 Arturo Muñoz-Calderón1
Arturo Muñoz-Calderón1 Zoraida Díaz-Bello2
Zoraida Díaz-Bello2 Belkisyolé Alarcón de Noya2
Belkisyolé Alarcón de Noya2 Oscar O. Noya-González2
Oscar O. Noya-González2 Alejandro G. Schijman1*
Alejandro G. Schijman1*- 1Laboratorio de Biología Molecular de la Enfermedad de Chagas, Instituto de Investigaciones en Ingeniería Genética y Biología Molecular (INGEBI), Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Buenos Aires, Argentina
- 2Instituto de Medicina Tropical “Dr Félix Pifano”, Universidad Central de Venezuela, Caracas, Venezuela
We aimed to characterize the genetic constitution of natural T. cruzi populations involved in an Oral Chagas Disease (OCD) outbreak at a rural school of the community of Chichiriviche de la Costa, Venezuela, which affected patients did not respond to the etiological treatment. Peripheral blood samples and/or hemocultures were obtained from twenty-nine OCD patients at time of diagnosis or along nine years of Post-treatment (Tx) follow-up. The IgG serology, T. cruzi discrete typing units (DTU), satellite DNA-qPCR parasitic loads, and minicircle signatures were determined at Pre-Tx and after Tx. The serological titles and parasitic loads changed after treatment, with a significant decrease of IgG titers (Spearman’s r value= -0.961) and median parasite loads from 2.869 [IQR = 2.113 to 3.720] to 0.105 [IQR = -1.147 to 1.761] log10 par eq. /mL at Pre-Tx and Post-Tx, respectively, suggesting infection evolution from acute to chronic phase, without seroconversion or parasitological eradication, which was indicative of treatment failure. All patients were infected with T. cruzi DTU I populations. At Pre-Tx their median Jaccard genetic distances were 0.775 [IQR = 0.708 to 0.882], decreasing in genetic variability towards the end of follow-up (Mann-Whitney U test p= 0.0031). Interestingly, no Post-Tx minicircle signature was identical to its Pre-Tx counterpart population in a same patient, revealing selection of parasite subpopulations between the primary infection and Post-Tx. The parasitic populations isolated from hemocultures showed a lower number of bands in the minicircle signatures with respect to the signatures obtained directly from the patients’ blood samples, demonstrating a process of parasitic selection and reduction of the population variability that initially infected the patients. Decrease of parasitic loads after treatment as well as Pre- and Post-Tx intra-TcI diversity might be a consequence of both, natural evolution of the acute infection to the chronic phase and persistence of refractory populations due to Tx selection.
Introduction
Chagas disease (CD), caused by Trypanosoma cruzi affects mostly populations living in poor housing conditions in Latin America (Pereiro, 2019). Although initially considered transmitted mostly by triatomine bugs, the parasite can be transmitted by other routes, one of the most important in recent decades being oral transmission, as a result of contamination of food with feces of wild triatomines or secretions from reservoirs in endemic regions (Rueda et al., 2014; Alarcón de Noya et al., 2015).
Venezuela is the third country with oral Chagas disease (OCD) reports since 2007, and the first one with the most numerous OCD micro-epidemics (Alarcón de Noya et al., 2015). The first outbreaks were associated with consumption of guava (Psidium guajava) juice in an urban school in the Chacao Municipality (Caracas, Capital District) in 2007 (Alarcon de Noya et al., 2010) and the second in a rural school in the community of Chichiriviche de la Costa (Vargas State) (Alarcon de Noya et al., 2016) in 2009, with varied degree of disease severity and mortality (Alarcón de Noya et al., 2015; Alarcon de Noya et al., 2016).
In Chichiriviche de la Costa, a small tourist town nestled between mountains and on the shores of the Caribbean Sea, located on the north-central coast of Venezuela, there were simultaneous cases of fever and myocarditis in students, teachers, and administrative staff of the local school (10°31′53.97″N - 67°15′36.02″W). These clinical findings led to the serological screening of specific anti-T. cruzi IgM and IgG reactivities to 441 people, resulting in 89 infected people and mortality associated with acute T. cruzi infection in 5.6% of them (Alarcón de Noya and Martinez, 2009; Alarcon de Noya et al., 2016).
In these patients, the outcome observed by serological, parasitological and PCR-based monitoring two years after treatment with Benznidazole (Bnz) was disappointing. Around 70% of patients still had positive lytic antibodies; some presented anti-T.cruzi IgG antibody titers and positive T. cruzi PCR findings (Alarcón de Noya et al., 2011). Considering that Bnz is highly effective in the acute phase and during childhood in other endemic regions where other parasite DTUs prevail (Bianchi et al., 2015; Moscatelli et al., 2019), these data suggested therapeutic failure and consequently a second treatment was administered to people with persistence of lytic antibodies and/or positive PCR results, however without outcome improvement (Garcia-Bournissen, 2019; Molina-Morant et al., 2020).
It is crucial to carry out genetic studies on parasitic populations involved in OCD aiming to seek an explanation for the differences in Bnz efficacy with respect to other epidemiological settings. In assessing drug efficacy, in vitro susceptibility tests carried out in parasite isolates obtained at pre-treatment (Pre-Tx) and Post-treatment (Post-Tx) from these outbreaks displayed clones naturally less susceptible to Bnz and Nfx (Muñoz-Calderón et al., 2012; Muñoz-Calderón et al., 2019). Indeed, the genetic polymorphism of T. cruzi may be involved in its susceptibility to anti-parasitic drugs. However, genetic studies of natural parasite populations in CD patients exhibiting treatment failure have not been done so far. In this context, peripheral blood samples and hemocultures obtained from OCD patients at Chichiriviche de la Costa outbreak were analyzed at time of diagnosis and after Tx, attempting to detect and characterize fluctuations of T. cruzi genetic diversity and parasitic loads along the follow-up.
Methods
Ethics Statement
The study was approved by the Ethical Review Board of the Instituto de Medicina Tropical “Dr Félix Pifano”-Universidad Central de Venezuela, Caracas, Venezuela (CEC-IMT 019/2010 - December 10, 2010); following the principles expressed in the Declaration of Helsinki.
Written informed consent forms were signed by each participant or from their legal guardians (minor subjects were included). Samples were anonymized before being processed.
Patients and Samples
The study population consisted of students, teachers, school workers and outsiders involved in the preparation or transportation of food consumed in schools, and anyone considered as a potential at-risk “school contact”. The participants eligible for the study were 29 patients who presented positive anti-T. cruzi IgG antibody titers or kinetoplastid DNA-PCR amplification after nine years of Post-Tx follow-up, according to data provided by the Instituto de Medicina Tropical “Dr Félix Pifano”- Universidad Central de Venezuela, Caracas, Venezuela.
Table 1 shows epidemiological features of the study patients from the Chichiriviche de la Costa outbreak, hemocultures obtained, number of blood samples analyzed and Post-Tx time period of each sample. Additionally, the distribution of the number of samples for each of the variables studied in this work (Pre-treatment and Post-treatment points), is outlined in Figure 1.
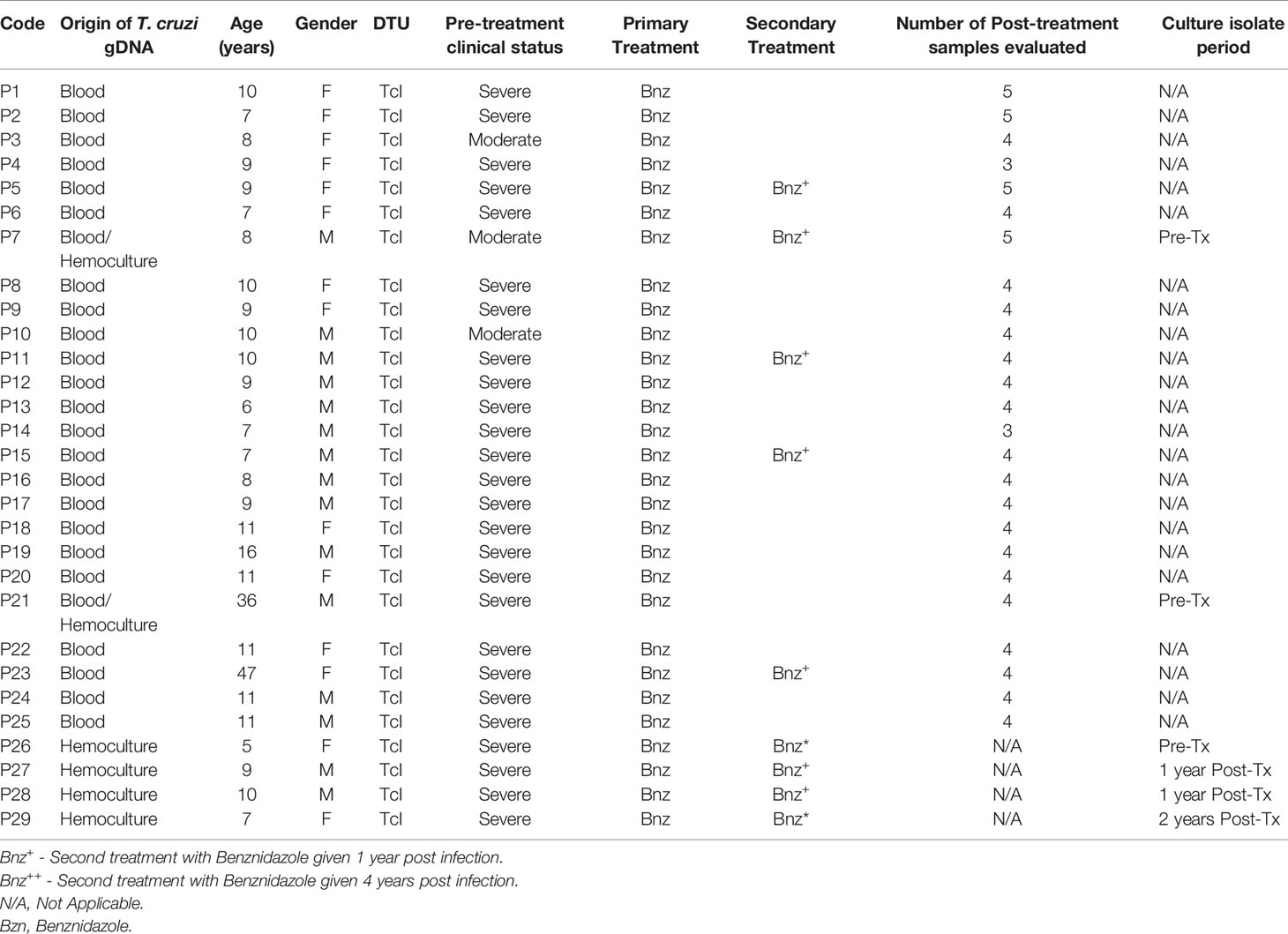
Table 1 Description of the Oral Chagas Disease patients´ cohort with Pre and Post treatment follow-up samples for 9 years.
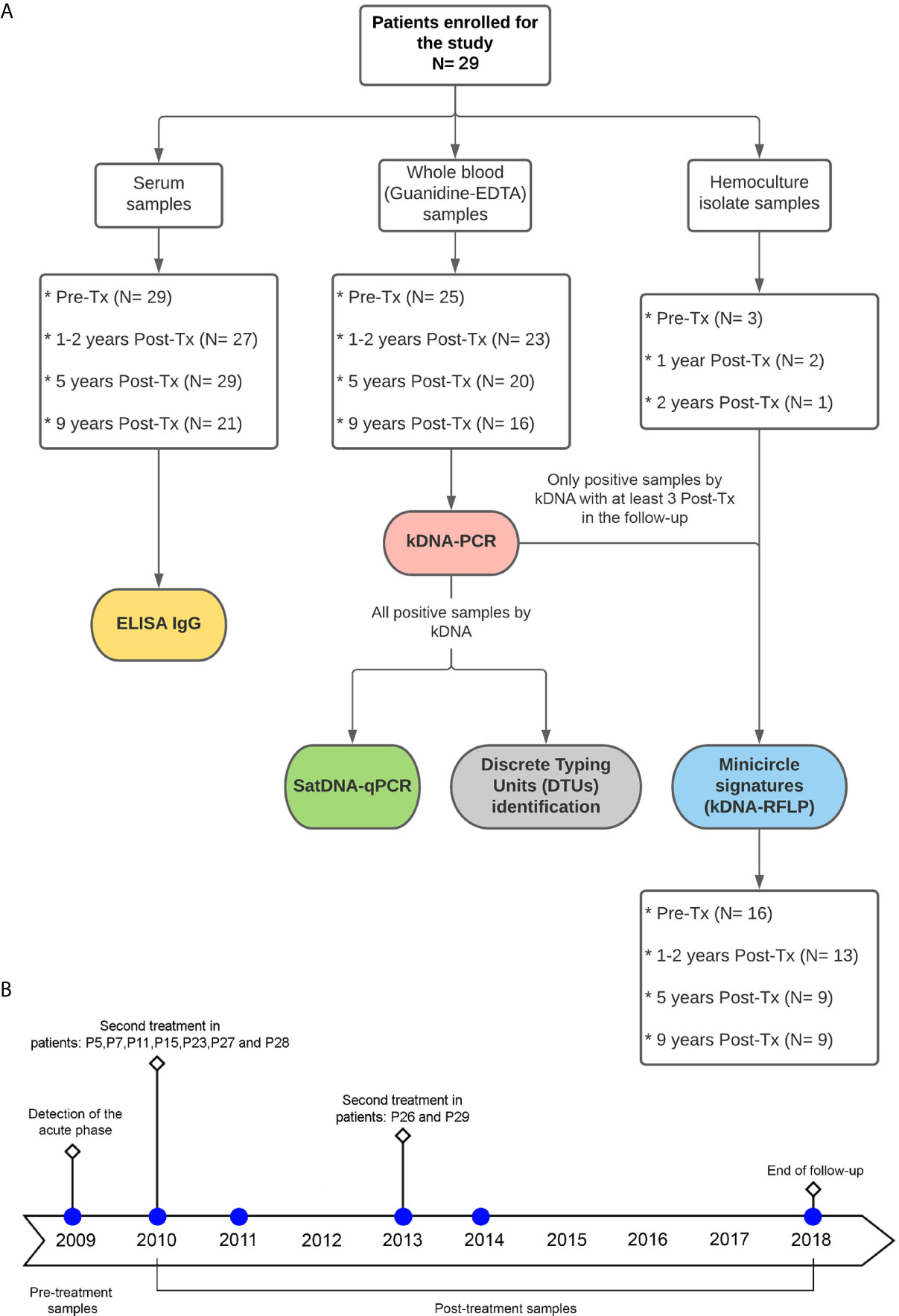
Figure 1 (A) Flowchart for the distribution of study patients´ samples and type of analysis performed. Tx: Treatment. (B) timeline of blood sample collection at follow-up time.
Clinical Classification
Patients with acute OCD were clinically classified with i) moderate symptoms whose clinical situation allowed them to continue with work activities (facial edema; dyspnea; abdominal pain; headache; arthralgia; myalgia; asthenia; drowsiness; headache; intraocular pain; rash; echocardiogram (ECHO): Supraventricular hypertrophy), and ii) severe symptoms who had to be hospitalized or deceased (ECHO: pericardial effusion or myocarditis; electrocardiogram (EKG): supraventricular arrhythmia or supraventricular hypertrophy; adenomegaly; cardiomegaly; tachycardia).
Antiparasitic Treatment
Patients were treated with Bnz (6 mg/kg/day) during 60 continuous days in three daily doses (Alarcón de Noya et al., 2011; Alarcon de Noya et al., 2016). During follow-up, nine treated patients who remained with evidence of parasite persistence (high values in anti-T. cruzi IgG titers, presence of lytic antibodies, and/or persistence of positive PCR) received a new course of treatment with Bnz (Amato Neto, 1999).
Laboratory Diagnosis
At each visit, a single peripheral blood sample was withdrawn by venipuncture. Five milliliters were dedicated to serum collection to perform IgG-based serological analyses. The remaining volume was used for molecular biology studies.
Immunoenzymatic Assay
An in-house immunoenzymatic assay (ELISA) was performed using a delipidized antigenic lysate obtained from the epimastigote stage of the T. cruzi “PM” strain (TcI) and processed using Maxisorp plates. ELISA tests involving anti-human IgG alkaline phosphatase conjugates were carried out simultaneously in all sera. ELISA IgG cut-off line was defined as an optical density (OD) value equal to 0.200 to determine a positive sample (Alarcón de Noya et al., 2011).
Preparation of Samples for Molecular Biology Studies
Five mL of patients’ peripheral blood was mixed with an equal volume of 6M Guanidine HCl/0.2 mM EDTA, pH 8.00 (GEB) and boiled according to Diaz-Bello et al. (2008). The GEB mixture was stored at 4°C; DNA was extracted from 300 µL aliquots using the High Pure PCR Template Preparation kit (Roche Diagnostics, Indianapolis, IN) and eluted in 100 µL elution buffer (Duffy et al., 2013).
Parasite Cultures
Epimastigote forms obtained from patients’ hemocultures and four TcI reference strains (Dm28c, SilvioX10, gal4, and CMA) were cultured in LIT medium (supplemented with 5% bovine fetal serum) and sub-cultured every 15 days during the exponential growth phase.
DNA Extraction and Quantitative Analysis
Parasites were collected at logarithmic phase and centrifuged at 3000 × g. The High Pure PCR Template Preparation kit (Roche Diagnostic, Indianapolis, IN) was used for DNA extraction following the manufacturer’s recommendations. The genomic DNA was then preserved at 4°C until use. DNA purity and concentration were determined in a Nanodrop ND-1000 (NanoDrop Technologies, Houston, TX, USA) at 260/280 nm wavelengths.
kDNA-PCR Amplification
A 330 bp sequence belonging to the hypervariable region of T. cruzi kDNA was amplified as reported (Schijman et al., 2011). Briefly, the master mix was composed by 1X Taq platinum amplification buffer, 200 µM dNTPs, 3 mM MgCl2 solution, 1.5 U Taq Platinum (Invitrogen, Brazil), 10 µM kDNA specific primers 121 (AAATAATGTACGGGKGAGATGCATGA) and 122 (GGTTCGATTGGGGTTGGTGTAATATA), 5 µl of template DNA, and a quantity of water sufficient to give a final volume of 50 µl. Cycling parameters were one step of 3 min denaturation at 94°C; 2 cycles of 1 min at 97.5°C, 2 min at 64°C; 33 cycles of 1 min at 94°C, 1 min at 62°C and one final extension step of 10 min at 72°C. The kDNA-PCR products were analyzed in 2% agarose gels stained with ethidium bromide.
Estimation of Parasitic Loads
A standardized duplex quantitative real-time PCR (qPCR) targeted to T. cruzi Satellite (Sat) DNA and Internal Amplification control (IAC) was used, under the reported conditions (Duffy et al., 2013). Standard curves were plotted with 1/10 serial dilutions of total DNA obtained from the same stock of a GEB seronegative sample spiked with 1x105 par.eq./mL of Silvio X10 (TcI) cultured epimastigotes. The reportable linear range of this qPCR was 1 to 6 log10par eq./mL for TcI-infected samples (Duffy et al., 2013).
Identification of Trypanosoma cruzi Discrete Typing Units (DTUs)
A Real-Time PCR-based algorithm for identification of T. cruzi DTUs was done by means of amplification of spliced-leader genes, 24Sα rRNA, 18Sα rRNA and COII genomic markers using TaqMan probes, as described (Cura et al., 2015).
Minicircle Signatures
To identify minicircle signatures (Ms) of bloodstream parasite populations, kDNA-PCR samples were studied. The 330 bp purified amplicon was subjected to HindFI, RsaI, and MspI digestions as reported (Burgos et al., 2010). Signatures were visualized and digitized using a Syngene ultraviolet light transilluminator (Synoptics Ltd., Cambridge, UK), at 498 nm. The sizes of the bands of each signature were determined by comparison with molecular weight markers included in each electrophoresis, using the GelAnalyzer v2010a program (http://www.gelanalyzer.com). To assess the reproducibility of the Ms, three independent kDNA-PCR experiments from a same DNA preparation were performed, and each product was digested on triplicate.
Genetic Distance Determination
Binary matrices were created from each Ms. Only sharp bands resolved by gel electrophoresis were used in the analysis. Visible amplified bands were scored as “1”, whereas the absence of bands of the same molecular weight was scored as “0”. All samples from a same patient were analyzed in a same gel. The degree of similarity, i.e. frequency of co-occurrence, between clinical samples was measured using the Jaccard’s coefficient (JC) between Pre-Tx and Post-Tx samples for each patient.
In order to estimate the overall variability of the Pre-Tx parasite populations using as marker the minicircle signatures, a median value of the Jaccard genetic distances among the pre-Tx samples was determined, and the profiles obtained from each sample at the different Post-Tx points were compared against that pre-Tx median value.
In addition, the minicircle profiles obtained from the hemocultures were compared among them, in order to obtain an overall JC sight of the cultured parasite strains and compare their JCs with those obtained among laboratory reference clones belonging to Tc I.
Statistical Analysis
A data analysis was performed on the follow-up panel to evaluate the change in parasite loads, IgG antibody values, Jaccard’s coefficient, and/or the clinical classification of the patients. The graphic comparisons between the different variables obtained from each patient: IgG serology (optical density), parasite load (Par.eq/mL) and JC, were carried out with a Z score data transformation. The continuous measurements were expressed and plotted as the median and interquartile range of 25–75%, whereas dichotomous variables were expressed and plotted as a percentage.
For all variables, the normal distributions were evaluated using the Shapiro-Wilks test. The Spearman’s rank correlation coefficient was calculated to assess correlations among the variables under study (parasitemia levels, ELISA IgG values and patients’ clinical classification). Correlation is stronger as the value of r approaches 1 for positive correlations or -1 for negative correlations. To evaluate the differences between Pre-Tx and Post-Tx follow-up points, the Mann-Whitney U test was used.
All analyses were performed with the RStudio Team software (2020), and calculations were determined with a p-value of <0.05 for significant statistical differences.
Results
Peripheral blood samples and/or hemocultures obtained from 29 OCD patients at Pre-Tx and/or at different Post-Tx visits were included in the study (Table 1). From each patient (except P26 to P29, in which only hemocultures were obtained), one Pre-Tx sample and Post-Tx samples collected at at least two different visits were analyzed by serological and molecular tools.
IgG-based Serological Findings and Parasitic Loads
All study patients were seropositive at Pre-Tx. The Post-Tx accumulated seropositivity was 82.26% (n = 21) (Figure 2A). After nine years of Post-Tx, seropositivity was 71.43% (n=15), with a median in optical density of 0.320 [IQR = 0.174 to 0.507]. Although there was not seroconversion, a trend towards antibodies titers decrease was observed (Figure 2C).
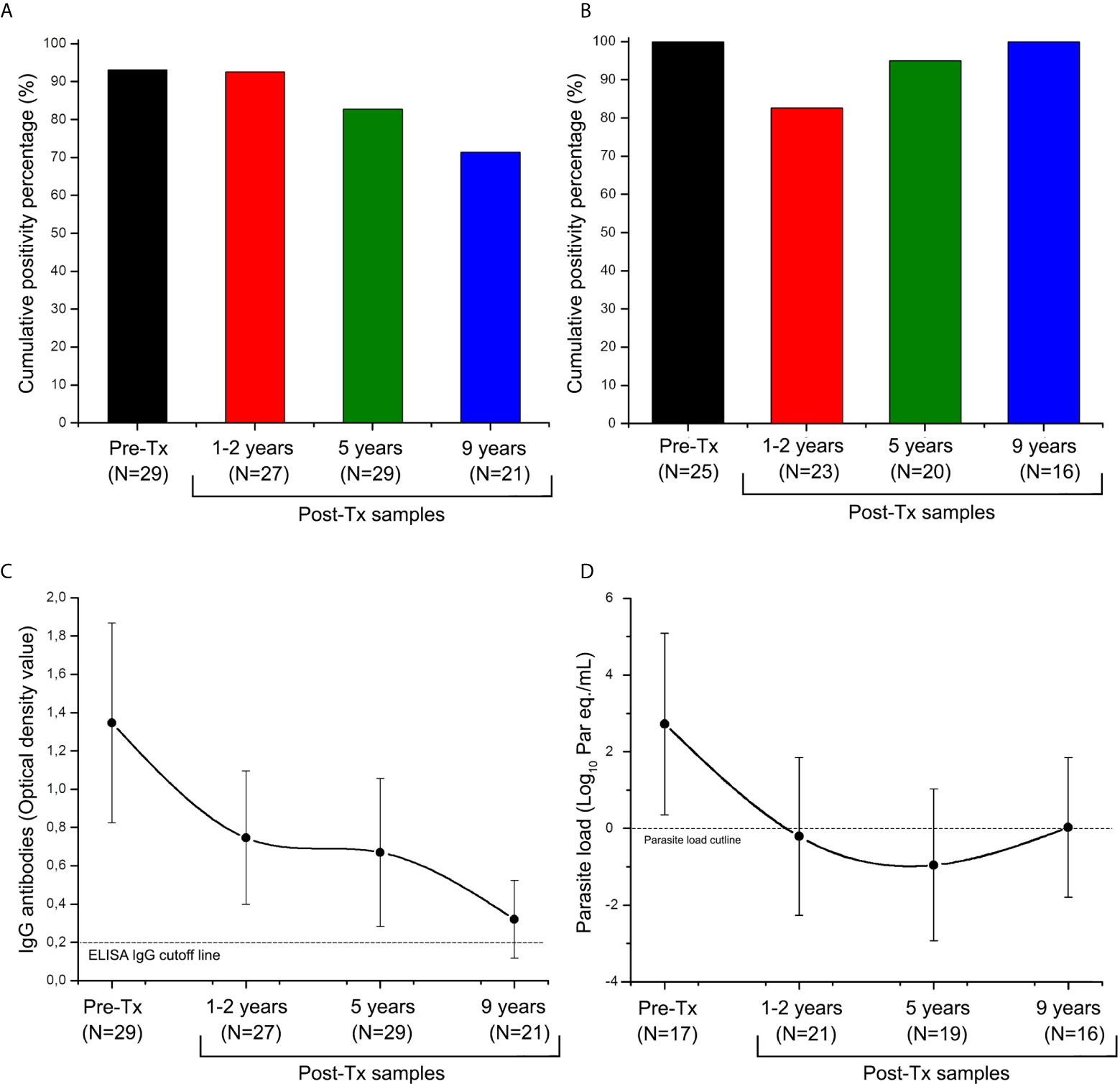
Figure 2 Distribution of anti-Trypanosoma cruzi ELISA-IgG reactivity and SatDNA-qPCR positivity during the follow-up of patients of the Chichiriviche de la Costa oral Chagas disease outbreak. (A) Cumulative positivity percentage of ELISA-IgG; (B) satDNA-qPCR cumulative positivity percentage; (C) Trend of optical density values for the ELISA-IgG; (D) Trend of parasitic load values for the satDNA-qPCR. Tx: Treatment. ELISA IgG cutline: Optical Density (OD) value reportable by a positive sample (Diaz-Bello et al., 2008); TcI load cutline: 1 par eq./mL, lowest value of linear reportable range of qPCR (Duffy et al., 2013). The data in Figures (C) and (D) are presented as median and interquartile range from 25 to 75%.
Out of the 29 study patients, whole blood samples were collected in 25. In patient P7 and P21 whole blood and a Pre-Tx hemoculture isolate were obtained. In patients P26 to P29, hemocultures but not blood were obtained at different follow-up stages, so qPCR quantification of parasitic loads was not performed.
Overall, 92.06% (116 out of 126 samples) agreement between kDNA-PCR and satDNA-qPCR findings was found (Supplementary Data). The percentage of positivity for Pre-Tx samples was 100% (n=25), with a median parasite load of 2.869 [IQR = 2.113 to 3.720] log10 par eq./mL (Figures 2B, D). In approximately one-third of patients, parasitic load fluctuations above and below the lower reportable range were detected during follow-up. Despite this, a 92.5% (n=14) cumulative qPCR positivity was obtained up to nine years Post-Tx (Figure 2B). During Post-Tx monitoring, a median of the parasite load of 0.105 [IQR = -1.147 to 1.761] log10 par eq./mL was obtained, which is close to qPCR detection limit (Figure 2D). At 9 years Post-Tx, 50% (n=8) of the samples remained quantifiable by qPCR, with a median of the parasite load of 0.028 [IQR = -0.502 to 1.783] log10 par eq./mL. Thus, SatDNA-qPCR was more sensitive than serological analysis to detect treatment failure, allowing detection of 28.57% (n=5) more cases of patients who did not respond to treatment. In sum, treatment failure of around 85% (n=16) was obtained.
Serological reactivity decreased during Post Tx follow-up (Spearman’s r value= -0.961, Figure 3A), whereas parasitic load showed a slight tendency to decrease at the end of follow-up (Spearman’s r value= -0.400), however, this trend was not statistically significant (P value of the Spearman’s rank correlation coefficient >0.05). Post-Tx parasitic loads showed a median of 0.105 [IQR = -1.147 to 1.761] Log10 par.eq/mL along monitoring (Figure 3B).
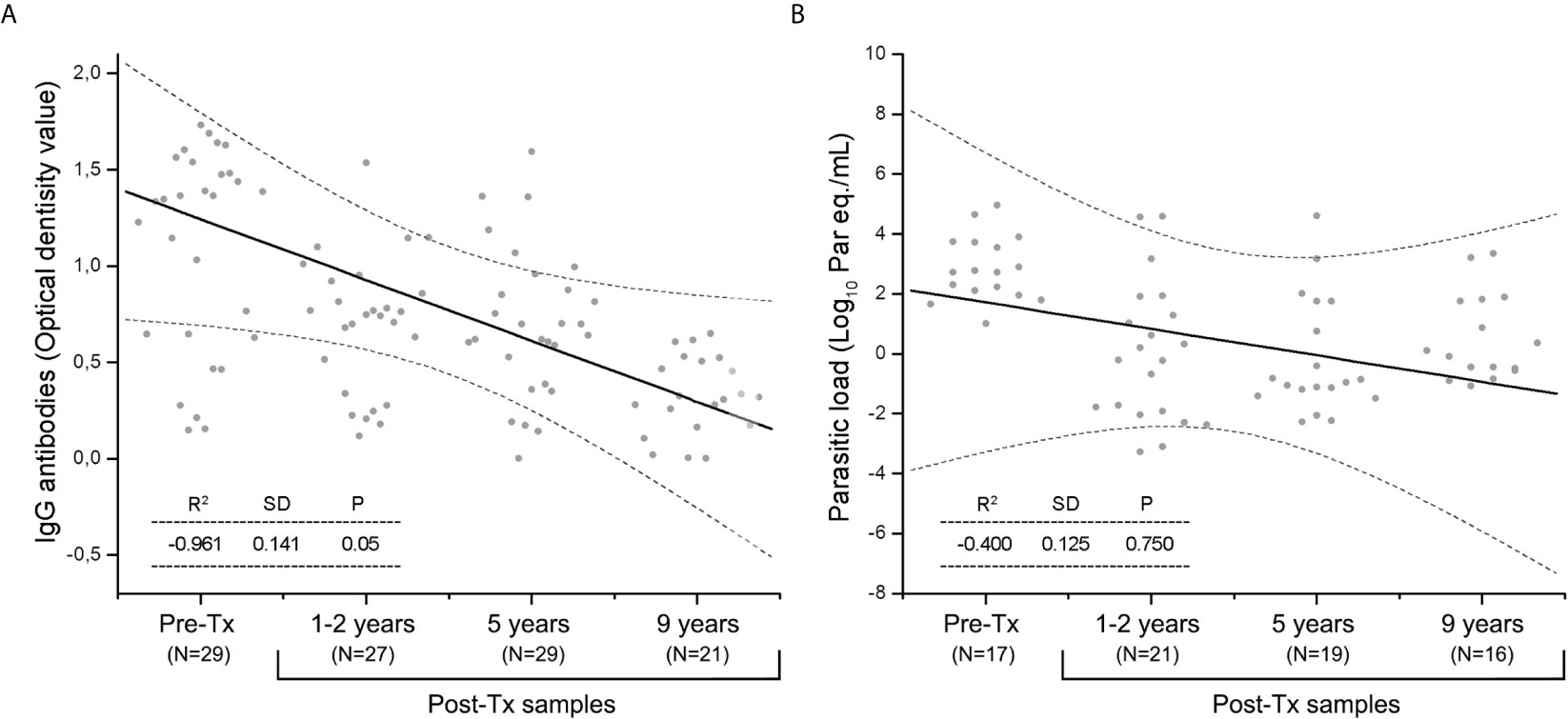
Figure 3 The Spearman’s rank correlation coefficient during Post-Tx follow-up in patients of the Chichiriviche de la Costa oral Chagas disease outbreak. (A) IgG serology; (B) SatDNA-qPCR parasitic loads. Dotted lines indicate the 95% confidence interval. P value <0.05 indicates the significance of the correlation between the variables.
Characterization of DTUs and Minicircle Signatures
Bloodstream DTUs were identified in kDNA-PCR positive samples, resulting all TcI (Table 1). The intra-TcI genetic composition was characterized by RFLP-PCR of kDNA amplicons obtained in 41 blood samples from 12 patients at different follow-up time points, and in the hemocultures isolated from 6 patients. The Ms exhibited between five and 14 digestion bands from ~30 to 330 bp of length (Figure 4A). A high degree of heterogeneity was observed among different patients` populations and between Pre-Tx and Post-Tx Ms signatures in the same patients.
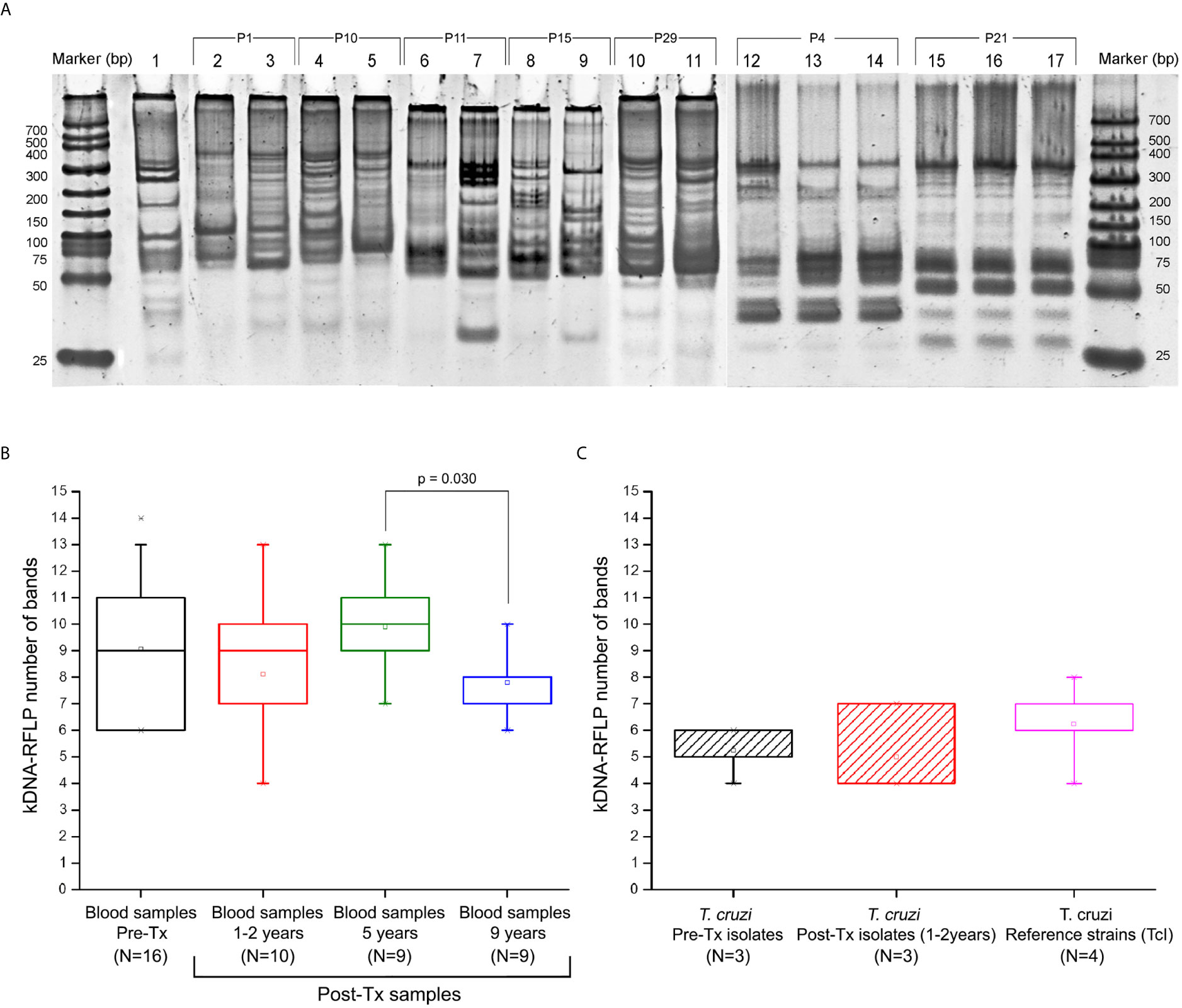
Figure 4 Profile of minicircle signatures in parasite populations by kDNA-PCR restriction fragment polymorphism (RFLP). (A) Example of comparative band analysis of minicircle profiles obtained by RFLP-PCR between patients’ parasite populations; (B) Boxplot distribution of number bands obtained by RFLP-PCR throughout the follow-up in the blood samples. (C) Distribution of the boxplot of number of digestion fragments obtained by RFLP-PCR in Trypanosoma cruzi isolates obtained from hemocultures and TcI reference strains. 1: Reference strain (DTU I - Dm28c); 2-3: Sample P1 (Pre-Tx and 5 years Post-Tx); 4-5: Sample P10 (Pre-Tx and 5 years Post-Tx); 6-7: Sample P11 (Pre-Tx and 5 years Post-Tx); 8-9: Sample P15 (Pre-Tx and 5 years Post-Tx); 10-11: Sample P29 (Pre-Tx culture isolate and 1 year Post-Tx culture isolate); 12-14: Sample P4 (Pre-Tx, 5 years and 9 years Post-Tx); 15-17: Sample P21 (Pre-Tx, 5 years and 9 years Post-Tx). The JC results shown in the box “reference strains” were obtained from the comparison of the Ms from the Dm28c, SilvioX10, gal4, and CMA strains. P value indicates significant differences between populations (Mann-Whitney U test; p<0.05). Data in the boxplot are presented as median and interquartile range of 25 to 75%.
Comparing the JC values, a slight difference in median values, 0.775 [IQR = 0.708 to 0.882] and 0.857 [IQR = 0.798 to 0.941] were observed in Pre-Tx populations analyzed from clinical samples or culture isolates, respectively (Figures 5A, B). However, the Mann-Whitney U test did not show differences between both types of samples. Out of the 25 Post-Tx Ms profiles evaluated, none presented 100% homology with the corresponding Pre-Tx Ms (Supplementary Data), revealing that parasite populations changed between the time of diagnosis and times of follow-up. When analyzing the overall T. cruzi variability between Pre-Tx and Post-Tx follow-up samples, fluctuations in the degree of polymorphism were observed.
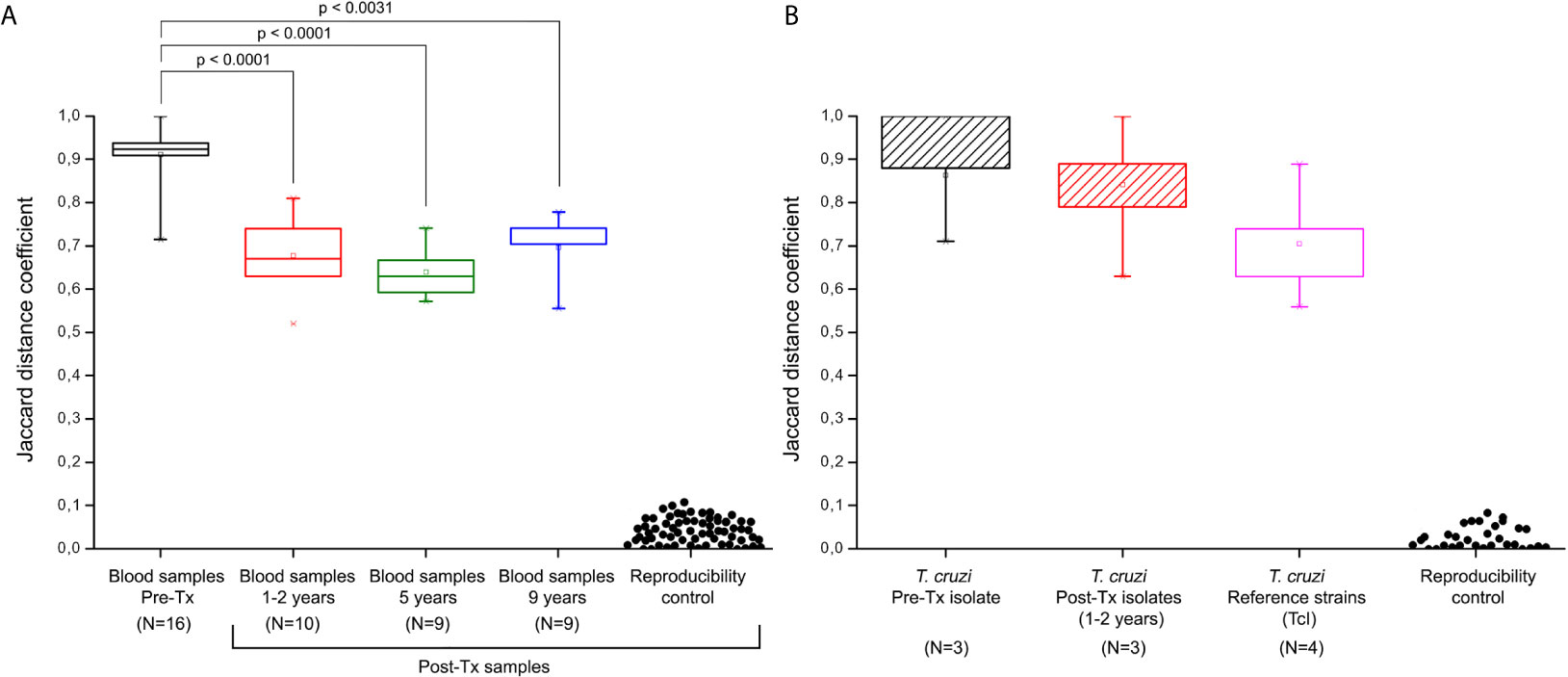
Figure 5 Jaccard distances of minicircle signatures in parasite populations by kDNA-PCR restriction fragment length polymorphism (RFLP). (A) Jaccard distances from bloodstream parasites populations in patients of the Chichiriviche de la Costa outbreak. (B) Jaccard distances of the hemocultures from the Chichiriviche de la Costa outbreak and TcI reference strains. Reproducibility controls were obtained by comparison of signatures between duplicate samples. Tx: treatment. The JC results shown in the box “reference strains” were obtained from the comparison of the Ms from the Dm28c, SilvioX10, gal4, and CMA strains. P values indicate significant differences between populations (Mann-Whitney U test; p<0.05). Data in the boxplot are presented as median and interquartile range of 25 to 75%.
The JC distances decreased respect to Pre-Tx values at 1-2 years and five years Post-Tx (Mann-Whitney U test p<0.0001, respectively). At 9 years Post-Tx, a slight increase in the JC values could be observed compared to those obtained in previous Post-Tx points, however, the difference in values was only significant when comparing the JC values against the Pre-Tx samples (Mann-Whitney U test p= 0.0031). The median of the JC among the three parasite isolates obtained at Pre-Tx was 0.92 and the JC values among the three isolates tested 1-2 years Post-Tx was 0.84 (Mann-Whitney U test p>0.05). Finally, the JC values among the Pre-Tx hemocultures obtained from this outbreak, suggested greater heterogeneity with respect to the JC values obtained in the four TcI reference strains, without reaching significant difference (Mann-Whitney U test p>0.05).
The number of Ms restriction fragments from the tested patients’ parasite populations was counted as an indirect measure of the degree of clonality of those populations along follow-up. Different patterns were observed (Figure 6). In four patients, the number of Ms bands persisted or showed a slight decrease between Pre-Tx and two years Post-Tx and decreased in complexity when tested five years Post-Tx (pattern 1), suggesting clonal selection. The parasitic populations of other four patients showed an increase in kDNA fragments (pattern 2), which suggests proliferation of a higher number of parasite clones. Finally, two patients (P4 and P21) showed no variation in the number and molecular weights of the Ms restriction fragments of their parasitic populations, except for differences in the intensity of some bands (Figure 4).
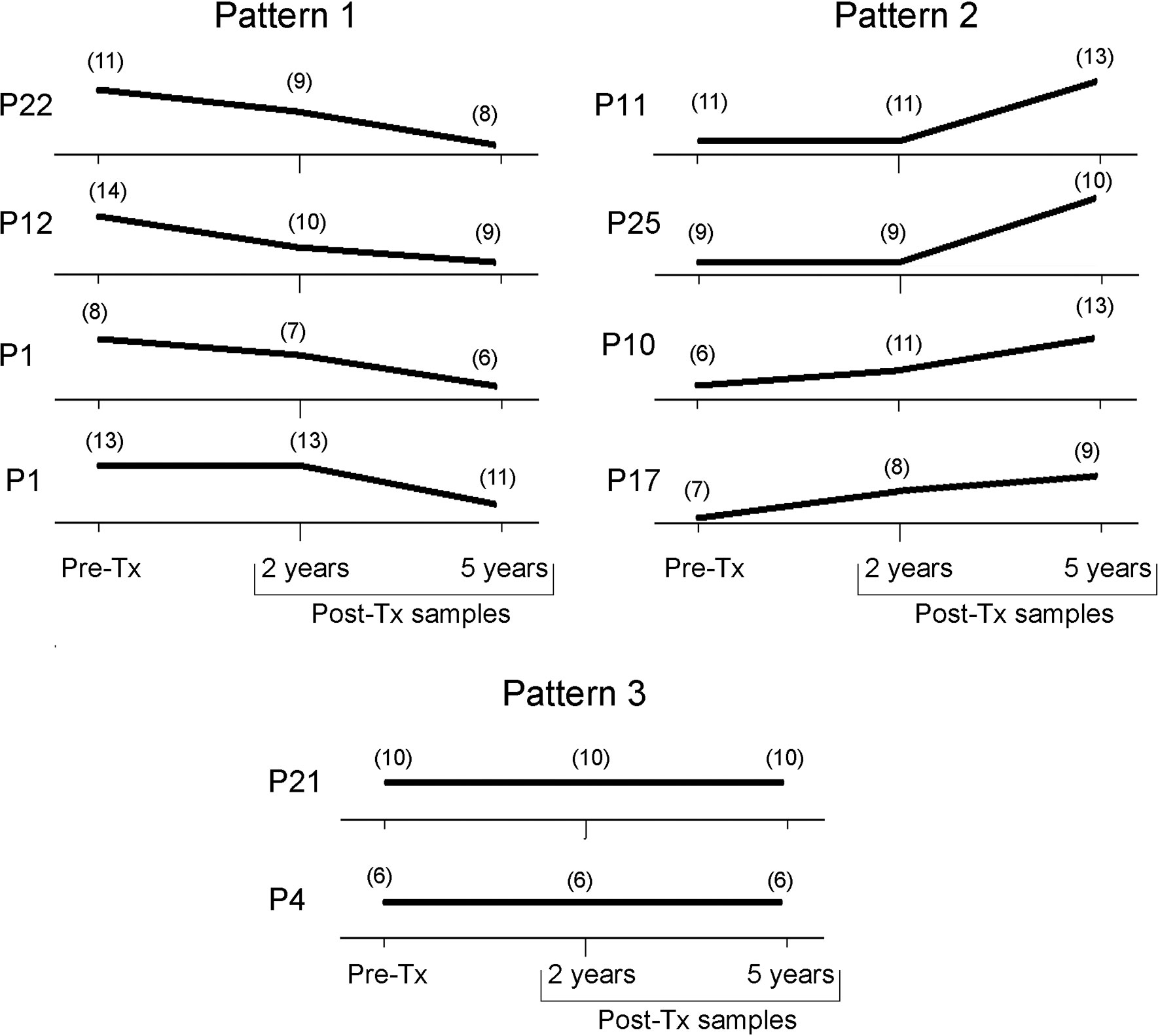
Figure 6 Trend of the genetic variability of natural T. cruzi populations by kDNA-RFLP. Each panel represents the count of the number of bands present at each time point along follow-up. Tx, treatment.
Discussion
This is the first study showing the dynamics of parasitic burden and genetic diversity of natural T. cruzi populations infecting OCD patients who did not respond to Bnz treatment after nine years of follow-up.
The fluctuations in ELISA-IgG titers and parasitic loads after etiological treatment showed negative Pearson correlation coefficients with a tendency to negativization. Parasite load values at nine years Post-Tx were very close to the lowest value of the qPCR reportable range (Figure 2D). However, no seroconversion nor persistent PCR negativization was observed, suggesting a partial parasitological response to Tx, which was even observed in the nine patients who received two rounds of Bnz.
When analyzing parasite loads, the first stands out was the heterogeneity observed in Post-Tx values. This could be associated with the evolution of the infection from its acute phase at time of diagnosis, to the chronic phase during the follow-up period. Low bloodstream parasitic loads are typical of the chronic stage, when parasite persists in tissues according to their clonal histotropism (Macedo and Segatto, 2010). The immune system plays a fundamental role in the control of circulating parasite levels once intracellular amastigotes replicate and differentiate into trypomastigotes that are released to the bloodstream (Macedo and Segatto, 2010), and this can be reflected in the bloodstream qPCR quantifications. Thus, the observed changes may be more related to the passage from one infection phase to the other one than to a selective pressure exerted by chemotherapy, or to a mixture of both factors.
Following the hypothesis proposed by Alarcón de Noya and co-authors, that “all individuals were infected by a common source for each outbreak, based in eco-epidemiological and molecular genetics studies” (Alarcon de Noya et al., 2010; Munoz-Calderon et al., 2013; Diaz-Bello et al., 2014; Alarcon de Noya et al., 2016), and identification of TcI DTU in all patients, similarly to what has been detected in other surveys in Venezuela (Carrasco et al., 2012), parasite genetic heterogeneity in patients from Chichiriviche de la Costa was revealed at the intra DTU level and between Pre-Tx and Post-Tx in a same patient. This is indicative of polyclonal populations as sources of oral infection in this outbreak. In fact, in food contaminated with triatomines feces, all metacyclic trypomastigotes present in the intestinal content of the triatomines are involved. It is estimated that a single fecal sample may contain between 3000 to 4000 trypomastigotes per microliter, thus a high clonal complexity may exist (Schaub, 1989).
Wild populations of T. cruzi may contain both susceptible and resistant clones to chemotherapeutic drugs, therefore destruction of susceptible forms by drugs leads to the selection and proliferation of resistant subpopulations (Noya et al., 2015). In a subgroup of patients whose parasite populations decreased or persisted in clonal complexity after Tx, suggested by the quantity of minicircle restriction fragments of different lengths, it is tempting to speculate that some drug-driven selection pressure could have played a role in subpopulations selection. (Ms patterns 1 and 3; Figure 6). Interestingly, parasite culture isolate from patient P21 harbored a truncated Nitroreductase protein sequence, with a putative role in drug resistance (Unpublished data).
In vitro susceptibility studies carried out with T. cruzi strains isolated from the patients of the Chacao outbreak showed high heterogeneity in IC50 values against Nfx, suggesting that the therapeutic failure could be due in part to a phenotypic variability extant in the original parasite source of oral transmission (Muñoz-Calderón et al., 2019). These findings, together with the high heterogeneity found between Pre-Tx and Post-Tx follow-up bloodstream T. cruzi populations of this outbreak are compatible to the existence of drug-driven selective pressure.
On the other side, clonal selection may occur in the natural development of T. cruzi infection from its acute to its chronic phase. Indeed, clones capable to escape from the host’s immune system are the ones to proliferate in the chronic phase. The median of the parasitic loads of 0.105 [IQR = -1.147 to 1.761] log10 par eq./mL observed during follow-up are typical of chronic infection. In four patients (P10, P11, P17, and P25) an increase in Ms polymorphism at some point in the Post-Tx follow-up was observed. In general, it is assumed that a higher number of minicircle bands is indicative of a higher level of clonality (Burgos et al., 2010). In all cases, as the JC value with respect to the Pre-Tx Ms is high, it can be hypothesized that the subpopulations proliferating after treatment were hidden into the tissues during treatment. In these particular cases, it could be attributed to possible reinfections, since the area is a region with a risk of active vector transmission (Noya et al., 2015).
A point to be highlighted is that the characterized parasitic populations correspond to bloodstream parasites. However, it is not possible to analyze the genetic diversity of parasites housed in tissues. Sánchez-Valdéz et al. (2018) showed for the first time that T. cruzi enters a dormant state, i.e. some amastigotes can interrupt their cellular replication during an in vitro infection (Thomas et al., 2018; Alonso-Padilla et al., 2020). This characteristic in bacteria and other protozoa has been related to the recurrence of infection or drug resistance (Sánchez-Valdéz et al., 2018). Therefore, the appearance of different parasite populations in the Post-Tx samples of OCD patients respect to their Pre-Tx populations could also be associated with this feature. Accordingly, we could have been identifying the genetic diversity of intracellular parasite populations that survived in an environment where the drug concentration was lower and they were not necessarily resistant clones. This could be the case for those patients who showed an increase in parasite variability after Tx. On the other hand, dormancy would not be so relevant in TcI strains, at least compared to other DTUs, as reported by Resende et al. (2020).
The limited sample size of the patients for each of the time points evaluated (Pre-Tx, 1-2 years, 5 years and 9 years Post-Tx) has been a study limitation, mainly influenced by the difficulty of extracting parasitic gDNA from blood samples in patients at the chronic phase of Chagas disease. Additionally, the lack of a placebo group limits the analysis of the genetic variability of parasitic populations when evolving from the acute phase to the chronic phase of the disease. The inclusion of a placebo group would have also allowed us to evaluate in greater detail the response of these parasitic populations after the etiological treatment. These limitations must be taken in account when designing future studies. In conclusion, our serological and molecular parasitological findings suggest that synergistic multiple factors, such as the existence of clones with natural drug resistance, selective pressure exerted by trypanomicidal treatment, existence of dormant subpopulations, and evolution of acute oral infection to its chronic phase may account for the persistence of seroreactivity and parasite diversity in this patient cohort after almost a decade of follow-up.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics Statement
The studies involving human participants were reviewed and approved by Ethical Review Board of the Instituto de Medicina Tropical “Dr Félix Pifano”-Universidad Central de Venezuela, Caracas, Venezuela (CEC-IMT 019/2010 - December 10, 2010). Written informed consent to participate in this study was provided by the participants’ legal guardian/next of kin.
Author Contributions
AM-C and AS were responsible for study design, data analysis, and writing up of the manuscript. ZD-B, BA, and ON-G were responsible for recruitment and clinical care of the patients. AM-C and ZD-B were responsible for the laboratory analysis. AS and BA supervised the molecular and serological related experiments. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by a Fellowship given by the Programme for biotechnology in Latin America and the Caribbean of the United Nations University (UNU-BIOLAC) [to AM-C], funds from the ANPCyT through PICT 2014-0274 [to AS] and financial support from the Immunology Section - Instituto de Medicina Tropical – Universidad Central de Venezuela.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This is a collaborative article of the Network NHEPACHA.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2021.665063/full#supplementary-material
References
Alarcon de Noya, B., Colmenares, C., Diaz-Bello, Z., Ruiz-Guevara, R., Medina, K., Munoz-Calderon, A., et al. (2016). Orally-Transmitted Chagas Disease: Epidemiological, Clinical, Serological and Molecular Outcomes of a School Microepidemic in Chichiriviche De La Costa, Venezuela. Parasite Epidemiol. Control 1 (2), 188–198. doi: 10.1016/j.parepi.2016.02.005
Alarcón de Noya, B., Díaz-Bello, Z., Colmenares, C., Muñoz-Calderón, A., Ruiz-Guevara, R., Balzano, L., et al. (2011). Eficacia Terapéutica En El Primer Brote De Transmisión Oral De La Enfermedad De Chagas En Venezuela. Biomédica 31 (sup 3), 3–315. doi: 10.7705/biomedica.v31i0.525
Alarcón de Noya, B., Díaz-Bello, Z., Colmenares, C., Ruiz-Guevara, R., Mauriello, L., Muñoz-Calderón, A., et al. (2015). Update on Oral Chagas Disease Outbreaks in Venezuela: Epidemiological, Clinical and Diagnostic Approaches. Memórias do Instituto Oswaldo Cruz 110 (3), 377–386. doi: 10.1590/0074-02760140285
Alarcon de Noya, B., Diaz-Bello, Z., Colmenares, C., Ruiz-Guevara, R., Mauriello, L., Zavala-Jaspe, R., et al. (2010). Large Urban Outbreak of Orally Acquired Acute Chagas Disease At a School in Caracas, Venezuela. J. Infect. Dis. 201 (9), 1308–1315. doi: 10.1086/651608
Alarcón de Noya, B., Martinez, J. (2009). Transmisión Oral De La Enfermedad De Chagas En Venezuela: Un Segundo Brote Escolar. Salus 13 (2), 9.
Alonso-Padilla, J., Abril, M., Alarcón de Noya, B., Almeida, I. C., Angheben, A., Araujo Jorge, T., et al. (2020). Target Product Profile for a Test for the Early Assessment of Treatment Efficacy in Chagas Disease Patients: An Expert Consensus. PloS Neglected Trop. Dis. 14 (4), e0008035. doi: 10.1371/journal.pntd.0008035
Amato Neto, V. (1999). Etiological Treatment for Infection by Trypanosoma Cruzi. Memórias do Instituto Oswaldo Cruz 94, 337–339. doi: 10.1590/S0074-02761999000700065
Bianchi, F., Cucunubá, Z., Guhl, F., González, N. L., Freilij, H., Nicholls, R. S., et al. (2015). Follow-Up of an Asymptomatic Chagas Disease Population of Children After Treatment With Nifurtimox (Lampit) in a Sylvatic Endemic Transmission Area of Colombia. PloS neglected Trop. Dis. 9 (2), e0003465. doi: 10.1371/journal.pntd.0003465
Burgos, J. M., Diez, M., Vigliano, C., Bisio, M., Risso, M., Duffy, T., et al. (2010). Molecular Identification of Trypanosoma Cruzi Discrete Typing Units in End-Stage Chronic Chagas Heart Disease and Reactivation After Heart Transplantation. Clin. Infect. Dis. 51 (5), 485–495. doi: 10.1086/655680
Carrasco, H. J., Segovia, M., Llewellyn, M. S., Morocoima, A., Urdaneta-Morales, S., Martínez, C., et al. (2012). Geographical Distribution of TrypanosomaCruzi Genotypes in Venezuela. PloS neglected Trop. Dis. 6 (6), e1707. doi: 10.1371/journal.pntd.0001707
Cura, C. I., Duffy, T., Lucero, R. H., Bisio, M., Peneau, J., Jimenez-Coello, M., et al. (2015). Multiplex Real-Time Pcr Assay Using Taqman Probes for the Identification of TrypanosomaCruzi Dtus in Biological and Clinical Samples. PloS Negl. Trop. Dis. 9 (5), e0003765. doi: 10.1371/journal.pntd.0003765
Diaz-Bello, Z., Thomas, M. d. C., Egui, A., Pérez-Antón, E., López López, M. C, et al. (2014). Trypanosoma Cruzi Genotyping Supports a Common Source of Infection in a School-Related Oral Outbreak of Acute Chagas Disease in Venezuela. Epidemiol. Infect. 142 (1), 156–162. doi: 10.1017/S0950268813000757
Diaz-Bello, Z., Zavala-Jaspe, R., Diaz-Villalobos, M., Mauriello, L., Maekelt, A., de Noya, B. A. (2008). A Confirmatory Diagnosis of Antibodies Anti-TrypanosomaCruzi in Donors Referred by Blood Banks in Venezuela. Investigacion clinica 49 (2), 141–150.
Duffy, T., Cura, C. I., Ramirez, J. C., Abate, T., Cayo, N. M., Parrado, R., et al. (2013). Analytical Performance of a Multiplex Real-Time PCR Assay Using Taqman Probes for Quantification of TrypanosomaCruzi Satellite DNA in Blood Samples. PloS Negl. Trop. Dis. 7 (1), e2000. doi: 10.1371/journal.pntd.0002000
Garcia-Bournissen, F. (2019). “Clinical Pharmacology of Drugs for the Treatment of Chagas Disease,” in Chagas Disease, vol. 299-312. (Cham, New York:Springer).
Macedo, A. M., Segatto, M. (2010). “Implications of Trypanosoma Cruzi Intraspecific Diversity in the Pathogenesis of Chagas Disease,” in American Trypanosomiasis (Amsterdam: Elsevier), 489–522.
Molina-Morant, D., Fernández, M., Bosch-Nicolau, P., Sulleiro, E., Bangher, M., Salvador, F., et al. (2020). Efficacy and Safety Assessment of Different Dosage of Benznidazol for the Treatment of Chagas Disease in Chronic Phase in Adults (Multibenz Study): Study Protocol for a Multicenter Randomized Phase Ii Non-Inferiority Clinical Trial. Trials 21, 1–10. doi: 10.1186/s13063-020-4226-2
Moscatelli, G., Moroni, S., García Bournissen, F., González, N., Ballering, G., Schijman, A., et al. (2019). Longitudinal Follow Up of Serological Response in Children Treated for Chagas Disease. PloS neglected Trop. Dis. 13 (8), e0007668. doi: 10.1371/journal.pntd.0007668
Muñoz-Calderón, A., Díaz-Bello, Z., Ramírez, J. L., Noya, O., Alarcón de Noya, B. (2019). Nifurtimox Response of TrypanosomaCruzi Isolates From an Outbreak of Chagas Disease in Caracas, Venezuela. J. Vector Borne Dis. 56 (3), 237–243. doi: 10.4103/0972-9062.289397
Munoz-Calderon, A., Diaz-Bello, Z., Valladares, B., Noya, O., Lopez, M. C., Alarcon de Noya, B., et al. (2013). Oral Transmission of Chagas Disease: Typing of TrypanosomaCruzi From Five Outbreaks Occurred in Venezuela Shows Multiclonal and Common Infections in Patients, Vectors and Reservoirs. Infect. Genet. Evol. 17, 113–122. doi: 10.1016/j.meegid.2013.03.036
Muñoz-Calderón, A., Santaniello, A., Pereira, A., Yannuzzi, J., Díaz-Bello, Z., Alarcón de Noya, B. (2012). Susceptibilidad in Vitro a Nifurtimox Y Benznidazol De Aislados De TrypanosomaCruzi Obtenidos De Pacientes Venezolanos Con Enfermedad De Chagas Infectados Por Mecanismos De Transmisión Oral Y Vectorial. Rev. Ibero-Latinoam Parasitol 71 (1), 14–22.
Noya, O., Ruiz-Guevara, R., Díaz-Bello, Z., Alarcón de Noya, B. (2015) “Epidemiología Y Clínica De La Transmisión Oral De Trypanosoma Cruzi,” in Rev Esp Epidem: XI Workshop on Chagas Disease (Barcelona Spain: Rev Esp Epidem), 23–34.
Pereiro, A. C. (2019). Guidelines for the Diagnosis and Treatment of Chagas Disease. Lancet 393 (10180), 1486–1487. doi: 10.1016/S0140-6736(19)30288-0
Resende, B. C., Oliveira, A. C. S., Guañabens, A. C. P., Repolês, B. M., Santana, V., Hiraiwa, P. M., et al. (2020). The Influence of Recombinational Processes to Induce Dormancy in TrypanosomaCruzi. Front. Cell. Infection Microbiol. 10, 5. doi: 10.3389/fcimb.2020.00005
Rueda, K., Trujillo, J. E., Carranza, J. C., Vallejo, G. A. (2014). Transmisión Oral De TrypanosomaCruzi: Una Nueva Situación Epidemiológica De La Enfermedad De Chagas En Colombia Y Otros Países Suramericanos. Biomédica 34 (4), 631–641. doi: 10.7705/biomedica.v34i4.2204
Sánchez-Valdéz, F. J., Padilla, A., Wang, W., Orr, D., Tarleton, R. L. (2018). Spontaneous Dormancy Protects TrypanosomaCruzi During Extended Drug Exposure. Elife 7, e34039. doi: 10.7554/eLife.34039
Schaub, G. (1989). Trypanosoma Cruzi: Quantitative Studies of Development of Two Strains in Small Intestine and Rectum of the Vector Triatoma Infestans. Exp. parasitology 68 (3), 260–273. doi: 10.1016/0014-4894(89)90108-2
Schijman, A. G., Bisio, M., Orellana, L., Sued, M., Duffy, T., Jaramillo, A. M. M., et al. (2011). International Study to Evaluate Pcr Methods for Detection of TrypanosomaCruzi DNA in Blood Samples From Chagas Disease Patients. PloS Negl. Trop. Dis. 5 (1), e931. doi: 10.1371/journal.pntd.0000931
Keywords: Trypanosoma cruzi, Oral Chagas disease, trypanocidal chemotherapy, therapeutic failure, genetic diversity, restriction fragment length polymorphism, parasite load, minicircle signature
Citation: Muñoz-Calderón A, Díaz-Bello Z, Alarcón de Noya B, Noya-González OO and Schijman AG (2021) Characterization and Follow-Up of Trypanosoma cruzi Natural Populations Refractory to Etiological Chemotherapy in Oral Chagas Disease Patients. Front. Cell. Infect. Microbiol. 11:665063. doi: 10.3389/fcimb.2021.665063
Received: 07 February 2021; Accepted: 12 April 2021;
Published: 28 April 2021.
Edited by:
Gustavo Benaim, Fundación Instituto de Estudios Avanzados (IDEA), VenezuelaReviewed by:
Sergio Andres Castañeda, Rosario University, ColombiaRodrigo Andrés López-Muñoz, Austral University of Chile, Chile
Copyright © 2021 Muñoz-Calderón, Díaz-Bello, Alarcón de Noya, Noya-González and Schijman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alejandro G. Schijman, c2NoaWptYW5AZG5hLnViYS5hcg==