 Meng Zhou
Meng Zhou Xiao-Hong Ji2
Xiao-Hong Ji2 Hong-Gao Liu
Hong-Gao Liu Kurt Miller
Kurt Miller Yuan Yuan
Yuan Yuan Josef Vlasák
Josef Vlasák- 1Institute of Microbiology, School of Ecology and Nature Conservation, Beijing Forestry University, Beijing, China
- 2College of Pharmacy and Life Sciences, Jiujiang University, Jiujiang, China
- 3Faculty of Agronomy and Life Sciences, Zhaotong University, Zhaotong, China
- 4Urb. Bellas Lomas, Mayaguez, Puerto Rico
- 5Biology Centre of the Academy of Sciences of the Czech Republic, České Budějovice, Czechia
Two new species in Hymenochaetaceae, Fulvifomes acaciae and Pyrrhoderma nigra, are illustrated and described from tropical Asia and America based on morphology and phylogenetic analyses. F. acaciae is characterized by perennial, pileate, and woody hard basidiomata when fresh; ash gray to dark gray, encrusted, concentrically sulcate, and irregularly cracked pileal surface; circular pores of 7–8 per mm with entire dissepiments; a dimitic hyphal system in trama and context; absence of setal element and presence of cystidioles; and broadly ellipsoid, yellowish brown, thick-walled, and smooth basidiospores measuring 5–6 μm × 4–5 μm. P. nigra is characterized by perennial and resupinate basidiomata with dark gray to almost black pore surface when fresh; small and circular pores of 7–9 per mm, a monomitic hyphal system with generative hyphae simple septate, hyphoid setae dominant in subiculum but not in tube trama, and absence of cystidia; and ellipsoid, hyaline, thin-walled basidiospores measuring 4–5 μm × 3–3.6 μm. The differences between the new species and morphologically similar and phylogenetically related species are discussed. Keys to Fulvifomes and Pyrrhoderma have also been provided.
1. Introduction
Fulvifomes is a monophyletic genus in Hymenochaetaceae (Wagner and Fischer, 2002; Wu et al., 2022). However, it has been treated as a synonym of Phellinus Quél. for several decades (Ryvarden and Johansen, 1980; Gilbertson and Ryvarden, 1986–1987a; Larsen and Cobb-Poulle, 1990; Núñez and Ryvarden, 2000). The genus is characterized by basidiomata annual to perennial, effused-reflexed; pileate or substipitate, corky to woody hard; hyphal system monomitic or dimitic; generative hyphae simple septate; setal elements absent; basidiospores subglobose to ellipsoid, yellowish to brown, fairly thick- to thick-walled, smooth; mostly on angiosperms and cause a white rot (Wu et al., 2022a). Recently, molecular analyses have detected new taxa in the genus (2015c; 2022b; Zhou, 2014; Ji et al., 2017; Salvador-Montoya et al., 2018; Wu et al., 2022a). So far, 49 species have been recorded in the genus (Wu et al., 2022a).
Pyrrhoderma Imazeki is another monophyletic genus in Hymenochaetaceae (Wu et al., 2022a) and was erected with Pyrrhoderma sendaiense as the generic type by Imazeki (1966). The genus was emended by Zhou et al. (2018). It is characterized by basidiomata annual to perennial, effused-reflexed, pileate to laterally stipitate, corky to woody hard when dry; pileal surface with a cuticle or crust; hyphal system monomitic; generative hyphae simple septate; hyphoid and hymenial setae present or absent; basidiospores ellipsoid to subglobose, hyaline, thin-walled; on angiosperm wood and cause a white rot. Previously, seven species were accepted in Pyrrhoderma (Wu et al., 2022a).
The pioneer phylogeny of Hymenochaetaceae was made by Wagner and Fischer (2001) based on limited samples, and more phylogenetic analyses were contributed based on more samples (2015b; 2016a; 2016b; Dai, 2010; Zhou, 2015a; Rajchenberg et al., 2015; Wu et al., 2016, 2022; Miettinen et al., 2019). Phylogenies of Fulvifomes were recently analyzed, and many new species were described (Zhou, 2015c; Ji et al., 2017; Tchoumi et al., 2020; Hattori, et al., 2022). Pyrrhoderma is a small genus in Hymenochaetaceae, and Zhou et al. (2018) published a comprehensive phylogeny on the genus.
During an investigation on tropical Asian and American hymenochaetaceous fungi, samples with morphological characteristics fit definitions of Fulvifomes and Pyrrhoderma. Phylogenetically, these have formed two distinct lineages within Fulvifomes and Pyrrhoderma, respectively. Therefore, in the present paper, we described two new species in Hymenochaetaceae.
2. Materials and methods
2.1. Morphological studies
Our studied specimens have been deposited in the herbarium of the Institute of Microbiology, Beijing Forestry University (BJFC), the private herbarium of Josef Vlasák (JV), and the National Museum Prague of Czech Republic (PRM). The sections were prepared in 5% potassium hydroxide (KOH), Melzer’s reagent (IKI), and cotton blue (CB). The following abbreviations are used: KOH, 5% potassium hydroxide; IKI, Melzer’s reagent; IKI–, neither amyloid nor dextrinoid; CB, cotton blue; CB–, acyanophilous; CB+, cyanophilous after 12 h stained with cotton blue; L, mean spore length (arithmetic average of spores); W, mean spore width (arithmetic average of spores); Q, variation in the ratios of L/W between specimens studied; and n, number of basidiospores measured from a given number of specimens. The microscopic procedure follows Dai (2010), and the special color terms follow Petersen (1996) and Anonymous (1969). Sections were studied at magnifications up to ×1,000 using a Nikon Eclipse 80i microscope with phase contrast illumination. Drawings were made with the aid of a drawing tube. Microscopic features, measurements, and illustrations were made from the slide preparations stained with CB. Basidiospores were measured from sections cut from the tubes.
2.2. DNA extraction, amplification, and sequencing
The extraction of total genomic DNA from frozen specimens followed Góes-Neto et al. (2005) using the protocol of Cetyltrimethyl Ammonium Bromide (CTAB) 2%. The CTAB rapid plant genome extraction kit-DN14 (Aidlab Biotechnologies Co., Ltd., Beijing) was used to obtain PCR products from dried specimens, following the manufacturer’s instructions with some modifications (2016; Chen et al., 2015). The internal transcribed spacer (ITS) region was amplified with the primer pairs ITS5 and ITS4 (White et al., 1990). For the large subunit nuclear ribosomal RNA gene (nLSU), the primer pairs LR0R and LR7 (Vilgalys and Hester, 1990) and LR0R and LR5 (White et al., 1990) were used for PCR amplification. The PCR procedure for ITS was as follows: initial denaturation at 95°C for 3 min, followed by 34 cycles of denaturation at 94°C for 40 s, annealing at 54°C for 45 s, and extension at 72°C for 1 min, followed by a final extension at 72°C for 10 min. The PCR procedure for 28S was as follows: initial denaturation at 94°C for 1 min followed by 35 cycles at 94°C for 30 s, 50°C for 1 min, 72°C for 1.5 min, and a final extension of 72°C for 10 min. The PCR products were purified and directly sequenced at Beijing Genomics Institute. The sequence quality was checked following Nilsson et al. (2012).
2.3. Phylogenetic analyses
The phylogenetic trees were constructed using sequences obtained in this study and additional sequences downloaded from GenBank (Tables 1, 2). Both ITS and 28S datasets were aligned within MAFFT version 7 (Katoh et al., 2019) and ClustalX (Thompson et al., 1997), followed by manual proofreading in BioEdit (Hall, 1999). Ambiguous regions were deleted, and gaps were manually adjusted to optimize alignment before phylogenetic analyses. Phellinus betulinus (Murrill) Parmasto and P. populicola Niemelä were used as outgroups in the phylogeny of Fulvifomes (Wu et al., 2022a; Figure 1). Oxyporus populinus (Schumach.) Donk was used as an outgroup in the phylogeny of Pyrrhoderma (Zhou et al., 2018; Figure 2). Each data matrix was edited in Mesquite version 3.70 (Maddison and Maddison, 2021).
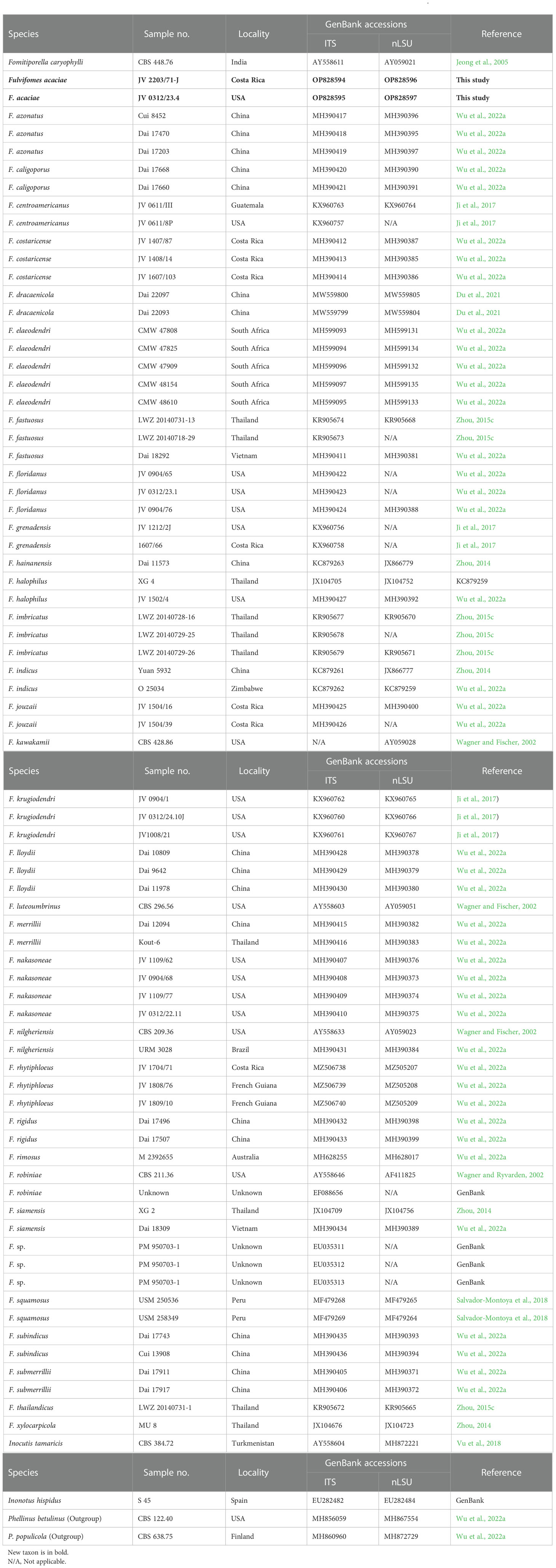
Table 1 Taxa, voucher specimens, and GenBank accession numbers of sequences used in the phylogeny of Fulvifomes.
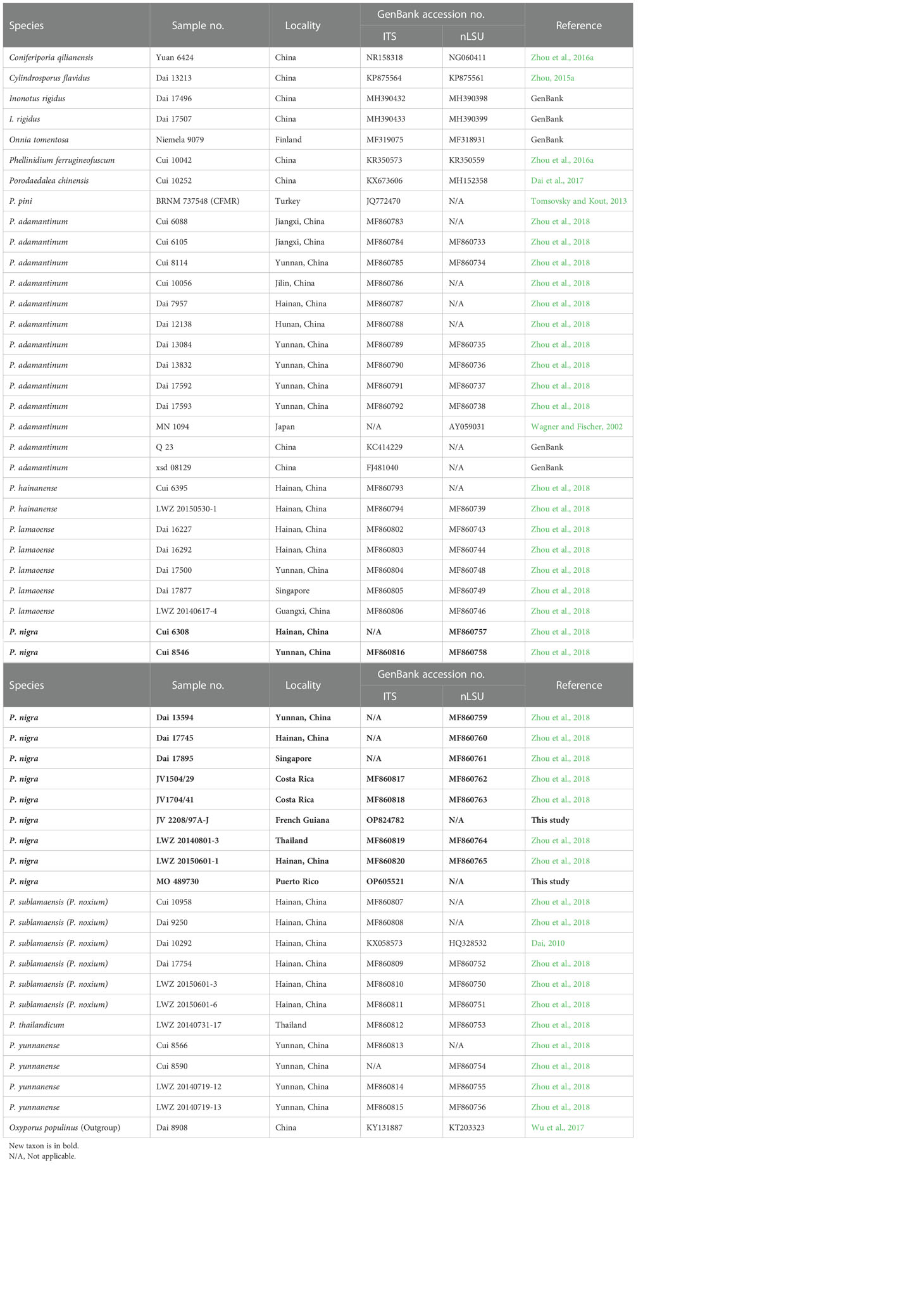
Table 2 Taxa, voucher specimens, and GenBank accession numbers of sequences used in the phylogeny of Pyrrhoderma.
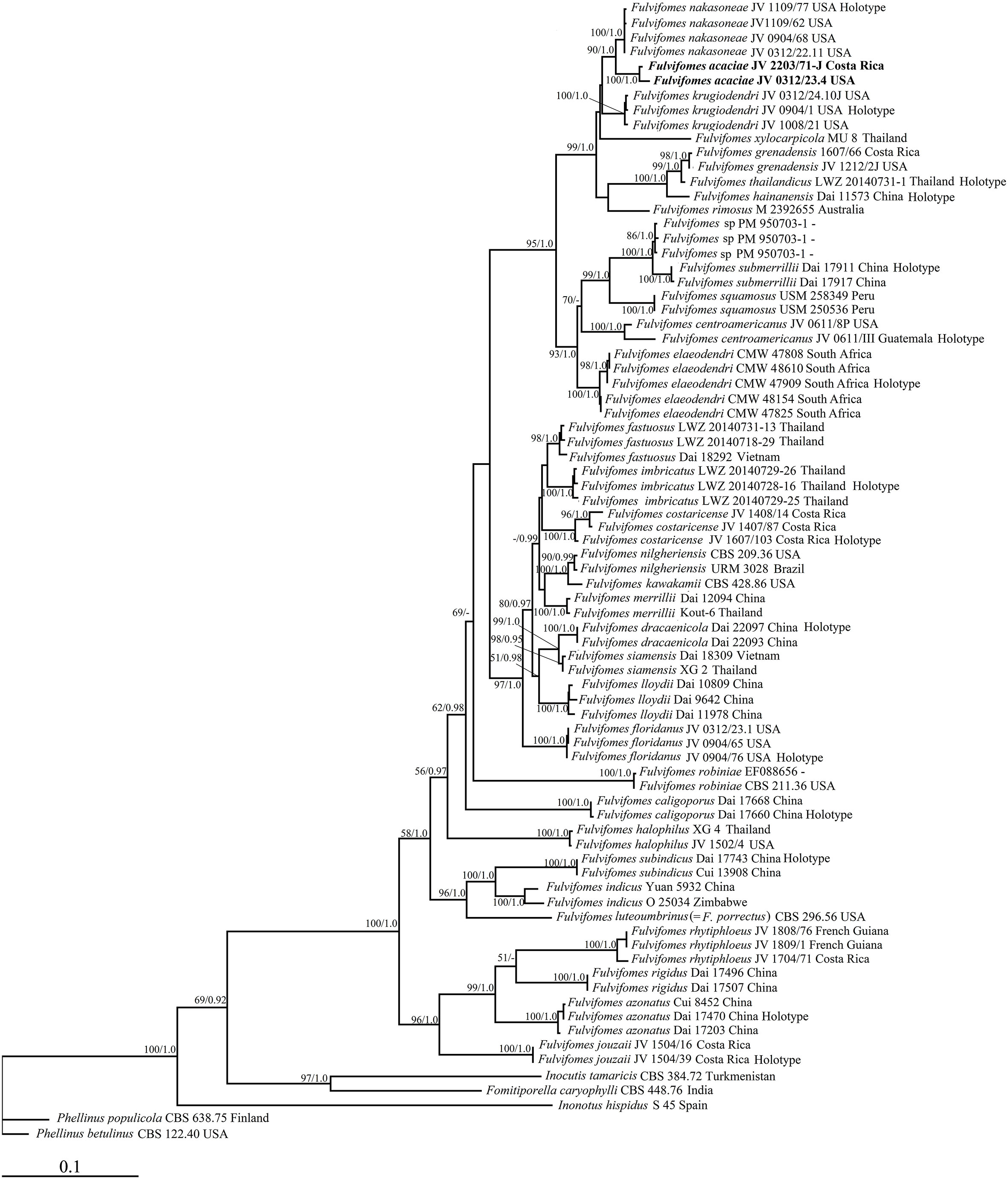
Figure 1 Maximum likelihood tree illustrating the phylogeny of Fulvifomes based on the combined dataset of ITS+28S sequences. Phellinus betulinus (MH856059; MH867554) and P. populicola (MH860960; MH872729) were used as outgroups. The maximum likelihood bootstrap values (≥50) and Bayesian posterior probability values (≥0.90) are indicated above the branches. The new species is in bold.
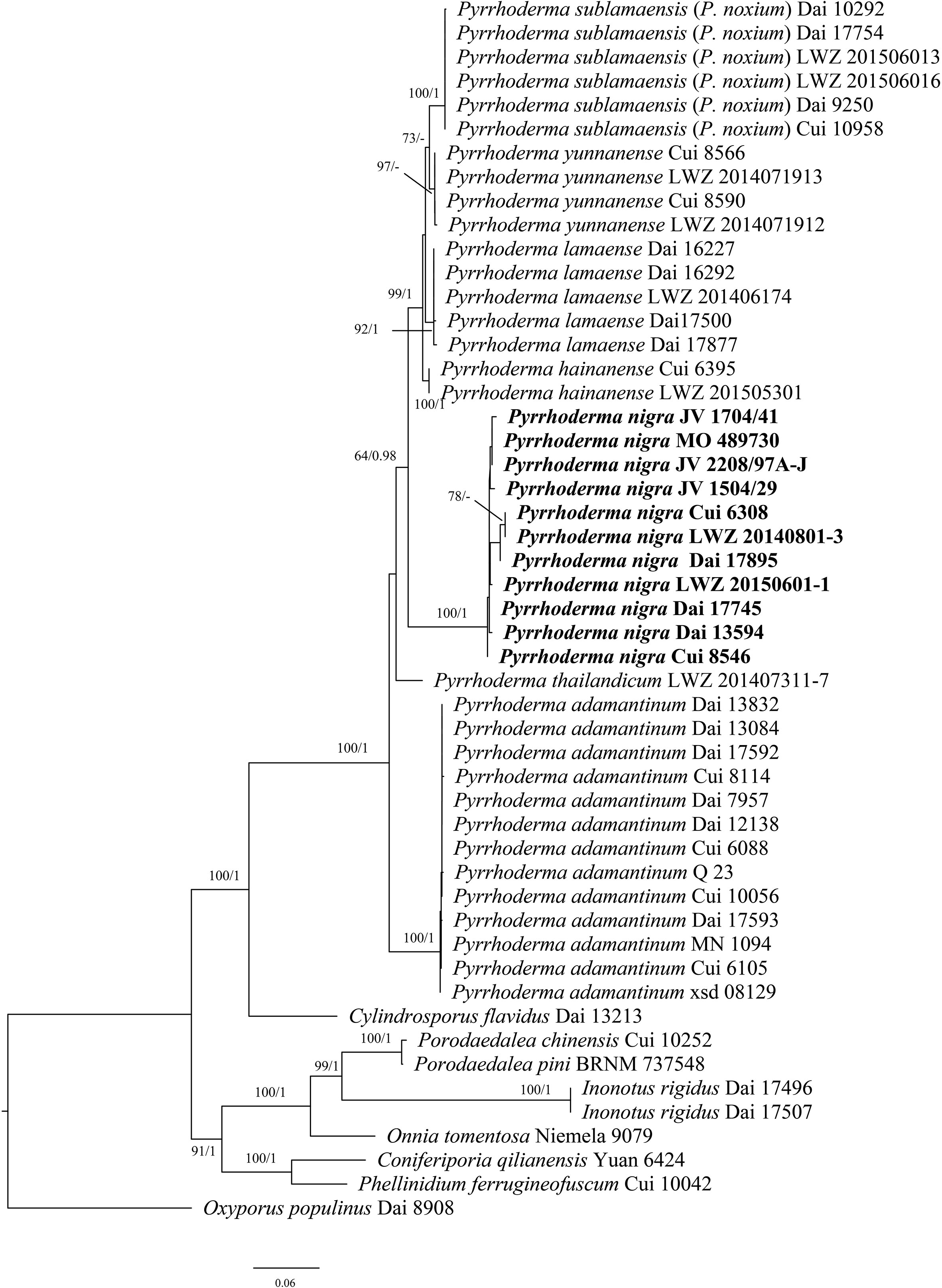
Figure 2 Maximum likelihood tree illustrating the phylogeny of Pyrrhoderma based on the combined dataset of ITS+28S sequences. Oxyporus populinus (KY131887; KT203323) was used as an outgroup. The maximum likelihood bootstrap values (≥50) and Bayesian posterior probability values (≥0.90) are indicated above the branches. The new species is in bold.
Phylogenetic analyses were conducted using maximum likelihood (ML) and Bayesian Inference (BI) based on ITS+28S aligned datasets using RAxML version 8.2.12 (Stamatakis, 2014) and MrBayes version 3.2.6 (Ronquist et al., 2012). Sequence alignments were deposited at TreeBase (http://purl.org/phylo/treebase; submission ID 29762 and 29862).
GTR+I+G was estimated as the best-fit evolutionary model for the resulting alignments from these two datasets with jModelTest (Guindon and Gascuel, 2003; Posada, 2008). RAxML version 8.2.12 (Stamatakis, 2014) was applied in the ML analysis. All parameters in the ML analysis were kept at default settings.
The BI was calculated with MrBayes version 3.2.6 (Ronquist et al., 2012) in two independent runs, each of which had four chains for 1.5 million generations that were initiated using random trees. Trees were sampled every 100 generations. The first 25% of the sampled trees were discarded as burn-in, whereas other trees were used to construct a 50% majority consensus tree and for calculating Bayesian posterior probabilities (BPPs).
The two methods constructed nearly congruent topologies for each alignment. Therefore, only the topology generated from the ML analysis is presented along with the bootstrap support for ML (BS) values and BPPs, simultaneously at the nodes. Phylogenetic trees were visualized using FigTree version 1.4.4 (Rambaut, 2018). Branches that received bootstrap support for ML (BS) and BPPs (≥75% for BS and 0.95 for BPPs) were considered as significantly supported.
3. Results
3.1. Phylogeny
Permission in the phylogenetic analysis of Fulvifomes (Figure 1), 79 fungal collections representing 37 taxa of Fulvifomes were included in the phylogenetic analyses and two samples of genus Phellinus were used as outgroups. The final alignment comprised a total of 1,922 base pairs (bp), including 1,032 bp of ITS and 890 bp of 28S. The best model for the combined ITS+28S dataset was estimated and applied in the Bayesian analysis: GTR+I+G, lset nst = 6, rates = invgamma; prset statefreqpr = dirichlet (1,1,1,1). Bayesian analysis resulted in an average standard deviation of split frequencies as 0.009512. As both ML and BI trees resulted in similar topologies, only the topology from the ML analysis is presented along with statistical values from the ML (≥50%) and BPP (≥0.9) algorithms (Figure 1).
In the phylogenetic analysis of Pyrrhoderma (Figure 2), the ITS+28S sequences from 51 fungal collections representing 15 species were used. The final alignment comprised a total of 1,686 bp, including 814 bp of ITS and 872 bp of 28S. The best model for the combined ITS+28S dataset was estimated and applied in the Bayesian analysis: GTR+I+G, lset nst = 6, rates = invgamma; prset statefreqpr = dirichlet (1,1,1,1). Bayesian analysis resulted in an average standard deviation of split frequencies = 0.006184. Both ML and BI trees resulted in similar topologies; thus, only the topology from the ML analysis is presented along with statistical values from the ML (≥50%) and BPP (≥0.9) algorithms (Figure 2).
3.2. Taxonomy
Fulvifomes acaciae Meng Zhou, Yuan & Vlasák, sp. nov. Figures 3, 4.

Figure 3 A basidiomata of Fulvifomes acaciae (holotype, JV 2203/71-J). Scale bar: 1 cm.
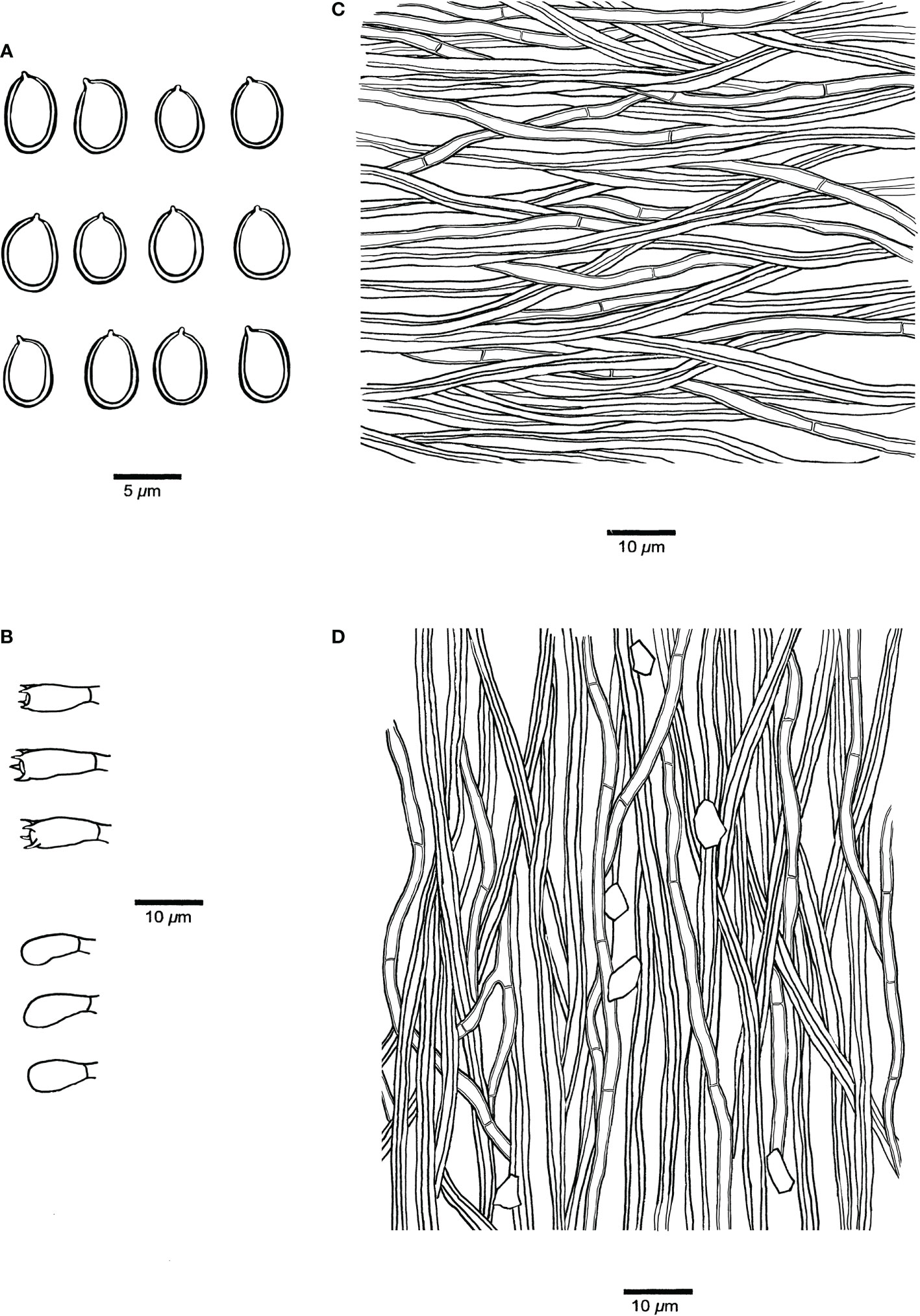
Figure 4 Microscopic structures of Fulvifomes acaciae (JV 0312/23.4-J). (A) Basidiospores. (B) Basidia and basidioles. (C) Hyphae from context. (D) Hyphae from tube trama.
MycoBank: MB xxxxxx
Type. Costa Rica, Mt. Rincon, Guachipelin, on living tree of Acacia, March 2022, JV 2203/71-J (isotype, BJFC).
Etymology. Acaciae (Lat.): referring to the species growing on Acacia.
Fruiting body. Basidiomata perennial, pileate, solitary, without distinctive odor or taste and woody hard when fresh, light in weight when dry. Pilei ungulate, projecting up to 20 cm and 15 cm wide and 7 cm thick at base. Pileal surface ash gray to dark gray when dry, encrusted, rough, concentrically sulcate, irregularly cracked; pileal margin dark gray, obtuse. Pore surface umber, glancing; sterile margin distinct, fulvous, up to 3 mm wide; pores circular, 7–8 per mm; dissepiments thick, entire. Context fulvous, woody hard, zonate, up to 5 mm thick. Tubes concolorous with context, woody hard, up to 6.5 cm long, tube layers indistinctly stratified.
Hyphal structure. Hyphal system dimitic in trama and context; generative hyphae simple septate; tissue becoming blackish brown in KOH.
Context. Generative hyphae hyaline to pale yellow, thin- to thick-walled, rarely branched, frequently simple septate, 2–3 μm in diameter; skeletal hyphae dominant, yellowish to golden yellow, thick-walled with a narrow to wide lumen, unbranched, aseptate, more or less straight, regularly arranged, 3–4 μm in diameter.
Tubes. Generative hyphae hyaline to pale yellow, thin- to slightly thick-walled, rarely branched, frequently simple septate, 2–3.5 μm in diameter; skeletal hyphae frequent, yellowish to golden yellow, thick-walled with a narrow to wide lumen, unbranched, aseptate, more or less straight, subparallel along tubes, 3–4.5 μm in diameter. Setae or setal hyphae absent; cystidioles absent; basidia barrel-shaped, with four sterigmata and a simple basal septum, 10–12 μm × 5–6 μm; basidioles in shape similar to basidia, slightly smaller than basidia. Big rhomboid crystals present in hymenia and trama.
Spores. Basidiospores broadly ellipsoid, yellowish brown, thick-walled, smooth, some collapsed, IKI–, CB–, (4.9–)5–6(–6.1) μm × (3.9–)4–5(–5.1) μm, L = 5.26 μm, W = 4.35 μm, Q = 1.19–1.23 (n = 60/2).
Additional specimen (paratype) examined. USA, Florida, Florida Keys, Key Largo, John Pennekamp Coral Reef State Park, December 2003, Josef Vlasák leg., on fallen trunk of Acacia, JV 0312/23.4-J (BJFC032898).
Pyrrhoderma nigra Meng Zhou, Yuan Yuan & Vlasák, sp. nov. Figures 5, 6

Figure 5 A basidiomata of Pyrrhoderma nigra (MO 489730). Scale bar: 1 cm.
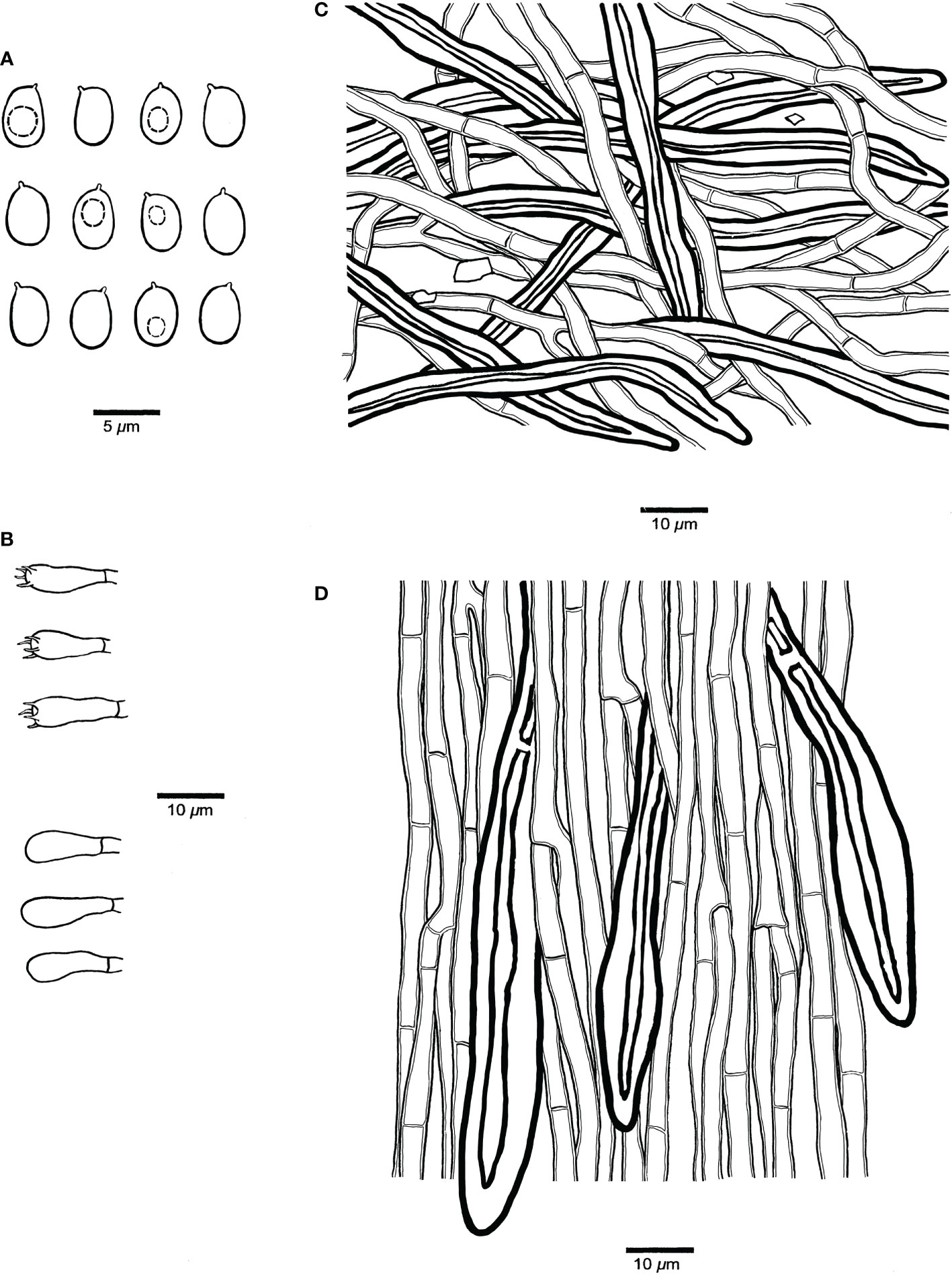
Figure 6 Microscopic structures of Pyrrhoderma nigra (holotype, Cui 8546). (A) Basidiospores. (B) Basidia and basidioles. (C) Hyphae from subiculum. (D) Hyphae from tube trama.
MycoBank: MB xxxxxx.
Type. China, Yunnan Province, Mengla County, Wangtianshu Forest Park, 2 November 2009, Bao-Kai Cui leg., on fallen angiosperm trunk, Cui 8546 (BJFC 007035).
Etymology. Nigra (Lat.): referring to having black pore surface when fresh.
Fruiting body. Basidiomata perennial, resupinate, firmly attached to the substrate, separable, up to 30 cm long, 16 cm wide and 8 mm thick at center, without odor or taste when fresh, woody hard and brittle when dry. Pore surface dark gray to almost black when fresh, becoming grayish brown when dry, glancing; sterile margin very narrow to almost absent, dark brown; pores mostly circular, 7–9 per mm; dissepiments thick, entire. Subiculum chestnut, woody hard, up to 3 mm thick. Tubes deep olive, woody hard to brittle, up to 3 mm long.
Hyphal structure. Hyphal system monomitic; generative hyphae simple septate; tissue darkening but otherwise unchanged in KOH.
Subiculum. Subicular hyphae yellowish to golden yellow, thick-walled with a wide lumen, occasionally branched, frequently simple septate, interwoven, some encrusted with fine crystals, 4–5.5 µm diameter; hyphoid setae dark brown, distinctly thick-walled with a narrow lumen, straight, apex obtuse and not encrusted, up to a few hundreds of µm long, 5–8 µm diameter.
Tubes. Tramal hyphae pale yellowish to yellow, thin- to thick-walled with a wide lumen, gelatinized, frequently branched, frequently simple septate, parallel along the tubes, 3–4.5 µm diameter; hyphoid setae frequent, but not dominant, dark brown, distinctly thick-walled with a narrow lumen, straight, apex obtuse or pointed, and sometimes encrusted with fine hyaline crystals, frequently projecting out of hymenium, up to a few hundreds of micrometers long, 8–12 µm diameter; cystidia and cystidioles absent; basidia barrel-shaped, with four sterigmata and a simple septum at the base, 7–9 µm × 4–4.5 µm; basidioles more or less pyriform, slightly smaller than basidia.
Spores. Basidiospores ellipsoid, hyaline, thin-walled, some with a big guttule, IKI–, CB–, 4–5 μm × 3–3.6(–4) μm, L = 4.43 µm, W = 3.34 µm, Q = 1.33 (n = 30/1).
Additional specimens (paratypes) examined. China: Hainan Province, Ledong County, Jianfengling National Nature Reserve, on fallen angiosperm trunk, 1 June 2015, LWZ 20150601-1 (IFP 019170). Costa Rica, Golfito, Playa Cacao, 19.IV.2015, JV 1504/29 (JV), Playa Nicuesa, 18.IV.2017, JV 1704/41(JV). French Guiana, Roura, Camp Cayman, 27.VIII.2018, JV 1808/107 (BJFC032959), St. Laurent du Maroni, Gite Moutouchi, on fallen angiosperm trunk, 31.VIII.2022, JV 2208/97A-J. (JV, BJFC) Puerto Rico, Mayagüez, Miradero, Papaya House, on fallen mango trunk, 4.8.2022, Kurt Miller MO 489730 (JV, BJFC).
4. Discussion
Macromorphologically, Fulvifomes krugiodendri has perennial, solitary, ungulate basidiomata; its pileal surface is dark gray, encrusted, concentrically sulcate with narrow zones, cracked with age; its pores as 7–9 per mm with thick and entire dissepiments. Microscopically, it has a dimitic hyphal system in both context and tube trama. Morphologically, F. krugiodendri is similar to F. acaciae, and both species are also closely related in our phylogeny (Figure 1). However, F. krugiodendri differs from F. acaciae by its subglobose basidiospores measuring 4.3–5 μm × 4–4.5 μm, L = 4.6 μm, W = 4.21 μm, Q = 1.08–1.09, interwoven tramal hyphae, and the absence of rhomboid crystals, and it lives on Krugiodendron (Ji et al., 2017). In addition, the nucleotide difference of ITS sequences between the two species is 3%.
Morphologically, Pyrrhoderma nigra is very similar to the resupinate Pyrrhoderma lamaoense (Murrill) L.W. Zhou & Y.C. Dai and P. sublamaensis (Lloyd) Y.C. Dai & F. Wu, but the latter two species have effused-reflexed to pileate basidiomata, the presence of cystidia, especially thinner basidiospores (2–2.4 μm vs. 3–3.6 µm, Wu et al., 2022). P. nigra and Pyrrhoderma thailandicum L. W. Zhou & Y.C. Dai share similar basidiospores (4–5 µm × 3–3.6 µm vs. 4–4.5 µm × 3–3.5 µm), but the latter differs from the former by annual basidiomata, bigger pores (3–5 per mm vs. 7–9 per mm) and the absence of setal elements.
Most species of Fulvifomes and Pyrrhoderma have been recorded in the tropics (Wu et al., 2022). The two new species described in the present study were found from tropical Asia and America. Similar to other polypores, species of Hymenochaetaceae is very rich in the tropics (Dai et al., 2021). So far, 50 and 8 species of Fulvifomes and Pyrrhoderma, respectively, have been identified, and identification keys to the species in the two genera are given below.
Key to species of Fulvifomes
1. Basidiocarps annual ..........................................................................2
1. Basidiocarps perennial .....................................................................5
2. Pores 4–5 per mm ....................F. indicus (Massee) L. W. Zhou
2. Pores 7–10 per mm ..........................................................................3
3. Basidiocarps resupinate; basidiospores ellipsoid...........................................F. rigidus (B.K. Cui & Y.C. Dai) X.H. Ji & Jia J. Chen
3. Basidiocarps pileate; basidiospores globose to subglobose......... 4
4. Pileal surface without a black cuticle; hyphal system dimitic........ F. aureobrunneus (J.E. Wright & Blumenf.) Y.C. Dai & F. Wu
4. Pileal surface with a black cuticle; hyphal system monomitic.............................F. luteoumbrinus (Romell) Y. C. Dai et al.
5. Chlamydospores present.................................................................. 6
5. Chlamydospores absent.....................................................................9
6. Basidiospores 5–8 µm long............................................................... 7
6. Basidiospores 4–5 µm long............................................................... 8
7. Pilei ungulate, tube layers separated by a thin context layer..... F. scaber (Berk.) Y.C. Dai & F. Wu
7. Pilei globose, tubes indistinctly stratified without context layer..........F. kravtzevii (Schwarzman) Y.C. Dai & F. Wu
8. Basidiocarps imbricate, pileal surface with a cuticle....................F. kawakamii (M.J. Larsen et al.) T. Wagner & M. Fisch.
8. Basidiocarps solitary, pileal surface without a cuticle.................F. durissimus (Lloyd) Bondartseva & S. Herrera
9. Basidiospores oblong-ellipsoid.......................................................10
9. Basidiospores ellipsoid, broadly ellipsoid, ovoid, subglobose or globose.......................................................................................................11
10. Pores 5–6 per mm; basidiospores 4.2–5.1 µm long....... F. collinus (Y.C. Dai & Niemelä) Y.C. Dai
10. Pores 7–8 per mm; basidiospores 3–3.6 µm long.......................F. fushanianus (T.T. Chang) Y.C. Dai & F. Wu
11. Tramal hyphae monomitic...........................................................12
11. Tramal hyphae dimitic..................................................................14
12. Basidiospores ellipsoid, <4 µm wide............................................F. caligoporus Y.C. Dai & X.H. Ji
12. Basidiospores ovoid or subgolobose, >4 µm wide....................13
13. Context without a granular core, pores 5–7 per mm.....F. lloydii (Cleland) Y.C. Dai & X.H. Ji
13. Context with a granular core, pores 3–4 per mm......................F. resinaceus (Kotl. & Pouzar) Y.C. Dai & F. Wu
14. Hyphae monomitic in context.....................................................15
14. Hyphae dimitic or subdimitic in context....................................25
15. Pileal surface uncracked................................................................16
15. Pileal surface cracked or rimose...................................................20
16. Hyphae at pileal surface with thin-walled and septate tips; on Newtonia buchananii; African species.................................F. newtoniae (Niemelä & Mrema) Y.C. Dai & F. Wu
16. Hyphae at pileal surface without thin-walled and septate tips; on an angiosperm other than Newtonia; Asian or American species............................17
17. Basidiospores >5.5 μm long...................................F. mangrovicus (Imazeki) T. Hatt.
17. Basidiospores <5.5 μm long..........................................................18
18. On Dracaena................................................F. dracaenicola Z.B. Liu & Y.C. Dai
18. On an angiosperm other than Dracaena....................................19
19. Pore surface not glancing; basidiospores CB+; Asian species.......F. subindicus Y.C. Dai & X.H. Ji
19. Pore surface glancing; basidiospores CB–; American species........F. floridanus Y.C. Dai & Vlasák
20. Pileal surface squamose with long scales.........F. squamosus Salvador-Montoya & Drechsler-Santos
20. Pileal surface glabrous or tomentose without long scales........21
21. Basidiospores globose, >5 µm wide...............................F. cedrelae (Murrill) Murrill
21. Basidiospores ovoid, broadly ellipsoid to subglobose, <5 µm wide..........................................................................................................22
22. Pileal surface with a black crust....................................................23
22. Pileal surface without crust...........................................................24
23. Pores 4–7 per mm; basidiospores 3–4 µm wide.................F. grenadensis (Murrill) Murrill
23. Pores 7–8 per mm; basidiospores 4–5 µm wide......................F. siamensis T. Hatt. et al.
24. Pore surface dull chocolate brown, pores 4–5 per mm................F. rimosus (Berk.) Fiasson & Niemelä
24. Pore surface yellowish to reddish brown, pores 7–8 per mm.....................F. robiniae (Murrill) Murrill
25. Pileal surface azonate.....................................................................26
25. Pileal surface concentrically zonate.............................................28
26. Basidiospores 4.5–6 μm wide........................................F. crocatus (Fr.) Y.C. Dai & F. Wu
26. Basidiospores 3–4 μm wide..........................................................27
27. Pore surface not glancing, pores 7–9 per mm...........F. azonatus Y.C. Dai & X.H. Ji
27. Pore surface glancing, pores 5–7 per mm................F. swieteniae Murrill
28. Pileal surface cracked.....................................................................29
28. Pileal surface uncracked................................................................37
29. Pores 7–11 per mm.........................................................................30
29. Pores 4–7 per mm...........................................................................32
30. Pilei triquetrous, pore surface dark brown, not glancing.........F. minutiporus (Bond. & Herrera) Y.C. Dai & F. Wu
30. Pilei ungulate, pore surface grayish brown to umber, glancing............31
31. Basidiospores subglobose, 4.3–5 μm × 4–4.5 μm......................F. krugiodendri Y.C. Dai et al.
31. Basidiospores subglobose broadly ellipsoid, 5–6 μm × 4–5 μm................................................................................................................................ F. acaciae
32. Growing on Pseudocedrela or Elaeodendron; African species..............................................................................................33
32. Growing on an angiosperm other than Pseudocedrela and Elaeodendron; Asian and American species............................................34
33. Context with a black line; on Elaeodendron croceum...............F. elaeodendri Tchotet et al.
33. Context without a black line; on Pseudocedrela kotschyi.........F. yoroui Olou & F. Langer
34. Basidiospores mostly 5–6 μm wide......................F. coffeatoporus (Kotl. & Pouzar) Y.C. Dai & F. Wu
34. Basidiospores mostly 3.5–5 μm wide..........................................35
35. Pore surface not glancing; American species.............................F. nakasoneae Y.C. Dai & Vlasák
35. Pore surface glancing; Asian species...........................................36
36. Pileal surface matted, not encrusted; cystidioles absent...........F. xylocarpicola T. Hatt. et al.
36. Pileal surface encrusted; cystidioles present........F. thailandicus L.W. Zhou
37. Pores 3–4 per mm.................................F. hainanensis L.W. Zhou
37. Pores 4–11 per mm.........................................................................38
38. Cystidioles present.........................................................................39
38. Cystidioles absent...........................................................................41
39. A thin black line present........................................between context and substrate F. allardii (Bres.) Bondartseva & S. Herrera
39. A thin black line absent.................................................................40
40. Pores 4–5 per mm; basidiospores ellipsoid to reniform..........F. merrillii (Murrill) Baltazar & Gibertoni
40. Pores 6–7 per mm; basidiospores ellipsoid...........F. submerrillii X.H. Ji & Jia J Chen
41. Growing on Abies; context very thin to almost lacking............F. acontextus (Ryvarden) Y.C. Dai & F. Wu
41. Growing angiosperm wood; distinct context present..............42
42. Basidiocarps usually effused-reflexed to pileate........................43
42. Basidiocarps distinctly pileate......................................................44
43. Pileal surface dark brown to black; growing exclusively on Xylocarpus........................................................F. halophilus T. Hatt. et al.
43. Pileal surface luteous brown; growing on an angiosperm other than Xylocarpus..........................................F. mcgregorii (Bres.) Y.C. Dai
44. Basidiospores 5–6 µm long...........................................F. fastuosus (Lév.) Bondartseva & S. Herrera
44. Basidiospores 4–5 µm long...........................................................45
45. Basidiospores globose..............................................F. rhytiphloeus (Mont.) Camp.-Sant. & Robledo
45. Basidiospores broadly ellipsoid to subglobose..........................46
46. Pilei ungulate; basidiospores <3.7 μm wide...................F. jouzaii Y.C. Dai & F. Wu
46. Pilei applanate, dimidiate or semicircular; basidiospores >3.7 μm wide.........................................................................................................47
47. Pileal surface encrusted.................................................................48
47. Pileal surface not encrusted..........................................................49
48. Basidiospores 3.9–4.5 μm long; Central American species......F. centroamericanus Y.C. Dai et al.
48. Basidiospores 4.6–5.1 μm long; Asian species.......F. imbricatus L.W. Zhou
49. Pores 9–11 per mm..................F. costaricense Y.C. Dai & Vlasák
49. Pores 7–9 per mm.....................................F. nilgheriensis (Mont.) Bondartseva & S. Herrera
Key to species of Pyrrhoderma
1. Hyphoid setae absent.........................................................................2
1. Hyphoid setae present........................................................................3
2. Pores 5–6 per mm; basidiospores 6–7 μm long...........................P. adamantinum (Berk.) Imazeki
2. Pores 3–5 per mm; basidiospores 4–4.5 μm long........................P. thailandicum L.W. Zhou & Y.C. Dai
3. Pores 2–4 per mm; dissepiments lacerate...............P. luteofulvum (Cleland & Rodway) Y.C. Dai & F. Wu
3. Pores 6–9 per mm; dissepiments entire..........................................4
4. Hymenial setae present..............................................P. yunnanense L.W. Zhou & Y.C. Dai
4. Hymenial setae absent........................................................................5
5. Basidiocarps annual.....................................................P. hainanense L.W. Zhou & Y.C. Dai
5. Basidiocarps perennial.......................................................................6
6. Basidiospores 3–3.6 µm wide................................................P. nigra
6. Basidiospores 2–2.4 μm wide............................................................7
7. Contextual hyphae interwoven, basidiospores oblong-ellipsoid, 3.2–4.3 μm long...............P. lamaoense (Murrill) L.W. Zhou & Y.C. Dai
7. Contextual hyphae regularly arranged, basidiospores ellipsoid, 2.6–3.3 μm long.....................P. sublamaensis (Lloyd) Y.C. Dai & F. Wu
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/supplementary material.
Author contributions
MZ, YY and JV coordinated the project and designed the experimental plan. MZ and YY analyzed the data with help from XHJ and JV. MZ, YY, HGL, KM and JV collected the samples from the field. MZ, X-HJ and YY writing the original draft preparation. MZ, X-HJ, YY and JV review and editing the manuscript. YY and JV acquire funding. All authors contributed to the article and approved the submitted version.
Funding
The research was financed by National Natural Science Foundation of China (Project Nos. 32161143013 and 32011540380) and by the institutional support of the Academy Sciences of the Czech Republic (RVO: 60077344).
Acknowledgments
We thank Prof. Bao-Kai Cui (Institute of Microbiology, School of Ecology and Nature Conservation, Beijing Forestry University) for allowing us to studying his specimens.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Anonymous (1969). Flora of British fungi. colour identification chart (London: Her Majesty’s Stationery Office), 1–3.
Chen, J. J., Cui, B. K., Dai, Y. C. (2016). Global diversity and molecular systematics of Wrightoporia s.l. (Russulales, basidiomycota). Persoonia 37, 21–36. doi: 10.3767/003158516X689666
Chen, J. J., Cui, B. K., Zhou, L. W., Korhonen, K., Dai, Y. C. (2015). Phylogeny, divergence time estimation, and biogeography of the genus Heterobasidion (Basidiomycota, russulales). Fungal Divers. 71, 185–200. doi: 10.1007/s13225-014-0317-2
Dai, Y. C. (2010). Hymenochaetaceae (Basidiomycota) in China. Fungal Divers. 45, 131–343. doi: 10.1007/s13225-010-0066-9
Dai, S. J., Vlasak, J., Tomsovsky (2017). Porodaedalea chinensis (Hymenochaetaceae, basidiomycota) - a new polypore from China. Mycospere 8, 986–993. doi: 10.5943/mycosphere/8/6/2
Dai, Y. C., Yang, Z. L., Cui, B. K., Wu, G., Yuan, H. S., Zhou, L. W., et al. (2021). Diversity and systematics of the important macrofungi in Chinese forests. Mycosystema 40, 770–805.
Du, P., Cao, T. X., Wu, Y. D., Zhou, M., Liu, Z. B. (2021). Two new species of Hymenochaetaceae on Dracaena cambodiana from tropical China. MycoKeys 80, 1–17. doi: 10.3897/mycokeys.80.63997
Gilbertson, R. L., Ryvarden, L. (1986–1987a). North American polypores 1-2. Oslo: Fungiflora, 1–885.
Góes-Neto, A., Loguercio-Leite, C., Guerrero, R. T. (2005). DNA Extraction from frozen field-collected and dehydrated herbarium fungal basidiomata: performance of SDS and CTAB-based methods. Biotemas 18, 19–32.
Guindon, S., Gascuel, O. (2003). A simple, fast and accurate method to estimate large phylogenies by maximum-likelihood. Syst. Biol. 52, 696–704. doi: 10.1080/10635150390235520
Hall, T. A. (1999). Bioedit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 41, 95–98.
Hattori, T., Ota, Y., Sotome, K. (2022). Two new species of Fulvifomes (Basidiomycota, hymenochaetaceae) on threatened or near threatened tree species in Japan. Mycoscience 63, 131–141. doi: 10.47371/mycosci.2022.04.002
Jeong, W. J., Lim, Y. W., Lee, J. S., Jung, H. S. (2005). Phylogeny of Phellinus and related genera inferred from combined data of ITS and mitochondrial SSU rDNA sequences. J. Microbiol. biot 15, 1028–1038. doi: 10.1007/s10295-005-0024-9
Ji, X. H., Wu, F., Dai, Y. C., Vlasák, J. (2017). Two new species of Fulvifomes (Hymenochaetales, basidiomycota) from America. MycoKeys 22, 1–13. doi: 10.3897/mycokeys.22.12380
Katoh, K., Rozewicki, J., Yamada, K. D. (2019). MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Briefings Bioinf. 20, 1160–1166. doi: 10.1093/bib/bbx108
Larsen, M., Cobb-Poulle, L. A. (1990). Phellinus (Hymenochaetaceae). a survey of the world taxa. Synopsis Fungorum 3, 1–206. doi: 10.2307/3760175
Maddison, W. P., Maddison, D. R. (2021)Mesquite: A modular system for evolutionary analysis (Accessed 4 October 2021).
Miettinen, O., Larsson, K. H., Spirin, V. (2019). Hydnoporia, an older name for Pseudochaete and Hymenochaetopsis, and typification of the genus Hymenochaete (Hymenochaetales, basidiomycota). Fungal Syst. Evol. 4, 77–96. doi: 10.3114/fuse.2019.04.0
Nilsson, R. H., Tedersoo, L., Abarenkov, K., Ryberg, M., Kristiansson, E., Hartmann, M., et al. (2012). Five simple guidelines for establishing basic authenticity and reliability of newly generated fungal ITS sequences. MycoKeys 4, 37–63. doi: 10.3897/mycokeys.4.3606
Núñez, M., Ryvarden, L. (2000). East Asian Polypores 1. ganodermataceae and hymenochaetaceae. Synopsis Fungorum 13, 1–168.
Petersen, J. H. (1996). The Danish mycological society’s colour-chart. Greve: Foreningen til Svampekundskabens Fremme, 1–6.
Posada, D. (2008). jModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 25, 1253–1256. doi: 10.1093/molbev/msn083
Rajchenberg, M., Pildain, M. B., Bianchinotti, M. V., Barroetaveña, C. (2015). The phylogenetic position of poroid hymenochaetaceae (Hymenochaetales, basidiomycota) from Patagonia, Argentina. Mycologia 107, 754–767. doi: 10.3852/14-170
Rambaut, A. (2018)Molecular evolution, phylogenetics and epidemiology. FigTree ver. 1.4.4 software (Accessed October 2022).
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D., Darling, A., Höhna, S., et al. (2012). MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61, 539–542. doi: 10.1093/sysbio/sys029
Ryvarden, L., Johansen, I. (1980). A preliminary polypore flora of East Africa. Fungiflora, 1–636. doi: 10.2307/3759822
Salvador-Montoya, C. A., Popoff, O. F., Reck, M., Drechsler-Santos, E. R. (2018). Taxonomic delimitation of Fulvifomes robiniae (Hymenochaetales, basidiomycota) and related species in America: F. squamosus sp. nov. Plant Syst. Evol. 304, 445–459. doi: 10.1007/s00606-017-1487-7
Stamatakis, A. (2014). RAxML version 8: A tool for phylogenetic analyses and post analyses of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Tchoumi, J. M. T., Coetzee, M. P. A., Rajchenberg, M., Roux, J. (2020). Poroid hymenochaetaceae associated with trees showing wood-rot symptoms in the garden route national park of south Africa. Mycologia 112, 722–741. doi: 10.1080/00275514.2020.1753160
Thompson, J. D., Gibson, T. J., Plewniak, F., Jeanmougin, F., Higgins, D. G. (1997). The clustal X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 25, 4876–4882. doi: 10.1093/nar/25.24.4876
Tomsovsky, M., Kout, J. (2013). Porodaedalea cedrina (Basidiomycota, agaricomycetes, hymenochaetaceae) - a new polypore from the Mediterranean area. Nova Hedwigia 96, 419–426. doi: 10.1127/0029-5035/2013/0087
Vilgalys, R., Hester, M. (1990). Rapid genetic identification and mapping of enzymatically amplified ribosomal DNA from several cryptococcus species. J. Bacteriol. 172, 4238–4246. doi: 10.1128/jb.172.8.4238-4246.1990
Vu, D., Groenewald, M., De, V. M., Gehrmann, T., Stielow, B., Eberhardt, U., et al. (2018). Large-Scale generation and analysis of filamentous fungal DNA barcodes boosts coverage for kingdom fungi and reveals thresholds for fungal species and higher taxon delimitation. Stud. Mycol. 92, 135–154. doi: 10.1016/j.simyco.2018.05.001
Wagner, T., Fischer, M. (2001). Natural groups and a revised system for the European poroid hymenochaetales (Basidiomycota) supported by nLSU rDNA sequence data. Mycol. Res. 105, 773–782. doi: 10.1017/S0953756201004257
Wagner, T., Fischer, M. (2002). Proceedings towards a natural classification of the worldwide taxa phellinus s.l. and inonotus s.l., and phylogenetic relationships of allied genera. Mycologia 94, 998–1016. doi: 10.1080/15572536.2003.11833156
Wagner, T., Ryvarden, L. (2002). Phylogeny and taxonomy of the genus Phylloporia (Hymenochaetales). Mycol. Prog. 1, 105–116. doi: 10.1007/s11557-006-0009-8
White, T. J., Bruns, T., Lee, S., Taylor, J. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics,” in PCR protocols: A guide to methods and applications. Eds. Innis, M. A., Gelfand, D. H., Sninsky, J. J., White, T. J. (New York, NY: Academic Press), 315–322. doi: 10.1016/B978-0-12-372180-8.50042-1
Wu, F., Chen, J. J., Ji, X. H., Vlasák, J., Dai, Y. C. (2017). Phylogeny and diversity of the morphologically similar polypore genera Rigidoporus, physisporinus, oxyporus and Leucophellinus. Mycologia 109, 749–765. doi: 10.1080/00275514.2017.1405215
Wu, F., Man, X. W., Tohtirjap, A., Dai, Y. C. (2022b). A comparison of polypore funga and species composition in forest ecosystems of China, north America, and Europe. For. Ecosyst. 9, 100051. doi: 10.1016/j.fecs.2022.100051
Wu, F., Zhou, L. W., Dai, Y. C. (2016). Neomensularia duplicata gen. et sp. nov. (Hymenochaetales, basidiomycota) and two new combinations. Mycologia 108, 891–898. doi: 10.3852/16-020
Wu, F., Zhou, L. W., Vlasák, J., Dai, Y. C. (2022a). Global diversity and systematics of hymenochaetaceae with poroid hymenophore. Fungal Divers. 113, 1–192. doi: 10.1007s13225-021-00496-4
Zhou, L. W. (2014). Fulvifomes hainanensis sp. nov. and F. indicus comb. nov. (Hymenochaetales, basidiomycota) evidenced by a combination of morphology and phylogeny. Mycoscience 55, 70–77. doi: 10.1016/j.myc.2013.05.006
Zhou, L. W. (2015a). Cylindrosporus flavidus gen. et comb. nov. (Hymenochaetales, basidiomycota) segregated from Onnia. Phytotaxa 219, 276–282. doi: 10.11646/phytotaxa.219.3.7
Zhou, L. W. (2015b). Four new species of Phylloporia (Hymenochaetales, basidiomycota) from tropical China with a key to Phylloporia species worldwide. Mycologia 107, 1184–1192. doi: 10.3852/14-254
Zhou, L. W. (2015c). Fulvifomes imbricatus and F. thailandicus (Hymenochaetales, basidiomycota): Two new species from Thailand based on morphological and molecular evidence. Mycol. Prog. 14, 1–8. doi: 10.1007/s11557-015-1116-1
Zhou, L. W., Ji, X. H., Vlasák, J., Dai, Y. C. (2018). Taxonomy and phylogeny of Pyrrhoderma: A redefinition, the segregation of Fulvoderma gen. nov. and four new species. Mycologia 110, 872–889. doi: 10.1080/00275514.2018.1474326
Zhou, L. W., Vlasák, J., Dai, Y. C. (2016a). Taxonomy and phylogeny of Phellinidium (Hymenochaetales, basidiomycota): A redefinition and the segregation of Coniferiporia gen. nov. for forest pathogens. Fungal Biol. 120, 988–1001. doi: 10.1016/j.funbio.2016.04.008
Zhou, L. W., Vlasák, J., Decock, C., Assefa, A., Stenlid, J., Abate, D., et al. (2016b). Global diversity and taxonomy of the Inonotus linteus complex (Hymenochaetales, basidiomycota): Sanghuangporus gen. nov., Tropicoporus excentrodendri and T. guanacastensis gen. et spp. nov., and 17 new combinations. Fungal Divers. 77, 335–347. doi: 10.1007/s13225-015-0335-8
Keywords: white rot, Hymenochaetaceae, polypore, taxonomy, wood-decaying fungi
Citation: Zhou M, Ji X-H, Liu H-G, Miller K, Yuan Y and Vlasák J (2023) Two new species of Hymenochaetaceae from tropical Asia and America. Front. Cell. Infect. Microbiol. 12:1100044. doi: 10.3389/fcimb.2022.1100044
Received: 16 November 2022; Accepted: 23 December 2022;
Published: 19 January 2023.
Edited by:
Jia-Jia Chen, Jiangsu Vocational College of Agriculture and Forestry, ChinaReviewed by:
Junzhi Qiu, Fujian Agriculture and Forestry University, ChinaChang-lin Zhao, Southwest Forestry University, China
Copyright © 2023 Zhou, Ji, Liu, Miller, Yuan and Vlasák. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuan Yuan, eXVhbnl1YW4xMDE4QGJqZnUuZWR1LmNu; Josef Vlasák, dmxhc2FrQHVtYnIuY2FzLmN6