
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Cell. Infect. Microbiol., 13 February 2023
Sec. Parasite and Host
Volume 13 - 2023 | https://doi.org/10.3389/fcimb.2023.1113896
This article is part of the Research TopicWomen in Parasite & Host vol. IIView all 4 articles
 Aryani Felixa Fajardo Martínez1
Aryani Felixa Fajardo Martínez1 Samuel Cota Teixeira1
Samuel Cota Teixeira1 Guilherme de Souza1
Guilherme de Souza1 Alessandra Monteiro Rosini1
Alessandra Monteiro Rosini1 Joed Pires de Lima Júnior1
Joed Pires de Lima Júnior1 Gabriel Nogueira Melo1
Gabriel Nogueira Melo1 Kelvin Orlando Espinoza Blandón2
Kelvin Orlando Espinoza Blandón2 Angelica Oliveira Gomes3
Angelica Oliveira Gomes3 Sergio Ricardo Ambrósio4
Sergio Ricardo Ambrósio4 Rodrigo Cassio Sola Veneziani4
Rodrigo Cassio Sola Veneziani4 Jairo Kenupp Bastos5
Jairo Kenupp Bastos5 Carlos Henrique Gomes Martins6
Carlos Henrique Gomes Martins6 Eloisa Amália Vieira Ferro1
Eloisa Amália Vieira Ferro1 Bellisa Freitas Barbosa1*
Bellisa Freitas Barbosa1*The conventional treatment of congenital toxoplasmosis is mainly based on the combination of sulfadiazine and pyrimethamine. However, therapy with these drugs is associated with severe side effects and resistance, requiring the study of new therapeutic strategies. There are currently many studies with natural products, including Copaifera oleoresin, showing actions against some pathogens, as Trypanosoma cruzi and Leishmania. In the present study, we investigated the effects of the leaf hydroalcoholic extract and oleoresin from Copaifera multijuga against Toxoplasma gondii in human villous (BeWo) and extravillous (HTR8/SVneo) trophoblast cells, as well as in human villous explants from third-trimester pregnancy. For this purpose, both cells and villous explants were infected or not with T. gondii, treated with hydroalcoholic extract or oleoresin from C. multijuga and analyzed for toxicity, parasite proliferation, cytokine and ROS production. In parallel, both cells were infected by tachyzoites pretreated with hydroalcoholic extract or oleoresin, and adhesion, invasion and replication of the parasite were observed. Our results showed that the extract and oleoresin did not trigger toxicity in small concentrations and were able to reduce the T. gondii intracellular proliferation in cells previously infected. Also, the hydroalcoholic extract and oleoresin demonstrated an irreversible antiparasitic action in BeWo and HTR8/SVneo cells. Next, adhesion, invasion and replication of T. gondii were dampened when BeWo or HTR8/SVneo cells were infected with pretreated tachyzoites. Finally, infected and treated BeWo cells upregulated IL-6 and downmodulated IL-8, while HTR8/SVneo cells did not change significantly these cytokines when infected and treated. Finally, both the extract and oleoresin reduced the T. gondii proliferation in human explants, and no significant changes were observed in relation to cytokine production. Thus, compounds from C. multijuga presented different antiparasitic activities that were dependent on the experimental model, being the direct action on tachyzoites a common mechanism operating in both cells and villi. Considering all these parameters, the hydroalcoholic extract and oleoresin from C. multijuga can be a target for the establishment of new therapeutic strategy for congenital toxoplasmosis.
Toxoplasma gondii is an obligate intracellular protozoan parasite belonging to the Apicomplexa phylum, responsible for serious morbidities mainly related to newborns and immunocompromised people (Blader et al., 2015; Zhang et al., 2019). It is estimated that a third of the world population is infected with this microorganism, making it one of the most successful microorganisms (Montoya and Liesenfeld, 2004). Congenital toxoplasmosis is considered as one of the most serious forms of the disease, caused by the transplacental passage of T. gondii tachyzoites acquired during or just before pregnancy. It triggers serious implications for the fetal development and may go unreported into adolescence or adulthood (Kodjikian, 2010). Primary infection during pregnancy can lead to spontaneous abortion, stillbirth, premature delivery, malformations and neurological or ocular disorders in newborns (Carlier et al., 2012; Oz, 2014).
An immune response, preferentially T helper 1 (Th1), is induced when the parasites invade the host cells, requiring components of the innate and adaptive immune response (Carruthers, 2002). Initially, T. gondii is recognized by innate immune response cells, which stimulate interleukin (IL)-12 release by dendritic cells, macrophages and neutrophils, and induce interferon (IFN)-γ production by natural killer (NK) cells (Fitch et al., 1975). The production of these proinflammatory cytokines is associated with the activation of the adaptive immune response mediated by CD4+ and CD8+ T cells, which produce high levels of IFN-γ as well as other proinflammatory cytokines, besides triggering the release of various inflammatory mediators, such as nitric oxide (NO) by macrophages (Buzoni-Gatel et al., 2006; Miller et al., 2009). In addition to IFN-γ and IL-12, other proinflammatory cytokines are of great importance in controlling T. gondii infection, such as IL-6 and macrophage migration inhibitory factor (MIF) (Castro et al., 2013; Souza et al., 2021). Therefore, the production of these proinflammatory cytokines stands out as a robust and classic defense mechanism against T. gondii infection in the host. In parallel, IL-8 has been considered an important cytokine during T. gondii infection. This cytokine is a member of the CXC chemokine family and it is an important mediator during innate immune response to several pathogens (Dong and Zheng, 2015). Studies showed that T. gondii has the ability to attract IL-8 and this cytokine can recruit inflammatory cells to local infection, attracting neutrophil and immature dendritic cells and, consequently, promoting a dissemination of the infection by host organism (Sommerville et al., 2013). Then, IL-8 can be an important mediator to favor the establishment of the infection. In addition, anti-inflammatory cytokines such as IL-10 and transforming growth factor (TGF)-β are necessary to prevent an exacerbated immune response that could be detrimental to the host (Fowler et al., 1976). The immune response is not sufficient to clear the infection. In this sense, the establishment of new drugs are mandatory for infected pregnant women and congenitally infected children.
When maternal infection is present, but without evidence of fetal infection, the classical therapy is based in spiramycin, a macrolide antibiotic that prevents congenital transmission (Elsheikha, 2008; Tamaru et al., 2011; Peyron et al., 2017). In the case of confirmed fetal infection, the first treatment option is the combination of pyrimethamine and sulfadiazine (Villena et al., 1998; Doliwa et al., 2013). The coadministration of folinic acid becomes necessary to minimize the toxic effects of pyrimethamine (Montoya and Remington, 2008). It should be noted that sulfadiazine is associated with gastrointestinal disturbances, and patients often do not tolerate this chemotherapy (Montoya and Remington, 2008). Furthermore, approximately half of the patients treated with spiramycin retained Toxoplasma DNA in their blood for a long time (Habib, 2008). Therefore, finding active and less toxic drugs as new therapeutic strategies to prevent or treat congenital toxoplasmosis is absolutely necessary and encouraging. Our previous studies have shown alternative drugs or potential molecular targets with great effects against T. gondii infection (Costa et al., 2009; Souza et al., 2021; Teixeira et al., 2021). Thus, it is important to highlight the importance of studying alternative treatments for the control of T. gondii infection.
In this scenario, the use of natural products and their isolated molecules has emerged as an interesting tool for the discovery of new relevant substances with a great antiparasitic action against T. gondii. Huang et al. (2021) used five essential oleoresins (EOs) and evaluated their antiparasitic activity against T. gondii in fibroblast cells (HFF). EOs from Eucalyptus globulus, Cupressus sempervirens, Citrus aurantifolia and Melaleuca alternifolia did not show any effect in T. gondii infection. However, EO from Pelargonium X. Asperum (PaEO) inhibited the growth of T. gondii in a dose-dependent manner. Also, another study conducted by Sharif et al. (2019) evaluated the effect of the hydroalcoholic extract from the stem of Tinospora crispa (EETC) against T. gondii using Vero cells. The authors observed that EETC induced more than 70% and 80% reduction in invasion rate and parasite intracellular proliferation, respectively. In addition, Miranda et al. (2021) investigated the action of the hydroalcoholic extract of Annona muricata (EtOHAm) and its fractions in the control of T. gondii, showing that the HexAm, CH2Cl2Am and EtOAcAm fractions exerted anti-Toxoplasma effect in vitro; however, only EtOHAm was effective in improving survival and decreasing tissue parasitism in C57BL/6 mice.
Copaifera genus, also popularly known as “pau d’oleo”, belongs to the Fabaceae family (Leguminosae). The representative species of this genus are found mainly in the American and African continents (Veiga-Júnior and Pinto, 2002). Copaifera oleoresin is a natural substance composed of a non-volatile solid resinous part formed by diterpenic acids responsible for 55 to 60% of its constitution, which is diluted in another volatile part, the essential oleoresin composed of sesquiterpenes (Cascon and Gilbert, 2000). Copaifera oleoresin has sesquiterpenes, diterpenes and β-caryophyllene as its main compound, possessing healing and antiseptic properties, in addition to being considered as anti-inflammatory and antibiotic (Santos et al., 2008; Souza et al., 2011; Arruda et al., 2019). Several studies have demonstrated the antiparasitic action of the oleoresin and isolated components from Copaifera or other members from the Fabaceae family, especially with trypanocidal, leishmanicidal and antiplasmodial activities, through direct action on parasites and/or by modulating the immune response (Izumi et al., 2013; Tomani et al., 2021). Extracts and purified molecules of various species of the genera Piper, Tanacetum, Porophyllum and Copaifera exhibited interesting antitrypanosomal and antileishmanial activities. These natural compounds affected different structures in the parasites, suggesting that the mitochondria are the strategic target to induce parasite death (Lazarin-Bidóia et al., 2022).
However, few studies have addressed the action of these compounds against T. gondii. Recently, Teixeira et al. (2020) demonstrated the ability of four Copaifera oleoresins (C. reticulata, C. duckei, C. paupera and C. pubiflora) to impair T. gondii infection in human villous trophoblast cells (BeWo lineage) and human villous explants from third-trimester pregnancy. However, there are no studies about the possible effects of the hydroalcoholic extract and oleoresin from C. multijuga against T. gondii infection in maternal-fetal interface. C. multijuga is found in the Amazon and is now being studied extensively due to its various properties, the most expressive being its anti-inflammatory, anticancer and antimicrobial (antiseptic) properties (Pinheiro et al., 2017).
In this sense, the present study aims to demonstrate, for the first time, the anti-T. gondii effects of the leaf hydroalcoholic extract and oleoresin from C. multijuga in human trophoblast cells. As experimental models, we used BeWo and HTR8/SVneo cells, representatives of human villous and extravillous trophoblasts, respectively, as well as human villous explants as an experimental model of maternal-fetal interface. Our research group widely uses these cells and explants in studies about human maternal-fetal interface (Barbosa et al., 2014; Barbosa et al., 2015; Teixeira et al., 2020; Souza et al., 2021; Teixeira et al., 2021).
BeWo cells were commercially obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA), while HTR8/SVneo cells were a gift from Dr. Estela Bevilacqua (University of São Paulo, São Paulo, SP, Brazil). Both cells were cultured in RPMI 1640 medium (Cultilab, Campinas, SP, Brazil) supplemented with 100 U/mL penicillin (Sigma Chemical Co., St. Louis, MO, USA), 100 μg/mL streptomycin (Sigma) and 10% fetal bovine serum (FBS) (Cultilab) at 37°C and 5% CO2.
Human third-trimester placentas (36 to 40 weeks of pregnancy), n = 4, were collected after elective cesarean deliveries at the Clinics Hospital of the Federal University of Uberlândia (HC-UFU), MG, Brazil. Placental tissues were collected based on exclusion criteria, as follows: pre-eclampsia, chronic hypertension, infectious diseases including toxoplasmosis, chorioamnionitis, chronic kidney disease, heart disease, connective tissue disease, pre-existing diabetes mellitus, gestational diabetes mellitus and other pathological manifestations. Briefly, placental tissues were washed in sterile PBS to remove excess blood, then aseptically dissected to remove endometrial tissue and fetal membranes up to 1 h after collection. Terminal chorionic villi containing five to seven free tips per explant were harvested and added to 96-well microplates (one villus per well) in 200 µL/well of fresh RPMI 1640 medium supplemented with 100 U/mL penicillin, 100 µg/mL streptomycin and 10% FBS for 24 h at 37°C in a humidified atmosphere containing CO2 (5%) (Silva et al., 2017).
Toxoplasma gondii tachyzoites (2F1 clone) constitutively expressing the β-galactosidase gene were maintained by serial passages in BeWo cells cultured in medium containing 2% FBS, 100 U/mL penicillin and 100 μg/mL streptomycin at 37°C and 5% CO2. 2F1 clone is derived from the highly virulent strain RH and was a gift from Dr. Vern Carruthers, School of Medicine from the University of Michigan (USA).
The samples of oleoresin and leaf hydroalcoholic extract from C. multijuga were provided by Professor Carlos Henrique Gomes Martins from the Department of Microbiology (DEMIC), Institute of Biomedical Sciences (ICBIM), Federal University of Uberlândia (UFU), Uberlândia, MG, Brazil. Authorization to carry out scientific studies with plant species from the Brazilian biodiversity was requested from the Council for Authorization and Information on Biodiversity (SIBIO/ICMBio/MMA/BRAZIL) and Genetic Heritage Management (CGEN/MMA/BRAZIL). Authorizations to carry out research activities with these plants were issued under numbers 35143-1 and 010225/2014-5, respectively.
Authentic oleoresin was collected in the North region, in the state of Amazonas, in the city of Manacapuru, by Jonas J. M. da Silva by drilling the tree trunks using a 2” drill bit. The exudates removed were stored in glass bottles. All the plant material collected was identified by Silvane Tavares Rodrigues, a voucher specimen (NID 03/2013 and 62/2013) was deposited in the Herbarium of the Brazilian Agricultural Research Corporation (Embrapa Eastern Amazon), by direct comparison with authentic herbarium vouchers, of which a taxonomic identity certificate is available upon request.
Air dried leaves (40 °C for 48 h) of C. multijuga (200 g) were grounded and then exhaustively extracted by maceration with 1.2 L of ethanol/H2O (7:3) at room temperature for 48 h to afford 50 g of C. multijuga hydroalcoholic extract of leaves after lyophilization (Pimenta et al., 2019).
We verified the toxicity of the compounds selected in BeWo and HTR8/SVneo cells. For this purpose, we performed the MTT [(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltertrazolin bromide)] colorimetric assay, following the protocol described by Mosmann (1983). BeWo (3×104 cells/well/200 µL) and HTR8/SVneo (1.5×104 cells/well/200 µL) cells were cultured in 96-well plates for 24 h in RPMI 1640 medium with 10% FBS at 37°C and 5% CO2. Next, cells were treated in serial dilutions with increasing concentrations of oleoresin or hydroalcoholic extract from C. multijuga in a medium with 10% FBS for 24 h. In parallel, both cells were treated with DMSO (0.8%), the percentage used to obtain higher dilution for both oleoresin and hydroalcoholic extract. As control, cells were not treated and received only medium. Then, the supernatants were removed and the cells incubated with 10 µL MTT (5 mg/mL) plus 90 µL of RPMI 10% FBS medium for 4 h at 37°C and 5% CO2. Formazan crystals resulting from cellular metabolism were solubilized by adding a solution containing 10% sodium dodecyl sulfate (SDS, Sigma) and 50% N,N-dimethyl formamide (Sigma) for 30 min (Mosmann, 1983). Optical densities were measured at 570 nm on a plate reader (Versa Max ELISA Microplate Reader, Molecular Devices, Sunnyvale, CA, USA). Data were expressed as the percentage of viable cells (cell viability %) in comparison to untreated cells (100% of cell viability). Three independent experiments were performed in eight replicates.
We selected different concentrations of oleoresin and hydroalcoholic extract from C. multijuga after MTT assay and tested their effect on T. gondii growth. BeWo (3×104 cells/well/200 µL) and HTR8/SVneo (1.5×104 cells/well/200 µL) cells were cultured in 96-well plates for 24 h in RPMI 1640 medium with 10% FBS at 37°C and 5% CO2. Next, both cells were infected with T. gondii tachyzoites at a multiplicity of infection (MOI) of 3:1 (parasites per cell). After 3 h, the medium was discarded, extracellular parasites were retired by washing with medium, and the cells were treated with non-toxic concentrations of oleoresins or hydroalcoholic extracts from C. multijuga for additional 24 h at 37°C and 5% CO2 as follows: for BeWo cells, 4 to 32 µg/mL; for HTR8/SVneo, 4 to 16 µg/mL. Sulfadiazine and pyrimethamine (SDZ + PYR, 200 + 8 µg/mL for BeWo, and 100 + 4 µg/mL for HTR8/SVneo) was used as positive control (Silva et al., 2017). BeWo or HTR8/SVneo cells infected by T. gondii and non-treated were used as negative controls. After 24 h of treatment, the culture supernatants were collected and stored at -80°C for further measurement of cytokines. In parallel, T gondii intracellular proliferation was analyzed using a colorimetric β-galactosidase assay (Silva et al., 2017; Souza et al., 2021). We quantified T. gondii intracellular proliferation and calculated the number of tachyzoites in comparison with a standard curve containing free tachyzoites (1×106 to 15,625×103 parasites). Infected BeWo or HTR8/SVneo cells treated only with medium (negative control) represented uninhibited parasite growth. We calculated the means of untreated cells (medium) that corresponded to 100% of T. gondii proliferation. Then, all values about number of tachyzoites obtained from all treatments were compared to 100%, and data demonstrated in % of T. gondii proliferation. Three independent experiments were performed in eight replicates.
In order to evaluate the maintenance of the antiparasitic effects of oleoresins and hydroalcoholic extracts in T. gondii growth, we firstly performed the reversibility test (Teixeira et al., 2020). Briefly, BeWo (3×104 cells/well/200 µL) or HTR8/SVneo (1.5×104 cells/well/200 µL) cells were seeded in 96-well plates as above described. After 24 h in culture, cells were infected with T. gondii tachyzoites (3:1) for 3 h, washed to remove extracellular parasites and treated for two conditions as follows: (1) BeWo cells were treated with oleoresin or hydroalcoholic extract (both 16 or 32 µg/mL), HTR8/SVneo cells were treated with oleoresin or hydroalcoholic extract (both 8 or 16 µg/mL), or both cells received SDZ + PYR (positive control) or only medium (negative control), then parasite intracellular proliferation was performed after 24 h in culture conditions; (2) cells and parasites were cultured in the same conditions as previously described, but after 24 h of treatment, the cells were washed, the medium replaced by new RPMI 1640 medium without treatments, and the parasites were allowed to grow for additional 24 h. In both situations, we quantified the T. gondii proliferation using the β-galactosidase assay as mentioned above. Finally, we measured the percentage reversibility rate (treatment reversibility %) at 24 h after treatment removal compared to the untreated group (considered as 100% reversibility) and the corresponding treatment condition at 24 h of treatment (baseline for comparison). Three independent experiments were performed in eight replicates.
To corroborate the reversibility data, we investigated whether the treatment with oleoresin or hydroalcoholic extract in infected BeWo and HTR8/SVneo cells would interfere with the ability of these parasites to invade and replicate within new fresh cells. Briefly, BeWo and HTR8/SVneo cells were seeded at 1.0x106 cells/well in 6-well plates. After 24 h, cells were infected with T. gondii tachyzoites (3:1) for 3 h at 37°C and 5% CO2. Then, we treated the cells with oleoresin or hydroalcoholic extract at 16 or 32 µg/mL for BeWo; or 8 or 16 µg/mL for HTR8/SVneo. Either SDZ + PYR or only medium was added as controls. After 24 h, we collected the intracellular parasites from infected cells by multiple passages through a 21- and 26-gauge needle. Finally, T. gondii tachyzoites from each experimental condition were then allowed to reinfect previously seeded BeWo (3×104 cells/well/200 µL) and HTR8/SVneo (1.5×104 cells/well/200 µL) cells monolayers in 96-well plates. The ability to infect the cells was analyzed after 3 h, when the β-galactosidase assay was used to quantify the total number of tachyzoites (% of T. gondii invasion), as described above. Three independent experiments were performed in eight replicates.
We performed an adherence test (Oliveira et al., 2006; Borges et al., 2016; Teixeira et al., 2020). Cells were seeded at a density of 1.0 × 105 (BeWo) and 6.0 × 104 (HTR8/SVneo) in 24-well plates containing 13-mm coverslips in each well. Cells were fixed with paraformaldehyde (4%) for 30 min. They were then washed three times with 1x phosphate buffered saline (PBS). Subsequently, T. gondii tachyzoites were preincubated for 1 h with oleoresin, hydroalcoholic extract or SDZ + PYR. Pretreated tachyzoites with 16 or 32 µg/ml of oleoresin or hydroalcoholic extract were used to adhere BeWo cells; while preincubated parasites with 8 or 16 µg/ml of oleoresin or hydroalcoholic extract were used to adhere HTR8/SVneo cells. SDZ + PYR (200 + 8 μg/mL for later adhesion in BeWo, or 100 + 4 μg/mL for later adhesion in HTR8/SVneo), or only medium, were used as controls. Pretreated tachyzoites by 1 h, as described, were resuspended in medium and incubated with fixed cells for 3 h. Subsequently, the coverslips were incubated overnight with mouse anti-T. gondii monoclonal primary antibody [SAG1/p30] (Abcam TP3 #ab8313) (diluted 1:500 in PGN-0.01% saponin solution). Coverslips were then rinsed three times with 1x PBS and incubated with Alexa Fluor 488 conjugated anti-mouse IgG (Invitrogen, USA #A11001) (diluted 1:500 in PGN-0.01% saponin solution), phalloidin conjugated with tetramethylrhodamine isothiocyanate (TRITC) (Sigma, P1951) (diluted 1:50 in PGN+saponin) and TOPRO-3 Iodide (Life Techonologies) (diluted 1:500 in PGN+saponin) for 1 h in the dark at room temperature to label tachyzoites, F-actin and nuclei, respectively. We mounted coverslips on glass slides and samples were analyzed by confocal fluorescence microscopy (Zeiss, LSM 510 Meta, Germany) with an inverted microscope (Zeiss Axiovert 200 M). We analyzed the following parameters: the number of BeWo and HTR8/SVneo cells with adhered tachyzoites and the total number of adhered tachyzoites per cell in a total of 20 randomly chosen fields. Two independent experiments were performed in three replicates.
After checking the adhesion of pretreated parasites on BeWo and HTR8/SVneo cells, we also investigated the intracellular replication and invasion of pretreated parasites with oleoresins or hydroalcoholic extract (Castanheira et al., 2015; Teixeira et al., 2020). Then, T. gondii tachyzoites were preincubated for 1 h at 37°C and 5% CO2 with oleoresin or ethanol extract (32 or 16 µg/mL for later infection in BeWo, or 16 or 8 µg/mL for later infection in HTR8/SVneo). In parallel, tachyzoites were also treated with SDZ + PYR (200 + 8 μg/mL for later infection in BeWo, or 100 + 4 μg/mL for later infection in HTR8/SVneo), or only medium. Subsequently, the pretreated parasites were added in previously adhered BeWo or HTR8/SVneo cells in 96-well plates. Finally, two different analyses were performed: (1) to verify the T. gondii invasion, both cells were maintained with pretreated tachyzoites for only 3 h; or (2) to verify parasite intracellular proliferation, the cell monolayers were washed after 3 h of infection, a fresh supplemented culture medium was added, and the culture maintained for another 24 h at 37°C and 5% CO2. In both experiments, intracellular T. gondii was quantified using the β-galactosidase assay, as described above (Silva et al., 2017; Souza et al., 2021). Three independent experiments were performed in eight replicates.
Toxicity of oleoresin and hydroalcoholic extract in human villous explants was performed using viability assays with lactate dehydrogenase (LDH) and MTT, according to previous protocols (Teixeira et al., 2020). In both assays, explants were treated with oleoresin or hydroalcoholic extract from C. multijuga at 64, 128 or 256 µg/mL or SDZ + PYR (150 + 200 µg/mL) (Silva et al., 2017). As viability control, explants were treated with culture medium alone. After 24 h of incubation, the culture supernatants were collected and LDH concentration measured according to the manufacturer’s instructions (Castro-Filice et al., 2014), with minor modifications (LDH Liquiform, Labtest Diagnostica S.A., Lagoa Santa, MG, Brazil). This assay is based on the consumption and decreased absorption of NADH at 340 nm, measured by a microplate reader (Versa Max ELISA Microplate Reader, Molecular Devices, Sunnyvale, CA, USA). LDH released in the culture the medium was expressed in U/L of LDH enzymatic activity and was used as a marker of tissue integrity. In parallel, in the same experimental condition, as described above, the viability of the tissue was also evaluated by the MTT test (Teixeira et al., 2020). Tissue viability was expressed in percentages (% viability by MTT incorporation), and the absorbance of villi incubated with culture medium alone (untreated villous explants) was considered to be 100% viable. In addition, we performed a morphological analysis of the treated explants. Tissue sections were included in paraffin, stained with hematoxylin/eosin and examined under a light microscope (BX40 Olympus, Tokyo, Japan). Two independent experiments were performed in six replicates.
Also, we quantified the T. gondii intracellular proliferation in explants treated with oleoresin or hydroalcoholic extract from C. multijuga using the β-galactosidase colorimetric assay (Silva et al., 2017; Souza et al., 2021), with minor modifications. Explants were cultured in 96-well microplates (one villus per well/200 µl) in supplemented culture medium for 24 h at 37°C and 5% CO2. Then, the villi were infected with T. gondii tachyzoites (1 × 106 parasites per well/200 µL) and incubated for 24 h at 37°C and 5% CO2. Subsequently, the villous explants were thoroughly rinsed with culture medium to remove non-adherent parasites. Following our tissue viability data (LDH and MTT assays), the villi were treated for additional 24 h with oleoresin or hydroalcoholic extract (256 and 128 µg/mL), or SDZ + PYR (150 + 200 µg/mL). Uninfected and untreated or infected and untreated explants were cultured with culture medium alone. Finally, the culture supernatants were collected and stored at -80°C for future cytokine measurement. In addition, the villous explants were collected and stored at −80°C for the following analyses: determination of protein content using Bradford reagent and T. gondii intracellular proliferation by β-galactosidase assay. Initially, frozen villous explants were homogenized by the addition of 150 µL of radioimmunoprecipitation assay buffer (RIPA) [50 mM Tris-HCl, 150 mM NaCl, 1% Triton X-100, 1% (w/v) sodium deoxycholate and 0.1% (w/v) sodium deoxycholate SDS, pH 7.5] containing protease inhibitor cocktail (Complete, Roche Diagnostic, Mannheim, Germany). Homogenate was centrifuged at 21,000 × g for 15 min at 4°C, and the supernatant was collected to measure the total amount of protein using the Bradford method (Bradford, 1976). T. gondii intracellular proliferation was carried out with 20 µL of supernatants from each sample (Teixeira et al., 2020), absorbance measured at 570 nm using a kinetic plate reader (Versa Max ELISA Microplate Reader, Molecular Devices, Sunnyvale, CA, USA), the number of tachyzoites was normalized according to total protein concentration (µg/mL) of each villus obtained by Bradford assay, and expressed by the number of parasites per µg of tissue. The data were presented as a percentage (% of T. gondii proliferation), as described above. Four independent experiments were performed in six replicates.
We measured cytokines in supernatants using a double-antibody sandwich enzyme-linked immunosorbent assay (ELISA). ELISA for IL-6, IL-8, TNF, IL-10, and MIF were purchased from OpTEIA, BD Bioscience, San Diego, CA, USA; or Duoset R&D Systems, Minneapolis, MN, USA, and tests were performed according to the manufacturer’s instructions. The detection limits of each cytokine were 4.7 pg/mL for IL-6; 31.2 pg/mL for IL-8; 7.8 pg/mL for TNF, IL-10 and MIF.
For villous explants, the data were normalized according to the total protein of each villous. Then, the data about cytokines were obtained by the ratio between concentration of cytokines (pg/mL) and concentration of total protein from Bradford assay (µg/mL), resulting in pg/mg of tissue (Silva et al., 2017; Souza et al., 2021).
The influence of oleoresin and hydroalcoholic extract in ROS production was also verified in BeWo and HTR8/SVneo cells. The assay was based on the peroxide-dependent oxidation of 2’,7’-dichlorodihydrofluorescein diacetate (H2DCF-DA) to form the fluorescent compound 2’,7’-dichlorofluorescein (DCF), with some modifications. Briefly, BeWo (3×104 cells/well/200 µL) or HTR8/SVneo (1.5×104 cells/well/200 µL) cells were seeded in 96-well black clear-bottom plate. Then, cells were infected or not with T. gondii tachyzoites (3:1) for 3 h; they were then washed abundantly with culture medium and treated with oleoresin or hydroalcoholic extract (16 or 32 µg/mL) for BeWo and (8 or 16 µg/mL) for HTR8/SVneo cells, or both cells received SDZ + PYR cells (200 + 8 μg/mL for BeWo, and 100 + 4 μg/mL for HTR8/SVneo) or only medium, as controls. Hydrogen peroxide (H2O2) was used as a positive control for ROS production. After treatment for 24 h, cells were harvested, washed with 1×PBS and incubated with 150 μL of H2DCF-DA (10 μM; diluted in 1×PBS containing 10% FBS) for 45 min at 37°C and 5% CO2 in the dark. Finally, the DCF fluorescence intensity was detected using a microplate reader (GloMax® Discover System, Promega Corporation) at excitation and emission wavelengths of 488 nm and 522 nm, respectively. The data are presented as median fluorescence intensity (MFI). Three independent experiments were performed in eight replicates.
The present research protocol using human tissue samples was approved by the Ethics Committee of the Federal University of Uberlandia, MG, Brazil, with approval number 3.679.426. A consent term was obtained from all subjects and/or their legal guardians.
All data were expressed as means ± standard deviations (SD) using GraphPad Prisma version 8.01. Significant differences were compared to controls by using One-way ANOVA, Bonferroni’s multiple comparisons post-test for the parametric data. Nonparametric data were analyzed by the Kruskal–Wallis test and Tukey’s multiple comparison post-test. Data were considered statistically significant when P < 0.05.
Firstly, we evaluated the cell viability in BeWo and HTR8/SVneo cells treated with hydroalcoholic extract or oleoresin in several concentrations.
Reduced cell viability was detected in BeWo cells exposed to hydroalcoholic extract or oleoresin from 64 to 512 µg/mL (*P < 0.05, ****P < 0.00001) in comparison to untreated cells (medium) (Figures 1A, B). DMSO-treated cells did not show any change in cell viability in relation to untreated cells (Figures 1A, B). On the other hand, HTR8/SVneo diminished cell viability when treated with hydroalcoholic extract or oleoresin from 32 (***P < 0.0001, **P < 0.001) to 512 µg/mL (****P < 0.00001) when compared to untreated cells (medium) (Figures 1C, D). As observed for BeWo, DMSO also did not induce change in cell viability for HTR8/SVneo (Figures 1C, D).
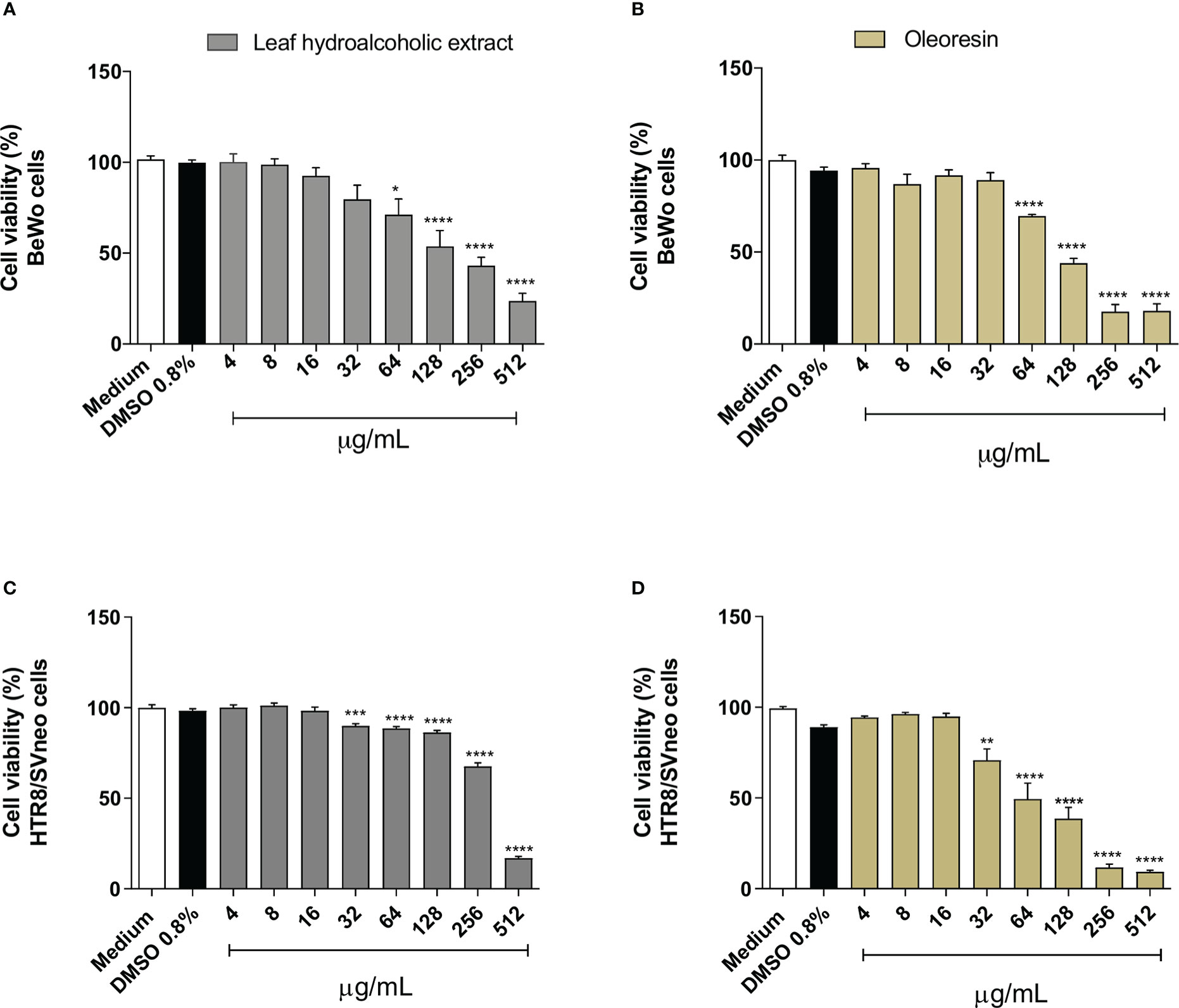
Figure 1 Cell viability. BeWo (A, B) and HTR8/SVneo (C, D) cells were treated or not with several concentrations of oleoresin or hydroalcoholic extract from C. multijuga for 24 h, and MTT assay was performed. Cell viability was expressed as percentage (cell viability %), considering the absorbance of cells incubated only with the medium as 100% viability. The results are expressed as means ± standard deviation of three independent experiments performed with eight replicates. Significant differences detected by One-Way ANOVA with Bonferroni’s multiple comparison post-test, or Kruskal-Wallis when appropriate. *P < 0.05, **P < 0.001, ***P < 0.0001, and ****P < 0.00001 in relation to the medium.
In our cell viability assay, we established concentrations of hydroalcoholic extract and oleoresin that did not alter the viability of BeWo and HTR8/SVneo cells for the subsequent set up of experiments.
Both hydroalcoholic extract and oleoresin reduced significantly the parasite proliferation in BeWo (Figure 2A) and HTR8/SVneo (Figure 2B) cells in all concentrations tested in comparison to untreated cells (medium) (***P < 0.0001; *****P < 0.00001). In addition, sulphadiazine and pyrimethamine (SDZ + PYR) also diminished T. gondii replication in relation to untreated cells (****P < 0.00001). These results demonstrated that both the hydroalcoholic extract and oleoresin had an antiproliferative effect against T. gondii.
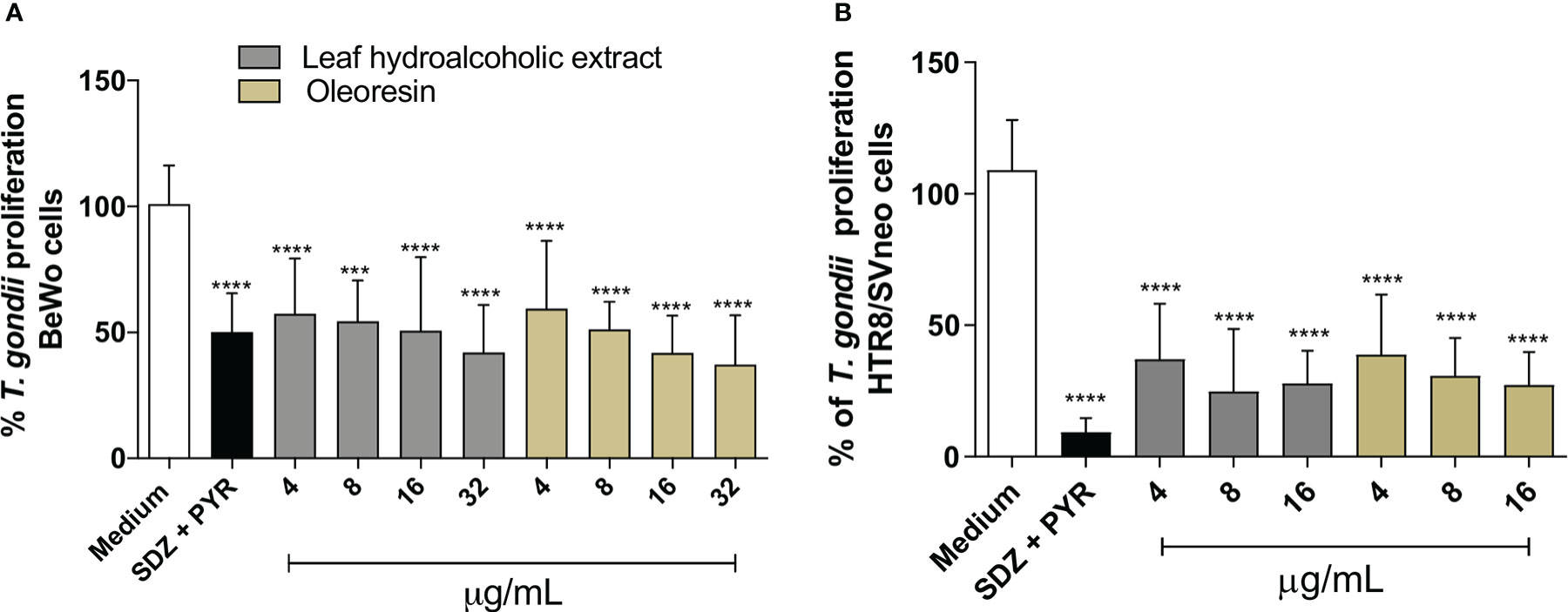
Figure 2 T. gondii intracellular proliferation. BeWo (A) and HTR8/SVneo (B) cells were infected by T. gondii and treated or not with non-toxic concentrations of hydroalcoholic extract or oleoresin from C. multijuga for 24 h. Untreated and infected cells (medium) were considered as 100% parasite proliferation, and SDZ + PYR was used as positive control. T. gondii intracellular proliferation was analyzed using a β-galactosidase colorimetric assay and expressed as percentage (% of T. gondii proliferation) in comparison to untreated cells (medium). The results were expressed as means ± standard deviation of three experiments performed in eight replicates. Significant differences detected by One-Way ANOVA, Bonferroni’s multiple comparisons post-test. ***P < 0.0001 or ****P < 0.00001 in relation to the medium.
In order to determine whether the antiparasitic effects promoted by the hydroalcoholic extract and oleoresin from C. multijuga would be irreversible, we exposed infected BeWo and HTR8/SVneo cells to the hydroalcoholic extract or oleoresin for 24 h, then cell monolayers were rinsed and incubated with medium free of treatment for additional 24 h. At the same time, we quantified the parasite proliferation after 24 h of treatment as a baseline for comparison. For BeWo cells, we choose 16 and 32 µg/mL for both the hydroalcoholic extract and oleoresin; for HTR8/SVneo, 8 and 16 µg/mL were the choice.
As previously detected (Figure 2), again both the hydroalcoholic extract and oleoresin reduced T. gondii intracellular proliferation after 24 h of treatment in BeWo (Figure 3A) and HTR8/SVneo cells (Figure 3B) when compared to untreated cells (medium) (**P < 0.001, ****P < 0.00001). Also, SDZ + PYR decreased parasite replication in relation to untreated cells (****P < 0.00001) (Figures 3A, B). Interestingly, both the hydroalcoholic extract and oleoresin showed an irreversible effect on tachyzoites, since a reduced parasite growth remained active in BeWo and HTR8/SVneo cells in comparison to untreated cells (considered as 100% reversibility), even after removal of treatments for additional 24 h (****P < 0.00001) (Figures 3A, B). For both cells, SDZ + PYR also showed irreversible effect (****P < 0.00001) (Figures 3A, B). These results suggest that both the hydroalcoholic extract and oleoresin maintained their antiproliferative effect even after the elimination of the treatment.
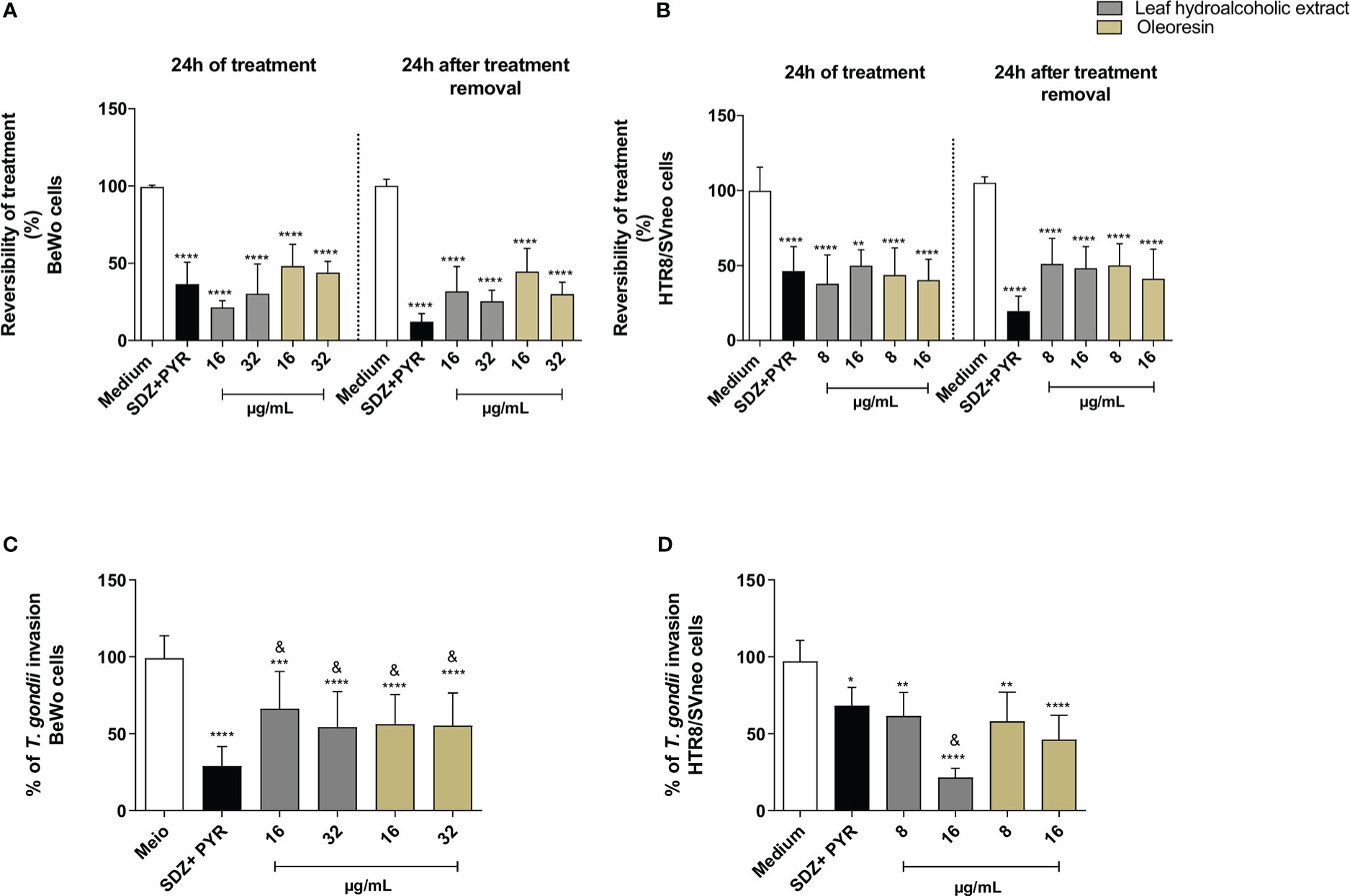
Figure 3 Reversibility test. BeWo (A) and HTR8/SVneo (B) cells were infected by T. gondii and treated or not with non-toxic concentrations of the hydroalcoholic extract or oleoresin from C. multijuga for 24 h. In parallel, both cells were infected, treated, washed to remove the treatments, and maintained for additional 24 h in the medium free of treatment. Untreated and infected cells (medium) were considered as 100% reversibility of treatment, and SDZ + PYR was used as positive control. T. gondii intracellular proliferation was analyzed using a β-galactosidase colorimetric assay and expressed as percentage (reversibility of treatment) in comparison to untreated cells (medium). The reversibility index measured the ability of parasites to recover from treatment and proliferate in new cells. Finally, parasites obtained directly from BeWo (C) and HTR8/SVneo (D) cells treated with the hydroalcoholic extract or oleoresin were collected and used to infect new cells for 3 h. The number of tachyzoites was determined using the β-galactosidase assay and expressed as % of T. gondii invasion. The results were expressed as means ± standard deviation of three experiments performed in eight replicates. Significant differences detected by the One-Way ANOVA, Bonferroni’s multiple comparisons post-test. *P < 0.05, **P < 0.001, ***P < 0.0001 or ****P < 0.00001 in relation to the medium. &P < 0.05 in relation to SDZ + PYR.
In order to obtain more information about the effect triggered by both the hydroalcoholic extract and oleoresin on T. gondii tachyzoites, we evaluated whether intracellular parasites obtained directly from treated-BeWo and HTR8/SVneo cells could maintain their ability to infect new host cells. For this purpose, tachyzoites were collected from treated-BeWo and HTR8/SVneo cells and used to infect new monolayer cells. For BeWo cells, all concentrations of hydroalcoholic extract and oleoresin reduced the ability of tachyzoites to infect new BeWo cells when compared to untreated cells (medium) (***P < 0.0001; ****P < 0.00001) (Figure 3C). SDZ + PYR also reduced the ability of parasites to infect new BeWo cells in relation to medium (****P < 0.00001) (Figure 3C). Although the hydroalcoholic extract and oleoresin have downmodulated the ability of parasites to infect new BeWo cells, this effect was lower if compared to classical treatment (SDZ + PYR) (&P < 0.05) (Figure 3C). Finally, for HTR8/SVneo cells, all treatments (hydroalcoholic extract, oleoresin and SDZ + PYR) dampened the capacity of T. gondii to infect new HTR8/SVneo cells in comparison to untreated cells (*P < 0.05; **P < 0.001; ****P < 0.00001) (Figure 3D). Interestingly, parasites derived from hydroalcoholic extract-treated-cells at 16 µg/mL demonstrated lower ability to infect new cells when compared to SDZ + PYR (&P < 0.05) (Figure 3D).
These findings indicate that the inhibitory effects promoted by the hydroalcoholic extract and oleoresin from C. multijuga dampen the ability of parasites to invade new host cells.
To assess whether hydroalcoholic extract and oleoresin have a direct action on tachyzoites, we performed a variety of assays. In the first set of experiments, we evaluated whether the hydroalcoholic extract and oleoresin treatments would affect parasite adhesion to host cells. For this purpose, T. gondii tachyzoites were pretreated for 1 h with oleoresin or hydroalcoholic extract and then incubated with previously fixed-BeWo or HTR8/SVneo cells.
As result for BeWo cells, only the hydroalcoholic extract or oleoresin at 32 µg/mL was able to reduce the number of cells with adhered parasites when compared to untreated parasites (medium) (***P < 0.0001, *P < 0.05) (Figure 4A). On the other hand, all concentrations of treatments, except for SDZ + PYR, reduced the total number of adhered parasites in relation to untreated parasites (****P < 0.00001, **P < 0.001) and SDZ + PYR-treated parasites (&P < 0.05) (Figure 4B). Representative images showing the effect of the hydroalcoholic extract and oleoresin on BeWo cells incubated with pretreated-tachyzoites can be observed in Figures 4C-F.

Figure 4 Adhesion assay in BeWo cells. T. gondii tachyzoites were preincubated for 1 h with the hydroalcoholic extract or oleoresin from C. multijuga, and then allowed to interact with previously fixed BeWo cells during 3 h. As control, tachyzoites were incubated with SDZ + PYR or only medium (medium). The number of cells with adhered parasites (A) and the total number of adhered parasites (B) were determined in a total of 20 fields examined randomly with 20X objective lens. Representative images highlighting the effect of treatments on the tachyzoite-host cell interaction: (C) untreated parasites, (D) 32 µg/mL hydroalcoholic extract, (E) 16 µg/mL oleoresin and (F) 32 µg/mL oleoresin. The results were expressed as means ± standard deviation of two experiment performed in three replicates. Significant differences detected by One-Way ANOVA and Bonferroni’s multiple comparisons post-test. *P < 0.05, **P < 0.001, ***P < 0.0001 or ****P < 0.00001 in relation to the medium. &P < 0.05 in relation to SDZ + PYR. White arrows indicate tachyzoites attached to BeWo cells. The cell nucleus is labeled with TOPRO-3 (blue). T. gondii tachyzoites labeled with Alexa Fluor 488-conjugated anti-mouse IgG (green). Phalloidin-TRITC labeled F-actin is shown in red. Scale bar: 50 µm.
Similarly, parasites pretreated with the hydroalcoholic extract or oleoresin diminished their ability to adhere in HTR8/SVneo cells, since the number of cells with adhered parasites as well as the total number of adhered parasites per field were significantly smaller when related to untreated parasites (medium) (****P < 0.00001) and SDZ + PYR-treated parasites (&P < 0.05) (Figure 5A, B). Also, SDZ + PYR reduced the number of cells with adhered parasites (***P < 0.0001) and the total number of adhered parasites (**P < 0.001) in relation to the medium (Figure 5A, B). Representative images showing the effect of the hydroalcoholic extract and oleoresin on HTR8/SVneo cells incubated with pretreated-tachyzoites can be observed in Figures 5C-F.
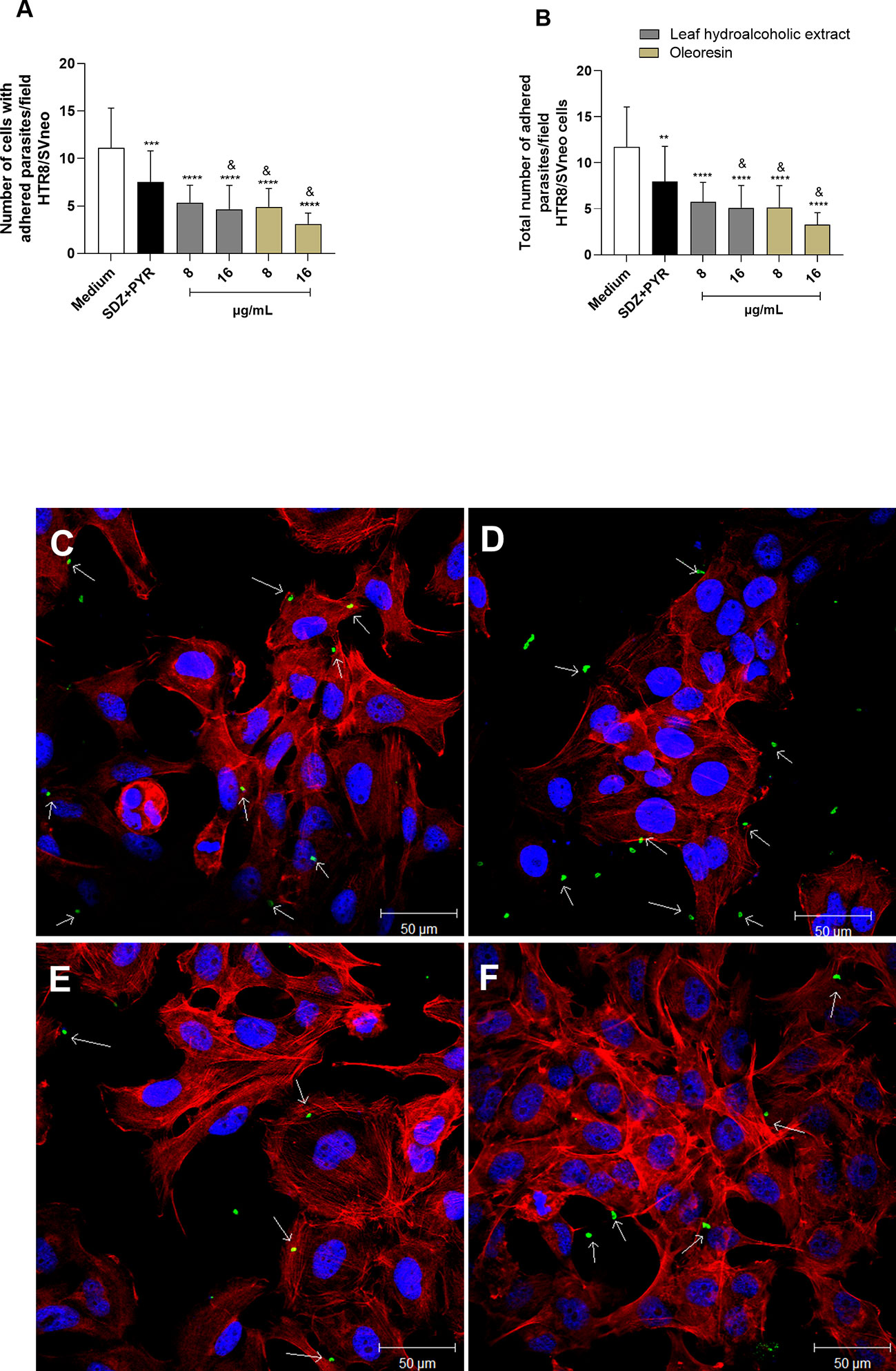
Figure 5 Adhesion assay in HTR8/SVneo cells. T. gondii tachyzoites were preincubated for 1 h with the hydroalcoholic extract or oleoresin from C. multijuga, and then allowed to interact with previously fixed HTR8/SVneo cells during 3 h. As control, tachyzoites were incubated with SDZ + PYR or only medium (medium). The number of cells with adhered parasites (A) and the total number of adhered parasites (B) were determined in a total of 20 fields examined randomly with 20X objective lens. Representative images highlighting the effect of the treatments on the tachyzoite-host cell interaction: (C) untreated parasites, (D) 16 µg/mL hydroalcoholic extract, (E) 8 µg/mL oleoresin, and (F) 16 µg/mL oleoresin. The results were expressed as means ± standard deviation of two experiment performed in three replicates. Significant differences detected by One-Way ANOVA and Bonferroni’s multiple comparisons post-test. **P < 0.001, ***P < 0.0001 or ****P < 0.00001 in relation to the medium. &P < 0.05 in relation to SDZ + PYR. White arrows indicate tachyzoites attached to HTR8/SVneo cells. The cell nucleus is labeled with TOPRO-3 (blue). T. gondii tachyzoites labeled with Alexa Fluor 488-conjugated anti-mouse IgG (green). Phalloidin-TRITC labeled F-actin is shown in red. Scale bar: 50 µm.
Considering that pretreated parasites had lower ability to adhere to host cells (Figures 4, 5), we now verified the capacity to invade and proliferate when also preincubated with the hydroalcoholic extract or oleoresin. Our data demonstrated that the pretreatment of T. gondii tachyzoites with all the hydroalcoholic extract or oleoresin concentrations triggered low rates of invasion and intracellular proliferation, regardless of the cell type (***P < 0.0001, ****P < 0.00001), in comparison to untreated parasites (medium) (Figures 6A-D). SDZ + PYR also reduced the invasion and replication of pretreated tachyzoites in BeWo and HTR8/SVneo cells in relation to the medium (**P < 0.001, ***P < 0.0001, ****P < 0.00001) (Figures 6A-D).
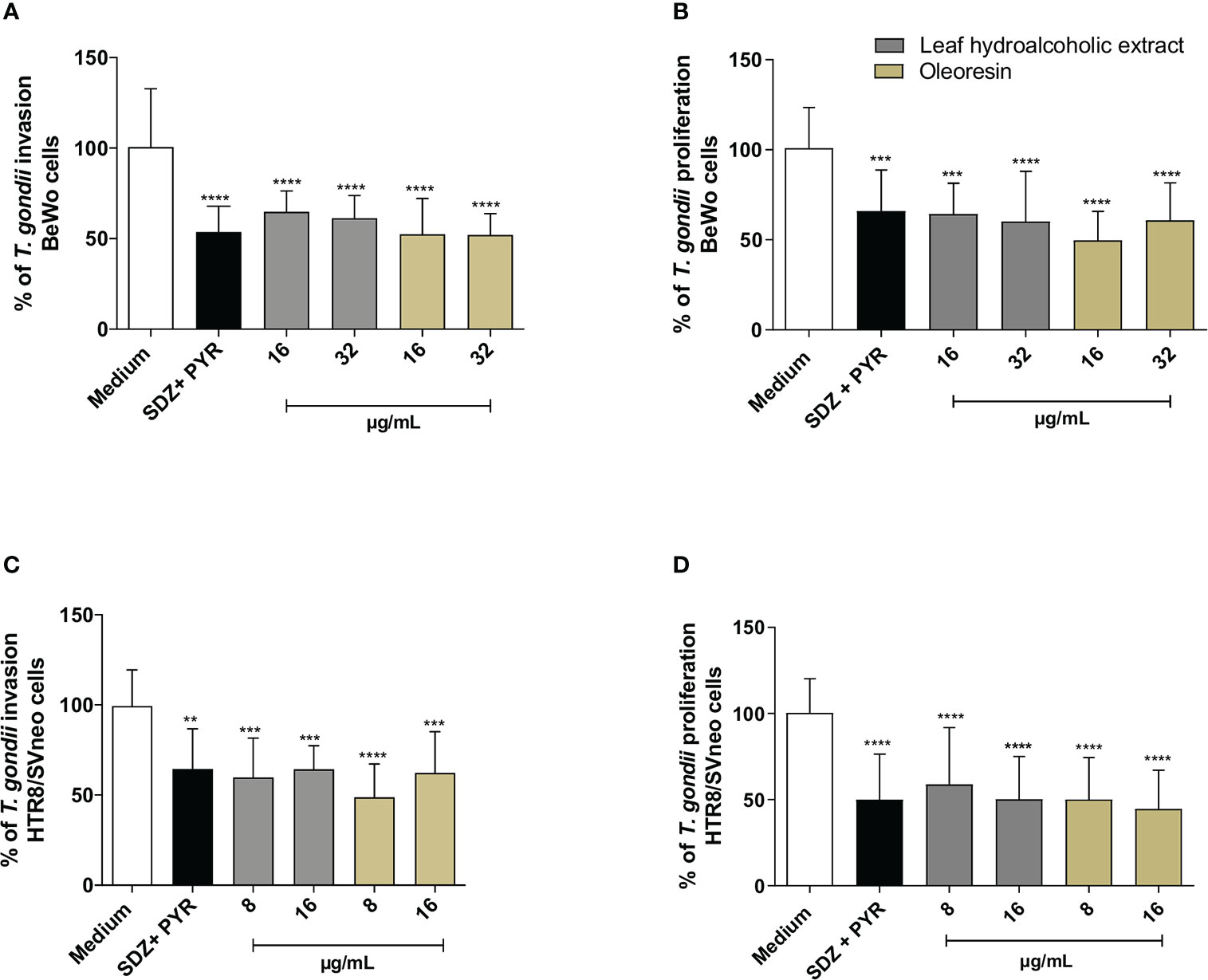
Figure 6 Invasion and proliferation assay. T. gondii tachyzoites were preincubated for 1 h with the hydroalcoholic extract or oleoresin from C. multijuga, and then allowed to interact with BeWo (A, B) or HTR8/SVneo (C, D) cells during 3 or 24 h to verify invasion and intracellular proliferation, respectively. As control, tachyzoites were incubated with SDZ + PYR or only medium (medium). The % of T. gondii invasion and the % of T. gondii proliferation were determined using the β-galactosidase activity. Untreated parasites (medium) were considered as 100% invasion and proliferation. The results were expressed as means ± standard deviation of three experiments performed in eight replicates. Significant differences detected by One-Way ANOVA and Bonferroni’s multiple comparisons post-test. **P < 0.001, ***P < 0.0001 or ****P < 0.00001 in relation to the medium.
These findings indicate that the hydroalcoholic extract and oleoresin from C. multijuga dampen the ability of parasites to adhere, invade and proliferate in host cells from maternal-fetal interface.
So far, our findings demonstrated that the hydroalcoholic extract and oleoresin from C. multijuga have a direct effect on the parasites, since treated parasites have reduced ability to adhere, invade and replicate into the cells. However, we could not exclude the possibility that these compounds may affect the host cell environment. Thus, we investigated the possible immunomodulatory effects of the hydroalcoholic extract and oleoresin by measuring cytokines in culture supernatant.
For BeWo cells, our results showed that in the absence of infection, concentrations of 32 µg/mL of the hydroalcoholic extract and oleoresin induced an increase in IL-6 levels in comparison to untreated cells (medium) (*P < 0.05, **P < 0.001) and SDZ + PYR (&P < 0.05) (Figure 7A). After T. gondii infection, levels of IL-6 were higher in untreated/infected cells (medium Tg) compared to untreated and uninfected cells (medium) (#P < 0.05). However, SDZ + PYR reduced IL-6 in relation to untreated/infected cells (**P < 0.001). In addition, all concentrations of the hydroalcoholic extract or oleoresin induced an upregulation of IL-6 in infected BeWo cells when compared to SDZ + PYR-treated cells (&P < 0.05). Also, 32 µg/mL of oleoresin induced an increase of IL-6 in comparison to untreated/infected cells (**P < 0.001) (Figure 7B).
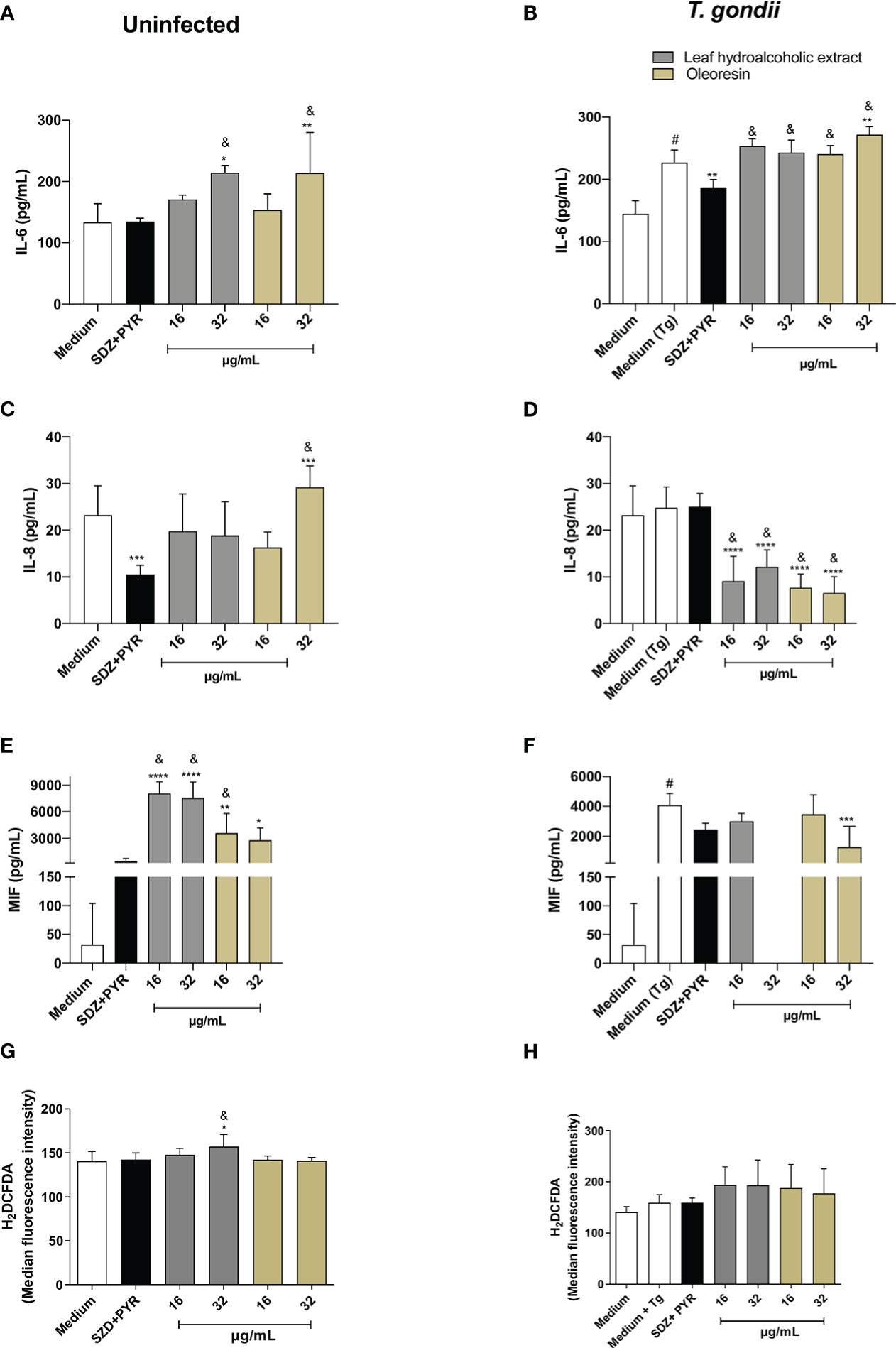
Figure 7 Cytokine and ROS production in BeWo cells. BeWo cells were infected and treated or not with the hydroalcoholic extract or oleoresin from C. multijuga for 24 h. Untreated and uninfected cells (medium), untreated and infected cells (medium Tg) and SDZ + PYR were used as controls. Then, supernatants were collected and used to measure IL-6 (A, B), IL-8 (C, D) and MIF (E, F). In parallel, BeWo cells were infected, treated or not with the hydroalcoholic extract or oleoresin from C. multijuga for 24 h, incubated with the probe 2′,7′-dichlorodihydrofluorescein diacetate (H2DCF-DA), ROS production was measured by a plate reader and data expressed as mean fluorescence intensity (MFI) (G, H). The results were expressed as means ± standard deviation of three experiments performed in eight replicates. Significant differences detected by One-Way ANOVA, Bonferroni’s multiple comparisons post-test. *P < 0.01, **P < 0.001, ***P < 0.0001 or ****P < 0.00001 in relation to medium (for uninfected cells) or to medium Tg (for infected cells). &P < 0.05 in relation to SDZ + PYR (for uninfected or infected cells). #P < 0.05 between medium and medium Tg.
Regarding IL-8, uninfected BeWo cells reduced or increased the level of this cytokine when treated with SDZ + PYR or 32 µg/mL, respectively (***P < 0.0001) in relation to untreated cells (medium) (Figure 7C). Interestingly, 32 µg/mL oleoresin triggered a higher release of IL-8 in comparison to SDZ + PYR-treated cells (&P < 0.05) (Figure 7C). However, all concentrations of the hydroalcoholic extract and oleoresin downmodulated IL-8 in infected BeWo cells in comparison to untreated/infected cells (medium Tg) (****P < 0.00001) or SDZ + PYR-treated cells (&P < 0.05) (Figure 7D).
For MIF production, uninfected BeWo cells augmented the release of this cytokine in all concentrations of the hydroalcoholic extract and oleoresin when compared to untreated cells (medium) (*P < 0.01, **P < 0.001, ****P < 0.00001) or SDZ + PYR-treated cells (&P < 0.05) (Figure 7E). However, in the presence of infection, untreated and 32 µg/mL oleoresin-treated cells upregulated or downmodulated MIF in comparison to medium (#P < 0.05) or medium Tg (***P < 0.0001), respectively (Figure 7F).
Finally, we measured ROS in BeWo cells infected or not with T. gondii and treated or not with the hydroalcoholic extract or oleoresin. We just observed that uninfected cells treated with 32 µg/mL of the hydroalcoholic extract augmented the level of ROS in relation to untreated cells (medium) (*P < 0.01) and SDZ + PYR-treated cells (&P < 0.05) (Figure 7G). Additionally, infected BeWo cells did not change ROS release in presence of any treatment (Figure 7H).
TNF and IL-10 were not detected in BeWo cell supernatants under any experimental conditions (data not shown).
We also verified cytokine and ROS production in HTR8/SVneo cells infected or not with T. gondii and treated with the hydroalcoholic extract or oleoresin.
In the absence of infection, HTR8/SVneo cells increased IL-6 release when treated with the hydroalcoholic extract (8 µg/mL) or oleoresin (16 µg/mL) in comparison to untreated cells (medium) (*P < 0.01, **P < 0.001) and SDZ + PYR-treated cells (&P < 0.05) (Figure 8A). In the presence of infection, untreated HTR8/SVneo cells, as well as cells treated with the hydroalcoholic extract or oleoresin, increased the levels of IL-6 in relation to the medium or SDZ + PYR-treated cells, respectively (#P < 0.05, &P < 0.05) (Figure 8B). Also, IL-6 was lower in SDZ + PYR-treated cells in comparison to untreated and infected cells (medium Tg) (**P < 0.001) (Figure 8B).
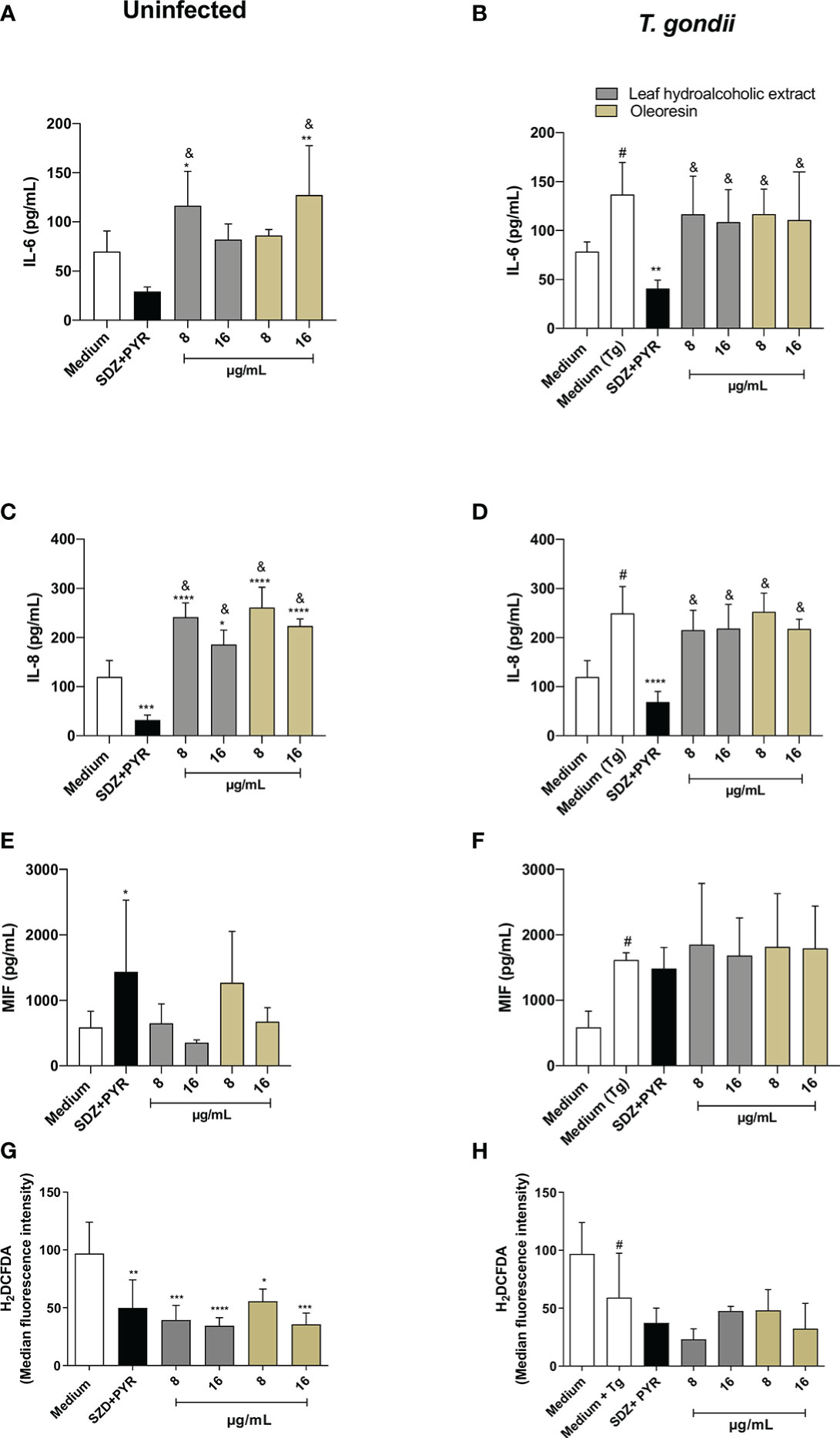
Figure 8 Cytokine and ROS production in HTR8/SVneo cells. HTR8/SVneo cells were infected and treated or not with the hydroalcoholic extract or oleoresin from C. multijuga for 24 h. Untreated and uninfected cells (medium), untreated and infected cells (medium Tg) and SDZ + PYR were used as controls. Then, supernatants were collected and used to measure IL-6 (A, B), IL-8 (C, D) and MIF (E, F). In parallel, HTR8/SVneo cells were infected, treated or not with the hydroalcoholic extract or oleoresin from C. multijuga for 24 h, incubated with the probe 2′,7′-dichlorodihydrofluorescein diacetate (H2DCF-DA), ROS production was measured by a plate reader and data were expressed as mean fluorescence intensity (MFI) (G, H). The results were expressed as means ± standard deviation of three experiments performed in eight replicates. Significant differences detected by One-Way ANOVA, Bonferroni’s multiple comparisons post-test. *P < 0.01, **P < 0.001, ***P < 0.0001 or ****P < 0.00001 in relation to the medium (for uninfected cells) or to the medium Tg (for infected cells). &P < 0.05 in relation to SDZ + PYR (for uninfected or infected cells). #P < 0.05 between medium and medium Tg.
Regarding IL-8 for uninfected cells, SDZ + PYR reduced IL-8 (***P < 0.0001) and all concentrations of the hydroalcoholic extract and oleoresin (*P < 0.01, ****P < 0.00001) increased IL-8 when compared to the medium (Figure 8C). Also, the hydroalcoholic extract and oleoresin presented higher levels of IL-8 in comparison to SDZ + PYR (&P < 0.05) (Figure 8C). When HTR8/SVneo cells were infected by T. gondii, IL-8 was augmented in relation to the medium (#P < 0.05), SDZ + PYR decreased IL-8 in comparison to only infected cells (medium Tg) (****P < 0.00001), and all treatments with the hydroalcoholic extract or oleoresin upregulated IL-8 in relation to SDZ + PYR (&P < 0.05) (Figure 8D).
The MIF production was not significantly changed in uninfected HTR8/SVneo cells, except for uninfected and SDZ + PYR-treated cells (*P < 0.01) (Figure 8E). Although not statistically different, the hydroalcoholic extract and oleoresin showed an increase in MIF release in infected HTR8/SVneo cells, while only infected cells augmented MIF in comparison to untreated and uninfected cells (#P < 0.05) (Figure 8F).
Finally, we measured ROS in HTR8/SVneo cells infected or not with T. gondii and treated or not with the hydroalcoholic extract or oleoresin. We observed that uninfected cells treated with the hydroalcoholic extract, oleoresin or SDZ + PYR presented low levels of ROS in relation to untreated cells (medium) (*P < 0.01, **P < 0.001, ***P < 0.0001, ****P < 0.00001) (Figure 8G). Although not statistically different, the hydroalcoholic extract and oleoresin showed a reduction in ROS release in infected HTR8/SVneo cells, while only infected cells diminished ROS in comparison to untreated and uninfected cells (#P < 0.05) (Figure 8H).
TNF and IL-10 were not detected in supernatants under any experimental conditions (data not shown).
First, to determine non-toxic concentration to use in the experiments, the villous explant viability after treatments with oleoresin or hydroalcoholic extract was performed by measuring LDH release and MTT assay. Our data showed that both oleoresin and hydroalcoholic extract did not alter tissue viability at any of the concentrations tested when compared to untreated explants (medium). Also, treatment with SDZ + PYR did not cause significant cytotoxicity in villous explants in relation to the untreated group (Figures 9A, B). In addition, treatments did not alter the tissue morphological structure, which was highlighted by the typical morphology of syncytiotrophoblast cells (black arrows) and mesenchyme (*) compared to untreated villous explants (Figures 9C-F).
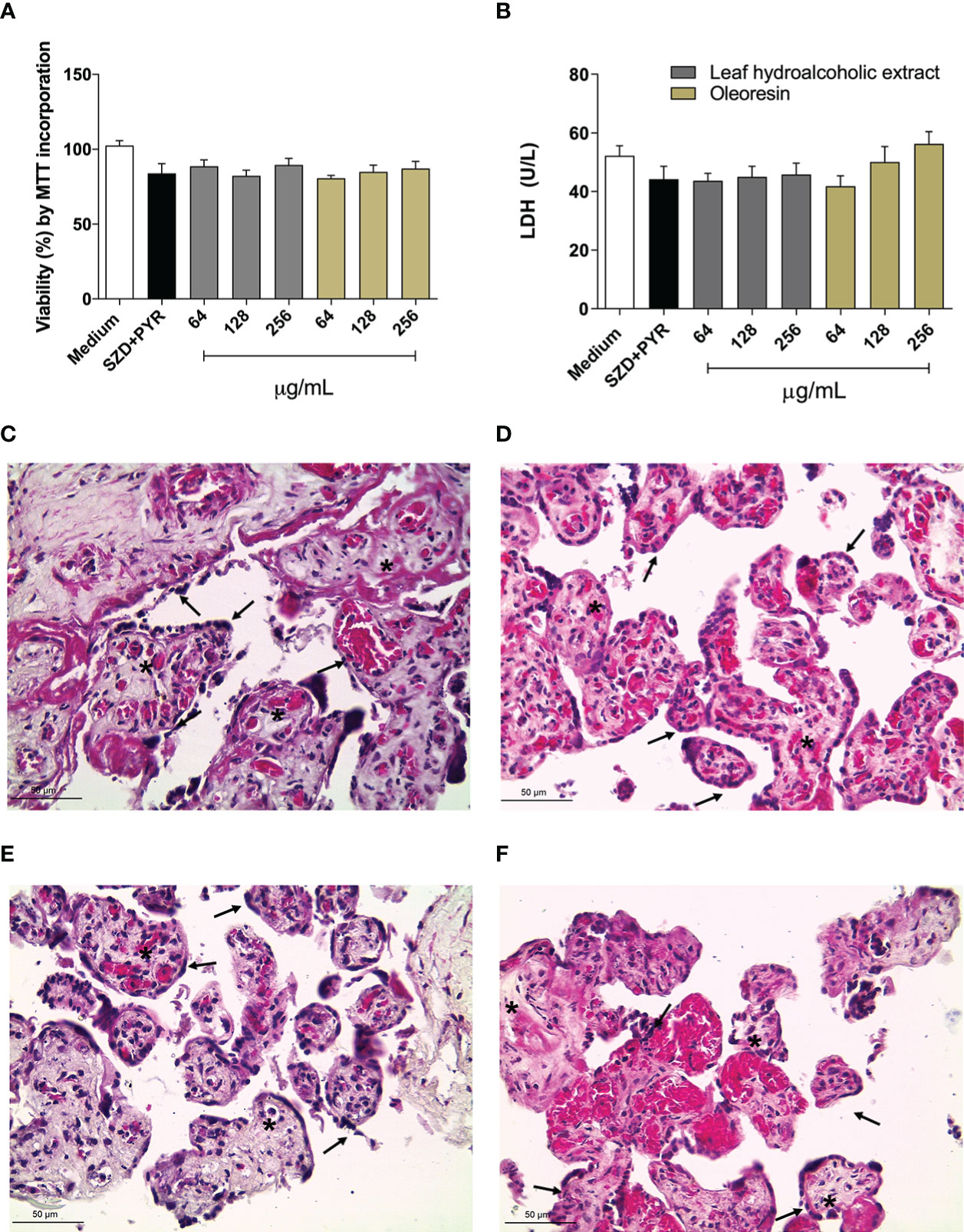
Figure 9 Viability of human villous explants. The villous explants were treated for 24 h with hydroalcoholic extract or oleoresin from C. multijuga, SDZ+PYR or culture medium alone (medium). (A) Tissue viability is shown in percentages (% viability by incorporation of MTT). (B) Supernatants were collected and used to measure LDH levels (U/L). Representative images of villi incubated with (C) culture medium alone, (D) SDZ+PYR, 256 µg/mL hydroalcoholic extract (E), 256 µg/mL oleoresin (F). Data are expressed as means ± standard deviation of two experiments performed in six replicates. Significant differences detected by One-way ANOVA, Bonferroni’s multiple comparisons post-test (statistically significant when P < 0.05). Hematoxylin-eosin (HE) stained histological sections show syncytiotrophoblast cells (black arrows) and mesenchyme (*). Scale bar: 50 µm.
We evaluated the effects of the hydroalcoholic extract and oleoresin (256 and 128 µg/mL) on the control of T. gondii intracellular proliferation in human chorionic villi using the β-galactosidase assay. We observed that treatments at both concentrations significantly reduced the percentage of intracellular parasites in comparison to untreated/infected explants (considered as 100% proliferation) (****P < 0.00001). As expected, treatment with SDZ + PYR also inhibited parasite growth (****P < 0.00001) when compared to the untreated group (medium) (Figure 10).
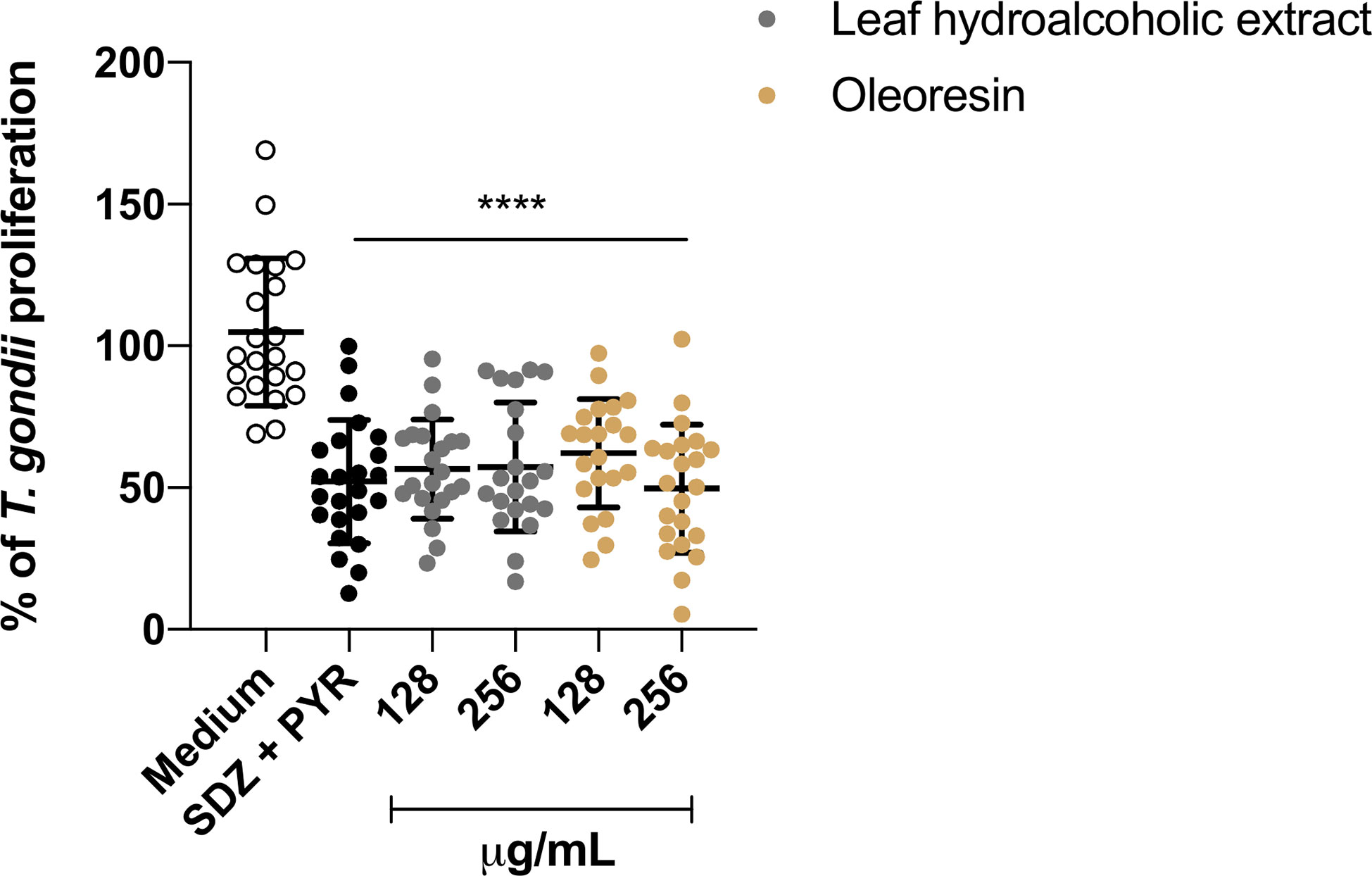
Figure 10 T. gondii intracellular proliferation. The human villous explants were infected with T. gondii followed by treatment for an additional 24 h with hydroalcoholic extract, oleoresin, SDZ+PYR or culture medium alone (medium). T. gondii intracellular proliferation was measured by β-galactosidase assay, and the number of tachyzoites was expressed as a percentage (% of T. gondii proliferation), and the untreated/infected (medium) was considered as 100% parasite proliferation. Data are expressed as means ± standard deviation of four experiments performed in six replicates. *Comparison with infected/untreated cells (****P < 0.00001). Significant differences detected by One-way ANOVA, Bonferroni’s multiple comparisons post-test differences were considered when P < 0.05.
In addition, we evaluated whether the hydroalcoholic extract and oleoresin would modulate the cytokine profile in chorionic villi. Our results showed that IL-6, IL-8, MIF and TNF showed no significant change in their production when uninfected explants were treated with both hydroalcoholic extract or oleoresin if compared to medium, except for TNF when 256 µg/mL hydroalcoholic extract was added in the samples (*P < 0.05) (Supplementary Figure 1). Finally, T. gondii infection reduced the levels of all cytokines in comparison to untreated explants (medium), regardless of hydroalcoholic extract and oleoresin treatments (#P < 0.05) (Supplementary Figure 1).
Toxoplasmosis is considered as a global health problem with nearly a billion people infected, reaching a prevalence of the disease in 71.5% (Ahmadpour et al., 2019). Regarding the treatment, a combination of sulfadiazine and pyrimethamine (SDZ + PYR) is the first choice to treat congenital toxoplasmosis, one of the most serious forms of the disease (Abugri et al., 2019). Despite the great clinical importance, the conventional treatment for congenital toxoplasmosis is currently limited, and it is also linked to serious side effects in both the mother and the child (Montoya and Remington, 2008; Halonen and Weiss, 2013). In this sense, the search for alternative therapeutic tools has gained attention.
In order to assess the impact of both the leaf hydroalcoholic extract and oleoresin from C. multijuga in the T. gondii infection, we used BeWo and HTR8/SVneo cells as villous and extravillous trophoblast cell models, respectively, being considered as excellent experimental models for the study of congenital toxoplasmosis in vitro (Barbosa et al., 2014; Souza et al., 2021; Teixeira et al., 2021). Firstly, the viability assay was performed. Next, the parasite replication revealed that both compounds efficiently inhibited the T. gondii intracellular proliferation and triggered an irreversible effect on tachyzoites.
In agreement with our data, studies have already demonstrated that oleoresins from Copaifera present activity against some pathogens. C. pubiflora and its compound (ent-hardwickiic acid) showed antibacterial properties and should be explored as new therapeutic alternatives to treat oral diseases such as dental caries and endodontic infections (Moraes et al., 2020). Pieri (2012) determined that copalic acid and β-caryophyllene are common components obtained from oleoresins and suggested that the bioactivity of the Copaifera oleoresin on Trypanosoma cruzi replication is due to this synergism. Additionally, terpenes isolated from Copaifera oleoresins promoted changes in oxidative metabolism, induced autophagy and decreased the replicative forms of T. cruzi, in particular amastigotes (Izumi et al., 2012). Izumi et al. (2013) tested oleoresins of eight species of Copaifera against T. cruzi, and oleoresin from C. martii and C. officinalis exhibited the best activity against replicative forms of the parasite. A more recent study demonstrated that oleoresin and kaurenoic acid from C. martii decreased T. cruzi amastigote proliferation in murine peritoneal macrophages and HeLa cells (Kian et al., 2018). In addition, murine peritoneal macrophages treated with C. reticulata oil controlled significantly the infection by Leishmania amazonensis (Santos et al., 2008), while C. martii oil promoted a reduction on the size lesion caused by L. amazonensis in BALB/c (Santos et al., 2011). Similarly, the treatment with nanoemulsion from Copaifera led to a reduction in L. infantum and L. amazonensis infection levels in macrophage cultures, as well as presented beneficial effects on the lesion size and parasite burden in BALB/c (De Moraes et al., 2018). Also, human fibroblast treated with oleoresin from C. reticulata showed activity against the Plasmodium falciparum W2 and 3D7 strains and reduced the parasitemia in BALB/c (Souza et al., 2017). Finally, our previous study showed that oleoresins from C. reticulata, C. duckei, C. paupera and C. pubiflora induced a significant reduction on T. gondii infection in BeWo cells (Teixeira et al., 2020). Until the present moment, no study was conducted to verify the effects of hydroalcoholic extracts from Copaifera against T. gondii, systemically or in maternal-fetal interface; and also there is no study about the effect of the hydroalcoholic extract and oleoresin from C. multijuga on T. gondii infection. Thus, the present study is the first to show these effects.
It is important to emphasize that HTR8/SVneo cells are experimental model of human extravillous trophoblast cells (Souza et al., 2021). This type of trophoblast is important to invade the myometrium in order to interpose between the endothelial cells and guarantee the adequate blood supply to the embryo (Aplin et al., 2020). Our previous study demonstrated that extravillous trophoblasts are more susceptible to T. gondii infection in comparison to villous trophoblast (Oliveira et al., 2021). Thus, it is possible to conclude that extravillous trophoblast cells are a potential entry for tachyzoites gain the embryonic and placental tissues, maybe the major transmission route. In the present study, we verified that oleoresin and hydroalcoholic extract from C. multijuga reduced significantly the infection in HTR8/SVneo cells. It means that these compounds are great tools to control the vertical transmission of T. gondii when it occurs by the extravillous trophoblast route. One more time, the present study is the first to show these effects.
T. gondii is an obligate intracellular protozoan parasite capable of infecting a wide variety of nucleated cells (Dubey and Jones, 2008). Therefore, the successful establishment of the infection requires the ability of the parasite to adhere, invade and proliferate within the host cell (Hall et al., 2011; Adeyemi et al., 2017). Therefore, the next question that we would like to determine was the direct action of the hydroalcoholic extract and oleoresin from C. multijuga on tachyzoites. Pretreated parasite demonstrated a lower capacity to adhere, invade and proliferate in both cells. These findings are in agreement with our previous study, in which preincubated tachyzoites with oleoresins from C. reticulata, C. duckei, C. paupera and C. pubiflora showed lower rates of adhesion, invasion and proliferation in BeWo cells (Teixeira et al., 2020). However, other studies have already demonstrated that parasites pretreated with other drugs also showed reduced ability to infection. Castro-Filice et al. (2014) showed that the pretreatment of T. gondii with a combination of SDZ + PYR + folinic acid or azithromycin reduced the parasite proliferation after 72 h in human villous explants. Similarly, tachyzoites from atypical strains of T. gondii pretreated with azithromycin or spiramycin had a lower capacity to invade and replicate in BeWo cells (Ribeiro et al., 2017). Thus, the pretreatment with oleoresin and hydroalcoholic extract from C. multijuga suggested that these compounds can directly affect the parasite structure and, consequently, decrease its functional capabilities. A recent study declared that these natural compounds can affect different structures in T. cruzi and Leishmania spp, and it is hypothesized that the mitochondrion is a strategic target to induce parasite death (Lazarin-Bidóia et al., 2022). Also, L. infantum and L. amazonensis promastigotes treated with nanoemulsion from Copaifera presented ultrastructural alterations as oval cell shape and retracted flagella (De Moraes et al., 2018). Our previous study demonstrated that C. paupera, C. reticulata, C. pubiflora and C. duckei induced tethered T. gondii tachyzoites, which dampen the cytokinesis (Teixeira et al., 2020). Future studies are necessary to investigate the types of structural changes that occur in T. gondii when treated with these compounds from C. multijuga.
In a second moment, after demonstrating the effect of C. multijuga on tachyzoites replication, as well as adhesion and invasion, we investigated the immune response of host cells, suggesting that these compounds could be involved in the modulation of the host’s cellular environment.
It was observed that BeWo cells upregulated IL-6 and downmodulated IL-8. It is widely known that IL-6 is an important cytokine involved in T. gondii control in various cell types, such as macrophages and monocytes (Pereira et al., 2019) and human and murine trophoblasts (Barbosa et al., 2014; Barbosa et al., 2015; Gomes et al., 2018). Thus, it is plausible to suggest that the higher IL-6 in BeWo cells infected and treated with hydroalcoholic extract or oleoresin was important to control T. gondii replication. Although MIF is a cytokine important during T. gondii infection (Silva et al., 2017), the level of this cytokine increased only in uninfected treated-BeWo cells or oleoresin treated-cell at 32 µg/mL, then it seems that MIF did not contribute to parasite control when oleoresin or hydroalcoholic extract are used in BeWo cells. IL-8 has emerged as a cytokine important to disseminate T. gondii infection (Sommerville et al., 2013). Our previous study showed low levels of IL-8 in THP-1 cells infected by T. gondii and inhibited for cyclooxygenase 2 (COX-2), which contributes to the control of parasite replication (Pereira et al., 2019). In this sense, it is possible to hypothesize that the low levels of IL-8 in BeWo cells treated with the hydroalcoholic extract and oleoresin from C. multijuga could have contributed to T. gondii control. Future studies are necessary to verify the influence of Copaifera on COX-2 modulation. Therefore, in BeWo cells, the T. gondii control mediated by the hydroalcoholic extract and oleoresin from C. multijuga was an association between the direct action of these compounds on tachyzoites, as well as the immune response of these cells. For HTR8/SVneo cells, cytokines were slightly modulated (not statistically significant) when compared to untreated/infected cells. Thus, it is possible to conclude that cytokines did not contribute to T. gondii control in HTR8/SVneo cells when treated with compounds from C. multijuga. Thus, in this case, the direct action of these compounds on tachyzoites was the only mechanism to downmodulate infection in these cell types. As HTR8/SVneo cells are representative of human extravillous trophoblast (Souza et al., 2021), a potent route for congenital toxoplasmosis, and both oleoresin and hydroalcoholic extract controlled T. gondii infection in these cells without change significantly the cytokine profile, it is possible to conclude that both compounds from C. multijuga are an excellent alternative strategy to prevent vertical transmission and maintain the immune profile required for a successful pregnancy. Finally, the ROS production was not significant in infected and treated BeWo and HTR8/SVneo cells.
Finally, we investigated the effect of oleoresin and hydroalcoholic extract in human villous explants from third-trimester pregnancy, and we also observed reduced T. gondii intracellular proliferation, even with no change in cytokine profile, suggesting a direct effect of both compounds on tachyzoites. One more time, both compounds from C. multijuga showed to be an excellent alternative strategy to prevent vertical transmission and maintain the immune profile required for a successful pregnancy. Cell cultures are notorious examples of in vitro studies. However, the placental explants are very important to validate the data obtained with cell lineages, since these cells can be very different from the natural environment. Then, in studies of maternal-fetal interface, the association between in vitro (cells) and ex vivo (explants) studies (Rosini et al., 2022) is mandatory, and we demonstrated that both oleoresin and hydroalcoholic extract are enough to control T. gondii infection in both trophoblast cells and villous explants.
Based on the results of the study, the leaf hydroalcoholic extract and oleoresin obtained from C. multijuga presented different antiparasitic activities in a dependent manner of the experimental model. For BeWo, representative of human villous trophoblast, C. multijuga presented a direct action in tachyzoite and modulated the immune response, whereas only the direct effect in tachyzoites participated of the parasite control in HTR8/SVneo cells, representative of human extravillous trophoblast, and human explants. Considering all these parameters, the leaf hydroalcoholic extract and oleoresin from C. multijuga can be a target for the establishment of new therapeutic strategies for congenital toxoplasmosis. However, more detailed studies are needed to evaluate the bioactive components and other possible mechanisms of these compounds.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
The studies involving human participants were reviewed and approved by Ethics Committee of the Federal University of Uberlandia, MG, Brazil. Approval number 3.679.426. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
ST and BB design the experiments. AM, ST, GS, AR. JJ, GM, and BB performed the experiments. ST, KB, AM, and BB analyzed the data. SA, RV, JB, and CM were responsible for the identification and collection of Copaifaera spp. oleoresin and hydroalcoholic extract. BB, ST, AG, CM, and EF, participated in the data interpretation. AM and BB discussed the findings. ST, GS, and BB, reviewed the manuscript. All authors approved the final version of the manuscript.
This work was supported by Brazilian Researcher Agencies: Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG, APQ-01629-22), and Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP, # 2017/02858-3). This study was financed in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brazil (CAPES) – Finance Code 001.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2023.1113896/full#supplementary-material
Supplementary Figure 1 | Cytokine production in villous explants. Explants were infected and treated or not with the hydroalcoholic extract or oleoresin from C. multijuga for 24 h. Untreated and uninfected explants (medium), untreated and infected explants (medium Tg) and SDZ + PYR were used as controls. Then, supernatants were collected and used to measure IL-6 (A, B), IL-8 (C, D), MIF (E, F) and TNF (G, H). The results were expressed as means ± standard deviation of four experiments performed in six replicates. Significant differences detected by One-Way ANOVA, Bonferroni’s multiple comparisons post-test. *P < 0.05 in relation to medium (for uninfected cells). #P < 0.05 between medium and medium Tg.
Abugri, D. A., Jaynes, J. M., Witola, W. H. (2019). Anti-Toxoplasma activity of sorghum bicolor-derived lipophilic fractions. BMC Res. Notes 12 (1), 688. doi: 10.1186/s13104-019-4732-z
Adeyemi, O. S., Murata, Y., Sugi, T., Kato, K. (2017). Inorganic nanoparticles kill Toxoplasma gondii via changes in redox status and mitochondrial membrane potential. Int. J. Nanomed. 12, 1647–1661. doi: 10.2147/IJN.S122178
Ahmadpour, E., Zargami, E., Mahami-Oskouei, M., Spotin, A., Shahbazi, A., Kafil, H. S., et al. (2019). Diagnosis of Toxoplasma gondii infection in pregnant women using automated chemiluminescence and quantitative real time PCR. Asian Pac. J. Trop. Med. 12 (1), 26–31. doi: 10.4103/1995-7645.250341
Aplin, J. D., Myers, J. E., Timms, K., Westwood, M. (2020). Tracking placental development in health and disease. Nat. Rev. Endocrinol. 16, 479–494. doi: 10.1038/s41574-020-0372-6
Arruda, C., Mejía, J. A. A., Ribeiro, V. P., Borges, C. H. G., Martins, C. H. G., Veneziani, R. C. S., et al. (2019). Occurrence, chemical composition, biological activities and analytical methods on Copaifera genus - a review. Biomed. Pharmacother. 109, 1–20. doi: 10.1016/j.biopha.2018.10.030
Barbosa, B. F., Lopes-Maria, J. B., Gomes, A. O., Angeloni, M. B., Castro, A. S., Franco, P. S., et al. (2015). IL10, TGF Beta1, and IFN gamma modulate intracellular signaling pathways and cytokine production to control Toxoplasma gondii infection in BeWo trophoblast cells. Biol. Reprod. 92 (3), 82. doi: 10.1095/biolreprod.114.124115
Barbosa, B. F., Paulesu, L., Ietta, F., Bechi, N., Romagnoli, R., Gomes, A. O., et al. (2014). Susceptibility to Toxoplasma gondii proliferation in BeWo human trophoblast cells is dose-dependent of macrophage migration inhibitory factor (MIF), via ERK1/2 phosphorylation and prostaglandin E2 production. Placenta 35 (3), 152–162. doi: 10.1016/j.placenta.2013.12.013
Blader, I. J., Coleman, B. I., Chen, C. T., Gubbels, M. J. (2015). Lytic cycle of Toxoplasma gondii 15 years later. Annu. Rev. Microbiol. 69, 463–485. doi: 10.1146/annurev-micro-091014-104100
Borges, I. P., Castanheira, L. E., Barbosa, B. F., de Souza, D. L. N., da Silva, R. J., Mineo, J. R., et al. (2016). Anti-parasitic effect on Toxoplasma gondii induced by BnSP-7, a Lys49-phospholipase A2 homologue from Bothrops pauloensis venom. Toxicon 119, 84–91. doi: 10.1016/j.toxicon.2016.05.010
Bradford, M. M. (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72, 248–254. doi: 10.1006/abio.1976.9999
Buzoni-Gatel, D., Schulthess, J., Menard, L. C., Kasper, L. H. (2006). Mucosal defences against orally acquired protozoan parasites, emphasis on Toxoplasma gondii infections. Cell. Microbiol. 8 (4), 535–544. doi: 10.1111/j.1462-5822.2006.00692.x
Carlier, Y., Truyens, C., Deloron, P., Peyron, F. (2012). Congenital parasitic infections: A review. Acta Trop. 121 (2), 55–70. doi: 10.1016/j.actatropica.2011.10.018
Carruthers, V. B. (2002). Host cell invasion by the opportunistic pathogen. Toxoplasma gondii. Acta Trop. 1 (2), 111–122. doi: 10.1016/S0001-706X(01)00201-7
Cascon, V., Gilbert, B. (2000). Characterization of the chemical composition of oleoresins of Copaifera guianensis desf., Copaifera duckei dwyer and Copaifera multijuga hayne. Phytochemistry 55 (7), 773–778. doi: 10.1016/s0031-9422(00)00284-3
Castanheira, L., de Souza, D. L. N., Silva, R. J., Barbosa, B. F., Mineo, J. R., Tudini, K. A., et al. (2015). Insights into anti-parasitism induced by a c-type lectin from Bothrops pauloensis venom on toxoplasma gondii. Int. J. Biol. Macromol. 74, 568–574. doi: 10.1016/j.ijbiomac.2014.11.035
Castro, A. S., Alves, C. M. O. S., Angeloni, M. B., Gomes, A. O., Barbosa, B. F., Franco, P. S., et al. (2013). Trophoblast cells are able to regulate monocyte activity to control Toxoplasma gondii infection. Placenta 34 (3), 240–247. doi: 10.1016/j.placenta.2012.12.006
Castro-Filice, L. S., Barbosa, B. F., Angeloni, M. B., Silva, N. M., Gomes, A. O., Alves, C. M. O. S., et al. (2014). Azithromycin is able to control Toxoplasma gondii infection in human villous explants. J. Transl. Med. 12, 132. doi: 10.1186/1479-5876-12-132
Costa, I. N., Angeloni, M. B., Santana, L. A., Barbosa, B. F., Silva, M. C. P., Rodrigues, A. A., et al. (2009). Azithromycin inhibits vertical transmission of Toxoplasma gondii in Calomys callosus (Rodentia: Cricetidae). Placenta 30 (10), 884–890. doi: 10.1016/j.placenta.2009.08.002
De Moraes, A. R. D. P., Tavares, G. D., Rocha, F. J. S., de Paula, E., Giorgio, S. (2018). Effects of nanoemulsions prepared with essential oils of copaiba- and andiroba against Leishmania infantum and Leishmania amazonensis infections. Exp. Parasitol. 187, 12–21. doi: 10.1016/j.exppara.2018.03.005
Doliwa, C., Escotte-Binet, S., Aubert, D., Sauvage, V., Velard, F., Schmid, A., et al. (2013). Sulfadiazine resistance in Toxoplasma gondii : no involvement of overexpression or polymorphisms in genes of therapeutic targets and ABC transporters. Parasite 20, 19. doi: 10.1051/parasite/2013020
Dong, R., Zheng, S. (2015). Interleukin-8: A critical chemokine in biliary atresia. J. Gastroenterol. Hepatol. 30 (6), 970–976. doi: 10.1111/jgh.12900
Dubey, J., Jones, J. (2008). Toxoplasma gondii infection in humans and animals in the united states. Int. J. Parasitol. 38 (11), 1257–1278. doi: 10.1016/j.ijpara.2008.03.007
Elsheikha, H. M. (2008). Congenital toxoplasmosis: Priorities for further health promotion action. Public Health 122 (4), 335–353. doi: 10.1016/j.puhe.2007.08.009
Fitch, W., MacKenzie, E. T., Harper, A. M. (1975). Effects of decreasing arterial blood pressure on cerebral blood flow in the baboon. influence of the sympathetic nervous system. Circ. Res. 37 (5), 550–557. doi: 10.1161/01.res.37.5.550
Fowler, N. O., McCall, D., Chou, T. C., Holmes, J. C., Hanenson, I. B. (1976). Electrocardiographic changes and cardiac arrhythmias in patients receiving psychotropic drugs. Am. J. Cardiol. 37 (2), 223–230. doi: 10.1016/0002-9149(76)90316-7
Gomes, A. O., Barbosa, B. F., Franco, P. S., Ribeiro, M., Silva, R. J., Gois, P. S. G., et al. (2018). Macrophage migration inhibitory factor (MIF) prevents maternal death, but contributes to poor fetal outcome during congenital toxoplasmosis. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.00906
Habib, F. A. (2008). Post-treatment assessment of acute Toxoplasma infection during pregnancy. J. Obstet. Gynaecol. 28 (6), 593–595. doi: 10.1080/01443610802344332
Hall, C. I., Reese, M. L., Weerapana, E., Child, M. A., Bowyer, P. W., Albrow, V. E., et al. (2011). Chemical genetic screen identifies Toxoplasma DJ-1 as a regulator of parasite secretion, attachment, and invasion. Proc. Natl. Acad. Sci. U.S.A. 108 (26), 10568–10573. doi: 10.1073/pnas.1105622108
Halonen, S. K., Weiss, L. M. (2013). Toxoplasmosis. Handb. Clin. Neurol. 114, 125–145. doi: 10.1016/B978-0-444-53490-3.00008-X
Huang, S. Y., Yao, N., He, J. K., Pan, M., Hou, Z. F., Fan, Y. M., et al. (2021). In vitro anti-parasitic activity of Pelargonium x. asperum essential oil against. Toxoplasma gondii. Front. Cell. Dev. Biol. 9. doi: 10.3389/fcell.2021.616340
Izumi, E., Ueda-Nakamura, T., Veiga-Júnior, V. F., Nakamura, C. V. (2013). Toxicity of oleoresins from the genus Copaifera in Trypanosoma cruzi: A comparative study. Planta. Med. 79 (11), 952–958. doi: 10.1055/s-0032-1328712
Izumi, E., Ueda-Nakamura, T., Veiga-Júnior, V. F., Pinto, A. C., Nakamura, C. V. (2012). Terpenes from Copaifera demonstrated in vitro antiparasitic and synergic activity. J. Med. Chem. 55 (7), 2994–3001. doi: 10.1021/jm201451h
Kian, D., Lancheros, C. A. C., Assolini, J. P., Arakawa, N. S., Veiga-Júnior, V. F., Nakamura, C. V., et al. (2018). Trypanocidal activity of copaiba oil and kaurenoic acid does not depend on macrophage killing machinery. Biomed. Pharmacother. 103, 1294–1301. doi: 10.1016/j.biopha.2018.04.164
Kodjikian, L. (2010). Toxoplasmose et grossesse. J. Français d’Ophtalmologie 33 (5), 362–367. doi: 10.1016/j.jfo.2010.03.002
Lazarin-Bidóia, D., Garcia, F. P., Ueda-Nakamura, T., Silva, S. O., Nakamura, C. V. (2022). Natural compounds based chemotherapeutic against chagas disease and leishmaniasis: mitochondrion as a strategic target. Mem. Inst. Oswaldo Cruz 117, e220396. doi: 10.1590/0074-02760220396
Miller, C. M., Boulter, N. R., Ikin, R. J., Smith, N. C. (2009). The immunobiology of the innate response to Toxoplasma gondii. Int. J. Parasitol. 39 (1), 23–39. doi: 10.1016/j.ijpara.2008.08.002
Miranda, N. C., Araujo, E. C. B., Justino, A. B., Cariaco, Y., Mota, C. M., Costa-Nascimento, L. A., et al. (2021). Anti-parasitic activity of Annona muricata l. leaf ethanolic extract and its fractions against Toxoplasma gondii in vitro and in vivo. J. Ethnopharmacol. 273, 114019. doi: 10.1016/j.jep.2021.114019
Montoya, J. G., Liesenfeld, O. (2004). Toxoplasmosis. Lancet 363 (9425), 1965–1976. doi: 10.1016/S0140-6736(04)16412-X
Montoya, J. G., Remington, J. S. (2008). Management of Toxoplasma gondii infection during pregnancy. Clin. Infect. Dis. 47 (4), 554–566. doi: 10.1086/590149
Moraes, T. S., Leandro, L. F., Santiago, M. B., Silva, L. O., Bianchi, T. C., Veneziani, R. C. S., et al. (2020). Assessment of the antibacterial, antivirulence, and action mechanism of Copaifera pubiflora oleoresin and isolated compounds against oral bacteria. Biomed. Pharmacother. 129, 110467. doi: 10.1016/j.biopha.2020.110467
Mosmann, T. (1983). Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 65 (1-2), 55–63. doi: 10.1016/0022-1759(83)90303-4
Oliveira, F. C., Silva, R. J., Ribeiro, M., Guirelli, P. M., Castro, A. S., Gomes, A. O., et al. (2021). ERK1/2 phosphorylation and IL-6 production are involved in the differential susceptibility to Toxoplasma gondii infection in three types of human (cyto/ syncytio/ extravillous) trophoblast cells. Tissue Cell 72, 101544. doi: 10.1016/j.tice.2021.101544
Oliveira, J. G., Silva, N. M., Santos, A. A. D., Souza, M. A., Ferreira, G. L. S., Mineo, J. R., et al. (2006). BeWo trophoblasts are unable to control replication of Toxoplasma gondii, even in the presence of exogenous IFN-γ. Placenta 27 (6-7), 691–698. doi: 10.1016/j.placenta.2005.06.006
Oz, H. S. (2014). Toxoplasmosis complications and novel therapeutic synergism combination of diclazuril plus atovaquone. Front. Microbiol. 5. doi: 10.3389/fmicb.2014.00484
Pereira, A. C. A., Silva, R. J., Franco, P. S., Gomes, A. O., Souza, G., Milian, I. C. B., et al. (2019). Cyclooxygenase (COX)-2 inhibitors reduce Toxoplasma gondii infection and upregulate the pro-inflammatory immune response in Calomys callosus rodents and human monocyte cell line. Front. Microbiol. 10. doi: 10.3389/fmicb.2019.00225
Peyron, F., Mc Leod, R., Ajzenberg, D., Contopoulos-Ioannidis, D., Kieffer, F., Mandelbrot, L., et al. (2017). Congenital toxoplasmosis in France and the united states: One parasite, two diverging approaches. PloS Negl. Trop. Dis. 11 (2), e0005222. doi: 10.1371/journal.pntd.0005222
Pieri, F. A. (2012). Synergistic activity between terpenes from Copaifera genus against the etiological agent of chagas disease. Rev. Virtual Quim. 4 (3), 343–346. doi: 10.5935/1984-6835.20120025
Pimenta, I. P., Abrão, F., da Silva, J. J. M., Oliveira, L. C., Rogez, H. L. G., Ambrósio, S. R., et al. (2019). Antibacterial profile of copaifera multijuga oleoresin and hydroalcoholic extract of leaves against oral pathogens. Curr. Dentistry 1 (1), 53–60. doi: 10.2174/2542579X01666180629100020
Pinheiro, J. G. O., Tavares, E. A., Silva, S. S., Silva, J. F., Carvalho, Y. M. B. G., Ferreira, M. R. A., et al. (2017). Inclusion complexes of copaiba (Copaifera multijuga hayne) oleoresin and cyclodextrins: Physicochemical characterization and anti-inflammatory activity. Int. J. Mol. Sci. 18 (11), 2388. doi: 10.3390/ijms18112388
Ribeiro, M., Franco, P. S., Lopes-Maria, J. B., Angeloni, M. B., Barbosa, B. F., Gomes, A. O., et al. (2017). Azithromycin treatment is able to control the infection by two genotypes of Toxoplasma gondii in human trophoblast BeWo cells. Exp. Parasitol. 181, 111–118. doi: 10.1016/j.exppara.2017.08.004
Rosini, A. M., Teixeira, S. C., Milian, I. C. B., Silva, R. J., de Souza, G., Luz, L. C., et al. (2022). LPS-mediated activation of TLR4 controls Toxoplasma gondii growth in human trophoblast cell (BeWo) and human villous explants in a dependent-manner of TRIF, MyD88, NF-κB and cytokines. Tissue Cell 78, 101907. doi: 10.1016/j.tice.2022.101907
Santos, A. O., Costa, M. A., Ueda-Nakamura, T., Dias-Filho, B. P., Veiga-Júnior, V. F., Lima, M. M. S., et al. (2011). Leishmania amazonensis: Effects of oral treatment with copaiba oil in mice. Exp. Parasitol. 129 (2), 145–151. doi: 10.1016/j.exppara.2011.06.016
Santos, A. O., Ueda-Nakamura, T., Dias Filho, B. P., Veiga-Júnior, V. F., Pinto, A. C., Nakamura, C. V. (2008). Effect of Brazilian copaiba oils on Leishmania amazonensis. J. Ethnopharmacol. 120 (2), 204–208. doi: 10.1016/j.jep.2008.08.007
Sharif, A. A., Unyah, N. Z., Nordin, N., Basir, R., Wana, M. N., Ahmad, A. A., et al. (2019). Susceptibility of Toxoplasma gondii to ethanolic extract of Tinospora crispa in vero cells. Evid. Based Complement. Alternat. Med. 2019, 2916547. doi: 10.1155/2019/2916547
Silva, R. J., Gomes, A. O., Franco, P. S., Pereira, A. S., Milian, I. C. B., Ribeiro, M., et al. (2017). Enrofloxacin and toltrazuril are able to reduce Toxoplasma gondii growth in human BeWo trophoblastic cells and villous explants from human third trimester pregnancy. Front. Cell. Infect. Microbiol. 7. doi: 10.3389/fcimb.2017.0034
Sommerville, C., Richardson, J. M., Williams, R. A. M., Mottram, J. C., Roberts, C. W., Alexander, J., et al. (2013). Biochemical and immunological characterization of Toxoplasma gondii macrophage migration inhibitory factor. J. Biol. Chem. 288 (18), 12733–12741. doi: 10.1074/jbc.M112.419911
Souza, G. A. G., da Silva, N. C., de Souza, J., de Oliveira, K. R. M., da Fonseca, A. L., Baratto, L. C., et al. (2017). In vitro and in vivo antimalarial potential of oleoresin obtained from Copaifera reticulata ducke (Fabaceae) in the Brazilian Amazon rainforest. Phytomedicine 24, 111–118. doi: 10.1016/j.phymed.2016.11.021
Souza, A. B., Martins, C. H. G., Souza, M. G. M., Furtado, N. A. J. C., Heleno, V. C. G., Sousa, J. P. B., et al. (2011). Antimicrobial activity of terpenoids from Copaifera langsdorffii desf. against cariogenic bacteria. Phytother. Res. 25, (2) 215–220. doi: 10.1002/ptr.3244
Souza, G., Silva, R. J., Milián, I. C. B., Rosini, A. M., de Araújo, T. E., Teixeira, S. C., et al. (2021). Cyclooxygenase (COX)-2 modulates Toxoplasma gondii infection, immune response and lipid droplets formation in human trophoblast cells and villous explants. Sci. Rep. 11 (1), 12709. doi: 10.1038/s41598-021-92120-3
Tamaru, S., Kikuchi, A., Takagi, K., Wakamatsu, M., Horikoshi, T., Ogiso, Y. (2011). Fetal therapy of severe symptomatic toxoplasmosis using azithromycin: Congenital toxoplasmosis. J. Obstet. Gynaecol. Res. 37 (7), 953–957. doi: 10.1111/j.1447-0756.2010.01459.x
Teixeira, S. C., de Souza, G., Borges, B. C., de Araújo, T. E., Rosini, A. M., Aguila, F. A., et al. (2020). Copaifera spp. oleoresins impair Toxoplasma gondii infection in both human trophoblastic cells and human placental explants. Sci. Rep. 10 (1), 15158. doi: 10.1038/s41598-020-72230-0
Teixeira, S. C., Silva, R. J., Lopes-Maria, J. B., Gomes, A. O., Angeloni, M. B., Fermino, M. L., et al. (2021). Transforming growth factor (TGF)-β1 and interferon (IFN)-γ differentially regulate ICAM-1 expression and adhesion of Toxoplasma gondii to human trophoblast (BeWo) and uterine cervical (HeLa) cells. Acta Trop. 224, 106111. doi: 10.1016/j.actatropica.2021.106111
Tomani, J. C. D., Bonnet, O., Nyirimigabo, A., Deschamps, W., Tchinda, A. T., Jansen, O., et al. (2021). In vitro antiplasmodial and cytotoxic activities of compounds from the roots of Eriosema montanum baker f. (Fabaceae). Molecules 26 (9), 1–9. doi: 10.3390/molecules26092795
Veiga-Júnior, V. F., Pinto, A. C. (2002). O Gênero Copaifera l. Quim. Nova 25 (2), 273–286. doi: 10.1590/S0100-40422002000200016
Villena, I., Aubert, D., Leroux, B., Dupouy, D., Talmud, M., Chemla, C., et al. (1998). Pyrimethamine-sulfadoxine treatment of congenital toxoplasmosis: follow-up of 78 cases between 1980 and 1997. Scand. J. Infect. Dis. 30 (3), 295–300. doi: 10.1080/00365549850160963
Keywords: Toxoplasma gondii, congenital toxoplasmosis (CT), trophoblast, Copaifera multijuga, hydroalcoholic extract, oleoresin
Citation: Martínez AFF, Teixeira SC, de Souza G, Rosini AM, Júnior JPdL, Melo GN, Blandón KOE, Gomes AO, Ambrósio SR, Veneziani RCS, Bastos JK, Martins CHG, Ferro EAV and Barbosa BF (2023) Leaf hydroalcoholic extract and oleoresin from Copaifera multijuga control Toxoplasma gondii infection in human trophoblast cells and placental explants from third-trimester pregnancy. Front. Cell. Infect. Microbiol. 13:1113896. doi: 10.3389/fcimb.2023.1113896
Received: 01 December 2022; Accepted: 30 January 2023;
Published: 13 February 2023.
Edited by:
Mariana De Niz, Universidade de Lisboa, PortugalReviewed by:
Paula Mello De Luca, Oswaldo Cruz Foundation (FIOCRUZ), BrazilCopyright © 2023 Martínez, Teixeira, de Souza, Rosini, Júnior, Melo, Blandón, Gomes, Ambrósio, Veneziani, Bastos, Martins, Ferro and Barbosa. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bellisa Freitas Barbosa, YmVsbGlzYWZiQHVmdS5icg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.