 Xueyu Hou
Xueyu Hou Na Wu
Na Wu Shimeng Ren
Shimeng Ren Xinjuan Wang2
Xinjuan Wang2 Weidong Yu
Weidong Yu- 1Department of Pediatrics, Peking University People’s Hospital, Beijing, China
- 2Department of Central Laboratory & Institute of Clinical Molecular Biology, Peking University People’s Hospital, Beijing, China
- 3Department of Pediatrics, The First Affiliated Hospital of Zhengzhou University, Zhengzhou, China
- 4Beijing Municipal Key Laboratory of Child Development and Nutriomics, Capital Institute of Pediatrics, Beijing, China
Introduction: Down syndrome (DS), the presence of a supernumerary chromosome 21, is associated with cognitive dysfunction caused by early neurodegenerative processes. Alterations in the gut microbiota were observed in Chinese children with DS, and the genus Blautia was associated with cognitive function in these children. Therefore, it is crucial to understand the detailed composition of this group at the species level and to explore the effect of specific species on cognitive function.
Methods: In this study, Blautia-specific amplicon sequencing was conducted to identify the specific Blautia species in 15 children with DS and 15 matched healthy children.
Results: The taxonomic analyses suggested that the Blautia taxa were clustered by disease status. The diversity of Blautia at the species level differed between DS patients and healthy controls, with the abundances of Blautia massiliensis and Blautia argi decreasing in DS children, while Blautia faecis was increased. Acetic acid, one of the metabolites of Blautia, was significantly reduced in the DS group. Of particular interest, Kyoto Encyclopaedia of Genes and Genomes analysis revealed decreased modules related to starch and sucrose metabolism and glycolysis. In addition, B. argi was positively related to DS cognitive scores, and B. faecis was negatively related to cognitive function, implying its role on the DS cognitive impairments.
Discussion: Our study has important implications for understanding the important effects of specific species of Blautia on cognitive function and thus possibly provides a new strategy for future studies of cognitive improvement in individuals with DS.
1 Introduction
Trisomy 21, the presence of a supernumerary chromosome 21, results in a collection of clinical features commonly known as Down syndrome (DS) (Antonarakis et al., 2020). According to the Global Burden of Disease Report, DS accounted for approximately 2930 live births in China in 2019 (The global burden of disease (GBD) 2019 data). Children with DS commonly present cognitive dysfunction (Grieco et al., 2015) and are more likely to develop certain health conditions, including intellectual disability and early-onset Alzheimer’s disease (AD). The cognitive profile of children with DS present with a distinct collection of symptoms, including cognitive dysfunction, bowel dysfunction and obesity (Squassante et al., 2015). Cognitive dysfunction greatly impairs patients’ quality of life and places heavy economic and mental pressure on their families and society.
The combined effects of genetic and environmental factors are thought to contribute to cognitive dysfunction. Recently, dysbiosis in gut microbiome composition has been suggested to be associated with neurodegenerative diseases, such as Alzheimer’s disease (AD) and Parkinson’s disease (PD). PD patients display distinct metabolic profiles in gut microbiota compared with those of healthy individuals, with alterations in metabolites associated with the gut microbiota, including reduced concentrations of short-chain fatty acids (SCFAs) (Unger et al., 2016; Ahmed et al., 2019). Alterations in gut microbiota composition have also been identified in individuals with AD, including a decreased abundance of Firmicutes, one of the core phyla of human gut microbiota (Biddle et al., 2013). As a genus within the Lachnospiraceae family in the Firmicutes phylum, Blautia has been of particular interest because of its contribution to metabolic diseases and for its antibacterial activity against specific microorganisms (Kalyana Chakravarthy et al., 2018). The Blautia genus has been found to have 25 subspecies (NCBI; Liu et al., 2021), which exhibit many differences in metabolic products and may play different roles in the disease process of neurodegenerative cognitive dysfunction (Fung et al., 2017). In our previous study, we observed the gut microbiota from DS patients showed different pattern compared with healthy controls. Besides, we identified that the Blautia genus was present at a lower relative abundance in the feces of children (Ren et al., 2022). As SCFA-producing bacteria, Blautia plays an important role in maintaining the environmental balance in the intestine and preventing inflammation. The microbiota-derived metabolite SCFAs were proven to have a protective role in cognitive functions Erny et al., 2021, and long-term acetic acid deficiency was suggested as a risk factor for cognitive decline (Zheng et al., 2021a), implying the potential of Blautia in cognitive function in individuals with DS (Chen et al., 2020).
However, most of these reports did not conduct in-depth studies at the species or even strain levels, and little is known about the detailed composition of this group in the gut microbiota of DS and healthy children. To avoid drawing general conclusions at the genus level, in this study, we used Blautia group-specific amplicon sequencing to investigate the composition of Blautia species in feces from the DS and healthy groups. Furthermore, a detailed correlation analysis was performed to provide important clinical clues based on the relative abundance of Blautia at the species level, levels of SCFAs and cognitive function in DS and healthy children.
2 Materials and methods
2.1 Ethical approval
The research scheme was approved by the Ethics Review Committee of Peking University People’s Hospital of China (Approval No:2019PHB110-01). All participants and their parents received oral and written information about the study and written consent was obtained before recruitment. All experiments were conducted in accordance with approved guidelines and regulations (Ren et al., 2022).
2.2 Participant recruitment
As described in our previous study (Ren et al., 2022), 15 children with DS were randomly recruited from integrated boarding schools in Beijing and its surrounding areas, and 15 age- and diet structure-matched children were recruited as the healthy group. Fifteen pairs of samples were collected for amplicon sequencing analysis. There were no differences in age and gender between DS group and healthy group. All the study participants had the same diet offered by school, which greatly reduces the impacts of diet, lifestyle and geography on gut microbiota profile. Besides, we excluded children who had received antibiotics within 3 months. Each participant’s height and weight were measured, and body mass index (BMI) was calculated by dividing weight by the square of height. The DS group had a lower average height (P = 0.037), a higher BMI (P = 0.009) than the healthy group. There was no difference in the weight between the two groups. Due to the limitation of the Chinese version of the Wechsler Intelligence Scale for Children (WISC-IV), only eleven pairs of participants over the age of 6 were subjected to the cognition test.
2.3 Assessments of cognitive function
To assess the participants’ cognitive function, we calculated the Full Scale IQ (FSIQ) test and measured the other four factor indices: the perceptual reasoning index (PRI), which was used to examine nonverbal fluid reasoning skills; the verbal comprehension index (VCI), which was used to measure verbal acquired knowledge and verbal reasoning; the working memory index (WMI), which was used to measure a person’s ability to manipulate linguistic information in short-term instant memory and then formulate responses; and the processing speed index (PSI), which was used to evaluate children’s ability to efficiently scan and understand visual information and use information to complete the work. Cognitive tests were conducted by two well-trained pediatricians in a hospital conference room with no interference. As described in our previous study (Ren et al., 2022), the scores of FSIQ, PRI, VCI, WMI and PSI were all decreased in the DS group (all P < 0.001). The correlation coefficient of this type is approximately 0.857.
2.4 Fecal sample collection and DNA extraction
Fecal bacterial DNA storage tubes (Longsee Biological Company, Guangzhou, China) were used to collect fecal samples. Before processing, samples are transferred and stored at − 80°C. The average moving time between sample collection and storage is 19 minutes at -80°C, and the maximum is 23 minutes. Total fecal DNA was extracted using a QIAamp DNA Stool Mini Kit (Qiagen, Hilden, Germany). The concentration of genomic DNA in each fecal sample was quantified using a NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, MA, USA). DNA integrity and sizes were assessed by 1% agarose gel electrophoresis. The DNA was resuspended in H2O and stored at −80°C prior to use.
The concentration detection results of the collected and extracted genomic DNA were all in line with the standards (Table S1). The purity, fragment size, and concentration of the amplified products of all samples met the sequencing requirements (Figure S1).
2.5 Quantitative PCR
Quantitative PCR (qPCR) amplifications were performed in a 20-μL reaction solution containing SYBR® Green Real-time PCR Master Mix (Toyobo, Code No. QPK-201, Osaka, Japan). The specific primers for Blautia sp. covers all species of Blautia (Figure S2). The cycling conditions were as follows: 95°C for 1 min and then 33 cycles of 95°C for 15 s, 56°C for 15 s, and 72°C for 45 s.
The relative abundance of Blautia was evaluated using the following formulae. First, the relative value (RV) of Blautia was determined by the 2−ΔΔct method with normalization to the total bacterial level. Then, the relative abundance (RA) of Blautia was scaled to a reasonable range with the following equation: RA = log2 (RV × 10000 + 1).
2.6 Blautia-specific amplicon sequencing
DNA extraction was performed for Blautia-specific PCR amplicon sequencing. Standard protocols were used to generate amplicons using a genus-specific primer from Blautia. Briefly, the hypervariable region of the 16S rRNA gene was amplified using a primer pair (477F: 5′-CGGTACCTGACTAAGAAGC-3′ and 719R: 5′-GTTCCTCCTAATATCTACGC-3′) with barcodes (Luu et al., 2017). All PCRs were performed with Phusion® High-Fidelity PCR Master Mix (New England Biolabs). After the purification of a Blautia-specific PCR amplicon, the libraries were generated with an Ion Plus Fragment Library kit, and paired-end sequencing (2 × 125 bp) was performed with an IonS5™XL platform (Thermo Fisher) from Novogene (Beijing, China).
2.7 Sequence analysis
These reads were analyzed with QIIME ver. 2.2020.6 (Quantitative Insights into Microbial Ecology) software package. Single-end FASTQ reads were imported into QIIME using a q2 manifest file import method and quality filtered using the q2-dada2 denoising method. The frequencies of amplicon sequence variants (ASVs) were identified at the 100% similarity level. The representative sequences of ASVs were aligned with the Silva database. The taxonomic composition of each sample at the species level was calculated.
2.8 GC−MS analysis
Gas chromatography−mass spectrometry (GC−MS) analysis was conducted using a SHIMADZU GC2030-QP2020 NX GC−MS system with an HP-FFAP capillary column. A 1-μL aliquot of the analyte was injected in split mode (5:1). Helium was used as the carrier gas, the front inlet purge flow was 3 mL min−1, and the gas flow rate through the column was 1 mL min−1. The initial temperature was kept at 80°C for 1 min, then raised to 200°C at a rate of 10°C min−1 for 5 min, and then kept for 1 min at 240°C at a rate of 40°C min−1. The injection, transfer line, quad, and ion source temperatures were 240°C, 240°C, 200°C, and 150°C, respectively. The energy was −70 eV in electron impact mode. The mass spectrometry data were acquired in Scan mode with a m/z range of 33–150 after a solvent delay of 3.5 min.
2.9 Statistical analysis
The core metric phylogenetic method in the q2 diversity plug-in of QIME was used for diversity analysis. To estimate α-diversity (within sample), different metrics were calculated: Shannon (a quantitative measure of the number of ASVs present in a given sample and their relative abundance or community richness) and Simpson metrics. Alpha diversity indices were compared using the Mann–Whitney test. Principal coordinate analysis (PCoA) and Principal component analysis (PCA) based on the weighted UniFrac measures was used to visualize beta diversity relationships.
The microbial comparisons between the DS and healthy groups were performed using the Mann–Whitney test. Associations between clinical indices and Blautia were evaluated by the MaAsLin analysis (Stewart et al., 2018). Statistical analysis of the clinical data was performed using SPSS (Statistical Package for Social Sciences) 22.0 software (SPSS Inc., Chicago, IL, USA). P < 0.05 was considered statistically significant.
3 Results
3.1 Blautia species spectrum in DS and healthy children
A total of 2,400,845 high-quality combined sequence reads were obtained after data processing (average, 80,028.2 reads). All sequences were clustered into 150 representative ASVs, and 10 of the 150 ASVs were identified as Blautia taxa. The representative sequence of each ASV was assigned to the BLASTN dataset to estimate the diversity of Blautia species in the DS and healthy groups. Six published Blautia species with the best hits are shown in Table 1, namely, B. massiliensis, B. wexlerae, B. faecis, B. stercoris, B. obeum, and B. argi. Of these, ASV1, assigned to B. massiliensis, was the dominant species, representing 49.75% of Blautia sequences. In addition, the proportions of B. wexlerae, B. faecis, B. stercoris, B. obeum, and B. argi (ASV34 and ASV53) in the Blautia sequences were 41.85%, 2.98%, 2.54%, 2.10%, and 0.62%, respectively (ASV34 = 0.54%, ASV53 = 0.08%).
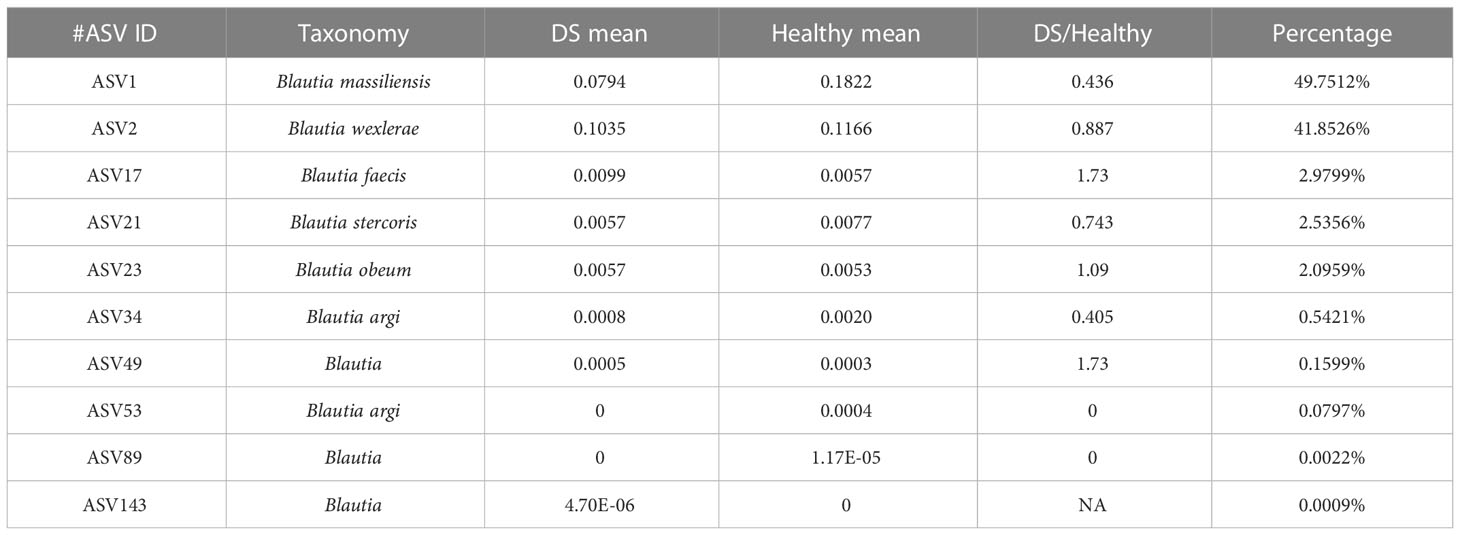
Table 1 Genomic DNA cluster numbering and taxonomic annotation.
These findings revealed the detailed phylogenetic inventory of the Blautia bacterial group in the human gut microbiota and showed that Blautia is prevalent in fecal samples from DS and healthy children.
3.2 Differences in Blautia diversity between DS and healthy children
Microbial alpha diversity was assessed by means of the Shannon and Simpson indices. The Shannon index was significantly increased in the DS group (Figure 1A) compared with the healthy group (4.104 vs. 3.984, P = 0.004), and there was no significant change of Simpson diversity index between the two groups (0.908 vs. 0.904, P = 0.1192) (Figure S3A), suggesting greater Blautia commensal diversity in the DS group. To further assess the alteration in the Blautia composition in the DS group, we used PCA to examine the observed discrete clustering of the intestinal microbiome in the DS and healthy groups. The results showed that the characteristics of the Blautia genus in DS patients were significantly different from those in healthy individuals (Figure S3B). Significant differences were also found by PCoA analysis, indicating that the Blautia genus structure differed significantly between the DS and healthy groups (Figure 1B). In addition, 7 of the 10 total ASVs were shared among all the samples. As shown in the Venn diagram, ASV143 was exclusively present in the DS group, while ASV53 (Blautia argi) and ASV89 were exclusively present in the healthy group (Figure 1C).
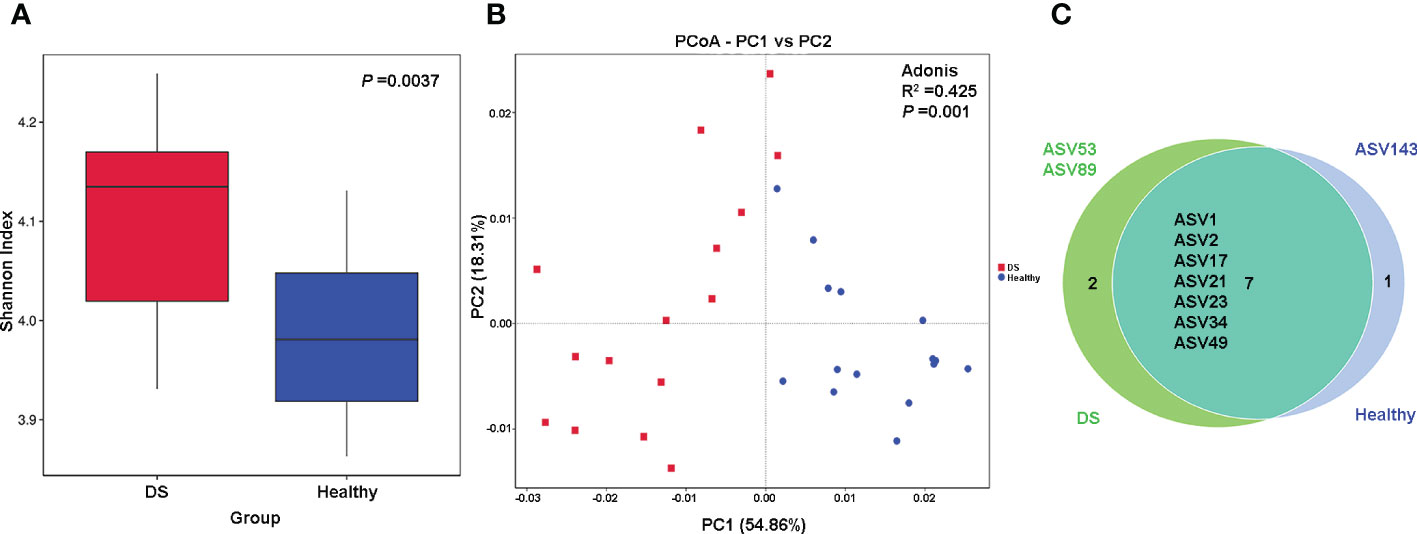
Figure 1 Differences in Blautia genus diversity between DS and healthy children. (A) Alpha diversity based on the Shannon index at the species level; (B) The principal coordinate analysis (PCoA) on the genus level; (C) Venn diagram illustrating the overlap of the ASVs identified in the fecal microbiota between the two groups.
3.3 Differences in the relative abundance of Blautia species between DS and healthy children
As a comparison of the group differences at the Blautia genus level, the quantitative PCR results showed that the relative abundance (RA) of Blautia was significantly decreased in the intestinal tract of children with DS (P < 0.05, Figure 2A). This result was consistent with our previous findings (Ren et al., 2022), which showed a significantly lower level of Blautia in children with DS. Taken together, these results indicate that the Blautia genus is disturbed in individuals with DS.

Figure 2 Differences in the relative abundance of Blautia species between DS and healthy children. (A) Relative abundance of Blautia between the two groups, *P<0.05 (B, E, G, H) Three species of Blautia were significantly decreased in children with DS, ***P < 0.0001, **P < 0.01; (C, F) Two species of Blautia were not significantly different between the two groups.
Next, we explored the RA of Blautia at the species level in DS and healthy children through Blautia-specific amplicon sequencing. The abundances of B. wexlerae and B. obeum were not significantly different between the two groups (Figures 2C, F).
Three species of Blautia were significantly decreased in children with DS: B. massiliensis (ASV1, P < 0.0001, Figure 2B), B. stercoris (ASV21, P = 0.0054, Figure 2E), and B. argi (ASV34, P = 0.0001, Figure 2G; ASV53, P = 0.0039, Figure 2H). Notably, B. argi (ASV53, P = 0.0039, Figure 2H) was nearly absent in children with DS. While, B. faecis significantly increased in children with DS (ASV17, P < 0.0001). While, B. faecis significantly increased in children with DS (ASV17, P < 0.0001, Figure 2D).
3.4 Relationship between Blautia species and cognitive function
To explore the factors contributing to the Blautia variation in the gut microbiome of DS and healthy children, canonical correspondence analysis (CCA) was performed. Some clinical parameters were identified as important contributors for differentiating DS samples from healthy samples (Figure 3A, Table S2). Among them cognitive indices had significant effects on healthy subject clustering, indicating that these clinical parameters are highly associated with the Blautia community variation between DS and healthy children. WMI had the largest effect on the healthy sample variation (R2 = 0.798105, P = 0.005).
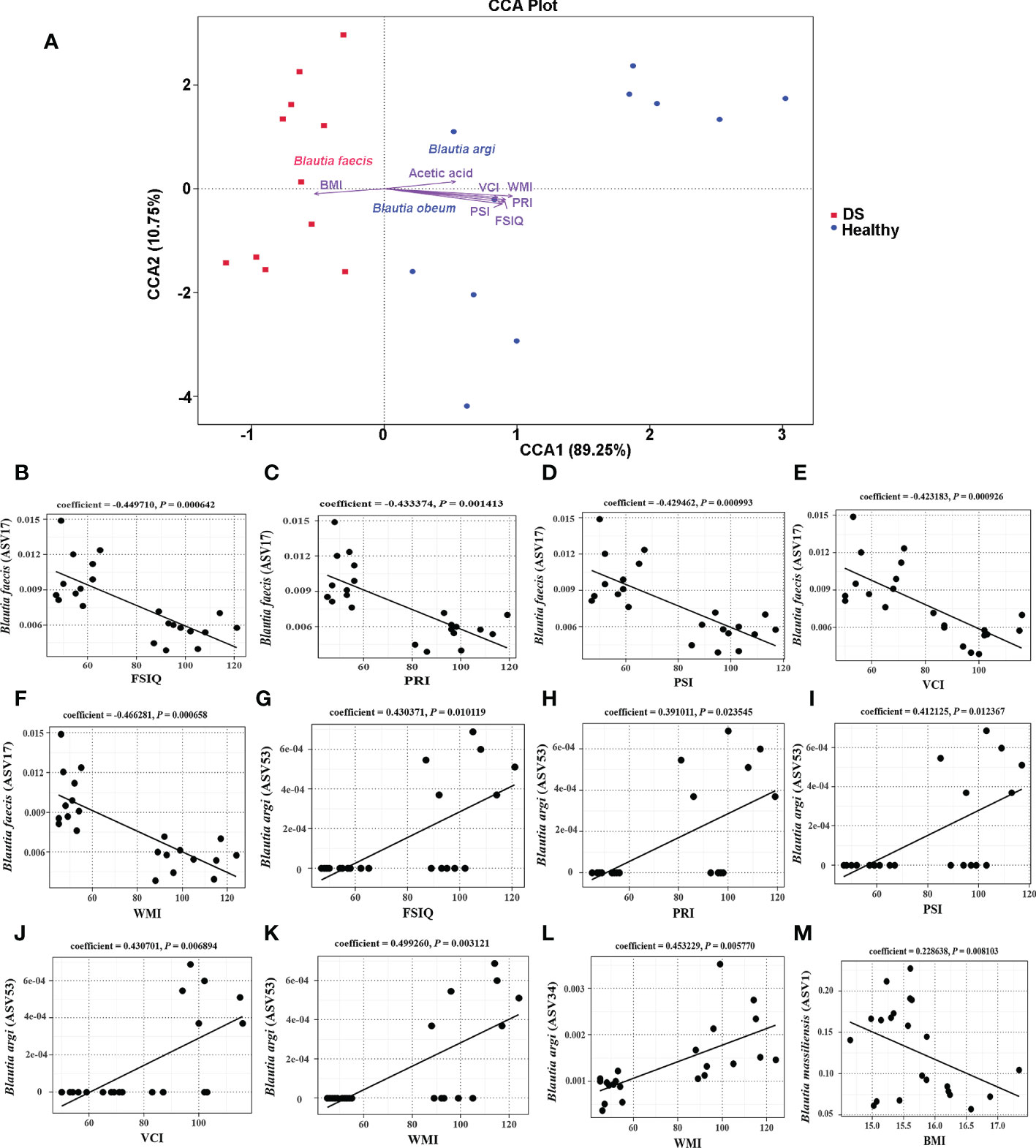
Figure 3 Correlations between Blautia species and cognitive function scores. (A) Key contributors to the Blautia variation were determined by CCA analysis. (B–F) The correlations between (B) faecis and all WISC-IV scores; (G–K) The correlations between (B) argi (ASV53) and all WISC-IV scores; (L) The correlation between (B) argi (ASV34) and the score of WMI; (M) The correlation between (B) massiliensis and BMI.
Furthermore, we examined correlations between the WISC-IV scores and the species level of Blautia. Significant negative correlations between B. faecis and all WISC-IV scores were observed by MaAsLin analysis (Figures 3B–F). B. argi (ASV53) had a significant positive association with all WISC-IV scores (Figures 3G–K), which is consistent with the findings of the CCA. While, B. argi (ASV34) was significant positively related to the score of WMI (Figure 3L). B. argi (ASV53) was indicated as an important contributor for differentiating healthy samples from DS samples. In addition, B. massiliensis had a significant positive association with BMI (Figure 3M). These results indicate that B. argi might be beneficial for cognitive function, while B. faecis might be a risk factor for cognitive function.
3.5 Relationship between Blautia species and acetic acid
SCFAs, particularly acetic acid, propionate, and butyrate, are the end products of the intestinal microbial fermentation of dietary fibers and resistant starch. The levels of SCFAs were determined by GC−MS analysis (Table S3), and the average level of acetic acid was significantly lower in the feces of the DS group than in that of the healthy samples (P < 0.05, Figure 4A). Notably, Blautia has been reported to be an acetic acid-producing bacterium (Aoki et al., 2021), and B. massiliensis (ASV1) and acetic acid contents had a significant positive correlation (P = 0.0014, Figure 4B), indicating its acetic acid-producing property. The relative abundance of Blautia species showed a positive correlation with a high level of acetic acid, which is consistent with previous studies (Fung et al., 2017).
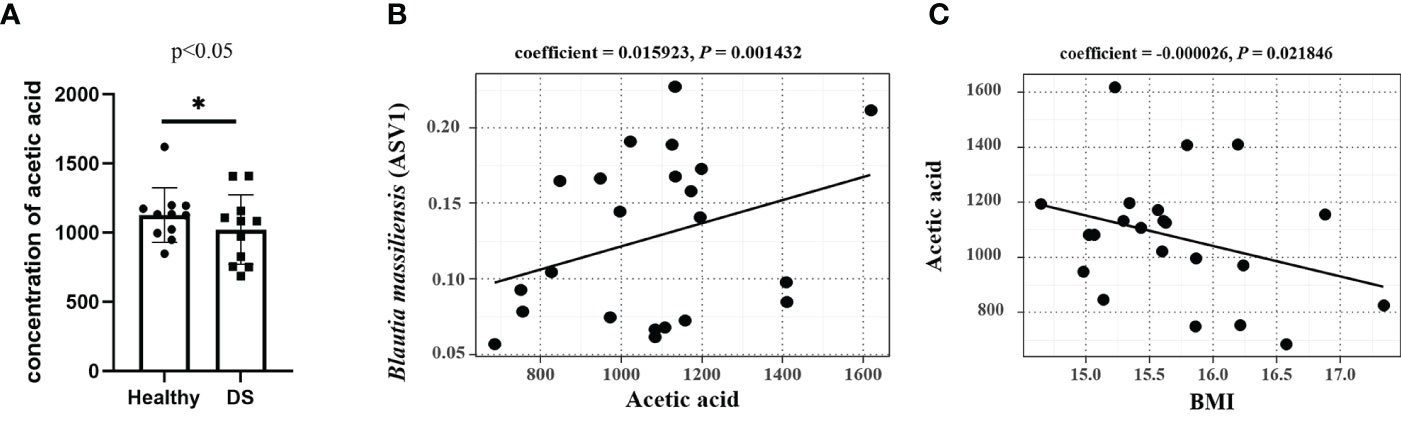
Figure 4 Correlations between (B) massiliensis and acetic acid. (A) The average level of acetic acid was significantly decreased in the feces of the DS group; (B) (B) massiliensis (ASV1) had a significant positive correlation with acetic acid contents; (C) The abundance of acetic acids had a significant negative correlation with BMI.
To further investigate the relationship between clinical indices and acetic acid, we used MaAsLin analysis to measure the correlation between WISC-IV scores, BMI and the level of acetic acid content. The findings indicated that acetic acid and B. massiliensis was negatively correlated with BMI (Figures 3M and 4C), suggesting that acetic acid plays an important role in over-weight of DS children. However, our findings indicated no significant differences between the abundance of acetic acids and the WISC-IV scores.
The importance of the association between the proportion of Blautia species and a high level of acetic acid is therefore supported by the acetic acid-producing effect, implying a beneficial impact of B. massiliensis as an acetic acid producing species on cognitive (Wen et al., 2020) and metabolic function in Chinese children by the metabolites.
3.6 Predictive function analysis
Tax4Fun, based on the closed-reference ASVs, was used to predict the functional categories of the KEGG Orthology (KO). The genomes of the Blautia-specific groups in DS children and healthy children were predicted, in conjunction with their physiological, cellular, and molecular functions (Figure 5). The genes of Blautia species in DS children were mainly enriched in molecular pathways, that is, pyruvate metabolism and glutamate metabolism. In contrast, the genes were deficient in the DS group in molecular pathways such as starch and sucrose and glycolysis metabolism. There were significant differences in the enrichment of the molecular pathways between the two groups of genes (Figure S4). Thus, functional genomics prediction suggested that Blautia may affect cognitive function by altering metabolites.
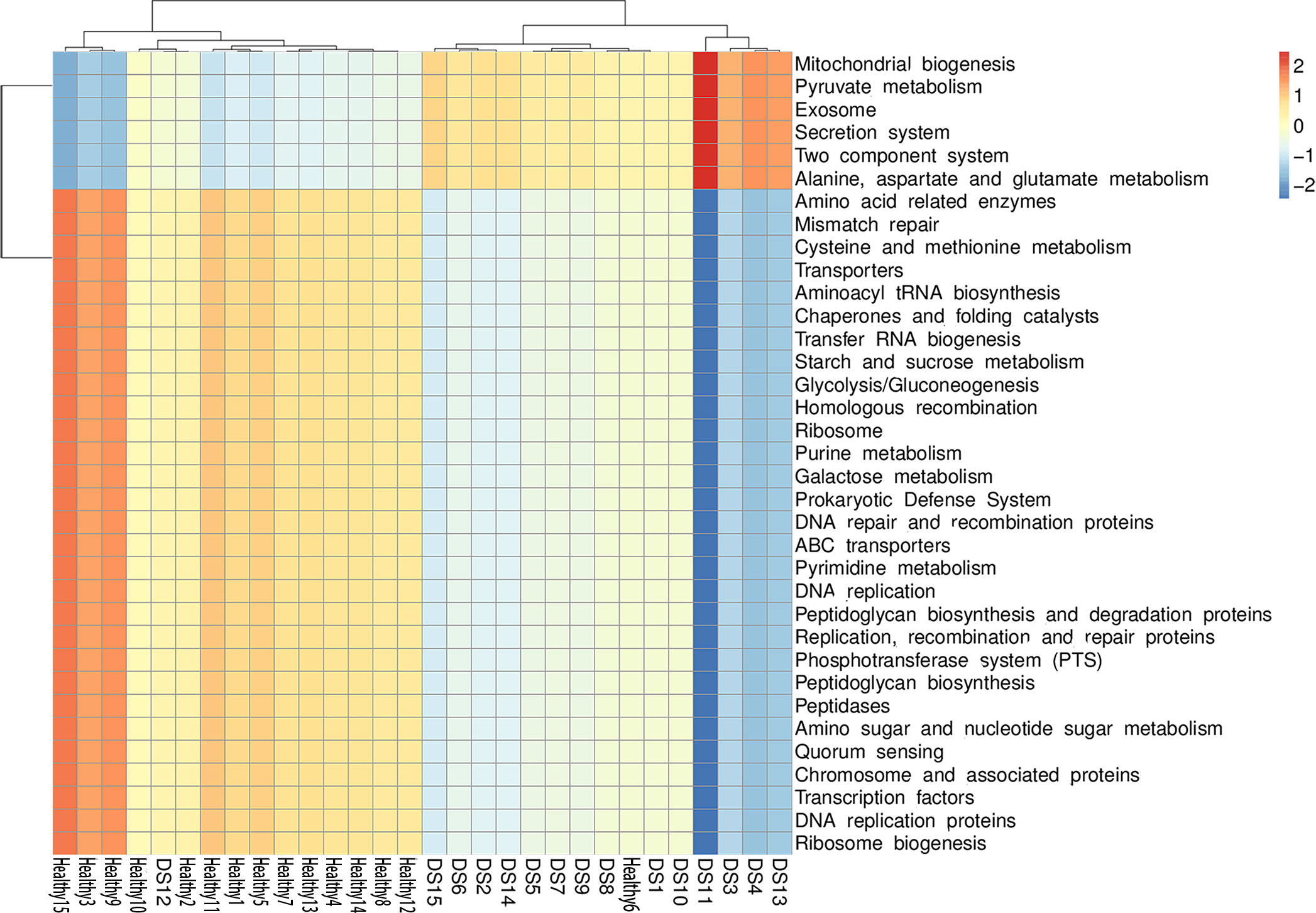
Figure 5 Functional predictions for the Blautia species of the DS and healthy control groups. Significantly enriched KEGG pathways at molecular level.
4 Discussion
Recent work has found that the gut microbiota play a key role in cognitive function in early neurodegenerative disease (Fung et al., 2017). In particular, the genus Blautia, one of the most abundant genera in the gut, has been identified to have an important relationship with DS and early-onset AD (Chen et al., 2020). In this study, the detailed composition of Blautia at species level in the gut microbiota of DS and healthy children was explored using Blautia group-specific amplicon sequencing. B. massiliensis were identified decreased in children with DS, and B. argi was nearly absent in children with DS, while B. faecis showed a higher level in DS children. In addition, we found that the level of B. argi was significantly correlated with cognitive WISC-IV scores. Our findings highlight the potential significance of B. faecis and B. argi in the development of cognitive function in children with DS and have implications for improving strategies in the gut-brain field. Molecular diversity at the species level with clinical cognitive function will help form the basis for future functional work of Blautia species on gut-brain neural mechanisms.
Blautia was proven to maintain the intestinal environmental balance by upregulating intestinal regulatory T cells and producing SCFAs (Chen et al., 2014). A cohort study (n = 108) reported that a lower proportion of B. hansenii could increase the risk of AD (Haran et al., 2019). A new study demonstrated that oral Blautia stercoris MRx0006 can attenuate some of the behavioral deficits in an autism-relevant genetic mouse model (Sen et al., 2022). Additionally, a prospective study showed that the abundance of Blautia in PD patients was decreased, and the scores of neuropathological indicators were improved after fecal microbial transplantation with an increased abundance of Blautia (Kuai et al., 2021). Notably, we previously found that the genus Blautia was decreased in DS children and positively related to cognitive function (Ren et al., 2022), which suggests that different species of Blautia may exert beneficial effects on children with DS. To conduct a more in-depth study of Blautia at the species or even strain levels in the development of cognitive function in children with DS, we performed Blautia-specific amplicon sequencing and identified the profile and alteration of Blautia at the species level in Chinese children with DS. As expected, a shift of the Blautia group in individuals with DS was observed, as shown by the PCoA plot, with higher α-diversity and observed species in the Venn diagram. Abundances of several Blautia species were detected to differ in the DS and healthy groups. Among them, B. massiliensis, and B. argi were significantly decreased in DS samples. In addition, B. massiliensis, the dominant genus in healthy children samples, showed remarkably positive correlations with BMI. Another absent Blautia species in DS samples was B. argi, which was found to have a significant explanatory effect on the variation across healthy samples and showed a significant positive link with cognitive function. B. argi is a new species of Blautia isolated from dog feces (Paek et al., 2019). However, B. argi was present extremely low in healthy fecal samples and was at trace levels or even absent from the DS group. Further experiments are needed to verify its effect on cognitive function. Of particular interest, we revealed higher level of B. faecis in DS gut microbiota, which was also inversely correlated with WISC-IV scores, suggesting its negative role on host cognitive function. Furthermore, we found that acetic acid, a metabolite of Blautia (Flint et al., 2008), was decreased in the DS group and negatively correlated with BMI. Acetic acid, one of the major SCFAs, is metabolized by gut bacteria from dietary fiber fermentation and exerts a variety of physiological functions (Nicholson et al., 2012). It has been widely observed that acetic acid can promote metabolic syndrome via the gut–microbiota–brain–β cell axis (Perry et al., 2016). Moreover, acetic acid may enhance the efficiency of the synaptic vesicle cycle and thereby protect against cognitive impairment in mice with type 1 diabetes (Zheng et al., 2021b). Here, we observed that the relative abundance of B. massiliensis was positively associated with the level of acetic acid, indicating its acetic acid-producing ability. These findings suggest a probiotic role for the Blautia species B. massiliensis in the development with over-weight issue in children with DS through its metabolite product, especially acetic acid. Overall, this study provides potential insight for the promising application of B. massiliensis and acetic acid as preservatives of DS children with obesity issue in the future.
To reveal the genomic features of the alerted Blautia group with DS, we performed Tax4Fun analysis and predicted the genes of Blautia species in children with DS compared with healthy children. The pathways of pyruvate metabolism and glutamate metabolism were mainly enriched in DS intestinal samples. Pyruvate accumulation was reported to contribute to acceleration-induced impairment of cognitive abilities (Mo et al., 2021). Importantly, indole-3-pyruvic acid was identified as a signature for the discrimination and prediction of AD (Wu et al., 2021). Severe cognitive deficit correlates with high levels of lactate and pyruvate in cerebrospinal fluid according to the senile dementia report (Pugliese et al., 2005). Pyruvate can be metabolized to glutamine in the brain (Gray et al., 2014). The synthesis of glutamine mainly occurs in astrocytes and is related to the immune inflammatory response of the nervous system (Storm-Mathisen et al., 1983). In addition, we also observed that the genes of Blautia species related to glycolysis were decreased in children with DS. The major function of aerobic glycolysis is to maintain high levels of glycolytic intermediates to support anabolic reactions in cells (Lunt and Vander Heiden, 2011). Recent studies revealed that changes in aerobic glycolysis prevail in the early phase of AD (An et al., 2018). Astrocytic glycolysis affects cognitive functions, and dysregulations of glycolysis cause synaptic plasticity and behavioral deficits in AD (Le Douce et al., 2020). The reduction in glycolysis led by the decreased abundance of Blautia species further provides potential insight for the possible application of Blautia species and metabolites in the future for delaying and ameliorating cognitive dysfunction in children with DS. Another interesting observation is the decreased starch and sucrose metabolism pathway in the DS group, which had the same trend as B. massiliensis in the DS group. B. massiliensis also present a significant negative correlation with BMI index. Comparative genomics studies also showed that the genes encoding B. massiliensis were mainly enriched in carbohydrate transport and metabolism functions compared with other species within the genus Blautia (Durand et al., 2017). The final products of glucose fermentation by B. massiliensis include acetic acid, succinic acid, and lactic acid (Liu et al., 2021). Taken together, the decrease in B. massiliensis may affect the cognitive function and weight of children with DS by reducing carbohydrate metabolism, especially acetic acid from glucose metabolism. These findings indicate that pyruvate and glutamate metabolism abnormalities due to Blautia species deletion may have a major impact on cognitive dysfunction in DS.
In summary, cognitive improvements made during a small window early in life are important for individuals with DS (Antonarakis et al., 2020), and the gut microbiota has potential in the prevention and treatment of early neurodegenerative disease (Heiss and Olofsson, 2019). Our study profiled Blautia at the species level and suggested that two Blautia species, B. faecis and B. argi, have a close relationship with cognitive function in children with DS. Uniquely, in the present study, the data indicate that the Blautia group in subjects with DS is correlated with cognitive function, providing a deeper understanding that modifying the proportion of B. faecis and B. argi, may help ameliorating cognitive dysfunction for children with DS. Additionally, we highlight the specific probiotic and Blautia-derived product acetic acid and B. massiliensis, with plausible effects on obesity for DS children. Modulating the profile of the acetate-producing Blautia taxa, could be a promising strategy in the search for alternatives for the improvement of cognitive function and metabolic disorders in children with DS.
Our research is limited (1) by the lack of macro genome sequencing data to identify important species’ metabolic pathways, (2) by the limited sample size, and further studies with more individuals needed to confirm our findings. Furthermore, our suggestion on the prevalence of specific species with different metabolites requires future research to reveal the metabolic pathways of these species in the DS gut microbiota. Long-term observation may be helpful to study the dynamic changes in the Blautia group in the intestinal microbiota of children with DS at the species level during the growth process.
5 Conclusion
In conclusion, our data extend our previous work and provide evidence for the molecular diversity of Blautia species in the fecal microbiota of DS patients compared with healthy subjects. Importantly, we identified three species, B. massiliensis, B. argi and B. faecis, that may play an important role in the development of cognitive function in children with DS.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA885084/, PRJNA885084.
Ethics statement
The studies involving human participants were reviewed and approved by the Ethics Review Committee of Peking University People’s Hospital of China. Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author contributions
JG, XH, and SR contributed to conception and design of the study. SR and XW collected samples. XH, QM, and YZ extracted the sample DNA. XH and NW performed the statistical analysis. XH and NW wrote the first draft of the manuscript. XH, NW, WY, SW, and JG wrote sections of the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No: 31671066) and Natural Science Foundation of Beijing Municipality (Grant No: 7222205).
Acknowledgments
We thank all of the participants and their parents who made this study possible.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2023.1109889/full#supplementary-material
References
Ahmed, S., Busetti, A., Fotiadou, P., Vincy Jose, N., Reid, S., Georgieva, M., et al. (2019). In vitro characterization of gut microbiota-derived bacterial strains with neuroprotective properties. Front. Cell Neurosci., 13:402. doi: 10.3389/fncel.2019.00402
An, Y., Varma, V. R., Varma, S., Casanova, R., Dammer, E., Pletnikova, O., et al. (2018). Evidence for brain glucose dysregulation in alzheimer's disease. Alzheimers Dement. 14 (3), 318–329. doi: 10.1016/j.jalz.2017.09.011
Antonarakis, S. E., Skotko, B. G., Rafii, M. S., Strydom, A., Pape, S. E., Bianchi, D. W., et al. (2020). Down syndrome. Nat. Rev. Dis. Primers. 6 (1), 9. doi: 10.1038/s41572-019-0143-7
Aoki, R., Onuki, M., Hattori, K., Ito, M., Yamada, T., Kamikado, K., et al. (2021). Commensal microbe-derived acetate suppresses NAFLD/NASH development via hepatic FFAR2 signalling in mice. Microbiome. 9 (1), 188. doi: 10.1186/s40168-021-01125-7
The global burden of disease (GBD) 2019 data. Available at: http://ghdx.healthdata.org/gbd-results-tool.
Biddle, A., Stewart, L., Blanchard, J., Leschine, S. (2013). Untangling the genetic basis of fibrolytic specialization by lachnospiraceae and ruminococcaceae in diverse gut communities. Diversity (Basel). 5 (3), 627–640. doi: 10.3390/d5030627
Chen, Y., Fang, L., Chen, S., Zhou, H., Fan, Y., Lin, L., et al. (2020). Gut microbiome alterations precede cerebral amyloidosis and microglial pathology in a mouse model of alzheimer's disease. BioMed. Res. Int. 2020, 8456596. doi: 10.1155/2020/8456596
Chen, L., Wang, W., Zhou, R., Ng, S. C., Li, J., Huang, M., et al. (2014). Characteristics of fecal and mucosa-associated microbiota in Chinese patients with inflammatory bowel disease. Med. (Baltimore). 93 (8), e51. doi: 10.1097/MD.0000000000000051
Durand, G. A., Pham, T., Ndongo, S., Traore, S. I., Dubourg, G., Lagier, J. C., et al. (2017). Blautia massiliensis sp. nov., isolated from a fresh human fecal sample and emended description of the genus Blautia. Anaerobe. 43, 47–55. doi: 10.1016/j.anaerobe.2016.12.001
Erny, D., Dokalis, N., Mezö, C., Castoldi, A., Mossad, O., Staszewski, O., et al. (2021). Microbiota-derived acetate enables the metabolic fitness of the brain innate immune system during health and disease. Cell Metab. 33 (11), 2260–2276.e7. doi: 10.1016/j.cmet.2021.10.010
Flint, H. J., Bayer, E. A., Rincon, M. T., Lamed, R., White, B. A. (2008). Polysaccharide utilization by gut bacteria: potential for new insights from genomic analysis. Nat. Rev. Microbiol. 6 (2), 121–131. doi: 10.1038/nrmicro1817
Fung, T. C., Olson, C. A., Hsiao, E. Y. (2017). Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 20 (2), 145–155. doi: 10.1038/nn.4476
Gray, L. R., Tompkins, S. C., Taylor, E. B. (2014). Regulation of pyruvate metabolism and human disease. Cell Mol. Life Sci. 71 (14), 2577–2604. doi: 10.1007/s00018-013-1539-2
Grieco, J., Pulsifer, M., Seligsohn, K., Skotko, B., Schwartz, A. (2015). Down syndrome: Cognitive and behavioral functioning across the lifespan. Am. J. Med. Genet. C Semin. Med. Genet. 169 (2), 135–149. doi: 10.1002/ajmg.c.31439
Haran, J. P., Bhattarai, S. K., Foley, S. E., Dutta, P., Ward, D. V., Bucci, V., et al. (2019). Alzheimer's disease microbiome is associated with dysregulation of the anti-inflammatory p-glycoprotein pathway. mBio. 10 (3), e00632–e00619. doi: 10.1128/mBio.00632-19
Heiss, C. N., Olofsson, L. E. (2019). The role of the gut microbiota in development, function and disorders of the central nervous system and the enteric nervous system. J. Neuroendocrinol. 31 (5), e12684. doi: 10.1111/jne.12684
Kalyana Chakravarthy, S., Jayasudha, R., Sai Prashanthi, G., Ali, M. H., Sharma, S., Tyagi, M., et al. (2018). Dysbiosis in the gut bacterial microbiome of patients with uveitis, an inflammatory disease of the eye. Indian J. Microbiol. 58 (4), 457–469. doi: 10.1007/s12088-018-0746-9
Kuai, X. Y., Yao, X. H., Xu, L. J., Zhou, Y. Q., Zhang, L. P., Liu, Y., et al. (2021). Evaluation of fecal microbiota transplantation in parkinson's disease patients with constipation. Microb. Cell Fact. 20 (1), 98. doi: 10.1186/s12934-021-01589-0
Le Douce, J., Maugard, M., Veran, J., Matos, M., Jégo, P., Vigneron, P. A., et al. (2020). Impairment of glycolysis-derived l-serine production in astrocytes contributes to cognitive deficits in alzheimer's disease. Cell Metab. 31 (3), 503–517.e8. doi: 10.1016/j.cmet.2020.02.004
Liu, X., Mao, B., Gu, J., Wu, J., Cui, S., Wang, G., et al. (2021). Blautia-a new functional genus with potential probiotic properties? Gut Microbes 13 (1), 1–21. doi: 10.1080/19490976.2021.1875796
Lunt, S. Y., Vander Heiden, M. G. (2011). Aerobic glycolysis: meeting the metabolic requirements of cell proliferation. Annu. Rev. Cell Dev. Biol. 27, 441–464. doi: 10.1146/annurev-cellbio-092910-154237
Luu, T. H., Michel, C., Bard, J. M., Dravet, F., Nazih, H., Bobin-Dubigeon, C. (2017). Intestinal proportion of Blautia sp. is associated with clinical stage and histoprognostic grade in patients with early-stage breast cancer. Nutr. Cancer. 69 (2), 267–275. doi: 10.1080/01635581.2017.1263750
Mo, F., Zhang, H., Tang, Y., Qi, R., Nie, S., Shen, H., et al. (2021). Pyruvate accumulation may contribute to acceleration-induced impairment of physical and cognitive abilities: an experimental study. Biosci. Rep. 41 (4), BSR20204284. doi: 10.1042/BSR20204284
NCBI. Available at: https://www.ncbi.nlm.nih.gov/data-hub/taxonomy/tree/?taxon=572511.
Nicholson, J. K., Holmes, E., Kinross, J., Burcelin, R., Gibson, G., Jia, W., et al. (2012). Host-gut microbiota metabolic interactions. Science. 336 (6086), 1262–1267. doi: 10.1126/science.1223813
Paek, J., Shin, Y., Kook, J. K., Chang, Y. H. (2019). Blautia argi sp. nov., a new anaerobic bacterium isolated from dog faeces. Int. J. Syst. Evol. Microbiol. 69 (1), 33–38. doi: 10.1099/ijsem.0.002981
Perry, R. J., Peng, L., Barry, N. A., Cline, G. W., Zhang, D., Cardone, R. L., et al. (2016). Acetic acid mediates a microbiome-brain-β-cell axis to promote metabolic syndrome. Nature. 534 (7606), 213–217. doi: 10.1038/nature18309
Pugliese, M., Carrasco, J. L., Andrade, C., Mas, E., Mascort, J., Mahy, N. (2005). Severe cognitive impairment correlates with higher cerebrospinal fluid levels of lactate and pyruvate in a canine model of senile dementia. Prog. Neuropsychopharmacol. Biol. Psychiatry 29 (4), 603–610. doi: 10.1016/j.pnpbp.2005.01.017
Ren, S., Wang, X., Qin, J., Mu, Q., Ye, S., Zhang, Y., et al. (2022). Altered gut microbiota correlates with cognitive impairment in Chinese children with down's syndrome. Eur. Child Adolesc. Psychiatry 31 (1), 189–202. doi: 10.1007/s00787-021-01799-2
Sen, P., Sherwin, E., Sandhu, K., Bastiaanssen, T. F. S., Moloney, G. M., Golubeva, A., et al. (2022). The live biotherapeutic Blautia stercoris MRx0006 attenuates social deficits, repetitive behaviour, and anxiety-like behaviour in a mouse model relevant to autism. Brain Behav. Immun. 106, 115–126. doi: 10.1016/j.bbi.2022.08.007
Squassante, L., Spiridigliozzi, G., Visootsak, J., Heller, J., Khwaja, O. (2015). Assessment of cognitive scales to examine memory, executive function and language in individuals with down syndrome: Implications of a 6-month observational study. Front. Behav. Neurosci. 9, 300. doi: 10.3389/fnbeh.2015.00300
Stewart, C. J., Ajami, N. J., O'Brien, J. L., Hutchinson, D. S., Smith, D. P., Wong, M. C., et al. (2018). Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature. 562 (7728), 583–588. doi: 10.1038/s41586-018-0617-x
Storm-Mathisen, J., Leknes, A. K., Bore, A. T., Vaaland, J. L., Edminson, P., Haug, F. M., et al. (1983). First visualization of glutamate and GABA in neurones by immunocytochemistry. Nature. 301 (5900), 517–520. doi: 10.1038/301517a0
Unger, M. M., Spiegel, J., Dillmann, K. U., Grundmann, D., Philippeit, H., Bürmann, J., et al. (2016). Short chain fatty acids and gut microbiota differ between patients with parkinson's disease and age-matched controls. Parkinsonism Relat. Disord. 32, 66–72. doi: 10.1016/j.parkreldis.2016.08.019
Wen, C., Xie, T., Pan, K., Deng, Y., Zhao, Z., Li, N., et al. (2020). Acetate attenuates perioperative neurocognitive disorders in aged mice. Aging (Albany NY). 12 (4), 3862–3879. doi: 10.18632/aging.102856
Wu, L., Han, Y., Zheng, Z., Peng, G., Liu, P., Yue, S., et al. (2021). Altered gut microbial metabolites in amnestic mild cognitive impairment and alzheimer's disease: Signals in host-microbe interplay. Nutrients. 13 (1), 228. doi: 10.3390/nu13010228
Zheng, H., Xu, P., Jiang, Q., Xu, Q., Zheng, Y., Yan, J., et al. (2021a). Depletion of acetate-producing bacteria from the gut microbiota facilitates cognitive impairment through the gut-brain neural mechanism in diabetic mice. Microbiome. 9 (1), 145. doi: 10.1186/s40168-021-01088-9
Keywords: Blautia massiliensis, Blautia argi, cognitive dysfunction, acetic acid, Down syndrome, Blautia faecis
Citation: Hou X, Wu N, Ren S, Wang X, Mu Q, Zhang Y, Wang S, Yu W and Guo J (2023) Profiling Blautia at high taxonomic resolution reveals correlations with cognitive dysfunction in Chinese children with Down syndrome. Front. Cell. Infect. Microbiol. 13:1109889. doi: 10.3389/fcimb.2023.1109889
Received: 28 November 2022; Accepted: 30 January 2023;
Published: 10 February 2023.
Edited by:
Daniel Czyz, University of Florida, United StatesCopyright © 2023 Hou, Wu, Ren, Wang, Mu, Zhang, Wang, Yu and Guo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jingzhu Guo, amluZ3podS5ndW9AYmptdS5lZHUuY24=; Weidong Yu, d2VpZG9uZ3l1QGJqbXUuZWR1LmNu
†These authors have contributed equally to this work and share first authorship