 Guillermo Quindós1*§
Guillermo Quindós1*§ Katherine Miranda-Cadena1§Rosario San-Millán1†§Katyna Borroto-Esoda2Emilia Cantón3María José Linares-Sicilia4
Katherine Miranda-Cadena1§Rosario San-Millán1†§Katyna Borroto-Esoda2Emilia Cantón3María José Linares-Sicilia4 Axel Hamprecht5Isabel Montesinos6‡
Axel Hamprecht5Isabel Montesinos6‡ Anna Maria Tortorano7
Anna Maria Tortorano7 Anna Prigitano7
Anna Prigitano7 Matxalen Vidal-García8‡Cristina Marcos-Arias1
Matxalen Vidal-García8‡Cristina Marcos-Arias1 Andrea Guridi1
Andrea Guridi1 Ferran Sanchez-Reus9Jesús Machuca-Bárcena10
Ferran Sanchez-Reus9Jesús Machuca-Bárcena10 Manuel Antonio Rodríguez-Iglesias10Estrella Martín-Mazuelos11Carmen Castro-Méndez11
Manuel Antonio Rodríguez-Iglesias10Estrella Martín-Mazuelos11Carmen Castro-Méndez11 Leyre López-Soria12Alba Ruiz-Gaitán3Marcelo Fernandez-Rivero3
Leyre López-Soria12Alba Ruiz-Gaitán3Marcelo Fernandez-Rivero3 Damaris Lorenzo13
Damaris Lorenzo13 Javier Capilla13Antonio Rezusta8
Javier Capilla13Antonio Rezusta8 Javier Pemán3Josep Guarro13Joana Pereira14
Javier Pemán3Josep Guarro13Joana Pereira14 Célia Pais14
Célia Pais14 Orazio Romeo15Guillermo Ezpeleta16Nerea Jauregizar17David Angulo2
Orazio Romeo15Guillermo Ezpeleta16Nerea Jauregizar17David Angulo2 Elena Eraso1
Elena Eraso1- 1Laboratorio de Micología Médica, UFI 11/25, Departamento de Inmunología, Microbiología y Parasitología, Facultad de Medicina y Enfermería, Universidad del País Vasco/Euskal Herriko Unibertsitatea (UPV/EHU), Bilbao, Spain
- 2SCYNEXIS, Inc., Jersey City, NJ, United States
- 3Instituto de Investigación Sanitaria, Hospital Universitario y Politécnico La Fe, Valencia, Spain
- 4Research Group GC24, Maimonides Biomedical Research Institute of Cordoba (IMIBIC), Department of Microbiology, Facultad de Medicina y Enfermería, Universidad de Córdoba, Córdoba, Spain
- 5University Hospital Cologne, Cologne and Institute for Medical Microbiology and Virology, University of Oldenburg, Oldenburg, Germany
- 6Microbiology Department, LHUB-ULB, Hôpital Erasme, Brussels, Belgium
- 7Dipartimento di Scienze Biomediche per la Salute, Università degli Studi di Milano, Milan, Italy
- 8Servicio de Microbiología, Hospital Universitario Miguel Servet, Zaragoza, Spain
- 9Servei de Microbiologia, Hospital de la Santa Creu i Sant Pau, Barcelona, Spain
- 10Área de Microbiología, Departamento de Biomedicina, Biotecnología y Salud Pública, Facultad de Medicina, Universidad de Cádiz, Cádiz, Spain
- 11Servicio de Microbiología, Hospital Universitario de Valme, Sevilla, Spain
- 12Servicio de Microbiología, Hospital Universitario de Cruces and BioCruces Bizkaia, Barakaldo, Spain
- 13Microbiology Unit, Medical School, Universitat Rovira i Virgili, Reus, Spain
- 14Centre of Molecular and Environmental Biology (CBMA), Department of Biology, University of Minho, Braga, Portugal
- 15Department of Chemical, Biological, Pharmaceutical and Environmental Sciences, University of Messina, Messina, Italy
- 16Servicio de Microbiología, Complejo Hospitalario de Navarra, Pamplona and Departamento de Medicina Preventiva y Salud Pública, Facultad de Medicina y Enfermería, Universidad del País Vasco/Euskal Herriko Unibertsitatea (UPV/EHU), Bilbao, Spain
- 17Departamento de Farmacología, Facultad de Medicina y Enfermería, Universidad del País Vasco/Euskal Herriko Unibertsitatea (UPV/EHU), Bilbao, Spain
Background: Ibrexafungerp (SCY-078) is the newest oral and intravenous antifungal drug with broad activity, currently undergoing clinical trials for invasive candidiasis.
Objective: The aim of this study was to assess the in vitro activity of ibrexafungerp and comparators against a collection of 434 European blood isolates of Candida.
Methods: Ibrexafungerp, caspofungin, fluconazole, and micafungin minimum inhibitory concentrations (MICs) were collected from 12 European laboratories for 434 blood isolates, including 163 Candida albicans, 108 Candida parapsilosis, 60 Candida glabrata, 40 Candida tropicalis, 29 Candida krusei, 20 Candida orthopsilosis, 6 Candida guilliermondii, 2 Candida famata, 2 Candida lusitaniae, and 1 isolate each of Candida bracarensis, Candida catenulata, Candida dubliniensis, and Candida kefyr. MICs were determined by the EUCAST broth microdilution method, and isolates were classified according to recommended clinical breakpoints and epidemiological cutoffs. Additionally, 22 Candida auris from different clinical specimens were evaluated.
Results: Ibrexafungerp MICs ranged from 0.016 to ≥8 mg/L. The lowest ibrexafungerp MICs were observed for C. albicans (geometric MIC 0.062 mg/L, MIC range 0.016–0.5 mg/L) and the highest ibrexafungerp MICs were observed for C. tropicalis (geometric MIC 0.517 mg/L, MIC range 0.06–≥8 mg/L). Modal MICs/MIC50s (mg/L) against Candida spp. were 0.125/0.06 for C. albicans, 0.5/0.5 for C. parapsilosis, 0.25/0.25 for C. glabrata, 0.5/0.5 for C. tropicalis, 1/1 for C. krusei, 4/2 for C. orthopsilosis, and 0.5/0.5 for C. auris. Ibrexafungerp showed activity against fluconazole- and echinocandin-resistant isolates. If adopting wild-type upper limits, a non-wild-type phenotype for ibrexafungerp was only observed for 16/434 (3.7%) isolates: 11 (4.6%) C. parapsilosis, 4 (5%) C. glabrata, and 1 (2.5%) C. tropicalis.
Conclusion: Ibrexafungerp showed a potent in vitro activity against Candida.
Introduction
Invasive candidiasis (IC) is the most common healthcare-associated invasive mycosis, being a major cause of human morbidity and mortality. Candida albicans is the most prevalent etiology, but other species, such as Candida glabrata, Candida parapsilosis, Candida krusei (Pichia kudriavzevii), and, more recently, Candida auris, are increasing causes of IC. These emergent species are usually less susceptible to current antifungal drugs. Although Candida isolates displaying antifungal resistance are still uncommon, they are increasingly reported worldwide. Therapy of IC is an unsolved clinical challenge and, for this reason, monitoring antifungal susceptibility patterns and resistance mechanisms is of utmost importance. Moreover, new antifungal drugs are needed as the number of available antifungal drug classes, and particularly those for oral administration, is limited (Arendrup and Patterson, 2017; Quindós et al., 2018; Fuller et al., 2019; Pfaller et al., 2019).
Ibrexafungerp (formerly SCY-078) is a semisynthetic triterpenoid glycoside derived from enfumafungin, which is structurally different from echinocandins and form a new class of antifungal drugs called “fungerps” that strongly inhibit fungal 1,3-β-glucan synthase (Davis et al., 2020). Even ibrexafungerp and echinocandins share similar mechanisms of action, and their binding sites to the target enzyme is not the same, resulting in very limited cross-resistance (Jiménez-Ortigosa et al., 2017; Pfaller et al., 2017). Ibrexafungerp displays significant in vitro and in vivo activities against azole- and echinocandin-resistant isolates of Candida species, including biofilm-forming strains (Jiménez-Ortigosa et al., 2014; Pfaller et al., 2017; Schell et al., 2017; Gamal et al., 2021).
Ibrexafungerp aims to be the first orally and intravenously available glucan synthase inhibitor useful in the treatment of life-threatening fungal infections (Davis et al., 2020) as well as superficial ones, such as vulvovaginal candidiasis (Schwebke et al., 2021; Sobel et al., 2022). Currently, there are 13 listed clinical trials for ibrexafungerp, eight of which have been completed (https://ClinicalTrials.gov/; accessed on March 8, 2022).
In the current study, we have determined the anti-Candida in vitro activity of ibrexafungerp, caspofungin, fluconazole, and micafungin against 434 European Candida blood isolates analyzed in 12 European laboratories.
Materials and Methods
Microorganisms
In vitro susceptibility of a collection of 434 Candida blood isolates (2016–2018) from 434 patients was determined at 12 laboratories from Belgium, Germany, Italy, Portugal, and Spain. Each laboratory studied its own clinical isolates. The collection included 163 C. albicans, 108 C. parapsilosis, 60 C. glabrata, 40 C. tropicalis, 29 C. krusei, 20 Candida orthopsilosis, 6 Candida guilliermondii (Meyerozyma guilliermondii), 2 Candida famata (Debaryomyces hansenii), 2 Candida lusitaniae (Clavispora lusitaniae), and 1 isolate each of Candida bracarensis, Candida catenulata (Diutina catenulata), Candida dubliniensis, and Candida kefyr (Kluyveromyces marxianus). Additionally, 22 C. auris from different clinical specimens were evaluated: Eight isolates were from blood, seven from oral specimens, and 7 from urine (Ruiz-Gaitán et al., 2017; Ruiz-Gaitán et al., 2018). Isolates were identified by phenotypic methods, MALDI-TOF (proteomic method), and, when needed, internal transcribed spacer (ITS) sequencing (genotypic method) (Miranda-Zapico et al., 2011). C. parapsilosis ATCC 22019 and C. krusei ATCC 6258 were included as quality control (QC) strains. Reference strain C. albicans ATCC 64550 was also included as recommended by EUCAST for detecting variation in echinocandin activities. Prior to testing, each isolate was cultured onto Sabouraud dextrose agar and/or CHROMagar Candida medium (Becton Dickinson, Sparks, MD, USA) to ensure purity and viability. QC and reference strain were included every testing day and all their results were within published ranges.
Antifungal Drugs and Susceptibility Testing
All isolates were tested for in vitro susceptibility to ibrexafungerp, caspofungin, fluconazole, and micafungin using the EUCAST broth microdilution method (Arendrup et al., 2020a; Arendrup et al., 2020b; EUCAST, 2020; EUCAST, 2021). The reference powder of ibrexafungerp was obtained from the manufacturer (SCYNEXIS Inc., Jersey City, NJ, USA). Stock solutions were prepared in dimethyl sulfoxide (DMSO), and the final range of ibrexafungerp, caspofungin, and micafungin concentrations tested was 0.008 to 8 mg/L except for fluconazole, which ranged from 0.125 to 128 mg/L. The EUCAST method was performed by using RPMI 1640 with 2% glucose and buffered to pH 7.0 with 0.165 M morpholinopropane sulphate (MOPS). Panels were inoculated with a final standardized cell concentration of 0.5–2.5 × 105 cells/ml. Panels were read spectrophotometrically at 450 nm after 24 h of incubation at 36 ± 1°C. Although caspofungin presents excessive inter-laboratory variation in minimum inhibitory concentration (MIC) results by the EUCAST broth microdilution method (Espinel-Ingroff et al., 2013), MICs were calculated for comparison purposes.
Data Analysis
MIC values were determined as the lowest concentration of antifungal drug that inhibited 50% of growth compared to that of the growth control. For individual species of Candida for which ten or more isolates were tested, summary descriptive statistics, including MIC ranges, modal MIC (MM) (the most reported MIC), geometric mean MIC (GM), MIC at which 50% of the isolates was inhibited (MIC50), and MIC at which 90% of the isolates was inhibited (MIC90), were calculated for each species and drug. High off-scale MIC results were converted to the next highest concentration, and low off-scale MIC results were left unchanged. Epidemiological cutoff values (ECVs) were used to differentiate wild-type (WT) (without acquired resistance mechanisms) from non-wild-type (NWT) (may harbor acquired resistance mechanisms) isolates. For antifungal drugs, such as ibrexafungerp and caspofungin, or/and species, such as C. auris and C. orthopsilosis, without EUCAST ECVs, WT upper limits (WTUL) were used as the susceptibility cutoff values and MICs > 2 dilution steps above the MM were regarded as NWT (Table 1). Comparison of efficacy among antifungal drugs was based on MIC90 differences: Those antifungal drugs with a difference of one double dilution were considered to have a similar activity (Wiederhold, 2021). This comparison should not be considered absolute, as other pharmacokinetics and pharmacodynamics (PK/PD) parameters should be taken into account. The study was approved by the Ethics Committee of the Universidad del País Vasco/Euskal Herriko Unibertsitatea (UPV/EHU, Bilbao, Spain, CEIAB Ethics reference number M30_2015_248).
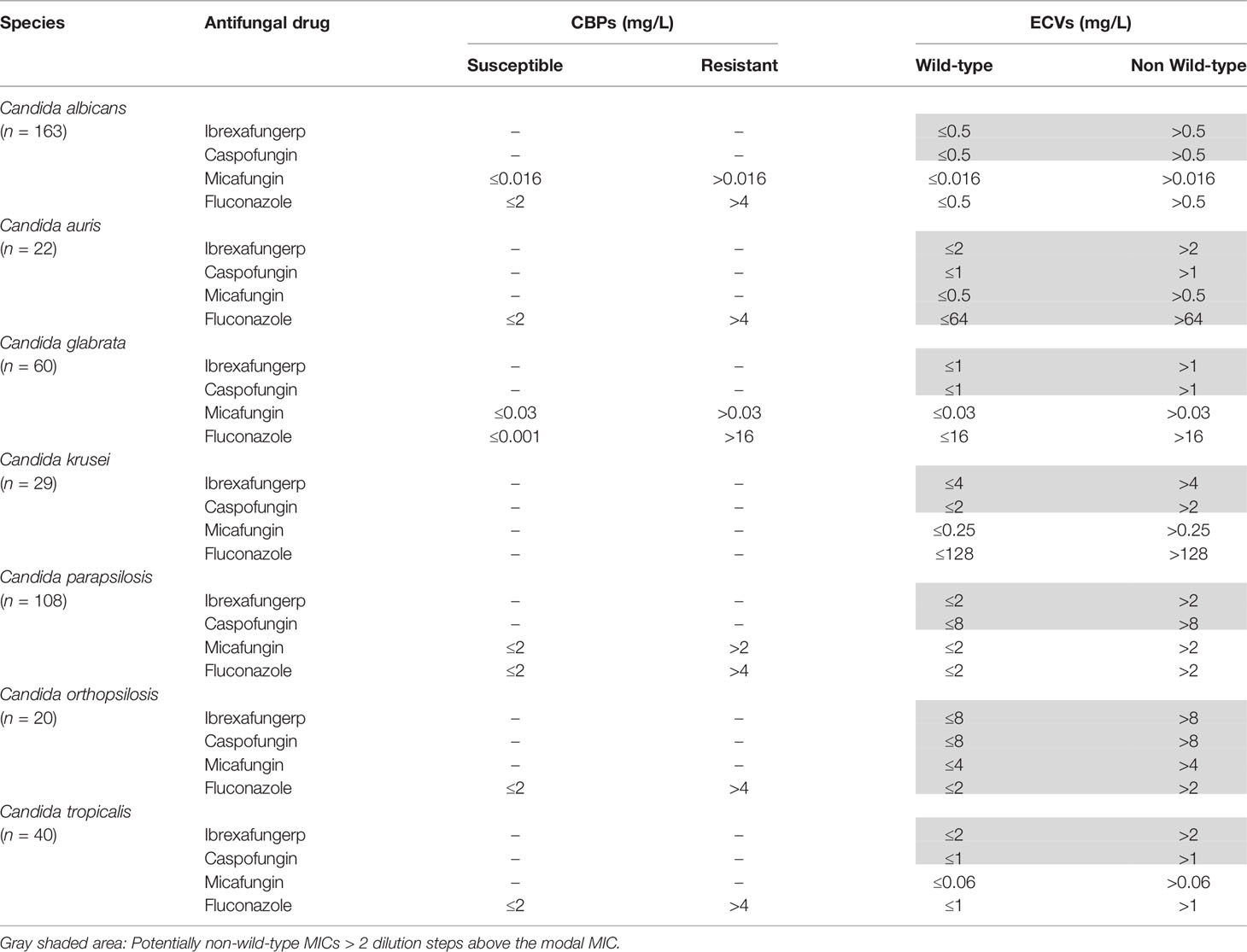
Table 1 EUCAST clinical breakpoints (CBPs) and epidemiological cutoff values (ECVs).
Results
MICs of ibrexafungerp against all species were tested in vitro ranging from 0.016 to ≥8 mg/L (MIC90 2 mg/L). These values were comparable to those of caspofungin and micafungin (MIC90 2 mg/L). The lowest ibrexafungerp MICs were observed for C. albicans (GM 0.062 mg/L, MIC range 0.016–0.5 mg/L, MIC90 0.125 mg/L) and the highest ibrexafungerp MICs were observed for C. tropicalis (GM 0.517 mg/L, MIC range 0.06–≥8 mg/L, MIC90 2 mg/L) (Table 2). Echinocandins MIC values for C. albicans ranged from ≤ 0.008 mg/L to 2 mg/L for micafungin (MM 0.008 mg/L, GM 0.001 mg/L, MIC90 0.016 mg/L) and from 0.016 mg/L to 2 mg/L for caspofungin (MM 0.125 mg/L, GM 0.104 mg/L, MIC90 0.125 mg/L). For the three isolates with elevated caspofungin or/and micafungin MICs, ibrexafungerp MIC range was 0.06 mg/L to 0.25 mg/L. In the current report, ibrexafungerp MICs ranged from 0.016 mg/L to 8 mg/L for 108 C. parapsilosis isolates (MM 0.5 mg/L, GM 0.660 mg/L, MIC90 4 mg/L) (Table 2). If we consider ibrexafungerp WTULs (>2 mg/L) obtained in this study, 11 C. parapsilosis isolates were NWT (4.6%). These NWT isolates were inhibited by ≤1 mg/L of fluconazole and by ≤4 mg/L of caspofungin or micafungin. We also observed that for six C. parapsilosis NWT (5.6%) and two resistant isolates to fluconazole (0.9%), the MIC range of ibrexafungerp was 0.5–2 mg/L. According to MIC90s, ibrexafungerp showed comparable values with caspofungin (MM 1 mg/L, GM 0.846 mg/L, MIC90 2 mg/L) and micafungin (MM 2 mg/L, GM 0.933 mg/L, MIC90 2 mg/L). Moreover, two isolates resistant to micafungin were inhibited by 2 mg/L of ibrexafungerp.
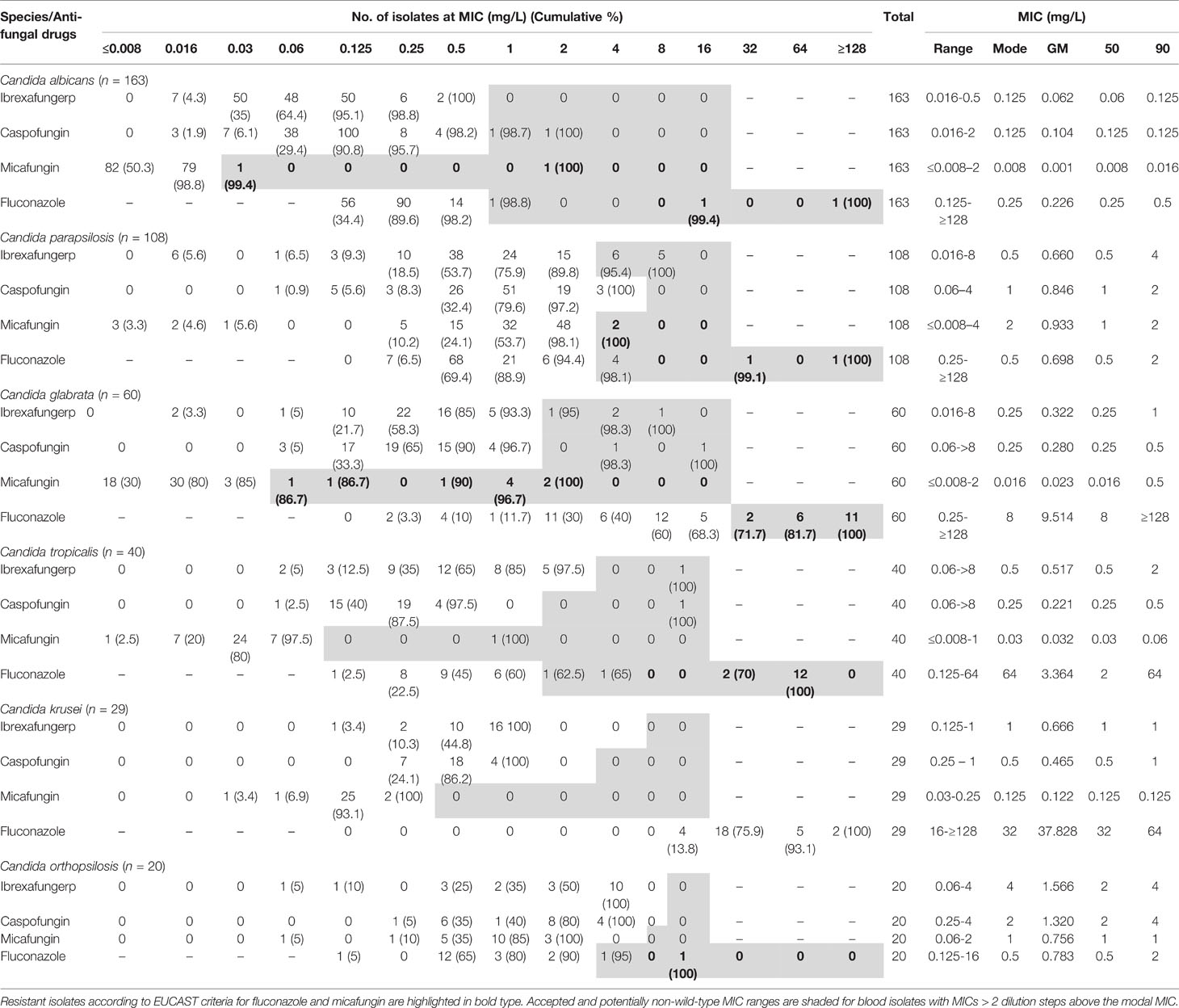
Table 2 Summary of ibrexafungerp and comparators’ in vitro antifungal activities against blood isolates from medically relevant species of Candida.
Ibrexafungerp also displayed potent in vitro activity against 60 C. glabrata isolates (MIC range 0.016–8 mg/L, MM 0.25 mg/L, GM 0.322 mg/L, MIC90 1 mg/L) (Table 2). Sixteen C. glabrata isolates resistant to fluconazole were inhibited by ≤1 mg/L of ibrexafungerp. Among 9 C. glabrata isolates with elevated MICs to caspofungin or/and micafungin, the MIC range for ibrexafungerp was 0.25 mg/L to 4 mg/L with a MIC50 of 1 mg/L. It was notable that whereas the increase in MM between C. glabrata WT and NWT isolates was 4-fold for caspofungin and 63-fold for micafungin, MM values for ibrexafungerp did not increase (Table 3).
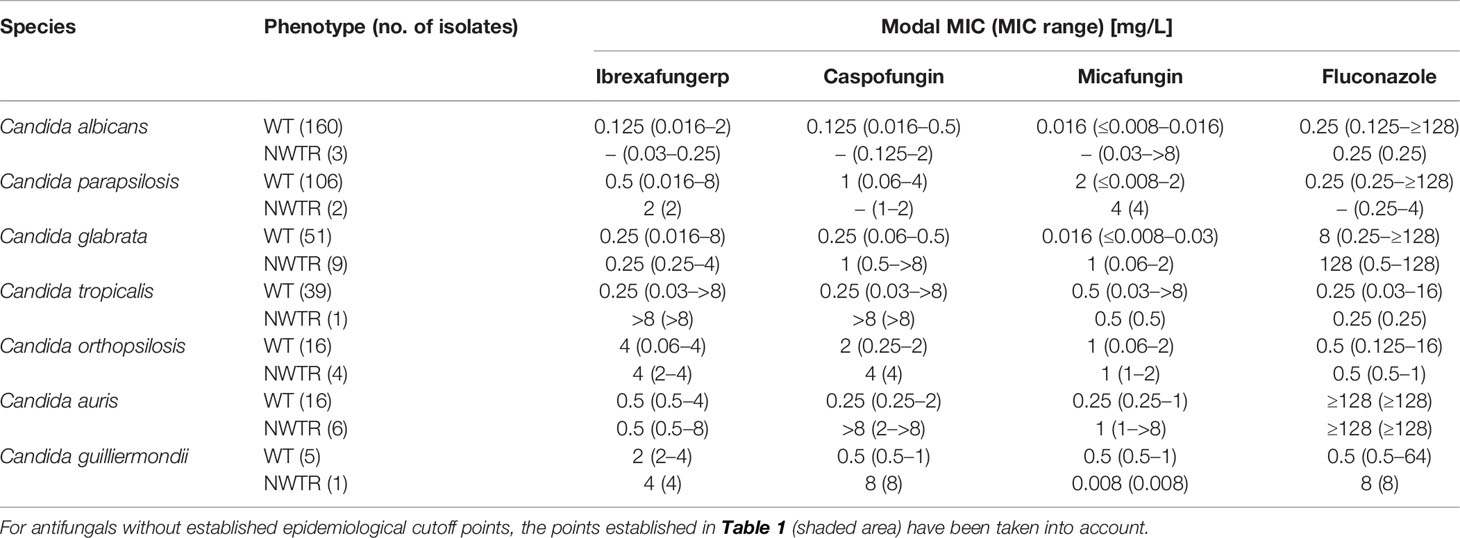
Table 3 MICs distribution of ibrexafungerp and comparator antifungal drugs against echinocandin wild-type (WT) and non-wild-type/resistant (NWTR) Candida spp. Isolates.
In vitro activity of ibrexafungerp was also observed against 40 C. tropicalis blood isolates and MICs ranged from 0.06 mg/L to ≥8 mg/L (MM 0.5 mg/L, GM 0.517 mg/L, MIC90 2 mg/L) (Table 2). Against 14 C. tropicalis resistant to fluconazole, ibrexafungerp MIC range was 0.25–2 mg/L (MIC90 1 mg/L).
Moreover, ibrexafungerp showed high in vitro activity against 29 C. krusei (MIC range 0.125–1 mg/L, MM 1 mg/L, GM 0.666 mg/L, MIC90 were 1 mg/L). MIC values of micafungin ranged from 0.03 mg/L to 0.25 mg/L (MM 0.125 mg/L, GM 0.122 mg/L, MIC90 0.125 mg/L) and from 0.25 mg/L to 1 mg/L for caspofungin (MM 0.5 mg/L, GM 0.465 mg/L, MIC90 1 mg/L). Additionally, all C. auris isolates were resistant in vitro to fluconazole (MIC ≥ 128 mg/L) while ibrexafungerp showed activity (MIC range 0.5 mg/L to 8 mg/L, MM 0.5 mg/L, GM 0.753 mg/L, MIC90 2 mg/L) (Table 4). C. auris urinary isolates showed higher MICs (data not shown). Against this species, the activity of ibrexafungerp was similar to the activity of micafungin (MIC range 0.125–>8 mg/L, MM 0.125 mg/L, GM 0.377 mg/L, MIC90 4 mg/L) and 8-fold more active than caspofungin (MIC range 0.25–>8 mg/L, MM 0.25 mg/L, GM 0.585 mg/L, MIC90 >8 mg/L). Among six isolates with elevated caspofungin or/and micafungin MICs, ibrexafungerp MICs ranged from 0.5 mg/L to 8 mg/L (MIC50 0.5 mg/L).
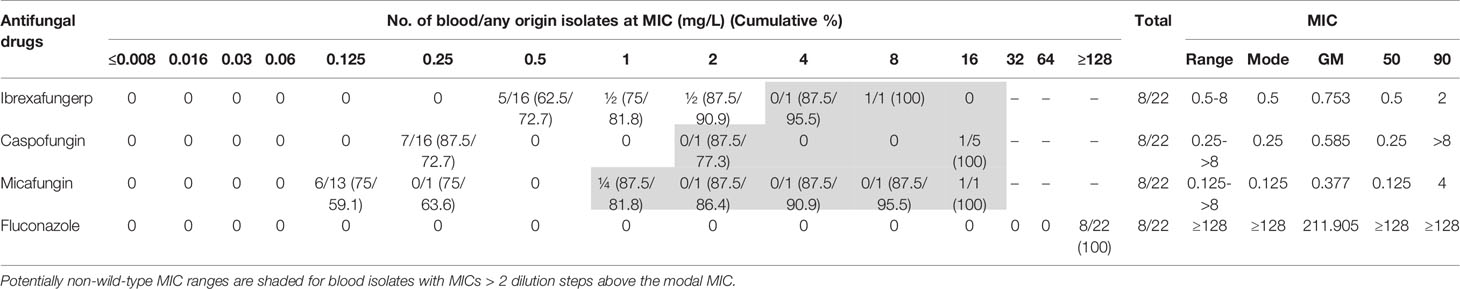
Table 4 Summary of ibrexafungerp and comparators’ in vitro antifungal activities against 22 clinical isolates of Candida auris.
MICs of ibrexafungerp were ≤ 1 mg/L for the 2 C. famata isolates and 1 isolate each of C. bracarensis, C. catenulata, C. dubliniensis, and C. kefyr (Table 5). Ibrexafungerp MIC50 was 2 mg/L for the 6 C. guilliermondii blood isolates with a MIC range from 2 to 4 mg/L, being twofold less active than caspofungin or micafungin.
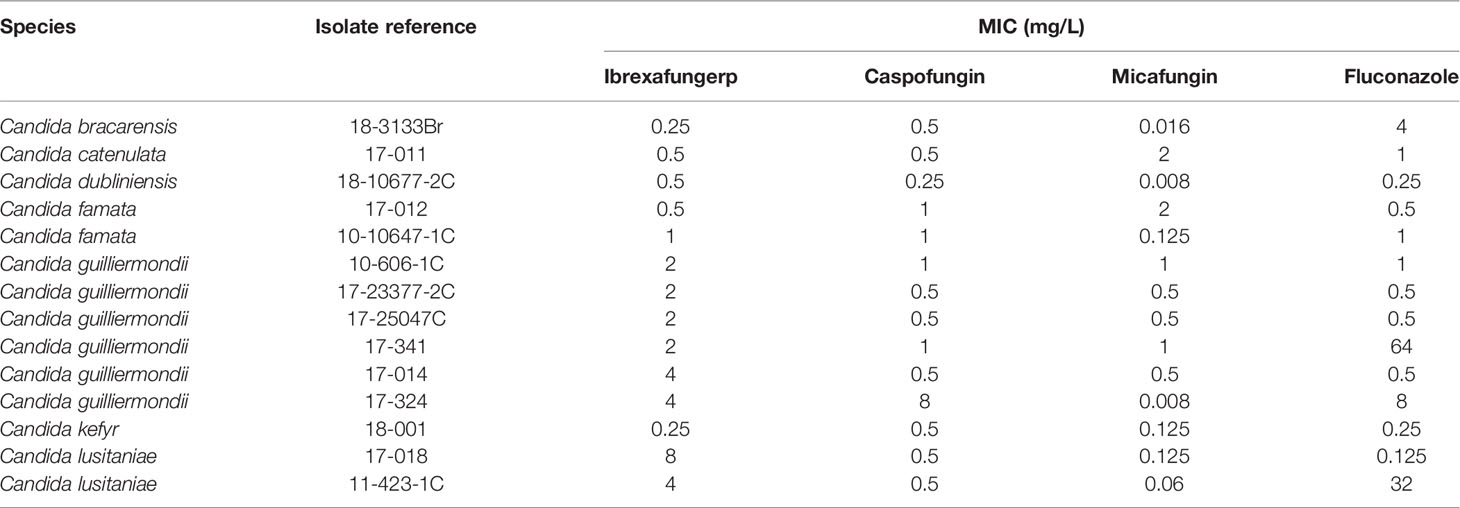
Table 5 Activities of ibrexafungerp and comparator antifungal drugs against other species of Candida blood isolates.
Discussion
The present study aimed to evaluate the in vitro activity of ibrexafungerp against a collection of 434 European blood isolates of Candida. Ibrexafungerp showed a potent in vitro activity against Candida, with MICs ranging from 0.016 to 16 mg/L. The lowest ibrexafungerp MICs were observed against C. albicans and the highest ibrexafungerp MICs were observed against C. tropicalis Moreover, ibrexafungerp also displayed remarkable activity against fluconazole- and echinocandin-resistant isolates.
Five species of Candida (C. albicans, C. parapsilosis, C. glabrata, C. tropicalis, and C. krusei) cause more than 90% of IC (Quindós, 2014; Arendrup and Patterson, 2017; Quindós et al., 2018). C. albicans remains the predominant cause, but there is an evident shift in the etiology, and IC caused by other species less susceptible to current antifungal drugs is becoming more frequent (Fuller et al., 2019; Pfaller et al., 2019). Echinocandins, such as anidulafungin, caspofungin, and micafungin, are considered first-line therapy for IC because they possess fungicidal activity against Candida (Gil-Alonso et al., 2015a; Gil-Alonso et al., 2015b). However, emergence of azole- and echinocandin-resistant [multidrug-resistant (MDR)] isolates has been reported for C. auris, C. glabrata, C. guilliermondii, C. lusitaniae, and C. parapsilosis (Lortholary et al., 2011; Pemán et al., 2012; Pfaller et al., 2012; Alexander et al., 2013; Pham et al., 2014; Dudiuk et al., 2017; Ruiz-Gaitán et al., 2019; Tortorano et al., 2021). MDR Candida isolates complicate clinical decision-making and are associated with treatment failure and high mortality rates (Lortholary et al., 2011; Alexander et al., 2013; Shields et al., 2013; Quindós et al., 2018; Tortorano et al., 2021).
Ibrexafungerp is a novel orally bioavailable semi-synthetic derivative of the terpenoid enfumafungin that inhibits glucan synthase, decreasing 1,3-β-D-glucan polymers and weakening fungal cell wall (Jiménez-Ortigosa et al., 2014; Pfaller et al., 2017; Schell et al., 2017; Davis et al., 2020). The current study that has evaluated European blood isolates confirms and extends the observations published in previous studies (Pfaller et al., 2013; Jiménez-Ortigosa et al., 2014; Berkow et al., 2017; Larkin et al., 2017; Marcos-Zambrano et al., 2017; Pfaller et al., 2017; Schell et al., 2017; Nunnally et al., 2019; Gamal et al., 2021). However, these previous studies have mostly tested American isolates. Our study with European isolates is in line with the conclusion that ibrexafungerp displays potent in vitro activity against the most clinically relevant species of Candida.
Ibrexafungerp showed an excellent activity against the C. albicans isolates included in the present study (MICs range 0.016–0.5 mg/L). These results confirm previous findings by other authors, such as Jiménez-Ortigosa et al. (2014); Schell et al. (2017), and Mesquida et al. (2021), demonstrating that ibrexafungerp showed activity against most FKS-mediated echinocandin-resistant C. albicans and against azole-resistant C. albicans.
Although there are no clinical breakpoints (CBPs) or ECVs available for ibrexafungerp, the study by Mesquida et al. (2022) as well as the present work have proposed WTULs. There are no differences in the proposed limits for C. glabrata (1 mg/L). However, our study suggests higher WTUL for C. albicans (0.5 mg/L vs. 0.25 mg/L by Mesquida et al.), for C. parapsilosis and C. tropicalis (2 mg/L vs. 1 mg/L by Mesquida et al.), and for C. krusei (4 mg/L vs. 2 mg/L by Mesquida et al.).
C. parapsilosis is the first or second etiology of IC in China, Japan, Latin America, and the Mediterranean countries of Africa, Asia, and Europe, such as Italy, Portugal, and Spain (Quindós et al., 2018). In the current report, ibrexafungerp MICs ranged from 0.016 mg/L to 8 mg/L. Our results are in accordance to those by Marcos-Zambrano et al. (2017). In both studies, ibrexafungerp displayed remarkably lower MICs than micafungin against C. parapsilosis. Although Mesquida et al. (2022) did not differentiate among C. parapsilosis species complex, they also reported lower MICs for ibrexafungerp than for echinocandins. These high MIC values reported for echinocandins against C. parapsilosis have been associated with substitutions in the hs1 region of FKS1 (Garcia-Effron et al., 2008; Dudiuk et al., 2017; Marcos-Zambrano et al., 2017; Mesquida et al., 2022). Schell et al. (2017) also reported lower ibrexafungerp MIC90 values for 19 C. parapsilosis blood isolates (0.25 mg/L) compared with echinocandins, suggesting that changes in FKS1 may not affect the capacity of ibrexafungerp to inhibit glucan synthase in this species.
C. glabrata has increased its etiological importance in IC in Australia, Canada, the USA, and countries in Central and Northern Europe, such as Belgium and Germany (Quindós, 2014; Trouvé et al., 2017; Quindós et al., 2018). In our study, ibrexafungerp displayed potent activity (MIC range 0.016–8 mg/L) also against fluconazole-resistant isolates. No cross-resistance has been found between ibrexafungerp and fluconazole, as previously noted by Marcos-Zambrano et al. (2017). The incidence of echinocandin resistance in C. glabrata is generally considered low, approximately 3%–4%, but can be as high as 30% in specific institutions (Alexander et al., 2013; Pham et al., 2014; Arendrup and Patterson, 2017). In the current study, ibrexafungerp MIC was ≤4 mg/L against four C. glabrata resistant to both fluconazole and micafungin. Pfaller et al. (2013) found similar results to ours but, in their report, ibrexafungerp was 8-fold more active than caspofungin against C. glabrata. Furthermore, ibrexafungerp showed activity against 31 C. glabrata strains with mutations in the hs of FKS1 or FKS2 (MIC ≤ 2 mg/L) and against 14 strains resistant to both caspofungin and fluconazole. In a later study by the same authors (Pfaller et al., 2017), 20 out of 25 FKS mutant C. glabrata isolates (80%) were NWT to one or more echinocandins, but only six (24%) were NWT to ibrexafungerp. Isolates of C. glabrata for which the ibrexafungerp MIC was > 2 mg/L (NWT) all were NWT and either intermediate or resistant to anidulafungin, caspofungin, and micafungin. In our study, 3 out of 4 isolates potentially NWT for ibrexafungerp were inhibited by ≤1 mg/L of caspofungin and one by 0.03 mg/L of micafungin. However, three of these four ibrexafungerp NWT isolates were resistant to micafungin.
Schell et al. (2017) evaluated 34 echinocandin-resistant C. glabrata isolates along with 34 paired control C. glabrata isolates, observing that ibrexafungerp MICs for individual C. glabrata isolates tended to be three to five dilutions higher than those for the echinocandins. However, ibrexafungerp MICs trended in agreement with those for the echinocandins. These authors detected that C. glabrata isolates with FKS1 or FKS2 mutations or echinocandin resistance were inhibited by ≤4 mg/L of ibrexafungerp. Nunnally et al. (2019) also reported good ibrexafungerp activity against 89 C. glabrata isolates with FKS1 or FKS2 mutations that conferred resistance to at least one echinocandin. Ibrexafungerp MIC values ranged from <0.03 mg/L to 4 mg/L while caspofungin and micafungin MICs ranged from 0.03 to >16 mg/L and 0.008 to >16 mg/L, respectively. In the study by Mesquida et al. (2022), an isolate of C. glabrata that displayed an ibrexafungerp MIC of 2 mg/L and echinocandin NWT phenotype harbored a mutation at FKS2. The spectrum of resistance mutations found in C. glabrata suggested a partially overlapping but independent binding site for ibrexafungerp relative to echinocandins on glucan synthase as these drugs are structurally dissimilar and interact differently with the target (Jiménez-Ortigosa et al., 2014). Consequently, this potent in vitro activity for ibrexafungerp has been reported against C. glabrata isolates harboring FKS1 and FKS2 point mutations that cause echinocandin resistance (Marcos-Zambrano et al., 2017; Pfaller et al., 2017; Schell et al., 2017; Nunnally et al., 2019; Mesquida et al., 2022). Moreover, the in vitro efficacy of ibrexafungerp has been supported by successful treatments in murine IC caused by C. glabrata resistant to echinocandins (Lepak et al., 2015; Wiederhold et al., 2018). In a murine model of IC caused by C. albicans, C. parapsilosis, or C. glabrata using an oral therapy with ibrexafungerp, Lepak et al. (2015) demonstrated that the AUC/MIC was the best pharmacodynamics parameter predicting clinical response. A MIC ≤ 1 mg/L obtained by CLSI would predict a clinical response using oral ibrexafungerp (Marcos-Zambrano et al., 2017).
In Asia, C. tropicalis was the second etiological agent of IC in many hospitals from China, India, Singapore, Thailand, and Taiwan (Quindós et al., 2018). We observed that ibrexafungerp MICs ranged from 0.06 mg/L to ≥ 8 mg/L for 40 C. tropicalis blood isolates, which confirms the consistently low ibrexafungerp MICs for azole-resistant C. tropicalis reported by Schell et al. (2017). Mesquida et al. (2022) found mutations in the FKS1 gene of two NWT isolates of C. tropicalis that resulted, in both cases, in ibrexafungerp MICs between 0.5 and 1 mg/L.
We observed high activity of ibrexafungerp against 29 C. krusei (MIC range 0.125–1 mg/L). Schell et al. (2017) reported ibrexafungerp MICs for six isolates of C. krusei (MIC range from 0.5 mg/L to 4 mg/L), which were higher than those for the other Candida species. These authors suggested that a naturally occurring unidentified substitution may be responsible for the reduced activity of ibrexafungerp on glucan synthase in C. krusei. However, it is unknown if this translates into clinical failure.
In a recent Spanish nationwide study on candidemia, C. orthopsilosis was the fifth most frequently isolated species, preceding C. krusei (Pemán et al., 2012). In the present study, ibrexafungerp showed good activity against 20 C. orthopsilosis (MIC range 0.06–4 mg/L). To our knowledge, this is the first study on in vitro activity of ibrexafungerp against C. orthopsilosis. The lack of previous reports in this regard precludes comparison. Differences in ibrexafungerp activity against the closely related species C. orthopsilosis and C. parapsilosis highlights the importance of a correct identification of the Candida species involved in invasive infections.
C. auris is an emerging pathogen that has been identified in many countries associated with high mortality and a marked ability to develop resistance to multiple commonly used antifungal agents and to withstand standard infection control practices (Larkin et al., 2017; Ruiz-Gaitán et al., 2017; Ruiz-Gaitán et al., 2018; Ruiz-Gaitán et al., 2019). Data from Larkin et al. (2017) show that the C. auris isolates exhibited multidrug resistance against fluconazole and amphotericin B. Moreover, some isolates also exhibited high MIC values for voriconazole and itraconazole. In the present report, all C. auris isolates were resistant to fluconazole while ibrexafungerp showed notable activity (MIC range 0.5 mg/L to 8 mg/L). The in vitro activity of ibrexafungerp against European C. auris isolates from our study was similar to that described by Berkow et al. (2017) who studied 100 isolates of this species from Asia, Africa, and America. Of note, in the current study, MM values between C. auris WT and NWT isolates increased 64-fold for caspofungin and 4-fold for micafungin, whereas MM values for ibrexafungerp did not change. Interestingly, ibrexafungerp has been shown to be effective in vivo in the treatment of experimental infections with C. auris (Wiederhold et al., 2021).
Ibrexafungerp was twofold less active than caspofungin or micafungin against six C. guilliermondii blood isolates. Accordingly, Mesquida et al. (2022) also found higher ibrexafungerp MICs for 12 C. guilliermondii isolates, from 0.125 to 8 mg/L. Naturally occurring high echinocandins MICs against C. guilliermondii have been recognized since these antifungal agents were first introduced probably in association to substitutions in the hs1 region of FKS1 (Dudiuk et al., 2017; Schell et al., 2017). In addition, Schell et al. (2017) reported that ibrexafungerp MIC ranges for three C. lusitaniae blood isolates were 1 to 2 mg/L and ibrexafungerp MICs for C. lusitaniae were three to five dilutions higher than those for the other species of Candida. Ibrexafungerp MICs for the two isolates of C. lusitaniae (MICs 4 and 8 mg/L) tested in the current study were higher than those observed for the other Candida species.
Aside from emerging resistance, an important limitation of echinocandins is that they must be administered daily by intravenous infusion, potentially prolonging hospital stays for patients undergoing echinocandin therapy and limiting them to inpatient settings in most instances. Of note, Wring et al. (2018) reported that the risk for interactions of ibrexafungerp with drugs metabolized via the cytochrome P450 family of enzymes is low. Ibrexafungerp exhibited concentration- and time-dependent fungicidal activity against C. albicans, C. glabrata, C. krusei, C. parapsilosis, and C. tropicalis in time-kill curve studies (Scorneaux et al., 2017). Moreover, ibrexafungerp has demonstrated efficacy for IC in phase 2 and 3 clinical studies (Spec et al., 2019).
The current study demonstrates that ibrexafungerp shows potent in vitro activity against Candida blood isolates and its activity is comparable to that of micafungin. Ibrexafungerp even exhibits good activity against fluconazole-resistant Candida isolates. Moreover, echinocandin-resistant isolates exhibit ibrexafungerp MICs consistent with those of echinocandin-susceptible isolates. However, direct comparisons of ibrexafungerp MICs with those of other antifungal drugs should be interpreted with caution, as different drugs may produce diverse ranges of MICs and yet have equivalent clinical efficacy because of their differences in bioavailability and in PK/PD properties. Although Pfaller et al. (2013) reported >90% essential agreement between both methods, CLSI and EUCAST, the comparison between our results and those of American authors should consider that the EUCAST method tends to yield higher MICs than CLSI, regardless of the studied species. Although the most clinically relevant species of Candida have been included in the current study, it would be interesting to assess the activity of ibrexafungerp against additional species as well as more NWT isolates.
In conclusion, we demonstrated that ibrexafungerp, a potent inhibitor of glucan synthase, could be an important acquisition to the antifungal toolbox for the therapy of patients suffering from IC caused by MDR species, such as C. auris, C. glabrata, or C. krusei. Considering the excellent pharmacokinetic properties of ibrexafungerp (oral availability and excellent tissue distribution and concentrations) as well as the potent activity observed against the main species causing candidiasis, ibrexafungerp should be regarded as a potential candidate for the therapy of these important diseases.
Data Availability Statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author Contributions
GQ, KM-C, and RS-M designed the research and participated in manuscript writing. KB-E, EC, MJL-S, AH, IM, AT, AP, MV-G, CM-A, AG, FS-R, JM-B, MR-I, EM-M, CC-M, LL-S, AR-G, MF-R, DL, JC, AR, JaP, JG, JoP, CP, OR, GE, NJ, DA, and EE conducted sampling and clinical measures, carried out the experiments, analyzed data, and drafted the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This study received funding from SCYNEXIS. The funder was not involved in the study design, collection, analysis, interpretation of data, the writing of the article, or the decision to submit it for publication. CM-A is a recipient of a grant from Fundación ONCE (Oportunidad al Talento). EE, AG, NJ, CM-A, and GQ have received grant support from Consejería de Educación, Universidades e Investigación del Gobierno Vasco (GIC15 IT-990-16), Fondo de Investigación Sanitaria del Gobierno de España (FIS PI11/00203), and UPV/EHU (UFI 11/25). All authors declare no other competing interests.
Conflict of Interest
Outside the current study, we declare the following potential conflicts: GQ has received research grants from Astellas Pharma, Pfizer, Merck Sharp & Dohme, and SCYNEXIS. GQ has served on advisory/consultant boards for Merck, Sharp & Dohme, and SCYNEXIS, and he has received speaker honoraria from Abbvie, Astellas Pharma, Merck Sharp & Dohme, Pfizer, and SCYNEXIS. KB-E and DA are employed by SCYNEXIS.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2022.906563/full#supplementary-material
Abbreviations
ATCC, American Type Culture Collection; CBP, clinical breakpoints; DMSO, dimethyl sulfoxide; ECV, epidemiological cutoff values; EUCAST, European Committee on Antimicrobial Susceptibility Testing; GM, geometric mean; IC, invasive candidiasis; ITS, internal transcribed spacer; MALDI-TOF, matrix-assisted laser desorption/ionization–time-of-flight; MM, modal MIC; MIC, minimum inhibitory concentration; MOPS, morpholinopropane sulphate; NWT, non-wild type; PK/PD, pharmacokinetics and pharmacodynamics; QC, quality control; UPV/EHU, Universidad del País Vasco/Euskal Herriko Unibertsitatea; WT, wild type; WTUL, wild-type upper limits.
References
Alexander, B. D., Johnson, M. D., Jiménez-Ortigosa, C., Catania, J., Booker, R., Castanheira, M., et al. (2013). Increasing Echinocandin Resistance in Candida Glabrata: Clinical Failure Correlates With Presence of FKS Mutations and Elevated Minimum Inhibitory Concentrations. Clin. Infect. Dis. 56, 1724–1732. doi: 10.1093/cid/cit136
Arendrup, M. C., Friberg, N., Mares, M., Kahlmeter, G., Meletiadis, J., Guinea, J. (2020a). How to Interpret MICs of Antifungal Compounds According to the Revised Clinical Breakpoints V. 10.0 European Committee on Antimicrobial Susceptibility Testing (EUCAST). Clin. Microbiol. Infect. 26, 1464–1472. doi: 10.1016/j.cmi.2020.06.007
Arendrup, M. C., Meletiadis, J., Mouton, J. W., Lagrou, K., Hamal, P., Guinea, J. (2020b) EUCAST Definitive Document E.DEF 7.3.2 Method for the Determination of Broth Dilution Minimum Inhibitory Concentrations of Antifungal Agents for Yeasts. Available at: https://www.eucast.org/astoffungi/methodsinantifungalsusceptibilitytesting/susceptibility_testing_of_yeasts/ (Accessed 07 March 2022).
Arendrup, M. C., Patterson, T. F. (2017). Multidrug-Resistant Candida: Epidemiology, Molecular Mechanisms, and Treatment. J. Infect. Dis. 2163), S445–S451. doi: 10.1093/infdis/jix131
Berkow, E. L., Angulo, D., Lockhart, S. R. (2017). In Vitro Activity of a Novel Glucan Synthase Inhibitor, SCY-078, Against Clinical Isolates of Candida Auris. Antimicrob. Agents Chemother. 61, e00435-17. doi: 10.1128/AAC.00435-17
Davis, M. R., Donnelley, M. A., Thompson, G. R. (2020). Ibrexafungerp: A Novel Oral Glucan Synthase Inhibitor. Med. Mycol. 58, 579–592. doi: 10.1093/mmy/myz083
Dudiuk, C., Macedo, D., Leonardelli, F., Theill, L., Cabeza, M. S., Gamarra, S., et al. (2017). Molecular Confirmation of the Relationship Between Candida Guilliermondii Fks1p Naturally Occurring Amino Acid Substitutions and Its Intrinsic Reduced Echinocandin Susceptibility. Antimicrob. Agents Chemother. 61, e02644-16. doi: 10.1128/AAC.02644-16
Espinel-Ingroff, A., Arendrup, M. C., Pfaller, M. A., Bonfietti, L. X., Bustamante, B., Canton, E., et al. (2013). Interlaboratory Variability of Caspofungin MICs for Candida Spp. Using CLSI and EUCAST Methods: Should the Clinical Laboratory Be Testing This Agent? Antimicrob. Agents Chemother. 57, 5836–5842. doi: 10.1128/AAC.01519-13
EUCAST (2020) European Committee on Antimicrobial Susceptibility Testing Breakpoint. Antifungal Agents: Tables for Interpretation of MICs Version 10.0. Available at: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/AFST/Clinical_breakpoints/AFST_BP_v10.0_200204_updatd_links_200924.pdf (Accessed March 2022).
EUCAST (2021) MIC and Zone Diameter Distributions and ECOFFs. Available at: https://mic.eucast.org/search/ (Accessed March 2022).
Fuller, J., Dingle, T. C., Bull, A., Shokoples, S., Laverdière, M., Baxter, M. R., et al. (2019). Species Distribution and Antifungal Susceptibility of Invasive Candida Isolates From Canadian Hospitals: Results of the CANWARD 2011-16 Study. J. Antimicrob. Chemother. 74 (Suppl.4), iv48–iv54. doi: 10.1093/jac/dkz287
Gamal, A., Chu, S., McCormick, T. S., Borroto-Esoda, K., Angulo, D. (2021). And Ghannoum, MIbrexafungerp, A Novel Oral Triterpenoid Antifungal in Development: Overview of Antifungal Activity Against Candida Glabrata. A Front. Cell Infect. Microbiol. 11. doi: 10.3389/fcimb.2021.642358
Garcia-Effron, G., Katiyar, S. K., Park, S., Edlind, T. D., Perlin, D. S. (2008). A Naturally Occurring Proline-to-Alanine Amino Acid Change in Fks1p in Candida Parapsilosis, Candida Orthopsilosis, and Candida Metapsilosis Accounts for Reduced Echinocandin Susceptibility. Antimicrob. Agents Chemother. 52, 2305–2312. doi: 10.1128/AAC.00262-08
Gil-Alonso, S., Jauregizar, N., Cantón, E., Eraso, E., Quindós, G. (2015a). Comparison of the In Vitro Activity of Echinocandins Against Candida Albicans, Candida Dubliniensis, and Candida Africana by Time-Kill Curves. Diagn. Microbiol. Infect. Dis. 82, 57–61. doi: 10.1016/j.diagmicrobio.2015.01.010
Gil-Alonso, S., Jauregizar, N., Cantón, E., Eraso, E., Quindós, G. (2015b). In Vitro Fungicidal Activities of Anidulafungin, Caspofungin, and Micafungin Against Candida Glabrata, Candida Bracarensis, and Candida Nivariensis Evaluated by Time-Kill Studies. Antimicrob. Agents Chemother. 59, 3615–3618. doi: 10.1128/AAC.04474-14
Jiménez-Ortigosa, C., Paderu, P., Motyl, M. R., Perlin, D. S. (2014). Enfumafungin Derivative MK-3118 Shows Increased In Vitro Potency Against Clinical Echinocandin-Resistant Candida Species and Aspergillus Species Isolates. Antimicrob. Agents Chemother. 58, 1248–1251. doi: 10.1128/AAC.02145-13
Jiménez-Ortigosa, C., Perez, W. B., Angulo, D., Borroto-Esoda, K., Perlin, D. S. (2017). De Novo Acquisition of Resistance to SCY-078 in Candida Glabrata Involves FKS Mutations That Both Overlap and Are Distinct From Those Conferring Echinocandin Resistance. Antimicrob. Agents Chemother. 61, e00833–e00817. doi: 10.1128/AAC.00833-17
Larkin, E., Hager, C., Chandra, J., Mukherjee, P. K., Retuerto, M., Salem, I., et al. (2017). The Emerging Pathogen Candida Auris: Growth Phenotype, Virulence Factors, Activity of Antifungals, and Effect of SCY-078, A Novel Glucan Synthesis Inhibitor, on Growth Morphology and Biofilm Formation. Antimicrob. Agents Chemother. 61, e02396-16. doi: 10.1128/AAC.02396-16
Lepak, A. J., Marchillo, K., Andes, D. R. (2015). Pharmacodynamic Target Evaluation of a Novel Oral Glucan Synthase Inhibitor, SCY-078 (MK-3118), Using an In Vivo Murine Invasive Candidiasis Model. Antimicrob. Agents Chemother. 59, 1265–1272. doi: 10.1128/AAC.04445-14
Lortholary, O., Desnos-Ollivier, M., Sitbon, K., Fontanet, A., Bretagne, S., Dromer, F., et al. (2011). Recent Exposure to Caspofungin or Fluconazole Influences the Epidemiology of Candidemia: A Prospective Multicenter Study Involving 2,441 Patients. Antimicrob. Agents Chemother. 55, 532–538. doi: 10.1128/AAC.01128-10
Marcos-Zambrano, L. J., Gómez-Perosanz, M., Escribano, P., Bouza, E., Guinea, J. (2017). The Novel Oral Glucan Synthase Inhibitor SCY-078 Shows In Vitro Activity Against Sessile and Planktonic Candida Spp. J. Antimicrob. Chemother. 72, 1969–1976. doi: 10.1093/jac/dkx010
Mesquida, A., Díaz-García, J., Sánchez-Carrillo, C., Muñoz, P., Escribano, P., Guinea, J. (2022). In Vitro Activity of Ibrexafungerp Against Candida Species Isolated From Blood Cultures. Determination of Wild-Type Populations Using the EUCAST Method. Clin. Microbiol. Infect. 28, 140.e1–140.e4. doi: 10.1016/j.cmi.2021.09.030
Mesquida, A., Vicente, T., Reigadas, E., Palomo, M., Sánchez-Carrillo, C., Muñoz, P., et al. (2021). In Vitro Activity of Ibrexafungerp and Comparators Against Candida Albicans Genotypes From Vaginal Samples and Blood Cultures. Clin. Microbiol. Infect. 27, 915.e5–915.e8. doi: 10.1016/j.cmi.2021.02.006
Miranda-Zapico, I., Eraso, E., Hernández-Almaraz, J. L., López-Soria, L. M., Carrillo-Muñoz, A. J., Hernández-Molina, J. M., et al. (2011). Prevalence and Antifungal Susceptibility Patterns of New Cryptic Species Inside the Species Complexes Candida Parapsilosis and Candida Glabrata Among Blood Isolates From a Spanish Tertiary Hospital. J. Antimicrob. Chemother. 66, 2315–2322. doi: 10.1093/jac/dkr298
Nunnally, N. S., Etienne, K. A., Angulo, D., Lockhart, S. R., Berkow, E. L. (2019). In Vitro Activity of Ibrexafungerp, A Novel Glucan Synthase Inhibitor Against Candida Glabrata Isolates With FKS Mutations. Antimicrob. Agents Chemother. 63, e01692-19. doi: 10.1128/AAC.01692-19
Pemán, J., Cantón, E., Quindós, G., Eraso, E., Alcoba, J., Guinea, J., et al. (2012). Epidemiology, Species Distribution, and In Vitro Antifungal Susceptibility of Fungaemia in a Spanish Multicentre Prospective Survey. J. Antimicrob. Chemother. 67, 1181–1187. doi: 10.1093/jac/dks019
Pfaller, M. A., Castanheira, M., Lockhart, S. R., Ahlquist, A. M., Messer, S. A., Jones, R. N. (2012). Frequency of Decreased Susceptibility and Resistance to Echinocandins Among Fluconazole-Resistant Bloodstream Isolates of Candida Glabrata. J. Clin. Microbiol. 50, 1199–1203. doi: 10.1128/JCM.06112-11.O
Pfaller, M. A., Diekema, D. J., Turnidge, J. D., Castanheira, M., Jones, R. N. (2019). Twenty Years of the SENTRY Antifungal Surveillance Program: Results for Candida Species From 1997-2016. Open Forum Infect. Dis. 61), S79–S94. doi: 10.1093/ofid/ofy358
Pfaller, M. A., Messer, S. A., Motyl, M. R., Jones, R. N., Castanheira, M. (2013). Activity of MK-3118, A New Oral Glucan Synthase Inhibitor, Tested Against Candida Spp. By Two International Methods (CLSI and EUCAST). J. Antimicrob. Chemother. 68, 858–863. doi: 10.1093/jac/dks466
Pfaller, M. A., Messer, S. A., Rhomberg, P. R., Borroto-Esoda, K., Castanheira, M. (2017). Differential Activity of the Oral Glucan Synthase Inhibitor SCY-078 Against Wild-Type and Echinocandin-Resistant Strains of Candida Species. Antimicrob. Agents Chemother. 61, e00161-17. doi: 10.1128/AAC.00161-17
Pham, C. D., Iqbal, N., Bolden, C. B., Kuykendall, R. J., Harrison, L. H., Farley, M. M., et al. (2014). The Role of FKS Mutations in C. Glabrata: MIC Values, Echinocandin Resistance and Multidrug Resistance. Antimicrob. Agents Chemother. 58, 4690–4696. doi: 10.1128/AAC.03255-15
Quindós, G. (2014). Epidemiology of Candidaemia and Invasive Candidiasis. A Changing Face. Rev. Iberoam. Micol. 31, 42–48. doi: 10.1016/j.riam.2013.10.001
Quindós, G., Marcos-Arias, C., San-Millán, R., Mateo, E., Eraso, E. (2018). The Continuous Changes in the Aetiology and Epidemiology of Invasive Candidiasis: From Familiar Candida Albicans to Multiresistant Candida Auris. Int. Microbiol. 21, 107–119. doi: 10.1007/s10123-018-0014-1
Ruiz-Gaitán, A. C., Cantón, E., Fernández-Rivero, M. E., Ramírez, P., Pemán, J. (2019). Outbreak of Candida Auris in Spain: A Comparison of Antifungal Activity by Three Methods With Published Data. Int. J. Antimicrob. Agents 53, 541–546. doi: 10.1016/j.ijantimicag.2019.02.005
Ruiz-Gaitán, A. C., Moret, A., López Hontangas, J. L., Molina, J. M., Aleixandre López, A. I., Cabezas, A. H., et al. (2017). Nosocomial Fungemia by Candida Auris: First Four Reported Cases in Continental Europe. Rev. Iberoam. Micol. 34, 23–27. doi: 10.1016/j.riam.2016.11.002
Ruiz-Gaitán, A., Moret, A. M., Tasias-Pitarch, M., Aleixandre-López, A. I., Martínez-Morel, H., Calabuig, E., et al. (2018). An Outbreak Due to Candida Auris With Prolonged Colonisation and Candidaemia in a Tertiary Care European Hospital. Mycoses 61, 498–505. doi: 10.1111/myc.12781
Schell, W. A., Jones, A. M., Borroto-Esoda, K., Alexander, B. D. (2017). Antifungal Activity of SCY-078 and Standard Antifungal Agents Against 178 Clinical Isolates of Resistant and Susceptible Candida Species. Antimicrob. Agents Chemother. 61, e01102-17. doi: 10.1128/AAC.01102-17
Schwebke, J. R., Sobel, R., Gersten, J. K., Sussman, S. A., Lederman, S. N., Jacobs, M. A., et al. (2021). Ibrexafungerp Versus Placebo for Vulvovaginal Candidiasis Treatment: A Phase 3, Randomized, Controlled Superiority Trial (VANISH 303). Clin. Infect. Dis. Ciab. 750. doi: 10.1093/cid/ciab750
Scorneaux, B., Angulo, D., Borroto-Esoda, K., Ghannoum, M., Peel, M., Wring, S. (2017). SCY-078 Is Fungicidal Against Candida Species in Time-Kill Studies. Antimicrob. Agents Chemother. 61, e01961-16. doi: 10.1128/AAC.01961-16
Shields, R. K., Nguyen, M. H., Press, E. G., Updike, C. L., Clancy, C. J. (2013). Caspofungin MICs Correlate With Treatment Outcomes Among Patients With Candida Glabrata Invasive Candidiasis and Prior Echinocandin Exposure. Antimicrob. Agents Chemother. 57, 3528–3535. doi: 10.1128/AAC.00136-13
Sobel, R., Nyirjesy, P., Ghannoum, M. A., Delchev, D. A., Azie, N. E., Angulo, D., et al. (2022). Efficacy and Safety of Oral Ibrexafungerp for the Treatment of Acute Vulvovaginal Candidiasis: A Global Phase 3, Randomised, Placebo-Controlled Superiority Study (VANISH 306). BJOG 129, 412–420. doi: 10.1111/1471-0528.16972
Spec, A., Pullman, J., Thompson, G. R., Powderly, W. G., Tobin, E. H., Vazquez, J., et al. (2019). MSG-10: A Phase 2 Study of Oral Ibrexafungerp (SCY-078) Following Initial Echinocandin Therapy in Non-Neutropenic Patients With Invasive Candidiasis. J. Antimicrob. Chemother. 74, 3056–3062. doi: 10.1093/jac/dkz277
Tortorano, A. M., Prigitano, A., Morroni, G., Brescini, L., Barchiesi, F. (2021). Candidemia: Evolution of Drug Resistance and Novel Therapeutic Approaches. Infect. Drug Resist. 14, 5543–5553. doi: 10.2147/IDR.S274872
Trouvé, C., Blot, S., Hayette, M. P., Jonckheere, S., Patteet, S., Rodriguez-Villalobos, H., et al. (2017). Epidemiology and Reporting of Candidaemia in Belgium: A Multi-Centre Study. Eur. J. Clin. Microbiol. Infect. Dis. 36, 649–655. doi: 10.1007/s10096-016-2841-3
Wiederhold, N. P. (2021). Antifungal Susceptibility Testing: A Primer for Clinicians. Open Forum Infect. Dis. 8, ofab444. doi: 10.1093/ofid/ofab444
Wiederhold, N. P., Najvar, L. K., Jaramillo, R., Olivo, M., Pizzini, J., Catano, G., et al. (2018). Oral Glucan Synthase Inhibitor SCY-078 Is Effective in an Experimental Murine Model of Invasive Candidiasis Caused by WT and Echinocandin-Resistant Candida Glabrata. J. Antimicrob. Chemother. 73, 448–451. doi: 10.1093/jac/dkx422
Wiederhold, N. P., Najvar, L. K., Olivo, M., Morris, K. N., Patterson, H. P., Catano, G., et al. (2021). Ibrexafungerp Demonstrates In Vitro Activity Against Fluconazole-Resistant Candida Auris and In Vivo Efficacy With Delayed Initiation of Therapy in an Experimental Model of Invasive Candidiasis. Antimicrob. Agents Chemother. 65, e02694-20. doi: 10.1128/AAC.02694-20
Wring, S., Murphy, G., Atiee, G., Corr, C., Hyman, M., Willett, M., et al. (2018). Lack of Impact by SCY-078, a First-in-Class Oral Fungicidal Glucan Synthase Inhibitor, on the Pharmacokinetics of Rosiglitazone, A Substrate for CYP450 2C8, Supports the Low Risk for Clinically Relevant Metabolic Drug-Drug Interactions. J. Clin. Pharmacol. 58, 1305–1313. doi: 10.1002/jcph.1146
Keywords: antifungal testing, antifungal resistance, Candida, ibrexafungerp, SCY-078, EUCAST, caspofungin, micafungin
Citation: Quindós G, Miranda-Cadena K, San-Millán R, Borroto-Esoda K, Cantón E, Linares-Sicilia MJ, Hamprecht A, Montesinos I, Tortorano AM, Prigitano A, Vidal-García M, Marcos-Arias C, Guridi A, Sanchez-Reus F, Machuca-Bárcena J, Rodríguez-Iglesias MA, Martín-Mazuelos E, Castro-Méndez C, López-Soria L, Ruiz-Gaitán A, Fernandez-Rivero M, Lorenzo D, Capilla J, Rezusta A, Pemán J, Guarro J, Pereira J, Pais C, Romeo O, Ezpeleta G, Jauregizar N, Angulo D and Eraso E (2022) In Vitro Antifungal Activity of Ibrexafungerp (SCY-078) Against Contemporary Blood Isolates From Medically Relevant Species of Candida: A European Study. Front. Cell. Infect. Microbiol. 12:906563. doi: 10.3389/fcimb.2022.906563
Received: 28 March 2022; Accepted: 19 April 2022;
Published: 16 May 2022.
Edited by:
Jose L. Lopez-Ribot, University of Texas at San Antonio, United StatesReviewed by:
Hamid Badali, The University of Texas Health Science Center at San Antonio, United StatesMaría Guembe, Gregorio Marañón Hospital, Spain
Copyright © 2022 Quindós, Miranda-Cadena, San-Millán, Borroto-Esoda, Cantón, Linares-Sicilia, Hamprecht, Montesinos, Tortorano, Prigitano, Vidal-García, Marcos-Arias, Guridi, Sanchez-Reus, Machuca-Bárcena, Rodríguez-Iglesias, Martín-Mazuelos, Castro-Méndez, López-Soria, Ruiz-Gaitán, Fernandez-Rivero, Lorenzo, Capilla, Rezusta, Pemán, Guarro, Pereira, Pais, Romeo, Ezpeleta, Jauregizar, Angulo and Eraso. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guillermo Quindós, R3VpbGxlcm1vLlF1aW5kb3NAZWh1LmV1cw==
†Deceased
‡Present addresses: Isabel Montesinos, Laboratoire/Biologie Clinique, CHU UCL Namur, Université catholique de Louvain, Godinne, Belgium
Matxalen Vidal-García, Servicio de Microbiología Clínica y Control de Infección, Hospital Universitario Basurto & BioCruces Bizkaia, Bilbao, Spain
§These authors have contributed equally to this work and share first authorship