 Larissa K. Ratten1,2*
Larissa K. Ratten1,2* Erica L. Plummer1,2
Erica L. Plummer1,2 Catriona S. Bradshaw1,2,3Christopher K. Fairley1,2Gerald L. Murray4,5,6
Catriona S. Bradshaw1,2,3Christopher K. Fairley1,2Gerald L. Murray4,5,6 Suzanne M. Garland4,5,6Deborah Bateson7,8Gilda Tachedjian9,10,11
Suzanne M. Garland4,5,6Deborah Bateson7,8Gilda Tachedjian9,10,11 Lindi Masson1,9,12,13
Lindi Masson1,9,12,13 Lenka A. Vodstrcil1,2,3*
Lenka A. Vodstrcil1,2,3*- 1Central Clinical School, Monash University, Carlton, VIC, Australia
- 2Melbourne Sexual Health Centre, Alfred Health, Carlton, VIC, Australia
- 3Melbourne School of Population and Global Health, The University of Melbourne, Parkville, VIC, Australia
- 4Department of Obstetrics and Gynaecology, The University of Melbourne, Parkville, VIC, Australia
- 5Centre for Women’s Infectious Diseases Research, The Royal Women’s Hospital, Parkville, VIC, Australia
- 6Infection and Immunity, Murdoch Children’s Research Institute, Parkville, VIC, Australia
- 7Family Planning NSW, Sydney, NSW, Australia
- 8Discipline of Obstetrics, Gynaecology and Neonatology, Faculty of Medicine and Health, The University of Sydney, Sydney, NSW, Australia
- 9Life Sciences Discipline, Burnet Institute, Melbourne, VIC, Australia
- 10Department of Microbiology, Monash University, Clayton, VIC, Australia
- 11Department of Microbiology and Immunology, Peter Doherty Institute for Infection and Immunity, University of Melbourne, Melbourne, VIC, Australia
- 12Institute of Infectious Disease and Molecular Medicine and Department of Pathology, University of Cape Town, Cape Town, South Africa
- 13Centre for the AIDS Programme of Research in South Africa, Durban, South Africa
Background: Exogenous sex steroids within hormonal contraception and menopausal hormone therapy (MHT) have been used for family planning and management of menopausal symptoms, without consideration of their effects on the vaginal microbiota. This is largely because their use predates our understanding of the importance of the vaginal microbiome on human health. We conducted a systematic review (PROSPERO: CRD42018107730) to determine the influence of exogenous sex steroids, stratified by oestrogen-containing or progestin-only types of contraception, and MHT on the vaginal microbiome, as measured by molecular methods.
Methods: Embase, PubMed and Medline were searched for relevant literature published through to December 1st 2020. Eligible studies reported on the effect of specific exogenous sex steroids on the vaginal microbiome using a molecular method. Data regarding the ‘positive’, ‘negative’ or ‘neutral’ effect of each type of contraceptive or MHT on the vaginal microbiome was extracted and summarised. A positive effect reflected sex steroid exposure that was associated with increased abundance of lactobacilli, a change to, or maintenance of, an optimal vaginal microbiota composition, or a decrease in bacterial diversity (specifically reflecting a low-diversity optimal microbiota state), relative to the control group. An exogenous sex steroid was designated as having a negative effect on the vaginal microbiome if it resulted in opposing effects (i.e. loss of lactobacilli, a non-optimal microbiota state). When no significant change was found, this was considered neutral/inconclusive.
Results: We identified 29 manuscripts reporting on the effect of exogenous sex steroids on the vaginal microbiome; 25 investigating hormonal contraceptives, and 4 investigating MHT. Oestrogen-containing contraception, particularly reflecting the combined oestrogen and progestin-containing contraceptive pill, had a positive effect on the composition of the vaginal microbiota. Progestin-only contraception, particularly reflecting depo-medroxyprogesterone acetate, had mixed effects on the microbiota. Among post-menopausal women using MHT, exogenous oestrogen applied topically was associated with increased prevalence of lactobacilli.
Conclusion: Our findings suggest that oestrogen-containing compounds may promote an optimal vaginal microbiota, which could have clinical applications. The impact of progestin-only contraceptives on the vaginal microbiota is less clear; more data is needed to determine how progestin-only contraceptives contribute to adverse reproductive and sexual health outcomes.
Introduction
Hormonal contraceptives have been in use since the 1960s and until recently, have not been examined for their influence on the vaginal microbiota. A vaginal microbiota associated with optimal reproductive and sexual health outcomes is characterised by Lactobacillus spp., although microbiome composition varies across geographical locations and specific populations (Fredricks et al., 2005; Ravel et al., 2011; Aldunate et al., 2013). Conversely, a non-optimal vaginal microbiota is typically characterised by reduced abundance of Lactobacillus spp. and is associated with negative sexual and reproductive health outcomes such as preterm delivery, miscarriage, low birthweight, pelvic inflammatory disease, and STI and HIV acquisition (Koumans et al., 1999; Wiesenfeld et al., 2002; Brotman et al., 2010; Chetwin et al., 2019; McKinnon et al., 2019). A non-optimal vaginal microbiota abundant in Gardnerella spp., including G. vaginalis, Atopobium vaginae and other facultative and strict anaerobes, is associated with the most common vaginal dysbiosis, bacterial vaginosis (BV) (Fredricks et al., 2005; Ravel et al., 2011; McKinnon et al., 2019). Prior systematic reviews and meta-analysis of observational data have shown that exogenous sex steroids, delivered as either oestrogen-containing and progestin-only contraceptives may exert a positive effect on the microbiota, with a decrease in BV (van de Wijgert et al., 2013; Vodstrcil et al., 2013). However, some progestin-only contraceptives (i.e. depo-medroxyprogesterone acetate [DMPA]), have been scrutinised for a suspected link between their use and an increased risk of HIV acquisition and transmission (Polis and Curtis, 2013; Deese et al., 2015; Dabee et al., 2021). Among post-menopausal women, the composition of the vaginal microbiota is implicated in vaginitis, which is a common condition in this population (Diem et al., 2018; The 2020 Genitourinary Syndrome of Menopause Position Statement of The North American Menopause Society, 2020). Vaginitis (formerly atrophic vaginitis) and other peri-and post-menopausal symptoms are managed with exogenous oestrogen within menopausal hormone therapy (MHT, also known as hormonal replacement therapy) in some cases (Castelo-Branco et al., 2005; Chollet et al., 2009). However, less is known about what constitutes an optimal vaginal microbiota in this population (Mitchell et al., 2017).
The use of hormonal contraceptives and MHT is rising globally (Contraceptive Use by Method 2019: Data Booklet. 2019) and clearly, these compounds need to be examined for their largely unappreciated effects on the vaginal microbiota. Advancements in molecular technology have allowed for more accurate and high-throughput studies examining the vaginal microbiota to be conducted. Given the uncertainty about the impact of specific hormonal contraceptives on the vaginal microbiota, this timely systematic review summarises the effect of specific oestrogen-containing or progestin-only contraceptives as well as MHT on the vaginal microbiota, and critically evaluates the strength of the findings.
Methods
Protocol and Registration
This systematic review was conducted in line with the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-analyses) statement (Page et al., 2021) and was prospectively registered with PROSPERO on the 5/09/2018 (International Prospective Register of Systematic Reviews; CRD42018107730).
Search Process, Eligibility Criteria
We searched the electronic databases Embase, PubMed and Medline for relevant literature published through to December 1st 2020 using the following strings: [(vagina* AND bacter* OR vagina*) AND micro* OR vagina*] AND flora AND [(hormon* AND contracept* OR oral) AND contracept* OR estrogen OR oestrogen OR progest*]. Studies were uploaded to Covidence (Veritas Health Innovation, Melbourne, Australia, www.covidence.org) and screened by two authors (L.K.R and E.L.P); with any disputes resolved by a third author (L.A.V). Conference abstracts were included for screening; only English language studies were considered and only data from human participants was examined.
To be eligible for inclusion, studies were required to report on the most commonly used sex steroids (i.e. oestrogen and/or progestin delivered as hormonal contraceptives or MHT) and provide a measure of the composition, stability and/or diversity of the vaginal microbiota by molecular methods (Supplementary Table 1). Molecular methods included i) quantitative polymerase chain reaction (qPCR), ii) 16S microarray, iii) next-generation sequencing and/or iv) Sanger sequencing. Studies were excluded if they 1) did not report on sex steroid use; 2) only measured the vaginal microbiota by non-molecular methods (such as culture and microscopy); 3) were qPCR studies that did not include at least one Lactobacillus target or one BV-associated bacteria; 4) only included pregnant women or women undergoing in vitro fertilisation; or 5) were reviews. Studies were not excluded based on trial design and could include cross-sectional studies, cohorts, and randomised trials. Where two studies presented data on the same population, only the most recent study was retained, unless one presented baseline cross-sectional data and the second presented longitudinal data, in which case both were included. Where studies measured the microbiota with molecular methods and reported sex steroid use but did not present findings on the effect of sex steroids on the microbiota, the study authors were contacted for additional details. Authors from seven studies were contacted via email, all responded and two provided additional data for three studies.
Study Population and Interventions Assessed
We deliberately did not specify a population age, which resulted in two distinct study populations identified in the literature search; reproductive-aged women using hormonal contraception and post-menopausal women using MHT, which were analysed separately. In the reproductive-aged population, any kind of hormonal contraception type was included as a suitable intervention, however these predominantly reflected oestrogen-containing or progestin-only methods of hormonal contraception, as these are the most commonly used formulations. Where more than one sex steroid use was reported in the study population, all types were recorded. In the post-menopausal population, topical conjugated oestrogens were used. The comparator group varied between studies and included 1) baseline specimens prior to sex steroid initiation, women not using contraception, or MHT, or women using a non-hormonal contraceptive [i.e. copper IUD (Cu-IUD), condom use].
Data Extraction
Data was extracted by two authors (L.R. and E.P.). Any discrepancies were resolved through discussion with a third author (L.V.). The following data were extracted: i) study location, ii) population characteristics (e.g. HIV status, sex-worker status, age, and ethnicity), iii) frequency of sampling, iv) molecular technique applied, v) molecular outcomes reported on, vi) comparator group used, vii) sex steroid source, viii) molecular findings and ix) summarised effect of sex steroid exposure (positive, neutral or negative).
Data Analysis
For studies using next generation sequencing, the impact of exogenous sex steroids on vaginal microbiota was reported as relative abundance, bacterial diversity, community state type (CST)/vaginal microbiota cluster or group assignment, and change in these measurements over time during sex steroid exposure. All results were stratified by sex steroid type. Microarray and qPCR study findings were dependent on the panel targets tested. Most commonly, biologically significant Lactobacillus spp. (i.e. L. crispatus, L. iners, L. gasseri, L. jensenii, L. johnsonii and L. vaginalis) as well as BV-associated bacteria [i.e. G. vaginalis, A. vaginae, Megasphaera, Leptotrichia, Sneathia, Prevotella, Lachnovaginosum genomospecies (formerly BVAB1), BVAB2, and Mycoplasma] were investigated. For these studies, the impact of exogenous sex steroids on the vaginal microbiota was reported as presence/absence, prevalence, and bacterial load (as mean target copies and log concentration) of key bacteria and change in these measurements stratified by sex steroid exposure over time. Studies that measured the vaginal microbiota with denaturing gradient gel electrophoresis (DGGE) and Sanger sequencing reported types and number of bacterial species present, but not relative abundance.
Definitions of Outcomes
Findings were classified as positive, negative, or neutral and further qualified as significant or non-significant. Positive findings generally reflected an increase in Lactobacillus spp. among women with sex-steroid exposure. Despite controversy in the field surrounding the role of L. iners (Petrova et al., 2017), L. iners was grouped with other Lactobacillus spp. for the purposes of this review as it is still considered preferable over BV-associated bacteria. An exogenous sex steroid was designated to have a positive effect on the vaginal microbiota if they reported any of the following i) an increase in abundance and/or prevalence of Lactobacillus spp. (or decrease in BV-associated bacteria) following initiation of sex steroid use and/or relative to the control group (i.e. either baseline sample or no hormonal contraceptive-use group), ii) a change to a compositional state reflecting an optimal vaginal microbiota [defined as a vaginal microbiota dominated by lactobacilli such as CST-I, CST-II, CST-III (Ravel et al., 2011)], iii) maintenance of an optimal vaginal microbiota state over time, or iv) a decrease in bacterial diversity, specifically reflecting a low-diversity optimal vaginal microbiota state relative to the control group for that study. An exogenous sex steroid was designated as having a negative effect on the microbiota if it was associated with i) a loss of L. crispatus or any other Lactobacillus spp. relative to the control group, ii) an increase in prevalence and/or abundance of any BV-associated bacteria, iii) a change in compositional state reflecting a non-optimal state (e.g. one dominated/characterised by BV-associated bacteria such as CST-IV, CST-V), and iv) an increase in diversity associated with a higher diversity non-optimal state and/or instability of the vaginal microbiota, relative to the control group used in the study. When there was no significant change (or no difference between exposure and control group) in vaginal microbiota state, stability, or prevalence/abundance of bacterial species reported, an exogenous sex steroid was considered to have a neutral or inconclusive effect.
Assessment of Bias and Quality
Two authors (L.K.R. and E.L.P.) assessed the risk of bias within studies and reported on i) representation of the general population, ii) intervention allocation, iii) sample size, iv) comparator group/s, v) stratification by intervention, vi) analysis adjusted for confounding variables and vii) methodology, as further defined in Supplementary Table 2. Studies were not excluded based on the risk of bias assessment.
Results
Study Selection
Literature searches identified 5315 records from Embase, Ovid Medline and PubMed; and two records were found through other sources. Duplicate references (n=2668) were removed, generating 2649 unique references (Figure 1). Of these, 2384 were excluded by title and abstract review. Of the remaining 265 articles, 236 were excluded during full-text screening. The reasons for excluding full text were: molecular methods not used (n=119), review article (n=32), article not available in English (n=20), insufficient bacterial targets (i.e. qPCR did not target BV-associated bacteria or Lactobacillus spp.) (n=23), no sex steroid outcomes reported (n=28), study not in humans (n=8) and datasets already represented in another publication (n=6). Twenty-nine eligible studies were identified and data extracted; with 25 studies reporting on the effect of hormonal contraception among reproductive-aged women (Table 1) and four on the effect of MHT among post-menopausal women (Table 2). Two studies reported on the same cohort; however, these were treated as separate studies as one reported on baseline data (Tuddenham et al., 2019a) and the second presented longitudinal data (Tuddenham et al., 2019b).
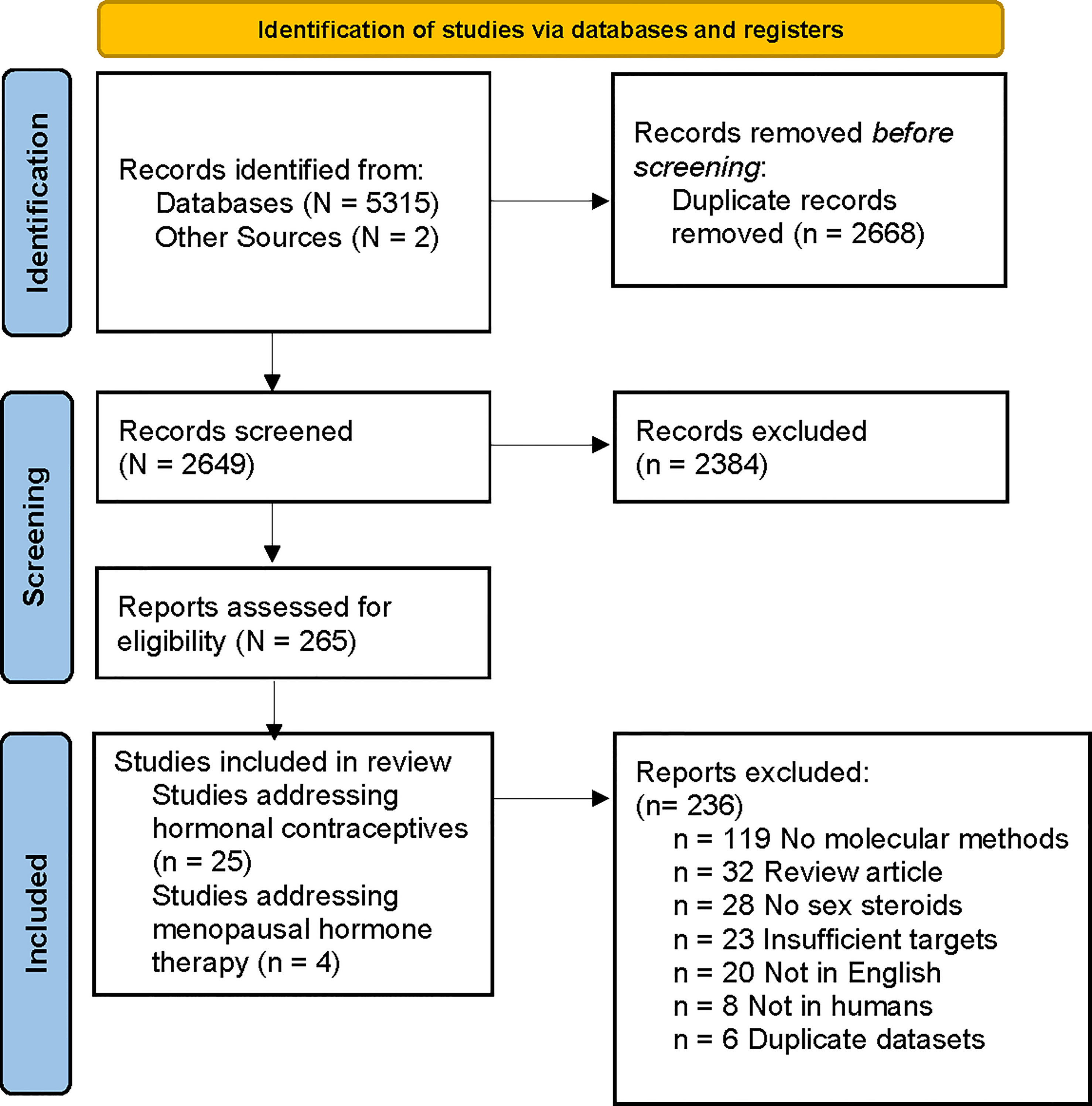
Figure 1 PRISMA flow diagram of literature search and article selection. Flow chart adapted from Page et al. (2021).
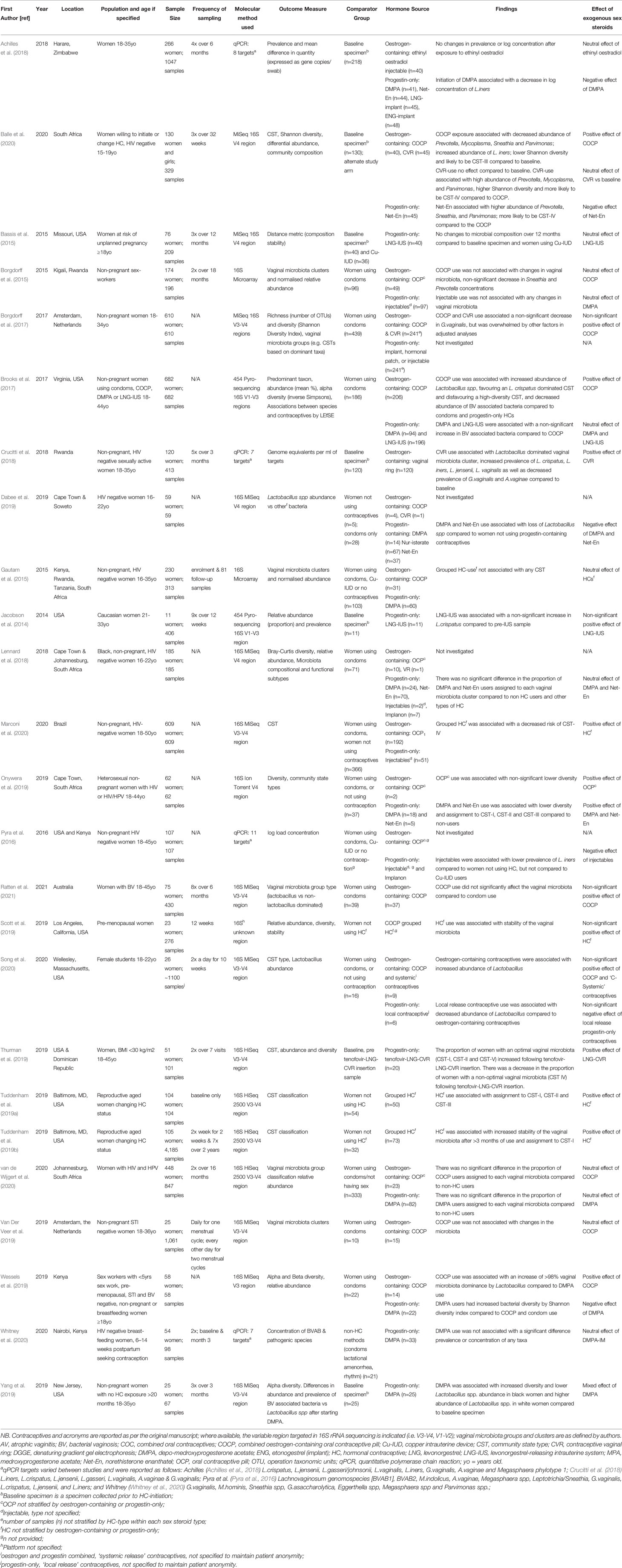
Table 1 Result summary of hormonal contraceptive studies.

Table 2 Summary of included menopausal hormone therapy studies.
Studies Reporting on the Effect of Hormonal Contraceptives on the Vaginal Microbiota
General Characteristics
A summary of the populations included in the studies identified is provided in Supplementary Table 3. Of the 25 studies addressing the effect of hormonal contraception on the vaginal microbiota, 20 of these were in HIV negative, non-sex workers without BV, two were in women with HIV and human papillomavirus (HPV), two were in sex workers with no other risk factors specified, and one was among women with symptomatic BV. Nine studies were cross-sectional and 16 were longitudinal cohort studies; the median follow-up time was 31 weeks (range 7 days to 24 months). Nineteen of the studies investigated the vaginal microbiota using next-generation sequencing technology, four used qPCR and two used microarray. Twelve studies were conducted in sub-Saharan Africa, eight were conducted in North America, two in Europe/Central Asia, two were in Latin America/Caribbean and one was conducted in the East-Asian and Pacific region; there were no studies in South Asia or the Middle East/North Africa. These studies represent approximately 14,000 specimens from approximately 4,400 women; of which 1,048 women were exposed to combined oestrogen- and progestin-containing contraceptives (termed oestrogen-containing contraceptives) and 968 were exposed to progestin-only contraceptives (Table 1). The combined-oestrogen containing contraceptive-types reported on included i) oral contraceptive pill (OCP), ii) contraceptive vaginal ring (CVR) and iii) ethinyl oestradiol-containing injectable. The progestin-only contraceptive types included i) injectable depo-medroxyprogesterone acetate (DMPA), ii) injectable norethisterone enanthate (Net-En), and iii) levonorgestrel (LNG) contained with an intrauterine system (LNG-IUS) or CVR (LNG-CVR). Other progestin-only contraceptives, including subdermal contraceptive implants and patch-use, were identified, but there was no or limited data reported and so they were not included in the review. The remaining women were either not exposed to contraceptives/or used non-hormonal contraceptive methods (including condoms or the Cu-IUD). Most of the studies (15/25) identified investigated more than one type of hormonal contraception, and we have stratified the findings by sex steroid type where possible (Table 1).
The Effect of Oestrogen-Containing Hormonal Contraceptive Types on the Vaginal Microbiome
Fourteen studies investigated the effect of combined oestrogen- and progestin-containing contraceptives, which included the combined oestrogen-containing OCP, an oestrogen-containing CVR and the ethinyl oestradiol-containing injectable. When grouped together, 8/14 (57%) identified a positive effect of oestrogen-containing hormonal contraception on the vaginal microbiota, 5/14 (36%) found neutral or no effect, and one study found a negative effect (7%; Table 3).

Table 3 Summary of beneficial, neutral/inconclusive and detrimental effects of exogenous sex steroids on the vaginal microbiota.
Oestrogen-Containing Oral Contraceptive Pills
Ten studies investigated the effect of the combined oestrogen-containing OCP on the vaginal microbiome (Borgdorff et al., 2015; Borgdorff et al., 2017; Brooks et al., 2017; Onywera et al., 2019; Van Der Veer et al., 2019; Wessels et al., 2019; Balle et al., 2020; Song et al., 2020; van de Wijgert et al., 2020; Ratten et al., 2021); six reported a positive effect on the vaginal microbiota, and four reported neutral findings (Tables 1 and 3). The OCPs were specified as oestrogen-containing by the authors in all studies, except for two (Borgdorff et al., 2015; Onywera et al., 2019). However, for the purpose of this review, these two studies were assigned as studies of oestrogen-containing OCP, and this was confirmed by the authors to be the most common formulation. Four studies found OCP-use had a positive effect on Lactobacillus spp. (Tables 1 and 3); three studies reported that OCP-use was associated with a higher relative abundance of Lactobacillus spp. compared to women not using the OCP (Brooks et al., 2017; Song et al., 2020), or women’s baseline (i.e. pre-OCP initiation) specimens (Balle et al., 2020), and one reported an increase in the number of women with a vaginal microbiota dominated by Lactobacillus spp. (i.e. >98% relative abundance of Lactobacillus spp.) compared to women using condoms (Wessels et al., 2019). Balle et al. specifically identified that OCP-use increased the relative abundance of L. iners compared to participants’ baseline specimens (Balle et al., 2020). Onywera et al. (2019) and Balle et al. (2020) both reported that OCP-use decreased bacterial diversity (measured by the Shannon diversity index), compared to non-OCP users and baseline specimens, respectively. Balle et al. (2020) also found that OCP-use decreased the relative abundance of several BV-associated bacteria, specifically Prevotella, Sneathia and Parvimonas relative to baseline and Net-En users, while Borgdorff et al. (2017) found a non-significant decrease in the relative abundance of G. vaginalis in OCP-users vs non-OCP users. Four studies found no effect of the OCP on the vaginal microbiota compared to women using condoms (Borgdorff et al., 2015; Van Der Veer et al., 2019; van de Wijgert et al., 2020; Ratten et al., 2021).
Oestrogen-Containing Contraceptive Vaginal Ring
Three studies investigated the effect of the oestrogen-containing CVR (Borgdorff et al., 2017; Crucitti et al., 2018; Balle et al., 2020). Compared to baseline specimens, Borgdorff et al. (2017) found that CVR-use was associated with a non-significant decrease in the relative abundance of G. vaginalis and Crucitti et al. (2018) found CVR-use was associated with a significant decrease in the prevalence G.vaginalis and A.vaginae (Table 1). Crucitti et al. (2018) also found that CVR-use was associated with an increase in the prevalence and load of Lactobacillus spp. in the vaginal microbiota, specifically L. crispatus, L. iners, L. jensenii and L. vaginalis. Conversely, Balle et al. (2020) found that oestrogen-containing CVR use had no impact on bacterial diversity compared to baseline. However, compared to OCP users, CVR-use was associated with a higher relative abundance of Prevotella, Mycoplasma, and Parvimonas, higher bacterial diversity and increased likelihood of having a vaginal microbiota composition designated to CST-IV (Balle et al., 2020).
Ethinyl Oestradiol Containing Injectable
Only one study investigated the effect of the ethinyl oestradiol-containing injectable among 40 women in Zimbabwe (Achilles et al., 2018). There were no significant changes to the composition of the vaginal microbiota following ethinyl oestrodial-containing injectable use relative to baseline specimens.
The Effect of Progestin-Only Contraceptives on the Vaginal Microbiotas
Twenty studies investigated the effect of progestin-only contraceptives, representing DMPA, Net-En and the levonorgestrel CVR and IUS. When looked at together, four studies identified a positive effect of progestin-only contraceptives on the vaginal microbiota, nine described a negative effect on the microbiota, one study reported different effects across patient sub-populations, and seven reported neutral or inconclusive effects (Tables 1 and 3).
Depot-Medroxyprogesterone Acetate Injectable
Of the ten studies that investigated DMPA (Borgdorff et al., 2015; Brooks et al., 2017; Achilles et al., 2018; Lennard et al., 2018; Dabee et al., 2019; Onywera et al., 2019; Wessels et al., 2019; Yang et al., 2019; van de Wijgert et al., 2020; Whitney et al., 2020), three reported a negative effect of DMPA on the vaginal microbiota, one reported a positive effect, five reported neutral findings and one reported mixed findings. DMPA-use was associated with a decrease in the relative abundance of Lactobacillus spp. in two studies (additional data provided by K. Lennard) (Dabee et al., 2019; Yang et al., 2019). In the study by Yang et al, DMPA-use had a different effect in the two study populations; it was associated with a decreased abundance of Lactobacillus spp. among African American/black women and an increase in relative abundance of Lactobacillus spp. among Caucasian/white women (Yang et al., 2019). Achilles et al. (2018) also reported a loss of Lactobacillus spp., specifically a decrease in log concentration of L. iners over six months of DMPA-use. Brooks et al. (2017) described a non-significant increase in the relative abundance of BV-associated bacteria associated with DMPA-use, which was classified as an inconclusive finding. Wessels et al. (2019) found that DMPA users had an increase in bacterial diversity (measured by Shannon diversity index). Conversely, Onywera et al. (2019) found that DMPA-use was associated with lower bacterial diversity (measured by Shannon diversity index) and a vaginal microbiota assigned to Lactobacillus spp. dominated CSTs (CST-I, CST-II and CST-III) relative to women using condoms or no contraceptives. One study found that DMPA had no effect on the prevalence and concentrations of BV-associated bacteria Sneathia, M. hominis and Parvimonas species type 1 vs women who did not use contraception/used non-hormonal contraceptives (Whitney et al., 2020). However, this study did not specifically look at the effect of DMPA on lactobacilli. Three studies found that there was no significant difference in the proportion of DMPA-users assigned to specific vaginal microbiota groups associated with optimal or non-optimal compositions compared to non-hormonal contraceptive users and/or users of other hormonal contraceptive types (additional data provided by [Lennard et al. (2018), van de Wijgert et al. (2020)] (Borgdorff et al., 2015; Lennard et al., 2018; van de Wijgert et al., 2020). In addition, Pyra et al. (2016) investigated the effect of injectable contraceptives but did not specify the type used. They defined injectable contraceptives as having a negative effect, as their use was associated with a lower prevalence of L. iners. Overall, DMPA had mixed effects with nearly equal numbers of studies reporting positive or neutral effects and negative effects on the vaginal microbiota.
Norethisterone Enanthate Injectable
Four studies investigated the effect of Net-En on the vaginal microbiota (Lennard et al., 2018; Dabee et al., 2019; Onywera et al., 2019; Balle et al., 2020), two of which had negative findings, one had positive findings and one had neutral findings. Balle et al. (2020) compared the effects of Net-En with participants’ baseline samples on the vaginal microbiota and found no effect of Net-En initiation. However, Net-En users were also more likely to be assigned to CST-IV than participants using the combined oestrogen-containing OCP (Balle et al., 2020). Dabee et al. (2019) found use of Net-En was associated with lower relative abundance of Lactobacillus spp. compared to women not using any hormonal contraception. In contrast, Onywera et al. (2019) found that Net-En use was associated with lower bacterial diversity (measured by Shannon diversity index) and assignment to Lactobacillus dominated CSTs compared to women not using hormonal contraception, however this represented a small proportion of Net-En users captured in this review (n=5). Lennard et al. (2018), had the most Net-En users (n=70) in their study and found that Net-En use was not associated with any specific vaginal microbiota composition when compared to women not using hormonal contraception, or women using other types of hormonal contraception (additional data provided by K. Lennard).
Levonorgestrel-Containing Contraceptive Rings and Intra-Uterine Systems
Four studies investigated the effect of levonorgestrel-containing systems on the vaginal microbiota (Jacobson et al., 2014; Bassis et al., 2015; Brooks et al., 2017; Thurman et al., 2019), two of which had positive findings, one had negative findings and the other had neutral findings. Thurman et al. (2019) found that the proportion of women with an optimal vaginal microbiota (CST I, II and V) increased following tenofovir-LNG-CVR insertion. There was a decrease in the proportion of women with a non-optimal vaginal microbiota (CST IV) following tenofovir-LNG-CVR insertion (Thurman et al., 2019). Three studies investigated the LNG-IUS with discordant findings; Jacobson et al. (2014) found that following insertion of LNG-IUS, the relative abundance of L. crispatus increased. Conversely, Brooks et al. (2017) compared LNG-IUS use to OCP-use and found a non-significant change in the relative abundance of BV-associated bacteria, while Bassis et al. (2015) found no difference in the vaginal microbiota composition between women using LNG-IUS and those using a Cu-IUD.
Song et al. (2020) grouped contraceptives as either oestrogen-containing or combined progestin-only “local release” contraceptives due to the small sample size (n=7, n=4 respectively). Compared to women not using hormonal contraception, oestrogen-containing contraceptives were associated with a non-significant increase in Lactobacillus spp. and use of progestin-only contraceptives was associated with a non-significant decrease in on the abundance of Lactobacillus spp (Song et al., 2020).
Studies That Grouped All Hormonal Contraceptives Together and Did Not Specify Type
Five studies did not stratify findings by hormonal contraception type (Tables 1 and 3; four reported a positive effect of grouped hormonal contraception-use on the vaginal microbiota and one reported a neutral effect. Scott et al. (2019) and Tuddenham et al. (2019b) found that grouped hormonal contraception-use was associated with stability of the vaginal microbiota. Both the cross-sectional and longitudinal studies by Tuddenham et al. (Tuddenham et al., 2019a; Tuddenham et al., 2019b) found that hormonal contraception-use was associated with a vaginal microbiota defined by optimal CSTs, and Marconi et al. (2020) found that hormonal contraception-use was associated with a decreased risk of having CST-IV. Gautam et al. (2015) reported that hormonal contraception-use did not change the vaginal microbiota.
Studies Reporting on the Effect of Menopausal Hormone Therapy on the Vaginal Microbiota
General Characteristics
Four studies that investigated the effect of MHT on the vaginal microbiota were identified (Tables 2 and 3) (Devillard et al., 2004; Heinemann and Reid, 2005; Dahn et al., 2008; Shen et al., 2016). These studies represent 332 samples from 158 women, and all four assessed the effect of a topical conjugated-oestrogen containing formulation (topical Premarin®). All studies were in post-menopausal women, with three studies performed in Canada (Devillard et al., 2004; Heinemann and Reid, 2005; Dahn et al., 2008) and one in China (Shen et al., 2016) (Supplementary Table 3). One study was cross-sectional (Dahn et al., 2008) and the other three were longitudinal (Devillard et al., 2004; Heinemann and Reid, 2005; Shen et al., 2016); the median follow up time was 10 weeks (range 4-13 weeks). Three studies used DGGE based methods (Devillard et al., 2004; Heinemann and Reid, 2005; Dahn et al., 2008); two were in combination with Sanger sequencing (Devillard et al., 2004; Heinemann and Reid, 2005) one used microarray. Shen et al. (2016) was the only study to use next-generation sequencing (Supplementary Table 3).
The Effect of Menopausal Hormone Therapy on the Vaginal Microbiota
All four studies found that use of topical conjugated-oestrogens increased the prevalence of Lactobacillus spp. compared to women not using menopausal hormone therapy (Devillard et al., 2004; Heinemann and Reid, 2005; Dahn et al., 2008; Shen et al., 2016) (Tables 2 and 3). Shen et al. (2016) and Heinemann and Reid (2005) also found that conjugated-oestrogen use decreased the bacterial diversity of the vaginal microbiota and in addition, Heinemann and Reid (2005) reported that its use increased the stability of the vaginal microbiota over time.
Assessment of Study Bias
We assessed each of the studies for Selection Bias, Sample size and Measurement Bias across 6 domains, with a summary score for risk of bias generated (Supplementary Table 2). Studies with the lowest summary score were considered to have the lowest risk of study bias, however none of the included studies had a low risk of bias across all criteria assessed (Table 4).
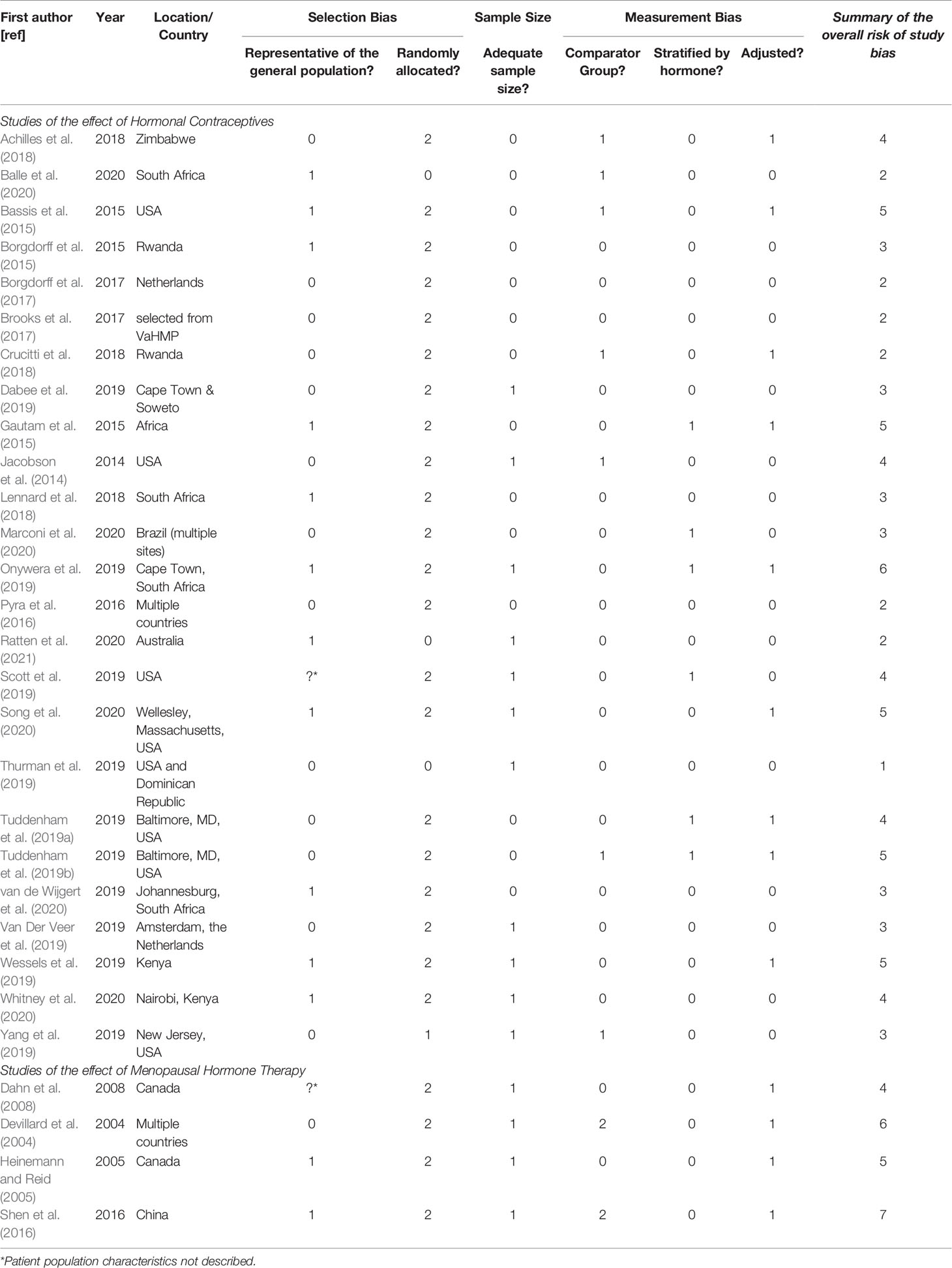
Table 4 Risk of Bias Summary Table.
Studies Reporting on the Effect of Hormonal Contraceptives on the Vaginal Microbiota Among Reproductive-Aged Women
When looking at selection bias across the 25 studies that investigated hormonal contraception, the study population was not described for only one study (Scott et al., 2019). One study was conducted on samples from a randomised control trial (Ratten et al., 2021) and in one, study participants were assigned treatment sequentially (Yang et al., 2019). For the remaining 22 studies, the participants self-selected their intervention. Eleven studies included a sample size of <100 participants total, which we assessed as high risk of bias (Jacobson et al., 2014; Dabee et al., 2019; Onywera et al., 2019; Scott et al., 2019; Van Der Veer et al., 2019; Yang et al., 2019; Song et al., 2020; Ratten et al., 2021). Eighteen studies used age-matched women not using hormonal contraceptives as the comparator group (e.g. no contraceptives or condom-use) (low risk), and seven used the baseline specimen prior to contraception initiation as the comparator group (low/medium risk) (Jacobson et al., 2014; Bassis et al., 2015; Achilles et al., 2018; Crucitti et al., 2018; Tuddenham et al., 2019b; Yang et al., 2019; Balle et al., 2020). Five studies did not stratify hormonal contraceptive exposure by oestrogen-containing or progestin-only (high risk) (Gautam et al., 2015; Scott et al., 2019; Tuddenham et al., 2019a; Tuddenham et al., 2019b; Marconi et al., 2020). Nine studies did not adjust analyses for confounding variables such as sex and douching practices (high risk) (Bassis et al., 2015; Gautam et al., 2015; Achilles et al., 2018; Crucitti et al., 2018; Tuddenham et al., 2019a; Tuddenham et al., 2019b; Onywera et al., 2019; Wessels et al., 2019; Song et al., 2020).
Studies Reporting on the Effect of Menopausal Hormone Therapy on the Vaginal Microbiota
Of the four MHT studies, one study did not define the study population (unknown risk) (Dahn et al., 2008), one was conducted in healthy women (classified as low risk or bias) (Devillard et al., 2004), and two were conducted in women with urogenital symptoms or infection (medium risk) (Heinemann and Reid, 2005; Shen et al., 2016). Women self-selected their therapy in all four studies, and all studies had fewer than 100 women and did not demonstrate sample size calculations for the effect of MHT use on the vaginal microbiota (high risk). Two studies had age-matched MHT-free women as comparators (low risk) (Dahn et al., 2008; Shen et al., 2016). In one study the comparator group was women who were not using MHT who had participated in another study (high risk) (Devillard et al., 2004). The fourth study compared findings to a group of women not using MHT, but these women had different symptoms to the MHT-exposed group (high risk) (Shen et al., 2016). All studies were stratified by hormone source (low risk), but no study adjusted for confounding variables (high risk).
Discussion
Exogenous sex steroids contained within hormonal contraceptives and menopausal hormone therapy have been used principally for family planning and management of menopausal symptoms, without consideration for potential effects on the vaginal microbiota. This is because their use in healthcare predates our current understanding of the importance of the vaginal microbiota in human health, largely due to the fact that they were adopted before the technology existed to assess the vaginal microbiota. Prior systematic reviews and a meta-analysis of observational data found that oestrogen-containing contraceptives in particular have a beneficial effect on the vaginal microbiota, as measured by non-molecular methods (van de Wijgert et al., 2013; Vodstrcil et al., 2013). In this systematic review, we aimed to determine the effects of exogenous sex steroid use on the vaginal microbiota, as measured using modern, molecular-based methods such as high-throughput sequencing. We found that oestrogen-containing contraceptives, particularly the combined oestrogen and progestin-containing OCP, had a positive effect on the composition of the vaginal microbiota. Among post-menopausal women using MHT, exogenous oestrogen also appeared to positively influence the vaginal microbiota. However, the significance of an optimal vaginal microbiota as defined in reproductive-aged women is not as well understood in the post-menopausal population. In contrast, contraceptives containing progestin alone had mixed effects on the vaginal microbiota of reproductive-aged women. Overall, our systematic review shows that oestrogen may play a role in supporting an optimal vaginal microbiota in both reproductive aged and peri/post-menopausal women. However, further well-powered studies with appropriate control groups are required to explore the specific effects of different oestrogen-containing and progestin-only contraceptives.
The Impact of Exogenous Oestrogen on the Vaginal Microbiota
We found that exogenous oestrogen, which predominantly reflected the use of the combined oestrogen and progestin-containing OCP, had a positive impact on the vaginal microbiota composition. Specifically, there was an increase in the prevalence and abundance of Lactobacillus spp. observed following oestrogen-exposure. Lactobacillus spp., particularly L. crispatus, characterise a vaginal microbiota associated with optimal reproductive and sexual health outcomes (Petrova et al., 2015; Anahtar et al., 2018). The positive effect of lactobacilli is proposed to be due to the production of lactic acid, which lowers the vaginal pH in addition to conferring antimicrobial and immunomodulatory benefits, and inhibiting the growth of anaerobic bacteria (Aldunate et al., 2015; Hearps et al., 2017; Tachedjian et al., 2017; Tyssen et al., 2018; Delgado-Diaz et al., 2020; Plummer et al., 2021). Different lactobacilli can produce two different lactic acid isomers, the L-isomer and D-isomer. The D-isomer is hypothesised to offer more protection against some bacterial upper genital tract infections such as chlamydia, however the L-isomer is more potent in inactivating HIV compared to the D-isomer at threshold concentrations in vitro (O’Hanlon et al., 2011; Aldunate et al., 2013; O’Hanlon et al., 2013; Witkin et al., 2013; Witkin, 2018; Edwards et al., 2019). In reproductive-aged women, endogenous oestrogen stimulates glycogen production by epithelial cells, which is then metabolised by Lactobacillus spp. as an energy source resulting in production of lactic acid (Boskey et al., 2001; O’Hanlon et al., 2011; O’Hanlon et al., 2013; van der Veer et al., 2019; Clabaut et al., 2021). Among women taking exogenous oestrogen, the amount of free glycogen available may increase or be more consistent throughout the menstrual cycle, and in turn indirectly increase lactic acid production (Mirmonsef and Spear, 2014; Nunn and Forney, 2016). Exogenous oestrogen could therefore have a therapeutic role among reproductive-aged women with a paucity of Lactobacillus spp. in their vaginal microbiota, such as in women with BV and vaginitis. In the one pilot randomised controlled trial to randomise women with BV receiving antibiotic treatment to adjunctive combined oestrogen-containing OCP-use or no OCP-use, there was no significant effect of sex steroid-exposure on BV recurrence rates (Vodstrcil et al., 2019). However, the findings of this study were impacted by the small sample size and attrition, and larger studies may be required to help us determine whether the use of oestrogen-containing contraceptives adjunctively or alone may positively influence the vaginal microbiome of women with BV.
The oestrogen-containing CVR had mixed impacts on the vaginal microbiota. Oestrogen-containing CVR-use was associated with an increase in several Lactobacillus spp. including L. crispatus, L. iners, L. vaginalis (Crucitti et al., 2018) and decrease in BV-associated bacteria such as G. vaginalis (Borgdorff et al., 2017; Crucitti et al., 2018). However, oestrogen-containing CVR-use was also shown to have no effect (Balle et al., 2020). Similarly, among women using progestin-only CVRs and intrauterine systems, the composition of the vaginal microbiota varied (Jacobson et al., 2014; Bassis et al., 2015; Brooks et al., 2017; Thurman et al., 2019). The high variability between these studies may be because of the limited number of studies, the different control groups used and/or the timing of the post-insertion specimens relative to when the contraceptive system was inserted. More studies are needed to understand the impact of CVR-use and IUS-use on vaginal microbiota composition.
Topical conjugated oestrogens in post-menopausal women was also associated with an increase in the abundance and prevalence of Lactobacillus spp. While there is abundant evidence to support the use of oestrogen to relieve post-menopausal symptoms such as vaginal dryness and discomfort (Hickey et al., 2016), we identified only four studies investigating menopausal hormone therapy on the vaginal microbiota. As there is limited information about the role of the vaginal microbiota in post-menopausal women, we do not know if there are benefits associated with re-establishing a Lactobacillus dominant vaginal microbiota in post-menopausal women.
The Effect of Progestin on the Vaginal Microbiota in Reproductive-Aged Women
There is global interest in how progestin-only hormonal contraception, especially DMPA, might impact the vaginal microbiota and may increase the risk of STI and/or HIV acquisition. Our systematic review found that the effect of progestin-only contraceptives on the vaginal microbiota was mixed. Of note, half (n=11) of these papers examined DMPA-use. In contrast to oestrogen-containing contraceptives, progestin-only contraceptives had an inconsistent effect on the abundance and prevalence of Lactobacillus spp. as well as other metrics such as bacterial diversity and prevalence/abundance of BV-associated bacteria. The inconsistent findings of the effect of progestin-only contraceptives on the vaginal microbiota within and/or between sub-populations may be explained by host genetics and gene polymorphisms (Dabee et al., 2021), however further research is needed. The effect of DMPA on the vaginal microbiota is of particular interest due to its high rate of use in sub-Saharan Africa, and concerns it may enhance HIV transmission/acquisition (Galvin and Cohen, 2004; Curtis et al., 2020; Smith et al., 2020). This was reflected in our study screening process, which identified most progestin-only studies investigated the impact of DMPA on the vaginal microbiota. These studies were predominantly of women in sub-Saharan Africa, where DMPA usage coincides with high rates of HIV. Other possible detrimental effects following initiation of DMPA include epithelium thinning in the vagina, tissue inflammation and altered cell-mediated immune responses. In fact all of these effects increase a woman’s susceptibility to BV, HIV and other STIs (van de Wijgert et al., 2013; Murphy et al., 2014). Using previously published microbiome data from the CAPRISA-004 trial (Klatt et al., 2017), women with a Lactobacillus-dominant vaginal microbiota (primarily reflecting L. iners) who were using DMPA had a 3-fold increased risk of HIV acquisition (relative to women using Net-En or OCP) (Noël-Romas et al., 2020). In addition, higher serum-MPA concentrations were associated with evidence of inflammation in the vaginal mucosal fluid of Lactobacillus-dominated women (Noël-Romas et al., 2020). Interestingly, these effects of DMPA and serum-MPA were not observed in non-Lactobacillus-dominant women. The authors concluded that there is an interaction between the microbiome, hormonal contraceptives, and HIV susceptibility, demonstrating the importance of the both the vaginal microbiota and hormonal contraceptives in assessing HIV risk. Indeed, specific BV-associated bacteria themselves have been shown to enhance inflammatory pathways, which further complicates our understanding of these relationships (Dabee et al., 2021). The Evidence for Contraceptive Options in HIV Outcomes (ECHO) trial went on to assess if the risk of HIV differed with the use of three contraceptive methods – DMPA, LNG implant and Cu-IUD - but found no substantial difference in risk of HIV acquisition by contraceptive method used (Ahmed et al., 2019). The results from subsequent molecular analyses from the ECHO trial are pending and will be of great interest due to the concern regarding the possible impact of contraceptive practices on the risk of HIV acquisition and transmission. Clearly the relationship between progestin-only contraceptive use and the vaginal microbiota are more complex than for oestrogen-containing contraceptives, and require further investigation in the context of different populations and settings, as observed in the study by Yang et al. (2020).
Strengths and Limitations
Our systematic review identified a significant number of studies that reported on hormonal contraception-use on the vaginal microbiota in reproductive aged women (n=25). We restricted our inclusion criteria to studies that measured the vaginal microbiota by molecular methods to reduce the significant bias which can be introduced by culture-dependant methods and microscopy. These methods only identify organisms that are able to be cultivated or are limited by taxonomic resolution. Despite the differences in methods used, next-generation sequencing was the most common measurement recorded in this review (n=20). However, all molecular methods have different biases, from DNA extraction, to data analysis, which should be considered. For example, different approaches to the steps used in 16S rRNA gene sequencing pipelines (i.e. DNA extraction method, primer selection, variable region selection, 16S rRNA gene reference database etc) can result in different findings when applied to the same samples (Pollock et al., 2018; Balkus et al., 2019). Additionally, microarray and qPCR results are dependent on the panel of targets tested. These factors further limited our ability to compare findings across studies. This study had a number of additional limitations. In the reproductive-aged cohort, most studies were conducted in sub-Saharan Africa (n=12) and North America (n=8) and therefore the effects of hormonal contraceptives may not be generalizable to other populations. This was even more apparent in the post-menopausal cohort where three of the four studies included were conducted in Canada, with the fourth from China. The small number of studied population groups, and the diverse findings across studies limited further comparison of our findings. Our analysis of bias also identified several other limitations specific to each of the included studies (Table 4). There was a high degree of variation in how the studies were conducted. For example, the contraceptive types assessed, how long contraceptives were used for, if there was a wash-out period from previous exogenous steroid exposure, the sample size, and how the data were analysed. The comparator groups also varied and included the participants’ baseline specimens or specimens from patients using non-hormonal contraceptives such as condoms or a Cu-IUD. The use of a Cu-IUD as the control group may have overestimated the benefit of the hormonal contraceptive assessed, as Cu-IUD use itself has been associated with dysbiosis (Erol et al., 2014; Achilles et al., 2018). Notably, many studies had small sample sizes, and, in most cases, patients self-selected their exposure and self-reported adherence to use. Furthermore, the effects of exogenous sex steroid exposure were not adjusted for behavioural practices. Despite contacting authors to retrieve more information, several studies were unable to specify which hormonal contraception-types were included in their hormonal contraception-use group/s, which prevented our ability to include the studies in analyses stratified by sex steroid-type. Regardless, of the five studies which included unspecified hormonal contraceptive-types, four found a positive influence of their use on the vaginal microbiota. Finally, there are newer formulations of hormonal contraceptives and menopausal hormone therapy that were not captured in this review.
Conclusions
Access to contraceptives is an invaluable part of modern women’s healthcare, but until recently there was little consideration as to how this might affect the vaginal microbiota. With increasing use of hormonal contraceptives and menopausal hormone therapy globally, and improved understanding of how the microbiota may contribute to negative reproductive and gynaecological outcomes, it is important we understand the immediate and long-term impact of specific exogenous sex steroid use on the vaginal microbiota. Advances in high throughput DNA sequencing has made molecular analyses of the vaginal microbiota accessible and allowed us to gain more insight into the effects exogenous sex steroids may have on specific organisms, as well as its overall composition.
Our findings suggest that oestrogen-containing contraceptives and MHT may promote an optimal vaginal microbiota in some populations, which could have clinical applications, such as adjunctive therapy to improve management of BV and vaginitis (atrophic). While research is needed to support the use of exogenous-oestrogen as an adjunctive therapy, our data suggests it does not impact the composition of the vaginal microbiota in a detrimental way. The impact of progestin-only hormonal contraceptives was less consistent as there was equal evidence that they have either a negative or neutral impact on the vaginal microbiota. Clearly, more data is needed in order to confirm that DMPA and other progestin-only contraceptives are not contributing to adverse reproductive and sexual health outcomes. The molecular results from the ECHO trial are greatly anticipated and may provide some answers. Additional prospective studies from a wider range of populations that identify the underlying mechanisms by which progestin-only contraceptives alter the vaginal microbiota are also needed.
In summary, this review highlights the complex nature of the relationship between progestin-only contraceptives and the vaginal microbiome, and confirms the potential benefits of exogenous oestrogen in conferring a vaginal microbiota associated with optimal health outcomes for women.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding authors.
Author Contributions
LR, LV, and CB conceived and designed the study. LR and EP conducted the formal analysis, extracted all data, and interpreted the data, with support from LV and CB. GM provided advice around molecular technologies used. LM, GT, and DB provided additional data interpretation. CB, LV, GM, CF, and SG provided supervision. LR drafted the initial manuscript with supervision from LV, and EP, CB, CF, GM, SG, DB, GT, and LM. All authors contributed to the article and approved the submitted version.
Funding
EP is supported by an Australian Government Research Training Program Scholarship. CF, CB, and SG are supported by Australian NHMRC Leadership Investigator Grants (GNT1172900, GNT1173361, and APP1197951, respectively). GT was supported by NHMRC Senior Research Fellowship (GNT1117748) and NHMRC Project Grant (GNT1164982).
Conflict of Interest
GT is a co-inventor on patent application AU201501042 and United States Patent No. US 9,801,839 B2 claiming the anti-inflammatory effects of lactic acid. DB has attended advisory meetings and provided educational updates for clinicians for MSD and Bayer Healthcare as part of her role as Medical Director at FPNSW but has never received personal remuneration for these services.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Janneke van de Wijgert and Katie Lennard for providing additional information or stratified data.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2021.732423/full#supplementary-material
References
Achilles, S. L., Austin, M. N., Meyn, L. A., Mhlanga, F., Chirenje, Z. M., Hillier, S. L. (2018). Impact of Contraceptive Initiation on Vaginal Microbiota. Am. J. Obstet. Reprod. 218 (6), 622.e1–622.e10. doi: 10.1016/j.ajog.2018.02.017
Ahmed, K., Baeten, J. M., Beksinska, M., Bekker, L.-G., Bukusi, E. A., Donnell, D., et al. (2019). HIV Incidence Among Women Using Intramuscular Depot Medroxyprogesterone Acetate, a Copper Intrauterine Device, or a Levonorgestrel Implant for Contraception: A Randomised, Multicentre, Open-Label Trial. Lancet 394 (10195), 303–313. doi: 10.1016/S0140-6736(19)31288-7
Aldunate, M., Srbinovski, D., Hearps, A. C., Latham, C. F., Ramsland, P. A., Gugasyan, R., et al. (2015). Antimicrobial and Immune Modulatory Effects of Lactic Acid and Short Chain Fatty Acids Produced by Vaginal Microbiota Associated With Eubiosis and Bacterial Vaginosis. Front. Physiol. 6 (JUN). doi: 10.3389/fphys.2015.00164
Aldunate, M., Tyssen, D., Johnson, A., Zakir, T., Sonza, S., Moench, T., et al. (2013). Vaginal Concentrations of Lactic Acid Potently Inactivate HIV. J. Antimicro. Chemother. 68 (9), 2015–2025. doi: 10.1093/jac/dkt156
Anahtar, M. N., Gootenberg, D. B., Mitchell, C. M., Kwon, D. S. (2018). Cervicovaginal Microbiota and Reproductive Health: The Virtue of Simplicity. Cell Host Microbe 23 (2), 159–168. doi: 10.1016/j.chom.2018.01.013
Balkus, J., Proll, S., Holm, J., Srinivasan, S., Dinwiddie, D., van der Pol, L., et al. (2019). A Multi-Site Comparative Study to Understand Sources of Variability in Studies of the Vaginal Microbiota. Sex. Transm. Infect. 95, A262–A2A3. doi: 10.1136/sextrans-2019-sti.659
Balle, C., Konstantinus, I. N., Jaumdally, S. Z., Havyarimana, E., Lennard, K., Esra, R., et al. (2020). Hormonal Contraception Alters Vaginal Microbiota and Cytokines in South African Adolescents in a Randomized Trial. Nat. Commun. 11 (1), 5578. doi: 10.1038/s41467-020-19382-9
Bassis, C. M., Allsworth, J. E., Wahl, H. N., Couasnon, M. T., Sack, D., Bell, J. D. (2015). The Effects of Contraception on the Vaginal Microbiota. Sex. Transmit. Infect. 91 (Suppl 2), A116–A1A7. doi: 10.1136/sextrans-2015-052270.308
Borgdorff, H., van der Veer, C., Van Houdt, R., Alberts, C. J., De Vries, H. J., Bruisten, S. M., et al. (2017). The Association Between Ethnicity and Vaginal Microbiota Composition in Amsterdam, the Netherlands. PloS One 12 (7), e0181135. doi: 10.1371/journal.pone.0181135
Borgdorff, H., Verwijs, M. C., Wit, F. W. N. M., Tsivtsivadze, E., Ndayisaba, G. F., Verhelst, R., et al. (2015). The Impact of Hormonal Contraception and Pregnancy on Sexually Transmitted Infections and on Cervicovaginal Microbiota in African Sex Workers. Sex. Transm. Dis. 42 (3), 143–152. doi: 10.1097/OLQ.0000000000000245
Boskey, E. R., Cone, R. A., Whaley, K. J., Moench, T. R. (2001). Origins of Vaginal Acidity: High D/L Lactate Ratio is Consistent With Bacteria Being the Primary Source. Hum. Reprod. 16 (9), 1809–1813. doi: 10.1093/humrep/16.9.1809
Brooks, J. P., Edwards, D. J., Blithe, D. L., Fettweis, J. M., Serrano, M. G., Sheth, N. U., et al. (2017). Effects of Combined Oral Contraceptives, Depot Medroxyprogesterone Acetate and the Levonorgestrel-Releasing Intrauterine System on the Vaginal Microbiome. Contraception 95 (4), 405–413. doi: 10.1016/j.contraception.2016.11.006
Brotman, R. M., Klebanoff, M. A., Nansel, T. R., Yu, K. F., Andrews, W. W., Zhang, J., et al. (2010). Bacterial Vaginosis Assessed by Gram Stain and Diminished Colonization Resistance to Incident Gonococcal, Chlamydial, and Trichomonal Genital Infection. J. Infect. Dis. 202 (12), 1907–1915. doi: 10.1086/657320
Castelo-Branco, C., Cancelo, M. J., Villero, J., Nohales, F., Julia, M. D. (2005). Management of Post-Menopausal Vaginal Atrophy and Atrophic Vaginitis. Maturitas 52 (Suppl 1), S46–S52. doi: 10.1016/j.maturitas.2005.06.014
Chetwin, E., Manhanzva, M. T., Abrahams, A. G., Froissart, R., Gamieldien, H., Jaspan, H., et al. (2019). Antimicrobial and Inflammatory Properties of South African Clinical Lactobacillus Isolates and Vaginal Probiotics. Sci. Rep. 9 (1), 1917. doi: 10.1038/s41598-018-38253-4
Chollet, J. A., Carter, G., Meyn, L. A., Mermelstein, F., Balk, J. L. (2009). Efficacy and Safety of Vaginal Estriol and Progesterone in Postmenopausal Women With Atrophic Vaginitis. Menopause 16 (5), 978–983. doi: 10.1097/gme.0b013e3181a06c80
Clabaut, M., Boukerb, A. M., Mlouka, A. B., Suet, A., Tahrioui, A., Verdon, J., et al. (2021). Variability of the Response of Human Vaginal Lactobacillus Crispatus to 17β-Estradiol. Sci. Rep. 11 (1), 11533. doi: 10.1038/s41598-021-91017-5
Crucitti, T., Hardy, L., van de Wijgert, J., Agaba, S., Buyze, J., Kestelyn, E., et al. (2018). Contraceptive Rings Promote Vaginal Lactobacilli in a High Bacterial Vaginosis Prevalence Population: A Randomised, Open-Label Longitudinal Study in Rwandan Women. PloS One 13 (7), e0201003. doi: 10.1371/journal.pone.0201003
Curtis, K. M., Hannaford, P. C., Rodriguez, M. I., Chipato, T., Steyn, P. S., Kiarie, J. N. (2020). Hormonal Contraception and HIV Acquisition Among Women: An Updated Systematic Review. BMJ Sex. Reprod. Health 46 (1), 8–16. doi: 10.1136/bmjsrh-2019-200509
Dabee, S., Barnabas, S. L., Lennard, K. S., Jaumdally, S. Z., Gamieldien, H., Balle, C., et al. (2019). Defining Characteristics of Genital Health in South African Adolescent Girls and Young Women at High Risk for HIV Infection. PloS One 14 (4), e0213975. doi: 10.1371/journal.pone.0213975
Dabee, S., Passmore, J. A. S., Heffron, R., Jaspan, H. B. (2021). The Complex Link Between the Female Genital Microbiota, Genital Infections, and Inflammation. Infect. Immun. 89 (5), e00487-20. doi: 10.1128/IAI.00487-20
Dahn, A., Saunders, S., Hammond, J. A., Carter, D., Kirjavainen, P., Anukam, K., et al. (2008). Effect of Bacterial Vaginosis, Lactobacillus and Premarin Estrogen Replacement Therapy on Vaginal Gene Expression Changes. Microbes Infect. 10 (6), 620–627. doi: 10.1016/j.micinf.2008.02.007
Deese, J., Masson, L., Miller, W., Cohen, M., Morrison, C., Wang, M., et al. (2015). Injectable Progestin-Only Contraception Is Associated With Increased Levels of Pro-Inflammatory Cytokines in the Female Genital Tract. Am. J. Reprod. Immunol. 74 (4), 357–367. doi: 10.1111/aji.12415
Delgado-Diaz, D. J., Tyssen, D., Hayward, J. A., Gugasyan, R., Hearps, A. C., Tachedjian, G. (2020). Distinct Immune Responses Elicited From Cervicovaginal Epithelial Cells by Lactic Acid and Short Chain Fatty Acids Associated With Optimal and Non-Optimal Vaginal Microbiota. Front. Cell Infect. Micro. 9, 446. doi: 10.3389/fcimb.2019.00446
Devillard, E., Burton, J. P., Hammond, J. A., Lam, D., Reid, G. (2004). Novel Insight Into the Vaginal Microflora in Postmenopausal Women Under Hormone Replacement Therapy as Analyzed by PCR-Denaturing Gradient Gel Electrophoresis. Eu J. Obstet. Gynecol. Reprod. Biol. 117 (1), 76–81. doi: 10.1016/j.ejogrb.2004.02.001
Diem, S. J., Guthrie, K. A., Mitchell, C. M., Reed, S. D., Larson, J. C., Ensrud, K. E., et al. (2018). Effects of Vaginal Estradiol Tablets and Moisturizer on Menopause-Specific Quality of Life and Mood in Healthy Postmenopausal Women With Vaginal Symptoms: A Randomized Clinical Trial. Menopause 25 (10), 1086–1093. doi: 10.1097/GME.0000000000001131
Edwards, V. L., Smith, S. B., McComb, E. J., Tamarelle, J., Ma, B., Humphrys, M. S., et al. (2019). The Cervicovaginal Microbiota-Host Interaction Modulates Chlamydia Trachomatis Infection. mBio 10 (4), e01548-19. doi: 10.1128/mBio.01548-19
Erol, O., Simavli, S., Derbent, A. U., Ayrim, A., Kafali, H. (2014). The Impact of Copper-Containing and Levonorgestrel-Releasing Intrauterine Contraceptives on Cervicovaginal Cytology and Microbiological Flora: A Prospective Study. Eu J. Contracept. Reprod. Health Care 19 (3), 187–193. doi: 10.3109/13625187.2014.900532
Fredricks, D. N., Fiedler, T. L., Marrazzo, J. M. (2005). Molecular Identification of Bacteria Associated With Bacterial Vaginosis. New Engl. J. Med. 353 (18), 1899–1911. doi: 10.1056/NEJMoa043802
Galvin, S. R., Cohen, M. S. (2004). The Role of Sexually Transmitted Diseases in HIV Transmission. Nat. Rev. Microbiol. 2, 33+. doi: 10.1038/nrmicro794
Gautam, R., Borgdorff, H., Jespers, V., Francis, S. C., Verhelst, R., Mwaura, M., et al. (2015). Correlates of the Molecular Vaginal Microbiota Composition of African Women. BMC Infect. Dis. 15 (1), 86. doi: 10.1186/s12879-015-0831-1
Hearps, A. C., Tyssen, D., Srbinovski, D., Bayigga, L., Diaz, D. J. D., Aldunate, M., et al. (2017). Vaginal Lactic Acid Elicits an Anti-Inflammatory Response From Human Cervicovaginal Epithelial Cells and Inhibits Production of Pro-Inflammatory Mediators Associated With HIV Acquisition. Mucosal Immunol. 10 (6), 1480–1490. doi: 10.1038/mi.2017.27
Heinemann, C., Reid, G. (2005). Vaginal Microbial Diversity Among Postmenopausal Women With and Without Hormone Replacement Therapy. Can. J. Microbiol. 51 (9), 777–781. doi: 10.1139/w05-070
Hickey, M., Marino, J. L., Tachedjian, G. (2016). Critical Review: Mechanisms of HIV Transmission in Depo-Provera Users: The Likely Role of Hypoestrogenism. J. Acquir. Immune Defic. Syndr. 71 (1), 1–7. doi: 10.1097/qai.0000000000000805
Jacobson, J. C., Turok, D. K., Dermish, A. I., Nygaard, I. E., Settles, M. L. (2014). Vaginal Microbiome Changes With Levonorgestrel Intrauterine System Placement. Contraception 90 (2), 130–135. doi: 10.1016/j.contraception.2014.04.006
Klatt, N. R., Cheu, R., Birse, K., Zevin, A. S., Perner, M., Noël-Romas, L., et al. (2017). Vaginal Bacteria Modify HIV Tenofovir Microbicide Efficacy in African Women. Science 356 (6341), 938–945. doi: 10.1126/science.aai9383
Koumans, E. H., Markowitz, L. E., Berman, S. M., St. Louis, M. E. (1999). A Public Health Approach to Adverse Outcomes of Pregnancy Associated With Bacterial Vaginosis. Int. J. Gynecol. Obstet. 67 (SUPPL. 1), S29–S33. doi: 10.1016/S0020-7292(99)00136-8
Lennard, K., Dabee, S., Barnabas, S. L., Havyarimana, E., Blakney, A., Jaumdally, S. Z., et al. (2018). Microbial Composition Predicts Genital Tract Inflammation and Persistent Bacterial Vaginosis in South African Adolescent Females. Infect. Immun. 86 (1), e00410-17. doi: 10.1128/IAI.00410-17
Marconi, C., El-Zein, M., Ravel, J., Ma, B., Lima, M. D., Carvalho, N. S., et al. (2020). Characterization of the Vaginal Microbiome in Women of Reproductive Age From 5 Regions in Brazil. Sex. Transm. Dis. 47 (8), 562–569. doi: 10.1097/OLQ.0000000000001204
McKinnon, L. R., Achilles, S. L., Bradshaw, C. S., Burgener, A., Crucitti, T., Fredricks, D. N., et al. (2019). The Evolving Facets of Bacterial Vaginosis: Implications for HIV Transmission. AIDS Res. Hum. Retroviruses 35 (3), 219–228. doi: 10.1089/aid.2018.0304
Mirmonsef, P., Spear, G. T. (2014). The Barrier to HIV Transmission Provided by Genital Tract Lactobacillus Colonization. Am. J. Reprod. Immunol. 71 (6), 531–536. doi: 10.1111/aji.12232
Mitchell, C. M., Srinivasan, S., Zhan, X., Wu, M. C., Reed, S. D., Guthrie, K. A., et al. (2017). Vaginal Microbiota and Genitourinary Menopausal Symptoms: A Cross-Sectional Analysis. Menopause 24 (10), 1160–1166. doi: 10.1097/GME.0000000000000904
Murphy, K., Irvin, S. C., Herold, B. C. (2014). Research Gaps in Defining the Biological Link Between HIV Risk and Hormonal Contraception. Am. J. Reprod. Immunol. 72 (2), 228–235. doi: 10.1111/aji.12209
Noël-Romas, L., Perner, M., Molatlhegi, R., Zuend, C. F., Mabhula, A., Hoger, S., et al. (2020). Vaginal Microbiome-Hormonal Contraceptive Interactions Associate With the Mucosal Proteome and HIV Acquisition. PloS Pathog. 16 (12), e1009097. doi: 10.1371/journal.ppat.1009097
Nunn, K. L., Forney, L. J. (2016). Unraveling the Dynamics of the Human Vaginal Microbiome. Yale J. Biol. Med. 89 (3), 331–337.
O’Hanlon, D. E., Moench, T. R., Cone, R. A. (2011). In Vaginal Fluid, Bacteria Associated With Bacterial Vaginosis Can be Suppressed With Lactic Acid But Not Hydrogen Peroxide. BMC Infect. Dis. 11, 200. doi: 10.1186/1471-2334-11-200
O’Hanlon, D. E., Moench, T. R., Cone, R. A. (2013). Vaginal pH and Microbicidal Lactic Acid When Lactobacilli Dominate the Microbiota. PloS One 8 (11), e80074. doi: 10.1371/journal.pone.0080074
Onywera, H., Williamson, A. L., Mbulawa, Z. Z. A., Coetzee, D., Meiring, T. L. (2019). Factors Associated With the Composition and Diversity of the Cervical Microbiota of Reproductive-Age Black South African Women: A Retrospective Cross-Sectional Study. PeerJ 2019 (8), e7488. doi: 10.7717/peerj.7488
Page, M. J., McKenzie, J. E., Bossuyt, P. M., Boutron, I., Hoffmann, T. C., Mulrow, C. D., et al. (2021). The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 372, n71. doi: 10.1136/bmj.n71
Petrova, M. I., Lievens, E., Malik, S., Imholz, N., Lebeer, S. (2015). Lactobacillus Species as Biomarkers and Agents That Can Promote Various Aspects of Vaginal Health. Front. Physiol. 6 (MAR). doi: 10.3389/fphys.2015.00081
Petrova, M. I., Reid, G., Vaneechoutte, M., Lebeer, S. (2017). Lactobacillus Iners: Friend or Foe? Trends Microbiol. 25 (3), 182–191. doi: 10.1016/j.tim.2016.11.007
Plummer, E. L., Bradshaw, C. S., Doyle, M., Fairley, C. K., Murray, G. L., Bateson, D., et al. (2021). Lactic Acid-Containing Products for Bacterial Vaginosis and Their Impact on the Vaginal Microbiota: A Systematic Review. PloS One 16, e0246953. doi: 10.1371/journal.pone.0246953
Polis, C. B., Curtis, K. M. (2013). Use of Hormonal Contraceptives and HIV Acquisition in Women: A Systematic Review of the Epidemiological Evidence. Lancet Infect. Dis. 13 (9), 797–808. doi: 10.1016/S1473-3099(13)70155-5
Pollock, J., Glendinning, L., Wisedchanwet, T., Watson, M. (2018). The Madness of Microbiome: Attempting to Find Consensus “Best Practice” for 16S Microbiome Studies. App. Enviro. Microbiol. 84 (7), e02627-17. doi: 10.1128/AEM.02627-17
Pyra, M., Srinivasan, S., Fredricks, D., Kimani, J., Schwebke, J., Anzala, O., et al. (2016). Associations Between Hormonal Contraceptive Methods and the Vaginal Microbiota. Am. J. Obstet. Gynecol. 215 (6), S815–S8S6. doi: 10.1016/j.ajog.2016.09.005
Ratten, L. K., Plummer, E. L., Murray, G. L., Danielewski, J., Fairley, C. K., Garland, S. M., et al. (2021). Sex is Associated With the Persistence of Non-Optimal Vaginal Microbiota Following Treatment for Bacterial Vaginosis: A Prospective Cohort Study. BJOG.: Int. J. Obstetrics. Gynaecol. 128 (4), 756–767. doi: 10.1111/1471-0528.16430
Ravel, J., Gajer, P., Abdo, Z., Schneider, G. M., Koenig, S. S. K., McCulle, S. L., et al. (2011). Vaginal Microbiome of Reproductive-Age Women. Proc. Natl. Acad. Sci. U.S.A. 108 (Suppl 1), 4680–4687. doi: 10.1073/pnas.1002611107
Scott, V. C. S., Thum, L. W., Ackerman, J. E., Khalique, M. U., Eilber, K. S., Anger, J. T., et al. (2019). Temporal Dynamics of the Genitourinary Microbiome. Neurourol. Urodynamics. 38 (Supplement 1), S9–S10. doi: 10.1002/nau.23949
Shen, J., Song, N., Williams, C. J., Brown, C. J., Yan, Z., Xu, C., et al. (2016). Effects of Low Dose Estrogen Therapy on the Vaginal Microbiomes of Women With Atrophic Vaginitis. Sci. Rep. 6, 24380. doi: 10.1038/srep24380
Smith, J. A., Beacroft, L., Abdullah, F., Buthelezi, B., Makua, M., Morroni, C., et al. (2020). Responding to the ECHO Trial Results: Modelling the Potential Impact of Changing Contraceptive Method Mix on HIV and Reproductive Health in South Africa. J. Int. AIDS Soc. 23 (10), e25620. doi: 10.1002/jia2.25620
Song, S. D., Acharya, K. D., Zhu, J. E., Deveney, C. M., Walther-Antonio, M. R. S., Tetel, M. J., et al. (2020). Daily Vaginal Microbiota Fluctuations Associated With Natural Hormonal Cycle, Contraceptives, Diet, and Exercise. mSphere 5 (4), e00593-20. doi: 10.1128/mSphere.00593-20
Tachedjian, G., Aldunate, M., Bradshaw, C. S., Cone, R. A. (2017). The Role of Lactic Acid Production by Probiotic Lactobacillus Species in Vaginal Health. Res. Microbiol. 168 (9–10), 782–792. doi: 10.1016/j.resmic.2017.04.001
The 2020 Genitourinary Syndrome of Menopause Position Statement of The North American Menopause Society. (2020). Menopause 27 (9), 976–992. doi: 10.1097/GME.0000000000001609
Thurman, A. R., Schwartz, J. L., Ravel, J., Gajer, P., Marzinke, M. A., Yousefieh, N., et al. (2019). Vaginal Microbiota and Mucosal Pharmacokinetics of Tenofovir in Healthy Women Using Tenofovir and Tenofovir/Levonorgestrel Vaginal Rings. PloS One 14 (5), e0199778. doi: 10.1371/journal.pone.0217229
Tuddenham, S., Ghanem, K. G., Caulfield, L. E., Rovner, A. J., Robinson, C., Shivakoti, R., et al. (2019a). Associations Between Dietary Micronutrient Intake and Molecular-Bacterial Vaginosis. Reprod. Health 16 (1), 151. doi: 10.1186/s12978-019-0814-6
Tuddenham, S., Ghanem, K., Gajer, P., Robinson, C., Ravel, J., Brotman, R. (2019b). The Effect of Hormonal Contraception on the Vaginal Microbiota Over 2 Years. Sex. Transm. Infect. 95, A263–A2A4. doi: 10.1136/sextrans-2019-sti.662
Tyssen, D., Wang, Y. Y., Hayward, J. A., Agius, P. A., DeLong, K., Aldunate, M., et al. (2018). Anti-HIV-1 Activity of Lactic Acid in Human Cervicovaginal Fluid. mSphere 3 (4), e00055-18. doi: 10.1128/mSphere.00055-18
United Nations, Department of Economic and Social Affairs, Population Division. (2019). Contraceptive Use by Method 2019: Data Booklet (ST/ESA/SER.A/435).
Van Der Veer, C., Bruisten, S. M., Van Houdt, R., Matser, A. A., Tachedjian, G., Van De Wijgert, J. H. H. M., et al. (2019). Effects of an Over-the-Counter Lactic-Acid Containing Intra-Vaginal Douching Product on the Vaginal Microbiota. BMC Microbiol. 19 (1). doi: 10.1186/s12866-019-1545-0
van der Veer, C., Hertzberger, R. Y., Bruisten, S. M., Tytgat, H. L. P., Swanenburg, J., de Kat Angelino-Bart, A., et al. (2019). Comparative Genomics of Human Lactobacillus Crispatus Isolates Reveals Genes for Glycosylation and Glycogen Degradation: Implications for In Vivo Dominance of the Vaginal Microbiota. Microbiome 7 (1), 49. doi: 10.1186/s40168-019-0667-9
van de Wijgert, J. H. H. M., Gill, A. C., Chikandiwa, A., Verwijs, M. C., Kelly, H. A., Omar, T., et al. (2020). Human Papillomavirus Infection and Cervical Dysplasia in HIV-Positive Women: Potential Role of the Vaginal Microbiota. AIDS (London England) 34 (1), 115–125. doi: 10.1097/QAD.0000000000002381
van de Wijgert, J. H., Verwijs, M. C., Turner, A. N., Morrison, C. S. (2013). Hormonal Contraception Decreases Bacterial Vaginosis But Oral Contraception may Increase Candidiasis: Implications for HIV Transmission. AIDS (London England) 27 (13), 2141–2153. doi: 10.1097/QAD.0b013e32836290b6
Vodstrcil, L. A., Hocking, J. S., Law, M., Walker, S., Tabrizi, S. N., Fairley, C. K., et al. (2013). Hormonal Contraception Is Associated With a Reduced Risk of Bacterial Vaginosis: A Systematic Review and Meta-Analysis. PloS One 8 (9), e73055. doi: 10.1371/journal.pone.0073055
Vodstrcil, L. A., Plummer, M. E., Fairley, C. K., Tachedjian, G., Law, M. G., Hocking, J. S., et al. (2019). Combined Oral Contraceptive Pill-Exposure Alone Does Not Reduce the Risk of Bacterial Vaginosis Recurrence in a Pilot Randomised Controlled Trial. Sci. Rep. 9 (1), 3555. doi: 10.1038/s41598-019-39879-8
Wessels, J. M., Lajoie, J., Cooper, M. I. J. H., Omollo, K., Felker, A. M., Vitali, D., et al. (2019). Medroxyprogesterone Acetate Alters the Vaginal Microbiota and Microenvironment in Women and Increases Susceptibility to HIV-1 in Humanized Mice. Dis. Models Mech. 12 (10), dmm039669. doi: 10.1242/dmm.039669
Whitney, B. M., Guthrie, B. L., Srinivasan, S., Tapia, K., Muriuki, E. M., Chohan, B. H., et al. (2020). Changes in Key Vaginal Bacteria Among Postpartum African Women Initiating Intramuscular Depot-Medroxyprogesterone Acetate. PloS One 15 (3), e0229586. doi: 10.1371/journal.pone.0229586
Wiesenfeld, H. C., Hillier, S. L., Krohn, M. A., Amortegui, A. J., Heine, R. P., Landers, D. V., et al. (2002). Lower Genital Tract Infection and Endometritis: Insight Into Subclinical Pelvic Inflammatory Disease. Obstetrics. Gynecol. 100 (3), 456–463. doi: 10.1097/00006250-200209000-00011
Witkin, S. S. (2018). Limitations of Treating Pregnant Women Based Solely on Vaginal Ph. BJOG.: Int. J. Obstetrics. Gynaecol. 125 (12), 1610. doi: 10.1111/1471-0528.15281
Witkin, S. S., Mendes-Soares, H., Linhares, I. M., Jayaram, A., Ledger, W. J., Forney, L. J. (2013). Influence of Vaginal Bacteria and D- and L-Lactic Acid Isomers on Vaginal Extracellular Matrix Metalloproteinase Inducer: Implications for Protection Against Upper Genital Tract Infections. mBio 4 (4), e00460-13. doi: 10.1128/mBio.00460-13
Yang, L., Hao, Y., Hu, J., Kelly, D., Li, H., Brown, S., et al. (2019). Differential Effects of Depot Medroxyprogesterone Acetate Administration on Vaginal Microbiome in Hispanic White and Black Women. Emerg. Microbes Infect. 8 (1), 197–210. doi: 10.1080/22221751.2018.1563458
Yang, S., Reid, G., Challis, J. R. G., Gloor, G. B., Asztalos, E., Money, D., et al. (2020). Effect of Oral Probiotic Lactobacillus Rhamnosus GR-1 and Lactobacillus Reuteri RC-14 on the Vaginal Microbiota, Cytokines and Chemokines in Pregnant Women. Nutrients 12 (2), 368–385. doi: 10.3390/nu12020368
Keywords: vaginal microbiota, hormonal contraceptives, menopausal hormone therapy, lactobacillus, Gardnerella vaginalis, oestrogen, progesterone, progestin
Citation: Ratten LK, Plummer EL, Bradshaw CS, Fairley CK, Murray GL, Garland SM, Bateson D, Tachedjian G, Masson L and Vodstrcil LA (2021) The Effect of Exogenous Sex Steroids on the Vaginal Microbiota: A Systematic Review. Front. Cell. Infect. Microbiol. 11:732423. doi: 10.3389/fcimb.2021.732423
Received: 29 June 2021; Accepted: 13 October 2021;
Published: 12 November 2021.
Edited by:
Maria De Lourdes Pereira, University of Aveiro, PortugalReviewed by:
Sonia M. Rodrigues Oliveira, University of Aveiro, PortugalTooba Mahboob, University of Malaya, Malaysia
Copyright © 2021 Ratten, Plummer, Bradshaw, Fairley, Murray, Garland, Bateson, Tachedjian, Masson and Vodstrcil. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Larissa K. Ratten, bGFyaXNzYS5yYXR0ZW5AbW9uYXNoLmVkdQ==; Lenka A. Vodstrcil, bGVua2Eudm9kc3RyY2lsQG1vbmFzaC5lZHU=