik, A., Kindrachuk, K. N., Trost, B., Napper, S., et al. (2012). Systems kinomics demonstrates congo basin monkeypox virus infection selectively modulates host cell signaling responses as compared to West African monkeypox virus. Mol. Cell. Proteomics 11, 1–12. doi: 10.1074/mcp.M111.015701
Kipkorir, V., Dhali, A., Srichawla, B., Kutikuppala, S., Cox, M., Ochieng, D., et al. (2022). The re-emerging monkeypox disease. Trop. Med. Int. Heal. 27, 961–969. doi: 10.1111/tmi.13821
Kmiec, D., Kirchhoff, F. (2022). Monkeypox: A new threat? Int. J. Mol. Sci. 23, 7866. doi: 10.3390/ijms23147866
Kotwal, G. J., Isaacs, S. N., Mckenzie, R., Frank, M. M., Moss, B. (1990). Inhibition of the complement cascade by the major secretory protein of vaccinia virus. Science 250, 827–830. doi: 10.1126/science.2237434
Krajcsi, P., Wold, W. S. M. (1998). Viral proteins that regulate cellular signalling. J. Gen. Virol. 79, 1323–1335. doi: 10.1099/0022-1317-79-6-1323
Kugelman, J. R., Johnston, S. C., Mulembakani, P. M., Kisalu, N., Lee, M. S., Koroleva, G., et al. (2014). Genomic variability of monkeypox virus among humans , democratic republic of the congo. Emerg. Infect. Dis. 20, 2–9. doi: 10.3201/eid2002.130118
Kumar, R., Nagar, S., Haider, S., Sood, U., Ponnusamy, K., Dhingra, G. G. (2023). Monkeypox virus: phylogenomics , host – pathogen interactome and mutational cascade. Microb Genom. 9, mgen00987. doi: 10.1099/mgen.0.000987
Ladnyj, I. D., Ziegler, P., Kima, E. (1972). A human infection caused by monkeypox virus in Basankusu Territory, Democratic Republic of the Congo. Bull. World Health Organ. 46, 593–597.
Lazear, E., Peterson, L. W., Nelson, C. A., Fremont, D. H. (2013). Crystal structure of the cowpox virus-encoded NKG2D ligand OMCP. J. Virol. 87, 840–850. doi: 10.1128/JVI.01948-12
Leung, J., Mccollum, A. M., Radford, K., Hughes, C., Lopez, A.S., Guagliardo, S.A.J., et al. (2019). Varicella in tshuapa province , Democratic Republic of Congo, 2009 – 2014. Trop. Med. Int. Heal. 24, 839–848. doi: 10.1111/tmi.13243
Li, M., Beg, A. A. (2000). Induction of necrotic-like cell death by tumor necrosis factor alpha and caspase inhibitors: novel mechanism for killing virus-infected cells. J. Virol. 74, 7470–7477. doi: 10.1128/JVI.74.16.7470-7477.2000
Li, K., Yuan, Y., Jiang, L., Liu, Y., Liu, Y., Zhang, L. (2023). Animal host range of mpox virus. J. Med. Virol. 95, e28513. doi: 10.1002/jmv.28513
Li, H., Zhang, H., Ding, K., Wang, X., Sun, G., Liu, Z. (2022). The evolving epidemiology of monkeypox virus. Cytokine Growth Factor Rev. 68, 1–12. doi: 10.1016/j.cytogfr.2022.10.002
Li, Y., Zhao, H., Wilkins, K., Hughes, C., Damon, I. K. (2010). Real-time PCR assays for the specific detection of monkeypox virus West African and Congo Basin strain DNA. J. Virol. Methods 169, 223–227. doi: 10.1016/j.jviromet.2010.07.012
Likos, A. M., Sammons, S. A., Olson, V. A., Frace, A. M., Li, Y., Olsen-Rasmussen, M., et al. (2005). A tale of two clades: Monkeypox viruses. J. Gen. Virol. 86, 2661–2672. doi: 10.1099/vir.0.81215-0
Liszewski, M. K., Leung, M. K., Hauhart, R., Buller, R. M. L., Bertram, P., Wang, X., et al. (2006). Structure and regulatory profile of the monkeypox inhibitor of complement: comparison to homologs in vaccinia and variola and evidence for dimer formation. J. Immunol. 176, 3725–3734. doi: 10.4049/jimmunol.176.6.3725
Liu, Q., Fu, L., Wang, B., Sun, Y., Wu, X., Peng, X., et al. (2023). Clinical characteristics of human mpox (Monkeypox) in 2022: A systematic review and meta-analysis. Pathogens 12, 146. doi: 10.3390/pathogens12010146
Liu, R., Moss, B. (2018). Vaccinia virus C9 ankyrin repeat/F-box protein is a newly identified antagonist of the type I interferon-induced antiviral state. J. Virol. 92, 1–17. doi: 10.1128/JVI.00053-18
Liu, J., Mucker, E. M., Chapman, J. L., Babka, A. M., Gordon, J. M., Bryan, A. V., et al. (2022). Retrospective detection of monkeypox virus in the testes of nonhuman primate survivors. Nat. Microbiol 7, 1980–1986. doi: 10.1038/s41564-022-01259-w
Lourie, B., Bingham, P. G., Evans, H. H., Foster, S. O., Nakano, J. H., Herrmann, K. L. (1972). Human infection with monkeypox virus: laboratory investigation of six cases in West Africa. Bull. World Health Organ. 46, 633–639.
Lum, F. M., Torres-Ruesta, A., Tay, M. Z., Lin, R. T. P., Lye, D. C., Rénia, L., et al. (2022). Monkeypox: disease epidemiology, host immunity and clinical interventions. Nat. Rev. Immunol. 22, 597–613. doi: 10.1038/s41577-022-00775-4
Luna, N., Ramírez, A. L., Muñoz, M., Ballesteros, N., Patiño, L. H., Castañeda, S. A., et al. (2022). Phylogenomic analysis of the monkeypox virus (MPXV) 2022 outbreak: Emergence of a novel viral lineage? Travel Med. Infect. Dis. 49, 102402. doi: 10.1016/j.tmaid.2022.102402
Luo, Q., Han, J. (2022). Preparedness for a monkeypox outbreak. Infect. Med. 1, 124–134. doi: 10.1016/j.imj.2022.07.001
Lysakova-Devine, T., Keogh, B., Harrington, B., Nagpal, K., Halle, A., Golenbock, D. T., et al. (2010). Viral inhibitory peptide of TLR4, a peptide derived from vaccinia protein A46, specifically inhibits TLR4 by directly targeting myD88 adaptor-like and TRIF-related adaptor molecule. J. Immunol. 185, 4261–4271. doi: 10.4049/jimmunol.1002013
Makkar, D. (2022). The latest news for may 2022 all you need to know on monkeypox. IP Int. J. Med. Microbiol. Trop. Dis. 8 (3), 183–195. doi: 10.18231/j.ijmmtd.2022.039
Malta, M., Mbala-Kingebeni, P., Rimoin, A. W., Strathdee, S. A. (2022). Monkeypox and global health inequities: A tale as old as time. Int. J. Environ. Res. Public Health 19, 10–13. doi: 10.3390/ijerph192013380
Mann, B. A., Huang, J. H., Li, P., Chang, H. C., Slee, R. B., O’Sullivan, A., et al. (2008). Vaccinia virus blocks Stat1-dependent and Stat1-independent gene expression induced by type I and type II interferons. J. Interf. Cytokine Res. 28, 367–379. doi: 10.1089/jir.2007.0113
Mansur, D. S., Maluquer de Motes, C., Unterholzner, L., Sumner, R. P., Ferguson, B. J., Ren, H., et al. (2013). Poxvirus targeting of E3 ligase β-trCP by molecular mimicry: A mechanism to inhibit NF-κB activation and promote immune evasion and virulence. PloS Pathog. 9, e1003183. doi: 10.1371/journal.ppat.1003183
Martin, D. P., Murrell, B., Golden, M., Khoosal, A., Muhire, B. (2015). RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 1, vev003. doi: 10.1093/ve/vev003
Mauldin, M. R., McCollum, A. M., Nakazawa, Y. J., Mandra, A., Whitehouse, E. R., Davidson, W., et al. (2022). Exportation of monkeypox virus from the african continent. J. Infect. Dis. 225, 1367–1376. doi: 10.1093/infdis/jiaa559
Mbala, P. K., Huggins, J. W., Riu-Rovira, T., Ahuka, S. M., Mulembakani, P., Rimoin, A. W., et al. (2017). Maternal and fetal outcomes among pregnant women with human monkeypox infection in the Democratic Republic of Congo. J. Infect. Dis. 216, 824–828. doi: 10.1093/infdis/jix260
McCarthy, M. W. (2022). Recent advances in the diagnosis monkeypox: implications for public health. Expert Rev. Mol. Diagn. 22, 739–744. doi: 10.1080/14737159.2022.2116979
McCollum, A., Nakazawa, Y., Ndongala, G. M., Pukuta, E., Karhemere, S., Lushima, R. S., et al. (2014). Human monkeypox in the Kivus, a conflict region of The Democratic Republic of the Congo. Int. J. Infect. Dis. 21, 192. doi: 10.1016/j.ijid.2014.03.820
Merouze, F., Lesoin, J. J. (1983). [Monkeypox: second human case observed in Ivory Coast (rural health sector of Daloa]. Med. Trop. 43, 145–147.
Mitjà, O., Ogoina, D., Titanji, B. K., Galvan, C., Muyembe, J.-J., Marks, M., et al. (2022). Monkeypox. Lancet. 401, 60–74. doi: 10.1016/S0140-6736(22)02075-X
Mogensen, T. H. (2009). Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 22, 240–273. doi: 10.1128/CMR.00046-08
Mohamed, M. R., McFadden, G. (2009). NFκB inhibitors: Strategies from poxviruses. Cell Cycle 8, 3125–3132. doi: 10.4161/cc.8.19.9683
Motwani, M., Pesiridis, S., Fitzgerald, K. A. (2019). DNA sensing by the cGAS–STING pathway in health and disease. Nat. Rev. Genet. 20, 657–674. doi: 10.1038/s41576-019-0151-1
Moulton, E. A., Atkinson, J. P., Buller, R. M. L. (2008). Surviving mousepox infection requires the complement system. PLoS Pathog. 4, e1000249. doi: 10.1371/journal.ppat.1000249
Nakazawa, Y., Emerson, G. L., Carroll, D. S., Zhao, H., Li, Y., Reynolds, M. G., et al. (2013). Phylogenetic and ecologic perspectives of a monkeypox outbreak, Southern Suda. Emerg. Infect. Dis. 19, 237–245. doi: 10.3201/eid1902.121220
Nakhaie, M., Arefinia, N., Charostad, J., Bashash, D., Haji Abdolvahab, M., Zarei, M. (2022). Monkeypox virus diagnosis and laboratory testing. Rev. Med. Virol. 33, e204. doi: 10.1002/rmv.2404
Nalca, A., Rimoin, A. W., Bavari, S., Whitehouse, C. A. (2005). Reemergence of monkeypox : prevalence , diagnostics , and countermeasures. Clin. Infect. Dis. 21702, 1765–1771. doi: 10.1086/498155
Ndodo, N., Ashcroft, J., Lewandowski, K., Yinka-Ogunleye, A., Chukwu, C., Ahmad, A., et al. (2023). Distinct monkeypox virus lineages co-circulating in humans before 2022. Nat. Med. 29, 2317–2324. doi: 10.1038/s41591-023-02456-8
Nigeria Centre for Disease Control (NCDC). (2022). UPDATE ON MONKEYPOX (MPX) IN NIGERIA. (Nigeria: Nigeria Centre for Disease Control (NCDC)).
Noe, S., Zange, S., Seilmaier, M., Antwerpen, M. H., Fenzl, T., Schneider, J., et al. (2022). Clinical and virological features of first human monkeypox cases in Germany. Infection. 51, 265–270. doi: 10.1007/s15010-022-01874-z
Nolen, L. D., Osadebe, L., Katomba, J., Likofata, J., Mukadi, D., Monroe, B., et al. (2015). Introduction of monkeypox into a community and household: Risk factors and zoonotic reservoirs in the democratic republic of the congo. Am. J. Trop. Med. Hyg. 93, 410–415. doi: 10.4269/ajtmh.15-0168
Noyce, R. S., Lederman, S., Evans, D. H. (2018). Construction of an infectious horsepox virus vaccine from chemically synthesized DNA fragments. PloS One 13, e0188453. doi: 10.1371/journal.pone.0188453
Nuzzo, J. B., Borio, L. L., Gostin, L. (2022). The WHO declaration of monkeypox as a global public health emergency. N. Engl. J. Med. 387, 679–691. doi: 10.1001/jama.2022.12513
Ortiz-Saavedra, B., León-Figueroa, D. A., Montes-Madariaga, E. S., Ricardo-Martínez, A., Alva, N., Cabanillas-Ramirez, C., et al. (2022). Antiviral treatment against monkeypox: A scoping review. Trop. Med. Infect. Dis. 7, 369. doi: 10.3390/tropicalmed7110369
O’Toole, A., Neher, R.A., Ndodo, N., Borges, V., Gannon, B., Gomes, J.P., et al. (2023). APOBEC3 deaminase editing in MPXV as evidence for sustained human transmission since at least 2016 Science. 382, 595–600. doi: 10.1126/science.adg8116
O’Toole, Á., Rambaut, A. (2022). Initial observations about putative APOBEC3 deaminase editing driving short-term evolution of MPXV since 2017 - Monkeypox / Evolution - Virological. Available at: https://virological.org/t/initial-observations-about-putative-apobec3-deaminase-editing-driving-short-term-evolution-of-mpxv-since-2017/830 (Accessed February 3, 2023).
Parker, S., Buller, R. M. (2013). A review of experimental and natural infections of animals with monkeypox virus between 1958 and 2012. Future Virol. 8, 129–157. doi: 10.2217/fvl.12.130
Payne, L. G. (1980). Significance of extracellular enveloped virus in the in vitro and in vivo dissemination of vaccinia. J. Gen. Virol. 50, 89–100. doi: 10.1099/0022-1317-50-1-89
Pears, C. (1995). Structure and function of the protein kinase C gene family. J. Biosci. 20, 311–332. doi: 10.1007/BF02703836
Petersen, E., Kantele, A., Koopmans, M., Asogun, D., Ogunlete, A., Ihekweazu, C., et al. (2019a). Human monkeypox –epidemiological and clinical characteristics, diagnosis and prevention. Infect. Dis. Clin. 33, 1027–1043. doi: 10.1016/j.idc.2019.03.001
Petersen, E., Kantele, A., Koopmans, M., Asogun, D., Yinka-Ogunleye, A., Ihekweazu, C., et al. (2019b). Human monkeypox: epidemiologic and clinical characteristics, diagnosis, and prevention. Infect. Dis. Clin. North Am. 33, 1027–1043. doi: 10.1016/j.idc.2019.03.001
Petersen, E., Zumla, A., Hui, D. S., Blumberg, L., Valdoleiros, S. R., Amao, L., et al. (2022). Vaccination for monkeypox prevention in persons with high-risk sexual behaviours to control on-going outbreak of monkeypox virus clade 3. Int. J. Infect. Dis. 122, 569–571. doi: 10.1016/j.ijid.2022.06.047
Phylogenetic pitchforks - Understanding Evolution. (2023). Available at: https://evolution.berkeley.edu/phylogenetic-systematics/reading-trees-a-quick-review/phylogenetic-pitchforks/ (Accessed February 3, 2023).
Poland, G. A., Kennedy, R. B., Tosh, P. K. (2022). Prevention of monkeypox with vaccines: a rapid review. Lancet Infect. Dis. 3099, 1–10. doi: 10.1016/S1473-3099(22)00574-6
Pourriyahi, H., Aryanian, Z., Afshar, Z. M., Goodarzi, A. (2023). A systematic review and clinical atlas on mucocutaneous presentations of the current monkeypox outbreak: With a comprehensive approach to all dermatologic and nondermatologic aspects of the new and previous monkeypox outbreaks. J. Med. Virol. 95, e28230. doi: 10.1002/jmv.28230
Rambaut, A., Drummond, A. J., Xie, D., Baele, G., Suchard, M. A. (2018). Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 67, 901–904. doi: 10.1093/sysbio/syy032
Rambaut, A., Lam, T. T., Carvalho, L. M., Pybus, O. G. (2016). Exploring the temporal structure of heterochronous sequences using TempEst (formerly Path-O-Gen). Virus Evol. 2, 1–7. doi: 10.1093/ve/vew007
Randall, R. E., Goodbourn, S. (2008). Interferons and viruses: An interplay between induction, signalling, antiviral responses and virus countermeasures. J. Gen. Virol. 89, 1–47. doi: 10.1099/vir.0.83391-0
Realegeno, S., Priyamvada, L., Kumar, A., Blackburn, J. B., Hartloge, C., Puschnik, A. S., et al. (2020). Conserved oligomeric golgi (COG) complex proteins facilitate orthopoxvirus entry, fusion and spread. Viruses 12, 707. doi: 10.3390/v12070707
Rehm, K. E., Connor, R. F., Jones, G. J. B., Yimbu, K., Roper, R. L. (2011). Vaccinia virus A35R inhibits MHC class II antigen presentation. Virology 23, 1–7. doi: 10.1016/j.virol.2009.11.008.Vaccinia
Reynolds, M. G., Davidson, W. B., Curns, A. T., Conover, C. S., Huhn, G., Davis, J. P., et al. (2007). Spectrum of infection and risk factors for human monkeypox, United State. Emerg. Infect. Dis. 13, 1332–1339. doi: 10.3201/eid1309.070175
Reynolds, M. G., Guagliardo, S. A. J., Nakazawa, Y. J., Doty, J. B., Mauldin, M. R. (2018). Understanding orthopoxvirus host range and evolution: from the enigmatic to the usual suspects. Curr. Opin. Virol. 28, 108–115. doi: 10.1016/j.coviro.2017.11.012
Rimoin, A. W., Kisalu, N., Kebela-Ilunga, B., Mukaba, T., Wright, L. L., Formenty, P., et al. (2011). Endemic human monkeypox, Republic of Congo, democratic 2001–2004. J. Phys. A Math. Theor. 44, 1689–1699. doi: 10.1088/1751-8113/44/8/085201
Rimoin, A. W., Mulembakani, P. M., Johnston, S. C., Lloyd Smith, J. O., Kisalu, N. K., Kinkela, T. L., et al. (2010). Major increase in human monkeypox incidence 30 years after smallpox vaccination campaigns cease in the Democratic Republic of Congo. Proc. Natl. Acad. Sci. U. S. A. 107, 16262–16267. doi: 10.1073/pnas.1005769107
Riopelle, J. C., Munster, V. J., Port, J. R. (2022). Atypical and unique transmission of monkeypox virus during the 2022 outbreak: an overview of the current state of knowledge. Viruses 14, 2012. doi: 10.3390/v14092012
Rizk, J. G., Lippi, G., Henry, B. M., Forthal, D. N., Rizk, Y. (2022). Prevention and treatment of monkeypox. Drugs 82, 957–963. doi: 10.1007/s40265-022-01742-y
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D. L., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: efficient bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Rubins, K. H., Hensley, L. E., Relman, D. A., Brown, P. O. (2011). Stunned silence: Gene expression programs in human cells infected with monkeypox or vaccinia virus. PloS One 6, e15615. doi: 10.1371/journal.pone.0015615
Sah, R., Abdelaal, A., Reda, A., Katamesh, B. E., Manirambona, E., Abdelmonem, H., et al. (2022a). Monkeypox and its possible sexual transmission: where are we now with its evidence? Pathogens 11, 1–10. doi: 10.3390/pathogens11080924
Sah, R., Mohanty, A., Reda, A., Padhi, B. K., Rodriguez-Morales, A. J. (2022b). Stigma during monkeypox outbreak. Front. Public Heal 10. doi: 10.3389/fpubh.2022.1023519
Saijo, M., Ami, Y., Suzaki, Y., Nagata, N., Iwata, N., Hasegawa, H., et al. (2009). Virulence and pathophysiology of the Congo Basin and West African strains of monkeypox virus in non-human primates. J. Gen. Virol. 90, 2266–2271. doi: 10.1099/vir.0.010207-0
Salter, J. D., Bennett, R. P., Smith, H. C. (2016). The APOBEC protein family: united by structure, divergent in function. Trends Biochem. Sci. 41, 578–594. doi: 10.1016/j.tibs.2016.05.001
Scarpa, F., Sanna, D., Azzena, I., Cossu, P., Locci, C., Angeletti, S., et al. (2022). Genetic variability of the monkeypox virus clade IIb B.1. J. Clin. Med. 11 6388. doi: 10.3390/jcm11216388
Schmidt, F. I., Mercer, J. (2012). Vaccinia virus egress: Actin OUT with ClathrIN. Cell Host Microbe 12, 263–265. doi: 10.1016/j.chom.2012.08.007
Schnierle, B. S. (2022). Monkeypox goes north: ongoing worldwide monkeypox infections in humans. Viruses 14, 1874. doi: 10.3390/v14091874
Seet, B. T., Johnston, J. B., Brunetti, C. R., Barrett, J. W., Everett, H., Cameron, C., et al. (2003). Poxviruses and immune evasion. Annu. Rev. Immunol. 21, 377–423. doi: 10.1146/annurev.immunol.21.120601.141049
Senkevich, T. G., Yutin, N., Wolf, Y. I., Koonin, E. V., Moss, B. (2021). Ancient gene capture and recent gene loss shape the evolution of orthopoxvirus-host interaction genes. MBio 12, e0149521. doi: 10.1128/mBio.01495-21
Shchelkunov, S. N., Totmenin, A. V., Babkin, I. V., Safronov, P. F., Ryazankina, O. I., Petrov, N. A., et al. (2001). Human monkeypox and smallpox viruses: Genomic comparison. FEBS Lett. 509, 66–70. doi: 10.1016/S0014-5793(01)03144-1
Shchelkunov, S. N., Totmenin, A. V., Safronov, P. F., Mikheev, M. V., Gutorov, V. V., Ryazankina, O. I., et al. (2002). Analysis of the monkeypox virus genome. Virology 297, 172–194. doi: 10.1006/viro.2002.1446
Shen-Gunther, J., Cai, H., Yufeng, W. (2023). A customized monkeypox virus genomic database (MPXV DB v1.0) for rapid sequence analysis and phylogenomic discoveries in CLC microbial genomics. Viruses 15, 40. doi: 10.3390/v15010040
Shisler, J. L., Jin, X.-L. (2004). The vaccinia virus K1L gene product inhibits host NF-κB activation by preventing IκBα Degradation. J. Virol. 78, 3553–3560. doi: 10.1128/JVI.78.7.3553-3560.2004
Siegrist, E. A., Sassine, J. (2022). Antivirals with activity against monkeypox: A clinically oriented review. Clin. Infect. Dis. 76, 155–164. doi: 10.1093/cid/ciac622
Singhal, T., Kabra, S. K., Lodha, R. (2022). Monkeypox : A review. Indian J. Pediatr. 89, 955–960. doi: 10.1007/s12098-022-04348-0
Sivan, G., Weisberg, A. S., Americo, J. L., Moss, B. (2016). Retrograde transport from early endosomes to the trans -golgi network enables membrane wrapping and egress of vaccinia virus virions. J. Virol. 90, 8891–8905. doi: 10.1128/JVI.01114-16
Sklenovská, N., Van Ranst, M. (2018). Emergence of monkeypox as the most important orthopoxvirus infection in humans. Front. Public Heal. 6. doi: 10.3389/fpubh.2018.00241
Slowinski, J. B. (2001). Molecular polytomies. Mol. Phylogenet. Evol. 19, 114–120. doi: 10.1006/mpev.2000.0897
Song, H., Josleyn, N., Janosko, K., Skinner, J., Reeves, R. K., Cohen, M., et al. (2013). Monkeypox virus infection of rhesus macaques induces massive expansion of natural killer cells but suppresses natural killer cell functions. PloS One 8, e77804. doi: 10.1371/journal.pone.0077804
Sousa, Á.F.L.d., Sousa, A.R.d., Fronteira, I. (2022). Monkeypox: between precision public health and stigma risk. Rev. Bras. Enferm. 75, e750501. doi: 10.1590/0034-7167.2022750501
Stephen, R., Alele, F., Olumoh, J., Tyndall, J., Okeke, M. I. (2022). The epidemiological trend of monkeypox and monkeypox-varicella zoster viruses co-infection in North-Eastern Nigeria. Front. Public Heal. 10, 1066589. doi: 10.3389/fpubh.2022.1066589
Stuart, J. H., Sumner, R. P., Lu, Y., Snowden, J. S., Smith, G. L. (2016). Vaccinia virus protein C6 inhibits type I IFN signalling in the nucleus and binds to the transactivation domain of STAT2. PloS Pathog. 12, 1–24. doi: 10.1371/journal.ppat.1005955
Suchard, M. A., Lemey, P., Baele, G., Ayres, D. L., Drummond, A. J., Rambaut, A. (2018). Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 4, 1–5. doi: 10.1093/ve/vey016
Sun, S.-C. (2017). The non-canonical NF-κB pathway in immunity and inflammation. Nat. Rev. Immunol. 17, 545–558. doi: 10.1038/nri.2017.52.The
Tchokoteu, P. F., Kago, I., Tetanye, E., Ndoumbe, P., Pignon, D., Mbede, J. (1991). Variola or a severe case of varicella? A case of human variola due to monkeypox virus in a child from the Cameroon. Ann. Soc. Belg Med. Trop. 71, 123–128.
Titanji, B. K., Tegomoh, B., Nematollahi, S., Konomos, M., Kulkarni, P. A. (2022). Monkeypox: A contemporary review for healthcare professionals. Open Forum Infect. Dis. 9, 1–13. doi: 10.1093/ofid/ofac310
Tu, S.-L. (2015). The bioinformatic characterization of five novel poxviruses. (Oak Bay and Saanich, British Columbia, Canada: University of Victoria).
Uvarova, E. A., Shchelkunov, S. N. (2001). Species-specific differences in the structure of orthopoxvirus complement-binding protein. Virus Res. 81, 39–45. doi: 10.1016/S0168-1702(01)00332-X
Van Vliet, K., Mohamed, M. R., Zhang, L., Villa, N. Y., Werden, S. J., Liu, J., et al. (2009). Poxvirus proteomics and virus-host protein interactions. Microbiol. Mol. Biol. Rev. 73, 730–749. doi: 10.1128/MMBR.00026-09
Vaughan, A., Aarons, E., Astbury, J., Balasegaram, S., Beadsworth, M., Beck, C. R., et al. (2018). Two cases of monkeypox imported to the United Kingdom, september 2018. Eurosurveillance 23, 1800509. doi: 10.2807/1560-7917.ES.2018.23.38.1800509
Velavan, T. P., Meyer, C. G. (2022). Monkeypox 2022 outbreak: An update. Trop. Med. Int. Heal. 27, 604–605. doi: 10.1111/tmi.13785
Walker, M. C. (2022). Monkeypox Virus Hosts and Transmission Routes : A Systematic Review of a Zoonotic Pathogen. Available at: https://scholarworks.uark.edu/biscuht/69%0AThis.
Wang, L., Shang, J., Weng, S., Aliyari, S. R., Ji, C., Cheng, G., et al. (2022). Genomic annotation and molecular evolution of monkeypox virus outbreak in 2022. J. Med. Virol. 95 (1), e28036. doi: 10.1002/jmv.28036
Wassenaar, T. M., Wanchai, V., Ussery, D. W. (2022). Comparison of Monkeypox virus genomes from the 2017 Nigeria outbreak and the 2022 outbreak. J. Appl. Microbiol. 133 (6), 3690–3698. doi: 10.1111/jam.15806
Weaver, J. R., Isaacs, S. N. (2008). Monkeypox virus and insights into its immunomodulatory proteins. Immunol. Revolut. 225, 96–113. doi: 10.1111/j.1600-065X.2008.00691.x.Monkeypox
Wenner, H. A., Macasaet, F. D., Kamitsuka, P. S., Kidd, P. (1968). Monkey pox. I. Clinical, virologic and immunologic studies. Am. J. Epidemiol. 87, 551–566. doi: 10.1093/oxfordjournals.aje.a120846
White, S. D., Jacobs, B. L. (2012). The amino terminus of the vaccinia virus E3 protein is necessary to inhibit the interferon response. J. Virol. 86, 5895–5904. doi: 10.1128/JVI.06889-11
Wolffe, E. J., Weisberg, A. S., Moss, B. (1998). Role for the vaccinia virus A36R outer envelope protein in the formation of virus-tipped actin-containing microvilli and cell-to-cell virus spread. Virology 244, 20–26. doi: 10.1006/viro.1998.9103
World Health Organization. (2022). 2022 Mpox (Monkeypox) Outbreak: Global Trends. Available at: https://worldhealthorg.shinyapps.io/mpx_global/ (Accessed February 2, 2023).
World Health Organization. (2022a). Disease Outbreak News; Multi-country monkeypox outbreak in non-endemic countries. Available at: https://www.who.int/emergencies/disease-outbreak-news/item/2022-DON385 (Accessed February 2, 2023).
World Health Organization. (2022b). Multi-country monkeypox outbreak: situation update. Available at: https://www.who.int/emergencies/disease-outbreak-news/item/2022-DON393 (Accessed February 2, 2023).
Xiang, Y., White, A. (2022). Monkeypox virus emerges from the shadow of its more infamous cousin: family biology matters. Emerg. Microbes Infect. 11, 1768–1777. doi: 10.1080/22221751.2022.2095309
Xuan, D. T. M., Yeh, I. J., Wu, C. C., Su, C. Y., Liu, H. L., Chiao, C. C., et al. (2022). Comparison of transcriptomic signatures between monkeypox-infected monkey and human cell lines. J. Immunol. Res. 2022, 3883822. doi: 10.1155/2022/3883822
Ye, F., Song, J., Zhao, L., Zhang, Y., Xia, L., Zhu, L., et al. (2019). Molecular evidence of human monkeypox virus infection, Sierra Leone. Emerg. Infect. Dis. 25, 1220–1222. doi: 10.3201/eid2506.180296
Yeh, T.-Y., Contreras, G. P. (2022). Recombination shapes 2022 monkeypox outbreak. Med. 3, 824–826. doi: 10.1101/2022.08.09.22278589
Yeh, T.-Y., Hsieh, Z.-Y., Feehley, M.C., Feehley, P.J., Contreas, G.P., Su, Y.-C., et al. (2022). Recombination shapes the 2022 monkeypox ( mpox ) outbreak. Med 3, 824–826. doi: 10.1016/j.medj.2022.11.003
Yinka-Ogunleye, A., Aruna, O., Dalhat, M., Ogoina, D., McCollum, A., Disu, Y., et al. (2019). Outbreak of human monkeypox in Nigeria in 2017–18: a clinical and epidemiological report. Lancet Infect. Dis. 19, 872–879. doi: 10.1016/S1473-3099(19)30294-4
Keywords: monkeypox, genomics, evolution, antivirals, epidemiology, infection biology, biosafety, one health
Citation: Alakunle E, Kolawole D, Diaz-Cánova D, Alele F, Adegboye O, Moens U and Okeke MI (2024) A comprehensive review of monkeypox virus and mpox characteristics. Front. Cell. Infect. Microbiol. 14:1360586. doi: 10.3389/fcimb.2024.1360586
Received: 23 December 2023; Accepted: 20 February 2024;
Published: 06 March 2024.
Edited by:
Mohd Ahmar Rauf, Wayne State University, United StatesReviewed by:
Ali Tavakoli Pirzaman, Babol University of Medical Sciences, IranCopyright © 2024 Alakunle, Kolawole, Diaz-Cánova, Alele, Adegboye, Moens and Okeke. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ugo Moens, dW1vMDAwQHBvc3QudWl0Lm5v; Malachy Ifeanyi Okeke, bWFsYWNoeS5va2VrZUBhdW4uZWR1Lm5n
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.