 Buyun Qian
Buyun Qian Yuan Li
Yuan Li
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Cell. Infect. Microbiol., 10 November 2022
Sec. Microbiome in Health and Disease
Volume 12 - 2022 | https://doi.org/10.3389/fcimb.2022.1059349
This article is part of the Research TopicRelationship Between Intestinal Microbiome and VasculitisView all 6 articles
In recent years, due to the development and widespread utilization of metagenomic sequencing and metabolomics, the relationship between gut microbiota and human cardiovascular diseases (CVDs) has received extensive attention. A growing number of studies have shown a strong relationship between gut microbiota and CVDs, such as coronary atherosclerosis, hypertension (HTN) and heart failure (HF). It has also been revealed that intestinal flora-related metabolites, such as trimethylamine-N-oxide (TMAO), short-chain fatty acids (SCFA) and bile acids (BAs), are also related to the development, prevention, treatment and prognosis of CVDs. In this review, we presented and summarized the recent findings on the relationship between gut microbiota and CVDs, and concluded several currently known gut microbiota-related metabolites and the occurrence and development of CVDs.
Cardiovascular diseases (CVDs), including coronary atherosclerosis, hypertension (HTN) and heart failure (HF), are important causes of death, resulting in a huge economic and health burden globally (Zoungas et al., 2014; Writing Group et al., 2016). As a chronic disease, in addition to inflammation, diabetes will affect the occurrence and development of CVDs (Haybar et al., 2019). Other factors such as diet and nutritional status also have a more profound effect on CVDs (Brown and Hazen, 2018). Moreover, microbial sequencing analysis has provided a wealth of information about the presence of characteristic gut microbiota associated with CVDs (Jäckel et al., 2017; Kiouptsi and Reinhardt, 2018; Lindskog Jonsson et al., 2018; Haghikia et al., 2022). Therefore, more and more studies have shown that the gut microbiota has a strong link with CVDs.
Human gut is a huge microbial habitat. The adult gut contains hundreds of species of bacteria. Gut flora plays an important role in maintaining body health (Wampach et al., 2017; Wu et al., 2020b; Li and Chen, 2022). Gut microbiota can produce certain biologically active metabolites in the gut. These gut microbiota-related metabolites function in various aspects of host physiology and are known as the ninth system of the human body (O’Hara and Shanahan, 2006; Qin et al., 2010). Intestinal flora can form the intestinal epithelial barrier, regulate immune function, digest nutrients, produce vitamins and prevent the invasion of pathogenic bacteria, which is essential for human health (DeGruttola et al., 2016; Thomas and Jobin, 2020; Wu et al., 2020a; Chen et al., 2021; Cui et al., 2021). When dietary habits, environmental factors, intestinal infections and other factors lead to changes in the human gut microbiome, it can lead to intestinal malnutrition, trigger inflammation and abnormal metabolism and then lead to the occurrence and development of CVDs (Yatsunenko et al., 2012; Naghavi et al., 2015). In this review, we introduced the role of gut microbiota in CVDs and summarized the related metabolites, which may provide new insight into the association between gut microbiota and CVDs (Table 1).

Table 1 The association between gut microbiota and CVDs.
Coronary heart disease (CHD) is based on coronary atherosclerosis and is an important representative of metabolic CVDs. The metagenomic sequencing showed that gut microbiota in patients with atherosclerotic cardiovascular disease differed from healthy individuals, with higher levels of Streptococcus and Enterobacteriaceae (Ott et al., 2006; Jie et al., 2017). It has been reported that co-abundant gut microbiota and serum metabolites are closely related to CHD severity, and gut microbiota such as Roseburia, Ruminococcaceae and Clostridium may regulate the metabolic activity of bile acids (BAs) and aromatic compounds, which will further affect the progression of coronary atherosclerosis (Liu et al., 2019a). It was revealed that gut microbiota and coronary atherosclerosis are correlated (Table 1).
Intestinal dysbiosis can also exert pro-atherosclerotic effects through metabolism-dependent pathways by altering the production of various metabolites, including Trimethylamine-N-oxide (TAMO), BAs, serum indoxylate, protocatechuic acid and lipopolysaccharide (LPS). TMAO is one of the most important metabolites associated with gut microbiota. Studies have shown that the body’s immune system regulation, cholesterol metabolism, oxidative stress and inflammatory responses will all be affected by TMAO to a certain extent, thereby increasing the risk of coronary atherosclerosis. Therefore, increased plasma TMAO concentration can increase the possibility of CVDs (Seldin et al., 2016; Geng et al., 2018; Xie et al., 2021). Wang et al. found that TMAO-dependent upregulation of macrophage scavenger receptor and CD36 expression impaired cholesterol metabolism in macrophages, thereby promoting foam cell production, one of the earliest cellular signals in the progression of coronary atherosclerosis (Wang et al., 2011). Studies have found that elevated levels of TMAO can induce activation of the NF-κB pathway and promote the release of inflammatory cytokines IL-18 and IL-1β, indicating that inflammatory mediators play a role in TMAO-induced endothelial dysfunction (Liu et al., 2019c). Zhu et al. found that TMAO could increase intracellular rectal calcium release from platelets, leading to platelet aggregation and thrombosis (Zhu et al., 2016). This study found that the choline analog 3,3-dimethylbutanol (DMB) has an inhibitory effect on choline TMA lyase activity and can reduce circulating TMAO, thereby attenuating the promoting effect of choline in coronary atherosclerosis (Wang et al., 2015).
TMAO plays an important role in the development of coronary atherosclerosis. Furthermore, coronary atherosclerosis is also affected by cholesterol metabolism, short-chain fatty acids and tryptophan metabolites (Paeslack et al., 2022; Roessler et al., 2022). It has been shown that deconjugated BAs are hydrophobic and can be excreted in feces, which will in turn lower circulating cholesterol and thus reduce the risk of coronary atherosclerosis (Dawson and Karpen, 2015). It was also found that serum indoxylate levels are positively correlated with coronary atherosclerosis and are predictive mechanistic biomarkers of coronary artery disease severity (Hsu et al., 2013). Additionally, the expression of atherosclerosis-related genes is also regulated by protocatechuic acid, including oxidative stress-related AOX1, CYP2E1 or TXNIP, adhesion molecule JAM-A, angiogenesis-related blood vessels endothelial growth factor receptor 2 and so on (Mauray et al., 2012). Moreover, LPS was reported to induce foam cell formation and cholesteryl ester accumulation from native low-density lipoprotein, suggesting that LPS has a pro-atherosclerotic effect (Lakio et al., 2006).
Gut microbiota metabolites are also involved in arterial thrombosis. It was revealed that fecal transplantation of TMAO-rich gut microbiota into germ-free mice can promote platelet function and arterial thrombosis (Huynh, 2020). Nemet et al. found that phenylacetylglutamine can induce platelet hyperresponsiveness through adrenergic receptors (Nemet et al., 2020). Besides, phytoestrogens may also have some prothrombotic or proinflammatory effects (Herrington, 2000).
HTN is the most common risk factor associated with CVDs, and as the main risk factor for stroke and CHD morbidity and mortality (Page, 1967), it has always been a hot topic. Recently, studies have shown that the gut microbiota is involved in blood pressure regulation and that abnormal bacterial populations are associated with HTN (Li et al., 2017; Wilck et al., 2017). Compared with healthy individuals, the abundance and diversity of gut microbes in hypertensive patients decreased, and the genus Prevotella was significantly increased (Li et al., 2017). In addition, a fecal microbiota transplantation (FMT) study confirmed that the fecal microbiota of patients with HTN can increase the blood pressure in germ-free mice, revealing a close link between gut microbiota and the regulation of blood pressure (Mell et al., 2015). Hence, there exists a link between gut microbiota and HTN (Table 1).
In addition to changes in gut microbiota composition, excessive formation of gut microbiota metabolites is also considered to be a key factor in the occurrence of HTN. Bacteria belonging to the genera Bifidobacterium, Lactobacillus, Streptococcus and Escherichia coli can produce neurotransmitters within the autonomic nervous system that will alter vascular tone, leading to HTN (Karbach et al., 2016). It was also shown that the higher levels of circulating TMAO are positively associated with the high risk of blood pressure (Ge et al., 2020). Liu et al. found that the use of Lactobacillus rhamnosus GG strain can prevent HTN deterioration by reducing the levels of TMAO (Liu et al., 2019b). Besides, short-chain fatty acids (SCFAs) are also of great significance in the regulation of blood pressure and are mainly involved in the regulation of blood pressure mainly through Olfactory receptor 78 (Olfr78) and G protein-coupled receptor 41 (GPR41) (Felizardo et al., 2019). Natarajan et al. found that GPR41 knockout mice exhibited HTN (Natarajan et al., 2016). The activation of GPR41 was found to reduce SCFA-producing bacteria in some patients with hypertensive gut dysregulation (Pluznick, 2014). Meanwhile, Miyamoto et al. showed that Olfr78-deficient mice also exhibited high blood pressure (Miyamoto et al., 2016). Furthermore, studies also have shown that the production of propionate depends on regulatory T cells, and blood pressure can be reduced through regulatory T cells 17 and angiotensin II-induced effectors (Bartolomaeus et al., 2019). Although there exist many studies on the mechanism how gut microbiota influence HTN, the specific mechanism remains unclear and more studies are needed. The regulation of blood pressure by intestinal flora metabolites such as TMAO, SCFA and propionate will provide a new idea for drugs to improve intestinal flora to treat HTN.
HF is an irreversible end-stage disease with high mortality, characterized by edema and dyspnea (Ponikowski et al., 2016). The gut hypothesis suggested that reduced cardiac output and increased systemic congestion can cause ischemia and edema of the intestinal mucosa, leading to increased bacterial translocation and increased circulating endotoxin, leading to HF (Krack et al., 2004; Sandek et al., 2007). Studies have found that patients with HF presented the increased levels of pathogenic bacteria such as Candida and the decreased levels of anti-inflammatory bacteria such as Faecalibacterium, therefore contributing to the development of HF by participating in the regulation of mucosal immune (Pasini et al., 2016). This indicated that there exists a correlation between gut microbiota and HF (Table 1).
Gut microbiota metabolites such as SCFAs, TMAO, indoxyl sulfate and LPS also play an important role in the development of HF. Savi et al. demonstrated that TMAO can promote calcium release in healthy mouse cardiomyocytes, thereby altering their contractility (Zabell and Tang, 2017; Savi et al., 2018). The direct dietary TMAO supplementation can lead to higher systemic TMAO levels, increase myocardial fibrosis and induce HF (Organ et al., 2016). It was revealed that TMAO has deleterious effects on adult cardiomyocytes by inducing T-tubule network damage and calcium-handling dysfunction (Jin et al., 2020). TMAO can promote myocardial fibrosis by activating NLRP3 inflammasome-related signaling, suggesting that TMAO may be a potential target for the treatment of HF (Siu et al., 2019). Schuett et al. demonstrated that TMAO increases patient susceptibility to HF by increasing myocardial fibrosis (Schuett et al., 2017). Similarly, Wang et al. found that 3,3-dimethyl-1-butanol (DMB) ameliorated adverse cardiac structural remodeling in overload-induced HF mice by downregulating TMAO levels (Wang et al., 2020). Indoxyl sulfate exacerbates cardiac fibrosis, cardiomyocyte hypertrophy, and atrial fibrillation (Yisireyili et al., 2013; Aoki et al., 2015). Besides, Mayerhofer et al. found that BAs can play a role in the cardiovascular function by reducing heart rate by modulating channel conductance and calcium dynamics in atrial and ventricular cardiomyocytes, as well as modulating vascular tone (Mayerhofer et al., 2017). SCFAs have gut protective effects. Decreased SCFAs will lead to disruption of the intestinal barrier, facilitating translocation of endotoxins into the blood circulation, ultimately leading to HF (Tang et al., 2012; Nagatomo and Tang, 2015). SCFAs can also promote cardiac repair after HF by inducing CX3CR1+ cells (Tang et al., 2019). Furthermore, LPS can damage the mucosal barrier function of the intestine, increase intestinal permeability, thereby increasing inflammatory cytokines, which is closely related to the occurrence of HF (Lam et al., 2012b).
There existed several microorganism-targeted therapies used in CVDs (Table 2). FMT, which refers to the replacement of enteric pathogens by introducing the fecal contents of healthy subjects into the gastrointestinal tract of patients, is an effective method to directly introduce the gut microbiota (Colman and Rubin, 2014; Zhang et al., 2019). Studies have shown that FMT can eliminate the increased Bacteroides/Firmicutes ratio and reduce inflammation in cardiomyocytes, thereby reducing myocarditis in mice (Kim et al., 2018). In addition, clinical studies have shown that FMT can quickly restore the gut microbiota of healthy people after the use of antibiotics (Taur et al., 2018).However, the use of FMT is currently limited due to the transfer of endotoxins or infectious agents that may lead to new gastrointestinal complications (Brandt, 2013; De Leon et al., 2013).
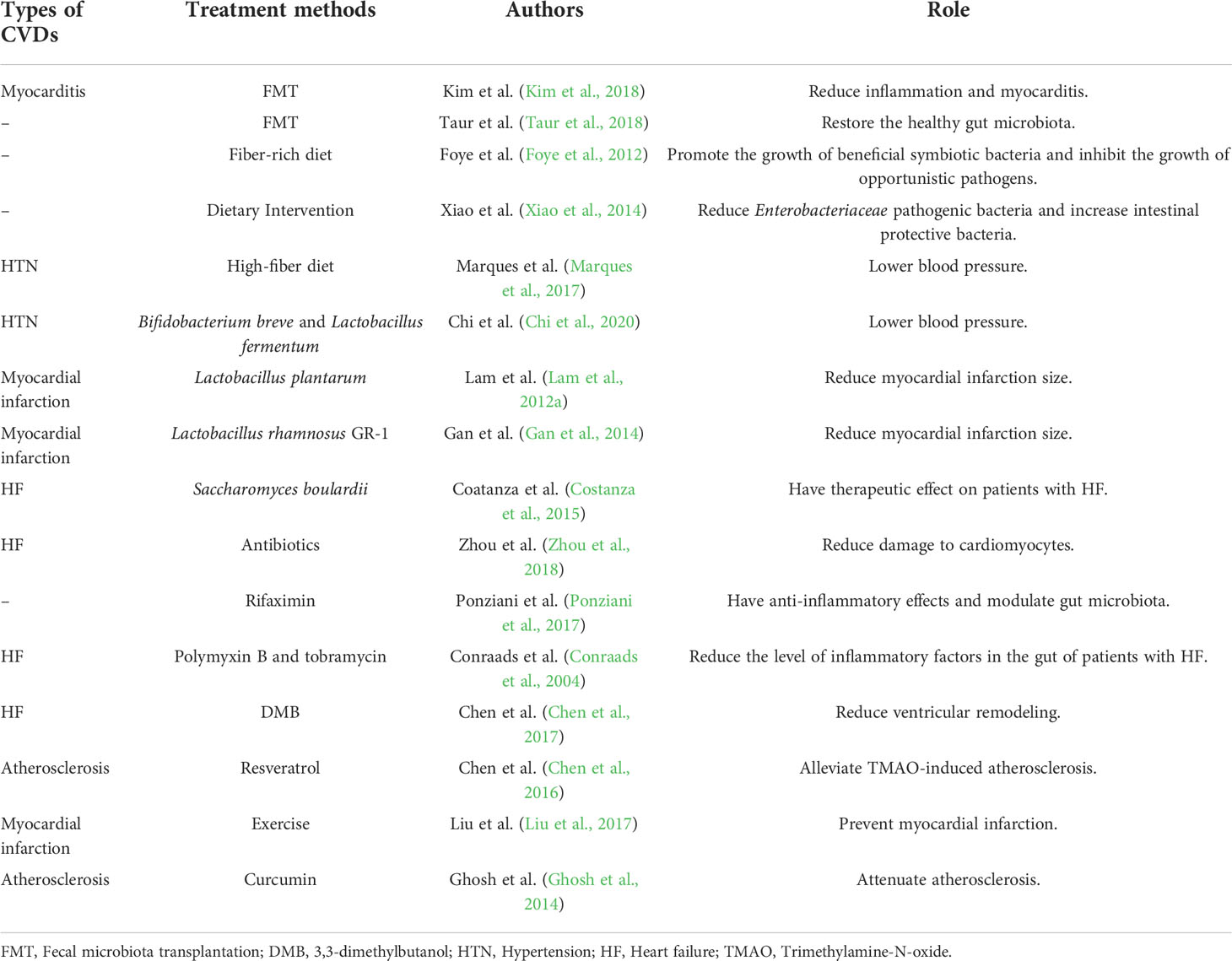
Table 2 Microorganism-targeted therapies.
Dietary intervention to regulate the treatment of CVDs has broad prospects (Appel et al., 1997; Estruch et al., 2013). Studies have shown fiber-rich diet can promote the growth of beneficial symbiotic bacteria and inhibit the growth of opportunistic pathogens (Foye et al., 2012). Xiao et al. found that dietary intervention with whole grains and traditional Chinese medicine foods can reduce Enterobacteriaceae pathogenic bacteria and increase intestinal protective bacteria such as Bifidobacterium (Xiao et al., 2014). In addition, a high-fiber diet can increase the acetic acid-producing microbiota, which in turn lowers blood pressure (Marques et al., 2017).
Among the numerous bacteria in the host gut, some are beneficial, and additional enhancement of these bacteria may lead to positive outcomes, resulting in the use of probiotics (Ojetti et al., 2009). It was found that the probiotics Bifidobacterium breve and Lactobacillus fermentum can have antihypertensive effects by restoring gut microbiota balance and preventing endothelial dysfunction (Chi et al., 2020). Lam et al. found that Lactobacillus plantarum could improve ventricular function and reduce myocardial infarction size (Lam et al., 2012a). In addition, similar results were obtained by treating myocardial ischemia rats with Lactobacillus rhamnosus GR-1 (Gan et al., 2014). Saccharomyces boulardii can also reduce the level of inflammatory markers and serum creatinine, which has therapeutic effect on patients with HF (Costanza et al., 2015). Although probiotics are safe, there is a lack of supervision, which may increase the risk of probiotics transferring to blood and lead to sepsis (Kochan et al., 2011).
Antibiotics affect the structure of the gut microbiota, which in turn treats CVDs. Study shows that antibiotic injections can eliminate shifts in gut microbiota, reduce damage to cardiomyocytes (Zhou et al., 2018). Several studies have found that antibiotics can reduce inflammation, such as rifaximin, which has anti-inflammatory effects and modulates gut microbiota (Ponziani et al., 2017), and polymyxin B and tobramycin can reduce the level of inflammatory factors in the gut of patients with HF (Conraads et al., 2004), which has important implications for the treatment of CVDs.
DMB treatment can reduce TMA production, limit the conversion of TMA to TMAO, and reduce ventricular remodeling (Chen et al., 2017). Furthermore, resveratrol from Polygonum cuspidatum can alleviate TMAO-induced atherosclerosis by remodeling the microbiota and reducing TMAO levels (Chen et al., 2016).
Besides, exercise can boost Firmicutes to Bacteroides ratio (Petriz et al., 2014; Lambert et al., 2015), increase levels of bacterial metabolites (Allen et al., 2018) and prevent myocardial infarction (Liu et al., 2017). But the effects of exercise on the gut microbiome are transient and reversible (Lira et al., 2010).
There are also other treatments, which can be used for CVDs. Ghosh et al. found that curcumin attenuates atherosclerosis by modulating intestinal barrier function (Ghosh et al., 2014). Additionally, berberine, derived from the Chinese herb Coptis chinensis, can modulate the gut microbiota, which in turn affects CVDs (Wang et al., 2019). In summary, Microorganism-targeted therapy mainly regulates CVDs through FMT, dietary interventions, and probiotics.
A large number of research results show that the gut microbiota is involved in the occurrence and development of CHD, HTN and HF, and plays an important role in it. The gut microbiota influence CVDs through immune regulation, the inflammatory response, gut barrier integrity, metabolic homeostasis. CVDs, in turn, also affect the structure and function of the gut microbiota (Figure 1). In addition to traditional culture methods, more advanced technologies such as metagenomics and metabolomics have become important means to study the human gut microbiota. Numerous studies have observed the relationship between gut microbiota and CVDs and proposed various potential mechanisms of action, especially mechanisms such as metabolic pathways. A better understanding of the human gut microbiota can provide more potential treatments for patients with CVDs on the basis of current clinical traditional drug treatments, such as dietary adjustments, rational use of probiotics and antibiotics, and even FMT, etc., bringing more possibilities for the prevention and treatment of CVDs. At present, most studies on the correlation between gut microbiota and CVDs are based on animal experiments and the mechanism between gut microbiota and CVDs is still not fully understood. Although TMAO is a potential biomarker for CVDs development, other gut microbiota or related metabolites should be explored as early CVDs markers. In addition, exploring the immune mechanisms of CVDs also helps us analyze how gut microbiota metabolites interfere with disease at the molecular level. More experiments are needed to explore the link between gut microbiota and CVDs, and further clinical studies are needed. Some approaches based on gut microbiota for the treatment of CVDs are still in clinical trials and have potential advantages as well as limitations. Therapeutic strategies to improve the gut microbiota are potential avenues for the treatment of CVDs.
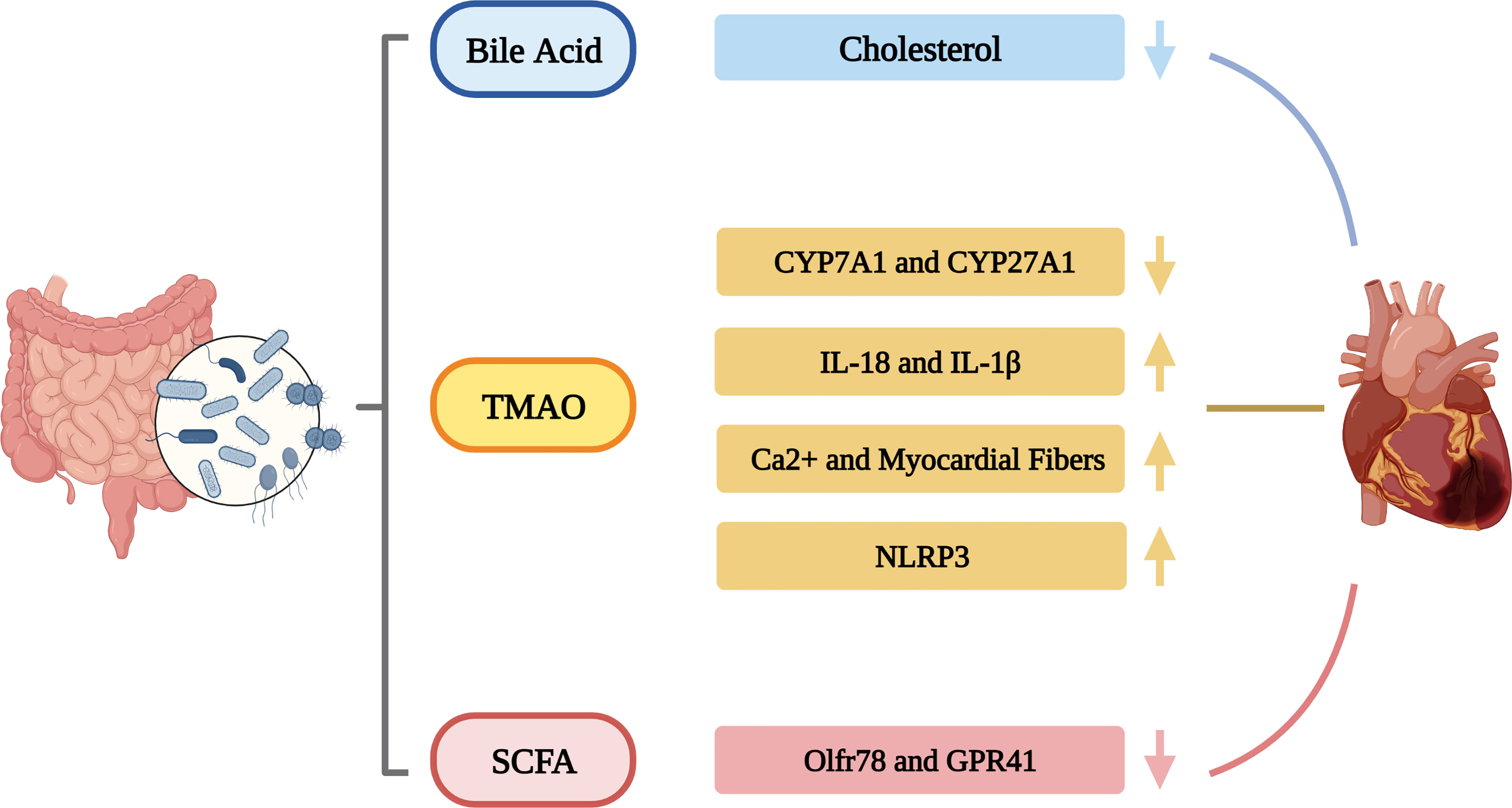
Figure 1 The roles of gut microbiota in CVDs.
BQ had the idea for the article. KZ performed the literature search and data analysis. YL and KS drafted and critically revised the work. All authors contributed to the article and approved the submitted version.
This study was supported by Nanjing Medical University Gusu College Scientific Research Fund (NO. GSKY20210202).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
CVDs, Cardiovascular diseases; TMAO, Trimethylamine-N-oxide; SCFAs, Short-chain fatty acids; BAs, Bile acids; CHD, Coronary atherosclerotic heart disease; DMB, 3,3-dimethylbutanol; LPS, Lipopolysaccharide; HTN, Hypertension; FMT, Fecal microbiota transplantation; HF, Heart failure
Allen, J. M., Mailing, L. J., Niemiro, G. M., Moore, R., Cook, M. D., White, B. A., et al. (2018). Exercise alters gut microbiota composition and function in lean and obese humans. Med. Sci. Sports Exerc. 50 (4), 747–757. doi: 10.1249/MSS.0000000000001495
Aoki, K., Teshima, Y., Kondo, H., Saito, S., Fukui, A., Fukunaga, N., et al. (2015). Role of indoxyl sulfate as a predisposing factor for atrial fibrillation in renal dysfunction. J. Am. Heart Assoc. 4 (10), e002023. doi: 10.1161/JAHA.115.002023
Appel, L. J., Moore, T. J., Obarzanek, E., Vollmer, W. M., Svetkey, L. P., Sacks, F. M., et al. (1997). A clinical trial of the effects of dietary patterns on blood pressure. DASH collaborative research group. N Engl. J. Med. 336 (16), 1117–1124. doi: 10.1056/NEJM199704173361601
Bartolomaeus, H., Balogh, A., Yakoub, M., Homann, S., Marko, L., Hoges, S., et al. (2019). Short-chain fatty acid propionate protects from hypertensive cardiovascular damage. Circulation 139 (11), 1407–1421. doi: 10.1161/Circulationaha.118.036652
Brandt, L. J. (2013). FMT: first step in a long journey. Am. J. Gastroenterol. 108 (8), 1367–1368. doi: 10.1038/ajg.2013.165
Brown, J. M., Hazen, S. L. (2018). Microbial modulation of cardiovascular disease. Nat. Rev. Microbiol. 16 (3), 171–181. doi: 10.1038/nrmicro.2017.149
Chen, Z.-R., Jin, S.-F., Ma, W.-B., Jiang, R.-L. (2021). Intestinal microecology: a crucial strategy for targeted therapy of liver diseases. Hepatobiliary Pancreat. Dis. Int. 20 (5), 499–500. doi: 10.1016/j.hbpd.2021.07.007
Chen, M. L., Yi, L., Zhang, Y., Zhou, X., Ran, L., Yang, J. N., et al. (2016). Resveratrol attenuates trimethylamine-N-Oxide (TMAO)-induced atherosclerosis by regulating TMAO synthesis and bile acid metabolism via remodeling of the gut microbiota. Mbio 7 (2). doi: 10.1128/mBio.02210-15
Chen, K., Zheng, X., Feng, M., Li, D., Zhang, H. (2017). Gut microbiota-dependent metabolite trimethylamine n-oxide contributes to cardiac dysfunction in Western diet-induced obese mice. Front. Physiol. 8. doi: 10.3389/fphys.2017.00139
Chi, C., Li, C., Wu, D., Buys, N., Wang, W., Fan, H., et al. (2020). Effects of probiotics on patients with hypertension: a systematic review and meta-analysis. Curr. Hypertens. Rep. 22 (5), 34. doi: 10.1007/s11906-020-01042-4
Colman, R. J., Rubin, D. T. (2014). Fecal microbiota transplantation as therapy for inflammatory bowel disease: a systematic review and meta-analysis. J. Crohns Colitis 8 (12), 1569–1581. doi: 10.1016/j.crohns.2014.08.006
Conraads, V. M., Jorens, P. G., De Clerck, L. S., Van Saene, H. K., Ieven, M. M., Bosmans, J. M., et al. (2004). Selective intestinal decontamination in advanced chronic heart failure: a pilot trial. Eur. J. Heart Fail 6 (4), 483–491. doi: 10.1016/j.ejheart.2003.12.004
Costanza, A. C., Moscavitch, S. D., Faria Neto, H. C., Mesquita, E. T. (2015). Probiotic therapy with saccharomyces boulardii for heart failure patients: a randomized, double-blind, placebo-controlled pilot trial. Int. J. Cardiol. 179, 348–350. doi: 10.1016/j.ijcard.2014.11.034
Cui, D., Tang, Y., Jiang, Q., Jiang, D., Zhang, Y., Lv, Y., et al. (2021). Follicular helper T cells in the immunopathogenesis of SARS-CoV-2 infection. Front. Immunol. 3806. doi: 10.3389/fimmu.2021.731100
Dawson, P. A., Karpen, S. J. (2015). Intestinal transport and metabolism of bile acids. J. Lipid Res. 56 (6), 1085–1099. doi: 10.1194/jlr.R054114
DeGruttola, A. K., Low, D., Mizoguchi, A., Mizoguchi, E. (2016). Current understanding of dysbiosis in disease in human and animal models. Inflammation Bowel Dis. 22 (5), 1137–1150. doi: 10.1097/MIB.0000000000000750
De Leon, L. M., Watson, J. B., Kelly, C. R. (2013). Transient flare of ulcerative colitis after fecal microbiota transplantation for recurrent clostridium difficile infection. Clin. Gastroenterol. Hepatol. 11 (8), 1036–1038. doi: 10.1016/j.cgh.2013.04.045
Estruch, R., Ros, E., Salas-Salvado, J., Covas, M. I., Corella, D., Aros, F., et al. (2013). Primary prevention of cardiovascular disease with a Mediterranean diet. N Engl. J. Med. 368 (14), 1279–1290. doi: 10.1056/NEJMoa1200303
Felizardo, R. J. F., Watanabe, I. K. M., Dardi, P., Rossoni, L. V., Camara, N. O. S. (2019). The interplay among gut microbiota, hypertension and kidney diseases: The role of short-chain fatty acids. Pharmacol. Res. 141, 366–377. doi: 10.1016/j.phrs.2019.01.019
Foye, O. T., Huang, I. F., Chiou, C. C., Walker, W. A., Shi, H. N. (2012). Early administration of probiotic lactobacillus acidophilus and/or prebiotic inulin attenuates pathogen-mediated intestinal inflammation and smad 7 cell signaling. FEMS Immunol. Med. Microbiol. 65 (3), 467–480. doi: 10.1111/j.1574-695X.2012.00978.x
Gan, X. T., Ettinger, G., Huang, C. X., Burton, J. P., Haist, J. V., Rajapurohitam, V., et al. (2014). Probiotic administration attenuates myocardial hypertrophy and heart failure after myocardial infarction in the rat. Circ. Heart Fail 7 (3), 491–499. doi: 10.1161/CIRCHEARTFAILURE.113.000978
Geng, J., Yang, C., Wang, B., Zhang, X., Hu, T., Gu, Y., et al. (2018). Trimethylamine n-oxide promotes atherosclerosis via CD36-dependent MAPK/JNK pathway. BioMed. Pharmacother. 97, 941–947. doi: 10.1016/j.biopha.2017.11.016
Ge, X., Zheng, L., Zhuang, R., Yu, P., Xu, Z., Liu, G., et al. (2020). The gut microbial metabolite trimethylamine n-oxide and hypertension risk: A systematic review and dose-response meta-analysis. Adv. Nutr. 11 (1), 66–76. doi: 10.1093/advances/nmz064
Ghosh, S. S., Bie, J., Wang, J., Ghosh, S. (2014). Oral supplementation with non-absorbable antibiotics or curcumin attenuates western diet-induced atherosclerosis and glucose intolerance in LDLR-/- mice–role of intestinal permeability and macrophage activation. PloS One 9 (9), e108577. doi: 10.1371/journal.pone.0108577
Haghikia, A., Zimmermann, F., Schumann, P., Jasina, A., Roessler, J., Schmidt, D., et al. (2022). Propionate attenuates atherosclerosis by immune-dependent regulation of intestinal cholesterol metabolism. Eur. Heart J. 43 (6), 518–533. doi: 10.1093/eurheartj/ehab644
Haybar, H., Shokuhian, M., Bagheri, M., Davari, N., Saki, N. (2019). Involvement of circulating inflammatory factors in prognosis and risk of cardiovascular disease. J. Mol. Cell Cardiol. 132, 110–119. doi: 10.1016/j.yjmcc.2019.05.010
Herrington, D. (2000). Role of estrogens, selective estrogen receptor modulators and phytoestrogens in cardiovascular protection. Can. J. Cardiol. 16 (Suppl E), 5E–9E.
Hsu, C. C., Lu, Y. C., Chiu, C. A., Yu, T. H., Hung, W. C., Wang, C. P., et al. (2013). Levels of indoxyl sulfate are associated with severity of coronary atherosclerosis. Clin. Invest. Med. 36 (1), E42–E49. doi: 10.25011/cim.v36i1.19404
Huynh, K. (2020). Novel gut microbiota-derived metabolite promotes platelet thrombosis via adrenergic receptor signalling. Nat. Rev. Cardiol. 17 (5), 265–265. doi: 10.1038/s41569-020-0367-y
Jäckel, S., Kiouptsi, K., Lillich, M., Hendrikx, T., Khandagale, A., Kollar, B., et al. (2017). Gut microbiota regulate hepatic von willebrand factor synthesis and arterial thrombus formation via toll-like receptor-2. Blood J. Am. Soc. Hematol. 130 (4), 542–553. doi: 10.1182/blood-2016-11-754416
Jie, Z. Y., Xia, H. H., Zhong, S. L., Feng, Q., Li, S. H., Liang, S. S., et al. (2017). The gut microbiome in atherosclerotic cardiovascular disease. Nat. Commun. 8, 845. doi: 10.1038/s41467-017-00900-1
Jin, B., Ji, F., Zuo, A., Liu, H., Qi, L., He, Y., et al. (2020). Destructive role of TMAO in T-tubule and excitation-contraction coupling in the adult cardiomyocytes. Int. Heart J. 61 (2), 355–363. doi: 10.1536/ihj.19-372
Karbach, S. H., Schonfelder, T., Brandao, I., Wilms, E., Hormann, N., Jackel, S., et al. (2016). Gut microbiota promote angiotensin II-induced arterial hypertension and vascular dysfunction. J. Am. Heart Assoc. 5 (9). doi: 10.1161/JAHA.116.003698
Kim, T. T., Parajuli, N., Sung, M. M., Bairwa, S. C., Levasseur, J., Soltys, C. M., et al. (2018). Fecal transplant from resveratrol-fed donors improves glycaemia and cardiovascular features of the metabolic syndrome in mice. Am. J. Physiol. Endocrinol. Metab. 315 (4), E511–E519. doi: 10.1152/ajpendo.00471.2017
Kiouptsi, K., Reinhardt, C. (2018). Contribution of the commensal microbiota to atherosclerosis and arterial thrombosis. Br. J. Pharmacol. 175 (24), 4439–4449. doi: 10.1111/bph.14483
Kochan, P., Chmielarczyk, A., Szymaniak, L., Brykczynski, M., Galant, K., Zych, A., et al. (2011). Lactobacillus rhamnosus administration causes sepsis in a cardiosurgical patient–is the time right to revise probiotic safety guidelines? Clin. Microbiol. Infect. 17 (10), 1589–1592. doi: 10.1111/j.1469-0691.2011.03614.x
Krack, A., Richartz, B. M., Gastmann, A., Greim, K., Lotze, U., Anker, S. D., et al. (2004). Studies on intragastric PCO2 at rest and during exercise as a marker of intestinal perfusion in patients with chronic heart failure. Eur. J. Heart Fail 6 (4), 403–407. doi: 10.1016/j.ejheart.2004.03.002
Lakio, L., Lehto, M., Tuomainen, A. M., Jauhiainen, M., Malle, E., Asikainen, S., et al. (2006). Pro-atherogenic properties of lipopolysaccharide from the periodontal pathogen actinobacillus actinomycetemcomitans. J. Endotoxin Res. 12 (1), 57–64. doi: 10.1179/096805106X89099
Lambert, J. E., Myslicki, J. P., Bomhof, M. R., Belke, D. D., Shearer, J., Reimer, R. A. (2015). Exercise training modifies gut microbiota in normal and diabetic mice. Appl. Physiol. Nutr. Metab. 40 (7), 749–752. doi: 10.1139/apnm-2014-0452
Lam, Y. Y., Ha, C. W., Campbell, C. R., Mitchell, A. J., Dinudom, A., Oscarsson, J., et al. (2012b). Increased gut permeability and microbiota change associate with mesenteric fat inflammation and metabolic dysfunction in diet-induced obese mice. PloS One 7 (3), e34233. doi: 10.1371/journal.pone.0034233
Lam, V., Su, J., Koprowski, S., Hsu, A., Tweddell, J. S., Rafiee, P., et al. (2012a). Intestinal microbiota determine severity of myocardial infarction in rats. FASEB J. 26 (4), 1727–1735. doi: 10.1096/fj.11-197921
Li, W., Chen, T. (2022). An insight into the clinical application of gut microbiota during anticancer therapy. Adv. Gut Microbiome Res. 2022. doi: 10.1155/2022/8183993
Lindskog Jonsson, A., Caesar, R., Akrami, R., Reinhardt, C., Fåk Hållenius, F., Borén, J., et al. (2018). Impact of gut microbiota and diet on the development of atherosclerosis in apoe–/– mice. Arterioscler. Thromb. Vasc. Biol. 38 (10), 2318–2326. doi: 10.1161/ATVBAHA.118.311233
Lira, F. S., Rosa, J. C., Pimentel, G. D., Souza, H. A., Caperuto, E. C., Carnevali, L. C., Jr., et al. (2010). Endotoxin levels correlate positively with a sedentary lifestyle and negatively with highly trained subjects. Lipids Health Dis. 9, 82. doi: 10.1186/1476-511X-9-82
Liu, H., Chen, X., Hu, X., Niu, H., Tian, R., Wang, H., et al. (2019a). Alterations in the gut microbiome and metabolism with coronary artery disease severity. Microbiome 7 (1), 68. doi: 10.1186/s40168-019-0683-9
Liu, M., Han, Q., Yang, J. (2019c). Trimethylamine-n-oxide (TMAO) increased aquaporin-2 expression in spontaneously hypertensive rats. Clin. Exp. Hypertens. 41 (4), 312–322. doi: 10.1080/10641963.2018.1481420
Liu, Z., Liu, H. Y., Zhou, H., Zhan, Q., Lai, W., Zeng, Q., et al. (2017). Moderate-intensity exercise affects gut microbiome composition and influences cardiac function in myocardial infarction mice. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.01687
Liu, J., Li, T., Wu, H., Shi, H., Bai, J., Zhao, W., et al. (2019b). Lactobacillus rhamnosus GG strain mitigated the development of obstructive sleep apnea-induced hypertension in a high salt diet via regulating TMAO level and CD4(+) T cell induced-type I inflammation. BioMed. Pharmacother. 112, 108580. doi: 10.1016/j.biopha.2019.01.041
Li, J., Zhao, F., Wang, Y., Chen, J., Tao, J., Tian, G., et al. (2017). Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 5 (1), 14. doi: 10.1186/s40168-016-0222-x
Marques, F. Z., Nelson, E., Chu, P. Y., Horlock, D., Fiedler, A., Ziemann, M., et al. (2017). High-fiber diet and acetate supplementation change the gut microbiota and prevent the development of hypertension and heart failure in hypertensive mice. Circulation 135 (10), 964–96+. doi: 10.1161/CIRCULATIONAHA.116.024545
Mauray, A., Felgines, C., Morand, C., Mazur, A., Scalbert, A., Milenkovic, D. (2012). Bilberry anthocyanin-rich extract alters expression of genes related to atherosclerosis development in aorta of apo e-deficient mice. Nutr. Metab. Cardiovasc. Dis. 22 (1), 72–80. doi: 10.1016/j.numecd.2010.04.011
Mayerhofer, C. C. K., Ueland, T., Broch, K., Vincent, R. P., Cross, G. F., Dahl, C. P., et al. (2017). Increased Secondary/Primary bile acid ratio in chronic heart failure. J. Card. Fail. 23 (9), 666–671. doi: 10.1016/j.cardfail.2017.06.007
Mell, B., Jala, V. R., Mathew, A. V., Byun, J., Waghulde, H., Zhang, Y. J., et al. (2015). Evidence for a link between gut microbiota and hypertension in the Dahl rat. Physiol. Genomics 47 (6), 187–197. doi: 10.1152/physiolgenomics.00136.2014
Miyamoto, J., Kasubuchi, M., Nakajima, A., Irie, J., Itoh, H., Kimura, I. (2016). The role of short-chain fatty acid on blood pressure regulation. Curr. Opin. Nephrol. Hypertens. 25 (5), 379–383. doi: 10.1097/MNH.0000000000000246
Nagatomo, Y., Tang, W. H. W. (2015). Intersections between microbiome and heart failure: Revisiting the gut hypothesis. J. Card. Fail. 21 (12), 973–980. doi: 10.1016/j.cardfail.2015.09.017
Naghavi, M., Wang, H. D., Lozano, R., Davis, A., Liang, X. F., Zhou, M. G., et al. (2015). Global, regional, and national age-sex specific all-cause and cause-specific mortality for 240 causes of death 1990-2013: a systematic analysis for the global burden of disease study 2013. Lancet 385 (9963), 117–171. doi: 10.1016/S0140-6736(14)61682-2
Natarajan, N., Hori, D., Flavahan, S., Steppan, J., Flavahan, N. A., Berkowitz, D. E., et al. (2016). Microbial short chain fatty acid metabolites lower blood pressure via endothelial G protein-coupled receptor 41. Physiol. Genomics 48 (11), 826–834. doi: 10.1152/physiolgenomics.00089.2016
Nemet, I., Saha, P. P., Gupta, N., Zhu, W., Romano, K. A., Skye, S. M., et al. (2020). A cardiovascular disease-linked gut microbial metabolite acts via adrenergic receptors. Cell 180 (5), 862–877.e822. doi: 10.1016/j.cell.2020.02.016
O’Hara, A. M., Shanahan, F. (2006). The gut flora as a forgotten organ. EMBO Rep. 7 (7), 688–693. doi: 10.1038/sj.embor.7400731
Ojetti, V., Lauritano, E. C., Barbaro, F., Migneco, A., Ainora, M. E., Fontana, L., et al. (2009). Rifaximin pharmacology and clinical implications. Expert Opin. Drug Metab. Toxicol. 5 (6), 675–682. doi: 10.1517/17425250902973695
Organ, C. L., Otsuka, H., Bhushan, S., Wang, Z. N., Bradley, J., Trivedi, R., et al. (2016). Choline diet and its gut microbe-derived metabolite, trimethylamine n-oxide, exacerbate pressure overload-induced heart failure. Circ.-Heart Fail. 9 (1). doi: 10.1161/CIRCHEARTFAILURE.115.002314
Ott, S. J., El Mokhtari, N. E., Musfeldt, M., Hellmig, S., Freitag, S., Rehman, A., et al. (2006). Detection of diverse bacterial signatures in atherosclerotic lesions of patients with coronary heart disease. Circulation 113 (7), 929–937. doi: 10.1161/CIRCULATIONAHA.105.579979
Paeslack, N., Mimmler, M., Becker, S., Gao, Z., Khuu, M. P., Mann, A., et al. (2022). Microbiota-derived tryptophan metabolites in vascular inflammation and cardiovascular disease. Amino Acids 1–18. doi: 10.1007/s00726-022-03161-5
Page, I. H. (1967). The mosaic theory of arterial hypertension–its interpretation. Perspect. Biol. Med. 10 (3), 325–333. doi: 10.1353/pbm.1967.0031
Pasini, E., Aquilani, R., Testa, C., Baiardi, P., Angioletti, S., Boschi, F., et al. (2016). Pathogenic gut flora in patients with chronic heart failure. JACC Heart Fail 4 (3), 220–227. doi: 10.1016/j.jchf.2015.10.009
Petriz, B. A., Castro, A. P., Almeida, J. A., Gomes, C. P., Fernandes, G. R., Kruger, R. H., et al. (2014). Exercise induction of gut microbiota modifications in obese, non-obese and hypertensive rats. BMC Genomics 15, 511. doi: 10.1186/1471-2164-15-511
Pluznick, J. (2014). A novel SCFA receptor, the microbiota, and blood pressure regulation. Gut Microbes 5 (2), 202–207. doi: 10.4161/gmic.27492
Ponikowski, P., Voors, A. A., Anker, S. D., Bueno, H., Cleland, J. G. F., Coats, A. J. S., et al. (2016). 2016 ESC guidelines for the diagnosis and treatment of acute and chronic heart failure: The task force for the diagnosis and treatment of acute and chronic heart failure of the European society of cardiology (ESC)Developed with the special contribution of the heart failure association (HFA) of the ESC. Eur. Heart J. 37 (27), 2129–2200. doi: 10.1093/eurheartj/ehw128
Ponziani, F. R., Zocco, M. A., D’Aversa, F., Pompili, M., Gasbarrini, A. (2017). Eubiotic properties of rifaximin: Disruption of the traditional concepts in gut microbiota modulation. World J. Gastroenterol. 23 (25), 4491–4499. doi: 10.3748/wjg.v23.i25.4491
Qin, J., Li, R., Raes, J., Arumugam, M., Burgdorf, K. S., Manichanh, C., et al. (2010). A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464 (7285), 59–65. doi: 10.1038/nature08821
Roessler, J., Leistner, D. M., Landmesser, U., Haghikia, A. (2022). Modulatory role of gut microbiota in cholesterol and glucose metabolism: Potential implications for atherosclerotic cardiovascular disease atherosclerosis. Atherosclerosis 359, 1–12. doi: 10.1016/j.atherosclerosis.2022.08.018
Sandek, A., Bauditz, J., Swidsinski, A., Buhner, S., Weber-Eibel, J., von Haehling, S., et al. (2007). Altered intestinal function in patients with chronic heart failure. J. Am. Coll. Cardiol. 50 (16), 1561–1569. doi: 10.1016/j.jacc.2007.07.016
Savi, M., Bocchi, L., Bresciani, L., Falco, A., Quaini, F., Mena, P., et al. (2018). Trimethylamine-N-Oxide (TMAO)-induced impairment of cardiomyocyte function and the protective role of urolithin b-glucuronide. Molecules 23 (3), 549. doi: 10.3390/molecules23030549
Schuett, K., Kleber, M. E., Scharnagl, H., Lorkowski, S., Marz, W., Niessner, A., et al. (2017). Trimethylamine-n-oxide and heart failure with reduced versus preserved ejection fraction. J. Am. Coll. Cardiol. 70 (25), 3202–3204. doi: 10.1016/j.jacc.2017.10.064
Seldin, M. M., Meng, Y., Qi, H., Zhu, W., Wang, Z., Hazen, S. L., et al. (2016). Trimethylamine n-oxide promotes vascular inflammation through signaling of mitogen-activated protein kinase and nuclear factor-kappaB. J. Am. Heart Assoc. 5 (2). doi: 10.1161/JAHA.115.002767
Siu, L. L., Even, C., Mesía, R., Remenar, E., Daste, A., Delord, J.-P., et al. (2019). Safety and efficacy of durvalumab with or without tremelimumab in patients with PD-L1-Low/Negative recurrent or metastatic HNSCC: The phase 2 CONDOR randomized clinical trial. JAMA Oncol. 5 (2), 195–203. doi: 10.1001/jamaoncol.2018.4628
Tang, T. W. H., Chen, H. C., Chen, C. Y., Yen, C. Y. T., Lin, C. J., Prajnamitra, R. P., et al. (2019). Loss of gut microbiota alters immune system composition and cripples postinfarction cardiac repair. Circulation 139 (5), 647–659. doi: 10.1161/CIRCULATIONAHA.118.035235
Tang, T. T., Yuan, J., Zhu, Z. F., Zhang, W. C., Xiao, H., Xia, N., et al. (2012). Regulatory T cells ameliorate cardiac remodeling after myocardial infarction. Basic Res. Cardiol. 107 (1), 232. doi: 10.1007/s00395-011-0232-6
Taur, Y., Coyte, K., Schluter, J., Robilotti, E., Figueroa, C., Gjonbalaj, M., et al. (2018). Reconstitution of the gut microbiota of antibiotic-treated patients by autologous fecal microbiota transplant. Sci. Transl. Med. 10 (460). doi: 10.1126/scitranslmed.aap9489
Thomas, R. M., Jobin, C. (2020). Microbiota in pancreatic health and disease: the next frontier in microbiome research. Nat. Rev. Gastroenterol. Hepatol. 17 (1), 53–64. doi: 10.1038/s41575-019-0242-7
Wampach, L., Heintz-Buschart, A., Hogan, A., Muller, E. E. L., Narayanasamy, S., Laczny, C. C., et al. (2017). Colonization and succession within the human gut microbiome by archaea, bacteria, and microeukaryotes during the first year of life. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.00738
Wang, A. L., Cai, D. C., Zhang, H., Li, Q. Y., Zhang, Y., Qiu, Y., et al. (2019). Using herbal medicine to target the “microbiota-metabolism-immunity” axis as possible therapy for cardiovascular disease. Pharmacol. Res. 142, 205–222. doi: 10.1016/j.phrs.2019.02.018
Wang, Z., Klipfell, E., Bennett, B. J., Koeth, R., Levison, B. S., Dugar, B., et al. (2011). Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 472 (7341), 57–63. doi: 10.1038/nature09922
Wang, G., Kong, B., Shuai, W., Fu, H., Jiang, X., Huang, H. (2020). 3,3-Dimethyl-1-butanol attenuates cardiac remodeling in pressure-overload-induced heart failure mice. J. Nutr. Biochem. 78, 108341. doi: 10.1016/j.jnutbio.2020.108341
Wang, Z., Roberts, A. B., Buffa, J. A., Levison, B. S., Zhu, W., Org, E., et al. (2015). Non-lethal inhibition of gut microbial trimethylamine production for the treatment of atherosclerosis. Cell 163 (7), 1585–1595. doi: 10.1016/j.cell.2015.11.055
Wilck, N., Matus, M. G., Kearney, S. M., Olesen, S. W., Forslund, K., Bartolomaeus, H., et al. (2017). Salt-responsive gut commensal modulates TH17 axis and disease. Nature 551 (7682), 585–589. doi: 10.1038/nature24628
Writing Group, M., Mozaffarian, D., Benjamin, E. J., Go, A. S., Arnett, D. K., Blaha, M. J., et al. (2016). Heart disease and stroke statistics-2016 update: A report from the American heart association. Circulation 133 (4), e38–360. doi: 10.1161/CIR.0000000000000350
Wu, J., Guo, Y., Lu, X., Huang, F., Lv, F., Wei, D., et al. (2020a). Th1/Th2 cells and associated cytokines in acute hepatitis e and related acute liver failure. J. Immunol. Res. 2020. doi: 10.1155/2020/6027361
Wu, J., Huang, F., Ling, Z., Liu, S., Liu, J., Fan, J., et al. (2020b). Altered faecal microbiota on the expression of Th cells responses in the exacerbation of patients with hepatitis e infection. J. Viral Hepat. 27 (11), 1243–1252. doi: 10.1111/jvh.13344
Xiao, S., Fei, N., Pang, X., Shen, J., Wang, L., Zhang, B., et al. (2014). A gut microbiota-targeted dietary intervention for amelioration of chronic inflammation underlying metabolic syndrome. FEMS Microbiol. Ecol. 87 (2), 357–367. doi: 10.1111/1574-6941.12228
Xie, G., Yan, A., Lin, P., Wang, Y., Guo, L. (2021). Trimethylamine n-oxide-a marker for atherosclerotic vascular disease. Rev. Cardiovasc. Med. 22 (3), 787–797. doi: 10.31083/j.rcm2203085
Yatsunenko, T., Rey, F. E., Manary, M. J., Trehan, I., Dominguez-Bello, M. G., Contreras, M., et al. (2012). Human gut microbiome viewed across age and geography. Nature 486 (7402), 222–227. doi: 10.1038/nature11053
Yisireyili, M., Shimizu, H., Saito, S., Enomoto, A., Nishijima, F., Niwa, T. (2013). Indoxyl sulfate promotes cardiac fibrosis with enhanced oxidative stress in hypertensive rats. Life Sci. 92 (24-26), 1180–1185. doi: 10.1016/j.lfs.2013.05.008
Zabell, A., Tang, W. H. (2017). Targeting the microbiome in heart failure. Curr. Treat Options Cardiovasc. Med. 19 (4), 27. doi: 10.1007/s11936-017-0528-4
Zhang, F., Zhang, T., Zhu, H., Borody, T. J. (2019). Evolution of fecal microbiota transplantation in methodology and ethical issues. Curr. Opin. Pharmacol. 49, 11–16. doi: 10.1016/j.coph.2019.04.004
Zhou, X., Li, J., Guo, J., Geng, B., Ji, W., Zhao, Q., et al. (2018). Gut-dependent microbial translocation induces inflammation and cardiovascular events after ST-elevation myocardial infarction. Microbiome 6 (1), 66. doi: 10.1186/s40168-018-0441-4
Zhu, W., Gregory, J. C., Org, E., Buffa, J. A., Gupta, N., Wang, Z., et al. (2016). Gut microbial metabolite TMAO enhances platelet hyperreactivity and thrombosis risk. Cell 165 (1), 111–124. doi: 10.1016/j.cell.2016.02.011
Keywords: gut microbiota, cardiovascular diseases (CVDs), metabolites, association, therapies
Citation: Qian B, Zhang K, Li Y and Sun K (2022) Update on gut microbiota in cardiovascular diseases. Front. Cell. Infect. Microbiol. 12:1059349. doi: 10.3389/fcimb.2022.1059349
Received: 01 October 2022; Accepted: 25 October 2022;
Published: 10 November 2022.
Edited by:
Yafeng Li, The Fifth Hospital of Shanxi Medical University, ChinaReviewed by:
Christoph Reinhardt, Johannes Gutenberg University Mainz, GermanyCopyright © 2022 Qian, Zhang, Li and Sun. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kangyun Sun, c3Vua2FuZ3l1bkBuam11LmVkdS5jbg==; Yuan Li, bGl5dWFuMTU5NjIxNUAxNjMuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.