 Efstathios S. Giotis
Efstathios S. Giotis- 1Section of Molecular Virology, Department of Infectious Diseases, Imperial College London, London, United Kingdom
- 2School of Life Sciences, University of Essex, Colchester, United Kingdom
Bats are considered natural reservoirs of various, potentially zoonotic viruses, exemplified by the influenza A-like viruses H17N10 and H18N11 in asymptomatic Neotropical bats. These influenza viruses are evolutionarily distinct, are poorly adapted to laboratory mice and ferrets and cannot reassort in vitro with conventional strains to form new influenza subtypes. However, they have attracted renewed attention following reports that their entry in host cells is mediated by the trans-species conserved MHC-II proteins, suggesting that they hold zoonotic potential. Despite the recent studies, the viruses' epidemiology and public health significance remain incompletely understood. Delineating the mechanistic basis of the interactions with their hosts and assessing their global distribution are essential in order to fully assess the zoonotic threat that these strains pose.
Introduction
The Severe Acute Respiratory Syndrome (SARS), Middle East Respiratory Syndrome (MERS), Nipah (NiV), Hendra (HeV), and Ebola (EBOV) viruses' outbreaks confirmed the inextricable nature of human and bat health and disease and highlighted that focusing on the “spillover” potential of known, and novel, bat viruses is critical to predict and prevent pandemics. The remarkable ability of bats to coexist with a wide range of viruses that would be pathogenic in flightless mammals (Fl−M) is not yet fully understood but possibly relates to their unique, flight-adapted antiviral immunity (Calisher et al., 2006; Hayman et al., 2013). Intriguingly, genetic material from viruses that resemble influenza type A viruses has been recovered from asymptomatic fruit bats of the Neotropic bat family Phyllostomidae (Sturnira lilium and Artibeus planirostris) in several countries of Central and South America (Figure 1) (Tong et al., 2012, 2013; Campos et al., 2019). Influenza A viruses (IAVs) are orthomyxoviruses with eight single-stranded negative-sense viral RNAs (vRNAs) encapsidated into viral ribonucleoproteins (vRNPs). IAVs emerge from aquatic birds, via genome reassortment and mutation, and are able to cause epidemics (and sporadic pandemics; Simonsen, 1999) in humans, lower animals and birds (Simonsen, 1999). The bat influenza viruses (BatIVs) are phylogenetically distinct from the conventional IAVs and they were designated as H17N10 and H18N11 (Table 1). Bats in Latin America, but not in Central Europe, have been found seropositive for BatIVs (Tong et al., 2013; Fereidouni et al., 2015). Antibodies against human H2N2 and H3N2, as well as classical H9, have also been found in bats elsewhere (L'vov et al., 1979; Kelkar et al., 1981; Isaeva et al., 1982), suggesting they are susceptible to IAV infection. The epithelial kidney cells of flying foxes (Pteropus alecto) co-express both avian (α2,3-Gal) and human (α2,6-Gal) sialic acid (SA) receptors (Chothe et al., 2017) and are thus susceptible to infection by both avian and human IAVs, but more importantly they allow reassortment between co-infecting influenza viruses (Dlugolenski et al., 2013). The N-terminal domain of the H17N10 PA subunit of the influenza virus polymerase complex possesses endonuclease activity comparable to that of IAVs (Tefsen et al., 2014). Equally intriguingly, in the position 627 of the polymerase gene PB2, one of the most commonly identified IAV virulence markers, BatIVs have a serine compared to glutamic acid in avian and lysine in mammalian influenza strains, suggestive of an alternative evolutionary pathway for avian IAV's adaptation in mammals (Mehle, 2014). This all raised the question whether novel IAVs could emerge from bats to which human and animal populations would be immunologically “naïve,” causing pandemics.
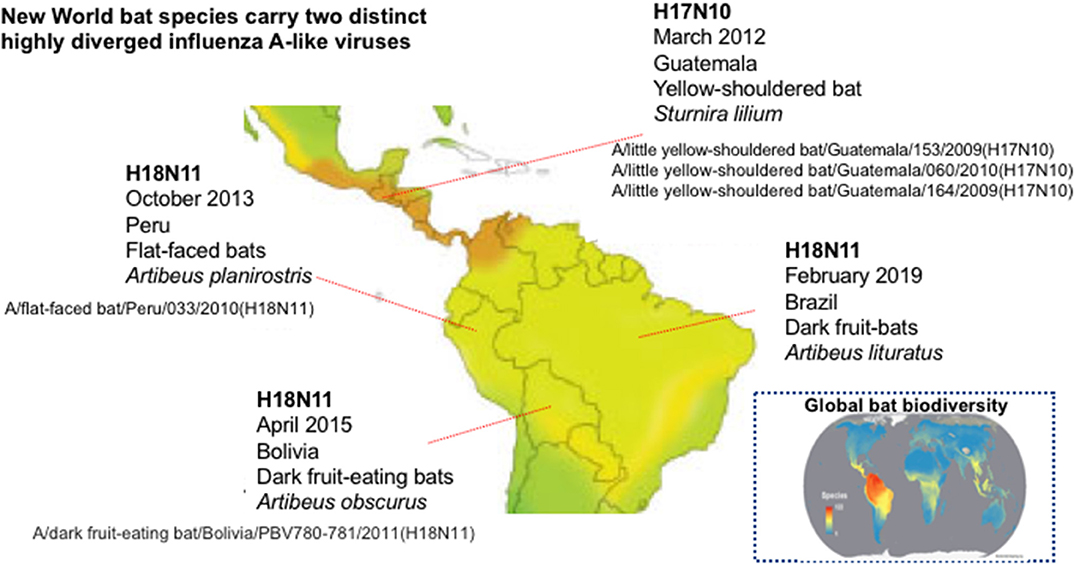
Figure 1. Countries of Central and South America where bat influenza A viruses have been reported. Map showing the global pattern of bat species richness was provided by Clinton Jenkins (see BiodiversityMapping.org) using species data from The IUCN Red List of Threatened Species (2018; https://www.iucnredlist.org) (Jenkins et al., 2013).
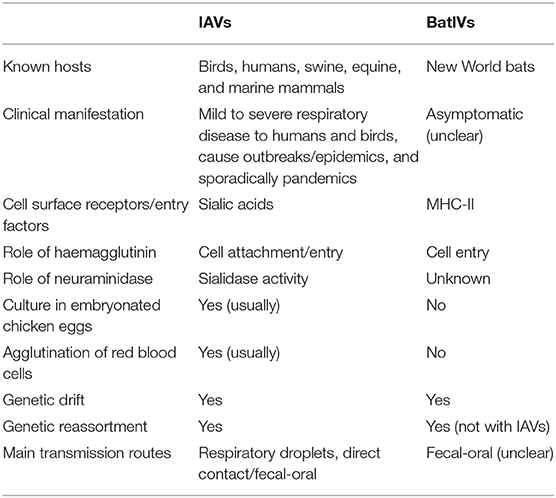
Table 1. List of distinctive features of IAVs and BatIVs.
BatIVs are Distinct and Employ Unconventional Receptors for Cell Entry
The genetic material of BatIVs is similar to classic flu viruses, but their surface glycoproteins haemagglutinin (HA) and neuraminidase (NA) are evolutionarily and functionally diverged (Li et al., 2012; Zhu et al., 2012; Sun et al., 2013; Tong et al., 2013). BatIVs cannot be cultured in embryonated chicken eggs and do not agglutinate red blood cells (Tong et al., 2012, 2013). Initial efforts by researchers to isolate live infectious BatIVs directly from bats failed, due to unavailability of permissive cell lines (Ciminski et al., 2017). In addition, research on these viruses was further complicated by the dearth of bat cell lines and the limited bat genomic data. Attempts to circumvent these limitations have included: (i) using HA-17 or HA-18 pseudotyped vesicular stomatitis virus (VSV) and HIV-1 based lentiviruses (Hoffmann et al., 2016; Maruyama et al., 2016; Carnell et al., 2018; Giotis et al., 2019), (ii) engineering BatIV/IAV chimaeric viruses (Juozapaitis et al., 2014; Zhou et al., 2014), and (iii) reconstructing authentic BatIVs using reverse genetics (Moreira et al., 2016; Sato et al., 2019; Zhong et al., 2019). HA17-VSV was able to infect bat cell lines (EidNi, HypNi, and EpoNi) but only a few of the common Fl−M cell lines, including human U-87 MG glioblastoma and SK-Mel-28 melanoma cells, canine RIE 1495 and MDCK II kidney cells (Hoffmann et al., 2016; Maruyama et al., 2016; Moreira et al., 2016). The identification of MDCK II, in particular, as susceptible cell lines to BatIVs opened the way for a more comprehensive characterization of these strains (Moreira et al., 2016; Giotis et al., 2019; Karakus et al., 2019).
Crystal structure analyses revealed that the bat haemagglutinins display typical HA protein folds but lack any obvious cavity to accommodate SA, which are the conventional receptors of IAVs. Recently, two independent studies demonstrated that the cell-entry of H17N10 (Giotis et al., 2019) and H18N11 (Karakus et al., 2019) is mediated by MHC-II receptors that are well-conserved in many species. Hence, immortalized cell lines that express MHC-II receptors on their surface such as several human leukemia and lymphoma cell lines Raji, Ramos, and BJAB B-lymphocytes could be used for the study of the viruses' biology (Giotis et al., 2019). Interestingly, ectopic expression of pig, mice, and chicken MHC-II have been shown to confer susceptibility to H18N11 in non-susceptible cells (Karakus et al., 2019) implying a potential role for the respective animals as intermediary hosts. It is as yet unclear whether MHC-II receptors function with other unknown factors to facilitate virus internalization and also whether the viruses remain cell associated following cell-entry and are being passed on by direct cell-cell contact.
MHC-II molecules occur as three highly polymorphic isotypes (HLA-DR, HLA-DP, and HLA-DQ). They are selectively expressed on the surface of professional antigen presenting cells (APCs), act as ligands for the T-cell receptor (TCR), and play a key role in the presentation of foreign antigens to CD4+ T helper cells and immune surveillance (Jones et al., 2006; Roche and Furuta, 2015). Bats contain all classical MHC class II gene families that are responsible for antigen presentation with an extra DRB2 gene copy located outside the MHC-II region in P. alecto (Ng et al., 2017). The MHC-II dependent cell entry suggests that BatIVs might hijack APCs such as B lymphocytes and dendritic cells for viral dissemination and/or survival perhaps in the early stages of infection. It is unknown to what degree their binding to APCs might influence the global outcome of the host immune responses. A blockade of TCR recognition by steric hindrance as described for Epstein-Barr virus (Ressing et al., 2003, 2005; Wiertz et al., 2007), could explain the asymptomatic status of the infection in the captured New World bats.
The Enigmatic Role of bat Influenza Viruses' Neuraminidases
The bat neuraminidases (NAs) are structurally similar to classical NAs but lack conserved amino acids for SA binding or cleavage (Li et al., 2012; Zhu et al., 2012). Moreover, unlike classical NAs, they display no enzymatic activity (Garcia-Sastre, 2012; Carnell et al., 2018), have a dispensable role in viral entry (Hoffmann et al., 2016; Maruyama et al., 2016; Giotis et al., 2019) and their function is not yet elucidated. Recent studies demonstrated that the passage of reverse-genetics-generated H18N11 virus in cell cultures, accumulates mutations in the N11 protein that increase virus titers in culture and may enhance organ tropism in vivo (Zhong et al., 2019). Ciminski et al. demonstrated by utilizing a chimeric bat influenza virus (PR8-H18N11) that viruses encoding the full-length N11 protein exhibited a growth advantage over viruses that encode a truncated protein version and also showed that N11 is essential for viral transmission (Ciminski et al., 2019b). Another study showed that the N10 protein facilitates heterosubtypic (H5 and H7) influenza hemagglutinin-bearing pseudotype release in the absence of another source of neuraminidase, indicating a possible role of N10 in viral release (Carnell et al., 2018). It has been proposed that N11 downregulates MHC-II, thereby facilitating virion release but mechanistic data for such function is as yet missing (Ciminski et al., 2019b). There is no evidence that a functional balance exists between bat HAs and NAs although it is possible that the proteins coordinate their actions. Despite the recent progress in our understanding, the exact function of bat NAs remains an enticing mystery.
Intraspecies and Interspecies Transmission of bat Influenza Viruses
It is now becoming more evident that BatIVs may transmit in a different manner than conventional flu strains. It has recently been shown that the H18N11 virus readily transmits between bats (Ciminski et al., 2019b). Following experimental intranasal H18N11 infection, the Neotropical Phylostomidae bat species Artibeus jamaicensis shed high viral loads via the fecal route and were able to infect naïve contact animals (Ciminski et al., 2019b). Histopathological analysis of the infected bats indicated that viral replication proceeds in the follicle-associated epithelium of gut-associated lymphoid tissue, suggesting virus uptake from the gastrointestinal lumen (Ciminski et al., 2019b), in line with the rich gut epithelial expression of MHC-II (Wosen et al., 2018). Furthermore, the researchers detected high loads of H18N11 viral transcripts in rectal swabs and excretions (Ciminski et al., 2019b). Collectively, these findings imply that an environmental (fecal-oral) mode of BatIVs transmission is more likely than an airborne one, albeit the latter is not yet compellingly disproved. Other infection routes including subcutaneous, transplacental, vaginal, intracranial infections have not yet been reported. In contrast to bats, H18N11 has been reported to have limited replicative ability in laboratory mice and more interestingly in ferrets which share similar lung physiology and SA distributions to humans (Ciminski et al., 2019b; Karakus et al., 2019; Zhong et al., 2019). Whether these observations are animal/infection-route-dependent or they actually reflect a low zoonotic risk for BatIVs remains to be seen.
In absence of conclusive scientific proof, the question remains as to whether BatIVs are confined to a sylvatic transmission cycle and perpetuate in Neotropical bat populations or are capable of urban adaptation. Anthropogenic disruptions of ecological habitats that led to urban transmission of other enzootic bat viruses (i.e., HeV, NiV) have been extensively described in the scientific literature. Despite the increasing disturbances in the fire-prone Neotropical forests, New World bats have not yet been implicated in the transmission of zoonotic viruses, other than rabies, to humans (Moratelli and Calisher, 2015). Even so, Latin America is home to the richest and most diverse bat fauna in the world (Figure 1) including almost 150 Phyllostomidae species (Jenkins et al., 2013) in which BatIVs have been detected. Unlike African and Asian bats which are consumed regularly, New World bats are only eaten by few native indigenous people (Moratelli and Calisher, 2015). A possible exposure to infected bat blood and body fluids may hypothetically create a pathway for disease transmission to humans. Another scenario may involve the accidental introduction of BatIVs to the local fauna or other Phyllostomidae species such as the widespread blood-eating vampire bat (Desmodus rotundus), which thrives in both native and anthropogenically transformed ecosystems (Bergner et al., 2020). Vampire bats have long been suspected of passing on rabies to humans and livestock in Latin America by biting and scratching (Rupprecht et al., 2002). It will be useful to explore in future studies whether haematophagous bats can act as maintenance hosts for BatIVs and if their biting can form a potential zoonotic transmission route either directly or through mammalian intermediate hosts.
The Host Factors Regulating BatIVs Replication are Poorly Understood
BatIVs, like all viruses, have to compromise with positive and negative genetic factors present in target cells for their survival at each replication stage. Little is known regarding the interaction of BatIV proteins and RNA with the host or viral factors even though such interactions may determine the fate and/or efficiency of infection, transmission, and epidemic potential of the viruses. Previous studies revealed that the bat Nonstructural NS1 proteins can act as interferon (IFN) antagonists in human cells, and likely inhibit induction of IFN at a pretranscriptional level (Ciminski et al., 2017). More recently, it has been shown that the IFN-induced human MxA protein controls the replication of H18N11, but it is not clear whether sufficient MxA-escape mutations in H18N11 NP can be acquired in vivo that could potentially result in full MxA resistance (Ciminski et al., 2019a). Nearly all lab work examining host and viral immune-modulating proteins is performed with human/rodent cell lines. The difficulty in interpreting these data is that evolutionarily-optimized immune factors behave differently in non-natural hosts. Certainly, comprehensive kinetic analyses of immune-responses to BatIVs using primary or immortalized bat cell lines will be particularly informative. For instance, a comparison of the transcriptome of BatIV infected versus uninfected bat cells could help us identify specific immune genes contributing to host resistance and the molecular mechanisms underlying the viral pathogenesis.
Do bat Influenza Viruses Pose a Zoonotic Risk?
Reassortment of gene segments between co-infecting viruses is a key process mediating the genetic evolution of influenza viruses and the generation of novel epidemic and pandemic strains. BatIVs are able to reassort between themselves but not with conventional IAVs in vitro (Juozapaitis et al., 2014; Zhou et al., 2014). Generation of bat chimeric viruses was only possible when the HA/NA coding regions were flanked with the authentic BatIV packaging signals demonstrating packaging incompatibilities between IAVs and BatIVs (Juozapaitis et al., 2014; Zhou et al., 2014). This finding dismisses the scenario of emergence of a new “reassortant” virus with human/avian IAVs unless the bat viruses undergo major genetic changes over time.
However, the abilities of BatIVs to (i) reassort between themselves, (ii) to mutate in order to infect and transmit sustainably among their hosts, and (iii) enter human HLA-DR+ cells, highlight that a zoonotic transmission of BatIVs is theoretically possible. The documented spillover of other non-reassortant bat-borne RNA viruses following continued host-pathogen interaction (i.e., NiV and EBOV) lends certain credence to this hypothesis, albeit clearly, supporting evidence is lacking.
To explore the ecological and evolutionary dynamics of these and possibly other unknown influenza-A-like viruses, further prevalence and serological studies in Neotropical bat populations are required coupled with the surveillance of bat-exposed humans and livestock. Surveys of bat colonies have previously led to identification of other zoonotic viruses, including HeV in Pteropus sp. in Australia, NiV in Pteropus lylei in Thailand, and MARV in Rousettus aegyptiacus in Uganda (Wacharapluesadee et al., 2010; Amman et al., 2012; Field et al., 2015). A computational study which used spatial empirical models to trace the steps of emergence of bat viruses and the transmission opportunities to humans pinpointed sub-Saharan Africa as the top-priority location for pathogen discovery in wildlife (Brierley et al., 2016). West Subsaharan Africa, in particular, hosts enormous populations of sedentary and migrating bats living in proximity to human and animal populations. Considering the number, the morbidity and mortality of emerging viruses that are hosted in African bats as well as the serological evidence against IAVs (Freidl et al., 2015), future surveillance and serological studies could lead to the identification of novel bat influenza subtypes.
Conclusions
In summary, BatIVs are unconventional influenza viruses that resemble to some extent more paramyxoviruses rather than typical orthomyxoviruses. Despite the recent findings on the cell entry factors and NAs of these viruses, it is clear that we only scratched the surface in terms of characterization of these viruses. The scientific evidence so far indicate a limited spillover risk but data is not conclusive enough to dismiss out of hand the possibility of zoonotic transmission. Forecasting viral spillover is a challenging task and additional interdisciplinary and more up-to-date approaches are warranted to fully appreciate the ecology and the implications of these viruses for public health. Future studies on BatIVs hold extra value as they can provide broader mechanistic insights into the molecular biology of influenza viruses and might inform translational studies.
Author Contributions
The author confirms being the sole contributor of this work and has approved it for publication.
Funding
EG was supported by funding from a Wellcome Trust New Investigator award to Marcus Dorner (104771/Z/14/Z).
Conflict of Interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Amman, B. R., Carroll, S. A., Reed, Z. D., Sealy, T. K., Balinandi, S., Swanepoel, R., et al. (2012). Seasonal pulses of Marburg virus circulation in juvenile rousettus aegyptiacus bats coincide with periods of increased risk of human infection. PLoS Pathog. 8:e1002877. doi: 10.1371/journal.ppat.1002877
Bergner, L. M., Orton, R. J., Benavides, J. A., Becker, D. J., Tello, C., Biek, R., et al. (2020). Demographic and environmental drivers of metagenomic viral diversity in vampire bats. Mol. Ecol. 29, 26–39. doi: 10.1111/mec.15250
Brierley, L., Vonhof, M. J., Olival, K. J., Daszak, P., and Jones, K. E. (2016). Quantifying global drivers of zoonotic bat viruses: a process-based perspective. Am. Nat. 187, E53–E64. doi: 10.1086/684391
Calisher, C. H., Childs, J. E., Field, H. E., Holmes, K. V., and Schountz, T. (2006). Bats: important reservoir hosts of emerging viruses. Clin. Microbiol. Rev. 19, 531–545. doi: 10.1128/CMR.00017-06
Campos, A. C. A., Goes, L. G. B., Moreira-Soto, A., De Carvalho, C., Ambar, G., Sander, A. L., et al. (2019). Bat influenza A(HL18NL11) virus in fruit bats, Brazil. Emerg. Infect. Dis. 25, 333–337. doi: 10.3201/eid2502.181246
Carnell, G., Giotis, E. S., Grehan, K., Ferrara, F., Mather, S., Molesti, E., et al. (2018). The bat influenza H17N10 can be neutralized by broadly- neutralizing monoclonal antibodies and its neuraminidase can facilitate viral egress. Biorxiv 499947. doi: 10.1101/499947
Chothe, S. K., Bhushan, G., Nissly, R. H., Yeh, Y. T., Brown, J., Turner, G., et al. (2017). Avian and human influenza virus compatible sialic acid receptors in little brown bats. Sci. Rep. 7:660. doi: 10.1038/s41598-017-00793-6
Ciminski, K., Pulvermuller, J., Adam, J., and Schwemmle, M. (2019a). Human MxA is a potent interspecies barrier for the novel bat-derived influenza A-like virus H18N11. Emerg. Microbes Infect. 8, 556–563. doi: 10.1080/22221751.2019.1599301
Ciminski, K., Ran, W., Gorka, M., Lee, J., Malmlov, A., Schinkothe, J., et al. (2019b). Bat influenza viruses transmit among bats but are poorly adapted to non-bat species. Nat. Microbiol. 4, 2298–2309. doi: 10.1038/s41564-019-0556-9
Ciminski, K., Thamamongood, T., Zimmer, G., and Schwemmle, M. (2017). Novel insights into bat influenza A viruses. J. Gen. Virol. 98, 2393–2400. doi: 10.1099/jgv.0.000927
Dlugolenski, D., Jones, L., Tompkins, S. M., Crameri, G., Wang, L. F., and Tripp, R. A. (2013). Bat cells from Pteropus alecto are susceptible to influenza A virus infection and reassortment. Influenza Other Respir. 7, 900–903. doi: 10.1111/irv.12128
Fereidouni, S., Kwasnitschka, L., Balkema Buschmann, A., Muller, T., Freuling, C., Schatz, J., et al. (2015). No virological evidence for an influenza A–like virus in European bats. Zoonoses Public Health 62, 187–189. doi: 10.1111/zph.12131
Field, H., Jordan, D., Edson, D., Morris, S., Melville, D., Parry-Jones, K., et al. (2015). Spatiotemporal aspects of Hendra virus infection in Pteropid bats (flying-foxes) in Eastern Australia. PLoS ONE 10:e0144055. doi: 10.1371/journal.pone.0144055
Freidl, G. S., Binger, T., Muller, M. A., De Bruin, E., Van Beek, J., Corman, V. M., et al. (2015). Serological evidence of influenza A viruses in frugivorous bats from Africa. PLoS ONE 10:e0127035. doi: 10.1371/journal.pone.0127035
Garcia-Sastre, A. (2012). The neuraminidase of bat influenza viruses is not a neuraminidase. Proc. Natl. Acad. Sci. U. S. A. 109, 18635–18636. doi: 10.1073/pnas.1215857109
Giotis, E. S., Carnell, G., Young, E. F., Ghanny, S., Soteropoulos, P., Wang, L. F., et al. (2019). Entry of the bat influenza H17N10 virus into mammalian cells is enabled by the MHC class II HLA-DR receptor. Nat. Microbiol. 4, 2035–2038. doi: 10.1038/s41564-019-0517-3
Hayman, D. T., Bowen, R. A., Cryan, P. M., Mccracken, G. F., O'shea, T. J., Peel, A. J., et al. (2013). Ecology of zoonotic infectious diseases in bats: current knowledge and future directions. Zoonoses Public Health 60, 2–21. doi: 10.1111/zph.12000
Hoffmann, M., Kruger, N., Zmora, P., Wrensch, F., Herrler, G., and Pohlmann, S. (2016). The hemagglutinin of bat-associated influenza viruses is activated by TMPRSS2 for pH-dependent entry into bat but not human cells. PLoS ONE 11:e0152134. doi: 10.1371/journal.pone.0152134
Isaeva, E. I., Belkina, T. S., Rovnova, Z. I., Kosiakov, P. N., and Selivanov Ia, M. (1982). Antigenic determinants of human influenza viruses among the influenza viruses isolated from animals. Vopr. Virusol. 27, 681–686.
Jenkins, C. N., Pimm, S. L., and Joppa, L. N. (2013). Global patterns of terrestrial vertebrate diversity and conservation. Proc. Natl. Acad. Sci. U. S. A. 110, E2602–E2610. doi: 10.1073/pnas.1302251110
Jones, E. Y., Fugger, L., Strominger, J. L., and Siebold, C. (2006). MHC class II proteins and disease: a structural perspective. Nat. Rev. Immunol. 6, 271–282. doi: 10.1038/nri1805
Juozapaitis, M., Aguiar Moreira, E., Mena, I., Giese, S., Riegger, D., Pohlmann, A., et al. (2014). An infectious bat-derived chimeric influenza virus harbouring the entry machinery of an influenza A virus. Nat. Commun. 5:4448. doi: 10.1038/ncomms5448
Karakus, U., Thamamongood, T., Ciminski, K., Ran, W., Gunther, S. C., Pohl, M. O., et al. (2019). MHC class II proteins mediate cross-species entry of bat influenza viruses. Nature 567, 109–112. doi: 10.1038/s41586-019-0955-3
Kelkar, S. D., Kadam, S. S., and Banerjee, K. (1981). Haemagglutination inhibition antibodies against influenza virus in bats. Indian J. Med. Res. 74, 147–152.
Li, Q., Sun, X., Li, Z., Liu, Y., Vavricka, C. J., Qi, J., et al. (2012). Structural and functional characterization of neuraminidase-like molecule N10 derived from bat influenza A virus. Proc. Natl. Acad. Sci. U. S. A 109, 18897–18902. doi: 10.1073/pnas.1211037109
L'vov, D. K., Easterday, B., Hinshow, W., Dandurov Iu, V., and Arkhipov, P. N. (1979). Isolation of strains of the Hong Kong complex (H3N2) influenza virus from Nyctalus noctula bats in Kazakhstan. Vopr. Virusol. 338–341.
Maruyama, J., Nao, N., Miyamoto, H., Maeda, K., Ogawa, H., Yoshida, R., et al. (2016). Characterization of the glycoproteins of bat-derived influenza viruses. Virology 488, 43–50. doi: 10.1016/j.virol.2015.11.002
Moratelli, R., and Calisher, C. H. (2015). Bats and zoonotic viruses: can we confidently link bats with emerging deadly viruses? Mem. Inst. Oswaldo Cruz 110, 1–22. doi: 10.1590/0074-02760150048
Moreira, E. A., Locher, S., Kolesnikova, L., Bolte, H., Aydillo, T., Garcia-Sastre, A., et al. (2016). Synthetically derived bat influenza A-like viruses reveal a cell type- but not species-specific tropism. Proc. Natl. Acad. Sci. U. S. A. 113, 12797–12802. doi: 10.1073/pnas.1608821113
Ng, J. H. J., Tachedjian, M., Wang, L. F., and Baker, M. L. (2017). Insights into the ancestral organisation of the mammalian MHC class II region from the genome of the pteropid bat, pteropus alecto. BMC Genomics 18:388. doi: 10.1186/s12864-017-3760-0
Ressing, M. E., van Leeuwen, D., Verreck, F. A., Gomez, R., Heemskerk, B., Toebes, M., et al. (2003). Interference with T cell receptor-HLA-DR interactions by Epstein-Barr virus gp42 results in reduced T helper cell recognition. Proc. Natl. Acad. Sci. U. S. A. 100, 11583–11588. doi: 10.1073/pnas.2034960100
Ressing, M. E., van Leeuwen, D., Verreck, F. A., Keating, S., Gomez, R., Franken, K. L., et al. (2005). Epstein-Barr virus gp42 is posttranslationally modified to produce soluble gp42 that mediates HLA class II immune evasion. J. Virol. 79, 841–852. doi: 10.1128/JVI.79.2.841-852.2005
Roche, P. A., and Furuta, K. (2015). The ins and outs of MHC class II-mediated antigen processing and presentation. Nat. Rev. Immunol. 15, 203–216. doi: 10.1038/nri3818
Rupprecht, C. E., Hanlon, C. A., and Hemachudha, T. (2002). Rabies re-examined. Lancet Infect. Dis. 2, 327–343. doi: 10.1016/S1473-3099(02)00287-6
Sato, M., Maruyama, J., Kondoh, T., Nao, N., Miyamoto, H., Takadate, Y., et al. (2019). Generation of bat-derived influenza viruses and their reassortants. Sci. Rep. 9:1158. doi: 10.1038/s41598-018-37830-x
Simonsen, L. (1999). The global impact of influenza on morbidity and mortality. Vaccine 17(Suppl.1), S3–S10. doi: 10.1016/S0264-410X(99)00099-7
Sun, X., Shi, Y., Lu, X., He, J., Gao, F., Yan, J., et al. (2013). Bat-derived influenza hemagglutinin H17 does not bind canonical avian or human receptors and most likely uses a unique entry mechanism. Cell Rep. 3, 769–778. doi: 10.1016/j.celrep.2013.01.025
Tefsen, B., Lu, G., Zhu, Y., Haywood, J., Zhao, L., Deng, T., et al. (2014). The N-terminal domain of PA from bat-derived influenza-like virus H17N10 has endonuclease activity. J. Virol. 88, 1935–1941. doi: 10.1128/JVI.03270-13
Tong, S., Li, Y., Rivailler, P., Conrardy, C., Castillo, D. A., Chen, L. M., et al. (2012). A distinct lineage of influenza A virus from bats. Proc. Natl. Acad. Sci. U. S. A 109, 4269–4274. doi: 10.1073/pnas.1116200109
Tong, S., Zhu, X., Li, Y., Shi, M., Zhang, J., Bourgeois, M., et al. (2013). New world bats harbor diverse influenza A viruses. PLoS Pathog. 9:e1003657. doi: 10.1371/journal.ppat.1003657
Wacharapluesadee, S., Boongird, K., Wanghongsa, S., Ratanasetyuth, N., Supavonwong, P., Saengsen, D., et al. (2010). A longitudinal study of the prevalence of Nipah virus in Pteropus lylei bats in Thailand: evidence for seasonal preference in disease transmission. Vector Borne Zoonotic Dis. 10, 183–190. doi: 10.1089/vbz.2008.0105
Wiertz, E. J., Devlin, R., Collins, H. L., and Ressing, M. E. (2007). Herpesvirus interference with major histocompatibility complex class II-restricted T-cell activation. J. Virol. 81, 4389–4396. doi: 10.1128/JVI.01525-06
Wosen, J. E., Mukhopadhyay, D., Macaubas, C., and Mellins, E. D. (2018). Epithelial MHC class II expression and its role in antigen presentation in the gastrointestinal and respiratory tracts. Front. Immunol. 9:2144. doi: 10.3389/fimmu.2018.02144
Zhong, G., Fan, S., Hatta, M., Nakatsu, S., Walters, K. B., Lopes, T. J. S., et al. (2019). Mutations in the NA-like protein of bat influenza H18N11 virus enhance virus replication in mammalian cells, mice, and ferrets. J. Virol. 95:01416-19. doi: 10.1128/JVI.01416-19
Zhou, B., Ma, J., Liu, Q., Bawa, B., Wang, W., Shabman, R. S., et al. (2014). Characterization of uncultivable bat influenza virus using a replicative synthetic virus. PLoS Pathog. 10:e1004420. doi: 10.1371/journal.ppat.1004420
Keywords: bats, influenza virus, haemagglutinin, neuraminidase, Major Histocompatibility Complex (MHC) class II, sialic acids
Citation: Giotis ES (2020) Inferring the Urban Transmission Potential of Bat Influenza Viruses. Front. Cell. Infect. Microbiol. 10:264. doi: 10.3389/fcimb.2020.00264
Received: 03 February 2020; Accepted: 04 May 2020;
Published: 03 June 2020.
Edited by:
Rosa M. Del Angel, Instituto Politécnico Nacional de México (CINVESTAV), MexicoReviewed by:
Teneema Kuriakose, St. Jude Children's Research Hospital, United StatesBinod Kumar, Loyola University Chicago, United States
Copyright © 2020 Giotis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Efstathios S. Giotis, ZS5naW90aXNAaW1wZXJpYWwuYWMudWs=