 Hanyu Qin
Hanyu Qin Bingbing Xiao
Bingbing Xiao
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Infect. Microbiol. , 25 March 2022
Sec. Biofilms
Volume 12 - 2022 | https://doi.org/10.3389/fcimb.2022.858155
This article is part of the Research Topic Vaginal Dysbiosis and Biofilms View all 10 articles
Bacterial vaginosis (BV) is the most common infectious disease of the reproductive tract in women of childbearing age. It often manifests as an imbalance in the vaginal microbiome, including a decrease in Lactobacillus and an increase in anaerobic bacteria. While Gardnerella spp. are considered a major cause of BV, they are also detected in the vaginal microbiome of healthy women. G. vaginalis was the only recognized species of Gardnerella until a recent study characterized three new species, G. leopoldii, G. piotii, and G. swidsinskii. This review describes the different types and genetic diversity of Gardnerella, as well as new findings on the correlation between different Gardnerella spp. and BV.
Bacterial vaginosis (BV) is the most common lower genital tract infection affecting approximately 30% of women in the general population and 50% of African American women (Ravel et al., 2011; Kenyon et al., 2013; Morrill et al., 2020). Variations in prevalence exist among different races and ethnicities (Allsworth and Peipert, 2007). BV is associated with a disruption of the optimal vaginal microbiota characterized by a decreased proportion of lactic acid-producing bacteria and an increased proportion of a wide array of strict and facultative anaerobes (Ravel et al., 2021). Bacteria commonly associated with BV include Gardnerella vaginalis (G. vaginalis), Megasphaera spp., Fannyhessea vaginae [previously known as Atopobium vaginae, (Rodriguez Jovita et al., 1999; Nouioui et al., 2018)], Dialister spp., Mobiluncus spp., Sneathia amnii, Sneathia sanguinegens, Porphyromonas spp., and Prevotella spp (Muzny et al., 2018; Rosca et al., 2020). Although BV is frequently asymptomatic, women with BV are more likely to report vaginal odor, itching, and discharge than those without (Klebanoff et al., 2004). In addition, most women do not report BV symptoms to their providers, even when there are clinical signs (Masson et al., 2019). Serious adverse health outcomes have been associated with BV, including increased risk of infertility (Ravel et al., 2021), adverse pregnancy outcomes (Giakoumelou et al., 2016; Tabatabaei et al., 2019), pelvic inflammatory disease (PID) (Wiesenfeld et al., 2002; Ravel et al., 2021) and sexually transmitted infections (STIs), including chlamydia (Shipitsyna et al., 2020), gonorrhea (Bautista et al., 2017), human papilloma virus (HPV) (Usyk et al., 2022) and human immunodeficiency virus (HIV) (Atashili et al., 2008).
BV etiology is controversial and not yet fully understood. According to Catlin’research, the G. vaginalis-associated vaginal syndrome was earlier called nonspecific vaginitis (NSV) in recognition of the absence of recognized agents of vaginitis (Catlin, 1992). In 1955, Gardner and Dukes showed that 90% of NSV cases were caused by a single microbe, Haemophilus vaginalis (H. vaginalis), and the name was changed to H. vaginalis vaginitis (Gardner and Dukes, 1955). However, subsequent studies found that because H. vaginalis does not require heme or nicotinamide adenine dinucleotide for growth, it may not be a member of the Haemophilus genus. H. vaginalis shows uncertainty in gram staining, also a characteristic of the Corynebacteria. As a result, H. vaginalis was reassigned to the Corynebacterium genus and renamed Corynebacterium vaginale (Zinnemann and Turner, 1963). This name also proved to be inaccurate because these bacteria are catalase-negative and do not have arabinose in their cell wall (Catlin, 1992). In 1980, two large taxonomic studies using DNA hybridization, electron microscopy, and various biochemical methods showed that the bacterium lacked close similarity to any previously established genus (Greenwood and Pickett, 1980; Piot et al., 1980). This resulted in the development of a new genus, Gardnerella, and Corynebacterium vaginitis was renamed Gardnerella vaginitis. Gardner believed that Gardnerella vaginitis was a specific vaginal infection with a clear cause and that most vaginitis previously classified as “nonspecific” was likely caused by Gardnerella spp. In 1984, the name was officially changed to “BV” because vaginitis is suggestive of an inflammatory response in the vaginal epithelium, which is usually absent in women with BV (Catlin, 1992). It wasn’t until 1984 that BV was officially named (Workshop on Anaerobic Curved Rods and Bacterial Vaginosis, Stockholm, January, 1984) (Bump et al., 1984).
Since Gardnerella spp. is highly detected in BV, it appears to have a special role in vaginal microbiota dysbiosis (Reid, 2018). While Gardnerella spp. is found in 95% to 100% of BV cases (Muzny et al., 2019), colonization does not always lead to BV (Hickey and Forney, 2014; Machado et al., 2015). Indeed, the role of Gardnerella spp. in BV has remained controversial because it is present in both healthy vaginal microbiota and in BV (Zozaya-Hinchliffe et al., 2010). As a result, researchers have speculated about whether there are different Gardnerella spp. (Cornejo et al., 2018) with distinct pathogenicities that can lead to different clinical outcomes (Janulaitiene et al., 2018). It is possible that while healthy women are colonized by a less virulent strain of Gardnerella spp., other more virulent strains promote the development of BV. Studies have used several Gardnerella spp. typing tests to explore the clinical characteristics of different species. Indeed, the taxonomy of Gardnerella spp. will need to be more completely defined in order to fully understand the mechanism of Gardnerella spp. in BV pathogenesis (Castro et al., 2019). This study reviews recent literature to explore the characteristics of Gardnerella spp., the typing methods used, and their clinical significance.
G. vaginalis was the first recognized Gardnerella species (Harwich et al., 2010), with its closest relatives in the Bifidobacterium genus (Harwich et al., 2010; Castro et al., 2019). The cells are small, nonmotile, nonencapsulated, non-spore-forming, pleomorphic rods with average dimensions of 0.4×1.0~1.5µm (Onderdonk et al., 2016). The cells are small, nonmotile, nonencapsulated, non-spore-forming, pleomorphic rods with average dimensions of 0.4×1.0~1.5µm (Sadhu et al., 1989). Known as a facultative anaerobic microorganism, G. vaginalis was also described as fastidious, as it grew better at 37°C in complex media in an atmosphere with 5–10% of carbon dioxide (CO2) or in a candleflame extinction jar (Catlin, 1992; Cereija et al., 2013). Nevertheless, it was demonstrated that certain G. vaginalis strains are strict anaerobes (Malone et al., 1975). The cellular surface of G. vaginalis is covered with fimbriae, which are responsible for the attachment of G. vaginalis to vaginal epithelial cells (Scott et al., 1989; Onderdonk et al., 2016). More recent biochemical testing has shown that G. vaginalis is catalase-, oxidase-, and b-glucosidase-negative (Catlin, 1992; Turovskiy et al., 2011). It can ferment starch, dextrin, sucrose, glucose, fructose, ribose, maltose and raffinose. Some strains can also ferment xylose and trehalose. Conversely, G. vaginalis is unable to ferment rhamnose, melibiose, mannitol and sorbitol (Harwich et al., 2010).
The important pathogenic characteristics of Gardnerella spp. include the production of sialidase, an enzyme that degrades cervical and vaginal mucus, and vaginolysin, a lysoid that induces the lysis of vaginal epithelial cells (Castro et al., 2019). Sialidase is associated with the degradation of key mucosal protective factors, such as mucin, that lead to the shedding of vaginal epithelial cells (Lewis et al., 2013; Hardy et al., 2017). Gardnerella spp. has three genes that encode sialidase, NanH1 (sialidase A gene), NanH2, and NanH3. NanH2 and NanH3 appear to be the main sources of sialidase activity in Gardnerella spp (Robinson et al., 2019). Vaginal hemolysin is a cholesterol-dependent cell hemolysin that was first discovered in the culture medium of Gardnerella spp. It has cytotoxic activity against human erythrocytes and can induce the dissolution of human erythrocytes, epithelial cells, and polymorphonuclear leukocytes (Zilnyte et al., 2015). Vaginal hemolysin and sialidase are the most widely studied virulence factors of Gardnerella spp. (Santiago et al., 2011; Janulaitiene et al., 2018), and are assumed to play a substantial role in the pathogenesis of BV (Pleckaityte et al., 2012). Swidsinski et al. used fluorescence in situ hybridization (FISH) specific for Gardnerella spp. and were the first to show that these species form biofilms on vaginal epithelium in women with BV. This finding revealed the nature of clue cells, epithelial cells covered by a Gardnerella spp. biofilm, and their etiological role in Gardnerella spp. infection (Swidsinski et al., 2005).
Gardnerella spp. was first divided into eight biotypes based on its metabolic characteristics (Piot et al., 1984). In 1894, Piot et al. established a simple and repeatable method for Gardnerella spp. biotyping, known as Pilot typing, that is based on the reaction of lipase, hippurate hydrolysis, and β-galactosidase. Benito et al. identified 17 Gardnerella spp. biotypes based on these characteristics in addition to fermentation of xylose, arabinose, and galactose (Benito et al., 1986). This method is based on enzyme reactions, however, so there is a risk that the wrong biotypes will be produced when conditions change. Thus, biotyping is not recommended to understand the epidemic etiology of Gardnerella spp. (Ingianni et al., 1997).
Four genotypes of Gardnerella spp. were identified by amplified ribosomal DNA restriction analysis (ARDRA) using different restriction enzymes (Ingianni et al., 1997). In 1997, Ingianni et al. first used ARDRA to identify 34 Gardnerella spp. strains. This method is more accurate than biotyping but still relies on the isolation and purification of Gardnerella spp. Each subtype has a distinct growth and metabolic pattern, however, and preferentially isolated strains may misrepresent the clinical distribution of Gardnerella subtypes.
Quantitative polymerase chain reaction (qPCR) typing can be used on uncultured clinical specimens, allowing for quantitative assessment of bacterial load and qualitative identification of Gardnerella subtypes (Balashov et al., 2014). Since the isolation of living bacterial cells is not involved, this approach can be performed on different types of samples, including frozen DNA or vaginal swab samples collected and stored under sub-optimal conditions. In 2012, Ahmed et al. conducted a comparative genomic analysis of 17 Gardnerella spp. clinical isolates and suggested that it would be more accurate to define Gardnerella spp. as four separate species (Ahmed et al., 2012). Balashov and his colleagues developed multiplex single-tube qPCR targeting genes encoding putative a-L-fucosidase (clade 1), a hypothetical protein (clade 2), thioredoxin (clade 3) and CIC family chloride transporter (clade 4), which was proven to be clade-specific and capable of strain typing and identification of the four G. vaginalis clades in noncultured clinical vaginal specimens (Balashov et al., 2014).
Chaperonin-60 is a molecular chaperone required for the folding and assembly of proteins and protein complexes in all eubacteria and in the plastids and mitochondria of eukaryotes (Hill et al., 2005). A 549-567 bp fragment of the cpn-60 gene was amplified by universal PCR primers, and the sequencing results were relatively stable (Jayaprakash et al., 2012). Gardnerella spp. comprises four subgroups A, B, C, and D, based on cpn60 barcode sequences and whole-genome sequences (Jayaprakash et al., 2012; Schellenberg et al., 2016). Cpn60 sequencing was also applied to vaginal microbes in a larger sample of African commercial sex workers (Schellenberg et al., 2011). Profiling of vaginal microbiomes using cpn60 barcode sequencing, and application of clade-specific PCR showed that the vagina is often colonized by multiple Gardnerella spp. subgroups simultaneously (Khan et al., 2019).
Whole genome sequencing refers to the analysis of the whole genome sequence of Gardnerella using digital DNA-DNA hybridization and average nucleotide identity. In 2019, Vaneechoutte et al. analyzed the whole genome sequence of 81 Gardnerella spp. and confirmed the existence of 13 subgroups, GSP01-GSP13, all of which are sufficiently different to be classified as independent species. Two subgroups were found in subtypes clade 1, clade 2, and clade 4, and three undefined subgroups were found in subtype clade 3 (Vaneechoutte et al., 2019). This was an important breakthrough in this field that resulted in an amended description of G. vaginalis and the characterization of three additional Gardnerella species, G. leopoldii, G. piotii, and G. swidsinskii (Potter et al., 2019).
The commonly used Gardnerella typing methods include ARDRA, cpn-60, qPCR, and whole genome sequencing (Castro et al., 2019). Since ARDRA typing of Gardnerella results does not specifically correspond to clinical status (Piot et al., 1984; Benito et al., 1986), it is not detailed here. Cpn-60 gene sequencing was used to divide Gardnerella into four subtypes A, B, C and D, based on the phylogenetic relationship between each cpn60 gene subsequence (Jayaprakash et al., 2012). Similarly, qPCR divided Gardnerella isolates into four clades 1, 2, 3 and 4 (Balashov et al., 2014). Schellenberg et al. compared cpn60 UT sequences from 112 G. vaginalis isolates from three continents with cpn60 UT sequences extracted from previously published whole genome sequences. All new isolates and previously published genomes fall into four cpn60 subgroups (Jayaprakash et al., 2012), including 17 isolates belonging to clades 1- 4 (Ahmed et al., 2012). These results indicate that similar phylogenetic resolution can be achieved using a partial single gene sequence (552 bp) as 473 full-length gene sequences common to all 17 genomes. Cpn60 subgroups A, B, C, and D correspond to clade 4, 2, 1, and 3, respectively (Schellenberg et al., 2016). Using a retrospective comparative analysis of 103 publicly available genomes and meta-transcriptomic bacterial vaginosis datasets, Potter et al. identified 9 genome species of Gardnerella, GS01-09 (Potter et al., 2019). Gardnerella species, labeled with names or numbers, were defined by analysis of 81 genome sequences by digital DNA-DNA hybridization, average nucleotide identity (ANI), and MALDI-MS protein signatures (Vaneechoutte et al., 2019). The specific corresponding relationship is shown in Table 1.
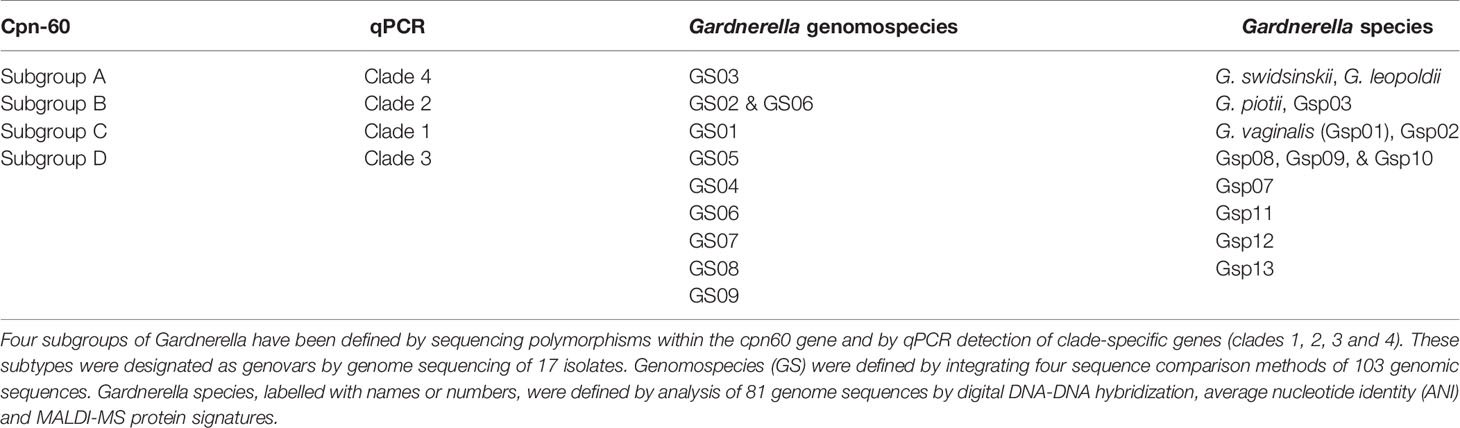
Table 1 The relationship between different Gardnerella species.
Gardnerella is detected in both the vaginal secretions of women with BV and healthy women. Using two qPCR assays, Balashov et al. analyzed G. vaginalis bacterial loads and clade distribution in 100 clinical vaginal-swab samples and showed that the prevalence of G. vaginalis was 100% of BV patients and 97% in healthy women; however, the G. vaginalis concentration was significantly lower in non-BV samples. The detection frequency of clades 1, 2, 3 and 4 was 53%, 25%, 32% and 83%, respectively. In addition, 70% of BV vaginal swab samples had multiple subtypes of Gardnerella (Balashov et al., 2014). An assessment of the vaginal microbiota of 413 non-pregnant, reproductive-age Canadian women showed that the number of Gardnerella spp. detected per Gardnerella-positive sample ranged from 1 to 10. Multiple Gardnerella spp. were detected in 63.8% of samples, consistent with a previous report of multiple Gardnerella clades in 70% of samples from women (Balashov et al., 2014), and the majority contained one or two species (Hill et al., 2019). Multiple subtypes of Gardnerella had a significant positive correlation with BV, suggesting that the co-occurrence of multiple subtypes may be one of the risk factors for BV.
There was no correlation between Gardnerella spp. biotyping and clinical symptoms (Pleckaityte et al., 2012). According to Hill and Albert’s research, the relative abundances of the more frequently occurring species (G. vaginalis, G. swidsinskii, G. leopoldii, G. piotii, and genome species) among groups based on clinical Nugent scores (negative, intermediate, and BV) and self-reported symptoms in the 2 weeks prior to the swab collection (odor, irritation, and discharge) were compared (Hill and Albert, 2019). Whole genome sequence analysis revealed a strong relationship between abnormal odor and discharge with higher relative abundance of G. vaginalis and G. swidsinskii. The cooccurrence of these Gardnerella spp. also showed proportionality, suggesting that their abundance is correlated. Thus, it remains unclear whether one of these species or both are associated with vaginal symptoms (Hill and Albert, 2019).
Early studies on Gardnerella typing indicated that biotypes 1, 2, and 5 were more common in females with BV (Piot et al., 1984), however subsequent studies could not confirm these findings. Balashov et al. found a positive correlation between BV and clade 1 and clade 3 in vaginal samples from 60 American women. Meanwhile, clade 2 was positively correlated with vaginal microbiota in an intermediate state of BV and C4 had no correlation with infection (Balashov et al., 2014). Janulaitiene et al. performed qPCR on vaginal swab samples from 109 Lithuanian women and confirmed that the microbial status of the vaginal microbiota was associated with the clade 1 and clade 2 subgroups (Nugent score 7-10). The clade 3 and clade 4 showed no association with high Nugent Scores (Janulaitiene et al., 2017). However, the results from a study on the vaginal microbiota of 299 Russian women of reproductive age were different from those of previous studies. Quantifying the four Gardnerella subtypes could more accurately distinguish BV from healthy microbiota than detecting the sialidase A gene and clade 4 was closely related to the status of the BV microbiome (Shipitsyna et al., 2019).
Plummer et al. studied the relationship between infection with the clade 1, clade 2, and clade 3 subtypes and Nugent scores in 101 Australian women of reproductive age. Multiple Gardnerella subtypes and the clade 1 subtype alone were associated with the absence of Lactobacillus in the vaginal microbiome. Clade 4 was not associated with BV or the absence of Lactobacillus, supporting the existence of symbiotic and pathogenic subtypes of Gardnerella spp (Plummer et al., 2020). Hill et al. used whole genome sequencing to compare the species abundance of Gardnerella subgroups between healthy women and those with BV. The relative abundance of G. vaginalis, G. swidsinskii, and G. piotii correlated significantly with Nugent scores. In addition, the association between G. piotii (B subgroup/clade 2) and the “intermediate state” microbiome was observed by cpn60 sequencing and qPCR (Hill and Albert, 2019). Gardnerella typing results from different studies are shown in Table 2.
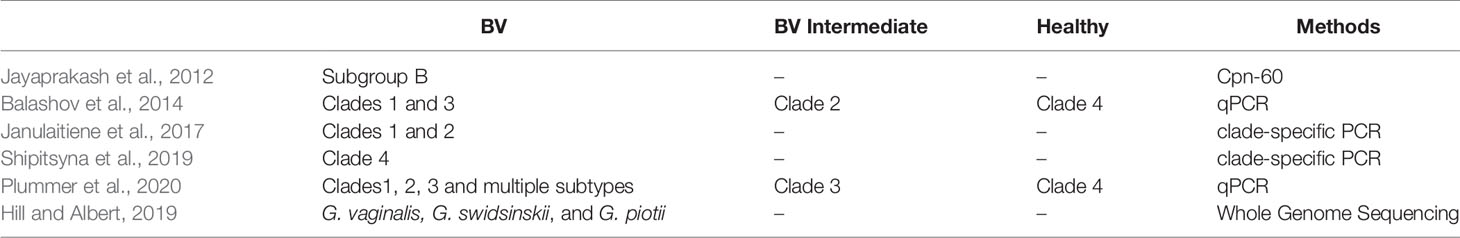
Table 2 Different Gardnerella subtypes predict distinct BV clinical outcomes.
In summary, current studies remain unable to determine the relationship between different Gardnerella genotypes and clinically relevant BV status.
Metronidazole resistance by Gardnerella isolates is likely responsible for refractory or recurrent BV. Women with BV are typically infected with multiple Gardnerella spp. subtypes, so metronidazole may eliminate sensitive Gardnerella subtypes but allow drug-resistant subtypes to survive. This could explain the presence of Gardnerella in the vaginal microbiota even after metronidazole treatment. BV is characterized by a thick vaginal multi-species biofilm, in which Gardnerella spp. is the predominant species. Since standard antibiotics, like metronidazole, are unable to fully eradicate the vaginal biofilm, this may explain the high recurrence rates of BV (Machado et al., 2015; Verstraelen and Swidsinski, 2019). Early identification of metronidazole-resistant Gardnerella subtypes helps to predict the prognosis of BV and inform an appropriate treatment plan. To explore the sensitivity of different Gardnerella subtypes to metronidazole, Schuyler et al. collected 88 Gardnerella strains and divided them into four groups, clades 1, 2, 3 and 4, by qPCR sequencing. Metronidazole resistance was defined as a minimum inhibitory concentration ≥32 μg/mL. A high correlation was found between metronidazole resistance and Gardnerella typing. While clade 3 and clade 4 strains showed 100% resistance, while clade 1 and clade 2 showed 35% and 7.1% resistance, respectively (Schuyler et al., 2016).
In 2017, Hilbert et al. conducted a short-term longitudinal study of 149 non-pregnant Canadian women of reproductive age. Vaginal swab samples were collected at first diagnosis, 7 days after treatment, and 40-45 days after follow-up to detect the abundance of different Gardnerella subtypes by qPCR. Higher prevalence of clade 1 and clade 4 were found in vaginal samples than clade 2 and clade 3. The abundance of each subtype increased as the Nugent score, or the severity of BV, worsened (Hilbert et al., 2017). The abundance of clade 1 and clade 4 decreased after clinical treatment regardless of the clinical and microbiological outcome, and clade 2 decreased in women who continued treatment for 40-45 days. Recurrent BV is characterized by increased clade 1 and clade 2 levels after treatment. The clade 1 and clade 4 subgroups were the dominant strains in vaginal specimens. While the clade abundance of Gardnerella was generally higher in vaginal samples that met the four Amsel criteria than those that did not, clade 1 was an exception. Thus, differences in qPCR subtype abundance were associated with Nugent score, Amsel criteria, sensitivity to treatment, and BV recurrence.
A recent study by Turner et al. associated sustained high abundance of the Gardnerella Gsp07 subtype with a refractory BV response and sustained low abundance of the Gardnerella Gsp07 subtype and G. Swidsinskii/G. Leopoldii with BV remission. In most patients with BV relapse or remission, the abundance of Lactobacillus species increased 4–14 days after initiation of treatment, and the increase was more obvious and sustained in patients with BV remission. These findings confirmed that Gardner’s Gsp07 subtypes and G. swidsinskii/G. leopoldii coinfection correlate with poor clinical outcomes. Alternatively, direct or indirect inhibition of lactic acid bacteria strains may interfere with clinical recovery. Treatment by clinicians targeting these marker subtypes of adverse outcomes may improve clinical outcomes in patients with BV (Turner et al., 2021).
Gardnerella pathogenicity is primarily mediated through vaginal hemolysin (VLY), sialidase and biofilm formation (Pleckaityte et al., 2012). Previous studies have shown that pathogenicity differs by Gardnerella subspecies (Janulaitiene et al., 2018). Zilnyte et al. found that VLY activity is dependent on the complement regulatory molecule, CD59, and showed that higher CD59 expression in hamsters correlated with increased vaginal hemolysin-soluble cell sensitivity (Zilnyte et al., 2015). In the cell culture model, the expression level of vaginal hemolysin was correlated with the level of cytotoxicity, but there was no any correlation between VLY production level and G. vaginalis genotype/biotype (Pleckaityte et al., 2012).
Sialidase lyses the terminal sialic acid residues of sialoglycan in the vaginal environment and plays a key role in providing nutrition for Gardnerella spp. through sialic acid catabolism, providing a site for bacteria adhesion to the epithelium, facilitating biofilm formation, and modulating immune responses (Lewis et al., 2013; Schellenberg et al., 2017). Harwich et al. (2010) and Janulaitiene et al. (2018) found significant differences in the sialidase activity of Gardnerella clades, however, with clade 2 having the highest levels followed by clade 1, and clade 4 having the lowest. In addition, the gene coding for sialidase was detected in all isolates of clade 1 and clade 2, but not in clade 4 isolates (Schellenberg et al., 2016; Janulaitiene et al., 2017). Shipitsyna also holds that clade 4 strains mostly lack the sialidase A gene (Shipitsyna et al., 2019). Sialidase activity is considered a marker of BV. Indeed, more than 50% of BV is asymptomatic, which may be caused by Gardnerella subspecies that lack sialidase (Janulaitiene et al., 2017). Sialidase acts on sugar chains with sialic acid residues, which are abundant on the mucosal surface of the reproductive tract. Sialidase activity can be used as a diagnostic marker of BV (Janulaitiene et al., 2018) and rapid clinical detection using products like BVBlue® (Sekisui Diagnostics, L.L.C., Birmingham, AL, USA) (Hill and Albert, 2019).
The exfoliated vaginal epithelial cells in BV patients are covered with multi-bacterial biofilms dominated by Gardnerella (Vestby et al., 2020). Biofilm formation is not only associated with increased antimicrobial resistance and disease recurrence but also increased risk of sexual transmission. However, there is no significant difference in biofilm quantity between Gardnerella subtypes (Janulaitiene et al., 2018).
In summary the relationship between the different subtypes of Gardnerella and bacterial vaginosis is unclear. A large number of studies show that different Gardnerella subtypes are possibly represent different drug resistance, virulence, bacterial load and indicate the clinical outcomes of BV. And the clinical significance of asymptomatic BV remains unclear, one possible explanation for its occurrence is the presence of high numbers of nonpathogenic Gardnerella or other morphologically similar species. This is especially likely given that Gardnerella is one of the key predictors of the Nugent score. Furthermore, metronidazole treatment for BV cure rate is not ideal, and the proportion of refractory and recurrent BV continues to rise. This study reviewed the relationship between Gardnerella subtypes species and BV clinical outcomes and evaluated patient prognosis according to Gardnerella typing. This is particularly important so that appropriate treatment can be given to improve the BV clinical cure rate and reduce adverse obstetric and gynecological complications as well as disease recurrence. Given the current diversity of Gardnerella phenotypes, especially virulence factors, genotypic diversity, and Gardnerella prevalence in women, understanding the clinical significance of these different strains is critical.
HQ and BX discussed the contents, wrote, reviewed, and edited the manuscript. All authors contributed to the article and approved the submitted version.
This present work was funded by the grants of the National Key Research and Development Program of China (2021YFC2301000) and the National Natural Science Foundation of China (81971342).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Ahmed, A., Earl, J., Retchless, A., Hillier, S. L., Rabe, L. K. (2012). Comparative Genomic Analyses of 17 Clinical Isolates of Gardnerella Vaginalis Provide Evidence of Multiple Genetically Isolated Clades Consistent With Subspeciation Into Genovars. J. Bacteriol. 194, 3922–3937. doi: 10.1128/JB.00056-12
Allsworth, J. E., Peipert, J. F. (2007). Prevalence of Bacterial Vaginosis: 2001-2004 National Health and Nutrition Examination Survey Data. Obstet Gynecol. 109 (1), 114–120. doi: 10.1097/01.Aog.0000247627.84791.91
Atashili, J., Poole, C., Ndumbe, P. M., Adimora, A. A., Smith, J. S. (2008). Bacterial Vaginosis and HIV Acquisition: A Meta-Analysis of Published Studies. Aids 22 (12), 1493–1501. doi: 10.1097/QAD.0b013e3283021a37
Balashov, S. V., Mordechai, E., Adelson, M. E., Gygax, E. G. (2014). Identification, Quantification and Subtyping of Gardnerella Vaginalis in Noncultured Clinical Vaginal Samples by Quantitative PCR. J. Med. Microbiol. 63 162–175. doi: 10.1099/jmm.0.066407-0
Bautista, C. T., Wurapa, E. K., Sateren, W. B., Morris, S. M., Hollingsworth, B. P., Sanchez, J. L. (2017). Association of Bacterial Vaginosis With Chlamydia and Gonorrhea Among Women in the U.S. Army. Am. J. Prev. Med. 52 (5), 632–639. doi: 10.1016/j.amepre.2016.09.016
Benito, R., Vazquez, J. A., Berron, S., Fenoll, A., Saez-Nieto, J. A. (1986). A Modified Scheme for Biotyping Gardnerella Vaginalis. J. Med. Microbiol. 21, 357–359. doi: 10.1099/00222615-21-4-357
Bump, R. C., Zuspan, F. P., Buesching, W. J., Ayers, L. W., Stephens, T. J. (1984). The Prevalence, Six-Month Persistence, and Predictive Values of Laboratory Indicators of Bacterial Vaginosis (Nonspecific Vaginitis) in Asymptomatic Women. Am. J. Obstet Gynecol. 150, 917–924. doi: 10.1016/0002-9378(84)90381-8
Castro, J., Jefferson, K. K., Cerca, N. (2019). Genetic Heterogeneity and Taxonomic Diversity Among Gardnerella Species. Trends Microbiol 28 (3), 202–211. doi: 10.1016/j.tim.2019.10.002
Catlin, B. W. (1992). Gardnerella Vaginalis: Characteristics, Clinical Considerations, and Controversies. Cinical Microbiol. Rev. 5 (3), 213–237. doi: 10.1128/CMR.5.3.213
Cereija, T. B., Castro, J., Alves, P., Cerca, N. (2013). Influence of Anaerobic Conditions on Vaginal Microbiota Recovery From Bacterial Vaginosis Patients. Sexually Transmitted Infections 89 (4), 307–307. doi: 10.1136/sextrans-2013-051100
Cornejo, O. E., Hickey, R. J., Suzuki, H., Forney, L. J. (2018). Focusing the Diversity of Gardnerella Vaginalis Through the Lens of Eco Types. Evolutionary Appl. 11, 312–324. doi: 10.1111/eva.12555
Gardner, H., Duke, C. (1955). Haemophilus Vaginalis Vaginitis: A Newly Defined Specific Infection Previously Classified “Nonspecific” Vaginitis. Am. J. Obstetrics Gynecol. 69, 962–976. doi: 10.1016/0002-9378(55)90095-8
Giakoumelou, S., Wheelhouse, N., Cuschieri, K., Entrican, G., Howie, S. E., Horne, A. W. (2016). The Role of Infection in Miscarriage. Hum. Reprod. Update 22 (1), 116–133. doi: 10.1093/humupd/dmv041
Greenwood, J. R., Pickett, M. J. (1980). Transfer of Haemophilus Vaginalis Gardner and Dukes to a New Genus, Gardnerella: G. Vaginalis (Gardner and Dukes) Comb. Nov. Int. J. Systematic Bacteriol. 30, 170–178. doi: 10.1099/00207713-30-1-170
Hardy, L., Jespers, V., Van den Bulck, M., Buyze, J., Mwambarangwe, L., Musengamana, V., et al. (2017). The Presence of the Putative Gardnerella Vaginalis Sialidase A Gene in Vaginal Specimens is Associated With Bacterial Vaginosis Biofilm. PLoS One 12 (2), e0172522. doi: 10.1371/journal.pone.0172522
Harwich, M. D., Alves, J. M., Buck, G. A., Strauss, J. F., Patterson, J. L., Oki, A. T., et al. (2010). Drawing the Line Between Commensal and Pathogenic Gardnerella Vaginalis Through Genome Analysis and Virulence Studies. BMC Genomics 11 (375), 3–12. doi: 10.1186/1471-2164-11-375
Hickey, R. J., Forney, L. J. (2014). Gardnerella Vaginalis Does Not Always Cause Bacterial Vaginosis. J. Infect. Dis. 210 (10), 1682–1683. doi: 10.1093/infdis/jiu303
Hilbert, D. W., Schuyler, J. A., Gygax, S. E., Adelson, M. E., Mordechai, E., Sobel, J. D. (2017). Gardnerella Vaginalis Population Dynamics in Bacterial Vaginosis. Eur. J. Clin. Microbiol. Infect. Dis. 36, 1269–1278. doi: 10.1007/s10096-017-2933-8
Hill, J. E., Albert, A. Y. K. (2019). Resolution and Cooccurrence Patterns of Gardnerella Leopoldii, G. Swidsinskii, G. Piotii and G. Vaginalis Within the Vaginal Microbiome. Infection Immun. 87 (12), e00532–00519. doi: 10.1128/IAI.00532-19
Hill, J. E., Goh, S. H., Money, D. M., Doyle, M., Li, A., Crosby, W. (2005). L,… Hemmingsen, S. M). Characterization of Vaginal Microflora of Healthy Nonpregnant Women by Sequencebased Methods. Am. J. Obstetrics Gynecol. 193, 682–692. doi: 10.1016/j.ajog.2005.02.094
Ingianni, A., Petruzzelli, S., Morandotti, G., Pompei, R. (1997). Genotypic Differentiation of Gardnerella Vaginalis by Amplified Ribosomal DNA Restriction Analysis (ARDRA). Immunol. Med. Microbiol. 18, 61–66. doi: 10.1111/j.1574-695X.1997.tb01028.x
Janulaitiene, M., Gegzna, V., Baranauskiene, L., Bulavaitė, A., Simanavicius, M., Pleckaityte, M. (2018). Phenotypic Characterization of Gardnerella Vaginalis Subgroups Suggests Differences in Their Virulence Potential. PLoS One 13 (7), e0200625. doi: 10.1371/journal.pone.0200625
Janulaitiene, M., Paliulyte, V., Grinceviciene, S., Zakareviciene, J., Vladisauskiene, A., Marcinkute, A., et al. (2017). Prevalence and Distribution of Gardnerella Vaginalis Subgroups in Women With and Without Bacterial Vaginosis. BMC Infect. Dis. 17 (394), 1–9. doi: 10.1186/s12879-017-2501-y
Jayaprakash, T. P., Schellenberg, J. J., Hill, J. E. (2012). Resolution and Characterization of Distinct Cpn60-Based Subgroups of Gardnerella Vaginalis in the Vaginal Microbiota. PLoS One 7 (8), e43009. doi: 10.1371/journal.pone.0043009
Kenyon, C., Colebunders, R., Crucitti, T. (2013). The Global Epidemiology of Bacterial Vaginosis: A Systematic Review. Am. J. Obstetrics Gynecol. 209 (6), 505–523. doi: 10.1016/j.ajog.2013.05.006
Khan, S., Voordouw, M. J., Hill, J. E. (2019). Competition Among Gardnerella Subgroups From the Human Vaginal Microbiome. Front. Cell. Infection Microbiol. 9. doi: 10.3389/fcimb.2019.00374
Klebanoff, M. A., Schwebke, J. R., Zhang, J., Nansel, T. R., Yu, K. F., Andrews, W. W. (2004). Vulvovaginal Symptoms in Women With Bacterial Vaginosis. Obstet Gynecol. 104 (2), 267–272. doi: 10.1097/01.AOG.0000134783.98382.b0
Lewis, W. G., Robinson, L. S., Gilbert, N. M., Perry, J. C., Lewis, A. L. (2013). Degradation, Foraging, and Depletion of Mucus Sialoglycans by the Vagina-Adapted Actinobacterium Gardnerella Vaginalis. J. Biol. Chem. 288, 12067–12079. doi: 10.1074/jbc.M113.453654
Machado, D., Castro, J., Palmeira-de-Oliveira, A., Martinez-de-Oliveira, J., Cerca, N. (2015). Bacterial Vaginosis Biofilms: Challenges to Current Therapies and Emerging Solutions. Front. Microbiol. 61528. doi: 10.3389/fmicb.2015.01528
Malone, B. H., Schreiber, M., Schneider, N. J., Holdeman, L. V. (1975). Obligately Anaerobic Strains of Corynebacterium Vaginale (Haemophilus Vaginalis). J. Clin. Microbiol. 2 (3), 272–275. doi: 10.1128/jcm.2.3.272-275.1975
Masson, L., Barnabas, S., Deese, J., Lennard, K., Dabee, S., Gamieldien, H., et al. (2019). Inflammatory Cytokine Biomarkers of Asymptomatic Sexually Transmitted Infections and Vaginal Dysbiosis: A Multicentre Validation Study. J. Sexually Transmitted Infections 95 (1), 5–12. doi: 10.1136/sextrans-2017-053506
Morrill, S., Gilbert, N. M., Lewis, A. L. (2020). Gardnerella Vaginalis as a Cause of Bacterial Vaginosis- Appraisal of the Evidence From In Vivo Models. Front. Cell. Infection Microbiol. 10. doi: 10.3389/fcimb.2020.00168
Muzny, C. A., Blanchard, E., Taylor, C. M., Aaron, K. J., Talluri, R., Griswold, M. E., et al. (2018). Identification of Key Bacteria Involved in the Induction of Incident Bacterial Vaginosis: A Prospective Study. J. Infect. Dis. 218 (6), 966–978. doi: 10.1093/infdis/jiy243
Muzny, C. A., Taylor, C. M., Swords, W. E., Tamhane, A., Chattopadhyay, D., Cerca, N., et al. (2019). An Updated Conceptual Model on the Pathogenesis of Bacterial Vaginosis. J. Infect. Dis. 220, 1399–1405. doi: 10.1093/infdis/jiz342
Nouioui, I., Carro, L., García-López, M., Meier-Kolthoff, J. P., Woyke, T., Kyrpides, N. C., et al. (2018). Genome-Based Taxonomic Classification of the Phylum Actinobacteria. Front. Microbiol. 9, 2007. doi: 10.3389/fmicb.2018.02007
Onderdonk, A. B., Delaney, M. L., Fichorova, R. N. (2016). The Human Microbiome During Bacterial Vaginosis. Clin. Microbiol. Rev. 29 (2), 223–238. doi: 10.1128/CMR.00075-15
Piot, P., Dyck, E. V., Peeters, M., Hale, J., Totten, P. A., Holmes, K. K. (1984). Biotypes of Gardnerella Vaginalis. J. Clin. Microbiol. 20, 677–679. doi: 10.1128/jcm.20.4.677-679.1984
Piot, P., Van Dyck, E., Goodfellow, M., Falkow, S. (1980). A Taxonomic Study of Gardnerella Vaginalis (Haemophilus Vaginalis) Gardner and Dukes 1955. Microbiology 119 (2), 373–396. doi: 10.1099/00221287-119-2-373
Pleckaityte, M., Janulaitiene, M., Lasickiene, R., Zvirbliene, A. (2012). Genetic and Biochemical Diversity of Gardnerella Vaginalis Strains Isolated From Women With Bacterial Vaginosis. FEMS Immunol. Med. Microbiol. 65, 69–77. doi: 10.1111/j.1574-695X.2012.00940.x
Plummer, E. L., Vodstrcil, L. A., Murray, G. L., Fairley, C. K., Danielewski, J. A., Garland, M. A. (2020). Gardnerella Vaginalis Clade Distribution Is Associated With Behavioural Practices and Nugent Score in Women Who Have Sex With Women. J. Infect. Dis. 221, 454–463. doi: 10.1093/infdis/jiz474
Potter, R. F., Burnham, C. D., Dantas, G. (2019). In Silico Analysis of Gardnerella Genomospecies Detected in the Setting of Bacterial Vaginosis. Clin. Chem. 65 (11), 1375–1387. doi: 10.1373/clinchem.2019.305474
Ravel, J., Gajer, P., Abdo, Z., Schneider, G. M., Koenig, S. S. K., McCulle, S. L., et al. (2011). Vaginal Microbiome of Reproductive-Age Women. J. Proc. Natl. Acad. Sci. 108 (Supplement 1), 4680–4687. doi: 10.1073/pnas.1002611107
Ravel, J., Moreno, I., Simón, C. (2021). Bacterial Vaginosis and its Association With Infertility, Endometritis, and Pelvic Inflammatory Disease. Am. J. Obstet Gynecol. 224 (3), 251–257. doi: 10.1016/j.ajog.2020.10.019
Reid, G. (2018). Is Bacterial Vaginosis a Disease? Appl. Microbiol. Biotechnol. 102 (2), 553–558. doi: 10.1007/s00253-017-8659-9
Robinson, L. S., Schwebke, J. R., Lewis, W. G., Lewis, A. L. (2019). Identification and Characterization of NanH2 and NanH3, Enzymes Responsible for Sialidase Activity in the Vaginal Bacterium Gardnerella Vaginalis. J. Biol. Chem. 294 (14), 5230 –55245. doi: 10.1074/jbc.RA118.006221
Rodriguez Jovita, M., Collins, M. D., Sjödén, B., Falsen, E. (1999). Characterization of a Novel Atopobium Isolate From the Human Vagina: Description of Atopobium Vaginae Sp. Nov. Int. J. Syst. Bacteriol. 49 Pt 4, 1573–1576. doi: 10.1099/00207713-49-4-1573
Rosca, A. S., Castro, J., Sousa, L. G. V., Cerca, N. (2020). Gardnerella and Vaginal Health: The Truth is Out There. FEMS Microbiol. Rev. 44 (1), 73–105. doi: 10.1093/femsre/fuz027
Sadhu, K., Domingue, P. A. G., Chow, A. W., Nelligan, J., Cheng, N., Costerton, J. W. (1989). Gardnerella Vaginalis has a Gram-Positive Cell-Wall Ultrastructure and Lacks Classical Cell-Wall Lipopolysaccharide. J. Med. Microbiol. 29 (3), 229–235. doi: 10.1099/00222615-29-3-229
Santiago, G. L., Deschaght, P., Aila, N. E., Kiama, T. N., Verstraelen, H., Jefferson, K. K., et al. (2011). Gardnerella Vaginalis Comprises Three Distinct Genotypes of Which Only Two Produce Sialidase. Am. J. Obstet Gynecol. 204 (405), 1–7. doi: 10.1016/j.ajog.2010.12.061
Schellenberg, J. J., Jayaprakash, T. P., Gamage, N. W., Patterson, M. H., Vaneechoutte, M., Hill, J. E. (2016). Gardnerella Vaginalis Subgroups Defined by Cpn60 Sequencing and Sialidase Activity in Isolates From Canada, Belgium and Kenya. PLoS One 11 (1), e0146510. doi: 10.1371/journal.pone.0146510
Schellenberg, J. J., Links, M. G., Hill, J. E., Dumonceaux, T. J. (2011). Molecular Definition of Vaginal Microbiota in East African Commercial Sex Workers. Am. Soc. Microbiol. 77 (12), 4066–4074. doi: 10.1128/AEM.02943-10
Schellenberg, J. J., Patterson, M. H., Hill, J. E. (2017). Gardnerella Vaginalis Diversity and Ecology in Relation to Vaginal Symptoms. Res. Microbiol. 168, 837–844. doi: 10.1016/j.resmic.2017.02.011
Schuyler, J. A., Mordechai, E., Adelson, M. E., Sobel, J. D., Gygax, S. E., Hilbert, D. W. (2016). Identification of Intrinsically Metronidazole-Resistant Clades of Gardnerella Vaginalis. Diagn. Microbiol. Infect. Dis. 84 (1), 1–3. doi: 10.1016/j.diagmicrobio.2015.10.006
Scott, T. G., Curran, B., Smyth, C. J. (1989). Electron Microscopy of Adhesive Interactions Between Gardnerella Vaginalis and Vaginal Epithelial Cells, McCoy Cells and Human Red Blood Cells. Microbiology 135 (3), 475–480. doi: 10.1099/00221287-135-3-475
Shipitsyna, E., Khusnutdinova, T., Budilovskaya, O., Krysanova, A., Shalepo, K., Savicheva, A., et al. (2020). Bacterial Vaginosis-Associated Vaginal Microbiota is an Age-Independent Risk Factor for Chlamydia Trachomatis, Mycoplasma Genitalium and Trichomonas Vaginalis Infections in Low-Risk Women, St. Petersburg, Russia. Eur. J. Clin. Microbiol. Infect. Dis. 39 (7), 1221–1230. doi: 10.1007/s10096-020-03831-w
Shipitsyna, E., Krysanova, A., Khayrullina, G., Shalepo, K., Savicheva, A., Guschin, A., et al. (2019). Quantitation of All Four Gardnerella Vaginalis Clades Detects Abnormal Vaginal Microbiota Characteristic of Bacterial Vaginosis More Accurately Than Putative G. Vaginalis Sialidase A Gene Count. Mol. Diagnosis Ther. 23, 139–147. doi: 10.1007/s40291-019-00382-5
Swidsinski, A., Mendling, W., Loening-Baucke, V., Ladhoff, A., Swidsinski, S., Hale, L. P., et al. (2005). Adherent Biofilms in Bacterial Vaginosis. Am. Coll. Obstetricians Gynecol. 106, 865–874. doi: 10.1097/01.AOG.0000183594.45524.d2
Tabatabaei, N., Eren, A. M., Barreiro, L. B., Yotova, V., Dumaine, A., Allard, C. (2019). Vaginal Microbiome in Early Pregnancy and Subsequent Risk of Spontaneous Preterm Birth: A Case–Control Study. Gynaecology 126 (3), 349–358. doi: 10.1111/1471-0528.15299
Turner, E., Sobel, J. D., Akins, A. A. (2021). Prognosis of Recurrent Bacterial Vaginosis Based on Longitudinal Changes in Abundance of Lactobacillus and Specific Species of Gardnerella. PLoSONE 16 (8), e0256445. doi: 10.1371/journal.pone.0256445
Turovskiy, Y., Sutyak Noll, K., Chikindas, M. L. (2011). The Aetiology of Bacterial Vaginosis. J. Appl. Microbiol. 110 (5), 1105–1128. doi: 10.1111/j.1365-2672.2011.04977.x
Usyk, M., Schlecht, N. F., Pickering, S., Williams, L., Sollecito, C. C., Gradissimo, A., et al. (2022). molBV Reveals Immune Landscape of Bacterial Vaginosis and Predicts Human Papillomavirus Infection Natural History. Nat. Commun. 13 (1), 233. doi: 10.1038/s41467-021-27628-3
Vaneechoutte, M., Guschin, A., Simaey, L. V., Gansemans, Y., Nieuwerburgh, F. V., Cools, P. (2019). Emended Description of Gardnerella Vaginalis and Description of Gardnerella Leopoldii Sp. Nov., Gardnerella Piotii Sp. Nov. And Gardnerella Swidsinskii Sp. Nov., With Delineation of 13 Genomic Species Within the Genus Gardnerella. Int. J. Syst. Evol. Microbiol. 69, 679–687. doi: 10.1099/ijsem.0.003200
Verstraelen, H., Swidsinski, A. (2019). The Biofilm in Bacterial Vaginosis: Implications for Epidemiology, Diagnosis and Treatment: 2018 Update. Curr. Opin. Infect. Dis. 32 (1), 38–42. doi: 10.1097/qco.0000000000000516
Vestby, L. K., Grønseth, T., Simm, R., Nesse, L. L. (2020). Bacterial Biofilm and its Role in the Pathogenesis of Disease. Antibiotics (Basel) 9 (2), 59. doi: 10.3390/antibiotics9020059
Wiesenfeld, H. C., Hillier, S. L., Krohn, M. A., Amortegui, A. J., Heine, R. P., Landers, D. V., et al. (2002). Lower Genital Tract Infection and Endometritis: Insight Into Subclinical Pelvic Inflammatory Disease. Obstet Gynecol. 100 (3), 456–463. doi: 10.1016/s0029-7844(02)02118-x
Zilnyte, M., Venclovas, Č., Zvirbliene, A., Pleckaityte, M. (2015). The Cytolytic Activity of Vaginolysin Strictly Depends on Cholesterol and Is Potentiated by Human Cd59. Toxins 7 (2072-6651), 110–128. doi: 10.3390/toxins7010110
Zinnemann, K., Turner, G. C. (1963). The Taxonomic Position of “Haemophilus Vaginalis” [Corynebacterium Vaginale]. J. PATH BACT 85, 213–219. doi: 10.1002/path.1700850120
Keywords: Gardnerella, bacterial vaginosis, clinical outcomes, subtypes, qPCR, cpn-60, whole genome sequencing
Citation: Qin H and Xiao B (2022) Research Progress on the Correlation Between Gardnerella Typing and Bacterial Vaginosis. Front. Cell. Infect. Microbiol. 12:858155. doi: 10.3389/fcimb.2022.858155
Received: 19 January 2022; Accepted: 01 March 2022;
Published: 25 March 2022.
Edited by:
António Machado, Universidad San Francisco de Quito, EcuadorReviewed by:
Alexander Swidsinski, Charité Universitätsmedizin Berlin, GermanyCopyright © 2022 Qin and Xiao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bingbing Xiao, ZG9jdG9yeGJiQDE2My5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.